Embed Size (px)
Citation preview

Indian Journal of Experimental Biology Vol. 38, June 2000, pp. 567-574
Regenerative ability of gastrocnemius muscle under diabetic condition with special reference to SDH & m-ATPase
C L Talesara & Nisha Vashishta
Muscle Physiology & Histochemistry Unit, Department of Zoology, University of Delhi, Delhi II 0007, India
Received 23 July /998; revised I March 2000
The present study analyzes the regeneration of skeletal muscle in streptozotocin (STZ) induced diabetic rats . Two weeks later the gastrocnemius muscle from di abetic rats were transplanted into diabetic and normal host to initiate regeneration and the normal gastrocnemius muscle was transplanted in normal and diabetic hosts for comparison . The regenerates were analyzed after 15 and 30 days of transplantation for histochemical (with respect to SOH and rn-A TPase) and 7, 14, 21 and 28 days after transplantation for biochemical studies (with respect to SOH and m-ATPase). Least enzymatic activity and the poorest regenerative ability in case of normal muscle in di abetic host (NM - DbH) and comparatively higher enzymatic activity and better regenerative ability in diabetic muscle in normal host (DbM -NH) was observed . The result of this study strongly supports that the normal host environment is crucial for the muscle recovery.
Diabetes mellitus is known to have a profound and differential effect on the three types of rat skeletal muscle fibres. Mammalian muscles are usually composed of fibres which differ not only in their gross colour i.e . red, white and intermediate (pink) but also in their histochemical , biochemical, physiological, metabolical and over all contractile properties1
-5
•
Mammalian skeletal muscle are comprised of three types of fibres with different population, i.e. fast oxidative glycolytic (FOG), fast glycolytic (FG) and slow oxidative (SO) fibres6
-8
.
Skeletal muscle requires insulin for normal growth and development9
-12
. A pathological deficiency of insulin as frequently investigated by experimentally induced diabetes mellitus in the rat, is characterized by a considerable disturbance of muscle protein tumoveru, a decrease of protein synthesis 14 and an increase in protein degradation 15
. The general loss of muscle protein seems also to include a loss of myofibrillar proteins. Decrease in succinate dehydrogenase (SOH) and myofibrillar adenosine triphosphatase (rn-A TPase) activity in the diabetic muscle has been reported earlier by many workers. This decrease in SOH activity can be attributed to the mitochondrial dysfunction in diabetic condition 16
, rn-A TPase activity has been correlated with speed of contraction of the muscle17
• Decrease in this enzyme activity has been observed in diabetic gastrocnemius muscle showing the decrease in contractile capacity of the muscle 18
.
Skeletal muscles show tremendous degree of adaptabi lity under varying specific functional demands 19.Tissue regeneration appears to be the common mode by which injured vertebrate muscles normally repair itself. It can occur in dimensions ranging from the repair of a single fibre or small group of fibres to the formation of entire muscle from minced fragments. A precise cascade of events that follows transplantation includes degeneration of a majority of myofibres, activation and proliferation of precursor myosatellite cells, differentiation of myoblasts, fusion to form myotubes and eventual maturation into
f "b 20 myo 1 res .
V . I . d. 2 )-?1 d h anous transp antat10n stu tes -· emonstrate t at skeletal muscle preserves its ability to regenerate and can recover from variety of experimental and metabolic alterations. In this experimental work an attempt has been made to study the regenerative abi lity of normal gastrocnemius muscle in normal and diabetic host and diabetic gastrocnemius muscle in normal and diabetic host. The study is mainly concerned with the two important contractile (m-ATPase) and mitochondrial (SOH) enzymes. According to Baran/7 m-ATPase corresponding to a given isomyosin, refl ects the speed of contraction of muscle thus relating to the mechanical performance and strength of the muscle. The deposition of diformazan as a result of SOH activity is mainly used as an indicator of aerobic potential m histochemically differentiated muscle fibre types 24
.

568 INDIAN J EXP BIOL, JUNE 2000
Regenerative ability of transplanted gastrocnemius muscle (from normal and streptozotocin induced diabetic rats) in normal and diabetic host has been studied with a view to probing the role of the host metabolic environment in determining the success of muscle regeneration.
Materials and Methods Adult (I 0-12 weeks) male albino rats Rattus
norvegicus weighing 150-200 g were used as a mammalian representative for the experiments. Rats were kept under standard laboratory conditions and fed on commercial food (Gold Mohur rat feed, Hindustan Lever Ltd.) and water ad libitum.
