Embed Size (px)
Citation preview

Regulation of gene Expression in Prokaryotes & Eukaryotes
1
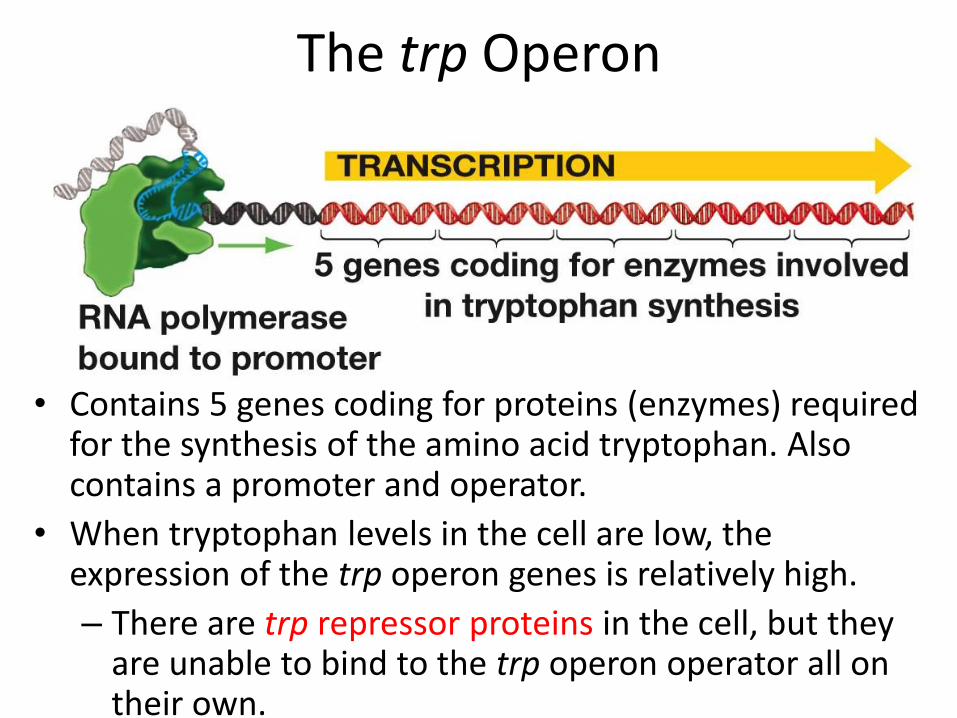
The trp Operon
• Contains 5 genes coding for proteins (enzymes) required for the synthesis of the amino acid tryptophan. Also contains a promoter and operator.
• When tryptophan levels in the cell are low, the expression of the trp operon genes is relatively high.
– There are trp repressor proteins in the cell, but they are unable to bind to the trp operon operator all on their own.

The trp Operon
• When tryptophan levels in the cell are high: – Tryptophan binds to the repressor and activates it. – The activated trp repressor can now bind to the trp
operator and block transcription of the trp operon genes.

The lac and trp
Operons

Positive Control of Transcription
• Occurs when a regulatory protein (an activator) binds to DNA and increases the rate of transcription of downstream genes.
• Let’s look at two examples of activator proteins at work: – AraC proteins can increase transcription of the ara operon genes.
– CAP proteins can increase transcription of the lac operon genes.

Positive Control of the ara Operon
• E. coli can also utilize arabinose, a pentose found in plant cell walls, as an energy source.
• The ara operon includes:
– 3 genes required for arabinose metabolism.
– A promoter to which RNA polymerase can bind.
– An initiator sequence to which an activator protein (AraC) can bind to stimulate transcription.

