Embed Size (px)
Citation preview

Journal of Bioscience and BioengineeringVOL. 108 No. 1, 41–46, 2009
www.elsevier.com/locate/jbiosc
Regioselective hydroxylation of isoflavones by Streptomyces avermitilis MA-4680
Changhyun Roh,1,§ Su-Hyun Seo,1,§ Kwon-Young Choi,1 Minho Cha,1 Bishnu Prasad Pandey,1
June-Hyung Kim,2 Jun-Seong Park,3 Duck Hee Kim,3 Ih Seop Chang,3 and Byung-Gee Kim1,⁎
⁎ CorrespondE-mail add
§ The first tw
1389-1723/$doi:10.1016/j
School of Chemical and Biological Engineering, Institute of Molecular Biology and Genetics, Institute of Bioengineering, Seoul National University, Seoul,South Korea1 Department of Chemical Engineering, Dong-A University, Hadan 840, Saha-Gu, Busan 604-714, South Korea2 and Skin Research Institute,
AmorePacific R&D Center, 314-1, Bora-dong, Giheung-gu, Yongin-si, Gyeonggi-do 449-729, South Korea3
Received 7 July 2008; accepted 6 February 2009
Screening of bacterial whole cells was performed for regioselective hydroxylation of daidzein and genistein. Amongthe strains examined, Streptomyces avermitilis MA-4680 showed high ortho-dihydroxylation activity to produce 3′,4′,7-trihydroxyisoflavone and 3′,4′,5,7-tetrahydroxyisoflavone from daidzein (4′,7-dihydroxyisoflavone) and genistein (4′,5,7-trihydroxyisoflavone), respectively. Using 100 mg cells (wet wt.) and 1% (v/v) Triton X100 in 1 ml of total reaction volume,where 100 μl of the substrate solution (0.5 mM in 10% (v/v) mixed solvent of DMSO:MeOH=3:7) was added to 900 μl ofpotassium phosphate buffer (100 mM, pH 7.2), a 16% molar conversion yield of 3′,4′,7-trihydroxyisoflavone was obtainedfrom 0.5 mM daidzein after 24 h of reaction time at 28 °C and 200 rpm. Ketoconazole significantly (ca. 90%) inhibited theortho-hydroxylation activity of daidzein, suggesting that cytochrome P450 enzymes putatively play roles in regiospecificdaidzein hydroxylation. The analysis of the reaction products was determined by gas chromatography/mass spectrometry(GC/MS) and 1H NMR.
© 2009, The Society for Biotechnology, Japan. All rights reserved.
[Key words: Cytochrome P450; Monohydroxylation; Streptomyces avermitilis MA-4680; Daidzein; Genistein]
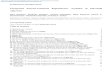
Daidzein (4′,7-dihydroxyisoflavone) and genistein (4′,5,7-trihy-droxyisoflavone) are diphenolic phytoestrogen compounds found innumerous plants and soybeans (1). They have been reported to act asantioxidants, antimicrobials, free radical scavengers, metal chelators,and antibacterial agents (1–3). In addition, isoflavones are known tohave medicinal and chemopreventive activities in human health.Isolation and synthesis of isoflavones have become frequent researchtopics due to their interesting biological activities (4–6). Recently,hydroxylated products of daidzein and genistein (Fig. 1) haveattracted considerable scientific interest because of their health-related qualities. Hydroxylated products have potent antioxidantproperties that contribute to their cholesterol-lowering effects,cardiovascular protection, antitumor effects, and anticarcinogenicproperties (7–13). Furthermore, compounds with the ortho-dihy-droxy group are known to exhibit anti-inflammatory and anti-allergicactivities (12), and express anticarcinogenic properties due to theinhibition of protein tyrosine kinases (14) by acting as potenttyrosinase inhibitors (15) and active inhibitors of lipoxygenases(16). Hydroxylated isoflavones are invaluable compounds for low-ering the incidence of cancer-related diseases (7, 17).
Regiospecific hydroxylation of aromatic compounds by chemicalsynthesis is difficult and involves diverse reaction steps. Theconversion of aromatic hydrocarbons into hydroxylated aromatic
ing author. Tel.: +82 2 880 6774; fax: +82 2 874 1206.ress: [email protected] (B.-G. Kim).o authors contributed equally to this work.
- see front matter © 2009, The Society for Biotechnology, Japan. All.jbiosc.2009.02.021
hydrocarbons, i.e., inserting an oxygen atom into a carbon-hydrogenbond, is one of the key features of oxidative metabolism of manyaromatic compounds (18). Regiospecific microbial hydroxylation of anon-activated carbon atom of aromatic compounds is a very attractiveand remarkable step in biosynthesis. The introduction of hydroxylgroups into isoflavones by the use of microorganisms represents anattractive alternative to conventional chemical synthesis. The selec-tive modification of isoflavones by microorganisms is a powerful toolthat deserves further analysis. Klus and Rufer et al. reported that theantioxidant capacity of hydroxylated compounds is higher than theoriginal isoflavones, which might affect their biological properties(7, 12). Among the metabolites, they identified 3′,4′,7-trihydroxyiso-flavone and 3′,4′,5,7-tetrahydroxyisoflavone, which exhibited higherantioxidant activity in the oxygen radical absorbance capacity assay aswell as the oxidation of low-density lipoproteins (12, 13). Thehydroxylated forms at the 3′ position of the isoflavone B-ring arelikely to improve their antioxidant properties (19, 20).