Rats were fasted overnight and induced diabetes by a single iv injection in the tail by a freshly prepared solution of streptozotocin (STZ) 40 mg/kg body weight, in 0.1 M citrate buffer pH 4.5 under anaesthesia25
. To screen for diabetes, plasma glucose levels were determined by Stangen Glucose Kit which is based on Trinders method in which glucose oxidase and peroxidase enzymes are used along with the chromogen 4-amino anti pyrine and phenol. STZ treated rats developed a markedly elevated plasma glucose level ( 180-250 mg/dl) that was measured two weeks after injection. Few rats that did not exhibit increase in plasma glucose level after STZ injection were excluded from the study. Two weeks after streptozotocin injection the rats were divided into 4 groups for minced muscle surgery/transplantation, 4 to 6 animals for each interval s in each group were used for a combined histochemical and biochemical study :
1st Group: Normal muscle in normal host (NM- NH) ; (autominced transplantation)
A part of gastrocnemius muscle of left leg from normal rat under anaesthesia was removed and then minced into I mm fragments and these were replaced into the site from where the muscle was removed without disturbing the related nerves (autominced transplantation). The muscle and the skin incisions were closed with a fine surgical thread and the wounds were cleaned with absolute alcohol. Nebasulf (Neomycin - Bacitracin - Sulphacetaniide) antiseptic powder was sprinkled on the wounds of operated animals and finally picric acid was rubbed on the wounds. 2nd group: Diabetic muscle in normal host (DbM -NH); (Isominced transplantation)
3rd group: Diabetic muscle in diabetic: host (DbM -DbH); (autominced transplantation) 4th group: Normal muscle in diabetic host (NM -DbH); (lsqminced transplantation)
Four muscle transplants from each group were removed for histochemical (at the intervals of 15 and 30 days) and biochemical (at the interval of 7, 14, 21, and 28 days) analysis after transplantation .
These muscle transplants were processed for the correlative histochemical and biochemical analysis. For histochemical studies the muscles were frozen in liquid nitrogen and immediately transferred to a cryostat (IEC- Model CTD - International Harris Cryostat) at -25°C. Cross sections (I 0 ~-tm thick) were cut and stained for SDH26
• Other sections were stained for alkali and acid stable forms of myofibrillar adenosine triphosphatase (m-ATPase) basically by the method of Padykula and Herman27 as modified by Guth and Samaha28
. For Biochemical, myofibrillar - ATPase assay, myofibrils were prepared according to Perry and Gre/9
. Assay was performed within 6 hr of myofibril preparations at pH 7.5 and at 37°C. Values were expressed as 11moles of Pi liberated/rng protein /min. The method of Rockstein and Herron 30 was used to calculate the amount of Pi liberated and the protein content was measured by the method of Gornall et a/. 31
.
SDH activity was measured in muscle homogenate using INT [2- (p- iodophenyl , 3 (p- nitrophenyl) - 5 -(phenyl tetrazolium chloride] as an electron acceptor and 0.2 M sodium succinate as a substrate32
. Values were expressed as ~-tg formazanlmg protein/min.
Statistical analysis All the data were analysed with two way ANOV A.
P< 0.05 was considered significant.
Results and Discussion The present study analyzes the regenerative ability
of gastrocnemius muscle under experimentally induced diabetic condition. The gastrocnemius muscle regenerates were analyzed histochemically (15 and 30 days) and biochemically (7, 14, 21 and 28 days) postoperatively with special reference to enzymes SDH and rn-A TPase.