Positive Control of the ara Operon
• Expression of the ara operon genes is high only when arabinose levels are high:
– (If you were E. coli, you wouldn’t want to waste precious resources expressing these genes unless there was arabinose around to break down!)
– How does the presence of arabinose stimulate expression of the ara operon genes?
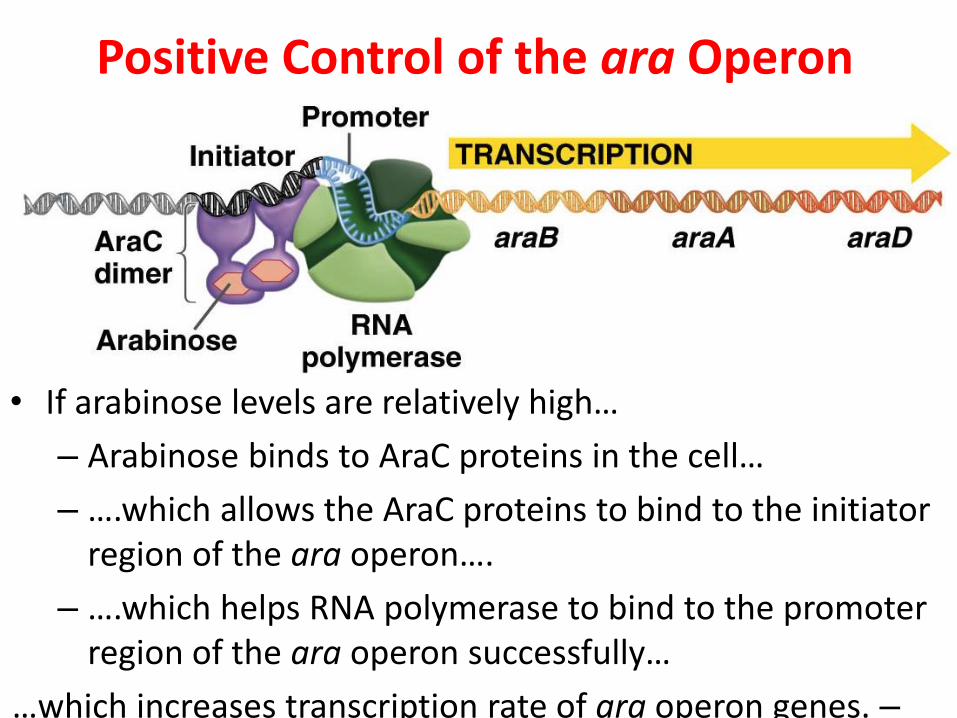
Positive Control of the ara Operon
• If arabinose levels are relatively high…
– Arabinose binds to AraC proteins in the cell…
– ….which allows the AraC proteins to bind to the initiator region of the ara operon….
– ….which helps RNA polymerase to bind to the promoter region of the ara operon successfully…
–…which increases transcription rate of ara operon genes.

Positive Control of the ara Operon
• Thus, AraC proteins, when bound to arabinose, can act as activators of gene expression at the ara operon:
– Example of positive control of gene expression.
• Let’s look at one more example of positive control of gene expression, this time going back to the lac operon.
– Gene expression at the lac operon is under both: • Negative control (by the lacI repressor protein)
• Positive control (by a protein called CAP)

10

• Eukaryotic cells have a much larger genome
• Eukaryotes have much greater cell specialization
• Thus eukaryotic cells contain an enormous amount of DNA that does not program the synthesis of RNA or protein
• This requires complex organization
• In eukaryotes expression of gene into proteins can be controlled at various locations
3/9/2017 11

Control of gene Expression
• Mammalian cells possess about 1000 times more genetic information than does the bacterium Escherichia coli.
• Much of this additional genetic information is probably involved in regulation of gene expression during the differentiation of tissues and biologic processes in the multicellular organism and in ensuring that the organism can respond to complex environmental challenges.
12

• Synthesis of proteins is controlled right from the chromatin stage.
• Expression of gene is controlled at many steps during the process of transcription and translation.
1. Transcriptional control. 2. RNA processing control. 3. RNA transport & localisation control. 4. Translation control. 5. mRNA degradation control. 6. Protein activator control.
CHECK POINTS FOR GENE EXPRESSION IN EUKARYOTES