The main objective of this study is to screen microorganismsparticipating in hydroxylations of daidzein and genistein, which arethemajor isoflavone compounds of phytoestrogen. We have identifiedthat Streptomyces avermitilis MA-4680, an actinomycetes, efficientlyconverts daidzein and genistein to 3′,4′,7-trihydroxyisoflavone and3′,4′,5,7-tetrahydroxyisoflavone, respectively.
MATERIALS AND METHODS
Chemicals Daidzein, genistein, 3′,4′,7-trihydroxyisoflavone, quinidine, cou-marin, and erythromycin were obtained from Sigma-Aldrich Chemical Co. (St. Louis,
rights reserved.
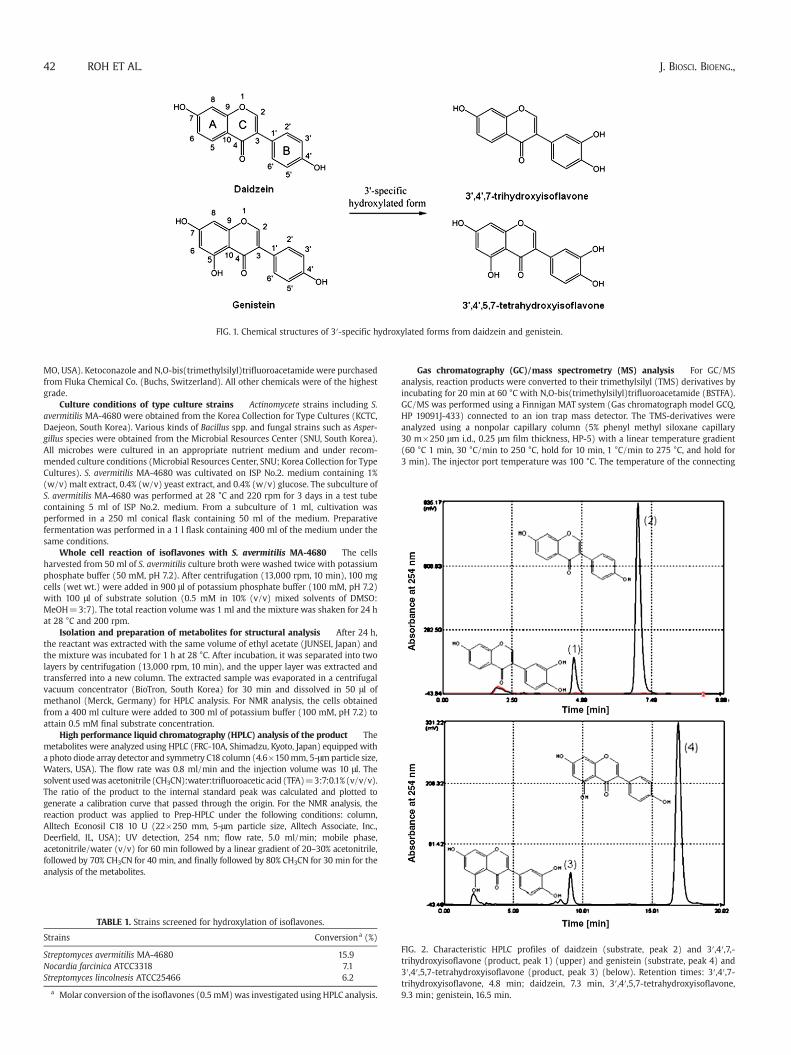
FIG. 1. Chemical structures of 3′-specific hydroxylated forms from daidzein and genistein.
42 ROH ET AL. J. BIOSCI. BIOENG.,
MO, USA). Ketoconazole and N,O-bis(trimethylsilyl)trifluoroacetamide were purchasedfrom Fluka Chemical Co. (Buchs, Switzerland). All other chemicals were of the highestgrade.
Culture conditions of type culture strains Actinomycete strains including S.avermitilis MA-4680 were obtained from the Korea Collection for Type Cultures (KCTC,Daejeon, South Korea). Various kinds of Bacillus spp. and fungal strains such as Asper-gillus species were obtained from the Microbial Resources Center (SNU, South Korea).All microbes were cultured in an appropriate nutrient medium and under recom-mended culture conditions (Microbial Resources Center, SNU; Korea Collection for TypeCultures). S. avermitilis MA-4680 was cultivated on ISP No.2. medium containing 1%(w/v) malt extract, 0.4% (w/v) yeast extract, and 0.4% (w/v) glucose. The subculture ofS. avermitilis MA-4680 was performed at 28 °C and 220 rpm for 3 days in a test tubecontaining 5 ml of ISP No.2. medium. From a subculture of 1 ml, cultivation wasperformed in a 250 ml conical flask containing 50 ml of the medium. Preparativefermentation was performed in a 1 l flask containing 400 ml of the medium under thesame conditions.
Whole cell reaction of isoflavones with S. avermitilis MA-4680 The cellsharvested from 50 ml of S. avermitilis culture broth were washed twice with potassiumphosphate buffer (50 mM, pH 7.2). After centrifugation (13,000 rpm, 10 min), 100 mgcells (wet wt.) were added in 900 μl of potassium phosphate buffer (100 mM, pH 7.2)with 100 μl of substrate solution (0.5 mM in 10% (v/v) mixed solvents of DMSO:MeOH=3:7). The total reaction volume was 1 ml and the mixture was shaken for 24 hat 28 °C and 200 rpm.
