Embed Size (px)
Citation preview

Proc. Nati. Acad. Sci. USAVol. 91, pp. 1219-1223, February 1994Biochemistry
Regulation of metallothionein genes by heavy metals appears to bemediated by a zinc-sensitive inhibitor that interacts with aconstitutively active transcription factor, MTF-1RICHARD D. PALMITERHoward Hughes Medical Institute and Department of Biochemistry, SL-15, University of Washington, Seattle, WA 98195
Contributed by Richard Palmiter, November 4, 1993
ABSTRACT A construct, MRE-PGeo, with five metalresponse elements fused to a selectable reporter gene wastransfected into BHK cells and a stable clone that could beinduced up to 100-fold by zinc, cadmium, bismuth, silver,cobalt, copper, mercury, or nickle was isolated. Some, andperhaps all, of these metals induce MRE-,Geo by displacingzinc. Transfection of these cells with a construct encoding thetranscriptional activator MTF-1 resulted in constitutive ex-pression of MRE-PGeo, whereas expression of an antisenseMTF-1 construct in these cells prevented induction by all of themetals. A variant cell line with high conktitutive expression inthe absence of added metals was isolated; normal regulationwas restored by cell fusion. These results suggest that regula-tion of metallothionein genes by metals is mediated by MTF-1interacting with metal response elements and that zinc func-tions to release MTF-1 from an inhibitor.
Metallothionein (MT) genes have been described for a widevariety of organisms ranging from yeast to man. These genesencode small, cysteine-rich, metal-binding proteins that pro-vide protection against metal toxicity and may play a role inhomeostasis of essential metals (1, 2). There is alto mountingevidence that they may help protect against reactive oxygen(3). MT gene expression is ugually inducible. For example,the mammalian MT-I and MT-II genes, which are expressedin most cells, are induced by a variety of metals, hoimones,and xenobiotics (1, 2, 4, 5).Most MT genes that have been studied are inducible by
metals. Induction of mammalian MT genes by metals ismediated by short DNA elements called metal responseelements (MREs) that are present in multiple copies in thepromoter region of these genes (5). For example, the mouseMT-I gene has five functional MREs within the 150 bpflanking the transcription start site, but they are not allfunctionally equivalent (6, 7). Genomic footprinting of nucleifrom induced and uninduced cells indicated that a factor isbound to the MREs only during induction, suggesting thatmetals allow a positively acting transcription factor to inter-act with the MREs (8, 9). Proteins have been detected innuclear extracts of mammalian cells that bind to radiolabeledMREs, but several different sizes for these proteins havebeen reported and the effects of zinc, cadmium, and copperon binding activity were variable; thus, the number of dif-ferent factors capable of binding MREs is unclear (7, 10-15).Recently, a clone encoding a MRE-binding protein wasisolated from a cDNA expression library by using labeledMREs as a probe (16). This protein, called MTF-1, has sixzinc fingers of the 2 Cys-2 His type and resembles transcrip-tion factors with this motif (17). It can be isolated in an activeform from uninduced cells. When the cDNA for this factorwas put under control ofa strong promoter and cotransfected
into cells along with a MRE-driven reporter gene, it resultedin constitutive expression of the reporter gene in the absenceof added metal inducer (16). Thus, MTF-1 is a candidatetranscription factor for interaction with MREs, but it is notclear how it is regulated by metals. The induction of mam-malian MTs by metals is compounded by the observation thata wide variety of metals are good inducers, including somemetals that are not bound by MT (18). Hence, it is unclearwhether all of these metals stimulate the same traiscriptionfactor and, if not, whether different metal-responsive tran-scription factors recognize different cis-acting elements inMT gene promoters. The cloning of MTF-1 (16) provides astarting point for trying to unravel the regulation of mamma-lian MT genes.
MATERIALS AND METHODSPlasmids. MRE-(3Geo. A BamHI-HindIII fragment con-
taining five MRE-d' elements (6) upstream of the basal mouseMT-I promoter (-42 to +60) was introduced between the NotI and Hind]Il sites of /Geo, a f-galactosidase-neomycinphosphotransferase (lacZ-neo) fusion gene (19). CMV-MTF-1. Two pairs of complementary oligonucleotides (25-mers with 6 bp of overlap) were synthesized with sequencescorresponding to the 5' and 3' ends of the published MTF-1clone (16). Each oligonucleotide pair was filled in by usingDNA polymerase and [a-32P]dNTPs and then combined toscreen a A ZAP mouse brain cDNA library (Stratagene). Fiveof nine clones that were plaque-purified hybridized to boththe 5' and 3' probes; the longest one extends 18 bp further 5'than the published sequence (16). The Nco I fragment thatincludes the entire MTF-I open reading frame was insertedbetween a cytomegalovirus (CMV) promoter/enhancer and asimian virus 40 (SV40) polyadenylylation region of a cDNAexpression plasmid. Then, a Not I fragment containing theCMV-MTF-1-SV40 cassette was introduced into pNUT, aplasmid vector with a dihydrofolate reductase gene driven bythe SV40 promoter/enhancer (20). CMV-MTF(AZF). Theregions encoding the zinc fingers were removed from theconstruct described above by restriction with ApaLI, whichcuts conveniently at the borders of the six fingers, and thereading frame was restored with an oligonucleotide linker.CMV-aMTF-1. When the 2.1-kb Nco I fragment containingthe MTF-1 sequence was inserted into the CMV vector, bothorientations were obtained. The antisense orientation wasmoved into pNUT as described above.