Two forms of chromatin
• Euchromatin – A lesser coiled, less methylation transcriptionally active region which can be easily accessed by the RNA polymerases.
• Heterochromatin – Large regions highly condensed methylated transcriptionally inactive region. The genes in this region cannot be accessed by the RNA polymerases for active transcription.
14
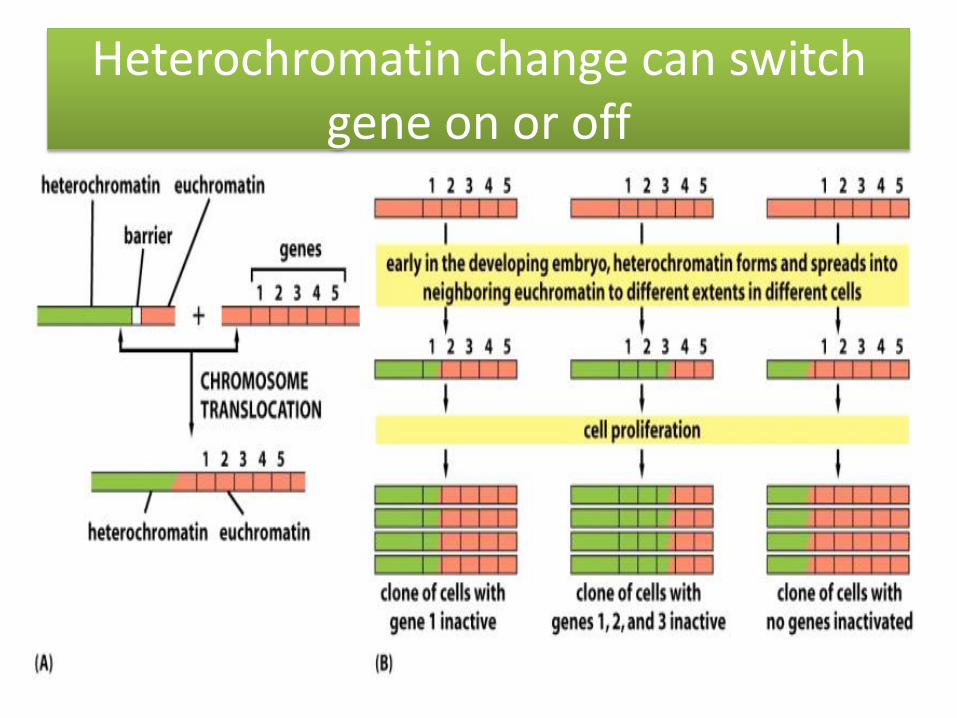
Heterochromatin change can switch gene on or off
16

Mechanism of regulation of gene expression- Details
1) Chromatin Remodeling • Chromatin structure provides an important level of
control of gene transcription. • With few exceptions, each cell contains the same
complement of genes (antibody-producing cells are a notable exception).
• The development of specialized organs, tissues, and cells and their function in the intact organism depend upon the differential expression of genes.
• Some of this differential expression is achieved by having different regions of chromatin available for transcription in cells from various tissues.
17

1) Chromatin Remodeling
• Large regions of chromatin are transcriptionally inactive in some cells while they are either active or potentially active in other specialized cells
• For example, the DNA containing the -globin gene cluster is in "active" chromatin in the reticulocytes but in "inactive" chromatin in muscle cells.
18

Formation and disruption of
nucleosome structure
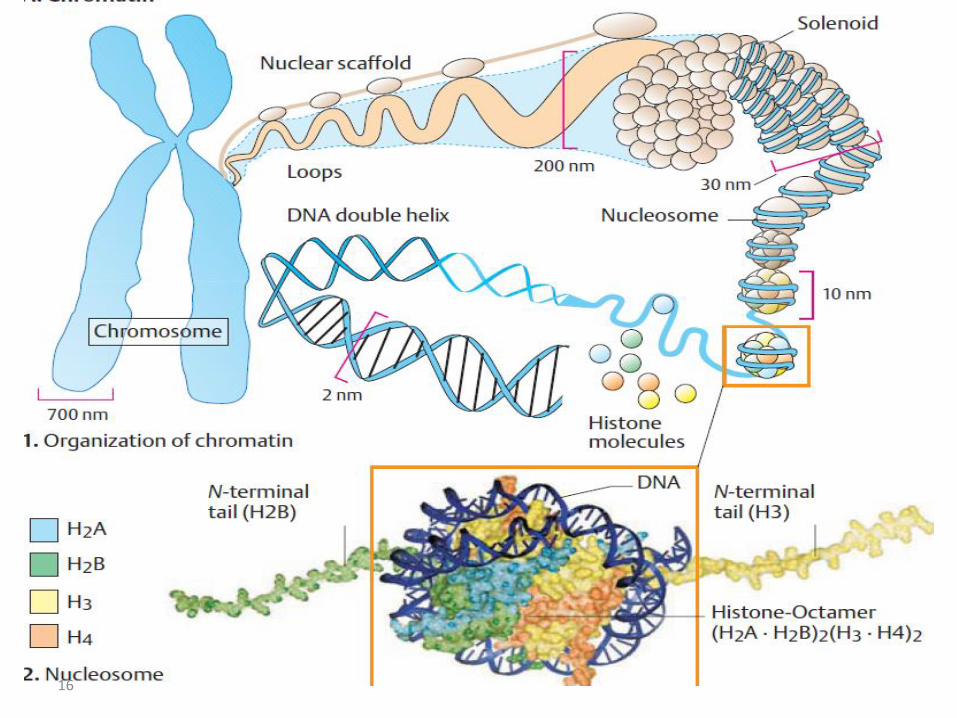
• The presence of nucleosomes and of complexes of histones and DNA provide a barrier against the ready association of transcription factors with specific DNA regions.
• The disruption of nucleosome structure is therefore an important part of eukaryotic gene regulation and the processes involved are as follows :
19

