Embed Size (px)
Citation preview

INTRODUCTION
Myelin, the fatty insulation surrounding vertebrate CNS axonsis produced by a specific class of macroglial cells termed oligo-dendrocytes (Peters et al., 1990). During development, the pro-liferation and differentiation of oligodendrocytes from cells ofthe neural tube is closely regulated such that in any particularregion of the CNS, mature oligodendrocytes appear at aspecific stage of development. For example, in rat optic nervesuch cells first appear around the day of birth (Skoff et al.,1976a,b), while in rat spinal cord they appear aroundembryonic day 17 (E17) (Warf et al., 1991).
During their cellular maturation, oligodendrocyte precursorsgo through a series of discrete differentiative steps which canbe identified by the expression of particular cell surfaceepitopes and proliferative responses (Raff, 1989; Gard andPfieffer, 1990). In neonatal rat optic nerve cultures, oligoden-drocytes develop from bipotential progenitor cells termed O-2A progenitors (Raff, 1989). These cells constitutively giverise to oligodendrocytes, but can be induced to differentiate invitro into a distinct class of astrocytes by exposure to CNTFand basal lamina components (Hughes et al., 1988; Lillien et
al., 1990). Initially, O-2A progenitors express tetrasialogan-gliosides recognized by the A2B5 antibody (Fredman et al.,1984; Eisenbarth et al., 1979). Subsequently, oligodendrocyteprecursors begin to express antigens recognized by the mono-clonal antibody O4 (Sommer and Schachner, 1981), whichappears to bind to sulfatide, seminolipid and an unidentifiedantigen (Sommer and Schachner, 1981; Bansal et al., 1989).This intermediate stage of development is maintained for 2-3days (Gard and Pfeiffer, 1990) before the expression of a thirdmarker, galactocerebroside (GC) (Ranscht et al., 1982) a majormyelin glycolipid. Anti-GC antibodies can be used to identifyspecifically oligodendrocytes as they mature in vitro (Raff etal., 1978).
Oligodendrocyte precursors proliferate in response to anumber of different mitogens at particular stages of their differ-entiation. A2B5-immunoreactive oligodendrocyte precursorsproliferate in response to mitogens present in both astrocyteand neuronal conditioned medium (Gard and Pfieffer, 1990),while O4-immunoreactive oligodendrocyte precursors prolif-erate in response to mitogens found only in neuronal condi-tioned medium (Gard and Pfieffer, 1990). Furthermore, A2B5-immunoreactive precursors proliferate in response to
649Development 120, 649-660 (1994)Printed in Great Britain © The Company of Biologists Limited 1994
During development, oligodendrocyte precursors undergosequential stages of differentiation characterized byexpression of distinct cell surface properties and prolifera-tive responses. Although both PDGF and bFGF aremitogenic for these cells, the factors that regulate the pro-gression of oligodendrocyte precursors through their dif-ferentiative program remain unclear.
One factor present in the embryonic spinal cord thatmay regulate differentiation of oligodendrocyte precursorsis retinoic acid. Here we show that retinoic acid inhibits thematuration of embryonic spinal cord oligodendrocyte pre-cursors in vitro at an early, highly motile stage of differ-entiation, characterized by the expression of A2B5immunoreactivity. Basic FGF acts both as a mitogen andan inhibitor of spinal cord oligodendrocyte precursor mat-uration, but at a significantly later stage of differentiation,characterized by the expression of O4 immunoreactivity.
In the presence of RA both the mitogenic and differen-tiation inhibiting effects of bFGF are abolished, consistentwith RA acting as an early regulator of oligodendrocytedifferentiation. During embryonic development, oligoden-drocyte precursors arise initially from a distinct group ofcells at the ventral ventricular zone of the spinal cord.Myelination of the entire spinal cord is dependent on themigration of immature precursor cells to peripheral devel-oping white matter. Since the embryonic spinal cord hasthe capacity to release relatively high levels of retinoids, wepropose that RA inhibits oligodendrocyte differentiationduring early embryonic development permitting theirdispersal throughout the entire spinal cord.
Key words: oligodendrocyte, retinoic acid, spinal cord, PDGF,bFGF, mitogen
SUMMARY
Regulation of oligodendrocyte differentiation: a role for retinoic acid in the
spinal cord
Elizabeth Noll* and Robert H. Miller†
Department of Neurosciences, Case Western Reserve University, School of Medicine, Cleveland, OH 44106, USA
*Present address: Department of Genetics, Harvard Medical School, 200 Longwood Ave, Boston MA 02115, USA†Author for correspondence

650
platelet-derived growth factor (PDGF) (Richardson et al.,1988; Raff et al., 1988; Fok-Seang and Miller, 1993). In spinalcord cultures, however, this proliferative response is lost ascells acquire O4 immunoreactivity (Fok-Seang and Miller,1993). In contrast, basic fibroblast growth factor (bFGF)promotes proliferation in both A2B5- and O4-immunoreactiveprecursors (Bogler et al., 1990; McKinnon et al., 1990; Fok-Seang and Miller, 1993). These observations suggest that theproliferative response of oligodendrocyte precursors is alteredduring the maturation step characterized by the acquisition ofO4 immunoreactivity (Gard and Pfieffer, 1990; Fok-Seang andMiller, 1993).
In late embryonic and early neonatal animals, oligodendro-cyte precursors intrinsically possess all the informationrequired for their subsequent differentiation. For example, thetiming of oligodendrocyte development does not appear todepend on positional information in the intact CNS. Oligo-dendrocytes develop GC immunoreactivity on the sameschedule in high density dissociated cell cultures derived fromeither brain or spinal cord as they do in the intact tissue (Abneyet al., 1981; Williams et al., 1985; Warf et al., 1991). Inaddition, oligodendrocyte development does not appear torequire the presence of other neural cell types. Single oligo-dendrocyte precursor cells grown in clonal isolation retain thecapacity to differentiate into GC+ cells (Temple and Raff,1985, 1986).
Although all the information necessary for oligodendrocytedifferentiation to a GC+ state is present in the precursor popu-lation, the temporal progression through the different stages ofmaturation appears to be influenced by environmental factors.For example, while in high density spinal cord cultures oligo-dendrocyte precursors in vitro undergo differentiationaccording to the in vivo schedule, in the absence of sufficientmitogen (PDGF), oligodendrocyte precursors will differentiateprematurely (Raff et al., 1988). Furthermore, when oligoden-drocyte precursors are grown in the presence of both PDGFand bFGF, precursor proliferation is stimulated, while theirdifferentiation is delayed at a maturational stage prior to theacquisition of GC immunoreactivity (Bogler et al., 1990;McKinnon et al., 1990).
Oligodendrocyte precursors are strikingly motile (LaChapelle et al., 1984; Noble et al., 1988) and may be influ-enced by several different tissue environments during theirearly differentiation (Small et al., 1987; Curtis et al., 1988;Hardy and Reynolds, 1991). The migrational behavior of oligo-dendrocyte precursors is more pronounced during the earlystages of their differentiation (Pfieffer et al., 1993; Warringtonet al., 1993). Thus, while A2B5 and O4-immunoreactive oligo-dendrocyte precursors are highly motile in vitro (Temple andRaff, 1986), this motile behavior is significantly reduced bothin vivo and in vitro as the cells acquire GC immunoreactivity(Curtis et al., 1988). During development of the rat optic nerve,oligodendrocyte precursors appear to originate in the brain andsubsequently migrate into the nerve (Small et al., 1987). In therat spinal cord, however, oligodendrocyte precursors appear tooriginate from a discrete cluster of cells at the ventral ventric-ular zone dorsal to the ventral midline (Warf et al., 1991;Pringle and Richardson, 1993; Noll and Miller, 1993) and sub-sequently migrate dorsally and laterally to the developingwhite matter (Warf et al., 1991; Noll and Miller, 1993). Thisventral origin of oligodendrocyte precursors in the embryonic
spinal cord is in close proximity to the notochord and floor-plate, a group of specialized cells important in the morpho-genetic development of the spinal cord (Van Straaten et al.,1989; Jessel and Dodd, 1992). Floorplate cells provide posi-tional information involved in the control of cell patterning inthe vertebrate central nervous system (Tessier-Levigne et al.,1988; Yamada et al., 1991; Ruiz i Altaba and Jessell, 1991)and have the capacity to synthesize relatively high levels ofretinoic acid (RA) and a precursor for the related retinoid 3,4-dihydroretinoic acid (Wagner et al., 1990). Other regions of theembryonic spinal cord also contain higher levels of retinoidsthan more anterior regions of the CNS (Wagner et al., 1993;Mendelsohn et al., 1991) suggesting that RA levels areelevated in embryonic spinal cord. We have examined theinfluence of RA on oligodendrocyte differentiation in vitrounder conditions that allow the differentiation of oligodendro-cyte precursors on the same schedule as that observed in vivo(Warf et al., 1991). We show that when embryonic spinal cordcultures are grown in the presence of exogenous retinoic acid,oligodendrocyte precursor cell maturation is inhibited early intheir differentiation. Removal of exogenous retinoic acidallowed the precursors to progress through their normal stagesof differentiation.
Since in developing spinal cord oligodendrocytes originatefrom a discrete region of the ventral cord, the successful myeli-nation of the entire cord depends critically on the subsequentmigration and differentiation of these cells. As the capacity formigration is closely correlated with an immature phenotype(Temple and Raff, 1986; Pfieffer et al., 1993), we propose thatretinoic acid present at relatively high levels in the embryonicspinal cord inhibits oligodendrocyte differentiation until theprecursor cells have migrated away from their localized originand uniformly populated the entire spinal cord.