Cells and Transfection. BHK cells (thymidine kinase-negative, MT-) were grown in Dulbecco's modified Eagle'smedium (DMEM) (Life Technologies, Gaithersburg, MD)plus 10o fetal bovine serum unless otherwise indicated. Fortransfection, a calcium phosphate precipitate with 20 ug ofplasmid DNA was applied to cells that were about 25%
Abbreviations: MT, metallothionein; MRE, metal response element;CMV, cytomegalovirus; SV40, simian virus 40; PDC, pyrrolidinedithiocarbonate.
1219
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.
Proc. Natl. Acad. Sci. USA 91 (1994)
confluent on a 10-cm plate in 10 ml of medium along with 100,uM chloroquine (21). The medium was changed 4 hr later.Cells were split the next day and selection with G418 (800pg/ml) and ZnSO4 (80 pM) or methotrexate (200 pM) wasbegun. Individual colonies were picked with cloning ringsabout 2 weeks later and then expanded in selection mediumfor about a month. For some experiments, cells were grownin a specially formulated Opti-MEM (Life Technologies) thatwas prepared without zinc, Opti-MEMAZn. This mediumcontains <0.1 MM zinc compared with 1.0 pLM in normalOpti-MEM. DMEM with 10%1 serum contains 3.8 pM zinc.Metal concentrations were measured by inductively coupledplasma emission spectroscopy.The MRE-,BGeo clone 3038-1-1 was mutagenized six times
with ICR-191 (5-10 pg/ml for 2-4 hr) as described byMcKendry et aL (22). Then the cells were selected with G418at 3.2 mg/ml in the absence of zinc. Clones were picked 2weeks later and tested individually for induction of MRE-(3Geo by zinc. One of the subclones (3286-8-8) was fused toa hygromycin-resistant clone of BHK cells by plating equalnumbers of the two cell types and treating them with 50%1polyethylene glycol for 1 min (23) and then selecting forhybrids with G418 (800 pg/ml), hygromycin (800 pg/ml), andZnSO4 (80 PM).
3-Galactosidase Assay. Cells were trypsinized and plated in24-well dishes 1-2 days before addition of inducers. Stocksolutions of metals (CdSO4, ZnSO4, AgNO3, CuSO4, HgCl2,bismuth ammonium citrate, CoCl2, NiCl2) or metal chelatorswere diluted about 100-fold into the tissue culture medium.About 20 hr later the cells were fixed for 5 min in 0.2%glutaraldehyde and 2% formaldehyde in phosphate-bufferedsaline at 4°C and then rinsed three times in phosphate-buffered saline. One milliliter of 4 mM o-nitrophenyl ,B-ga-lactopyranoside (Sigma) in 25 mM sodium phosphate, pH7.7/2mM MgCl2 was added to the fixed cells. The plates wereincubated at 23°C for 1-2 hr, and the absorbance at 405 nmwas measured. The number of cells was determined bysuspending unfixed cells in 0.5 ml 0.2% SDS, sonicatingbriefly, and measuring the amount of DNA by the fluores-cence of Hoechst 33258 with a TKO-100 fluorometer (Hoe-fer).RNA Analysis. Total nucleic acids were harvested by the
SDS-proteinase K method (24), followed by phenol/chloroform extraction and ethanol precipitation. The sampleswere dissolved and digested briefly with RNase-free DNase.The RNA was denatured, electrophoresed through a 1.4%agarose gel containing 2.2 M formaldehyde, and then trans-ferred to nylon membranes as described (25). The membraneswere prehybridized, hybridized with nick-translated lacZ ora-actin probes, and then washed as described (26) except thatall washes were at 68°C.