Isolation and preparation of metabolites for structural analysis After 24 h,the reactant was extracted with the same volume of ethyl acetate (JUNSEI, Japan) andthe mixture was incubated for 1 h at 28 °C. After incubation, it was separated into twolayers by centrifugation (13,000 rpm, 10 min), and the upper layer was extracted andtransferred into a new column. The extracted sample was evaporated in a centrifugalvacuum concentrator (BioTron, South Korea) for 30 min and dissolved in 50 μl ofmethanol (Merck, Germany) for HPLC analysis. For NMR analysis, the cells obtainedfrom a 400 ml culture were added to 300 ml of potassium buffer (100 mM, pH 7.2) toattain 0.5 mM final substrate concentration.
High performance liquid chromatography (HPLC) analysis of the product Themetabolites were analyzed using HPLC (FRC-10A, Shimadzu, Kyoto, Japan) equipped witha photo diode array detector and symmetry C18 column (4.6×150mm, 5-μmparticle size,Waters, USA). The flow rate was 0.8 ml/min and the injection volume was 10 μl. Thesolvent usedwas acetonitrile (CH3CN):water:trifluoroacetic acid (TFA)=3:7:0.1% (v/v/v).The ratio of the product to the internal standard peak was calculated and plotted togenerate a calibration curve that passed through the origin. For the NMR analysis, thereaction product was applied to Prep-HPLC under the following conditions: column,Alltech Econosil C18 10 U (22×250 mm, 5-μm particle size, Alltech Associate, Inc.,Deerfield, IL, USA); UV detection, 254 nm; flow rate, 5.0 ml/min; mobile phase,acetonitrile/water (v/v) for 60 min followed by a linear gradient of 20–30% acetonitrile,followed by 70% CH3CN for 40 min, and finally followed by 80% CH3CN for 30 min for theanalysis of the metabolites.
TABLE 1. Strains screened for hydroxylation of isoflavones.
Strains Conversiona (%)
Streptomyces avermitilis MA-4680 15.9Nocardia farcinica ATCC3318 7.1Streptomyces lincolnesis ATCC25466 6.2
a Molar conversion of the isoflavones (0.5mM)was investigated using HPLC analysis.
Gas chromatography (GC)/mass spectrometry (MS) analysis For GC/MSanalysis, reaction products were converted to their trimethylsilyl (TMS) derivatives byincubating for 20 min at 60 °C with N,O-bis(trimethylsilyl)trifluoroacetamide (BSTFA).GC/MS was performed using a Finnigan MAT system (Gas chromatograph model GCQ,HP 19091J-433) connected to an ion trap mass detector. The TMS-derivatives wereanalyzed using a nonpolar capillary column (5% phenyl methyl siloxane capillary30 m×250 μm i.d., 0.25 μm film thickness, HP-5) with a linear temperature gradient(60 °C 1 min, 30 °C/min to 250 °C, hold for 10 min, 1 °C/min to 275 °C, and hold for3 min). The injector port temperature was 100 °C. The temperature of the connecting
FIG. 2. Characteristic HPLC profiles of daidzein (substrate, peak 2) and 3′,4′,7,-trihydroxyisoflavone (product, peak 1) (upper) and genistein (substrate, peak 4) and3′,4′,5,7-tetrahydroxyisoflavone (product, peak 3) (below). Retention times: 3′,4′,7-trihydroxyisoflavone, 4.8 min; daidzein, 7.3 min, 3′,4′,5,7-tetrahydroxyisoflavone,9.3 min; genistein, 16.5 min.
ORTHO-SELECTIVE HYDROXYLATION OF ISOFLAVONES 43VOL. 108, 2009
parts was 300 °C and the electron energy for the EI mass spectra was 70 eV.Identification was performed by comparison of retention time and mass spectral data(recorded by full scan in the selected ion mode; m/z 100–600) of the sample with thatof authentic references.
Nuclear magnetic resonance (NMR) spectroscopy The NMR spectra wereobtained in DMSO-d6 (Aldrich, Chicago, IL, USA) using a Bruker Advance 400instrument (400 MHz, 9.4 T) (Bruker, Karlsruhe, Germany). For the 1H NMRexperiment, 32 transients were acquired with a spectral width of 8000 Hz. All NMRdata were processed using XWINNMR (Bruker). The structures of twometabolites fromdaidzein and genistein were determined based on the interpretation of the NMR data.
Effects of P450 inhibitors on hydroxylation of isoflavone The inhibitionstudy of daidzein hydroxylation by typical cytochrome P450 inhibitors was performedin the presence of 0.5 mM cytochrome P450 inhibitors. The P450 inhibitors used wereas follows: quinidine, coumarin, ketoconazole, and erythromycin (21–23). The extent ofinhibition was quantitatively measured by HPLC analysis after a 24 h reaction. For thecontrol, the same experiment was performed without inhibitors.
Effects of detergents on daidzein hydroxylation The effect of detergents onthe hydroxylation of daidzeinwas examined using 1% (v/v) of the final concentration ofthe following detergent: Tween 20, Tween 80, Triton X100, Triton X114, and Brij 35.