MATERIALS AND METHODS
Spinal cord dissociation and cell cultureEmbryonic rat spinal cord cultures were prepared from the thoracicand lumbar regions of embryonic day (E) E14, E16 and E18 Sprague-Dawley rat embryos. After removal of the meninges, spinal cords weredissociated into single cells using trypsin and EDTA as previouslydescribed (Warf et al., 1991). Dissociated cells were resuspended inDMEM medium with 10% fetal bovine serum (FBS), plated onto 12mm poly-L-lysine (0.1 mg/ml Sigma) coated coverslips at a density of0.5
×106 cells/coverslip for high density cultures. For low densitycultures, only the ventral third of the spinal cord was used to enrichfor oligodendrocyte precursors and the cells plated at a density of0.5×103 cells/coverslip. Approximately 60% of cells in such cultureswere oligodendrocyte precursors. All supplements were added tocultures at the time of plating. After 12 hours to allow cell attachment,cultures were switched to N2 medium (Bottenstein and Sato, 1979)containing 1% FBS and all supplements were replaced at that time.Each experiment consisted of sister cultures grown under five differentconditions: (1) control without any supplements, (2) PDGF with 8ng/ml PDGF, (human natural, AB heterodimer; #40113, CollaborativeBiomedical Products), (3) PDGF and FGF with 8 ng/ml PDGF+10ng/ml bFGF, (bFGF-bovine pituitary; #40002, Collaborative Biomed-ical Products), (4) PDGF, bFGF and RA (8 ng PDGF/ml+10 ngbFGF/ml+1 µM all-
trans retinoic acid/ml; (#R-2625, Sigma); and (5)PDGF and RA (8 ng PDGF/ml+1 µM all-trans retinoic acid). The con-centrations of supplements were selected based on previous studies.For example, PDGF and bFGF concentrations are similar to those used
E. Noll and R. H. Miller

651Inhibition of oligodendrocyte differentiation by RA
in analysis of purified progenitor cells (Bogler et al., 1990; McKinnonet al., 1990) while RA concentrations are similar to those that causeanteroposterior transformations in the developing CNS (Durston et al.,1989). All supplements were replaced every day for the duration of theexperiment and the complete medium was replaced every 2 or 3 days.Exogenous PDGF was added to all but control experiments toeliminate the possibility that any effects of RA were mediated indi-rectly through inhibiting the release of mitogen from other cells in theculture (Richardson et al., 1988; Yeh et al., 1991).
Immunofluorescence labelingCultures were fixed with 4% paraformaldehyde at 37°C for 3-5minutes and rinsed briefly in DMEM. Antibody incubations wereperformed at room temperature for 30 minutes in DMEM with 50%normal goat serum (NGS) added. Sister cultures were incubated withmonoclonal antibodies A2B5 (Eisenbarth et al., 1979; supernatant 1:1),O4 (Sommer and Schachner, 1981; supernatant 1:10) or anti-galacto-cerebroside (anti-GC, Ranscht et al., 1982; ascites fluid 1:200). Afterrinsing, cells were incubated for 30 minutes in either fluorescein-con-jugated goat anti-mouse IgM, (for A2B5 and O4) or rhodamine-con-jugated goat anti-mouse IgG (for anti-GC). Both secondary antibodies(Cappel) were diluted 1:50 in DMEM+50% NGS. Cultures were thenrinsed and mounted in 80% glycerol/PBS with 5% propyl gallate addedto prevent fluorochrome bleaching. In control cultures, deletion of theprimary antibody or substitution with normal mouse serum resulted inthe total absence of specific staining. Cells were viewed using a Nikonoptiphot microscope equipped with epifluorescent illumination andphotographed on Tri-X (400 ASA) film.
Quantitative analysisTo determine the number of immunoreactive cells that developed ineach culture, the total number of labeled cells were counted in 10 con-secutive fields under a ×20 objective using rhodamine or fluoresceinfilters. Counts were adjusted to reflect the area occupied by all cellson the coverslip. In all cases, the results represent the means ± s.d. ofcell counts from at least three separate experiments. For each exper-iment, the cells in control and supplement containing cultures werederived from a single dissociation procedure, grown on similarlytreated substrata under otherwise identical conditions and all supple-ments were added to individual cultures simultaneously to ensure con-sistency between cultures. Regardless of the initial age of the animalsused to establish the culture, all quantitative analysis was performedon cultures at an equivalent age of the first day after birth (P1), and4 days (P5) later in recovery experiments. Previous studies indicatedthat under these conditions reproducible numbers of oligodendrocytesdeveloped from known numbers of spinal cord cells in culture andcould be accurately quantified (Warf et al., 1991). Statistical com-parison of cell numbers in the presence of the different culture sup-plements were made using an unpaired t-test.
RESULTS
Spinal cord oligodendrocyte precursors go through a series ofmaturational stages, which can be identified by the sequentialexpression of cell surface epitopes. Discrete developmentalstages can be recognized by the antibodies A2B5 (detectableas early as E12), O4 (first detectable at E16) and anti-GC (firstdetectable at E17/18) in thoracolumbar rat spinal cord (Warfet al., 1991). This temporal sequence of oligodendrocyte mat-uration occurs on the same schedule both in vivo and in vitro,and, thus, it has been possible to examine the influence of avariety of putative cell regulatory factors such as PDGF, bFGFand retinoic acid on the differentiation of spinal cord oligo-dendrocytes in vitro.
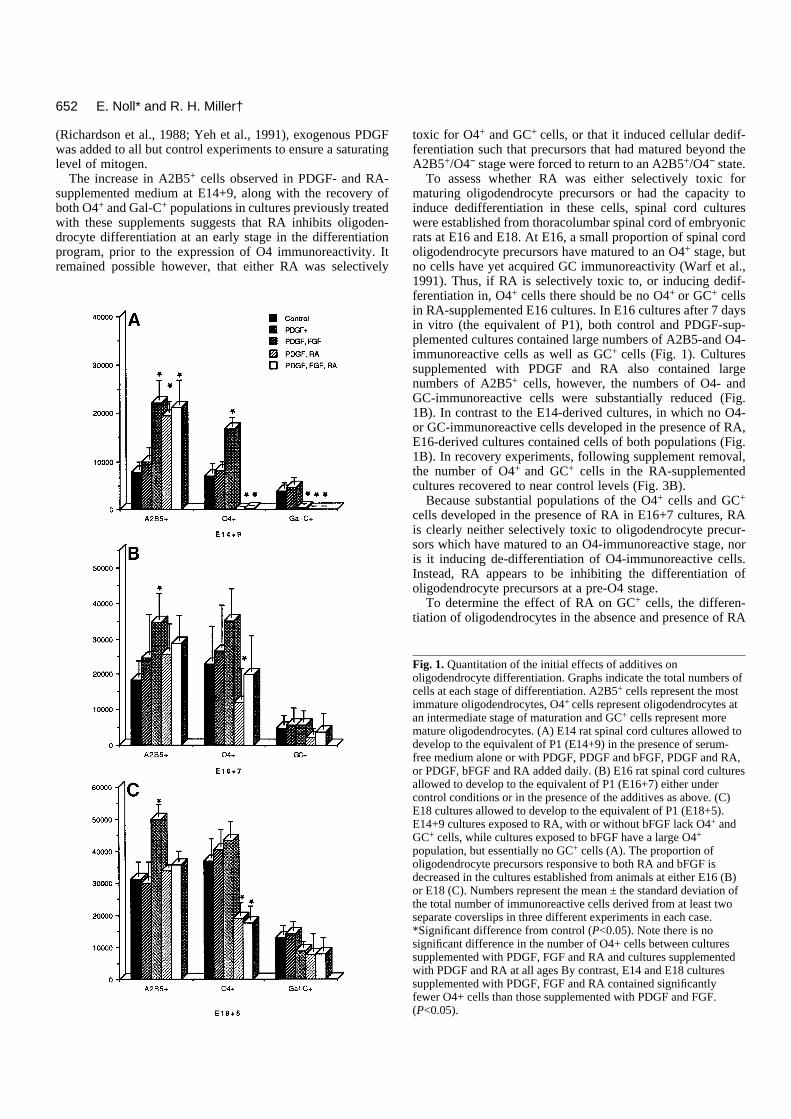
RA but not PDGF inhibits oligodendrocytedifferentiationTo assess the influence of PDGF and retinoic acid on the dif-ferentiative program of spinal cord oligodendrocytes in vitro,cultures derived from E14 thoracic and lumbar spinal cordwere grown either without supplements (control), in thepresence of PDGF or in the presence of PDGF and RA. After9 days in culture (the equivalent of P1), cultures were labeledwith either A2B5, O4 or anti-GC antibodies and the number ofimmunoreactive cells in the culture determined.