RESULTSZinc Induction of MRE-(Geo. Five MRE-d' elements were
fused to a basal promoter driving 3Geo, a selectable reportergene, and transfected into BHK cells, and stable clones wereselected in medium containing G418 and ZnSO4. Severalclones were isolated and one of them, 3038-1-1, which wasinduced about 100-fold by zinc, was chosen for furtheranalysis. Southern analysis indicated a single copy of MRE-(3Geo. Half-maximal induction of -galactosidase activity byzinc was obtained in 6 hr, and maximal activity was reachedin 20 hr. Upon removal of zinc, (3-galactosidase activity fellby halfwithin 10 hr and reached basal levels by 30 hr. For theexperiments described below, cells were split into 24-welldishes so that they would be about 75%M confluent 1-2 dayslater, at which time inducers were added. Cells were assayedfor (3-galactosidase activity about 20 hr later.
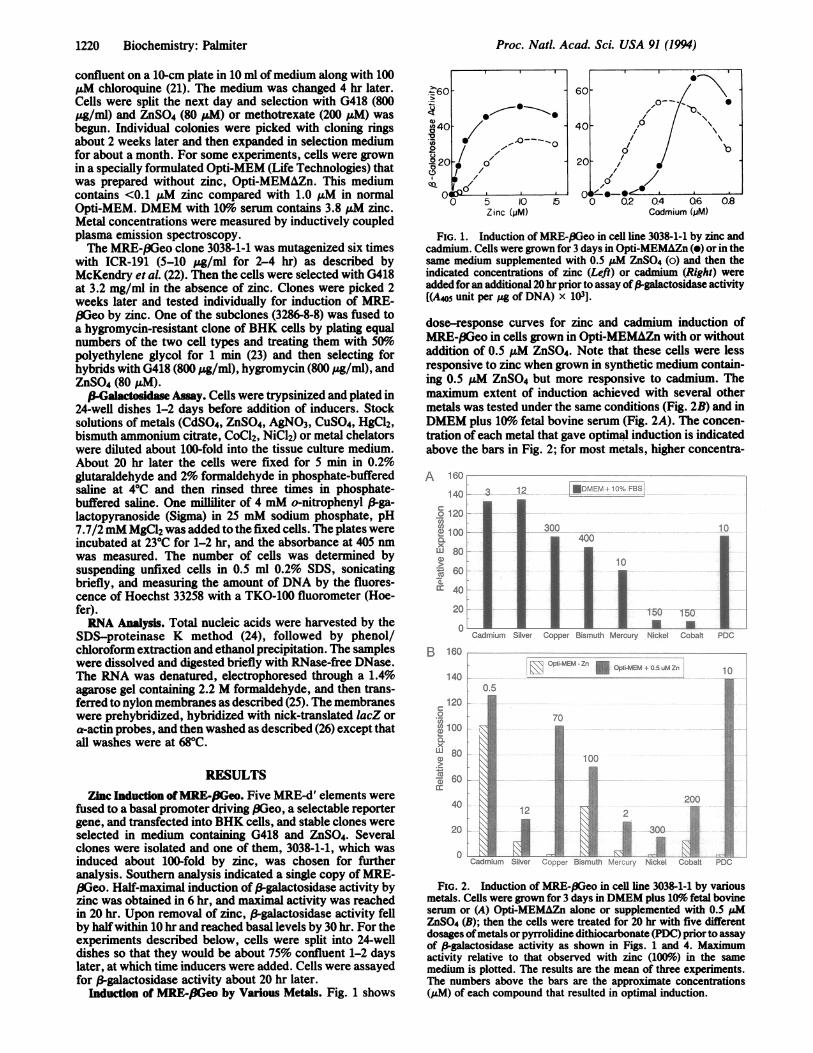
Induction of MRE-f3Geo by Various Metals. Fig. 1 shows
4~~60 60-10t_-. 0
(440-40 0
/O0 C(/-'0
0 5 10 15 0 0.2 0.4 Q6 0.8Zinc(/M) Cadmium (pM)
FIG. 1. Induction of MIRE-,8Geo in cell line 3038-1-1 by zinc andcadmium. Cells were grown for 3 days in Opti-MEMAZn (e) or in thesame medium supplemented with 0.5 AM ZnS04 (o) and then theindicated concentrations of zinc (Left) or cadmium (Right) wereadded for an additional 20 hr prior to assay of/-galactosidase activity[(A405 unit per pg of DNA) x 103].
