Embed Size (px)
Citation preview

ORIGINAL PAPER
Regional Characteristics of Histamine Uptake into Neonatal RatAstrocytes
Katja Perdan-Pirkmajer • Sergej Pirkmajer •
Andreja Raztresen • Mojca Krzan
Received: 16 January 2013 / Revised: 20 March 2013 / Accepted: 23 March 2013
� Springer Science+Business Media New York 2013
Abstract Histaminergic signalling constitutes an attrac-
tive target for treatment of neuropsychiatric disorders. One
obstacle to developing new pharmacological options has
been failure to identify putative specific histamine trans-
porter responsible for histamine clearance. Although high-
affinity histamine uptake was detected in neonatal cortical
astrocytes, its existence in other brain regions remains lar-
gely unexplored. We investigated whether cerebellar and
striatal astrocytes participate in histamine clearance and
evaluated the role of organic cation transporters (OCT) in
astroglial histamine transport. Kinetic and pharmacological
characteristics of histamine transport were determined in
cultured astrocytes derived from neonatal rat cerebellum,
striatum and cerebral cortex. As well as astrocytes of cortical
origin, cultured striatal and cerebellar astrocytes displayed
temperature-sensitive, high-affinity histamine uptake.
Exposure to ouabain or Na?-free medium, supplemented
with choline chloride markedly depressed histamine trans-
port in cortical astrocytes. Conversely, histamine uptake in
striatal and cortical astrocytes was ouabain-resistant and was
only partially diminished during incubation in the absence of
Na?. Also, histamine uptake remained unaltered upon
exposure to OCT inhibitor corticosterone, although OCTs
were expressed in cultured astrocytes. Finally, histamine
transport in cerebellar and striatal astrocytes was not sensi-
tive to antidepressants. Despite common characteristics,
cerebellar astrocytes had lower affinity, but markedly higher
transport capacity for histamine compared to striatal astro-
cytes. Collectively, we provide evidence to suggest that
cerebellar, striatal as well as cortical astrocytes possess sat-
urable histamine uptake systems, which are not operated by
OCTs. In addition, our data indicate that Na?-independent
histamine carrier predominates in cerebellar and striatal
astrocytes, whereas Na?-dependent transporter underlies
histamine uptake in cortical astrocytes. Our findings impli-
cate a role for histamine transporters in regulation of extra-
cellular histamine concentration in cerebellum and striatum.
Inhibition of histamine uptake might represent a viable
option to modulate histaminergic neurotransmission.
Keywords Rat � Astrocytes � Histamine uptake �Regional characteristics
Introduction
Histamine is a monoamine neurotransmitter in the central
nervous system, where it participates in regulation of sleep-
wake cycle, fluid balance and feeding as well as higher brain
functions, such as mood, memory and locomotion [1]. Tar-
geting histamine signalling in the brain is the focus of efforts
to expand pharmacopoeia against neuropsychiatric disorders
[2], although exact role of the histaminergic system in the
pathogenesis of specific diseases remains poorly defined [1].
Several pharmacological approaches have been undertaken
to modulate neurotransmission in other monoaminergic
systems, including inhibition of monoamine uptake through
the specific, high-affinity Na?/Cl--dependent transporters
(uptake1) for serotonin (SERT), noradrenaline (NET) and
dopamine (DAT). Monoamine uptake inhibitors include
several pharmacotherapeutic agents as well as recreational
K. Perdan-Pirkmajer � A. Raztresen � M. Krzan (&)
Department of Pharmacology and Experimental Toxicology,
Faculty of Medicine, University of Ljubljana, Korytkova 2,
1001 Ljubljana, Slovenia
e-mail: [email protected]
S. Pirkmajer
Faculty of Medicine, Institute of Pathophysiology, University
of Ljubljana, Zaloska 4, 1001 Ljubljana, Slovenia
123
Neurochem Res
DOI 10.1007/s11064-013-1028-x

substances, underscoring essential role of monoamine
transporters in regulation of the noradrenergic, dopaminer-
gic and serotoninergic systems and their physiological
functions [3]. In contrast, the role of histamine uptake in
histaminergic neurotransmission is incompletely character-
ized, not least because histamine remains the only mono-
amine neurotransmitter, whose putative specific transporter
has not been identified.
Neurons essentially lack plasmalemmal high-affinity
uptake system for histamine [4, 5], although histamine
shares the same vesicular transporter (VMAT2) with other
monoamines [6]. Indeed, synaptosomal studies have either
detected only low-efficiency histamine transport [7] or have
even failed to convincingly demonstrate saturable histamine
uptake [8], a unique exception among monoamine neuro-
transmitters [4]. In direct contrast, astrocyte-enriched brain
tissue fractions from adult rat cortex displayed high-affinity
histamine transport [9]. Moreover, high-affinity as well as
low-affinity histamine uptake was repeatedly demonstrated
in cultured astrocytes [10–14]. In addition, evidence sug-
gests glial cells participate in histamine uptake and/or
inactivation in vivo [15–17]. Taken together these findings
indicate that astrocytes may represent an important site of
histamine uptake. Yet carriers involved in astrocyte hista-
mine transport have been incompletely characterized.
Although selective uptake1 transporters, which mediate
high-affinity monoamine uptake into presynaptic terminals
[3], are expressed in astrocytes [18, 19] failure to observe
significant histamine uptake in neuronal terminals strongly
suggests histamine is not transported by SERT, DAT and/or
NET. Consistent with this notion, uptake studies have
demonstrated that exposure to histamine has either very low
or no inhibitory effect on serotonin and dopamine uptake by
SERT and DAT, respectively [20–22]. In fact, histamine is
not considered a substrate for SERT, NET or DAT and
histamine uptake assays have often been used to detect
and assess monoamine transport not dependent on uptake1
[17, 23, 24].
On the other hand, histamine is a prominent substrate for
the non-specific, corticosterone-sensitive organic cation
transporters (OCT) 2 (Slc22A2) and OCT 3 (extraneuronal
monoamine transporter (EMT) or Slc22A3) from the
Slc22A family of solute carriers [25, 26]. Although OCT2
and 3 mediate low-affinity, high-capacity transport of a
range of organic cations, they show relative preference
towards histamine compared to other monoamines [27, 28]
and may play an important role in histamine clearance in
the central nervous system. Importantly, OCT expression
has been detected in neurons as well as astrocytes [13, 19,
29, 30]. Furthermore, OCT inhibitors corticosterone and
decynium-22 (D22) reduce histamine uptake in periven-
tricular hypothalamic minces [23]. Similarly, D22 reduces
histamine clearance in murine hippocampus in vivo [17].
Thus, OCT2 and OCT3 are thought to represent a major
mechanism for low-affinity, high-capacity histamine
clearance in the brain. However, whereas neuronal OCT
expression is compatible with failure to identify efficient
high-affinity histamine uptake in synaptosomes [8], kinetic
parameters of OCTs [27, 28] markedly differ from those
observed for high-affinity histamine uptake in astrocytes
[10–12, 14]. These data, when taken together, suggest
histamine uptake in the central nervous system is not
exclusively mediated by OCTs.
Indeed, a significant fraction of total histamine clearance
in periventricular region of hypothalamus was not inhibited
by OCT inhibitors corticosterone and D22 [23], indicating
OCT-mediated histamine uptake is probably complemented
by other, unidentified transporters. In agreement with this
notion, striatal histamine levels during post-ischemic
reperfusion were unaltered between OCT3-deficient mice
and their wild type controls [31], although prominent OCT3
expression is normally detected in murine striatum [29]. On
the other hand, accentuated increase in histamine concen-
tration in reperfused cortex of OCT3 knock-out mice [31]
suggests impaired cortical histamine clearance and, by
extension, a role for OCT3 in histamine elimination in brain
cortex. Similarly, basal histamine concentration was affec-
ted only in cortex and thalamus/hypothalamus of OCT3-
deficient mice, while remaining unaltered in most other brain
regions, including striatum and cerebellum [32]. Available
evidence therefore indicates that relative contribution of
OCTs and/or other transporters to total histamine uptake may
substantially vary between brain regions.
Nevertheless, previous studies have focused almost
exclusively on histamine uptake in astrocytes from cerebral
cortex. Indeed, the potential role of astrocytes in subserv-
ing histamine elimination in most other brain regions,
including striatum and cerebellum, remains largely unex-
plored despite functional importance of histaminergic
neurotransmission in these areas [1]. We determined
whether cerebellar and striatal astrocytes could participate
in histamine clearance and evaluated the role of OCTs in
astroglial histamine transport. Our findings implicate a role
for histamine transporters in regulation of extracellular
histamine concentration in cerebellum and striatum.