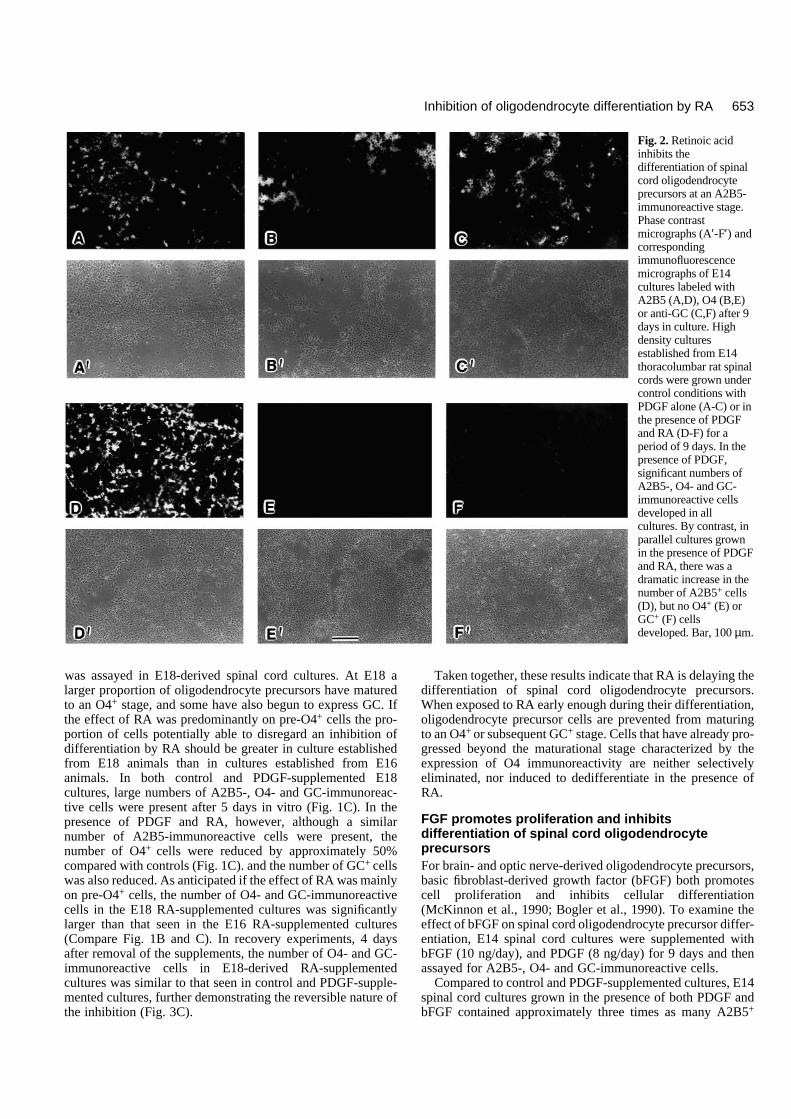
In both control and PDGF-supplemented cultures, similarnumbers of A2B5-, O4- and GC-immunoreactive cells werepresent (Fig. 1A), suggesting that addition of PDGF did notsubstantially alter the timing or extent of differentiation ofspinal cord oligodendrocytes. By contrast, cultures supple-mented with PDGF and RA contained virtually no O4- or GC-immunoreactive cells (Fig. 2), and the number of A2B5-immunoreactive cells was over two-fold that seen in controlcultures (Fig. 1A). A number of explanations may account forthe absence of O4+ and Gal-C+ cells and the increase in A2B5+
cells in PDGF- and RA-supplemented cultures. For example,supplement addition may (1) inhibit oligodendrocyte differen-tiation at an early A2B5+/O4−/GC− stage, (2) permanentlyblock the capacity of spinal cord cells to give rise to oligo-dendrocytes, (3) selectively kill oligodendrocyte precursorsthat have matured beyond an O4 stage, or, (4) induce dedif-ferentiation of O4+ or GC+ oligodendrocytes back to an A2B5+
stage. To determine if PDGF- and RA-treated E14 spinal cord
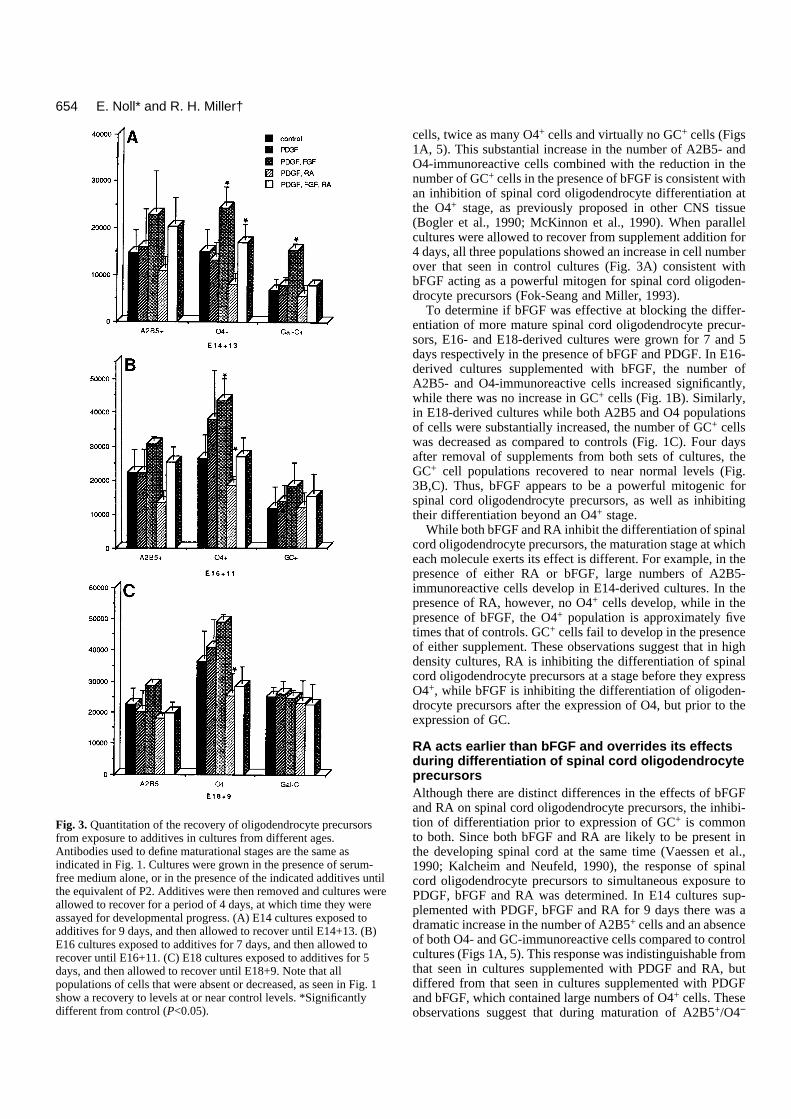
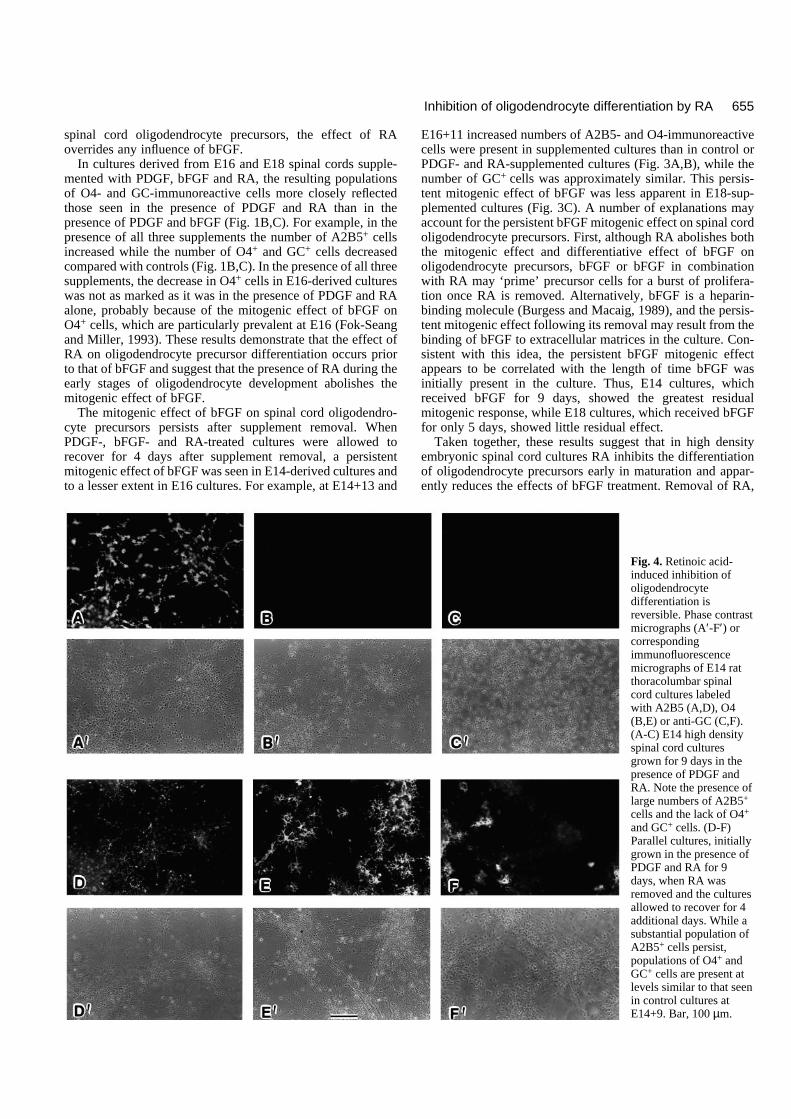
cultures retained the capacity to give rise to O4- and GC-immunoreactive cells, a recovery experiment was performed.Following labeling of the initial set of cultures at E14+9, allsupplements were removed from a parallel set of experimen-tal and control cultures by rinsing thoroughly and adding freshnon-supplemented medium. The cultures were then grown forfour additional days (until E14+13) and labeled with A2B5, O4and anti-GC antibodies. In control and previously PDGF-sup-plemented cultures, the number of A2B5-, O4- and GC-immunoreactive cells was approximately twice that seen atE14+9 (Fig. 3A), indicating both the normal differentiation andthe continued generation of oligodendrocyte precursors. Incultures previously supplemented with both PDGF and RA,however, there was a substantial increase in the number of O4-and GC-immunoreactive cells (Fig. 4) and a concomitantdecrease in the number of A2B5-immunoreactive cells (Fig.3A compare with Fig. 1A). These observations demonstratethat E14 spinal cord cultures grown in the presence of PDGFand RA for 9 days retain the capacity to give rise to oligoden-drocytes, even though such cells do not develop in the contin-uous presence of the supplements.