dose-response curves for zinc and cadmium induction ofM[EE-.BGeo in cells grown in Opti-MEMAZn with or withoutaddition of 0.5 pM ZnS04. Note that these cells were lessresponsive to zinc when grown in synthetic medium contain-ing 0.5 pM ZnS04 but more responsive to cadmium. Themaximum extent of induction achieved with several othermetals was tested under the same conditions (Fig. 2B) and inDMEM plus 10%o fetal bovine serum (Fig. 2A). The concen-tration of each metal that gave optimal induction is indicatedabove the bars in Fig. 2; for most metals, higher concentra-
A 160--140 3
o120co
2? 1001a
xLU 80L
560:a. '
20
0onCadmium
12 fDMlENP-- C - S
300400
1Silver Copper Bismuth Mercury
10
150 150
lt..Nickel Cobalt PDC
160 _-
140Kr\R OPTi-MEM Zn " Opti-MEM 0 5 uM Zn140
0.5
100
x
0) ~~~~~~10060
0 3200.,1t1,1.C.;12
0Cadmium Silver ,;j-ep Bismuth i r5Nickel Cobalt
10
.!.,
l.DiiiC[ii ;.:
eli
'i-,i
PDC
FIG. 2. Induction of MRE-.8Geo in cell line 3038-1-1 by variousmetals. Cells were grown for 3 days in DMEM plus 10%/ fetal bovineserum or (A) Opti-MEMAZn alone or supplemented with 0.5 ,uMZnSO4 (B); then the cells were treated for 20 hr with five differentdosages ofmetals or pyrrolidine dithiocarbonate (PDC) prior to assayof ,-galactosidase activity as shown in Figs. 1 and 4. Maximumactivity relative to that observed with zinc (100%) in the samemedium is plotted. The results are the mean of three experiments.The numbers above the bars are the approximate concentrations(uM) of each compound that resulted in optimal induction.
-
1220 Biochemistry: Palmiter

Proc. Natl. Acad. Sci. USA 91 (1994) 1221
tions were required in the presence of serum. The BHK cellsused for these studies do not synthesize MT, which increasestheir sensitivity to some metals. All of the metals tested gavesome induction in the medium containing serum (Fig. 2A),but only zinc, cadmium, bismuth, and silver were effective inOpti-MEMAZn. However, when 0.5 pM ZnSO4 was added tothe synthetic medium, induction by copper, mercury, andnickel was also observed (Fig. 2B). Hence, all of these metalsact via transcription factors that recognize MRE-d' and someof them probably induce expression by displacing zinc fromother components in the medium. PDC is also an effectiveinducer ofMRE-I3Geo (Fig. 2). Since PDC is a metal chelator,it probably acts by increasing the transport of metals intocells. Note that PDC did not induce MRE-,BGeo in thesynthetic medium unless 0.5 ,uM ZnSO4 was added (Fig. 2B).
Constitutive Expression of MRE-fIGeo by MTF-1. Thecoding region of mouse MTF-1 cDNA was cloned next to theCMV promoter/enhancer in an expression vector that alsocarries a dihydrofolate reductase gene which facilitates se-lection of stable clones with high numbers of transgenes (20).This construct was introduced into the 3038-1-1 cell line andfive stable clones were selected in the presence of 200 juMmethotrexate. The induction ofMRE-.BGeo by zinc in each ofthese clones was compared with that in the original 3038-1-1clone. All clones responded similarly (Fig. 3). The basalactivity in the absence of zinc was higher than that observedin the original 3038-1-1 clone when treated with 100 A&MZnSO4. MRE-/3Geo expression in all the MTF-1 clones wasinduced <1.3-fold by added zinc. These results confirm that
MRE-flGeo MREs IacZ neo bGHpA
AdditionalConstruct
none
Clone Uninduced MaximumActivity Zn-induced
Activity
3038-1-1 1 100
CMVp_-_ MTF-1 -SVpACMV-MTF-I -- ---
3256B-23256B-33256B-6
172 218200 241156 178
CMV-aMTrF- ZCMVp anti-MTF-1 <)SVpACMVaMTF- -a---4-- -4*-.
2.1 kb (NN) 3256C-53256C-63256C-9
0.69 kb (SB) 3269-3
1 133 221 9
0 13
CMVp _ MTFROF SVpACMV-MTF.AZF AZF
3283-POP 2 157
FIG. 3. Effect of CMV-driven MTF-1, antisense MTF-I, andMTF(AZF) on expression of MRE-pGeo. The various constructs(see Materials and Methods) were transfected into the 3038-1-1 BHKcell line carrying MRE-,3Geo, and stable clones or populations (POP)of cells resistant to 200 AuM methotrexate were selected. A zincdose-response experiment was performed as in Fig. 4 and themaximal P-galactosidase activity achieved relative to the parentalclone is indicated. bGHpA, bovine growth hormone polyadenylyla-tion signal; CMVp, CMV promoter; SVpA, SV40 polyadenylylationsignal; NN, Nco I-Nco I; SB, Sma I-BamHI.
expression of excess MTF-1 leads to constitutive expressionof MRE-driven reporter genes (16).To assess how much MTF-1 mRNA was produced in these
cells, RNA was isolated for Northern analysis and probedwith MTF-1 cDNA. The hybridization signal achieved with60 ng ofRNA from these transfected cells was comparable tothat from 20 pg of control BHK cells. Zinc treatment did notaffect the amount of endogenous MTF-I mRNA or thatderived from the CMV-MTF-1 construct. The amount ofMTF-1 mRNA detected in mouse liver, kidney, and spleenwas comparable to that in BHK cells; however, there wasabout 50-fold more MTF-1 mRNA in testis (data not shown).