After almost all forms of injury the damaged skeletal muscle fibres go through a characteristic degenerative phase before the regenerative response becomes obvious33
.34
. 15 days after traTI#plantation (Figs 1-5), the transplanted muscle shows large amount of
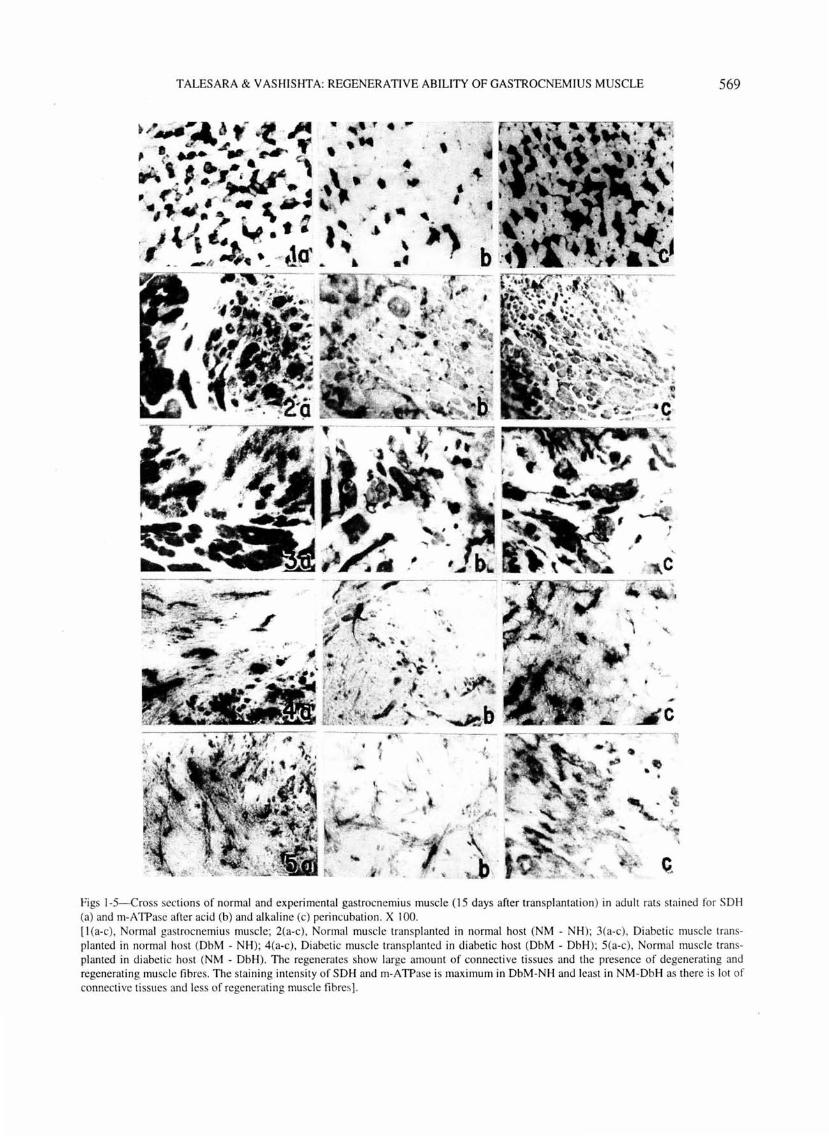
TALESARA & VASHISHTA: REGENERATIVE ABILITY OF GASTROCNEMI US MUSCLE 569
figs 1-5-Cross sections of normal and experimental gastrocnemiu s muscle (15 days after transpl antation) in adult rats stai ned for SOH (a) and rn-A TPase after ac id (b) and alkaline (c) perincubation. X I 00. [ I (a-c), Normal gastrocnemius muscle; 2(a-c), Normal muscle transpl anted in normal host (NM - NH); 3(a-c), Diabetic muscle transpl anted in normal host (DbM - NH); 4(a-c), Diabetic muscle transplanted in di abetic host (DbM - DbH); 5(a-c), Normal muscle transplanted in d iabetic host (NM - DbH). The regenerates show large amount of connecti ve tissues and the presence of degenerati ng and regenerating mu scle fibres. The staining intensity of SOH and rn-A TPase is maximum in DbM-NH and leas t in NM-DbH as there is lot of connect ive tissues and less of regenerating muscle fibres].