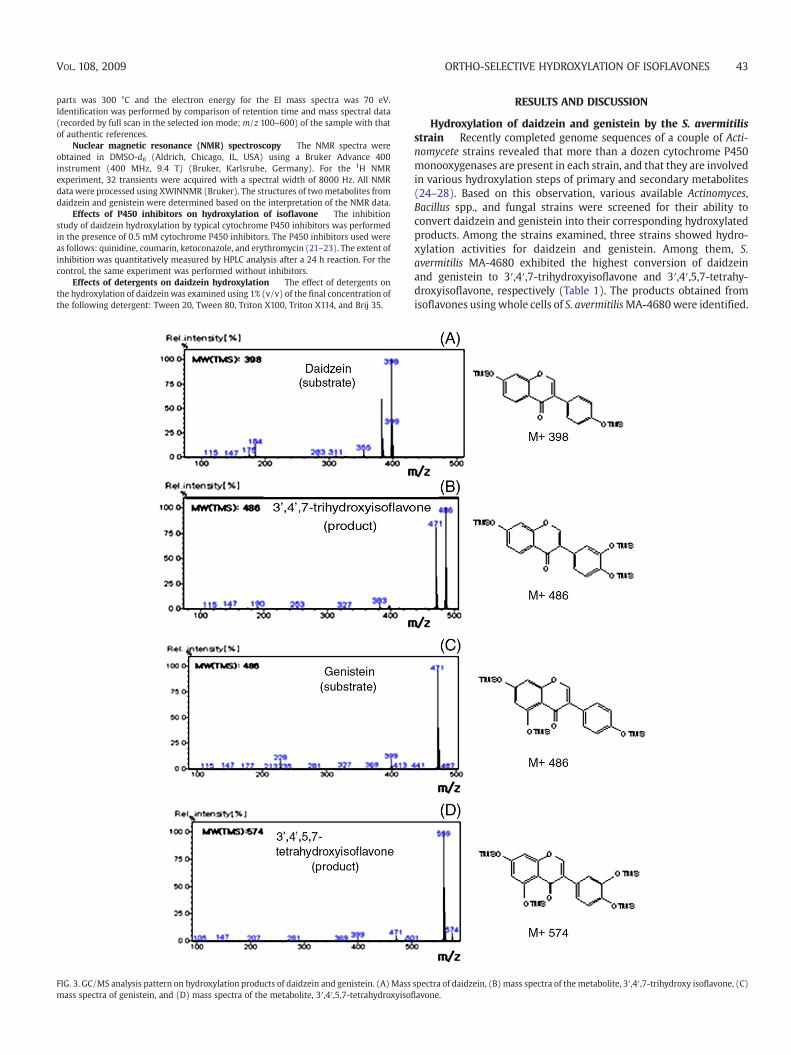
FIG. 3. GC/MS analysis pattern on hydroxylation products of daidzein and genistein. (A) Massmass spectra of genistein, and (D) mass spectra of the metabolite, 3′,4′,5,7-tetrahydroxyisof
RESULTS AND DISCUSSION
Hydroxylation of daidzein and genistein by the S. avermitilisstrain Recently completed genome sequences of a couple of Acti-nomycete strains revealed that more than a dozen cytochrome P450monooxygenases are present in each strain, and that they are involvedin various hydroxylation steps of primary and secondary metabolites(24–28). Based on this observation, various available Actinomyces,Bacillus spp., and fungal strains were screened for their ability toconvert daidzein and genistein into their corresponding hydroxylatedproducts. Among the strains examined, three strains showed hydro-xylation activities for daidzein and genistein. Among them, S.avermitilis MA-4680 exhibited the highest conversion of daidzeinand genistein to 3′,4′,7-trihydroxyisoflavone and 3′,4′,5,7-tetrahy-droxyisoflavone, respectively (Table 1). The products obtained fromisoflavones usingwhole cells of S. avermitilisMA-4680were identified.
spectra of daidzein, (B) mass spectra of the metabolite, 3′,4′,7-trihydroxy isoflavone, (C)lavone.