Antisense MTF-1 RNA Blocks Expression of MRE-PGeo.An antisense construct, CMV-aMTF-1, was made with the2.1-kb Nco I fragment and tested in a manner analogous tothat described above. Eight methotrexate-resistant cloneswere tested. There was variability among the clones in theextent of inhibition, but in the three best clones induction of(3-galactosidase activity was only 9-22% of control cells at allzinc concentrations tested (Fig. 3). An antisense constructwas also made with a 690-bp fiagment from the 5' end of thecDNA. Two of the six clones tested inhibited expressionabout 9-fold (Fig. 3). These results suggest that mouseantisense MTF-1 RNA can effectively block the synthesis ofhamster transcription factor(s) that mediate zinc responsive-ness. To ascertain whether all the metals that induce MRE-,SGeo act via MTF-1, clone 3256C-5 was tested with each ofthe metals at their optimal concentration in either the richmedium or Opti-MEMAZn. Induction by all metals wasinhibited about 10-fold in either medium, suggesting thatMTF-1 may be the only factor that binds to MREs (data notshown).MTF-1 May Be Regulated by an Inhibitory Factor. Because
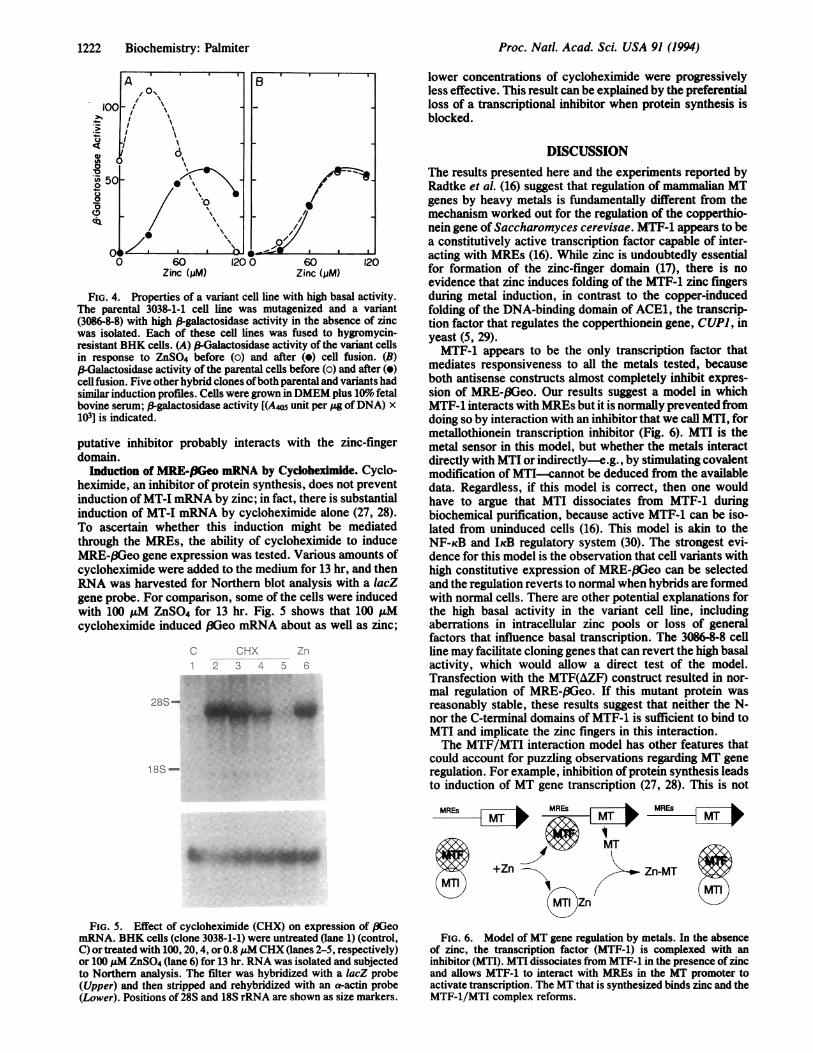
MTF-1 is active when isolated from uninduced cells (16), itmay dissociate from an inhibitor during purification. Trans-fection of CMV-MTF-1 into these cells may result in consti-tutive expression by overwhelming the inhibitor. To test thisidea, 3038-1-1 cells were mutagenized six times at 2- to3-week intervals with ICR-191 as described (22), and thencells that were resistant to G418 at 3.2 mg/ml were selectedin the absence of zinc. About 50 clones were picked andexamined for induction of ,-galactosidase by zinc. Oneclone, 3086-8, had a high basal expression that was stablytransmitted to several subclones. Elevated expression in theabsence of zinc could result from a variety of mechanisms,including dominant mutations in transcription factors inter-acting with MRE-,8Geo or in the reporter gene itself. Toeliminate some of these possibilities, a subclone (3086-8-8)was fused to hygromycin-resistant BHK cells and stablehybrids were selected and then tested for regulation ofMRE-(3Geo. This subclone had a high basal activity in theabsence of zinc and reverted to normal when cell hybridswere prepared (Fig. 4A). The parental clone (3038-1-1) hadlow basal activity before and after cell fusion (Fig. 4B). Notethat the dose-response curve for the variant is shifted to theleft relative to the parental cells. Overexpression of MTF-1produces a similar