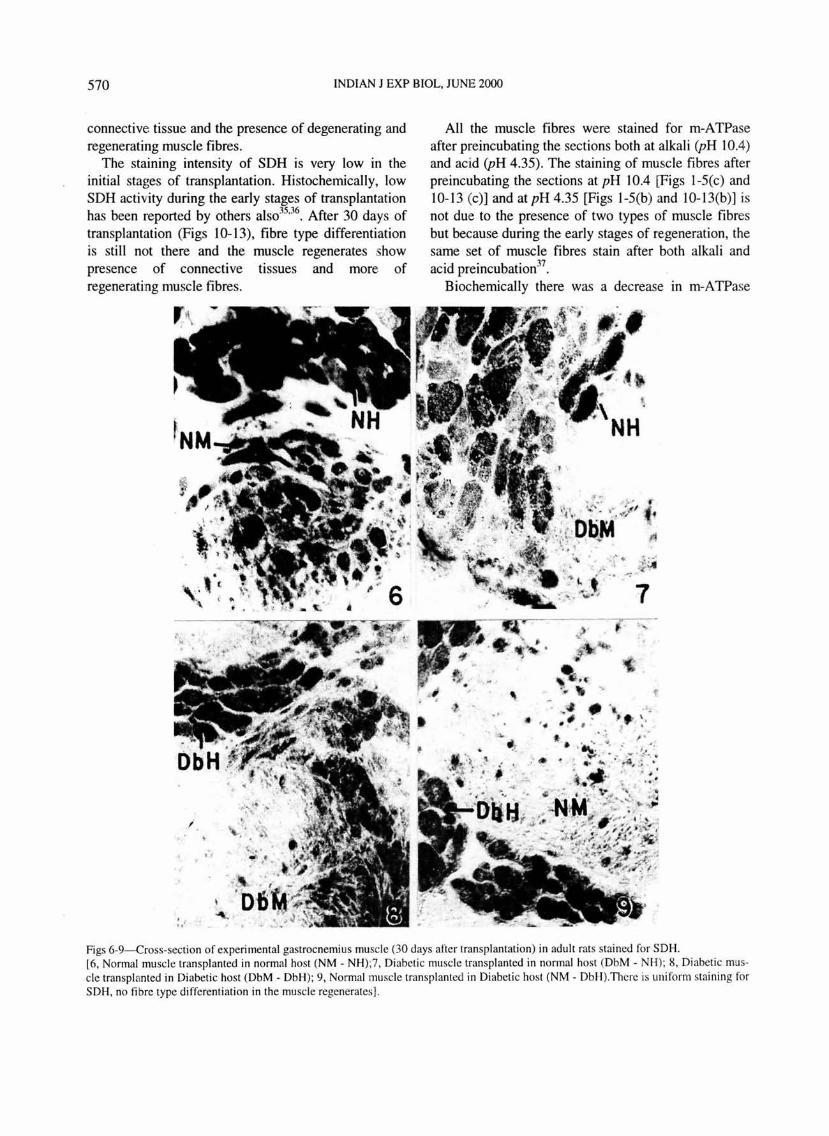
570 INDIAN J EXP BIOL, JUNE 2000
connective tissue and the presence of degenerating and regenerating muscle fibres .
The staining intensity of SOH is very low in the initial stages of transplantation. Histochemically, low SOH activity during the early stages of transplantation has been reported by others also35
'36
. After 30 days of transplantation (Figs 1 0-13), fibre type differentiation is still not there and the muscle regenerates show presence of connective tissues and more of regenerating muscle fibres.
All the muscle fibres were stained for rn-A TPase after preincubating the sections both at alkali (pH 10.4) and acid (pH 4.35). The staining of muscle fibres after preincubating the sections at pH 10.4 [Figs I-5(c) and 10-13 (c)] and at pH 4.35 [Figs 1-5(b) and 10-13(b)] is not due to the presence of two types of muscle fibres but because during the early stages of regeneration, the same set of muscle fibres stain after both alkali and acid preincubation37
.
Biochemically there was a decrease in m-ATPase
Figs 6-9-Cross-section of experimental gastrocnemius muscle (30 days after transplantation) in adult rats stained for SDH. [6, Normal muscle transpl anted in normal host (NM - NH);7, Di abeti c muscle transplanted in normal host (DbM- NH); 8, Diabetic muscle transplanted in Diabetic host (DbM - DbH); 9 , Normal muscle transplanted in Diabeti c host (NM - DbH ).Thcre is uniform staining for SDH, no fib re type di ffe renti ation in the muscle regenerates].