Methods
Materials and Reagents
All cell culture reagents, except fetal bovine serum (FBS),
Cambrex IEP GmbH (Wiesbaden, Germany) were obtained
from Gibco, Invitrogen (Paisley, Scotland, UK). Cell cul-
ture flasks and plates were from TPP (Trasandingen,
Switzerland). [3H]-histamine (525.4 GBq/mmol) was
Neurochem Res
123

purchased from Perkin Elmer (Massachusetts, USA),
RNeasy Kit from Qiagen (Valencia, Canada), High
Capacity cDNA Reverse Transcription Kit, TaqMan Gene
Expression Assays and TaqMan Gene Expression Master
Mix from Applied Biosystems (Carlsbad, CA, USA).
Bradford total protein assay was obtained from Bio-Rad
(Hercules, CA, USA). Decynium 22 (D22), ouabain, flu-
oxetine and corticosterone were from Sigma-Aldrich (St.
Louis, MO, USA), while desipramine and amitriptyline
were purchased from Sandoz (Cham, Switzerland). All
other reagents, unless otherwise specified, were of analyt-
ical grade and from Sigma-Aldrich.
Animal Housing and Care
Experiments were approved by Veterinary Administration
of Republic of Slovenia (34401-1/2010/8). Wistar rats were
maintained on 12-h day/12-h night cycle, received standard
rodent chow and were cared for in accordance with the
national guidelines for the care and use of laboratory ani-
mals. Three-day old neonatal rats were sacrificed for
removal of cerebellum, striatum or cerebral cortex.
Cultured Astrocytes
Astrocyte cultures were prepared, as described [12, 33, 34].
Briefly, primary cultures derived from cerebellum, striatum
or cerebral cortex, were grown in high-glucose Dulbecco’s
modified Eagle’s medium (DMEM), containing 10 % FBS,
1 mM pyruvate, 2 mM glutamine, and 25 lg/ml gentamy-
cin at 37 �C in humidified 95 % air/5 % CO2. To reduce the
number of contaminating cells, confluent cultures were
exposed to overnight shaking at 225 rpm. Medium con-
taining detached cells was removed the next morning and
fresh growth medium was added. The whole procedure was
repeated three times. After the third overnight shaking,
adherent cells were trypsinized and subsequently cultured
in the presence of 10 lM cytosine arabinoside for 24 h.
Upon reaching confluence, the cells were seeded into
12-well or 6-well plates and were grown for additional
3 weeks before being used for histamine uptake or real-time
PCR experiments, respectively. Cultures used for all
experiments contained 93–100 % type 1 astrocytes [33, 34].
Histamine Uptake in Cultured Astrocytes
Monolayer cultures in 12-well plates were preincubated for
30 min in the uptake buffer (25 mM HEPES, 125 mM
NaCl, 4.8 mM KCl, 1.2 mM KH2PO4, 1.2 mM MgSO4,
1.4 mM CaCl2, and 5.6 mM glucose, pH 7.4) at 37 �C
(total uptake) or at 4 �C (non-specific uptake). Cultured
astrocytes were incubated with 125 nM [3H]-histamine for
different time intervals to determine time dependence of
histamine uptake. Concentration dependence of histamine
uptake was determined by exposing cultured astrocytes to
different histamine concentrations, as indicated in the
Results, for 20 min. Experiments at 37 �C were terminated
by placing the plates on ice. [3H]-histamine was quickly
removed and the plates were washed twice with ice-cold
uptake buffer. The cells were subsequently lyzed in 300 ll
0.5 M NaOH. An aliquot (250 ll) of each sample was
transferred to a scintillation vial to measure the radioac-
tivity. The amount of transported histamine is normalized
to total protein content, which was determined in the
remaining aliquots (50 ll) of each sample with the Brad-
ford protein assay.
The Specific Histamine Uptake Rate and its Kinetics
The specific (temperature-sensitive) histamine uptake was
calculated as the difference between the total (37 �C) and
the non-specific histamine uptake (4 �C). Specific hista-
mine uptake rate data were linearly transformed and fitted
using linear regression to obtain Woolf, Eadie-Hofstee and
Lineweaver–Burk plots, which were used to determine the
apparent Michaelis–Menten constant (Km), the maximal
uptake rate (Vmax) and transport efficiency (Vmax/Km).
Kinetic parameters are reported as means with 95 % con-
fidence intervals.
Cell Treatments
To determine sensitivity of histamine transport to different
uptake (corticosterone, D22, antidepressants) or Na?/K?-
ATPase (ouabain) inhibitors, cultured astrocytes were first
preincubated in the uptake buffer for 30 min, followed by
incubation in the presence or absence of an inhibitor for
20 min at 37 �C. Astrocytes were then exposed to 125 nM
[3H]-histamine in the absence or presence of uptake or
Na?/K?-ATPase inhibitors for 20 min. To assess whether
histamine uptake is Na?-dependent, cultured astrocytes
were preincubated in the normal (Na?-containing) or Na?-
free uptake buffer, containing 125 mM choline chloride
((CH3)3N(Cl)CH2CH2OH) for 30 min. Astrocytes were
subsequently incubated with 125 nM [3H]-histamine in the
presence or absence of Na? for 20 min. Samples were
harvested and processed for radioactivity and protein
measurements as described above.
OCT mRNA Expression
The total RNA from rat cerebral cortex, neonatal kidney,
and cultured astrocytes was extracted using RNeasy kit
(Qiagen, Valencia, CA, U.S.A.) according to the manu-
facturer’s instructions. The RNA quantity and purity of
isolated total RNA was estimated by measuring the
Neurochem Res
123

absorbance at 260 and 280 nm with the Nanodrop 2000c
spectrophotometer, Thermo Fischer Scientific (Waltham,
MA, USA). cDNA was synthesized from 2 lg of total
RNA using High Capacity cDNA Reverse Transcription
Kit, Applied Biosystems (Carlsbad, CA, USA). Quantita-
tive real-time PCR was performed in the 96-well format,
using an ABI PRISM SDS 7500 sequence detection system
from the Applied Biosystems (Carlsbad, CA, USA). Taq-
Man gene expression assays were used to determine
mRNA expression of rat OCT1 (Rn00562250_m1), rat
OCT2 (Rn00580893_m1), rat OCT3 (Rn00570264_m1)
and rat beta actin (rACTB, 4352931E), which was used as
the endogenous control (reference gene). Samples were
analyzed in duplicates. 20 ll reaction mixture contained
10 ll TaqMan Gene Expression Master Mix, Applied
Biosystems (Carlsbad, CA, USA), 1 ll of the appropriate
Taqman Gene Expression Assay, 7 ll of diethylpyr-
ocarbonate-treated water and 2 ll of cDNA. Amplification
was carried out as follows: 2 min at 55 �C, denaturation for
10 min at 95 �C followed by 40 cycles at 95 �C and 40
cycles at 60 �C. Exponential phase of the amplification
reaction was used to determine the threshold cycle (Ct) for
target genes (OCT mRNA) and endogenous control
(ACTB). OCT mRNA levels were normalized to ACTB
mRNA and gene expression ratios are reported [35]. Brain
cortex and kidney from neonatal rats were used to validate
OCT gene expression assays.
Statistics
Results are presented as mean ± SEM. The kinetic
parameters Km, Vmax and transport efficiency (Vmax/Km)
were determined from Woolf, Eadie-Hofstee and Linewe-
aver–Burk plots using software Prism4 version 4.00
(GraphPad Software Inc., San Diego, USA). Comparisons
between groups were performed using Student’s t test or
one-way ANOVA, followed by Dunnett’s or Bonferroni’s
post hoc test, when several groups were compared. Sta-
tistical significance was established at P \ 0.05.