Formation and disruption of nucleosome structure (contd.)
A) Histone acetylation and deacetylation
Acetylation is known to occur on lysine residues in the amino terminal tails of histone molecules.
This modification reduces the positive charge of these tails and decreases the binding affinity of histone for the negatively charged DNA.
Accordingly, the acetylation of histones could result in disruption of nucleosomal structure and allow readier access of transcription factors of the same regulatory DNA elements.
20
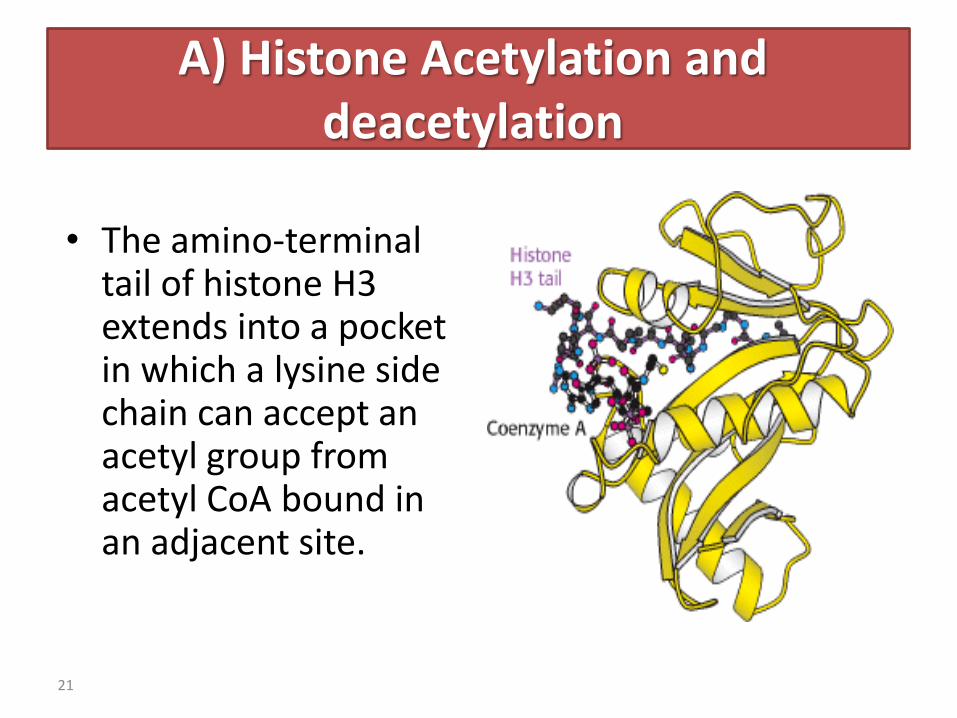
A) Histone Acetylation and deacetylation
• The amino-terminal tail of histone H3 extends into a pocket in which a lysine side chain can accept an acetyl group from acetyl CoA bound in an adjacent site.
21