Two observations indicate that retinoic acid is the activefactor in the inhibition of oligodendrocyte development. NoO4- or CG-immunoreactive cells develop in E14 spinal cordcultures supplemented with only RA even when maintained inculture for 14 days (data not shown). In addition, spinal cordoligodendrocytes develop on schedule in the presence of PDGFalone (Warf et al., 1991 and see above). To eliminate the pos-sibility that the effects of RA, were mediated indirectly throughinhibiting the release of PDGF (a major mitogen for oligoden-drocyte precursors) from other cell types in the culture
652
(Richardson et al., 1988; Yeh et al., 1991), exogenous PDGFwas added to all but control experiments to ensure a saturatinglevel of mitogen.
The increase in A2B5+ cells observed in PDGF- and RA-supplemented medium at E14+9, along with the recovery ofboth O4+ and Gal-C+ populations in cultures previously treatedwith these supplements suggests that RA inhibits oligoden-drocyte differentiation at an early stage in the differentiationprogram, prior to the expression of O4 immunoreactivity. Itremained possible however, that either RA was selectively
toxic for O4+ and GC+ cells, or that it induced cellular dedif-ferentiation such that precursors that had matured beyond theA2B5+/O4− stage were forced to return to an A2B5+/O4− state.
To assess whether RA was either selectively toxic formaturing oligodendrocyte precursors or had the capacity toinduce dedifferentiation in these cells, spinal cord cultureswere established from thoracolumbar spinal cord of embryonicrats at E16 and E18. At E16, a small proportion of spinal cordoligodendrocyte precursors have matured to an O4+ stage, butno cells have yet acquired GC immunoreactivity (Warf et al.,1991). Thus, if RA is selectively toxic to, or inducing dedif-ferentiation in, O4+ cells there should be no O4+ or GC+ cellsin RA-supplemented E16 cultures. In E16 cultures after 7 daysin vitro (the equivalent of P1), both control and PDGF-sup-plemented cultures contained large numbers of A2B5-and O4-immunoreactive cells as well as GC+ cells (Fig. 1). Culturessupplemented with PDGF and RA also contained largenumbers of A2B5+ cells, however, the numbers of O4- andGC-immunoreactive cells were substantially reduced (Fig.1B). In contrast to the E14-derived cultures, in which no O4-or GC-immunoreactive cells developed in the presence of RA,E16-derived cultures contained cells of both populations (Fig.1B). In recovery experiments, following supplement removal,the number of O4+ and GC+ cells in the RA-supplementedcultures recovered to near control levels (Fig. 3B).
Because substantial populations of the O4+ cells and GC+
cells developed in the presence of RA in E16+7 cultures, RAis clearly neither selectively toxic to oligodendrocyte precur-sors which have matured to an O4-immunoreactive stage, noris it inducing de-differentiation of O4-immunoreactive cells.Instead, RA appears to be inhibiting the differentiation ofoligodendrocyte precursors at a pre-O4 stage.
To determine the effect of RA on GC+ cells, the differen-tiation of oligodendrocytes in the absence and presence of RA
E. Noll* and R. H. Miller†
Fig. 1. Quantitation of the initial effects of additives onoligodendrocyte differentiation. Graphs indicate the total numbers ofcells at each stage of differentiation. A2B5+ cells represent the mostimmature oligodendrocytes, O4+ cells represent oligodendrocytes atan intermediate stage of maturation and GC+ cells represent moremature oligodendrocytes. (A) E14 rat spinal cord cultures allowed todevelop to the equivalent of P1 (E14+9) in the presence of serum-free medium alone or with PDGF, PDGF and bFGF, PDGF and RA,or PDGF, bFGF and RA added daily. (B) E16 rat spinal cord culturesallowed to develop to the equivalent of P1 (E16+7) either undercontrol conditions or in the presence of the additives as above. (C)E18 cultures allowed to develop to the equivalent of P1 (E18+5).E14+9 cultures exposed to RA, with or without bFGF lack O4+ andGC+ cells, while cultures exposed to bFGF have a large O4+
population, but essentially no GC+ cells (A). The proportion ofoligodendrocyte precursors responsive to both RA and bFGF isdecreased in the cultures established from animals at either E16 (B)or E18 (C). Numbers represent the mean ± the standard deviation ofthe total number of immunoreactive cells derived from at least twoseparate coverslips in three different experiments in each case.*Significant difference from control (P<0.05). Note there is nosignificant difference in the number of O4+ cells between culturessupplemented with PDGF, FGF and RA and cultures supplementedwith PDGF and RA at all ages By contrast, E14 and E18 culturessupplemented with PDGF, FGF and RA contained significantlyfewer O4+ cells than those supplemented with PDGF and FGF.(P<0.05).
653Inhibition of oligodendrocyte differentiation by RA
was assayed in E18-derived spinal cord cultures. At E18 alarger proportion of oligodendrocyte precursors have maturedto an O4+ stage, and some have also begun to express GC. Ifthe effect of RA was predominantly on pre-O4+ cells the pro-portion of cells potentially able to disregard an inhibition ofdifferentiation by RA should be greater in culture establishedfrom E18 animals than in cultures established from E16animals. In both control and PDGF-supplemented E18cultures, large numbers of A2B5-, O4- and GC-immunoreac-tive cells were present after 5 days in vitro (Fig. 1C). In thepresence of PDGF and RA, however, although a similarnumber of A2B5-immunoreactive cells were present, thenumber of O4+ cells were reduced by approximately 50%compared with controls (Fig. 1C). and the number of GC+ cellswas also reduced. As anticipated if the effect of RA was mainlyon pre-O4+ cells, the number of O4- and GC-immunoreactivecells in the E18 RA-supplemented cultures was significantlylarger than that seen in the E16 RA-supplemented cultures(Compare Fig. 1B and C). In recovery experiments, 4 daysafter removal of the supplements, the number of O4- and GC-immunoreactive cells in E18-derived RA-supplementedcultures was similar to that seen in control and PDGF-supple-mented cultures, further demonstrating the reversible nature ofthe inhibition (Fig. 3C).
Taken together, these results indicate that RA is delaying thedifferentiation of spinal cord oligodendrocyte precursors.When exposed to RA early enough during their differentiation,oligodendrocyte precursor cells are prevented from maturingto an O4+ or subsequent GC+ stage. Cells that have already pro-gressed beyond the maturational stage characterized by theexpression of O4 immunoreactivity are neither selectivelyeliminated, nor induced to dedifferentiate in the presence ofRA.
FGF promotes proliferation and inhibitsdifferentiation of spinal cord oligodendrocyteprecursorsFor brain- and optic nerve-derived oligodendrocyte precursors,basic fibroblast-derived growth factor (bFGF) both promotescell proliferation and inhibits cellular differentiation(McKinnon et al., 1990; Bogler et al., 1990). To examine theeffect of bFGF on spinal cord oligodendrocyte precursor differ-entiation, E14 spinal cord cultures were supplemented withbFGF (10 ng/day), and PDGF (8 ng/day) for 9 days and thenassayed for A2B5-, O4- and GC-immunoreactive cells.
Compared to control and PDGF-supplemented cultures, E14spinal cord cultures grown in the presence of both PDGF andbFGF contained approximately three times as many A2B5+
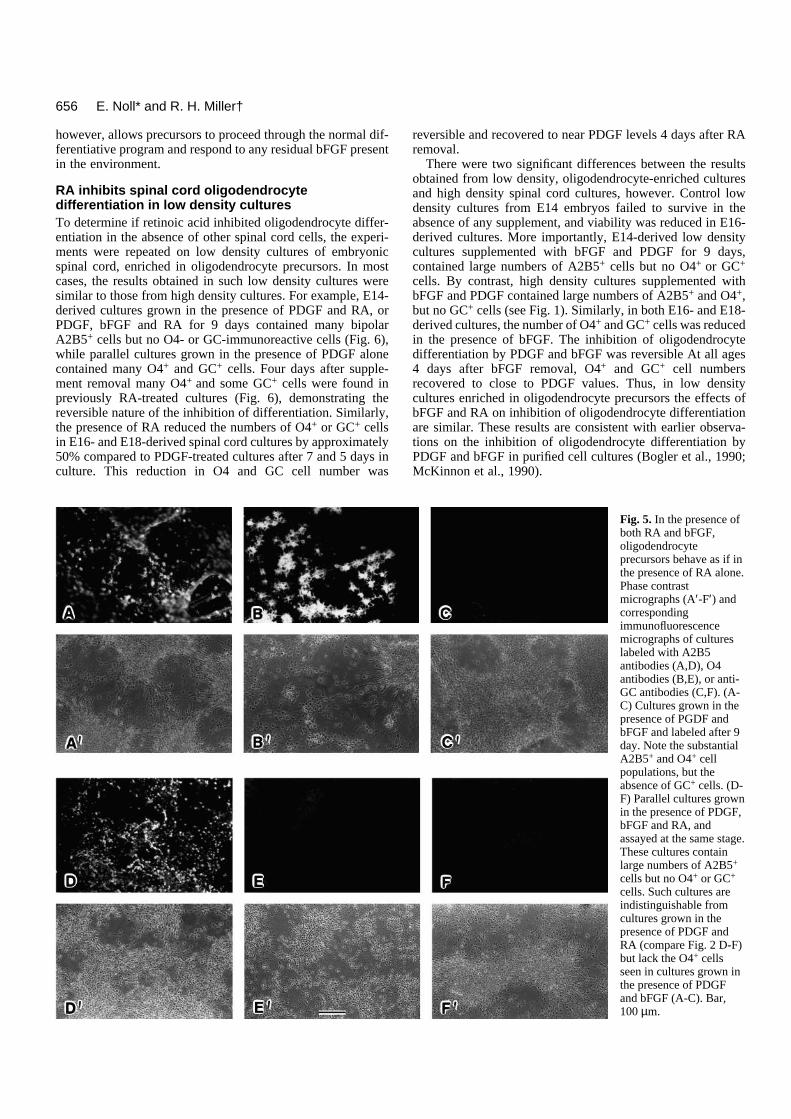
Fig. 2. Retinoic acidinhibits thedifferentiation of spinalcord oligodendrocyteprecursors at an A2B5-immunoreactive stage.Phase contrastmicrographs (A′-F′) andcorrespondingimmunofluorescencemicrographs of E14cultures labeled withA2B5 (A,D), O4 (B,E)or anti-GC (C,F) after 9days in culture. Highdensity culturesestablished from E14thoracolumbar rat spinalcords were grown undercontrol conditions withPDGF alone (A-C) or inthe presence of PDGFand RA (D-F) for aperiod of 9 days. In thepresence of PDGF,significant numbers ofA2B5-, O4- and GC-immunoreactive cellsdeveloped in allcultures. By contrast, inparallel cultures grownin the presence of PDGFand RA, there was adramatic increase in thenumber of A2B5+ cells(D), but no O4+ (E) orGC+ (F) cellsdeveloped. Bar, 100 µm.