Results
Cerebellar and Striatal Astrocytes Display
Time-Dependent and Temperature-Sensitive
Histamine Uptake
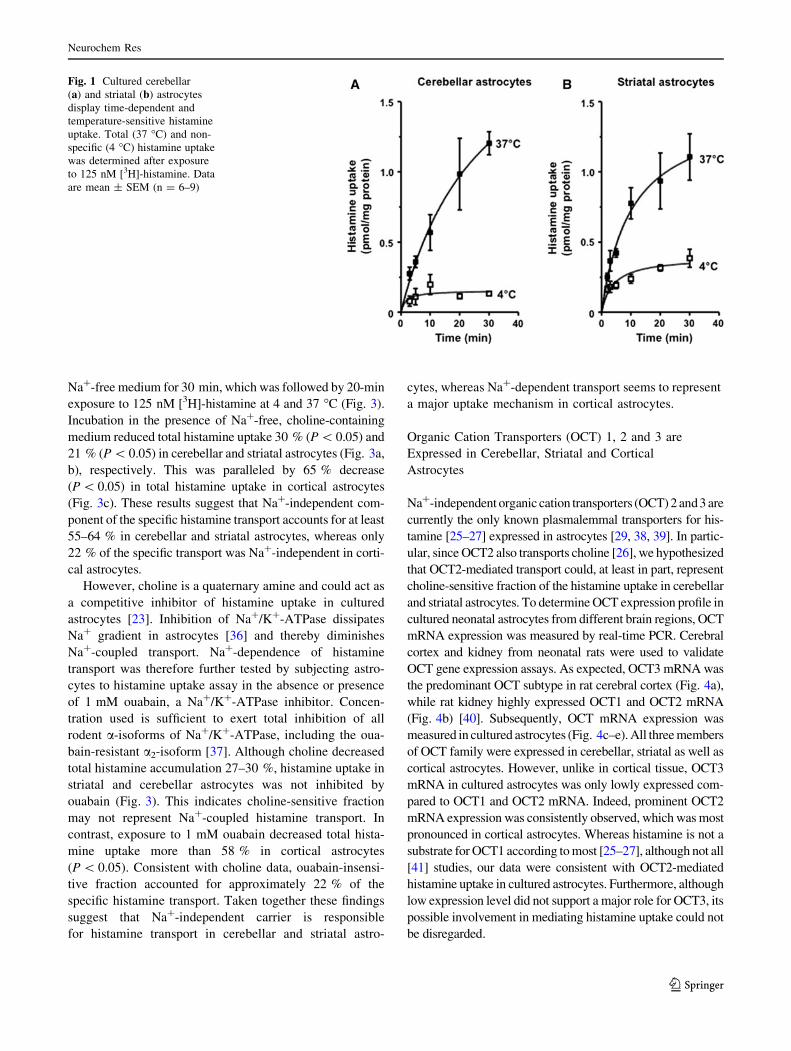
First of all, we determined whether carrier-mediated hista-
mine uptake exists in cultured astrocytes from neonatal rat
cerebellum and striatum. Astrocytes were exposed to 125 nM
[3H]-histamine for 2–30 min at 4 or 37 �C and the non-spe-
cific and total histamine uptake, respectively, were measured
(Fig. 1). Histamine accumulated in a time-dependent manner
and was clearly temperature-sensitive, as histamine uptake
decreased 70–85 % during exposure to 4 �C. Overall, these
data indicate that cultured cerebellar and striatal astrocytes
possess temperature-sensitive, carrier-mediated histamine
uptake.
Cerebellar, Striatal and Cortical Astrocytes Display
Differences in Histamine Uptake Characteristics
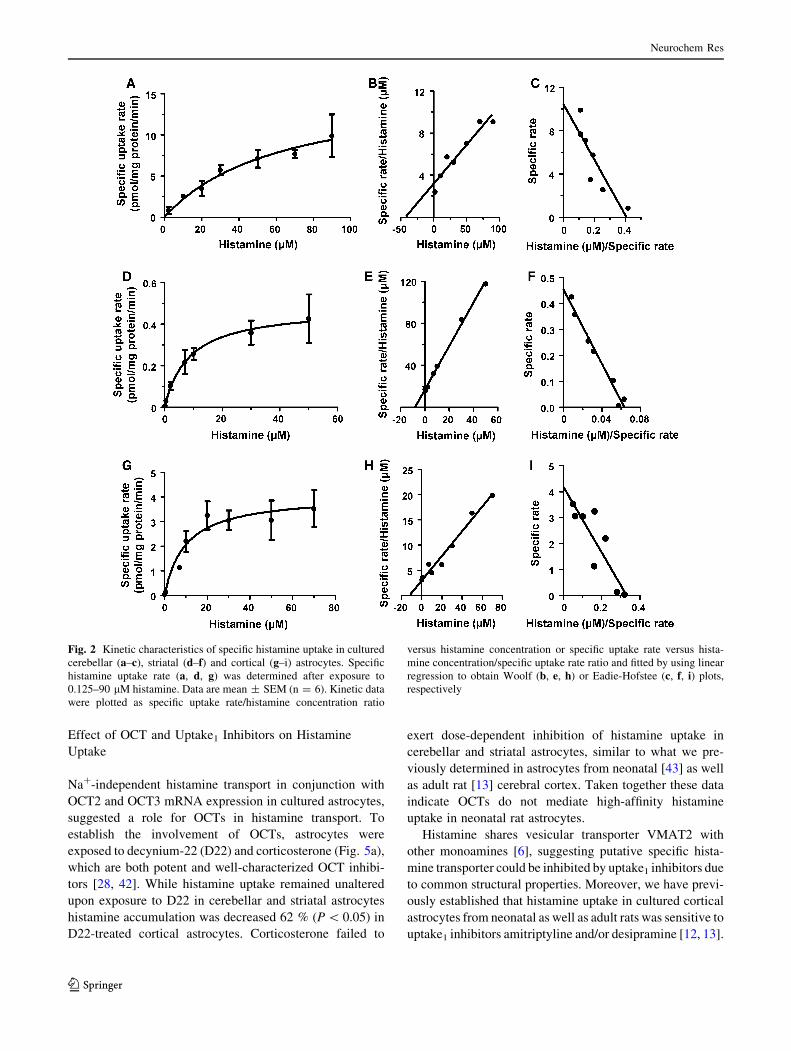
Next we evaluated concentration-dependence and kinetics of
histamine transport in striatal and cerebellar astrocytes. To
investigate whether regional differences in basic characteris-
tics of histamine transport exist, histamine uptake in cortical
astrocytes was assessed in parallel. We have previously
reported some basic properties of histamine transport in
neonatal cortical astrocytes [12, 14], here we provide further
and more detailed kinetic data. Astrocytes were incubated
with 0.125–90 lM histamine for 20 min and the specific
histamine uptake rate (Fig. 2) was determined from the spe-
cific, temperature-sensitive histamine uptake which was cal-
culated as the difference between the total (37 �C) and the
non-specific (4 �C) uptake. Uptake rate data were linearly
transformed and Woolf, Eadie-Hofstee (Fig. 2) and Linewe-
aver–Burk (not shown) plots were used to determine kinetic
parameters (Table 1). Saturable, concentration-dependent
histamine uptake was detected in cerebellar, striatal and, as
previously reported [14], cortical astrocytes. In comparison
with striatum or cortex, histamine uptake in cerebellar
astrocytes had the highest capacity (Vmax = 8.42-13.5 pmol/
mg protein/min) but somewhat lower affinity (Km =
18.5–43.1 lM). Transport efficiency (Vmax/Km) in cerebellar
astrocytes (0.31–0.46 ll/mg protein/min) was comparable to
histamine uptake in astrocytes from cerebral cortex. In con-
trast, striatal astrocytes were characterized by the highest
affinity for histamine (Km = 6.35–8.23 lM), but markedly
lower uptake efficiency (0.06–0.07 ll/mg protein/min). In
sum, these data demonstrate that cultured astrocytes from
neonatal cerebellum possess histamine uptake with moderate
affinity and comparatively high capacity, whereas striatal
astrocytes display high-affinity, low-capacity histamine
transport.
Histamine Uptake is Ouabain-Sensitive in Cortical,
but Not in Cerebellar and Striatal Astrocytes
Monoamine uptake is mediated by the specific, high-affinity/
low-capacity, Na?/Cl--dependent transporters (uptake1) [3]
as well as by the non-specific, low-affinity/high-capacity,
Na?/Cl--independent carriers (uptake2), which are thought to
represent a major histamine uptake system [17, 23, 27, 28]. To
determine, whether histamine uptake is Na?-dependent,
sodium chloride in the uptake buffer was substituted by
125 mM choline chloride. Astrocytes were preincubated in
Neurochem Res
123
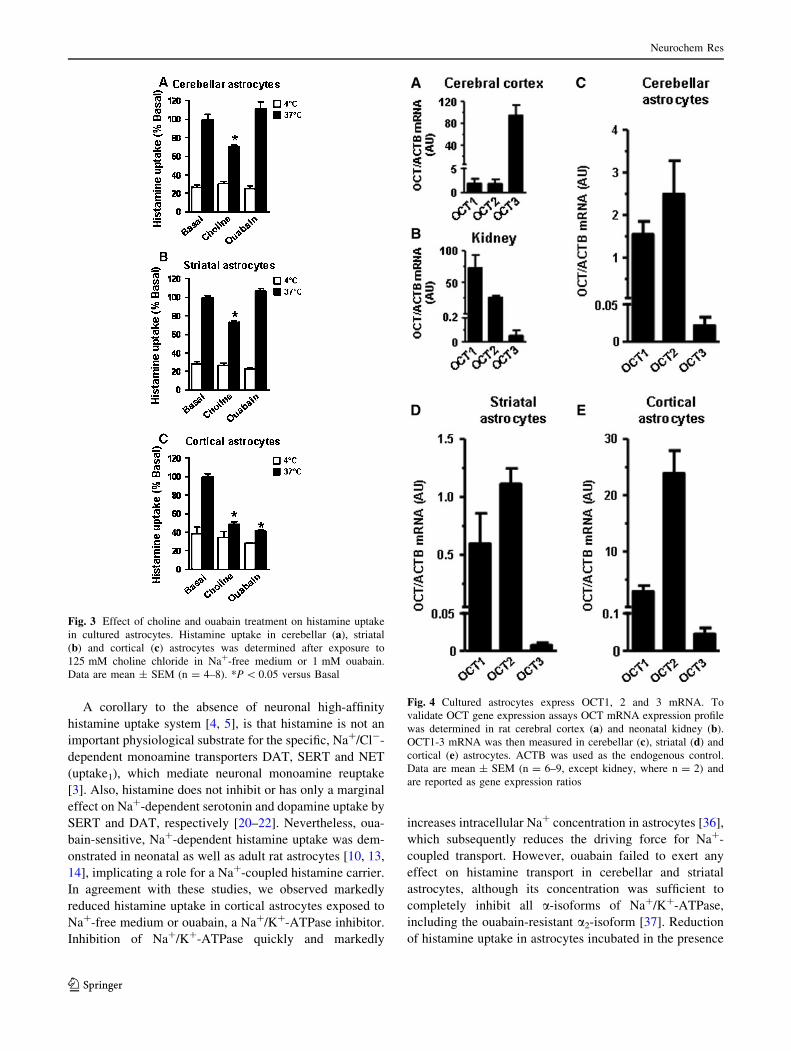
Na?-free medium for 30 min, which was followed by 20-min
exposure to 125 nM [3H]-histamine at 4 and 37 �C (Fig. 3).
