Embed Size (px)
Citation preview

Journal of Magnetic Resonance 216 (2012) 161–168
Contents lists available at SciVerse ScienceDirect
Journal of Magnetic Resonance
journal homepage: www.elsevier .com/locate / jmr
Reduced dimensionality 3D HNCAN for unambiguous HN, CA and N assignmentin proteins
Manoj Kumar Rout a, Pushpa Mishra a, Hanudatta S. Atreya b, Ramakrishna V. Hosur a,c,⇑a Department of Chemical Sciences, Tata Institute of Fundamental Research (TIFR), Mumbai 400 005, Indiab NMR Research Centre, Indian Institute of Science, Bangalore 560 012, Indiac UM-DAE Centre for Excellence in Basic Sciences, Mumbai University Campus, Kalina, Mumbai 400 098, India
a r t i c l e i n f o
Article history:Received 2 November 2011Revised 16 January 2012Available online 8 February 2012
Keywords:Reduced dimensionalityTriplet peak patternBackbone assignmentCheck pointsHNNNMR
1090-7807/$ - see front matter � 2012 Elsevier Inc. Adoi:10.1016/j.jmr.2012.01.022
⇑ Corresponding author at: Department of ChemicFundamental Research (TIFR), Homi Bhabha Road, CoFax: +91 22 22804610.
E-mail address: [email protected] (R.V. Hosur).
a b s t r a c t
We present here an improvisation of HNN (Panchal, Bhavesh et al., 2001) called RD 3D HNCAN for back-bone (HN, CA and 15N) assignment in both folded and unfolded proteins. This is a reduced dimensionalityexperiment which employs CA chemical shifts to improve dispersion. Distinct positive and negative peakpatterns of various triplet segments along the polypeptide chain observed in HNN are retained and theseprovide start and check points for the sequential walk. Because of co-incrementing of CA and 15N, peaksalong one of the dimensions appear at sums and differences of the CA and 15N chemical shifts. Thischanges the backbone assignment protocol slightly and we present this in explicit detail. The perfor-mance of the experiment has been demonstrated using Ubiquitin and Plasmodium falciparum P2 pro-teins. The experiment is particularly valuable when two neighboring amino acid residues have nearlyidentical backbone 15N chemical shifts.
� 2012 Elsevier Inc. All rights reserved.
1. Introduction
Sequence specific resonance assignment of proteins is theessential requirement for structure determination and study ofall the functional behaviors by NMR. During the last two decades,sequential assignments based on heteronuclear triple resonanceexperiments such as HNCA [1], HN(CO)CA [2], HNCO [1], CBCANH[3] and CBCA(CO)NH [4] has become established as a standard ap-proach, and many proteins have been assigned this way [5–8]. Inmost of these cases one bond or two-bond magnetization transfersoccur through the protein backbone and finally the amide protonmagnetization is detected. In recent years, however, the everincreasing demands for rapidity of assignment, and the desire toinvestigate folded, unfolded, partially folded and intrinsically dis-ordered proteins, all of which are seen to be playing importantroles in biological functions, has resulted in an enormous rushfor new methodological developments [9–25].
In this context, HNN and HN(C)N pulse sequences [14,15] haveproved to be very useful. Various improvisations of these experi-ments have also been reported for rapid resonance assignment[16–20]. Triplet specific patterns comprising positive and negativepeaks in these spectra lead to easier walk through the protein
ll rights reserved.
al Sciences, Tata Institute oflaba, Mumbai 400 005, India.
backbone. The main features of the HNN experiment are: (i) corre-lation of ith amide proton with 15N of three adjacent residues i � 1,i and i + 1 in the F3 � F1 plane, (ii) correlation of i � 1, i and i + 1amide protons with their corresponding 15N nuclei in the F3 � F2
plane; this acts as a triplet filter through the HSQC spectrum, (iii)special peak patterns arising from glycine in i or i � 1 positionswhich act as start points or check points during sequential walk,and (iv) absence of peak for proline in a triplet acts as a stop pointin the sequential walk [15]. The success of assignment from HNN isclearly based on good dispersion of the 15N chemical shifts, whichis generally very good in both folded and unfolded proteins. How-ever on rare occasions, poor dispersion in 15N chemical shifts cre-ates overlaps among the triplet peaks in the F3 � F1 plane. This, inturn, causes degenerate correlations in the F3 � F2 planes of theHNN spectrum leading to ambiguous amide proton assignments.Degenerate 15N chemical shifts of i and (i � 1 or i + 1) residuescan lead to cancellation of intensities as well. Hence new modifica-tions of HNN pulse-sequence along with corresponding assign-ment protocol are necessary to overcome this type of situation.