654
cells, twice as many O4+ cells and virtually no GC+ cells (Figs1A, 5). This substantial increase in the number of A2B5- andO4-immunoreactive cells combined with the reduction in thenumber of GC+ cells in the presence of bFGF is consistent withan inhibition of spinal cord oligodendrocyte differentiation atthe O4+ stage, as previously proposed in other CNS tissue(Bogler et al., 1990; McKinnon et al., 1990). When parallelcultures were allowed to recover from supplement addition for4 days, all three populations showed an increase in cell numberover that seen in control cultures (Fig. 3A) consistent withbFGF acting as a powerful mitogen for spinal cord oligoden-drocyte precursors (Fok-Seang and Miller, 1993).
To determine if bFGF was effective at blocking the differ-entiation of more mature spinal cord oligodendrocyte precur-sors, E16- and E18-derived cultures were grown for 7 and 5days respectively in the presence of bFGF and PDGF. In E16-derived cultures supplemented with bFGF, the number ofA2B5- and O4-immunoreactive cells increased significantly,while there was no increase in GC+ cells (Fig. 1B). Similarly,in E18-derived cultures while both A2B5 and O4 populationsof cells were substantially increased, the number of GC+ cellswas decreased as compared to controls (Fig. 1C). Four daysafter removal of supplements from both sets of cultures, theGC+ cell populations recovered to near normal levels (Fig.3B,C). Thus, bFGF appears to be a powerful mitogenic forspinal cord oligodendrocyte precursors, as well as inhibitingtheir differentiation beyond an O4+ stage.
While both bFGF and RA inhibit the differentiation of spinalcord oligodendrocyte precursors, the maturation stage at whicheach molecule exerts its effect is different. For example, in thepresence of either RA or bFGF, large numbers of A2B5-immunoreactive cells develop in E14-derived cultures. In thepresence of RA, however, no O4+ cells develop, while in thepresence of bFGF, the O4+ population is approximately fivetimes that of controls. GC+ cells fail to develop in the presenceof either supplement. These observations suggest that in highdensity cultures, RA is inhibiting the differentiation of spinalcord oligodendrocyte precursors at a stage before they expressO4+, while bFGF is inhibiting the differentiation of oligoden-drocyte precursors after the expression of O4, but prior to theexpression of GC.
RA acts earlier than bFGF and overrides its effectsduring differentiation of spinal cord oligodendrocyteprecursorsAlthough there are distinct differences in the effects of bFGFand RA on spinal cord oligodendrocyte precursors, the inhibi-tion of differentiation prior to expression of GC+ is commonto both. Since both bFGF and RA are likely to be present inthe developing spinal cord at the same time (Vaessen et al.,1990; Kalcheim and Neufeld, 1990), the response of spinalcord oligodendrocyte precursors to simultaneous exposure toPDGF, bFGF and RA was determined. In E14 cultures sup-plemented with PDGF, bFGF and RA for 9 days there was adramatic increase in the number of A2B5+ cells and an absenceof both O4- and GC-immunoreactive cells compared to controlcultures (Figs 1A, 5). This response was indistinguishable fromthat seen in cultures supplemented with PDGF and RA, butdiffered from that seen in cultures supplemented with PDGFand bFGF, which contained large numbers of O4+ cells. Theseobservations suggest that during maturation of A2B5+/O4−
E. Noll* and R. H. Miller†
Fig. 3. Quantitation of the recovery of oligodendrocyte precursorsfrom exposure to additives in cultures from different ages.Antibodies used to define maturational stages are the same asindicated in Fig. 1. Cultures were grown in the presence of serum-free medium alone, or in the presence of the indicated additives untilthe equivalent of P2. Additives were then removed and cultures wereallowed to recover for a period of 4 days, at which time they wereassayed for developmental progress. (A) E14 cultures exposed toadditives for 9 days, and then allowed to recover until E14+13. (B)E16 cultures exposed to additives for 7 days, and then allowed torecover until E16+11. (C) E18 cultures exposed to additives for 5days, and then allowed to recover until E18+9. Note that allpopulations of cells that were absent or decreased, as seen in Fig. 1show a recovery to levels at or near control levels. *Significantlydifferent from control (P<0.05).
655Inhibition of oligodendrocyte differentiation by RA
spinal cord oligodendrocyte precursors, the effect of RAoverrides any influence of bFGF.
In cultures derived from E16 and E18 spinal cords supple-mented with PDGF, bFGF and RA, the resulting populationsof O4- and GC-immunoreactive cells more closely reflectedthose seen in the presence of PDGF and RA than in thepresence of PDGF and bFGF (Fig. 1B,C). For example, in thepresence of all three supplements the number of A2B5+ cellsincreased while the number of O4+ and GC+ cells decreasedcompared with controls (Fig. 1B,C). In the presence of all threesupplements, the decrease in O4+ cells in E16-derived cultureswas not as marked as it was in the presence of PDGF and RAalone, probably because of the mitogenic effect of bFGF onO4+ cells, which are particularly prevalent at E16 (Fok-Seangand Miller, 1993). These results demonstrate that the effect ofRA on oligodendrocyte precursor differentiation occurs priorto that of bFGF and suggest that the presence of RA during theearly stages of oligodendrocyte development abolishes themitogenic effect of bFGF.
The mitogenic effect of bFGF on spinal cord oligodendro-cyte precursors persists after supplement removal. WhenPDGF-, bFGF- and RA-treated cultures were allowed torecover for 4 days after supplement removal, a persistentmitogenic effect of bFGF was seen in E14-derived cultures andto a lesser extent in E16 cultures. For example, at E14+13 and
E16+11 increased numbers of A2B5- and O4-immunoreactivecells were present in supplemented cultures than in control orPDGF- and RA-supplemented cultures (Fig. 3A,B), while thenumber of GC+ cells was approximately similar. This persis-tent mitogenic effect of bFGF was less apparent in E18-sup-plemented cultures (Fig. 3C). A number of explanations mayaccount for the persistent bFGF mitogenic effect on spinal cordoligodendrocyte precursors. First, although RA abolishes boththe mitogenic effect and differentiative effect of bFGF onoligodendrocyte precursors, bFGF or bFGF in combinationwith RA may ‘prime’ precursor cells for a burst of prolifera-tion once RA is removed. Alternatively, bFGF is a heparin-binding molecule (Burgess and Macaig, 1989), and the persis-tent mitogenic effect following its removal may result from thebinding of bFGF to extracellular matrices in the culture. Con-sistent with this idea, the persistent bFGF mitogenic effectappears to be correlated with the length of time bFGF wasinitially present in the culture. Thus, E14 cultures, whichreceived bFGF for 9 days, showed the greatest residualmitogenic response, while E18 cultures, which received bFGFfor only 5 days, showed little residual effect.
Taken together, these results suggest that in high densityembryonic spinal cord cultures RA inhibits the differentiationof oligodendrocyte precursors early in maturation and appar-ently reduces the effects of bFGF treatment. Removal of RA,
Fig. 4. Retinoic acid-induced inhibition ofoligodendrocytedifferentiation isreversible. Phase contrastmicrographs (A′-F′) orcorrespondingimmunofluorescencemicrographs of E14 ratthoracolumbar spinalcord cultures labeledwith A2B5 (A,D), O4(B,E) or anti-GC (C,F).(A-C) E14 high densityspinal cord culturesgrown for 9 days in thepresence of PDGF andRA. Note the presence oflarge numbers of A2B5+
cells and the lack of O4+
and GC+ cells. (D-F)Parallel cultures, initiallygrown in the presence ofPDGF and RA for 9days, when RA wasremoved and the culturesallowed to recover for 4additional days. While asubstantial population ofA2B5+ cells persist,populations of O4+ andGC+ cells are present atlevels similar to that seenin control cultures atE14+9. Bar, 100 µm.
656
however, allows precursors to proceed through the normal dif-ferentiative program and respond to any residual bFGF presentin the environment.
