Embed Size (px)
Citation preview
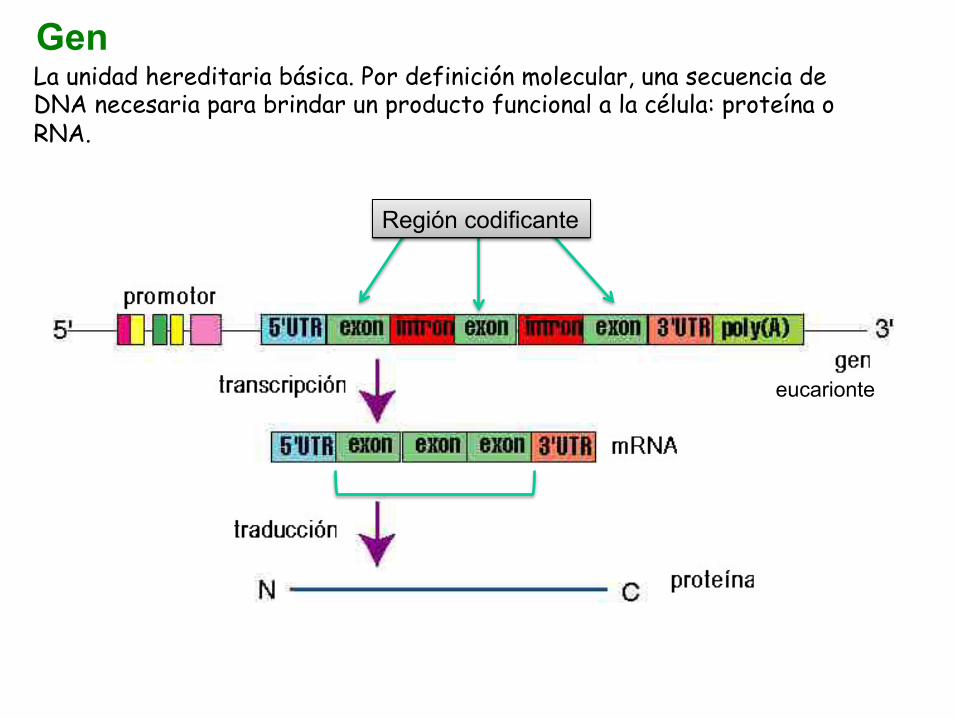
La unidad hereditaria básica. Por definición molecular, una secuencia de DNA necesaria para brindar un producto funcional a la célula: proteína o RNA.
Gen
eucarionte
Región codificante