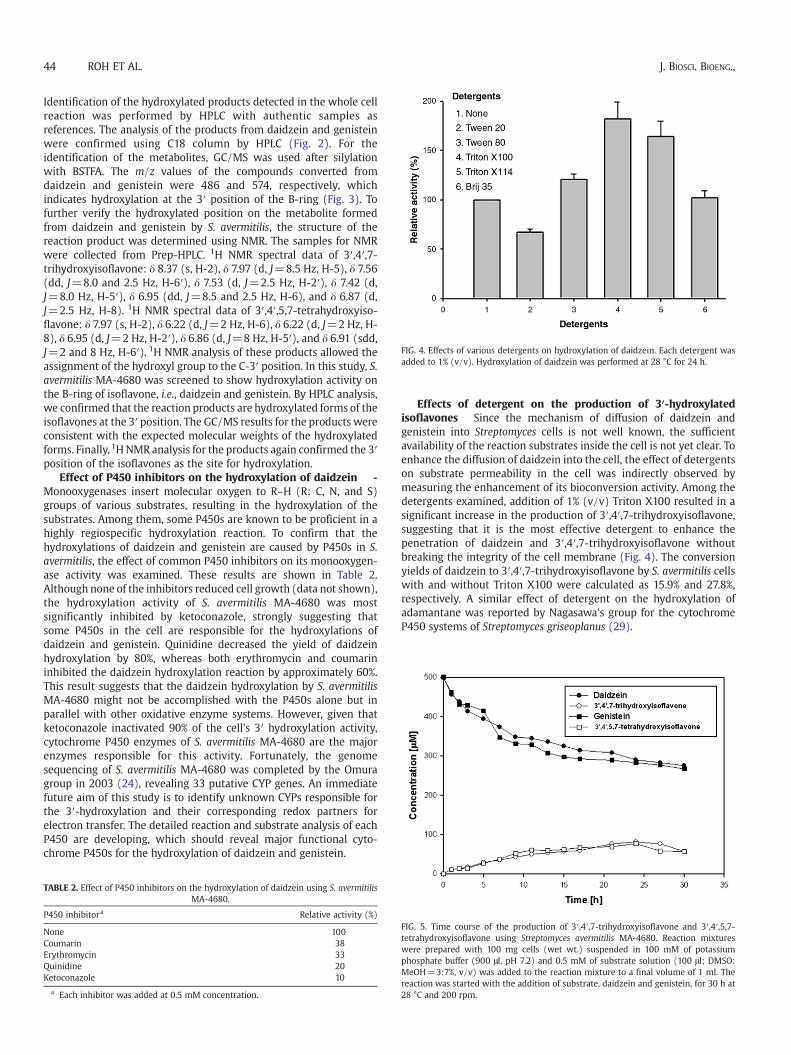
FIG. 4. Effects of various detergents on hydroxylation of daidzein. Each detergent wasadded to 1% (v/v). Hydroxylation of daidzein was performed at 28 °C for 24 h.
44 ROH ET AL. J. BIOSCI. BIOENG.,
Identification of the hydroxylated products detected in the whole cellreaction was performed by HPLC with authentic samples asreferences. The analysis of the products from daidzein and genisteinwere confirmed using C18 column by HPLC (Fig. 2). For theidentification of the metabolites, GC/MS was used after silylationwith BSTFA. The m/z values of the compounds converted fromdaidzein and genistein were 486 and 574, respectively, whichindicates hydroxylation at the 3′ position of the B-ring (Fig. 3). Tofurther verify the hydroxylated position on the metabolite formedfrom daidzein and genistein by S. avermitilis, the structure of thereaction product was determined using NMR. The samples for NMRwere collected from Prep-HPLC. 1H NMR spectral data of 3′,4′,7-trihydroxyisoflavone: δ 8.37 (s, H-2), δ 7.97 (d, J=8.5 Hz, H-5), δ 7.56(dd, J=8.0 and 2.5 Hz, H-6′), δ 7.53 (d, J=2.5 Hz, H-2′), δ 7.42 (d,J=8.0 Hz, H-5′), δ 6.95 (dd, J=8.5 and 2.5 Hz, H-6), and δ 6.87 (d,J=2.5 Hz, H-8). 1H NMR spectral data of 3′,4′,5,7-tetrahydroxyiso-flavone: δ 7.97 (s, H-2), δ 6.22 (d, J=2 Hz, H-6), δ 6.22 (d, J=2 Hz, H-8), δ 6.95 (d, J=2 Hz, H-2′), δ 6.86 (d, J=8 Hz, H-5′), and δ 6.91 (sdd,J=2 and 8 Hz, H-6′). 1H NMR analysis of these products allowed theassignment of the hydroxyl group to the C-3′ position. In this study, S.avermitilis MA-4680 was screened to show hydroxylation activity onthe B-ring of isoflavone, i.e., daidzein and genistein. By HPLC analysis,we confirmed that the reaction products are hydroxylated forms of theisoflavones at the 3′ position. The GC/MS results for the products wereconsistent with the expected molecular weights of the hydroxylatedforms. Finally, 1H NMR analysis for the products again confirmed the 3′position of the isoflavones as the site for hydroxylation.
Effect of P450 inhibitors on the hydroxylation of daidzein -Monooxygenases insert molecular oxygen to R–H (R: C, N, and S)groups of various substrates, resulting in the hydroxylation of thesubstrates. Among them, some P450s are known to be proficient in ahighly regiospecific hydroxylation reaction. To confirm that thehydroxylations of daidzein and genistein are caused by P450s in S.avermitilis, the effect of common P450 inhibitors on its monooxygen-ase activity was examined. These results are shown in Table 2.Although none of the inhibitors reduced cell growth (data not shown),the hydroxylation activity of S. avermitilis MA-4680 was mostsignificantly inhibited by ketoconazole, strongly suggesting thatsome P450s in the cell are responsible for the hydroxylations ofdaidzein and genistein. Quinidine decreased the yield of daidzeinhydroxylation by 80%, whereas both erythromycin and coumarininhibited the daidzein hydroxylation reaction by approximately 60%.This result suggests that the daidzein hydroxylation by S. avermitilisMA-4680 might not be accomplished with the P450s alone but inparallel with other oxidative enzyme systems. However, given thatketoconazole inactivated 90% of the cell's 3′ hydroxylation activity,cytochrome P450 enzymes of S. avermitilis MA-4680 are the majorenzymes responsible for this activity. Fortunately, the genomesequencing of S. avermitilis MA-4680 was completed by the Omuragroup in 2003 (24), revealing 33 putative CYP genes. An immediatefuture aim of this study is to identify unknown CYPs responsible forthe 3′-hydroxylation and their corresponding redox partners forelectron transfer. The detailed reaction and substrate analysis of eachP450 are developing, which should reveal major functional cyto-chrome P450s for the hydroxylation of daidzein and genistein.