Another important development has been the emergence of a newconcept, namely, co-evolution of two different types of nuclei alongwith reduced dimensionality approach; this makes use of chemicalshifts of two types of nuclei at the same time to disperse the peakswithout actually increasing the experimental time [21–25]. This in-cludes experiments such as (4,2)D HN(COCA)NH, (6D) APSY-seq-HNCOCANH, (7,2)D and (5,2)D APSY described recently. We haveincorporated this strategy in the HNN experiment, wherein Ca spins

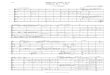
Fig. 1. Schematic of HNCAN pulse sequence. 1H, 15N, 13C and gradient channels are shown in various rows of the figure. Empty and solid rectangular bars represent 90� and180� hard pulses respectively. Empty and solid lobes in carbon channel indicate selective 90� and 180� pulses, respectively. All the pulses not indicated by / are applied withphase x. Proton decoupling is done using waltz-16 decoupling sequence [33,34] with field strength of 6.3 kHz during t1, t2 and 13C evolution period. 15N decoupling using theGARP-1 sequence [35] with the field strength 0.9 kHz is applied during acquisition. The 1H and 15N carrier frequencies are set at 4.71 ppm (water) and 117.5 ppm, respectivelyfor Ubiquitin. The 13C carrier frequency is set at 35 ppm. The strength of the 13Ca pulses (standard Gaussian cascade Q3 (180�) and Q5 (90�) pulses) (Emsley and Bodenhausen,1990) is adjusted so that they cause minimal excitation of carbonyl carbons. The 180� 13CO shaped pulse (width 200 ls) had a standard Gaussian cascade Q3 pulse profilewith minimal excitation of 13Ca The values for the individual periods containing t1 in 3D HNCAN are: A = t1/2, B = TN, and C = TN � t1/2. The values for the individual periodscontaining t2 are: D = TN � t2/2, E = TN, and F = t2/2. The delays are set as follows: k ¼ 2:7 ms, j = 5.4 ms, d = 2.7 ms. Delays containing 13Ca are: 2P = sCN � t2/2 and 2Q = sCN + t2/2. The sCN must be optimized and is around 12–15 ms. The phase cycling for the experiment is: U1 = 2(x), 2(�x); U2 = U3 = x, �x; U4 = 4(x), 4(�x); U5 = x; and Ureceiver = 2(x),4(�x), 2(x). Frequency discrimination in t1 and t2 has been achieved using States-TPPI phase cycling [32] of U1 and U4, respectively, along with the receiver phase. Thegradient (sine-bell shaped; 1 ms) levels are as follows: G1 = 30%, G2 = 30%, G3 = 30%, G4 = G5 = 80%, of the maximum strength of 53 G/cm in the z-direction. WATERGATEsequence [36] has been employed for better water suppression.
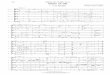
Fig. 2. Schematic diagram showing the magnetization transfer pathway of 3DHNCAN experiment. The flow of magnetization is shown by arrows. t1, t2 and t3 areincrementing time variables in the pulse sequence whereas F1, F2 and F3 are thenomenclature of the three different frequency axes in the 3D spectrum.
162 M.K. Rout et al. / Journal of Magnetic Resonance 216 (2012) 161–168
are made to co-evolve with 15N spins along one of the dimensions.Reduction in dimensionality [22,26–28] is achieved by joint samplingof Ca and 15N in a single indirect dimension of the multidimensionalNMR experiment and thus the resulting signals correspond to linearcombination of the jointly sampled chemical shifts. Due to this infor-mation the peak patterns and the assignment protocol get changedtoo. This experiment is termed as 3D HNCAN and its performancehas been demonstrated for the proteins, human Ubiquitin (BMRB en-try number-15410) and Plasmodium falciparum P2 (BMRB entrynumber-17616). The 3D HNCAN has the particular advantage thatthe dispersion of peaks in the sum and difference frequency regionsare different and this along with positive and negative peak patternshelps resolve ambiguities in situations of overlaps. The special peakpatterns present here in this and other spectra derived from HNNspeed up the analysis quite substantially.