Alelo La forma alternativa de un gen.
8 Chapter 1 • Genetics and the Organism
the endoplasmic reticulum and Golgi apparatus, and or-ganelles such as mitochondria and chloroplasts. The nu-cleus contains most of the DNA, but note that mitochon-dria and chloroplasts also contain small chromosomes.
Each gene encodes a separate protein, each with spe-cific functions either within the cell (for example, thepurple-rectangle proteins in Figure 1-9) or for export toother parts of the organism (the purple-circle proteins).The synthesis of proteins for export (secretory proteins)takes place on ribosomes that are located on the surfaceof the rough endoplasmic reticulum, a system of large,flattened membrane vesicles. The completed amino acidchains are passed into the lumen of the endoplasmicreticulum, where they fold up spontaneously to take ontheir three-dimensional structure. The proteins may bemodified at this stage, but they eventually enter thechambers of the Golgi apparatus and from there, the se-cretory vessels, which eventually fuse with the cell mem-brane and release their contents to the outside.
Proteins destined to function in the cytoplasm andmost of the proteins that function in mitochondria andchloroplasts are synthesized in the cytoplasm on ribo-somes not bound to membranes. For example, proteinsthat function as enzymes in the glycolysis pathway fol-low this route. The proteins destined for organelles arespecially tagged to target their insertion into specific or-ganelles. In addition, mitochondria and chloroplastshave their own small circular DNA molecules. The syn-thesis of proteins encoded by genes on mitochondrial orchloroplast DNA takes place on ribosomes inside theorganelles themselves. Therefore the proteins in mito-chondria and chloroplasts are of two different origins:either encoded in the nucleus and imported into the or-ganelle or encoded in the organelle and synthesizedwithin the organelle compartment.
1.2 Genetic variationIf all members of a species have the same set of genes,how can there be genetic variation? As indicated earlier,the answer is that genes come in different forms calledalleles. In a population, for any given gene there can befrom one to many different alleles; however, becausemost organisms carry only one or two chromosome setsper cell, any individual organism can carry only one ortwo alleles per gene. The alleles of one gene will alwaysbe found in the same position along the chromosome.Allelic variation is the basis for hereditary variation.
Types of variationBecause a great deal of genetics concerns the analysis ofvariants, it is important to understand the types of varia-tion found in populations. A useful classification is intodiscontinuous and continuous variation (Figure 1-10). Al-lelic variation contributes to both.
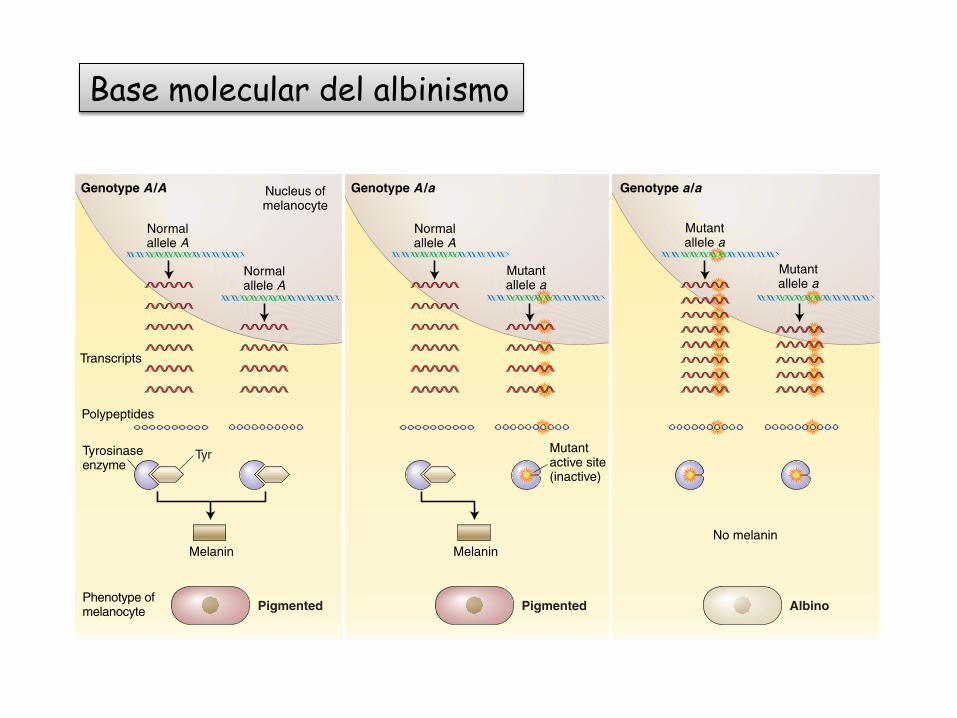
DISCONTINUOUS VARIATION Most of the research ingenetics in the past century has been on discontinuousvariation because it is a simpler type of variation, and itis easier to analyze. In discontinuous variation, a charac-ter is found in a population in two or more distinct andseparate forms called phenotypes. “Blue eyes” and“brown eyes” are phenotypes, as is “blood type A” or“blood type O.” Such alternative phenotypes are oftenfound to be encoded by the alleles of one gene. A goodexample is albinism in humans, which concerns pheno-types of the character of skin pigmentation. In most peo-ple, the cells of the skin can make a dark-brown or blackpigment called melanin, the substance that gives our skinits color ranging from tan color in people of Europeanancestry to brown or black in those of tropical and sub-tropical ancestry. Although always rare, albinos, whocompletely lack pigment in their skin and hair, are foundin all races (Figure 1-11). The difference between pig-
MESSAGE The flow of information from DNA to RNA toprotein is a central focus of modern biology.
Wingless fruit Winged fruit
2 m
m
Figure 1-10 Examples of discontinuous and continuous variation in natural populations.(a) Fruits of the sea blush, Plectritis congesta, have one of two distinct forms. Any oneplant has either all winged or all wingless fruits. (b) Variation in height, branch number,and flower number in the herb Achillea. [Part b, Carnegie Institution of Washington.]
(a) (b)
44200_01_p1-26 3/2/04 4:01 PM Page 8
Muchas variantes del gen
Polimorfismo