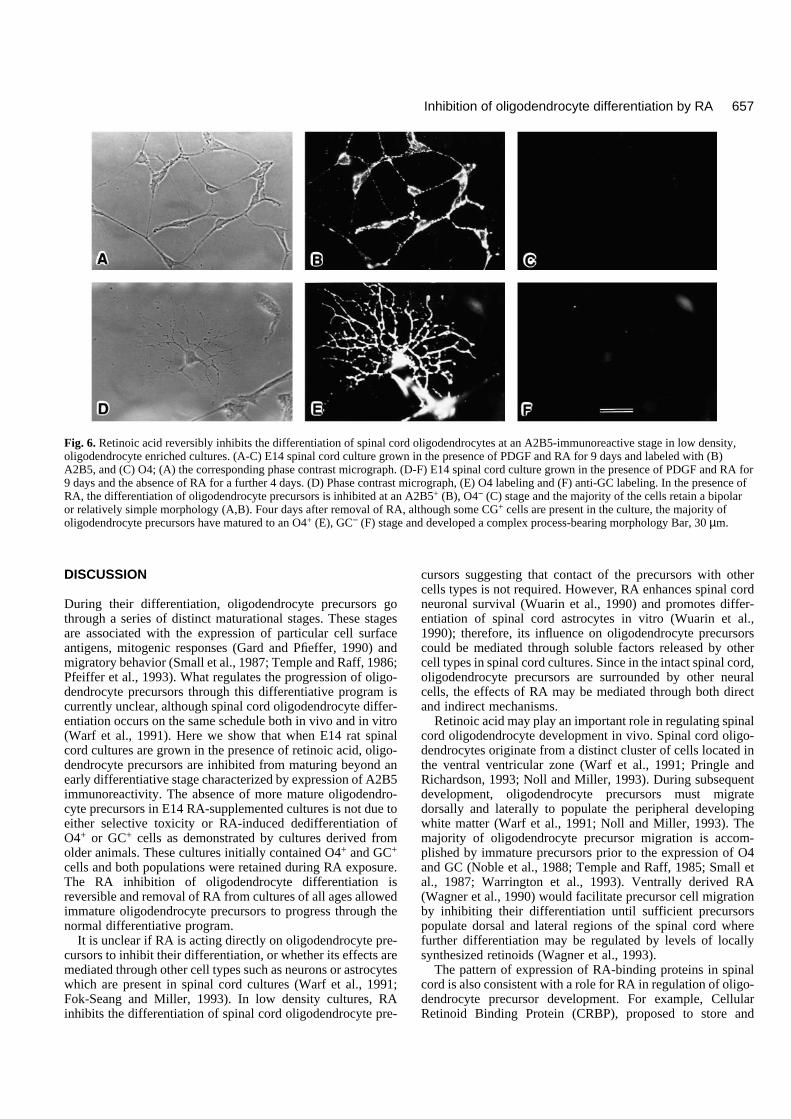
RA inhibits spinal cord oligodendrocytedifferentiation in low density culturesTo determine if retinoic acid inhibited oligodendrocyte differ-entiation in the absence of other spinal cord cells, the experi-ments were repeated on low density cultures of embryonicspinal cord, enriched in oligodendrocyte precursors. In mostcases, the results obtained in such low density cultures weresimilar to those from high density cultures. For example, E14-derived cultures grown in the presence of PDGF and RA, orPDGF, bFGF and RA for 9 days contained many bipolarA2B5+ cells but no O4- or GC-immunoreactive cells (Fig. 6),while parallel cultures grown in the presence of PDGF alonecontained many O4+ and GC+ cells. Four days after supple-ment removal many O4+ and some GC+ cells were found inpreviously RA-treated cultures (Fig. 6), demonstrating thereversible nature of the inhibition of differentiation. Similarly,the presence of RA reduced the numbers of O4+ or GC+ cellsin E16- and E18-derived spinal cord cultures by approximately50% compared to PDGF-treated cultures after 7 and 5 days inculture. This reduction in O4 and GC cell number was
reversible and recovered to near PDGF levels 4 days after RAremoval.
There were two significant differences between the resultsobtained from low density, oligodendrocyte-enriched culturesand high density spinal cord cultures, however. Control lowdensity cultures from E14 embryos failed to survive in theabsence of any supplement, and viability was reduced in E16-derived cultures. More importantly, E14-derived low densitycultures supplemented with bFGF and PDGF for 9 days,contained large numbers of A2B5+ cells but no O4+ or GC+
cells. By contrast, high density cultures supplemented withbFGF and PDGF contained large numbers of A2B5+ and O4+,but no GC+ cells (see Fig. 1). Similarly, in both E16- and E18-derived cultures, the number of O4+ and GC+ cells was reducedin the presence of bFGF. The inhibition of oligodendrocytedifferentiation by PDGF and bFGF was reversible At all ages4 days after bFGF removal, O4+ and GC+ cell numbersrecovered to close to PDGF values. Thus, in low densitycultures enriched in oligodendrocyte precursors the effects ofbFGF and RA on inhibition of oligodendrocyte differentiationare similar. These results are consistent with earlier observa-tions on the inhibition of oligodendrocyte differentiation byPDGF and bFGF in purified cell cultures (Bogler et al., 1990;McKinnon et al., 1990).
E. Noll* and R. H. Miller†
Fig. 5. In the presence ofboth RA and bFGF,oligodendrocyteprecursors behave as if inthe presence of RA alone.Phase contrastmicrographs (A′-F′) andcorrespondingimmunofluorescencemicrographs of cultureslabeled with A2B5antibodies (A,D), O4antibodies (B,E), or anti-GC antibodies (C,F). (A-C) Cultures grown in thepresence of PGDF andbFGF and labeled after 9day. Note the substantialA2B5+ and O4+ cellpopulations, but theabsence of GC+ cells. (D-F) Parallel cultures grownin the presence of PDGF,bFGF and RA, andassayed at the same stage.These cultures containlarge numbers of A2B5+
cells but no O4+ or GC+
cells. Such cultures areindistinguishable fromcultures grown in thepresence of PDGF andRA (compare Fig. 2 D-F)but lack the O4+ cellsseen in cultures grown inthe presence of PDGFand bFGF (A-C). Bar,100 µm.
657Inhibition of oligodendrocyte differentiation by RA
DISCUSSION
During their differentiation, oligodendrocyte precursors gothrough a series of distinct maturational stages. These stagesare associated with the expression of particular cell surfaceantigens, mitogenic responses (Gard and Pfieffer, 1990) andmigratory behavior (Small et al., 1987; Temple and Raff, 1986;Pfeiffer et al., 1993). What regulates the progression of oligo-dendrocyte precursors through this differentiative program iscurrently unclear, although spinal cord oligodendrocyte differ-entiation occurs on the same schedule both in vivo and in vitro(Warf et al., 1991). Here we show that when E14 rat spinalcord cultures are grown in the presence of retinoic acid, oligo-dendrocyte precursors are inhibited from maturing beyond anearly differentiative stage characterized by expression of A2B5immunoreactivity. The absence of more mature oligodendro-cyte precursors in E14 RA-supplemented cultures is not due toeither selective toxicity or RA-induced dedifferentiation ofO4+ or GC+ cells as demonstrated by cultures derived fromolder animals. These cultures initially contained O4+ and GC+
cells and both populations were retained during RA exposure.The RA inhibition of oligodendrocyte differentiation isreversible and removal of RA from cultures of all ages allowedimmature oligodendrocyte precursors to progress through thenormal differentiative program.
It is unclear if RA is acting directly on oligodendrocyte pre-cursors to inhibit their differentiation, or whether its effects aremediated through other cell types such as neurons or astrocyteswhich are present in spinal cord cultures (Warf et al., 1991;Fok-Seang and Miller, 1993). In low density cultures, RAinhibits the differentiation of spinal cord oligodendrocyte pre-
cursors suggesting that contact of the precursors with othercells types is not required. However, RA enhances spinal cordneuronal survival (Wuarin et al., 1990) and promotes differ-entiation of spinal cord astrocytes in vitro (Wuarin et al.,1990); therefore, its influence on oligodendrocyte precursorscould be mediated through soluble factors released by othercell types in spinal cord cultures. Since in the intact spinal cord,oligodendrocyte precursors are surrounded by other neuralcells, the effects of RA may be mediated through both directand indirect mechanisms.
Retinoic acid may play an important role in regulating spinalcord oligodendrocyte development in vivo. Spinal cord oligo-dendrocytes originate from a distinct cluster of cells located inthe ventral ventricular zone (Warf et al., 1991; Pringle andRichardson, 1993; Noll and Miller, 1993). During subsequentdevelopment, oligodendrocyte precursors must migratedorsally and laterally to populate the peripheral developingwhite matter (Warf et al., 1991; Noll and Miller, 1993). Themajority of oligodendrocyte precursor migration is accom-plished by immature precursors prior to the expression of O4and GC (Noble et al., 1988; Temple and Raff, 1985; Small etal., 1987; Warrington et al., 1993). Ventrally derived RA(Wagner et al., 1990) would facilitate precursor cell migrationby inhibiting their differentiation until sufficient precursorspopulate dorsal and lateral regions of the spinal cord wherefurther differentiation may be regulated by levels of locallysynthesized retinoids (Wagner et al., 1993).
The pattern of expression of RA-binding proteins in spinalcord is also consistent with a role for RA in regulation of oligo-dendrocyte precursor development. For example, CellularRetinoid Binding Protein (CRBP), proposed to store and
Fig. 6. Retinoic acid reversibly inhibits the differentiation of spinal cord oligodendrocytes at an A2B5-immunoreactive stage in low density,oligodendrocyte enriched cultures. (A-C) E14 spinal cord culture grown in the presence of PDGF and RA for 9 days and labeled with (B)A2B5, and (C) O4; (A) the corresponding phase contrast micrograph. (D-F) E14 spinal cord culture grown in the presence of PDGF and RA for9 days and the absence of RA for a further 4 days. (D) Phase contrast micrograph, (E) O4 labeling and (F) anti-GC labeling. In the presence ofRA, the differentiation of oligodendrocyte precursors is inhibited at an A2B5+ (B), O4− (C) stage and the majority of the cells retain a bipolaror relatively simple morphology (A,B). Four days after removal of RA, although some CG+ cells are present in the culture, the majority ofoligodendrocyte precursors have matured to an O4+ (E), GC− (F) stage and developed a complex process-bearing morphology Bar, 30 µm.