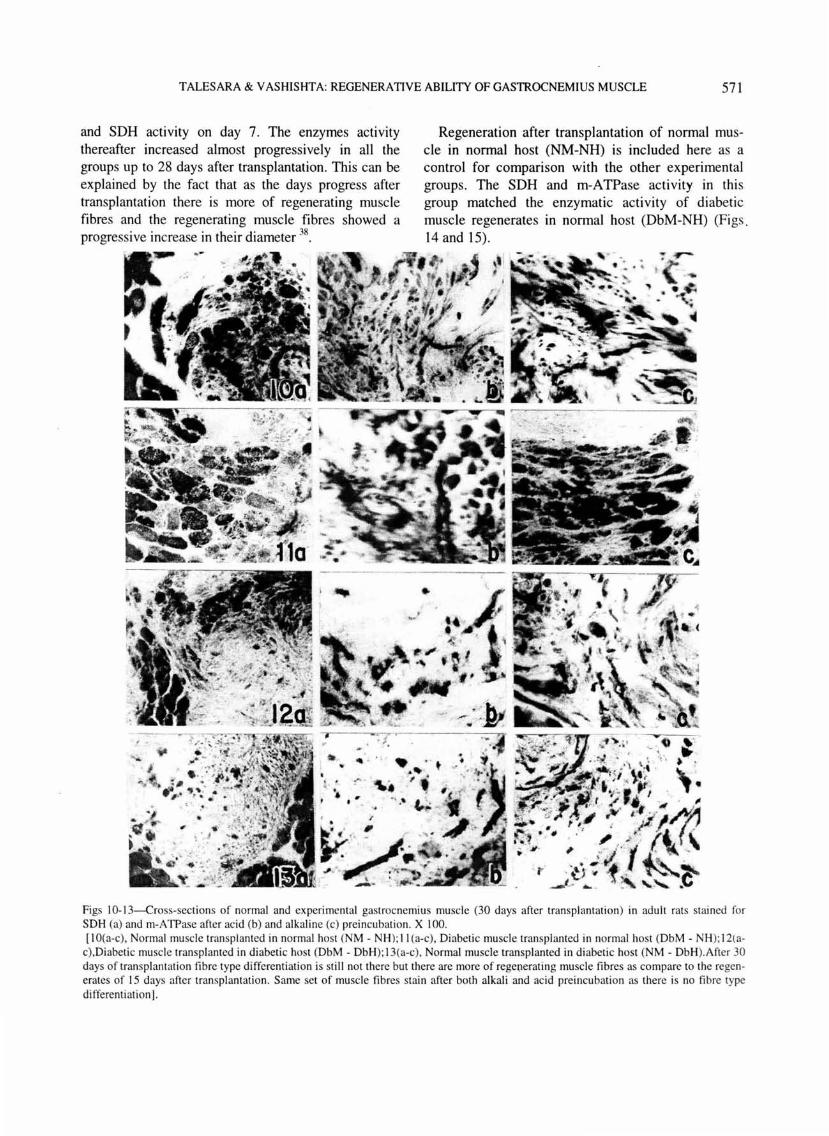
TALESARA & VASHISHTA: REGENERATIVE ABILITY OF GASTROCNEMIUS MUSCLE 571
and SDH actiVIty on day 7. The enzymes actiVIty thereafter increased almost progressively in all the groups up to 28 days after transplantation . This can be explained by the fact that as the days progress after transplantation there is more of regenerating muscle fibres and the regenerating muscle fibres showed a progressive increase in their diameter 38
•
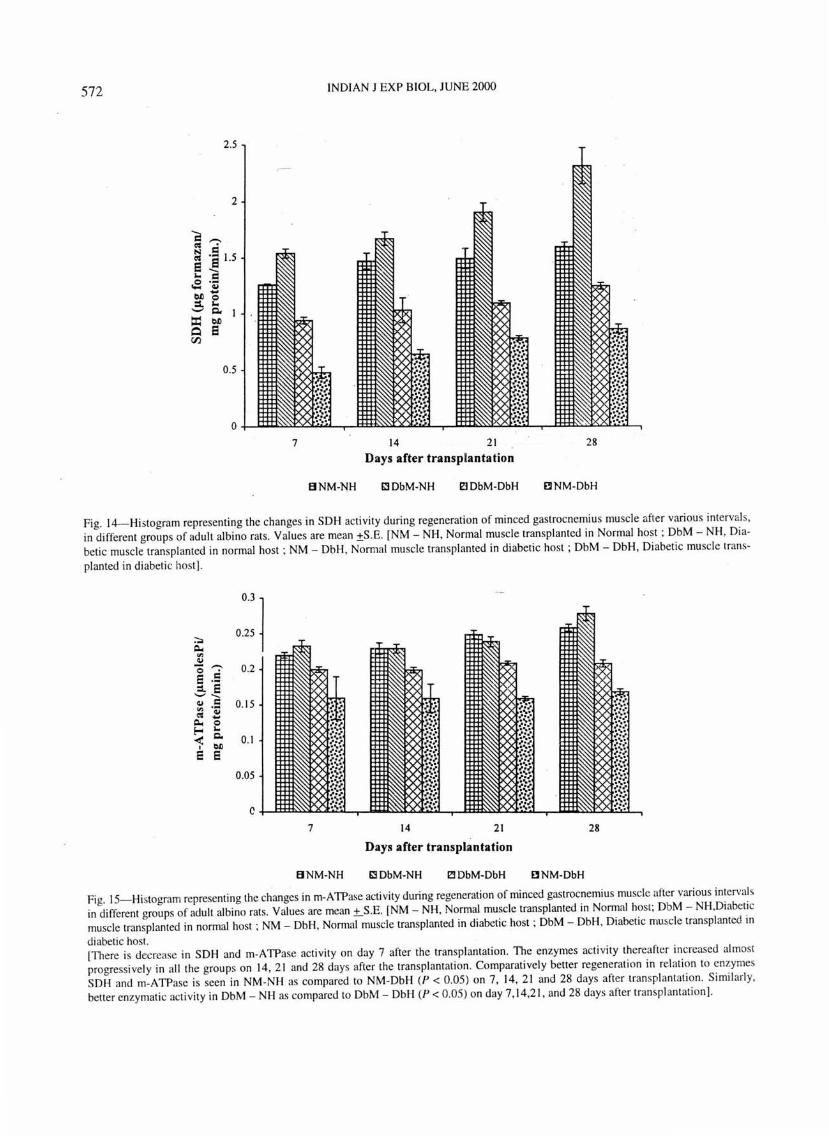
Regeneration after transplantation of normal muscle in nonpal host (NM-NH) is included here as a control for comparison with the other experimental groups. The SDH and m-ATPase activity in this group matched the enzymatic activity of diabetic muscle regenerates m normal host (DbM-NH) (Figs. 14 and 15).