91.2 Genetic variation
mented and unpigmented skin is caused by different al-leles of a gene that encodes an enzyme involved inmelanin synthesis.
The alleles of a gene are conventionally designatedby letters. The allele that codes for the normal form ofthe enzyme involved in making melanin is called A, andthe allele that codes for an inactive form of that enzyme(resulting in albinism) is designated a, to show that theyare related. The allelic constitution of an organism is itsgenotype, which is the hereditary underpinning of thephenotype. Because humans have two sets of chromo-somes in each cell, genotypes can be either A/A, A/a, ora/a (the slash shows that the two alleles are a pair). Thephenotype of A/A is pigmented, that of a/a is albino,and that of A/a is pigmented. The ability to make pig-ment is expressed over inability (A is said to be domi-nant, as we shall see in Chapter 2).
Although allelic differences cause phenotypic differ-ences such as pigmented and albino coloration, this doesnot mean that only one gene affects skin color. It isknown that there are several, although the identity and number of these genes are currently unknown.However, the difference between pigmented, of whatevershade, and albinism is caused by the difference in the al-leles of one gene—the gene that determines the abilityto make melanin; the allelic composition of other genesis irrelevant.
In some cases of discontinuous variation, there is apredictable one-to-one relation between genotype and
phenotype under most conditions. In other words, thetwo phenotypes (and their underlying genotypes) canalmost always be distinguished. In the albinism exam-ple, the A allele always allows some pigment forma-tion, whereas the a allele always results in albinismwhen present in two copies. For this reason, discontinu-ous variation has been successfully used by geneticiststo identify the underlying alleles and their role in cellu-lar functions.
Geneticists distinguish two categories of discontin-uous variation. In a natural population, the existence oftwo or more common discontinuous variants is calledpolymorphism (Greek; many forms). The various formsare called morphs. It is often found that differentmorphs are determined by different alleles of a singlegene. Why do populations show genetic polymor-phism? Special types of natural selection can explain afew cases, but, in other cases, the morphs seem to beselectively neutral.
Rare, exceptional discontinuous variants are calledmutants, whereas the more common “normal” pheno-type is called the wild type. Figure 1-12 shows an ex-ample of a mutant phenotype. Again, in many cases,the wild-type and mutant phenotypes are determinedby different alleles of one gene. Both mutants and poly-morphisms originally arise from rare changes in DNA(mutations), but somehow the mutant alleles of a poly-morphism become common. These rare changes inDNA may be nucleotide-pair substitutions or smalldeletions or duplications. Such mutations change theamino acid composition of the protein. In the case ofalbinism, for example, the DNA of a gene that encodesan enzyme involved in melanin synthesis is changed,such that a crucial amino acid is replaced by anotheramino acid or lost, yielding a nonfunctioning enzyme.Mutants (such as those that produce albinism) can oc-cur spontaneously in nature, or they can be producedby treatment with mutagenic chemicals or radiation.Geneticists regularly induce mutations artificially to
Figure 1-11 An albino. The phenotype is caused by two dosesof a recessive allele, a /a. The dominant allele A determinesone step in the chemical synthesis of the dark pigmentmelanin in the cells of skin, hair, and eye retinas. In a /aindividuals, this step is nonfunctional, and the synthesis ofmelanin is blocked. [Copyright Yves Gellie/Icone.]