2. Materials and methods
The 3D HNCAN experiment was tested with 1.6 mM uniformly15N/13C labeled human Ubiquitin dissolved in phosphate buffer
pH 6.5 in 90% H2O and 10% D2O (a product from Cambridge IsotopeLaboratories). All NMR experiments were performed at 25 �C on aBruker Avance spectrometer equipped with a CryoProbe, operatingat 1H frequency of 800 MHz. For both HNN and 3D HNCAN exper-iments, the delays 2TN, and 2sCN were set to 30 and 25 ms, respec-tively. 60 complex points were used along F1(15N) and F2(15N)dimensions in case of HNN. In case of 3D HNCAN experiment120 complex points were used in the F2 dimension. Sixteen scanswere recorded per fid. The acquisition time for a 16 scan experi-ment having 120 and 40 complex points in the indirect dimensionsF2 and F3 is 24 h.
The data was processed using NMRpipe [29] and Bruker soft-ware Topspin 2.1 (http://www.bruker.com/) and analyzed usingCARA and [30] SPARKY [31] (http://www.production.rbvi.ucsf.edu/home/sparky/).
3. Results and discussion
3.1. Magnetization transfer in HNCAN experiment
The pulse sequence for 3D HNCAN (Fig. 1) differs slightly fromthe original HNN pulse sequence described previously [14]. The ex-tra frequency labeling of Ca is done during sCN and this is jointlysampled with 15N in the t2 of HNN. Frequency selection along allthe indirect dimensions was done by States-TPPI method [32].The theoretical description of the pulse sequence goes identicallyto that of HNN except that the frequencies along one of the dimen-sions will be sums and differences of frequencies of the jointlysampled nuclei. In our experiments we placed the 13C carrier at35 ppm to achieve best separation of sum and difference frequen-cies. This is a simplistic strategy; it is also possible to use otherstrategies, perhaps better ones, such as TPPI for achieving thisobjective.
The magnetization transfer pathway of 3D HNCAN is shown inFig. 2. The amide magnetization originating from the ith residue is

Fig. 3. Schematic peak patterns in F3 � F2, F2 � F1 and F3 � F1 planes of a triplet sequence (XYZ) having no glycine or proline amino acids. Black and gray colors representpositive and negative peaks. The self and sequential peaks are shown in the respective labels in the figures.
Table 1Position of various peaks that are not seen in presence of proline in F3 � F2 plane of 3DHNCAN experiment.
Position of proline in the triplet Peaks not seen
PYZ ðHi�1;Ni�1 � Cai�1Þ
XYP ðHiþ1;Niþ1 � Cai Þ
M.K. Rout et al. / Journal of Magnetic Resonance 216 (2012) 161–168 163
partly transferred back to itself and the rest transferred to theamides of residues (i � 1) and (i + 1) (see Appendix A for theoreti-cal details). Thus, the peaks appear at the following coordinates:
F1 ¼ Ni; ðF3; F2Þ ! ðHi;Ni � Cai Þ; ðHi;Ni � Ca
i�1Þ ð1ÞðHi�1;Ni�1 � Ca
i�1Þ; ðHiþ1;Niþ1 � Cai Þ ð2Þ
F3 ¼ Hi; ðF1; F2Þ ! ðNi;Ni � Cai Þ; ðNi;Ni � Ca
i�1Þ ð3ÞðNiþ1;Ni � Ca
i Þ; ðNi�1;Ni � Cai�1Þ ð4Þ
F2 ¼ Ni � Cai ; ðF3; F1Þ ! ðHi;NiÞ; ðHi;Niþ1Þ ð5Þ
F2 ¼ Ni � Cai�1; ðF3; F1Þ ! ðHi;NiÞ; ðHi;Ni�1Þ ð6Þ
The schematic peak patterns for different planes are shown inFig. 3. In F3 � F2 plane at F1 = Ni, there are four peaks present (Eq.(1)) at F3 ¼ HN
i corresponding to F2 chemical shifts Ni � Cai and
Ni � Cai�1 (Fig. 3A). The peaks at Ni � Ca
i have higher intensity thanthose at Ni � Ca
i�1 [1] (the relative intensities are not shown infigure). Except these four peaks, two equal intensity peaks areobserved (Eq. (2)) at HN
i�1 and HNiþ1 corresponding to F2 values
Ni�1 � Cai�1 and Niþ1 � Ca
iþ1 respectively. Similarly in F2 � F1 planeat F3 = Hi, there are four peaks present (Eq. (3)) at ðNi;Ni � Ca
i Þand ðNi;Ni � Ca
i�1Þ (Fig. 3B). Two pairs of peaks are also present atðNiþ1;Ni � Ca
i Þ and ðNi�1;Ni � Cai�1Þ (Eq. (4)). Hence from this plane
direct information about sequential connection between Ni, Ni+1
and Ni�1 can be obtained.Like in HNN the F3 � F1 plane also provides direct sequential
correlations in 3D HNCAN (Fig. 3C). At F2 ¼ Ni � Cai there are two
peaks at ðHNi ;NiÞ and ðHN
i ;Niþ1Þ as shown in Eq. (5). Similarly atF2 ¼ Ni � Ca
i�1 there are two peaks corresponding to ðHNi ;NiÞ and
ðHNi ;Ni�1Þ. From all the four F3 � F1 planes at Ni � Ca
i and Ni � Cai�1
the chemical shift values of Ni�1, Ni and Ni+1 can be obtained.