658
release RA where high levels need to be maintained (Ruberteet al., 1991) appears concentrated in ventral (Maden et al.,1990; Ruberte et al., 1993) and central regions of the spinalcord, along the dorsoventral axis (Dollé et al., 1990). This dis-tribution is coincident with one of the observed pathways ofoligodendrocyte precursor migration (Noll and Miller, 1993).Conversely, most spinal cord oligodendrocyte precursors dif-ferentiate into mature oligodendrocytes in the peripheral whitematter (Gilmore, 1971; Hirano and Goldman, 1988). In devel-oping white matter, RA concentrations would have to be main-tained at low levels in order to allow oligodendrocyte differ-entiation to occur. A second RA binding protein, cellularretinoic acid binding protein (CRABP) proposed to sequesterRA where low levels are required (Ruberte et al., 1991)appears concentrated in the developing white matter of theembryonic rat spinal cord (Vaessen et al., 1990) as well as inmouse developing white matter (Dencker et al., 1990; Madenet al., 1991). Thus, local mechanisms exist to spatially regulatethe levels of retinoids within the spinal cord, and the distribu-tion of RA binding proteins is consistent with a role for RA inregulation of oligodendrocyte differentiation.
Further support for a role of RA in oligodendrocyte devel-opment comes from studies on RA-deficient rats. Developingrats maintained on a vitamin A-deficient diet show a decreasein myelin content in the CNS (Bhat and Rao, 1978). Bio-chemical analysis of myelin isolated from these animalsindicated a relatively normal composition, although histologi-cal analysis showed areas of CNS hypomyelination (Bhat andRao, 1978). Such results may reflect the premature cessationof oligodendrocyte migration and premature differentiation,due to low RA concentrations resulting in decreased amountsof myelin which is normal in composition.
Retinoic acid is not the only regulator of spinal cord oligo-dendrocyte differentiation. In the presence of bFGF, oligoden-drocyte precursor differentiation is inhibited at the stage of O4expression, prior to expression of GC immunoreactivity in highdensity cultures. In low density cultures, however, bFGFinhibits oligodendrocyte precursor at an A2B5 stage prior tothe expression of O4. It is unclear why bFGF inhibits oligo-dendrocyte differentiation earlier in low density cultures. Onepossibility is that positive differentiation signals are providedby other spinal cord cells at high density and these override theearliest effects of bFGF allowing oligodendrocyte precursorsto mature to an O4+ stage. In addition to inhibiting differen-tiation, bFGF also appears to have a strong mitogenic effect onspinal cord oligodendrocyte precursors. It seems likely that theeffects of bFGF will be common to oligodendrocytes frommany regions of the CNS. For example, purified oligodendro-cyte precursors from brain and optic nerve are stimulated toproliferate in the presence of bFGF, and their differentiation isinhibited prior to the expression of GC immunoreactivity(Bogler et al., 1990). In such cultures, as in low density spinalcord cultures, combinations of PDGF and bFGF block differ-entiation prior to O4 expression (Collarini et al., 1992; Pfiefferet al., 1993) directly paralleling the effect of RA in high densityembryonic spinal cord cultures.
Why the differentiation of oligodendrocyte precursors in thespinal cord should be regulated by both RA and bFGF iscurrently unclear. Since the proliferation of oligodendrocyteprecursors is especially protracted during spinal cord develop-ment, different regulators of oligodendrocyte differentiation
may be required during the early and late stages of develop-ment. Labeling of mitotic spinal cord neural precursors withBrdU indicates that oligodendrocyte precursors are rapidlyproliferating at E16.5 (Noll and Miller, 1993). Retroviralmediated gene transfer studies suggest they may be proliferat-ing as early as E12 (Zhang and Miller, unpublished observa-tion). Previous studies have documented the continued prolif-eration of spinal cord oligodendrocyte precursors during thefirst and second postnatal week (Gilmore, 1971), that is somethree weeks after their initial period of proliferation. Since bothbFGF (Enfors et al., 1990; Kalcheim and Neufeld, 1990) andFGF receptors (Josef et al., 1990; Wanaka et al., 1991) arepresent during much of spinal cord development, the differen-tiation of spinal cord oligodendrocyte precursors in the earlyembryonic spinal cord could be regulated by a combination ofRA and bFGF while during later embryonic and postnataldevelopment the differentiation of oligodendrocyte precursorscould be regulated primarily by bFGF levels.
The inhibition of spinal cord oligodendrocyte differentiationby RA is likely to be transient. RA levels appear relatively highin early embryonic spinal cord, but as development proceedsthey decrease (Wagner et al., 1993), possibly below the levelsrequired for the inhibition of oligodendrocyte differentiation.Furthermore, the notochord, one of the proposed ventralsources of RA in the early embryonic animal is a transientstructure present only during early embryological stages. Thisconcept of multiple regulators of cell proliferation and differ-entiation is not restricted to development of the CNS. In thehematopoietic system, for example, within a single cell lineagedifferent cytokines influence cell proliferation at differentstages of development (Metcalf, 1989).
In conclusion, we propose that retinoic acid plays a primaryrole in the regulation of differentiation of oligodendrocyte pre-cursors in the early embryonic spinal cord. The RA-inducedinhibition of oligodendrocyte differentiation allows immatureoligodendrocyte precursors to migrate from their point oforigin in the ventral ventricular zone to populate the develop-ing white matter throughout the entire spinal cord.
We thank Drs A. Hall, H. Zhang and J. Fok-Seang for helpful dis-cussions on this work. This study was supported in part by Grant NIH-25597 and the National Multiple Sclerosis Society.
REFERENCES
Abney, E., Bartlett, P. F. and Raff, M. C. (1981). Astrocytes, ependymal cellsand oligodendrocytes develop on schedule in dissociated cell cultures ofembryonic brain. Dev. Biol. 83, 301-310.
Bansal, R., Warrington, A. E., Gard, A. L., Ranscht, B. and Pfeiffer, S. E.(1989). Multiple and novel specificities of monoclonal antibodies O1, O4,and R-mAb used in the analysis of oligodendrocyte development. J.Neurosci. Res. 24, 548-557.
Bhat, P. V. and Rama Rao, P. B. (1978). Vitamin A nutrition in relation togangliosides and myelination in the developing brain. Wld. Rev. Nutr. Diet.31, 100-106.
Bogler, O., Wren, D., Barnett, S., Land, H. and Noble, M. (1990).Cooperation between growth factors promotes extended self-renewal andinhibits differentiation of oligodendrocyte-type-2 astrocyte (0-2A)progenitor cells. Proc. Natl. Acad. Sci. USA 87, 6368-6372.
Bottenstein, J. E. and Sato, G. H. (1979). Growth of a rat neuroblastoma cellline in serum-free supplemented medium. Proc. Natl. Acad. Sci. USA 76,514-517.
E. Noll* and R. H. Miller†

659Inhibition of oligodendrocyte differentiation by RA
Burgess, W. H. and Macaig, T. (1989). The heparin-binding (fibroblast)growth factor family of proteins. Annu. Rev. Biochem. 58, 575-606.
Collarini, E. J., Kuhn, R., Marshall, C. J., Monuki, E. S., Lemke, G. andRichardson, W. D. (1992). Down regulation of the POU transcription factorSCIP is an early event in oligodendrocyte differentiation in vitro.Development 116, 193-200.
Curtis, R., Cohen, J., Fok-Seang, J., Hanley, M. R., Gregson, N. A.,Reynolds, R. and Wilkin, G. P. (1988). Development of macroglial cells inrat cerebellum 1. Use of antibodies to follow early in vivo development andmigration of oligodendrocytes. J. Neurocytol. 17, 43-54.
Dencker, L., Annerwall, E., Busch, E. and Eriksson, U. (1990). Localizationof specific retinoid-binding sites and expression of cellular retinoic acidbinding protein (CRABP) in the early mouse embryo. Development 110,343-352.
Dollé, P., Ruberte, E., Leroy, P., Morriss-Kay, G. and Chambon, P. (1990).Retinoic acid receptors and celluar retinoid binding proteins I. A systematicstudy of their differential pattern of transcription during mouseorganogenesis. Development 110, 1133-1151.
Durston, A. J., Timmermans, P. M., Hage, W. J., Hendriks, H. F. J.,deVries, N. J., Heideveld, M. and Nieuwkoop, P. D. (1989). Retinoic acidcauses an anteroposterior tranformation in the developing central nervoussystem. Nature 340, 140141.
Eisenbarth, G. S., Walsh, F. S. and Nirenberg, M. (1979). Monoclonalantibody to a plasma membrane antigen antigen of neurons. Proc. Natl.Acad. Sci.USA 76, 4913-4917.
Ernfors, P., Lonnerberg, P., Ayer-LeLievre, C. and Persson, H. (1990).Developmental and regional expression of basic fibroblast growth factormRNA in the rat central nervous system. J. Neurosci. Res. 27, 10-15.
Fredman, P., Magnani, J. L., Nirenberg, M. and Ginsburg, V. (1984).Monoclonal antibody A2B5 reacts with many gangliosides in neuronaltissue. Arch. Biochem. Biophys. 33, 661-666.
Fok-Seang, J. and Miller, R. H. (1993). Distribution and differentiation ofA2B5+ glial precursors in the developing rat spinal cord. J. Neurosci. Res.(In Press).