Figure 1-12 Wild type and mutant Drosophila. A Drosophilamutant with abnormal wings and a normal fly (wild type) forcomparison. In both cases, the two phenotypes are caused bythe alleles of one gene.
Wild type Vestigial wings
44200_01_p1-26 3/2/04 4:01 PM Page 9
91.2 Genetic variation
mented and unpigmented skin is caused by different al-leles of a gene that encodes an enzyme involved inmelanin synthesis.
The alleles of a gene are conventionally designatedby letters. The allele that codes for the normal form ofthe enzyme involved in making melanin is called A, andthe allele that codes for an inactive form of that enzyme(resulting in albinism) is designated a, to show that theyare related. The allelic constitution of an organism is itsgenotype, which is the hereditary underpinning of thephenotype. Because humans have two sets of chromo-somes in each cell, genotypes can be either A/A, A/a, ora/a (the slash shows that the two alleles are a pair). Thephenotype of A/A is pigmented, that of a/a is albino,and that of A/a is pigmented. The ability to make pig-ment is expressed over inability (A is said to be domi-nant, as we shall see in Chapter 2).
Although allelic differences cause phenotypic differ-ences such as pigmented and albino coloration, this doesnot mean that only one gene affects skin color. It isknown that there are several, although the identity and number of these genes are currently unknown.However, the difference between pigmented, of whatevershade, and albinism is caused by the difference in the al-leles of one gene—the gene that determines the abilityto make melanin; the allelic composition of other genesis irrelevant.
In some cases of discontinuous variation, there is apredictable one-to-one relation between genotype and
phenotype under most conditions. In other words, thetwo phenotypes (and their underlying genotypes) canalmost always be distinguished. In the albinism exam-ple, the A allele always allows some pigment forma-tion, whereas the a allele always results in albinismwhen present in two copies. For this reason, discontinu-ous variation has been successfully used by geneticiststo identify the underlying alleles and their role in cellu-lar functions.
Geneticists distinguish two categories of discontin-uous variation. In a natural population, the existence oftwo or more common discontinuous variants is calledpolymorphism (Greek; many forms). The various formsare called morphs. It is often found that differentmorphs are determined by different alleles of a singlegene. Why do populations show genetic polymor-phism? Special types of natural selection can explain afew cases, but, in other cases, the morphs seem to beselectively neutral.
Rare, exceptional discontinuous variants are calledmutants, whereas the more common “normal” pheno-type is called the wild type. Figure 1-12 shows an ex-ample of a mutant phenotype. Again, in many cases,the wild-type and mutant phenotypes are determinedby different alleles of one gene. Both mutants and poly-morphisms originally arise from rare changes in DNA(mutations), but somehow the mutant alleles of a poly-morphism become common. These rare changes inDNA may be nucleotide-pair substitutions or smalldeletions or duplications. Such mutations change theamino acid composition of the protein. In the case ofalbinism, for example, the DNA of a gene that encodesan enzyme involved in melanin synthesis is changed,such that a crucial amino acid is replaced by anotheramino acid or lost, yielding a nonfunctioning enzyme.Mutants (such as those that produce albinism) can oc-cur spontaneously in nature, or they can be producedby treatment with mutagenic chemicals or radiation.Geneticists regularly induce mutations artificially to