Histone Acetylation Acetylation of the lysine makes RNA polymerase and transcription factors easier to access the promoter region. Therefore, in most cases, histone acetylation enhances transcription while histone deacetylation represses transcription. Histone acetylation is catalyzed by histone acetyltransferases (HATs) and histone deacetylation is catalyzed by histone deacetylases (denoted by HDs or HDACs).
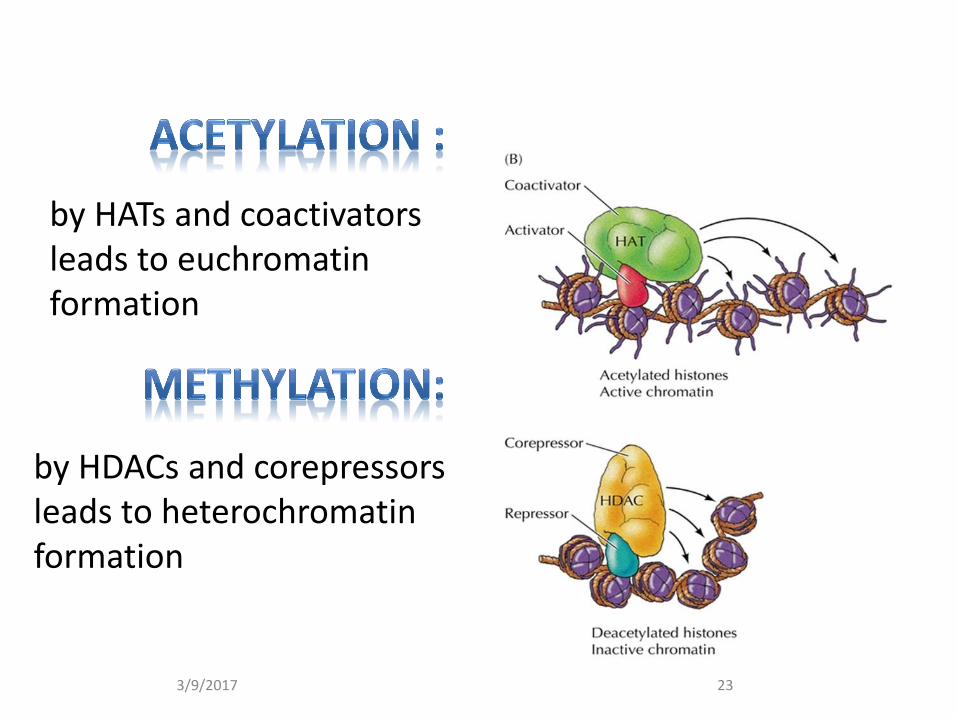
by HDACs and corepressors leads to heterochromatin formation
by HATs and coactivators leads to euchromatin formation
3/9/2017 23
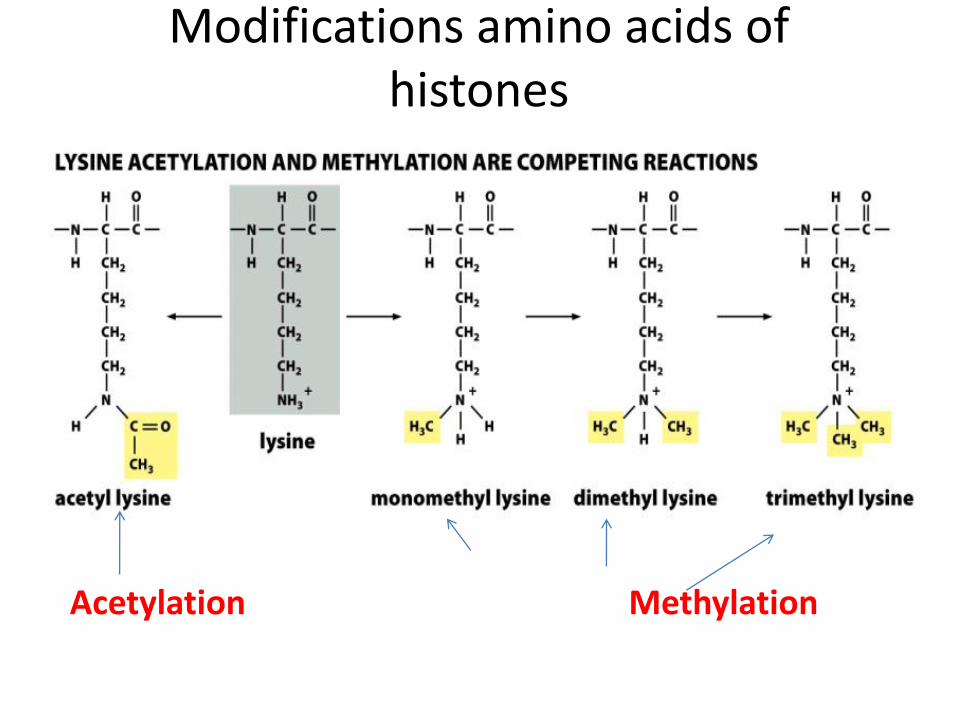
Modifications amino acids of histones
Acetylation Methylation