Figs 10-13-Cross-sections of normal and experimental gastrocnemius muscle (30 days after transplantation) in adult rats stained for SOH (a) and rn-A TPase after acid (b) and alkaline (c) preincubation . X I 00. [I O(a-c), Normal muscle transplanted in normal host (NM - NH); II (a-c), Diabetic muscle transplanted in normal host (DbM - NH); 12(a
c),Diabetic muscle transpl anted in diabetic host (DbM - Db H); 13(a-c), Normal muscle transplanted in diabetic host (NM - DbH).After 30 days of transplantation fibre type differentiation is stil l not there but there are more of rege(lerating muscle fibres as compare to the regenerates of 15 days after transplantation . Same set of muscle fibres stain after both alkali and aci d preincubation as there is no fibre type differentiation].
572 INDIAN J EXP BIOL, JUNE 2000
7 14 21 28 Days after transplantation
BNM-NH ~DbM-NH 21 DbM-DbH E3 NM-DbH
Fig. 14--Histogram representing the changes in SOH activity during regeneration of minced gastrocnemius mu scle afte r various intervals, in different groups of adult albino rats. Values are mean ±S .E. [NM - NH , Normal muscle transplanted in Normal host ; DbM - NH, Diabetic muscle transplanted in normal host ; NM- DbH , Normal muscle transpl anted in diabetic host ; DbM - DbH, Diabetic muscle trans-
planted in diabetic host] .
OJ 1 0.25
0.2
0.15
0.1
0.05
c 7 14 21 28
Days after transpl~ntation
B NM-NH ~DbM-NH ~DbM-DbH E3 NM-DbH
Fig. IS-Histogram representing the changes in m-ATPase acti vity during regeneration of minced gastrocnemius muscle after various intervals in di fferent groups of adult albino rats. Values are mean ±_S.E. [NM- NH , Normal muscle transplanted in Normal host; DbM- NH,Diabetic muscle transplanted in normal host ; NM - DbH, Normal muscle transplanted in diabetic host ; DbM - DbH, Diabetic muscle transplanted in
diabetic host. [There is decrease in SOH and m-ATPase activity on day 7 after the transplantation. The enzymes acti vity thereafter increased almost progressively in all the groups on 14, 2 1 and 28 days after the transplantat ion. Comparatively better regeneration in relation to enzymes SOH and m-ATPase is seen in NM -NH as compared to NM -DbH (P < 0.05 ) on 7, 14 , 21 and 28 days after transplantati on. Simil arl y, better enzymatic activi ty in DbM - NH as compared to DbM - DbH (P < 0.05) on day 7 , 14,2 1, and 28 days after transplantation].

TALESARA & YASHISHTA: REGENERATIVE ABILITY OF GASTROCNEMIUS MUSCLE 573
There IS maximum enzyme activity therefore maximum regeneration and more of regenerating muscle fibres when the diabetic muscle is transplanted in the normal host (DbM-NH) . The results show that the changes in skeletal muscle that accompany diabetes are reyersed when the diabetic muscle is transplanted into normal host.