Figure 1-11 An albino. The phenotype is caused by two dosesof a recessive allele, a /a. The dominant allele A determinesone step in the chemical synthesis of the dark pigmentmelanin in the cells of skin, hair, and eye retinas. In a /aindividuals, this step is nonfunctional, and the synthesis ofmelanin is blocked. [Copyright Yves Gellie/Icone.]
Figure 1-12 Wild type and mutant Drosophila. A Drosophilamutant with abnormal wings and a normal fly (wild type) forcomparison. In both cases, the two phenotypes are caused bythe alleles of one gene.
Wild type Vestigial wings
44200_01_p1-26 3/2/04 4:01 PM Page 9
AA AA AA
AA
AA
Aa
AA
aa Mutación Variante rara del gen
Silvestre Mutante
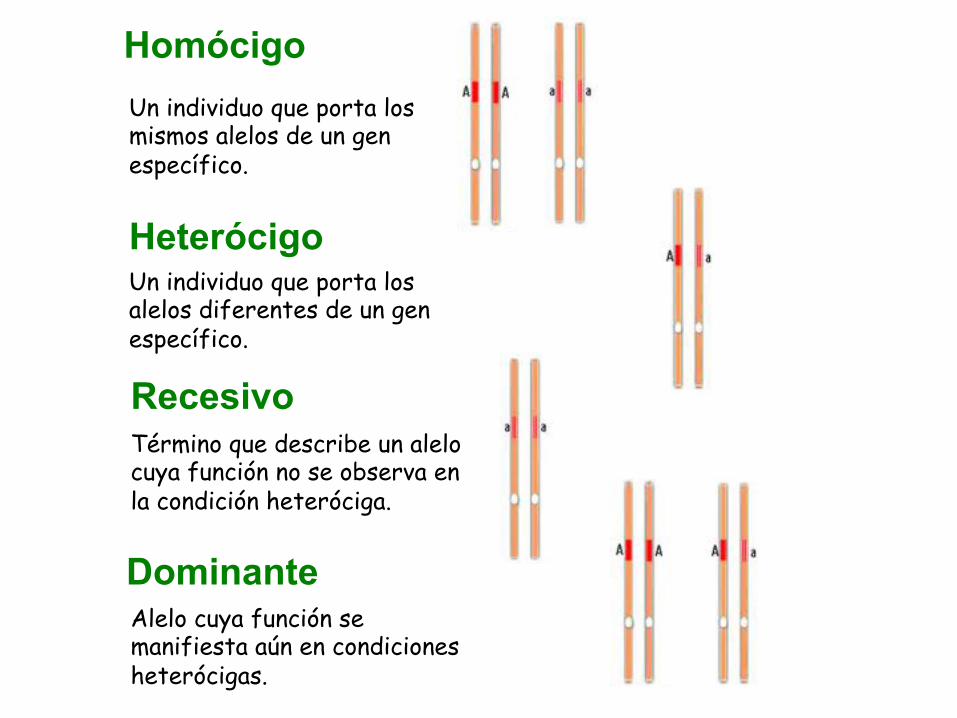
Heterócigo
Homócigo Un individuo que porta los mismos alelos de un gen específico.
Un individuo que porta los alelos diferentes de un gen específico.
Dominante Alelo cuya función se manifiesta aún en condiciones heterócigas.
Término que describe un alelo cuya función no se observa en la condición heteróciga.
Recesivo
111.3 Methodologies used in genetics
Polypeptides
Transcripts
Melanin
Normalallele A
Genotype A/A Genotype A/a Genotype a/a
Tyrosinaseenzyme
Phenotype ofmelanocyte
Tyr
Pigmented
Normalallele A
Normalallele A
Nucleus ofmelanocyte
Pigmented
Mutantallele a
Mutantallele a
Mutantallele a
Albino
MelaninNo melanin
Mutantactive site(inactive)
change in shape or size can result in an absence of bio-logical function (which would be the basis of a null al-lele) or reduced function. More rarely, mutation can leadto new function of the protein product.
only hereditary mechanisms, but all biological mecha-nisms. Many different methodologies are used to studygenes and gene activities, and these methodologies canbe summarized briefly as follows:
1. Isolation of mutations affecting the biologicalprocess under study. Each mutant gene reveals agenetic component of the process, and together themutant genes show the range of proteins thatinteract in that process.
2. Analysis of progeny of controlled matings (“crosses”)between mutants and wild-type individuals or otherdiscontinuous variants. This type of analysisidentifies genes and their alleles, their chromosomallocations, and their inheritance patterns. Thesemethods will be introduced in Chapter 2.
3. Genetic analysis of the cell’s biochemical processes.Life is basically a complex set of chemical reactions,so studying the ways in which genes are relevant tothese reactions is an important way of dissecting thiscomplex chemistry. Mutant alleles underlyingdefective function (see method 1) are invaluable in
Figure 1-13 Molecular basis of albinism. Left: Melanocytes (pigment-producing cells)containing two copies of the normal tyrosinase allele (A) produce the tyrosinase enzyme,which converts the amino acid tyrosine into the pigment melanin Center: Melanocytescontaining one copy of the normal allele make enough tyrosinase to allow production ofmelanin and the pigmented phenotype. Right: Melanocytes containing two copies ofthe mutant null allele (a) are unable to produce any of the enzyme.
MESSAGE New alleles formed by mutation can result inno function, less function, more function, or new functionat the protein level.
1.3 Methodologies used in geneticsAn overviewThe study of genes has proved to be a powerful ap-proach to understanding biological systems. Becausegenes affect virtually every aspect of the structure andfunction of an organism, being able to identify and de-termine the role of genes and the proteins that theyspecify is an important step in charting the variousprocesses that underlie a particular character under in-vestigation. It is interesting that geneticists study not
44200_01_p1-26 3/2/04 4:01 PM Page 11
Base molecular del albinismo