Methylation of deoxycytidine residues in DNA may effect gross changes in chromatin so as to prevent its active transcription.
Acute demethylation of deoxycytidine residues in a specific region of the tyrosine aminotransferase gene—in response to glucocorticoid hormones—has been associated with an increased rate of transcription of the gene.
25
B) Modification of DNA

C) DNA Binding Proteins
The binding of specific transcription factors to certain DNA elements may result in disruption of nucleosomal structure. Many eukaryotic genes have multiple protein-
binding DNA elements. The serial binding of transcription factors to these
elements may either directly disrupt the structure of the nucleosome or prevent its re-formation. These reactions result in chromatin-level structural
changes that in the end increase DNA accessibility to other factors and the transcription machinery.
26

• Histone modifications – These modifications make a region of gene either transcriptionally active or inactive.
Acetylation
• ↑Acetylation ↓ Condensation of DNA ↑ Transcription of genes in that region
Ubiquitination Ubiquitination of H2A – Transcriptional inactivation
Ubiquitination of H2B - Transcriptional activation
3/9/2017 27

DNA methylation: is the addition or removal of a methyl group predominately where cytosine bases occur consecutively. Methylation occurs most often in symmetrical CG sequences.
3/9/2017 28
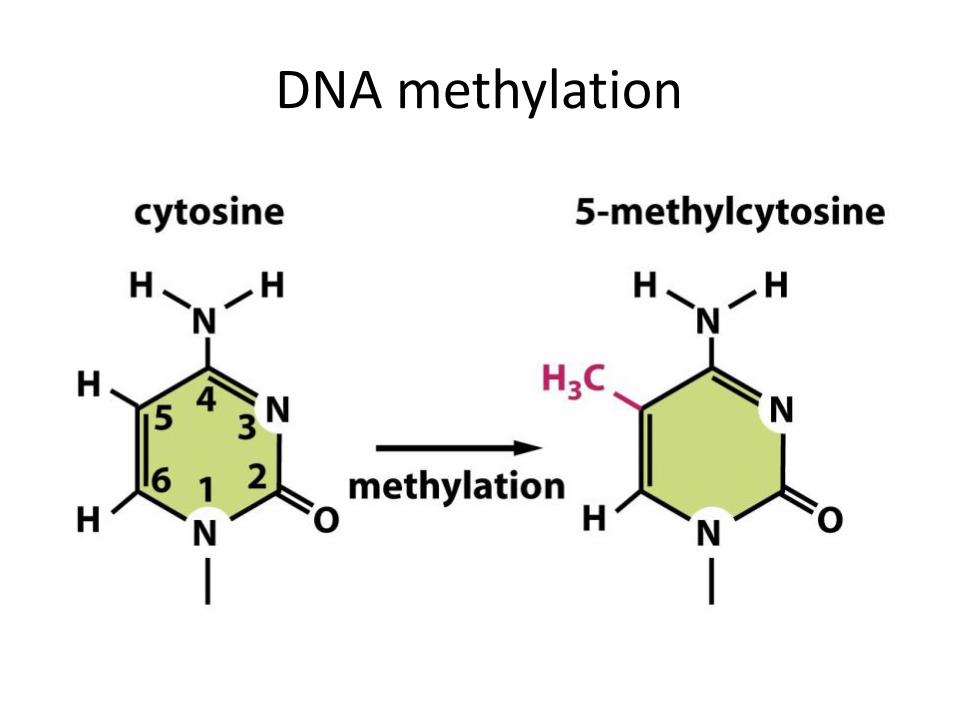
DNA methylation
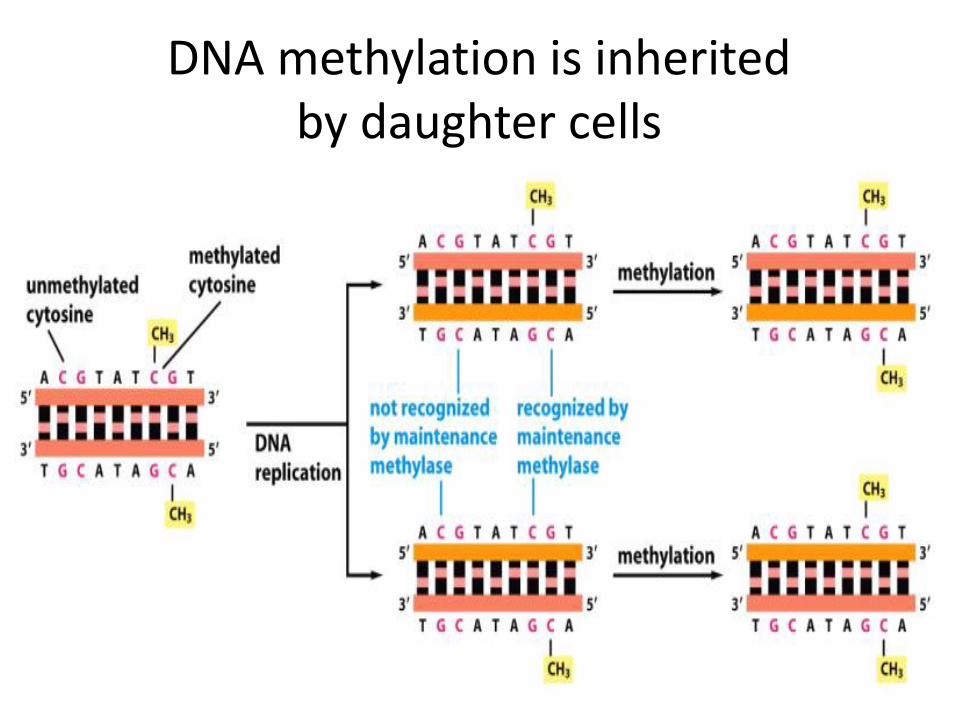
DNA methylation is inherited by daughter cells