result (data not shown). Because thismight reflect a change in zinc transport, the zinc content ofthe variant cells was compared with that of the parental cloneby growing them for 3 days in DMEM plus 10% fetal bovineserum with 65Zn (5 ,uM); there was no significant differencein total cellular zinc (data not shown). These results indicatethat the high basal activity is recessive and suggest that itcould be due to the loss of a transcriptional inhibitor.An MTF-1 construct, CMV-MTF(AZF), that cannot bind
to DNA was prepared by deleting the zinc-finger domains,with the idea that it might compete with the inhibitor andthereby liberate endogenous MTF-1. However, when thisconstruct was transfected into clone 3038-1-1, the inductionof MRE-f3Geo activity by zinc was normal (Fig. 3). Thus, the
Biochemistry: Palmiter
Proc. Natl. Acad. Sci. USA 91 (1994)
ei 50o
Ia
0 60 1200 60 120Zinc (jjM) Zinc (;jM)
FIG. 4. Properties of a variant cell line with high basal activity.The parental 3038-1-1 cell line was mutagenized and a variant(3086-8-8) with high P-galactosidase activity in the absence of zincwas isolated. Each of these cell lines was fused to hygromycin-resistant BHK cells. (A) ,B Galactosidase activity of the variant cellsin response to ZnS04 before (o) and after (o) cell fusion. (B)P-alactosidase activity of the parental cells before (o) and after (o)cell fusion. Five other hybrid clones ofboth parental and variants hadsimilar induction profiles. Cells were grown in DMEM plus 109o' fetalbovine serum; ,B galactosidase activity [(A405 unit per jig ofDNA) x103] is indicated.
putative inhibitor probably interacts with the zinc-fingerdomain.
'Inducffon of MRE-fiGe mRNA by Cydioheximide. Cyclo-heximide, an inhibitor of protein synthesis, does not preventinduction ofMT-I mRNA by zinc; in fact, there is substantialinduction of MT-I mRNA by cycloheximide alone (27, 28).To ascertain whether this induction might be mediatedthrough the MREs, the ability of cycloheximide to induceMRE-,BGeo gene expression was tested. Various amounts ofcycloheximide were added to the medium for 13 hr. and thenRNA was harvested for Northern blot analysis with a lacZgene probe. For comparison, some of the cells were inducedwith 100 uM ZnS04 for 13 hr. Fig. S shows that 100 ,uMcycloheximide induced SGeo mRNA about as well as zinc;
C CHX Z11.1 2 3 4 5 6
28S-
1 8S -
lower concentrations of cycloheximide were progressivelyless effective. This result can be explained by the preferentialloss of a transcriptional inhibitor when protein synthesis isblocked.