Incubation in the presence of Na?-free, choline-containing
medium reduced total histamine uptake 30 % (P \ 0.05) and
21 % (P \ 0.05) in cerebellar and striatal astrocytes (Fig. 3a,
b), respectively. This was paralleled by 65 % decrease
(P \ 0.05) in total histamine uptake in cortical astrocytes
(Fig. 3c). These results suggest that Na?-independent com-
ponent of the specific histamine transport accounts for at least
55–64 % in cerebellar and striatal astrocytes, whereas only
22 % of the specific transport was Na?-independent in corti-
cal astrocytes.
However, choline is a quaternary amine and could act as
a competitive inhibitor of histamine uptake in cultured
astrocytes [23]. Inhibition of Na?/K?-ATPase dissipates
Na? gradient in astrocytes [36] and thereby diminishes
Na?-coupled transport. Na?-dependence of histamine
transport was therefore further tested by subjecting astro-
cytes to histamine uptake assay in the absence or presence
of 1 mM ouabain, a Na?/K?-ATPase inhibitor. Concen-
tration used is sufficient to exert total inhibition of all
rodent a-isoforms of Na?/K?-ATPase, including the oua-
bain-resistant a2-isoform [37]. Although choline decreased
total histamine accumulation 27–30 %, histamine uptake in
striatal and cerebellar astrocytes was not inhibited by
ouabain (Fig. 3). This indicates choline-sensitive fraction
may not represent Na?-coupled histamine transport. In
contrast, exposure to 1 mM ouabain decreased total hista-
mine uptake more than 58 % in cortical astrocytes
(P \ 0.05). Consistent with choline data, ouabain-insensi-
tive fraction accounted for approximately 22 % of the
specific histamine transport. Taken together these findings
suggest that Na?-independent carrier is responsible
for histamine transport in cerebellar and striatal astro-
cytes, whereas Na?-dependent transport seems to represent
a major uptake mechanism in cortical astrocytes.
Organic Cation Transporters (OCT) 1, 2 and 3 are
Expressed in Cerebellar, Striatal and Cortical
Astrocytes
Na?-independent organic cation transporters (OCT) 2 and3 are
currently the only known plasmalemmal transporters for his-
tamine [25–27] expressed in astrocytes [29, 38, 39]. In partic-
ular, since OCT2 also transports choline [26], we hypothesized
that OCT2-mediated transport could, at least in part, represent
choline-sensitive fraction of the histamine uptake in cerebellar
and striatal astrocytes. To determine OCT expression profile in
cultured neonatal astrocytes from different brain regions, OCT
mRNA expression was measured by real-time PCR. Cerebral
cortex and kidney from neonatal rats were used to validate
OCT gene expression assays. As expected, OCT3 mRNA was
the predominant OCT subtype in rat cerebral cortex (Fig. 4a),
while rat kidney highly expressed OCT1 and OCT2 mRNA
(Fig. 4b) [40]. Subsequently, OCT mRNA expression was
measured in cultured astrocytes (Fig. 4c–e). All three members
of OCT family were expressed in cerebellar, striatal as well as
cortical astrocytes. However, unlike in cortical tissue, OCT3
mRNA in cultured astrocytes was only lowly expressed com-
pared to OCT1 and OCT2 mRNA. Indeed, prominent OCT2
mRNA expression was consistently observed, which was most
pronounced in cortical astrocytes. Whereas histamine is not a
substrate for OCT1 according to most [25–27], although not all
[41] studies, our data were consistent with OCT2-mediated
histamine uptake in cultured astrocytes. Furthermore, although
low expression level did not support a major role for OCT3, its
possible involvement in mediating histamine uptake could not
be disregarded.
Fig. 1 Cultured cerebellar
(a) and striatal (b) astrocytes
display time-dependent and
temperature-sensitive histamine
uptake. Total (37 �C) and non-
specific (4 �C) histamine uptake
was determined after exposure
to 125 nM [3H]-histamine. Data
are mean ± SEM (n = 6–9)
Neurochem Res
123
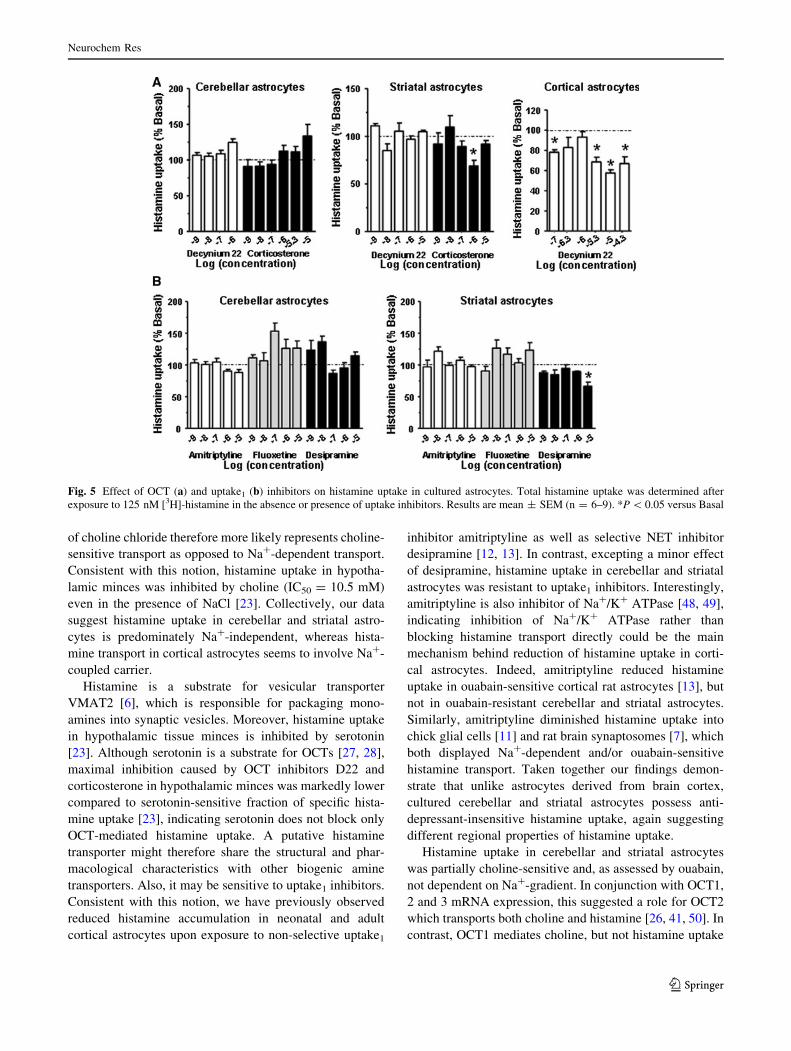
Effect of OCT and Uptake1 Inhibitors on Histamine
Uptake
Na?-independent histamine transport in conjunction with
OCT2 and OCT3 mRNA expression in cultured astrocytes,
suggested a role for OCTs in histamine transport. To
establish the involvement of OCTs, astrocytes were
exposed to decynium-22 (D22) and corticosterone (Fig. 5a),
which are both potent and well-characterized OCT inhibi-
tors [28, 42]. While histamine uptake remained unaltered
upon exposure to D22 in cerebellar and striatal astrocytes
histamine accumulation was decreased 62 % (P \ 0.05) in
D22-treated cortical astrocytes. Corticosterone failed to
exert dose-dependent inhibition of histamine uptake in
cerebellar and striatal astrocytes, similar to what we pre-
viously determined in astrocytes from neonatal [43] as well
as adult rat [13] cerebral cortex. Taken together these data
indicate OCTs do not mediate high-affinity histamine
uptake in neonatal rat astrocytes.
