Embed Size (px)
Citation preview

Regulation of the a 2 p l and a3(31 Isozymes of the Na,K-ATPase by Ca2+,
PKA, and PKC" GUSTAVO BLANCOb AND ROBERT W. MERCER
Department of Cell Biology and Physiology Washington University School of Medicine
St. Louis, Missouri 63110
The molecular mechanisms underlying the regulation of the Na,K-ATPase are poorly understood. Of the various isoforms of the catalytic a ( a l , a2, a3 , and a4) and glycosylated f3 (PI and p2) subunits that comprise the Na,K-ATPase of mammalian cells,' the a 1 p 1 isozyme has generally been used to study the modulation of activity by intracellular Ca'+ and regulatory phosphorylation/dephosphorylation.z~3 On the other hand, there is relatively little evidence for the regulation of the a2 and a 3 isoforms by second messengers. Here we show the effects of Ca2+ and activators of protein kinase A and C (PKA, PKC) on the activity of the Na,K-ATPase isozymes of the rat, expressed in Sf-9 insect cells using recombinant baculoviruses.
METHODS
Sf-9 cells grown in 150-mm petri dishes were infected with recombinant baculovi- ruses containing the rat al , 012, 013, and p l Na,K-ATPase cDNAs." At 40 hours postinfection, cells were treated with the PKA and PKC modulators. For PKA activation, 2 mM of N6,2'-0-dibutyryladenosine 3':5'-cyclic monophosphate (dibu- tyryl CAMP) was used for 1 hour. As an antagonist of PKA, 8-bromoadenosine 3'5' cyclic monophosphothioate, Rp-isomer was added at a concentration of 50 pM I hour before the addition of the dibutyryl CAMP. PKC activation was obtained with phorbol 12-myristate 13-acetate (PMA) at 1 pM for 1 hour. Staurosporine (200 nM) or 50 pM 1 -(5-isoquinolinesulfonyl)-2-methylpiperazine (H7) was used as PKC inhibitors 1 hour before and during treatment with PMA. After treatment, cells were harvested, centrifuged at 1,500 X g for 10 minutes, and resuspended in 10 mM imidazole hydrochloride (pH 7.3, 1 mM EGTA, with 100 nM tautomycin to inhibit protein phosphatases.
Enzyme activity from permeabilized intact cells was determined by the initial rate of release of 32P1 from [y-3zP]ATP.4 For Ca2+ dose-response curves, a cellular membrane fraction was ~ repa red .~ Maximal binding of [3H]ouabain was performed as
'This work was supported by National Institutes of Health grant GM 39746 and the George M. O'Brien Kidney and Urological Diseases Center at Washington University School of Medicine.
bAddress for correspondence: Dr. Custavo Blanco, Department of Cell Biology and Physiology, Washington University School of Medicine, 660 S . Euclid Avenue, St. Louis, MO 63 I10 (tel: 3 14-362-6922; fax: 3 14-362-7463; e-mail: [email protected]).
572

BLANCO & MERCER: REGULATION OF ISOZYMES 573
before4 and was calculated from Scatchard plots of the data, which in all cases showed a single population of binding sites. Curve fitting of the experimental data was carried out using the equations described previo~sly.~ Free Ca2+ concentrations were calcu- lated with a program written by M. Kurzmak (University of Maryland), which was based on the formulations of Fabiato and Fabiato.s
RESULTS AND DISCUSSION
To investigate the effect of Ca2+ on Na,K-ATPase isozyme activity, the activity of the isozymes in the presence of various concentrations of Ca2+ was determined. The Ca2+ dose-response curves indicated that the individual isozymes are differentially inhibited by Ca2+. For example, the calculated inhibition constants of the alpl , a2P1, and a3Pl isozymes were: 1.0 Z 0.2 X M, 7.3 ? 4.6 X lop6 M, and 1.9 +- 1.0 X M, respectively. This suggests that in excitable cells after depolarization, when the intracellular Ca2+ concentration approaches 5-10 $4, the a1 isozyme remains active, while the a 2 and a3 are functioning at approximately half their maximal capability. Thus, intracellular Ca2+ may regulate cellular contractility and excitability by selectively inhibiting specific Na,K-ATPase isozymes. This may be particularly relevant in the heart, where the rise in intracellular Ca2+ elicited by the cardiotonic steroids may be enhanced by further inhibition of the ouabain-sensitive a 2 and a3 isoforms.
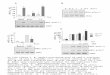
Phosphorylation of discrete Serflhr residues of the a1 subunit influences the activity of the enzyme. Although the effect is cell-type dependent, phosphorylation has primarily been associated with inhibition of the N ~ , K - A T P ~ s ~ . ~ To determine if protein kinases also influence the function of the other isoforms, the effects of activators of PKA and PKC on the specific activity of Na-pump isozymes were investigated. FIGURE 1 shows that activation of PKC with the phorbol ester, phorbol 12-myristate 13-acetate (PMA), results in a decrease in activity of the a 1 P I , a2P 1, and a3P 1 isozymes expressed in Sf-9 insect cells. The effect of PMA could partially be reversed by treating the cells with the specific inhibitors of PKC, staurosporin and H7. Moreover, Na,K-ATPase activity could be restored by treatment with protein phosphatase 2A. To assess whether the effect of PMA was a result of complete inactivation of a fraction of the Na,K-ATPase or partial inactivation of the total Na,K-ATPase, the turnover numbers of the a 2 and a3 isoforms were determined. The molecular activity of the isozymes was estimated from the maximal Na,K-ATPase activity and the amount of equilibrium [3H]ouabain binding. These values indicated that PMA treatment of the a2 and a 3 isoforms reduced molecular activity by 71 Z 4% and 55 2 2% of the control, respectively. This decrease in molecular activity accounts for the similar reduction in ATP hydrolysis. In contrast to these results, PKA activation by dibutyryl CAMP resulted in an increase in a3P 1 activity and a decrease in a 1 P 1 and a 2 p 1. This effect was blocked by the PKA competitive inhibitor 8-bromoadeno- sine cyclic monophosphothioate Rp-isomer (FIG. 2). Once more, the variation in activity paralleled changes in the molecular activity of the treated isozymes, with values of 88 +- 6% and 123 ? 4% for the a 2 and a 3 isoforms, respectively. The mechanisms by which PKA and PKC regulate the Na,K-ATPase isozymes are difficult to explain. For the rat a1 isoform, phosphorylation by PKA has been mapped to Ser943, while PKC phosphorylates the polypeptide on Serl6 and Ser23. Set943 is conserved in the a 2 and a3 polypeptides; however, Serl6 is not and Ser23 is present