DISCUSSIONThe results presented here and the experiments reported byRadtke et al. (16) suggest that regulation of mammalian MTgenes by heavy metals is fundamentally different from themechanism worked out for the regulation of the copperthio-nein gene of Saccharomyces cerevisae. MTF-1 appears to bea constitutively active transcription factor capable of inter-acting with MREs (16). While zinc is undoubtedly essentialfor formation of the zinc-finger domain (17), there is noevidence that zinc induces folding of the MTF-1 zinc fingersduring metal induction, in contrast to the copper-inducedfolding of the DNA-binding domain of ACE1, the transcrip-tion factor that regulates the copperthionein gene, CUP), inyeast (5, 29).MTF-1 appears to be the only transcription factor that
mediates responsiveness to all the metals tested, becauseboth antisense constructs almost completely inhibit expres-sion of MRE-(3Geo. Our results suggest a model in whichMTF-1 interacts with MREs but it is normally prevented fromdoing so by interaction with an inhibitor that we call MTI, formetallothionein transcription inhibitor (Fig. 6). MTI is themetal sensor in this model, but whether the metals interactdirectly with MTI or indirectly-e.g., by stimulating covalentmodification of MTI-cannot be deduced from the availabledata. Regardless, if this model is correct, then one wouldhave to argue that MTI dissociates from MTF-1 duringbiochemical purification, because active MTF-1 can be iso-lated from uninduced cells (16). This model is akin to theNF-KB and IicB regulatory system (30). The strongest evi-dence for this model is the observation that cell variants withhigh constitutive expression of MRE-,8Geo can be selectedand the regulation reverts to normal when hybrids are formedwith normal cells. There are other potential explanations forthe high basal activity in the variant cell line, includingaberrations in intracellular zinc pools or loss of generalfactors that influence basal transcription. The 3086-8-8 cellline may facilitate cloning genes that can revert the high basalactivity, which would allow a direct test of the model.Transfection with the MTF(AZF) construct resulted in nor-mal regulation of MRE-,fGeo. If this mutant protein wasreasonably stable, these results suggest that neither the N-nor the C-terminal domains of MTF-1 is sufficient to bind toMTI and implicate the zinc fingers in this interaction.The MTF/MTI interaction model has other features that
could account for puzzling observations regarding MT generegulation. For example, inhibition of protein synthesis leadsto induction of MT gene transcription (27, 28). This is not
MRWs MR-sILMRrsM T
A%~~A MT
FIG. 5. Effect of cycloheximide (CHX) on expression of BGeomRNA. BHK cells (clone 3038-1-1) were untreated (lane 1) (control,C) or treated with 100, 20, 4, or 0.8 iM CHX (lanes 2-5, respectively)or 100 ,uM ZnSO4 (lane 6) for 13 hr. RNA was isolated and subjectedto Northern analysis. The filter was hybridized with a lacZ probe(Upper) and then stripped and rehybridized with an a-actin probe(Lower). Positions of 28S and 18S rRNA are shown as size markers.
FIG. 6. Model of MT gene regulation by metals. In the absenceof zinc, the transcription factor (MTF-1) is complexed with aninhibitor (MTI). MTI dissociates from MTF-1 in the presence of zincand allows MTF-1 to interact with MREs in the MT promoter toactivate transcription. The MT that is synthesized binds zinc and theMTF-1/MTI complex reforms.
.",Frfmmw
1222 Biochemistry: Palmiter

Proc. Natl. Acad. Sci. USA 91 (1994) 1223
easily explained if MTF-1 is regulated directly by metals.However, if MTI has a shorter half-life than MTF-1, theninhibition of protein synthesis could liberate MTF-1 fortranscriptional activation. Another observation that is neatlyexplained by this model is the constitutive expression ofMTgenes in certain organs. For example, the highest levels ofMT expression in uninduced adult mice are found in testesand brain. Perhaps MTI is not expressed in those organs.How do different metals cause MTI to dissociate from
MTF-1? A conservative view is that MTI is sensitive only tozinc, either because it has binding sites for zinc or becausezinc activates a system leading to MTI dissociation. Intra-cellular zinc may exist in several pools. It is tightly bound tothe many zinc metalloproteins; however, it is also weaklybound to a wide variety of proteins and small molecules.Perhaps all the metals capable of inducing MT do so bydisplacing zinc from the weakly bound pool, and this dis-placed zinc influences the interaction of MTI with MTF-1.Thus, if MTI responds directly to zinc, it presumably has anaffinity for zinc that is below that of the structural proteinsbut higher than that of the weakly bound zinc. Hence, anincrease in "free" zinc would lead to liberation of MTF-1.The resulting induction of MT would lead to synthesis ofapo-MT that would bind the available zinc and restore thesystem to equilibrium (Fig. 6). The conceptual simplicity ofthis model is its most attractive feature. However, variationson this theme can easily be envisaged. For example, theremay be several different MTIs with different properties, somemetals other than zinc may directly interact with MTI, andMTI may not bind metals directly.Copper, mercury, nickel, and PDC fail to induce MRE-
pGeo in Opti-MEMAZn (Fig. 2). However, PDC and coppercan induce maximal expression, and mercury and nickel canpartially induce MRE-BGeo, when the cells are grown in aslittle as 0.5 uM ZnSO4, which by itself results in no expres-sion of MRE-pGeo. Thus, PDC and these metals clearly actby displacing extracellular zinc. The same argument mayapply intracellularly, where zinc is always present. Cadmiummay also act by displacing zinc. When cells are deprived ofzinc they become very sensitive to zinc and relatively insen-sitive to cadmium (Fig. 1). The increased sensitivity to zinccould be due either to increased transport of zinc or to achange in the relative amounts or affinities of MTF-1 andMTI. The decreased sensitivity to cadmium is predicted ifzinc is the only effective inducer, because during zinc star-vation the low-affinity pool of zinc would be depleted first;thus, addition of small amounts of cadmium would fill thispool without liberating any zinc. Addition of more cadmiumwould displace zinc from higher-affinity pools. In the case ofextreme zinc depletion, addition of cadmium leads to celldeath without induction (data not shown).