Histamine shares vesicular transporter VMAT2 with
other monoamines [6], suggesting putative specific hista-
mine transporter could be inhibited by uptake1 inhibitors due
to common structural properties. Moreover, we have previ-
ously established that histamine uptake in cultured cortical
astrocytes from neonatal as well as adult rats was sensitive to
uptake1 inhibitors amitriptyline and/or desipramine [12, 13].
Fig. 2 Kinetic characteristics of specific histamine uptake in cultured
cerebellar (a–c), striatal (d–f) and cortical (g–i) astrocytes. Specific
histamine uptake rate (a, d, g) was determined after exposure to
0.125–90 lM histamine. Data are mean ± SEM (n = 6). Kinetic data
were plotted as specific uptake rate/histamine concentration ratio
versus histamine concentration or specific uptake rate versus hista-
mine concentration/specific uptake rate ratio and fitted by using linear
regression to obtain Woolf (b, e, h) or Eadie-Hofstee (c, f, i) plots,
respectively
Neurochem Res
123

Histamine uptake in cerebellar and striatal astrocytes was
therefore measured in the absence or presence of amitripty-
line (non-selective monoamine reuptake inhibitor), fluoxe-
tine (selective serotonin reuptake inhibitor) and desipramine
(selective noradrenaline reuptake inhibitor) (Fig. 5b).
Whereas desipramine showed some inhibitory activity in
striatal astrocytes, amitriptyline and fluoxetine did not alter
histamine uptake neither in cerebellar nor striatal astrocytes.
Overall these results demonstrate that histamine transport
in striatal and cerebellar astrocytes is resistant to anti-
depressants.
Discussion
High-affinity uptake of serotonin, noradrenaline and
dopamine in neurons plays a key role in regulation of
monoaminergic neurotransmission and represents an
important target for pharmacotherapy of neuropsychiatric
disorders [3]. In contrast, histamine is the only monoamine
neurotransmitter lacking high-affinity neuronal uptake [4,
5]. Astrocytes may therefore represent a major site of
histamine uptake in the brain [9, 44, 45]. However, most
previous studies have focused on histamine uptake in
cortical or hypothalamic astrocytes [10–14], although his-
taminergic system spreads throughout the entire central
nervous system with pronounced regional variation in the
anatomical and functional characteristics [1, 4, 46]. In
addition, while dysfunction of histamine transport may be
associated with pathological alterations of local histamine
concentration in specific brain regions such as striatum [1,
47], whether and how astrocytes from different brain
regions participate in histamine uptake has not been thor-
oughly explored. Here we provide evidence that carrier-
mediated histamine transport exists in cerebellar and
striatal astrocytes. In addition, our data suggest regional
differences in characteristics of astroglial histamine uptake.
Neurons lack high-affinity histamine uptake [4, 5].
Furthermore, some synaptosomal studies even failed to
demonstrate saturable histamine transport [8]. In direct
contrast, we detected high-affinity, concentration-depen-
dent histamine uptake in cultured cerebellar and striatal
astrocytes, implicating a role for astrocytes in regulation of
extracellular histamine concentration and histaminergic
neurotransmission in these regions. Since high-affinity
histamine carrier has not been identified, low-affinity, high-
capacity organic cation transporters OCT2 and especially
OCT3 are often considered a major histamine uptake sys-
tem in the brain [17, 23, 31]. However, not only OCT
inhibitors failed to depress histamine uptake in cerebellar
and striatal astrocytes, also its kinetic properties are
markedly different from those reported for OCT2 and
OCT3. In particular, Km values (0.54–0.89 mM) for OCT-
mediated histamine transport [27, 28] are 10- to 100-fold
higher compared to what we observed in cerebellar
(18.5–43.1 lM) and striatal (6.4–8.2 lM) astrocytes.
These findings are consistent with the existence of high-
affinity histamine uptake in astrocyte-enriched brain frac-
tions as well as in cultured cortical or hypothalamic astro-
cytes, where even lower Km values (\1 lM) were observed
[9, 10]. Although transport efficiencies (0.05–0.46 ll/mg
protein/min), especially in astrocytes derived from stria-
tum, were lower than previously reported for rat astro-
cytes (2.25–16.42 ll/mg/min) [10, 12], they were still
significantly higher compared to efficiency of histamine
uptake in rat brain synaptosomes (0.015 and 0.0028 ll/
mg/min for the high and low affinity uptake process,
respectively) [7]. Taken together our results suggest
astrocytes in cerebellum and striatum possess high-
affinity histamine uptake system.
Table 1 Kinetic parameters of specific histamine uptake in neonatal cerebellar, striatal and cortical astrocytes
Cell type Km (CI) (lM) Vmax (CI) (pmol/mg/min) Vmax/Km (CI) (ll/mg protein/min)
Cerebellar astrocytes 43.1 (20.0–89.1)1
25.8 (9.14–42.5)2
18.5 (11.9–34.6)3
13.5 (10.0–20.8)1
10.5 (6.76–14.2)2
8.42 (5.90–14.7)3
0.313 (0.224–0.520)1
0.406 (0.310–0.798)2
0.456 (0.407–0.518)3
Striatal astrocytes 8.23 (6.16–10.6)1
7.21 (5.82–8.59)2
6.35 (5.69–7.18)3
0.48 (0.45–0.52)1
0.46 (0.40–0.51)2
0.43 (0.39–0.48)3
0.059 (0.049–0.074)1
0.063 (0.058–0.071)2
0.068 (0.067–0.069)3
Cortical astrocytes 12.3 (5.84–20.9)1
12.6 (5.65–19.6)2
9.65 (5.11–70.6)3
4.11 (3.51–4.96)1
4.19 (2.84–5.54)2
3.11 (1.67–22.5)3
0.334 (0.228–0.623)1
0.331 (0.260–0.546)2
0.322 (0.313–0.332)3
Vmax: maximal specific uptake rate, CI: 95 % confidence interval1 Woolf plot2 Eadie-Hofstee plot3 Lineweaver-Burk plot, Km: Michaelis–Menten constant
Neurochem Res
123
A corollary to the absence of neuronal high-affinity
histamine uptake system [4, 5], is that histamine is not an
important physiological substrate for the specific, Na?/Cl--
dependent monoamine transporters DAT, SERT and NET
(uptake1), which mediate neuronal monoamine reuptake
[3]. Also, histamine does not inhibit or has only a marginal
effect on Na?-dependent serotonin and dopamine uptake by
SERT and DAT, respectively [20–22]. Nevertheless, oua-
bain-sensitive, Na?-dependent histamine uptake was dem-
onstrated in neonatal as well as adult rat astrocytes [10, 13,
14], implicating a role for a Na?-coupled histamine carrier.
In agreement with these studies, we observed markedly
reduced histamine uptake in cortical astrocytes exposed to
Na?-free medium or ouabain, a Na?/K?-ATPase inhibitor.
Inhibition of Na?/K?-ATPase quickly and markedly
increases intracellular Na? concentration in astrocytes [36],
which subsequently reduces the driving force for Na?-
coupled transport. However, ouabain failed to exert any
effect on histamine transport in cerebellar and striatal
astrocytes, although its concentration was sufficient to
completely inhibit all a-isoforms of Na?/K?-ATPase,
including the ouabain-resistant a2-isoform [37]. Reduction
of histamine uptake in astrocytes incubated in the presence
Fig. 3 Effect of choline and ouabain treatment on histamine uptake
in cultured astrocytes. Histamine uptake in cerebellar (a), striatal
(b) and cortical (c) astrocytes was determined after exposure to
125 mM choline chloride in Na?-free medium or 1 mM ouabain.
Data are mean ± SEM (n = 4–8). *P \ 0.05 versus Basal
Fig. 4 Cultured astrocytes express OCT1, 2 and 3 mRNA. To
validate OCT gene expression assays OCT mRNA expression profile
was determined in rat cerebral cortex (a) and neonatal kidney (b).
OCT1-3 mRNA was then measured in cerebellar (c), striatal (d) and
cortical (e) astrocytes. ACTB was used as the endogenous control.
Data are mean ± SEM (n = 6–9, except kidney, where n = 2) and
are reported as gene expression ratios
Neurochem Res
123
of choline chloride therefore more likely represents choline-
sensitive transport as opposed to Na?-dependent transport.