Locus
El lugar donde se localiza el gen en el cromosoma. El locus normalmente incluye varios genes que se encuentran físicamente muy cerca uno del otro. Los genes localizados en un mismo locus tienen poca probabilidad de sufrir eventos de recombinación.

Célula haploide especializada involucrada en la reproducción sexual.
Gameto
Cruza Apareamiento entre dos individuos que conduce a la fusión de gametos.
Cigoto Célula producto de la fusión de gametos.

Constitución genética de un individuo, bien colectivamente en todos los loci o, más frecuentemente, en un solo locus
Genotipo
Fenotipo Expresión observable del genotipo, como rasgo morfológico, bioquímico o molecular.
Geno%po = bb Geno%po = BB o Bb
Recesivo = b Dominante = B
Feno%po = ojos cafés Feno%po = ojos azules

Influencia del ambiente
Frecuentemente los genes son influenciados por el ambiente y el fenotipo se afectará por este

Influencia del desrrollo
Aún bajo el mismo ambiente, genotipos idénticos pueden tener “ruido del desarrollo” y resultar en fenotipos diferentes

20 Chapter 1 • Genetics and the Organism
of flies raised at lower temperatures. These norms of re-action indicate that the relation between genotype andphenotype is complex rather than simple.
MESSAGE A single genotype may produce differentphenotypes, depending on the environment in whichorganisms develop. The same phenotype may be produced by different genotypes, depending on the environment.
If we know that a fruit fly has the wild-type genotype,this information alone does not tell us whether its eyehas 800 or 1000 facets. On the other hand, the knowl-edge that a fruit fly’s eye has 170 facets does not tell uswhether its genotype is ultrabar or infrabar. We cannoteven make a general statement about the effect of tem-perature on eye size in Drosophila, because the effect isopposite in two different genotypes. We see from Figure1-20 that some genotypes do differ unambiguously inphenotype, no matter what the environment: any wild-type fly has larger eyes than any ultrabar or infrabar fly.But other genotypes overlap in phenotypic expression:the eyes of an ultrabar fly may be larger or smaller thanthose of an infrabar fly, depending on the temperaturesat which the individuals developed.
To obtain a norm of reaction such as the norms ofreaction in Figure 1-20, we must allow different individ-uals of identical genotype to develop in many differentenvironments. To carry out such an experiment, wemust be able to obtain or produce many fertilized eggswith identical genotypes. For example, to test a humangenotype in 10 environments, we would have to obtaingenetically identical sibs and raise each individual in adifferent milieu. However, that is possible neither bio-logically nor ethically. At the present time, we do notknow the norm of reaction of any human genotype forany character in any set of environments. Nor is it clearhow we can ever acquire such information without theunacceptable manipulation of human individuals.
For a few experimental organisms, special geneticmethods make it possible to replicate genotypes and thusto determine norms of reaction. Such studies are particu-larly easy in plants that can be propagated vegetatively(that is, by cuttings). The pieces cut from a single plant allhave the same genotype, so all offspring produced in thisway have identical genotypes. Such a study has been doneon the yarrow plant, Achillea millefolium (Figure 1-21a).The experimental results are shown in Figure 1-21b.Many plants were collected, and three cuttings weretaken from each plant. One cutting from each plant wasplanted at low elevation (30 meters above sea level), oneat medium elevation (1400 meters), and one at high ele-vation (3050 meters). Figure 1-21b shows the mature in-dividuals that developed from the cuttings of sevenplants; each set of three plants of identical genotype isaligned vertically in the figure for comparison.
0
50High elevation
Hei
ght (
cm)
50Medium elevation
0 1 2 3 4 5 6 7
50Low elevation
Parental plant (source of cuttings)
0
(b)
Figure 1-21 A comparison of genotype versus phenotype inyarrow. (a) Achillea millefolium. (b) Norms of reaction toelevation for seven different Achillea plants (seven differentgenotypes). A cutting from each plant was grown at low,medium, and high elevations. [Part a, Harper Horticultural SlideLibrary; part b, Carnegie Institution of Washington.]
(a)
44200_01_p1-26 3/2/04 4:01 PM Page 20
Genotipos
Am
bien
tes
Reacción de la planta Achillea millefolium a diferentes alturas