In this view, the key to understanding the regulation ofmammalian MT genes by metals will be the isolation andcharacterization ofMTI and determination ofhow it interactswith MTF-1 and responds to metals.
I thank Peter Searle for constructing and testing the promoter withfive MRE-d' elements. I am grateful to Seth Findley for isolating the
MTF-I clone and to Phil Soriano for the (3Geo construct. I appreciatethe advice of Helen Blau on cell fusion and the constructive sugges-tions ofmy colleagues. This work was supported in part by NationalInstitutes of Health Grant HD09172.
1. Kagi, J. H. R. & Kojima, Y. (1987) Experientia Suppl. 52,25-80.
2. Suzuki, K. T., Imura, N. & Kimura, M. (1993) MetallothioneinIII: Biological Roles and Medical Implications (Birkhaeuser,Basel).
3. Tamai, K. T., Gralla, E. B., Ellerby, L. M., Valentine, J. S. &Thiele, D. J. (1993) Proc. Natl. Acad. Sci. USA 90, 8013-8017.
4. Waalkes, M. P. & Goering, P. L. (1990) Chem. Res. Toxicol. 3,281-288.
5. Hamer, D. H. (1986) Annu. Rev. Biochem. 55, 913-951.6. Stuart, G. W., Searle, P. F. & Palmiter, R. D. (1985) Nature
(London) 317, 828-831.7. Searle, P. F. (1990) Nucleic Acids Res. 18, 4683-4690.8. Anderson, R. D., Taplitz, S. J., Wong, S., Bristol, G., Larkin,
B. M. & Herschman, H. R. (1987) Mol. Cell. Biol. 7, 3574-3581.
9. Mueller, P. R., Salser, S. J. & Wold, B. (1988) Genes Dev. 2,412-427.
10. Westin, W. & Schaffner, W. (1988) EMBO J. 7, 3763-3770.11. Imbert, J., Zafarullah, M., Culotta, V. C., Gedamu, L. &
Hamer, D. (1989) Mol. Cell. Biol. 9, 5315-5323.12. Anderson, R. D., Taplitz, S. J., Oberbauer, A. M., Calame,
K. L. & Herschman, H. R. (1990) Nucleic Acids Res. 18,6049-6055.
13. Sequin, C. & Prevost, J. (1988) Nucleic Acids Res. 16, 10547-10560.
14. Koizumi, S., Suzuki, K. & Otsuka, F. (1992) J. Biol. Chem.267, 18659-18664.
15. Czupryn, M., Brown, W. E. & Vallee, B. L. (1992) Proc. Natl.Acad. Sci. USA 89, 10395-10399.
16. Radtke, F., Heuchel, R., Georgiev, O., Hergersberg, M.,Gariglio, M., Dembic, Z. & Schaffner, W. (1993) EMBO J. 12,1355-1362.
17. Vallee, B. L. (1993) Physiol. Rev. 73, 79-118.18. Durnam, D. M. & Palmiter, R. D. (1984) Mol. Cell. Biol. 4,
484-491.19. Friedrich, G. & Soriano, P. (1991) Genes Dev. 5, 1513-1523.20. Palmiter, R. D., Behringer, R. R., Quaife, C. J., Maxwell, F.,
Maxwell, I. H. & Brinster, R. D. (1987) Cell 50, 435-443.21. Searle, P. F., Stuart, G. W. & Palmiter, R. D. (1985) Mol. Cell.
Biol. 5, 1480-1489.22. McKendry, R., John, J., Flavell, D., Muller, M., Kerr, I. M. &
Stark, G. R. (1991) Proc. Natl. Acad. Sci. USA 88, 11455-11459.
23. Blau, H. M., Chiu, C.-P. & Webster, C. (1983) Cell 32, 1171-1180.
24. Durnam, D. M. & Palmiter, R. D. (1983) Anal. Biochem. 131,385-393.
25. Sambrook, J., Fritsch, E. F. & Maniatis, T. (1989) MolecularCloning: A Laboratory Manual (Cold Spring Harbor Lab.Press, Plainview, NY), 2nd Ed.
26. Palmiter, R. D., Chen, H. Y. & Brinster, R. L. (1982) Cell 29,701-710.
27. Mayo, K. E. & Palmiter, R. D. (1981) J. Biol. Chem. 256,2621-2624.
28. Alam, J. & Smith, A. (1992) J. Biol. Chem. 267, 16379-16384.29. Furst, P., Hu, S., Hackett, R. & Hamer, D. (1988) Cell 55,
705-717.30. Baeuerle, P. A. & Baltimore, D. (1988) Science 242, 540-546.
Biochemistry: Palmiter





