574 ANNALS NEW YORK ACADEMY OF SCIENCES
in 011 and 013. Thus, the effect of PKA and PKC activators may depend on phosphorylation of additional groups on the a polypeptides, on differential modifica- tions of the structure and hence function of the proteins, or on the involvement of another second messenger(s). In any case, because the Na,K-ATPase isoforms have unique kinetic characteristics? the differential modulation of their activity may be important in adapting Na-pump function to specific cellular requirements.
120 A -
40
20 s
10
T T
T
PMA PMA PMA PMA PPPA Staurosporin H7
FIGURE 1. Effect of phorbol 12-myristate 13-acetate (PMA) on the Na,K-ATPase activity of the rat a l p l . a2p1, and a3pl Na,K-ATPase isozymes. Sf-9 cells expressing the a l p l , (u2pl. or a301 isofoms were treated 40 hours after infection with PMA alone (1 pM for 1 hour) or with the addition of either staurosporine (200 nM) or H7 (50 pM). Cells were then processed for Na,K-ATPase activity. Aliquots of the cells treated with only PMA were homogenized and incubated for 1 hour at 25°C in the absence and presence of 2 U/mg protein of protein phosphatase 2A (PP2A). Na,K-ATPase activity was assayed in a reaction medium containing: 120 mM NaCI, 30 mM KCI, 3 mM MgCI2, 0.2 mM EGTA, 3 rnM [y-32P]ATP-cold ATP, 30 mM Tris-HCI (pH 7.4) in the absence or presence of 1 mM ouabain. Values are expressed as a percentage of the untreated cells, taken as 100%. Each value is the mean, and error bars represent the standard errors of the mean of three to seven experiments performed in triplicate on samples obtained from different infections.

BLANCO & MERCER: REGULATION OF ISOZYMES
150
’401 130
Dibutyryl cAMP
1
T
Dibutytyl cAMP 8-Bromo cAMPT Rp-isomer
575
FIGURE 2. Effect of N6,2’-O-dibutyryladenosine 3’:5’-cyclic monophosphate (dibutyryl CAMP) on the Na,K-ATPase activity of the rat cxIpI, a2pl, and a 3 p l Na,K-ATPase isozymes. Sf9 cells expressing the a l p l , a2pl . or a3p l isoforms were treated 40 hours after infection with dibutyryl cAMP alone (2 mM for 1 hour) or with the addition of 50 pM of 8-bromo adenosine cyclic monophosphothioate, Rp-isomer (8-bromo cAMF’T). Na,K-ATPase activity was deter- mined as described in Figure I . Values are expressed as percentage of the untreated cells, taken as 100%. Each value is the mean, and error bars represent the standard errors of the mean of three to seven experiments performed in triplicate on samples obtained from different infections.
REFERENCES
1.
2.
3.
SWEADNER, K. J. 1989. lsozymes of the Na,K-ATPase. Biochim. Biophys. Acta 988: 185- 220.
BEAUCE, L. & M. A. CAMPOS. 1983. Calcium inhibition of the ATPase and phosphatase activities of Na,K-ATPase. Biochim. Biophys. Acta 729: 137-149.
BERTORELLO, A. M. & A. I. KATZ. 1993. Short term regulation of renal Na,K-ATPase activity: Physiological relevance and cellular mechanisms. Am. J. Physiol. 265: F743- F755.
BLANCO. G., G. SANCHEZ & R. W. MERCER. 1995. Comparison of the enzymatic properties of the Na,K-ATPase a 3 p l and a3P2 isozymes. Biochemistry 34: 9897-9903.
FABIATO, A. & F. FABIATO. 1979. Calculator programs for computing the composition of the solutions containing multiple metals and ligands used for experiments in shnned muscle cells. J. Physiol. 75: 463-505.
4.
5.