Consistent with this notion, histamine uptake in hypotha-
lamic minces was inhibited by choline (IC50 = 10.5 mM)
even in the presence of NaCl [23]. Collectively, our data
suggest histamine uptake in cerebellar and striatal astro-
cytes is predominately Na?-independent, whereas hista-
mine transport in cortical astrocytes seems to involve Na?-
coupled carrier.
Histamine is a substrate for vesicular transporter
VMAT2 [6], which is responsible for packaging mono-
amines into synaptic vesicles. Moreover, histamine uptake
in hypothalamic tissue minces is inhibited by serotonin
[23]. Although serotonin is a substrate for OCTs [27, 28],
maximal inhibition caused by OCT inhibitors D22 and
corticosterone in hypothalamic minces was markedly lower
compared to serotonin-sensitive fraction of specific hista-
mine uptake [23], indicating serotonin does not block only
OCT-mediated histamine uptake. A putative histamine
transporter might therefore share the structural and phar-
macological characteristics with other biogenic amine
transporters. Also, it may be sensitive to uptake1 inhibitors.
Consistent with this notion, we have previously observed
reduced histamine accumulation in neonatal and adult
cortical astrocytes upon exposure to non-selective uptake1
inhibitor amitriptyline as well as selective NET inhibitor
desipramine [12, 13]. In contrast, excepting a minor effect
of desipramine, histamine uptake in cerebellar and striatal
astrocytes was resistant to uptake1 inhibitors. Interestingly,
amitriptyline is also inhibitor of Na?/K? ATPase [48, 49],
indicating inhibition of Na?/K? ATPase rather than
blocking histamine transport directly could be the main
mechanism behind reduction of histamine uptake in corti-
cal astrocytes. Indeed, amitriptyline reduced histamine
uptake in ouabain-sensitive cortical rat astrocytes [13], but
not in ouabain-resistant cerebellar and striatal astrocytes.
Similarly, amitriptyline diminished histamine uptake into
chick glial cells [11] and rat brain synaptosomes [7], which
both displayed Na?-dependent and/or ouabain-sensitive
histamine transport. Taken together our findings demon-
strate that unlike astrocytes derived from brain cortex,
cultured cerebellar and striatal astrocytes possess anti-
depressant-insensitive histamine uptake, again suggesting
different regional properties of histamine uptake.
Histamine uptake in cerebellar and striatal astrocytes
was partially choline-sensitive and, as assessed by ouabain,
not dependent on Na?-gradient. In conjunction with OCT1,
2 and 3 mRNA expression, this suggested a role for OCT2
which transports both choline and histamine [26, 41, 50]. In
contrast, OCT1 mediates choline, but not histamine uptake
Fig. 5 Effect of OCT (a) and uptake1 (b) inhibitors on histamine uptake in cultured astrocytes. Total histamine uptake was determined after
exposure to 125 nM [3H]-histamine in the absence or presence of uptake inhibitors. Results are mean ± SEM (n = 6–9). *P \ 0.05 versus Basal
Neurochem Res
123

[25–27, 51]. On the other hand, while OCT3 could con-
tribute to histamine uptake [27, 28], its lowly expression
did not indicate a major role for OCT3-mediated transport.
However, selective OCT inhibitors D22 and corticosterone
did not alter histamine uptake in cerebellar and striatal
astrocytes. Furthermore, OCTs are electrogenic, potential-
sensitive transporters [26]. Depolarization due to inhibition
of Na?/K?-ATPase by ouabain [37] could therefore be
expected to affect uptake of histamine, which is positively
charged under physiological conditions [14]. These results
essentially precluded major involvement of OCTs at least
in histamine concentration range tested in this study, which
was well below Km value for OCT2 and OCT3 [27, 28].
Consistent with our findings, histamine concentration in
cerebellum remained similar between OCT3 knock-out
mice and controls [32]. Similarly, histamine concentration
in striatum of OCT3-deficient mice was unaltered com-
pared with wild type controls under basal conditions as
well as during post-ischemic reperfusion [31]. Collectively,
these data indicate OCT2 and OCT3 do not mediate high-
affinity histamine uptake observed in cultured cerebellar
and striatal astrocytes.
Another interesting candidate for histamine transporter
in the brain is plasma membrane monoamine transporter
(PMAT), a Na?/Cl--independent, potential-sensitive non-
specific organic cation transporter [28, 52]. PMAT medi-
ates low-affinity histamine uptake and is sensitive to D22
and choline, but is not inhibited by corticosterone [28, 52].
Our pharmacological data in cortical astrocytes are there-
fore consistent with possible involvement of PMAT.
However, whereas PMAT is expressed in human astrocy-
toma cells [53], its expression in rodent astroglia has not
been demonstrated [38, 54], indicating PMAT does not
underlie histamine uptake in cultured rat astrocytes.
Although other non-specific organic cation carriers might
be involved in histamine uptake, it is difficult to speculate
on their identity since PMAT and OCTs are the only
known histamine carriers aside from the vesicular trans-
porter VMAT2. Nevertheless, our data suggest that hista-
mine and choline might share transporters other than
OCT2. This idea is tangible since a substantial fraction of
choline-sensitive histamine uptake in hypothalamic minces
appears to be resistant to OCT inhibitors [23]. Interest-
ingly, astrocytes express carnitine/organic cation trans-
porter (OCTN) 2 and choline transporter-like protein 1
(CTL1) [55, 56], which can mediate Na?-independent
uptake of a range of organic cations, including choline and
several OCT substrates. Also, transport of some organic
cations by OCTN2 is potential-insensitive, which resem-
bles our observations in ouabain-treated striatal and cere-
bellar astrocytes. Furthermore, CTL1-mediated transport is
sensitive to D22 and desipramine, but is not inhibited by
corticosterone [56], which mimics properties of histamine
uptake in cortical astrocytes. Despite these similarities it
remains to be seen if OCTN2 and/or CTL1 could play a
role in histamine uptake, not least because histamine does
not depress OCTN2-mediated uptake of carnitine, its pri-
mary substrate [57].
Cell culture represents an artificial milieu, where cul-
tured astrocytes are deprived of normal environmental
cues. Moreover, differences in media compositions or sera
used may directly affect phenotypic properties of cultured
astrocytes [58], including expression of neurotransmitter
carriers [59]. For example, cell culture factors provide one
possible reason why OCT mRNA expression pattern was
different between brain cortex and cultured cortical astro-
cytes. On the other hand, observed differences in OCT
mRNA expression may simply reflect the fact that cortical
homogenates contain heterogeneous cell population. As
well as astrocytes, brain tissue homogenates contain neu-
rons, microglia, oligodendroglia and endothelial cells,
while we investigated OCT expression and histamine
uptake in primary astrocyte cultures. Importantly, pro-
nounced regional differences in OCT distribution between
neurons and astrocytes have been described [29]. Further-
more, primary astrocytes in cell culture seem to retain
biological characteristics related to their area of origin.
Indeed, variations in uptake of serotonin and glutamate in
cultured astrocytes derived from different brain regions
corresponded to regional differences under in vivo condi-
tions [60]. Nevertheless, only limited extrapolations are
possible in our experimental model owing to limited
knowledge of regional variation in histamine uptake in
neonatal rat brain. Also, since ontogenetic development
affects organization of histaminergic system and brain
histamine levels in brain [61], cultured neonatal astrocyte
may not reflect the situation in adult rats. Notwithstanding
these caveats, its tempting to speculate that high-capacity,
intermediate-affinity histamine uptake in cerebellar astro-
cytes may reflect low histamine turnover rate, an indication
of low activity of histaminergic neurons, in cerebellum
[62–64]. In contrast, markedly higher histamine turnover in
striatum [62–64] may necessitate high-affinity uptake for
timely termination of histaminergic neurotransmission.
To summarize, we have demonstrated that astrocytes
derived from cerebellum and striatum possess intermediate-
or high-affinity, saturable histamine uptake. Furthermore, at
least two high-affinity transporters appear to be involved in
astrocytes derived from different brain regions. Na?-inde-
pendent/choline-sensitive carrier predominates in cerebellar
and striatal astrocytes, whereas Na?-dependent transporter
underlies histamine uptake in cortical astrocytes. Collec-
tively, our data therefore indicate existence of regional
variations in characteristics of astrocyte histamine uptake.
These observations allow us to predict subtle differences
in the fine tuning of histaminergic neurotransmission in
Neurochem Res
123

different neonatal rat brain regions. Moreover, our
findings implicate a role for histamine transporters in
regulation of extracellular histamine concentration in
cerebellum and striatum. Inhibition of histamine uptake
might therefore represent a viable option to modulate
histaminergic neurotransmission.