TABLE 2. Effect of P450 inhibitors on the hydroxylation of daidzein using S. avermitilisMA-4680.
P450 inhibitora Relative activity (%)
None 100Coumarin 38Erythromycin 33Quinidine 20Ketoconazole 10
a Each inhibitor was added at 0.5 mM concentration.
Effects of detergent on the production of 3′-hydroxylatedisoflavones Since the mechanism of diffusion of daidzein andgenistein into Streptomyces cells is not well known, the sufficientavailability of the reaction substrates inside the cell is not yet clear. Toenhance the diffusion of daidzein into the cell, the effect of detergentson substrate permeability in the cell was indirectly observed bymeasuring the enhancement of its bioconversion activity. Among thedetergents examined, addition of 1% (v/v) Triton X100 resulted in asignificant increase in the production of 3′,4′,7-trihydroxyisoflavone,suggesting that it is the most effective detergent to enhance thepenetration of daidzein and 3′,4′,7-trihydroxyisoflavone withoutbreaking the integrity of the cell membrane (Fig. 4). The conversionyields of daidzein to 3′,4′,7-trihydroxyisoflavone by S. avermitilis cellswith and without Triton X100 were calculated as 15.9% and 27.8%,respectively. A similar effect of detergent on the hydroxylation ofadamantane was reported by Nagasawa's group for the cytochromeP450 systems of Streptomyces griseoplanus (29).
FIG. 5. Time course of the production of 3′,4′,7-trihydroxyisoflavone and 3′,4′,5,7-tetrahydroxyisoflavone using Streptomyces avermitilis MA-4680. Reaction mixtureswere prepared with 100 mg cells (wet wt.) suspended in 100 mM of potassiumphosphate buffer (900 μl, pH 7.2) and 0.5 mM of substrate solution (100 μl; DMSO:MeOH=3:7%, v/v) was added to the reaction mixture to a final volume of 1 ml. Thereaction was started with the addition of substrate, daidzein and genistein, for 30 h at28 °C and 200 rpm.

TABLE 3. Investigation of known hydroxylated compounds of isoflavone in whole cellreactions.
Sources Substrates Products References
Micrococcus spp. Daidzein 4′,7,8-trihydroxyisoflavone (7)Arthrobacter spp. 4′,6,7-trihydroxyisoflavone
4′,6,7,8-tetrahydroxyisoflavone3′,4′,7,8-tetrahydroxyisoflavone3′,4′,6,7-tetrahydroxyisoflavone
Fusarium spp. Daidzein 3′,4′,7-trihydroxyisoflavone (32)Ascochyta spp.Streptomyces spp. Daidzein 4′,7,8-trihydroxyisoflavone (31)Nocardia spp. NRRL5646 Daidzein 4′,7,8-trihydroxyisoflavone (30)Mortierella isabellina
ATCC38063Streptomyces avermitilis
MA-4680Daidzein 3′,4′,7-trihydroxyisoflavone This study
Genistein 3′,4′,5,7-tetrahydroxyisoflavone
ORTHO-SELECTIVE HYDROXYLATION OF ISOFLAVONES 45VOL. 108, 2009
Biotransformation reaction profile Time course reactionprofiles of the formation of 3′-hydroxylated metabolites fromdaidzein and genistein were obtained using whole cell biotrans-formation. When 0.5 mM daidzein and genistein were used, ca.40% of the substrates were consumed within 24 h after addition.However, all the substrates consumed were not converted intotheir corresponding 3′-hydroxylated products. The maximum con-version to 3′-hydroxylated forms from each isoflavone was up to16% (Fig. 5), and the rate of hydroxylated product formation wasca. 0.85 μg/h/g cell wet weight. Like Nocardia spp., which showedhydroxylation and subsequent O-methylation at the same position(30), S. avermitilis MA-4680 also accomplished both hydroxylationand subsequent O-methylation by putative methyltransferase atthe same C-3′ position (data not shown). The specific compoundsfor the hydroxylation of the isoflavones are summarized in Table 3.Although the Arthrobacter spp. strain showed mono-hydroxylationof daidzein at C-6 position (4′,6,7-trihydroxyisoflavone) as well asdihydroxylation of daidzein at C-6 and C-8 (4′,6,7,8-tetrahydrox-yisoflavone), C-3′ and C-8 (3′,4′,7,8-tetrahydroxyisoflavone), andC-3′ and C-6 positions (3′,4′,6,7-tetrahydroxyisoflavone) as pre-viously reported, the Streptomyces strain only catalyzed hydroxyla-tion of daidzein at position C-8 (31). In the case of S. avermitilisMA-4680, C-3′ mono-hydroxylation (3′,4′,7-trihydroxyisoflavone)of daidzein and C-3′ and C-5 dihydroxylation (3′,4′,5,7-tetrahy-droxyisoflavone) of genistein were detected. This is the first reportindicating that S. avermitilis MA-4680 has the capability of hydro-xylation of isoflavones 3′ position. Using 100 g/l of S. avermitiliswet cell mass, 3′,4′,7-trihydroxyisoflavone and 3′,4′,5,7-tetrahy-droxyisoflavone were produced to the level of 2.03 mg/l in100 mM potassium phosphate buffer (pH 7.2) with 0.5 mM ofdaidzein and genistein, respectively. This biotransformation resultdemonstrates that ortho-dihydroxylated isoflavones exhibitingmore potent antioxidant activity can be produced on a large scaleusing microbial biotransformation.