GENÉTICA MENDELIANA

¿Cómo son heredadas las características de
un organismo?
1866 – “Experimentos sobre hibridación de
plantas” (Versuche über Plflanzenhybriden)
Gregor Mendel 1822 - 1884

Planteamiento experimental de Mendel Organismo: Pisum sativum “Guisante de Jardín”

“Guisante de Jardín”
• Dife ren tes semi l l as d i s p o n i b l e s c u y o s p r o d u c t o s e r a n fácilmente identificables y analizables.
• Autopo l in i zac ión o polinización cruzada
• 5 caracteres (7 PARES DE FENOTIPOS para analizar)
• Obtención de líneas puras (Homocigotos)

Procedimiento experimental – Polinización
cruzada
Observaciones: El resultado era el mismo no
importando que planta era la receptora o donadora de
polen (Cruza Recíproca)

Cruza Monohíbrida
Generación F1: • Todas Altas (Fenotipo Dominante)
Mendel: De alguna forma el fenotipo “enano”
debía estar presente pero sin expresarse
Autopolinización F1
Obtención de F2: • Re-aparecía el fenotipo Enano (Recesivo) • # Plantas altas:enanas 787:277
Proporción 3:1
(Sólo se estudia un rasgo/característica)

Resultados Cruzas Monohíbridas

Genes Dominante (B)
Recesivo (b)
Alelos
Explicación Mendel: cada carácter estaba controlado por un factor heredable (Genes).
Los genes
estaban en
pares Cada Parental tenia dos copias idénticas de un gen. Homocigotos Los gametos emergen de la MEIOSIS portando una sola
copia del gen. F1: dos alelos diferentes
(padre y madre). Heterocigoto
Fenotipos Genotipos Proporción genotípica
Proporción fenotípica
Alto DD 1
3 Dd 2
Enano dd 1 1
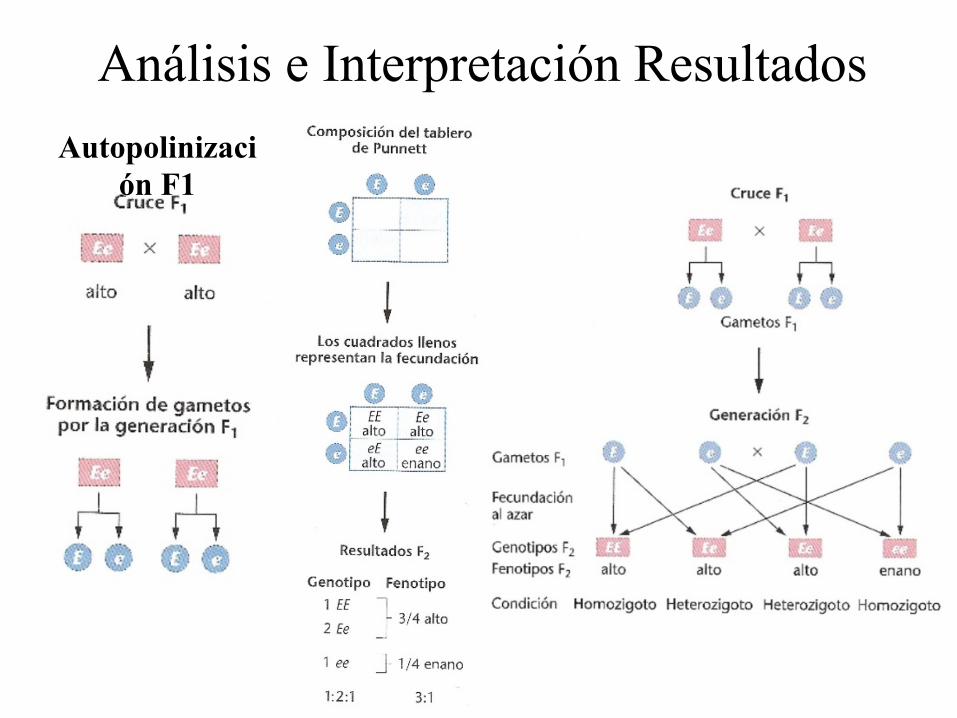
Análisis e Interpretación Resultados Autopolinizaci
ón F1