Acknowledgments This work was supported by the research grant
P3-067, J1-2014 of Ministry of Higher Education, Science and
Technology, Republic of Slovenia. We greatly appreciate technical
assistance of Mrs Jozica Kosir.
Conflict of interest The authors declare that they have no conflict
of interest.
References
1. Haas HL, Sergeeva OA, Selbach O (2008) Histamine in the
nervous system. Physiol Rev 88(3):1183–1241. doi:10.1152/
physrev.00043.2007
2. Passani MB, Blandina P (2011) Histamine receptors in the CNS
as targets for therapeutic intervention. Trends Pharmacol Sci
32(4):242–249. doi:10.1016/j.tips.2011.01.003
3. Iversen L (2006) Neurotransmitter transporters and their impact
on the development of psychopharmacology. Br J Pharmacol
147(Suppl 1):S82–S88. doi:10.1038/sj.bjp.0706428
4. Schwartz JC, Arrang JM, Garbarg M, Pollard H, Ruat M (1991)
Histaminergic transmission in the mammalian brain. Physiol Rev
71(1):1–51
5. Haas H, Panula P (2003) The role of histamine and the tubero-
mamillary nucleus in the nervous system. Nat Rev Neurosci
4(2):121–130. doi:10.1038/nrn1034
6. Merickel A, Edwards RH (1995) Transport of histamine by
vesicular monoamine transporter-2. Neuropharmacology 34(11):
1543–1547
7. Sakurai E, Oreland L, Nishiyama S, Kato M, Watanabe T, Yanai
K (2006) Evidence for the presence of histamine uptake into the
synaptosomes of rat brain. Pharmacology 78(2):72–80. doi:
10.1159/000095637
8. Barnes WG, Hough LB (2002) Membrane-bound histamine N-
methyltransferase in mouse brain: possible role in the synaptic
inactivation of neuronal histamine. J Neurochem 82(5):1262–1271
9. Rafalowska U, Waskiewicz J, Albrecht J (1987) Is neurotrans-
mitter histamine predominantly inactivated in astrocytes? Neu-
rosci Lett 80(1):106–110
10. Huszti Z, Imrik P, Madarasz E (1994) [3H]histamine uptake and
release by astrocytes from rat brain: effects of sodium depriva-
tion, high potassium, and potassium channel blockers. Neuro-
chem Res 19(10):1249–1256
11. Huszti Z, Rimanoczy A, Juhasz A, Magyar K (1990) Uptake,
metabolism, and release of [3H]-histamine by glial cells in pri-
mary cultures of chicken cerebral hemispheres. Glia 3(3):
159–168. doi:10.1002/glia.440030303
12. Osredkar D, Burnik-Papler T, Pecavar B, Kralj-Iglic V, Krzan M
(2009) Kinetic and pharmacological properties of [(3)H]-hista-
mine transport into cultured type 1 astrocytes from neonatal rats.
Inflamm Res 58(2):94–102. doi:10.1007/s00011-009-8103-4
13. Perdan-Pirkmajer K, Pirkmajer S, Cerne K, Krzan M (2012)
Molecular and kinetic characterization of histamine transport into
adult rat cultured astrocytes. Neurochem Int 61(3):415–422. doi:
10.1016/j.neuint.2012.05.002
14. Perdan-Pirkmajer K, Mavri J, Krzan M (2010) Histamine
(re)uptake by astrocytes: an experimental and computational
study. J Mol Model 16(6):1151–1158. doi:10.1007/s00894-
009-0624-9
15. Huszti Z, Magyar K, Kalman M (1990) Contribution of glial cells
to histamine inactivation. Agents Actions 30(1–2):237–239
16. Huszti Z, Prast H, Tran MH, Fischer H, Philippu A (1998) Glial
cells participate in histamine inactivation in vivo. Naunyn
Schmiedebergs Arch Pharmacol 357(1):49–53
17. Baganz NL, Horton RE, Calderon AS, Owens WA, Munn JL,
Watts LT, Koldzic-Zivanovic N, Jeske NA, Koek W, Toney GM,
Daws LC (2008) Organic cation transporter 3: keeping the brake
on extracellular serotonin in serotonin-transporter-deficient mice.
Proc Natl Acad Sci USA 105(48):18976–18981. doi:10.1073/
pnas.0800466105
18. Pickel VM, Chan J (1999) Ultrastructural localization of the
serotonin transporter in limbic and motor compartments of the
nucleus accumbens. J Neurosci 19(17):7356–7366
19. Takeda H, Inazu M, Matsumiya T (2002) Astroglial dopamine
transport is mediated by norepinephrine transporter. Naunyn
Schmiedebergs Arch Pharmacol 366(6):620–623. doi:10.1007/
s00210-002-0640-0
20. Giros B, el Mestikawy S, Bertrand L, Caron MG (1991) Cloning
and functional characterization of a cocaine-sensitive dopamine
transporter. FEBS Lett 295(1–3):149–154
21. Ramamoorthy S, Bauman AL, Moore KR, Han H, Yang-Feng T,
Chang AS, Ganapathy V, Blakely RD (1993) Antidepressant- and
cocaine-sensitive human serotonin transporter: molecular clon-
ing, expression, and chromosomal localization. Proc Natl Acad
Sci USA 90(6):2542–2546
22. Demchyshyn LL, Pristupa ZB, Sugamori KS, Barker EL, Blakely
RD, Wolfgang WJ, Forte MA, Niznik HB (1994) Cloning,
expression, and localization of a chloride-facilitated, cocaine-
sensitive serotonin transporter from Drosophila melanogaster.
Proc Natl Acad Sci USA 91(11):5158–5162
23. Gasser PJ, Lowry CA, Orchinik M (2006) Corticosterone-sensi-
tive monoamine transport in the rat dorsomedial hypothalamus:
potential role for organic cation transporter 3 in stress-induced
modulation of monoaminergic neurotransmission. J Neurosci
26(34):8758–8766. doi:10.1523/JNEUROSCI.0570-06.2006
24. Baganz N, Horton R, Martin K, Holmes A, Daws LC (2010)
Repeated swim impairs serotonin clearance via a corticosterone-
sensitive mechanism: organic cation transporter 3, the smoking
gun. J Neurosci 30(45):15185–15195. doi:10.1523/JNEURO
SCI.2740-10.2010
25. Grundemann D, Liebich G, Kiefer N, Koster S, Schomig E
(1999) Selective substrates for non-neuronal monoamine trans-
porters. Mol Pharmacol 56(1):1–10
26. Schomig E, Lazar A, Grundemann D (2006) Extraneuronal mono-
amine transporter and organic cation transporters 1 and 2: a review of
transport efficiency. Handb Exp Pharmacol 175:151–180
27. Amphoux A, Vialou V, Drescher E, Bruss M, Mannoury La Cour
C, Rochat C, Millan MJ, Giros B, Bonisch H, Gautron S (2006)
Differential pharmacological in vitro properties of organic cation
transporters and regional distribution in rat brain. Neurophar-
macology 50(8):941–952. doi:10.1016/j.neuropharm.2006.01.005
28. Duan H, Wang J (2010) Selective transport of monoamine neu-
rotransmitters by human plasma membrane monoamine trans-
porter and organic cation transporter 3. J Pharmacol Exp Ther
335(3):743–753. doi:10.1124/jpet.110.170142
29. Cui M, Aras R, Christian WV, Rappold PM, Hatwar M, Panza J,
Jackson-Lewis V, Javitch JA, Ballatori N, Przedborski S, Tieu K
(2009) The organic cation transporter-3 is a pivotal modulator of
neurodegeneration in the nigrostriatal dopaminergic pathway.
Proc Natl Acad Sci USA 106(19):8043–8048. doi:10.1073/
pnas.0900358106
Neurochem Res
123

30. Gasser PJ, Orchinik M, Raju I, Lowry CA (2009) Distribution of
organic cation transporter 3, a corticosterone-sensitive mono-
amine transporter, in the rat brain. J Comp Neurol 512(4):
529–555. doi:10.1002/cne.21921
31. Zhu P, Hata R, Ogasawara M, Cao F, Kameda K, Yamauchi K,
Schinkel AH, Maeyama K, Sakanaka M (2012) Targeted dis-
ruption of organic cation transporter 3 (Oct3) ameliorates ische-
mic brain damage through modulating histamine and regulatory T
cells. J Cereb Blood Flow Metab 32(10):1897–1908. doi:10.1038/
jcbfm.2012.92
32. Vialou V, Balasse L, Callebert J, Launay JM, Giros B, Gautron S
(2008) Altered aminergic neurotransmission in the brain of
organic cation transporter 3-deficient mice. J Neurochem 106(3):
1471–1482. doi:10.1111/j.1471-4159.2008.05506.x
33. Krzan M, Schwartz JP (2006) Histamine transport in neonatal and
adult astrocytes. Inflamm Res 55(Suppl 1):S36–S37. doi:10.1007/
s00011-005-0031-3
34. Schwartz JP, Wilson DJ (1992) Preparation and characterization
of type 1 astrocytes cultured from adult rat cortex, cerebellum,
and striatum. Glia 5(1):75–80. doi:10.1002/glia.440050111
35. Tuomi JM, Voorbraak F, Jones DL, Ruijter JM (2010) Bias in the
Cq value observed with hydrolysis probe based quantitative PCR
can be corrected with the estimated PCR efficiency value.