DNA methylation in gene control region inactivates gene expression
CpG Island: DNA region with many CG sequences, of which C can become methylated. In mammals, 70% to 80% of CpG cytosines are methylated.

Effects of DNA methylation
- Inactivation of a single gene expression
- Inactivation of genes from part of chromosome
- Inactivation of genes of whole chromosome (X)
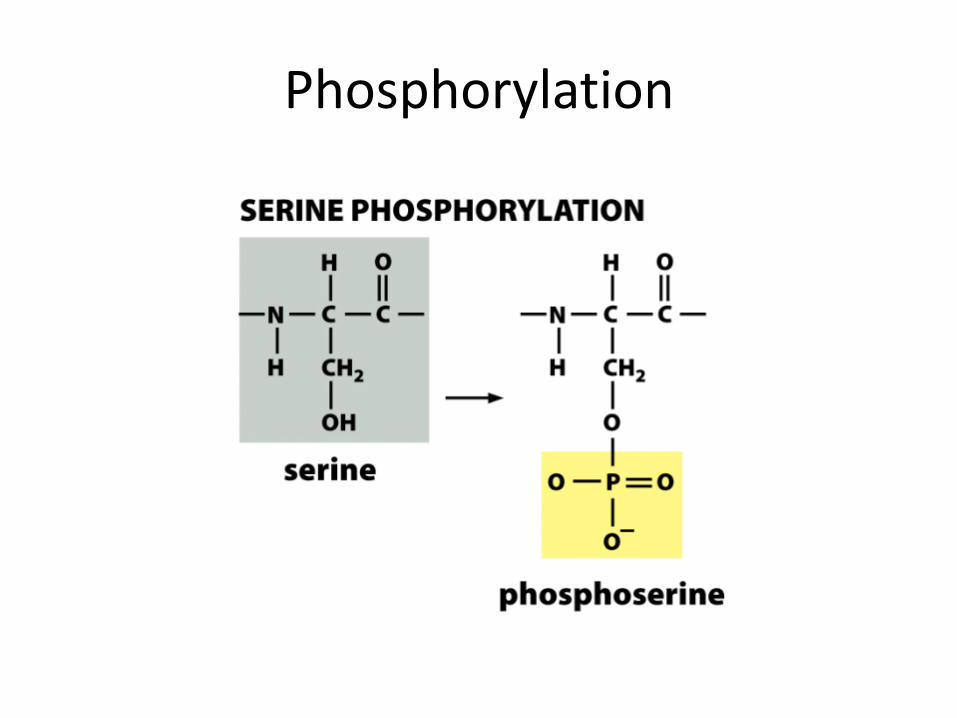
Phosphorylation