Regeneration pattern of diabetic muscle into diabetic host (DbM-DbH) was almost similar to that observed for normal muscle into diabetic host (NM-DbH). There is low SDH and m-ATPase activity in the group DbMDH with respect to NM-NH and DbM - NH but slightly higher enzymatic activity than NM - DbH (Figs 14 and 15).
The major differences in the enzymatic activity between the groups DbM-NH and NM-DbH was seen and the likely explanation for thi s is that the diabetic grafts show a very rapid and uniform cycle of degeneration of old muscle fibres and regeneration of new ones whereas normal grafts reacted more slowly therefore there was greater enzymatic activity in the diabetic grafts than in the normal ones.
The acceleration in the regenerative phases of the diabetic muscle has been reported earlier also by many investigators studying minced muscle regeneration39
•40
.
A likely explanation for such a faster rate of regeneration given by Carlson41 is that predenervation or any other trauma increases the population of satellite cells within a muscle thereby increas ing the precurssor population for regeneration and these satellite cells are more resistant to ischemia43
.
Recovery of diabetic muscle after transplantation into normal host provides an evidence that the muscle itself is not permanently affected by the di abetic state and that the outcome of regeneration is primarily determined by the host environment.
Acknowledgement Thanks are due to UGC, New Delhi for the award of
SRF to N.Y. and to Mr. E.A. Daniels for ass istance in photomicrography.
References I Armstrong R B, Gollnick P D & lanuzzo C D, Hi stochemi cal
properties skeletal mu scle fibres in STZ- diabeti c rats. Cell Tissue Res , 162 ( 1975) 387.
2 Armstrong R B & lanuzzo C D, Decay o f succinate dehydrogenase acti vity in rat skeletal muscle fo ll owing STZ injection. Harm Met . Res, 8 ( 1976) 392.
3 Armstrong R B & lanuzzo C D, Co mpensatory hypertroph y o f skeletal mu scle fibres in st reptozotocin d iabeti c rats . Cell Tissue Res, 18 1 ( 1977) 255.
4 Dang A Q, Faas F H & Carter W J, Effects of Streptozotocin induced diabetes on microsome long chain fatty acyl-CoA synthetase and hydrolase. Lipids, 19 ( 1984) 578.
5 Rutschmann , M, Dahlmann B & Reinaur I-1 , Loss of fast twitch iso myosins in skeletal muscles of the di abetic rats. Biochem. J, 221 ( 1984) 645 .
6 Peter J B, Barnard R J, Edgerton V R, Gilli spie A & Stempel K E, Metabolic profile of three fibre types of skeletal mu scle in guinea pigs and rabbits. Biochemistry, 11 ( 1972) 2627.
7 Allbrook D, Skeletal muscle regeneration. Muscle Nerve, 4 (1981) 234.
8 Talesara C L & Shashi Kiran, Cytophysiology of vertebrate skeletal muscle fibres. Indian J Exp Bioi, 24 ( 1986) 331.
9 Cheek D B & Graystone 1 E, Insulin and growth hormone. Regeneration of growth with parti cul ar reference to mu scle. Kidney lnt , 14 ( 1970) 3 17.
10 Trenkle A, Hormonal and nutrit ional interrel ationships and their e ffects o n skeletal mu scle. J Anim Sci, 38 ( 1974) I 142.
II Young V R, Regu lation of protein synthesis and skeletal muscle growth . J Anim Sci, 38 ( 1974) 1054.
12 Allen R E, Marker R A & Young R B, Cellul ar aspect of muscle growth-myogenic cell proliferati on. J Anim Sci, 49 ( 1979) 115 .
13 Je fferson L S, Flaim K E & Peavy D E, in Handbook of diabetes mellitus, 4 ( 198 1) 133.
14 Pain V M & Garli ck P J , Effect of streptozotocin di abetes and insulin treatment on the rate of protei n synthesis in ti ssue of the rat in vivo. J Bioi Chem, 249 ( 1974) 45 10
15 Garlick P J, Albertse E.C, McN url an M A & Pain Y M, Protei n synthesis and breakdown in muscle and ki dney of di abetic and insulin treated rats. Acta Biol Med Ger, 40 ( 198 1) 130 1.
16 Reinil a A & Akerblom H K, Ultrastru cture of heart muscle in short term di abetic rats: Innuence of insulin treatment. Diabetologia, 27 ( 1984) 397 .
17 Barany M., A TPase acti vity o f myosin corre lated with speed of muscle shortening. J Gen Physiol, 50 ( 1967) 197.
18 Jas ra P K & T alesara C L, Effects of alloxan di abetes in innervated and denervated young rat muscles: A correl ati ve hi stochemical and electrophoretic study. Indian J Exp Bio/,24 (1986) 163 .