Gard, A. L. and Pfeiffer, S. E. (1990). Two proliferative stages of theoligodendrocyte lineage (A2B5+O4- and O4+Gal-C-) under differentmitogenic control. Neuron 5, 615-625.
Gilmore, S. A. (1971). Neuroglial population in the spinal white matter ofneonatal and early postnatl rats: an autographic study of numbers ofneuroglia and changes in their proliferative acitivity. Anat. Res. 171, 383-292.
Hardy, R. and Reynolds, R. (1991). Proliferation and differentiation potentialof rat forebrain oligodendroglial progenitors both in vivo and in vitro.Development 111, 1061-1080.
Hirano, M and Goldman, J. E. (1988). Gliogenesis in rat spinal cord:evidence for origin of astrocytes and oligodendrocytes from radialprecursors. J. Neurosci. Res. 21, 155-167.
Hughes, S. M., Lillien, L. E., Raff, M. C., Roher, H. and Sendtner, M.(1988). Ciliary neurotrophic factor induces type-2 astrocyte differentiation inculture. Nature 335, 70-73.
Jessell, T. M. and Dodd, J. (1992). Floor plate-derived signal and the controlof neural cell pattern in vertebrates. Harvey Lect. 86, 87-127.
Josef, G., Heuer, C., von Bartheld, S., Kinoshita, Y., Evers, P. C. andBothwell, M. (1990). Alternating phases of FGF receptor and NGF receptorexpression in the developing chicken nervous system. Neuron 5, 283-296.
Kalcheim, C. and Neufeld, G. (1990). Expression of basic fibroblast growthfactor in the nervous system of early avian embryos. Development 109, 203-215.
La Chapelle, F., Gumple, M., Baluac, M., Jacque, C., Due, P. andBaumann, N. (1984). Transplantation of CNS fragments into the brain ofshiverer mutant mice: Extensive myelination by implantedoligodendrocytes. 1. Immunological studies. Dev. Neurosci. 6, 325-334.
Lillien, L. E., Sendtner, M. and Raff, M. C. (1990). Extracellular matrix-associated molecules collaborate with ciliary neurotrophic factor to inducetype-2 astrocyte development. J. Cell Biol. 111, 635-644.
Maden, M., Hunt, P., Eriksson, U., Kuroiwa, A., Krumlauf, R. andSummerbell, D. (1991). Retinoic acid-binding protein, rhombomeres andthe neural crest. Development 111, 35-44.
McKinnon, R. D., Matsui, T., Dubois-Dalque, M. and Aaronson, S. (1990).FGF modulates the PDGF-driven pathway of oligodendrocyte development.Neuron 5, 603-614.
Mendelsohn, C., Ruberte, E, LeMeur, M., Morriss-Kay, G. and Chambon,P. (1991). Developmental analysis of the retinoic acid-inducible RAR-B2promoter in transgenic animals. Development 113, 723-735.
Metcalf, D. (1989). The molecular control of cell division, differentiationcommitment and maturation in haemopoietic cells. Nature 339, 27-30.
Noble, M., Murray, K., Stroobant, P., Waterfield, M. D. and Riddle, P.(1988). Platelet derived growth factor promotes division and motility andinhibits premature differentation of the oligodendrocyte/type-2 astrocyteprogenitor cell. Nature 333, 560-562.
Noll, E. and Miller, R. H. (1993). Oligodenrocyte precursors originate at theventral ventricular zone dorsal to the ventral midline in the embryonic ratspinal cord. Development 118, 563-573.
Peters, A., Palay, S. L. and Webster, H. deF. (1990). The Fine Structure of theNervous System. 3rd edition. New York: Oxford University Press.
Pfieffer, S. E., Warrington, A. E. and Bansal, R. (1993). The oligodendrocyteand its many processes. Trends in Cell Biol. 3, 191-197.
Pringle, N. and Richardson, W. D. (1993). A singularity of PDGF alpha-receptor expression in the dorsoventral axis of the neural tube may define theorigin of the oligodendrocyte lineage. Development 117, 525-533.
Raff, M. C. (1989). Glial cell diversification in the rat optic nerve. Science 243,1450-1455.
Raff, M. C., Lillien, L. E., Richardson, W. D., Burne, J. F. and Noble, M. D.(1988). Platelet-derived growth factor from astrocytes drives the clock thattimes oligodendrocyte development in culture. Nature 333, 562-565.
Raff, M. C., Mirsky, R., Fields, K. L., Lisak, R. P., Dorfman, S. H.,Silberberg, D. H., Gregson, N. A., Liebowitz, S. and Kennedy, M. C.(1978). Galactocerebroside is a specific cell surface antigenic marker foroligodenrocytes in culture. Nature 274, 813-816.
Ranscht, B., Clapshaw, P. A., Price, J., Noble, M. and Seifert, W. (1982).Development of oligodendrocytes and Schwann cells studied with amonoclonal antibody against galactocerebroside. Proc. Natl. Acad. Sci. 79,2709-2713.
Richardson, W. D., Pringle, N., Mosley, M. J. Westermark, B. and Dubois-Dalcq, M. (1988). A role for platelet-derived growth factor in normalgliogenesis in the central nervous system. Cell 53, 309-319.
Ruberte, E., Friederich, V., Chambon, P. and Morriss-Kay, G. (1993).Retinoic acid receptors and cellular retinoid binding proteins. 111 Theirdifferential transcript distribution during mouse nervous systemdevelopment. Development 118, 267-282.
Ruberte, E., Dollé, P., Chambon, P. and Morriss-Kay, G. (1991). Retinoicacid receptors and cellular retinoid binding proteins II. Their differentialpattern of transcription during early morphogenesis in mouse embryos.Development 111, 45-60.
Ruiz i Altaba, A. and Jessell, T. M. (1991). Retinioc acid modifies the patternof cell differentiation in the central nervous system of neurula stage Xenopusembryos. Development 112, 945-958.
Skoff, R., Price, D. and Stocks, A. (1976a). Electron microscopticautoradiographic studies of gliogenesis in rat optic nerve. I. Cellproliferation. J. Comp. Neurol. 169, 291-312.
Skoff, R., Price, D. and Stocks, A. (1976b). Electron microscopticautoradiographic studies of gliogenesis in rat optic nerve II. Time of origin. J.Comp. Neurol. 196, 313-333.
Small, R. K., Riddle, P. and Noble, M. (1987). Evidence for migration ofoligodendrocyte-type-2 astrocyte progenitor cells into the developing ratoptic nerve. Nature 328, 155-157.
Sommer, I. and Schachner, M. (1981). Monoclonal antibodies (O1 to O4) tooligodendrocyte cell surfaces: an immunocytological study in the centralnervous system. Dev. Biol. 83, 311-327.
Temple, S. and Raff, M. C. (1985). Differentiation of a bipotential glialprogenitor cell in single cell microwell. Nature 313, 223-225.
Temple, S. and Raff, M. C. (1986). Clonal analysis of oligodendrocytedevelopment in culture. Evidence for a developmental clock that counts celldivisions. Cell 44, 773-779.
Tessier-Lavigne, M., Placzek, M., Lumsden, A. G. S., Dodd, J. and Jessell,T. M. (1988). Chemotropic guidance of developing axons in the mammaliannervous sytem. Nature 336, 775-778.
Van Straaten, H. W. M., Hekking, J. W. M., Beursgens, J. P. W. M.,Terwindt-Rouwenhorst, E. and Drukker, J. (1989). Effect of thenotochord on proliferation and differentiation in the neural tube of the chickembryo. Development 107, 793-803.
Vaessen, M., Meijers, J. H. C., Bootsma, D. and Van Kessel, A. G. (1990).The cellular retinoic acid-binding protein is expressed in tissues associatedwith retinoic acid induced malformations. Development 110, 371-378.
Wagner, M., Thaller, C., Jessell, T. and Eichele, G. (1990). Polarizingactivity and retinoid synthesis in the floor plate of the neural tube. Nature345, 819-822.
Wagner, M., Han, B. and Jessell, T. M. (1993). Regional differences in

660
retinoid release from embryonic neural tissue detected by an in vitro reporterassay. Development 116, 55-66.
Wanaka, A., Milbrandt, J. and Johnson, E. M. (1991). Expression of FGFreceptor gene in rat development. Development 111, 455-468.
Warf, B. C., Fok-Seang, J. and Miller, R. H. (1991). Evidence for the ventralorigin of oligodendrocytes in the rat spinal cord. J. Neurosci. 11, 2477-2488.
Warrington, A. E, Barbarese, E. and Pfieffer, S. E. (1993). Differentialmyelinogenic capacity of specific stages of the oligodendrocyte lineage upontransplantation into hypomyelinating hosts. J. Neurosci. Res. 34, 1-13.
Williams, B. P., Abney, E. R. and Raff, M. C. (1985). Macroglial celldevelopment in embyonic rat brain: Studies using monoclonal antibodies,fluorecence activated cell sorting and cell culture. Dev. Biol. 112, 126-134.
Wuarin, L., Sidell, N. and DeVellis, J. (1990). Retinoids increase perinatalspinal cord neuronal survival and astroglial differentiation. Int. J. Devl.Neuroscience 8:3, 317-326.
Yamada, T., Placzek, M., Tanaka, H., Dodd, J. and Jessell, T. M. (1991).Control of cell pattern in the developing nervous system: polarizing activityof the floor plate and notochord. Cell 64, 635-647.
Yeh, H. J., Ruit, K. G., Wangh, Y., Parks, W. C., Snider, W. D. and Deuel,T. F. (1991). PDGF-A chain is expressed by mammalian neurons duringdevelopment and in maturity. Cell 64, 209-216.
(Accepted 1 December 1993)
E. Noll* and R. H. Miller†





