Most important histone modifications
• Acetylation
• Methylation
• Phosphorylation
• Ubiquitination
For all these modifications, special enzymes are needed!

Histone-modification sites

Effects of histone-modifications on gene expression (histone H3)

Humans have Hox genes in four clusters:
Genes
Chromosome Cluster
HOXA1, HOXA2, HOXA3, HOXA4, HOXA5, HOXA6, HOXA7, HOXA9, HOXA10, HOXA11, HOXA13
chromosome 7 HOXA
HOXB1, HOXB2, HOXB3, HOXB4, HOXB5, HOXB6, HOXB7, HOXB8, HOXB9, HOXB13
chromosome 17 HOXB
HOXC4, HOXC5, HOXC6, HOXC8, HOXC9, HOXC10, HOXC11, HOXC12, HOXC13
chromosome 12 HOXC
HOXD1, HOXD3, HOXD4, HOXD8, HOXD9, HOXD10, HOXD11, HOXD12, HOXD13
chromosome 2 HOXD

Epigenetics ( above genetics)
Science studying heritable changes in gene expression, not resulting from changes in nucleotide sequence of DNA
• Reversible modifications of chromatin organization
- DNA
- Histones
• mRNA modifications
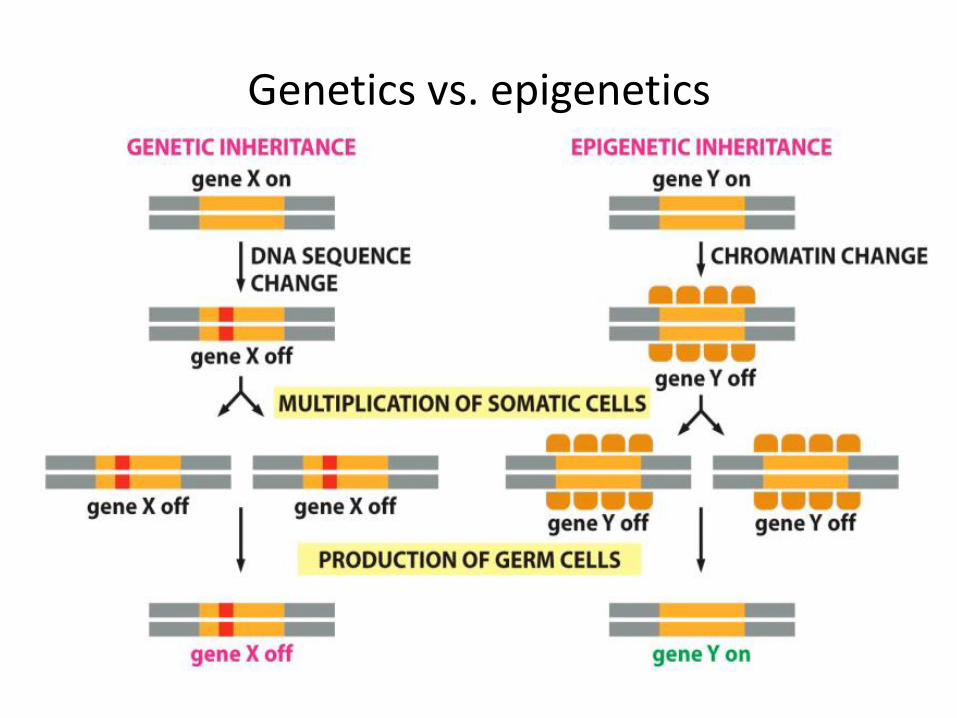
Genetics vs. epigenetics

• END Part II





