Considering that S. avermitilis MA-4680 has 33 P450s, it is a quiteformidable task to identify the ones responsible for the 3′ dihydrox-ylation. Since P450s from Streptomyces strains cloned into the E. coliBL21(DE3) expression system with camA and camB from Pseudomo-nas putida do not often show P450 activities for these substrates,finding the right partners of electron transfer for monooxygenationincreases the complexity of this identification procedure (data notshown). The current 3′ ortho-dihydroxylation activity for isoflavonesusing wild type S. avermitilis MA-4680 is currently approximately300–500 times higher than the best E. coli recombinant systemexpressing P450 genes from various organisms, such as P. putida, B.subtilis, and B. licheniformis (data not shown). Therefore, the cloningand overexpression of the right P450s in recombinant Streptomycesstrains, especially S. coelicolor or S. avermitilis, can become a good
substitute model system replacing the recombinant E. coli BL21(DE3)system in industrial biotransformation of isoflavones. In addition,since Streptomyces are generally used as a natural resource formedicines and antibiotics requiring additional hydroxylation reac-tions for improved activities, identifying the substrate specificity of allthe sequenced P450s in Streptomyces, P450 library construction, anddevelopment of recombinant P450s in Streptomyces expressionsystems would be of great benefit.
ACKNOWLEDGMENTS
This research was partially supported by the Brain Korea 21program of the Ministry of Education, Republic of Korea (Grant No.A05-0572-A11101-06A2-00020B) and by the National ResearchLaboratory Program (Grant No. R0A-2007-000-10007-0), KoreaMinistry of Science and Technology. We thank Prof. Yoongho Limand Younghee Park from Konkuk University for their help in theinterpretation of the NMR analysis.
References
1. Dixon, R. A. and Ferreira, D.: Molecules of interest genistein, Phytochemistry, 60,205–211 (2002).
2. Middleton Jr., E., and Kandaswami, C.: Effects of flavanoids on immune andinflammatory cell functions, Biochem. Pharmacil., 43, 1167–1179 (1992).
3. Heinonen, S. M., Waehaelae, H. K., and Adlercreutz, H.:Metabolism of isoflavonesin human subjects, Phytochem. Rev., 1, 175–182 (2002).
4. Komiyama, K., Funayama, S., Anraku, Y., Mita, A., Takahashi, Y., Omura, S., andShimasaki, H.: Isolation of isoflavonoids possessing antioxidant activity from thefermentation broth of Streptomyces sp, J. Antibiot., 42, 1344–1349 (1989).
5. Mazur,W. M., Duke, J. A., Wahala, K., Rasku, S., and Adlercreutz, H.: Isoflavonoidsand lignans in legumes: nutritional and health aspects in humans, J. Nutr. Biochem.,9, 193–200 (1998).
6. Foti, P., Erba, D., Riso, P., Spadafranca, A., Criscuoli, F., and Testolin, G.:Comparison between daidzein and genistein antioxidant activity in primary andcancer lymphocytes, Arch. Biochem. Biophys., 33, 421–427 (2005).
7. Klus, K. and Barz,W.: Formation of polyhydroxylated isoflavones from the soybeanseed isoflavones daidzein and glycitein by bacteria isolated from tempe, Arch.Microbiol., 164, 428–434 (1995).
8. Esaki, H., Watanabe, R., and Osaka, T.: Formation mechanism for potentantioxidative o-dihydroxyisoflavones in soybeans fermented with Aspergillussaitori, Biosci. Biotechnol. Biochem., 63, 851–858 (1999).
9. Kulling, S. E., Honig, D. M., Simat, T. J., and Metzler, M.: Oxidative in vitrometabolism of the soy phytoestrogens daidzein and genistein, J. Agric. Food. Chem.,48, 4963–4972 (2000).
10. Kulling, S. E., Honig, D. M., and Metzler, M.: Oxidative metabolism of the soyisoflavones daidzein and genistein in humans in vitro and in vivo, J. Agric. Food.Chem., 49, 3024–3033 (2001).
11. Hirota, A., Inaba, M., Chen, Y. C., Abe, N., Taki, S., Yano, M., and Kawaii, S.:Isolation of 8-hydroxyglycitein and 6-hydroxydaidzein from soybean miso, Biosci.Biotechnol. Biochem., 68, 1372–1374 (2004).
12. Rufer, C. E. and Kulling, S. E.: Antioxidant activity of isoflavones and their majormetabolites using different in vitro assays, J. Agric. Food. Chem., 54, 2926–2931(2006).
13. Rufer, C. E., Glatt, H., and Kulling, S. E.: Structural elucidation of hydroxylatedmetabolites of the isoflavan equol by gas chromatography-mass spectrometry andhigh-performance liquid chromatography-mass spectrometry, Drug. Metab. Dis-pos., 34, 51–60 (2006).
14. Akiyama, T., Ishida, J., Nakagawa, S., Ogawara, H., Watanabe, S., Itoh, N., Shibuya,M., and Fukami, Y.: Genistein, a specific inhibitor of tyrosine specific proteinkinases, J. Biol. Chem., 262, 5592–5595 (1987).