Methods 50(4):313–322. doi:10.1016/j.ymeth.2010.02.003
36. Rose CR, Waxman SG, Ransom BR (1998) Effects of glucose
deprivation, chemical hypoxia, and simulated ischemia on Na?
homeostasis in rat spinal cord astrocytes. J Neurosci 18(10):3554–3562
37. Chibalin AV, Heiny JA, Benziane B, Prokofiev AV, Vasiliev AV,
Kravtsova VV, Krivoi II (2012) Chronic nicotine modifies skel-
etal muscle Na, K-ATPase activity through its interaction with
the nicotinic acetylcholine receptor and phospholemman. PLoS
ONE 7(3):e33719. doi:10.1371/journal.pone.0033719
38. Dahlin A, Xia L, Kong W, Hevner R, Wang J (2007) Expression
and immunolocalization of the plasma membrane monoamine
transporter in the brain. Neuroscience 146(3):1193–1211. doi:
10.1016/j.neuroscience.2007.01.072
39. Daws LC (2009) Unfaithful neurotransmitter transporters: focus
on serotonin uptake and implications for antidepressant efficacy.
Pharmacol Ther 121(1):89–99. doi:10.1016/j.pharmthera.2008.
10.004
40. Wu X, Kekuda R, Huang W, Fei YJ, Leibach FH, Chen J,
Conway SJ, Ganapathy V (1998) Identity of the organic cation
transporter OCT3 as the extraneuronal monoamine transporter
(uptake2) and evidence for the expression of the transporter in the
brain. J Biol Chem 273(49):32776–32786
41. Arndt P, Volk C, Gorboulev V, Budiman T, Popp C, Ulzheimer-
Teuber I, Akhoundova A, Koppatz S, Bamberg E, Nagel G,
Koepsell H (2001) Interaction of cations, anions, and weak base
quinine with rat renal cation transporter rOCT2 compared with
rOCT1. Am J Physiol Renal Physiol 281(3):F454–F468
42. Hayer-Zillgen M, Bruss M, Bonisch H (2002) Expression and
pharmacological profile of the human organic cation transporters
hOCT1, hOCT2 and hOCT3. Br J Pharmacol 136(6):829–836.
doi:10.1038/sj.bjp.0704785
43. Perdan K, Kobe Z, Krzan M (2009) Nature of histamine transport
in neonatal rat cultured type 1 astrocytes–organic cation trans-
porters are not involved. Inflamm Res 58(Suppl 1):32–33. doi:
10.1007/s00011-009-0655-9
44. Huszti Z (1998) Carrier-mediated high affinity uptake system for
histamine in astroglial and cerebral endothelial cells. J Neurosci Res
51(5):551–558. doi:10.1002/(SICI)1097-4547(19980301)51:5\551:
AID-JNR1[3.0.CO;2-E
45. Huszti Z (2003) Histamine uptake into non-neuronal brain cells.
Inflamm Res 52(Suppl 1):S03–S06
46. Giannoni P, Passani MB, Nosi D, Chazot PL, Shenton FC, Medhurst
AD, Munari L, Blandina P (2009) Heterogeneity of histaminergic
neurons in the tuberomammillary nucleus of the rat. Eur J Neurosci
29(12):2363–2374. doi:10.1111/j.1460-9568.2009.06765.x
47. Rinne JO, Anichtchik OV, Eriksson KS, Kaslin J, Tuomisto L,
Kalimo H, Roytta M, Panula P (2002) Increased brain histamine
levels in Parkinson’s disease but not in multiple system atrophy.
J Neurochem 81(5):954–960
48. Carfagna MA, Muhoberac BB (1993) Interaction of tricyclic drug
analogs with synaptic plasma membranes: structure-mechanism
relationships in inhibition of neuronal Na?/K(?)-ATPase activ-
ity. Mol Pharmacol 44(1):129–141
49. Sanganahalli BG, Joshi PG, Joshi NB (2000) Differential effects
of tricyclic antidepressant drugs on membrane dynamics–a fluo-
rescence spectroscopic study. Life Sci 68(1):81–90
50. Sweet DH, Miller DS, Pritchard JB (2001) Ventricular choline trans-
port: a role for organic cation transporter 2 expressed in choroid plexus.
J Biol Chem 276(45):41611–41619. doi:10.1074/jbc.M108472200
51. Koepsell H, Lips K, Volk C (2007) Polyspecific organic cation
transporters: structure, function, physiological roles, and bio-
pharmaceutical implications. Pharm Res 24(7):1227–1251. doi:
10.1007/s11095-007-9254-z
52. Engel K, Wang J (2005) Interaction of organic cations with a
newly identified plasma membrane monoamine transporter. Mol
Pharmacol 68(5):1397–1407. doi:10.1124/mol.105.016832
53. Engel K, Zhou M, Wang J (2004) Identification and characteriza-
tion of a novel monoamine transporter in the human brain. J Biol
Chem 279(48):50042–50049. doi:10.1074/jbc.M407913200
54. Vialou V, Balasse L, Dumas S, Giros B, Gautron S (2007)
Neurochemical characterization of pathways expressing plasma
membrane monoamine transporter in the rat brain. Neuroscience
144(2):616–622. doi:10.1016/j.neuroscience.2006.09.058
55. Inazu M, Takeda H, Maehara K, Miyashita K, Tomoda A,
Matsumiya T (2006) Functional expression of the organic cation/
carnitine transporter 2 in rat astrocytes. J Neurochem
97(2):424–434. doi:10.1111/j.1471-4159.2006.03757.x
56. Inazu M, Takeda H, Matsumiya T (2005) Molecular and func-
tional characterization of an Na ? -independent choline trans-
porter in rat astrocytes. J Neurochem 94(5):1427–1437. doi:
10.1111/j.1471-4159.2005.03299.x
57. Ohashi R, Tamai I, Nezu Ji J, Nikaido H, Hashimoto N, Oku A,
Sai Y, Shimane M, Tsuji A (2001) Molecular and physiological
evidence for multifunctionality of carnitine/organic cation trans-
porter OCTN2. Mol Pharmacol 59(2):358–366
58. Codeluppi S, Gregory EN, Kjell J, Wigerblad G, Olson L,
Svensson CI (2011) Influence of rat substrain and growth con-
ditions on the characteristics of primary cultures of adult rat
spinal cord astrocytes. J Neurosci Methods 197(1):118–127. doi:
10.1016/j.jneumeth.2011.02.011
59. Kimelberg HK, Goderie SK, Conley PA, Higman S, Goldschmidt
R, Amundson RH (1992) Uptake of [3H]serotonin and [3H]glu-
tamate by primary astrocyte cultures. I. Effects of different sera
and time in culture. Glia 6 (1):1–8. doi:10.1002/glia.440060102
60. Amundson RH, Goderie SK, Kimelberg HK (1992) Uptake of
[3H]serotonin and [3H]glutamate by primary astrocyte cultures.
II. Differences in cultures prepared from different brain regions.
Glia 6(1):9–18. doi:10.1002/glia.440060103
61. Molina-Hernandez A, Diaz NF, Arias-Montano JA (2012) His-
tamine in brain development. J Neurochem 122(5):872–882. doi:
10.1111/j.1471-4159.2012.07863.x
62. Nishibori M, Oishi R, Saeki K (1984) Histamine turnover in the
brain of different mammalian species: implications for neuronal
histamine half-life. J Neurochem 43(6):1544–1549
63. Oishi R, Nishibori M, Saeki K (1984) Regional differences in the
turnover of neuronal histamine in the rat brain. Life Sci
34(7):691–699
64. Hough LB, Khandelwal JK, Green JP (1984) Histamine turnover
in regions of rat brain. Brain Res 291(1):103–109
Neurochem Res
123
![Directed establishment phenotypic 1 · Uptake of [3H]GABA. Primary astrocytes have been re-ported to display high-affinity uptake of GABA that is sensitive toinhibition by,B-alanine](https://img.pdfslide.us/doc/110x75/60f92f39b6f8ee3ebb361326/directed-establishment-phenotypic-1-uptake-of-3hgaba-primary-astrocytes-have.jpg)




