Cruza Prueba Distinguir el genotipo de una planta que expresa el fenotipo dominante.
La cruza se debe hacer con un individuo homocigoto recesivo.

Principios de Mendel 1. Principio de Dominancia. En un heterocigoto,
un alelo podrá ocultar la presencia de otro. Este es un principio acerca de la función génica. Algunos alelos controlan evidentemente el fenotipo incluso cuando están presentes en una sola copia.
2. Principio de segregación. En un heterocigoto, dos diferentes alelos se separan uno del otro durante la formación de gametos.
Este es un principio acerca de la transmisión génica. Aunque se observe sólo el fenotipo dominante siempre se transmiten ambos alelos (dominante y recesivo). Las bases biológicas de este fenómeno es el apareamiento y subsecuente separación de cromosomas homólogos durante la MEIOSIS.

Resumen: Modelo de Mendel 1. Para producir un carácter observable es necesario un factor hereditario
llamado GEN. 2. Cada planta tiene dos copias para un gen determinado (cromosomas
homólogos). 3. Existen formas alternativas del gen llamadas alelos. Una planta puede
ser bien B/B, B/b o b/b. La barra representa que los alelos forman una pareja (cromosomas homólogos).
4. En la planta B/b el alelo B domina. Por lo tanto, el fenotipo de la planta B/b se define como dominante. El alelo b es recesivo y en presencia de B no se manifiesta en el fenotipo.
5. Durante la MEIOSIS los miembros de un par génico se distribuyen equitativamente entre los gametos. Esta distribución se conoce como primera Ley de Mendel o Ley de la Segregación equitativa.
6. Un gameto contiene un solo miembro de un par génico. 7. Durante la fecundación, los gametos se unen al azar,
independientemente de que alelos porten.

Problemas 1. Dos líneas puras de ratón, una con piel negra y otra con piel
gris, fueron cruzadas, y toda la descendencia tenia piel negra. ¿Cuál es el genotipo y fenotipo de la Filial 2 (F2)?
2. Las palomas pueden presentar un patrón ajedrezado o liso. En una serie de cruces controlados, se obtuvieron los siguientes resultados:
Descendencia F1
Cruce P1 Ajedrezados Lisos
a) Ajedrezado x Ajedrezado 36 0
b) Ajedrezado x liso 38 0
c) liso x liso 0 35
Luego, los descendientes F1 se cruzaron selectivamente, con los siguientes resultados. Descendencia F2
Cruce F1 x F1 Ajedrezados Lisos
d) Ajedrezado (a) x liso (c) 34 0
e) Ajedrezado (b) x liso (c) 17 14
f) Ajedrezado (b) x Ajedrezado (b) 28 9
g) Ajedrezado (a) x Ajedrezado (b) 39 0

• ¿Cómo se heredan los patrones ajedrezado y liso?
• Definir los genotipos parentales de cada cruza • Representar las cruzas y genotipos de los
descendientes
Problemas