15. Chang, T. S., Ding, H. Y., and Lin, H. C.: Identifying 6,7,4′-trihydroxyisoflavone as apotent tyrosinase inhibitor, Biosci. Biotechnol. Boichem., 69, 1999–2001 (2005).
16. Voss, C., Sepulveda-Boza, S., and Zillike, F. W.: New isoflavonoids as inhibitors ofporcine 5-lipoxygenase, Biochem. Pharm., 44, 157–162 (1992).
17. Coward, L., Barnes, N. C., Setchell, K. D. R., and Barnes, S.: Genistein, daidzein, andtheir beta-glycoside conjugates: antitumor isoflavones in soybean foods fromAmerica and Asian diets, J. Agric. Food. Chem., 41, 1961–1967 (1993).
18. Holland, H. and Weber, H. K.: Enzymatic hydroxylation reactions, Curr. Opin.Biotechnol., 11, 547–553 (2000).
19. Briviba, K., Sepulveda-Boza, S., Zilliken, F., and Sies, H.: Isoflavonoids asinhibitors of lipid peroxidation and quenchers of singlet oxygen, in: C. A. Rice-Evans, L. Packer (Eds.), Flavonoids in Health and Disease, Marcel Dekker, Inc., NewYork, 1997, pp. 295–302 (1997).

46 ROH ET AL. J. BIOSCI. BIOENG.,
20. Arora, A., Nair, M. G., and Strasburg, G. M.: Antioxidant activities of isoflavonesand their biological metabolites in a liposomal system, Arch. Biochem. Biophys.,356, 133–141 (1998).
21. Liu, W. and Rosazza, J. P.: A soluble Bacillus cereus cytochrome P-450cin systemcatalyzes 1,4-cineole hydroxylations, Appl. Environ. Microbiol., 59, 3889–3893(1993).
22. Orita, M., Yamamoto, S., Katayama, N., Aoki, M., Takayama, K., Yamagiwa, Y.,Seki, N., Suzuki, H., Kurihara, H., Sakashita, H., Takeuchi, M., Fujita, S., Yamada,T., and Tanaka, A.: Coumarin and chromen-4-one analogues as tautomeraseinhibitors of macrophage migration inhibitory factor: discovery and X-ray crystal-lography, J. Med. Chem., 44, 540–547 (2001).
23. Harris, R. M., Wood, D. M., Bottomley, L., Blagg, S., Owen, K., Hughes, P. J., andWaring, R. H.: Phytoestrogens are potent inhibitors of estrogen sulfation:implications for breast cancer risk and treatment, J. Clin. Endocrin. Metab., 89,1779–1787 (2005).
24. Ikeda, H., Ishikawa, J., Hanamoto, A., Shinose, M., Kikuchi, H., Shiba, T., Sakaki,Y., Hattori, M., and Omura, S.: Complete genome sequence and comparativeanalysis of the industrial microorganism Streptomyces avermitilis, Nat. Biotechnol.,21, 526–531 (2003).
25. Bentley, S. D., Chater, K. F., Cerdeño-Tárraga, A. M., Challis, G. L., Thomson, N. R.,James, K. D., Harris, D. E., Quail, M. A., Kieser, H., and Harper, D., et al.: Completegenome sequence of themodel actinomycete Streptomyces coelicolor A3(2), Nature,417, 141–147 (2002).
26. Udwary, D. W., Zeigler, L., Asolkar, R. N., Singan, V., Lapidus, A., Fenical, W.,Jensen, P. R., and Moore, B. S.: Genome sequencing reveals complex secondarymetabolome in the marine actinomycete Salinispora tropica, Proc. Natl. Acad. Sci.U. S. A., 104, 10376–10381 (2007).
27. Ohnishi, Y., Ishikawa, J., Hara, H., Suzuki, H., Ikenoya,M., Ikeda, H., Yamashita, A.,Hattori, M., and Horinouchi, S.: Genome sequence of the streptomycin-producingmicroorganism Streptomyces griseus IFO 13350, J. Bacteriol.,190, 4050–4060 (2008).
28. Oliynyk, M., Samborskyy, M., Lester, J. B., Mironenko, T., Scott, N., Dickens, S.,Haydock, S. F., and Leadlay, P. F.: Complete genome sequence of the erythromycin-producing bacterium Saccharopolyspora erythraea NRRL23338, Nat. Biotechnol., 25,129–428 (2007).
29. Mitsukura, K., Kondo, Y., Yoshida, T., and Nagasawa, T.: Regioselective hydro-xylation of adamantane by Streptomyces griseoplanus cells, Appl. Microbiol.Biotechnol., 71, 502–504 (2006).
30. Maatooq, G. T. and Rosazza, J. P. N.: Metabolism of daidzein by Nocardia speciesNRRL 5646 and Mortierella isabellina ATCC 38063, Phytochem., 66, 1007–1011(2005).
31. Funayama, S., Anraku, Y., Mita, R., Komiyanma, Y., and Omura, S.: Structuralstudy of isoflavonoids possessing antioxidant activity isolated from fermentationbroth of Streptomyces sp, J. Antiobiot., 42, 1350–1355 (1989).
32. Barz, W. and Weltring, K. M.: Biodegradation of aromatic extractives of wood, in:T. Higuchi (Ed.), Biosynthesis and Biodegradation of Wood Components,Academic Press, New York, 1985, pp. 607–666 (1985).