19 Tal esara C L & Jasra P K, Course o f denervation atrophy in young and adult rat so leus muscles. Indian J Exp Bioi, 24 ( 1986) 274.
20 Gulati A K, Regenerati on, rei nnervatio n and recovery of denervated skeletal muscle after transplantation. Anal. Rec, 223 ( 1989) 46A.
21 Carlson B M & Faulkner 1 A. Reinnervation of long term denervated rat muscle freely gra fted into an innervated limb. Exp Neural, I 02 ( 1988) 50.
22 Denervation of skeletal muscle and dependency of late differenti ation on innervati on. Anal Rec, 220 ( 1988) 429.
23 Talesara C L & Mahant P, SDI-1 & m-ATPase in the denervated gastrocnemius muscle of young adu lt and middle aged albino rats (Rattus norvegicus). A correlative hi stochemical and biochemi cal study. Cell Mol Bioi, 29 ( 1983) 531.
24 George 1 C & Talcsara C L, The SDI-1 levels of pecto ral muscle of a few representative types of birds and bat in rel ation to fibre di ameter, muscle wei ght and body weight. .I Cell Camp Physiol, 58 ( 1961) 283.

574 INDIAN 1 EXP BIOL, JUNE 2000
25 Peter J B, Barnard R J , Edgerton V R , Gilli spie, A & Stempet K E, Metabolic profile of three fibre types o f skeletal muscle in guinea pigs and rabbi ts. Biochemistry, II ( 1972) 2627.
26 Nachlas M M, Tsou K K, DeSouza E, Cheng C S & Seli gman A M, Cytochemical demonstration of succinic dehydrogenase by the use of new para nitrophenyl subst ituted ditet razole. J Histochem Cytochem, 5 ( 1957) 420.
27 Padykul a H A & Herman E, The specifi city of the histochemical method for adenosine triphosphatase. J Histochem. Cytochem, 3 ( 1955) 170.
28 Guth L, Samaha F J, Procedure for the hi stochemical demonstrat ion of acto myosin ATPase. Exp Neural, 28 ( 1970) 365 .
29 Perry S V & Grey T C, A study of effects of substrate concentration and certain releas ing factors o n the magnesium activated myo librillar ATPase. Biochem. J, 64 ( 1956) 184.
30 Rockstei n M. & Herron P W, Colorimetric determin:Jt ion of inorgani c phosphate in microgram qu an titi es. Anal Chem , 23 (195 1) 1500.
3 1 Gornall A G. Bardawill C J & David M M, Determin ati on of serum proteins by means of biurette react ion . .I Bioi Chem, 177 ( 1949) 751.
32 Nachlas M M, Murgulies S I & Seligman A M, A colorimetri c method for the estimati on o f succi nic dehydrogenase acti vi ty. J Bioi Chem, 235 ( 1960) 499.
33 Carlson B M, Regeneratio n of the completely excised g:Jstrocnemius mu scle in the frog and rat fro m minced muscle fragments. J Morpho/, 125 ( 1968) 447.
34 Mastagli a F L, Dawkins R L & Papadimitriou J M, Morphological changes in skeletal muscle after transplantation. J Neural Sci, 25 ( 1975) 227.
35 Snow M H, Metabolic ac ti vity during dege nerati ve and early regenerative stages of minced skeletal muscle. Anat Rec, 176 ( 1973) 185 .
36 Magon D K, Basso n M D & Carlson B M, Hi stochemical patterns of dehydrogenase act ivity in the development of free muscle grafts in the rat. A nat Rec, 199 ( 198 1) 34 1.
37 Riley D A, Hi stochemi cal changes in ATPase act ivity du ring regeneration of adu lt skeletal muscle fibres. Expl Neural, 41 ( 1973) 690.
38 Carl son B M, The regeneration of skeletal muscle - A review. Am J Anal, 137 ( 1978) 11 9.
39 Carlson B M & Gutmann E, Regenerati on in free ly transpl anted intact muscles of the rat. Anal Rec. 183 ( 1975)47.
40 Mong F S F, Hi sto logical and hi stochemical studies on the nervous influence o f minced mu sc le regeneration of triceps surae of the rat. J Morpho/, 15 ( 1977) 45 1.
41 Snow M H, M yogenic cell formati on in regenerat ing rat skeletal muscle injured by mincing. A line structu ral study. A nat Rec, 188 ( 1977) 18 1.