3.2. Peak patterns in presence of glycine and prolines in the triplets ofresidues
The positive and negative peaks in the triplet sequence providespecific peak pattern in the HNN spectrum. Similar peak patternsare also present in 3D HNCAN experiment. The intensities of vari-ous peaks corresponding to self and sequential HN can be derivedfrom product operator methods (see Appendix A; Eqs.(A1)–(A12)). The absence of Cb carbon in glycine changes its mag-netization components compared to non-glycine residues. Thisresults in different triplet peak patterns in presence of glycine atself and sequential positions. The absence of amide proton inproline results in the absence of the corresponding peak. Thus,there will be different patterns of peaks for triplets of residues con-taining glycines and prolines at different positions. The variouspossibilities of patterns containing glycines are: (i) GYZ, (ii) XGZ,(iii) XYG, (iv) GGZ, (v) XGG and (vi) GGG. Schematic representationof various peak patterns in F3 � F2, F2 � F1 and F3 � F1 plane areshown in Fig. S1 A–C, respectively.
Two important points to consider here are the signs of the peakscorresponding to (i) self-amide i.e. HN
i and (ii) sequential amidesi.e. HN
i�1 and HNiþ1. As discussed before for the general case, XYZ,
all the peaks at HNi are positive while the peaks at HN
i�1 and HNiþ1
are negative in the F3 � F2 plane (Fig. 3A). Presence of glycine atith (central) position in the triplet (XGZ) will alter the signs ofpeaks at ðHi;Ni � Ca
i Þ and ðHiþ1;Niþ1 � Cai Þ (Fig. S1A). Glycine at
(i � 1)th position (GYZ) will change the signs of peaks atðHi;Ni � Ca
i�1Þ and ðHi�1;Ni�1 � Cai�1Þ. Glycines at position (i) and
(i � 1) will alter the signs of all the peaks (GGZ). Glycine at position(i + 1) i.e. XYG will be similar to the general case XYZ.
All the above schematic illustrations of different peak patternsfor F3 � F1 and F3 � F2 planes of 3D HNCAN experiment are demon-strated for Ubiquitin in Fig. S2. Fig. S2A shows the peak pattern ofF3 � F2 planes of the fragment 29–36. Residue 35 is a glycine. Thusresidues at locations 36, 35 and 34 behave as central residues inGYZ, XGZ and XYG triplets respectively. Here only the quartet peakpattern at F3 ¼ HN
i is shown. The specific peak patterns of residues35 and 36 are clearly distinguishable from those of others. Fig. S2Bshows the peak pattern of F3 � F1 strips of the fragment 34–36 ofthe same protein. Strips at F2 ¼ Ni � Ca
i and Ni � Cai�1 are shown
here. The sequential connections among Ni, Ni�1 and Ni+1 are quite

Fig. 4. Schematic assignment protocol for three sequentially connected residues: (A) HSQC spectrum. The sequential N15 chemical shifts provided by F2 � F1 plane are shownin (B). These are confirmed by the presence of Self-sequential connected peaks in three F3 � F2 planes at F1 = Ni�1, Ni and Ni+1 (C–E). From these planes the sequential amideproton chemical shifts ðHN
i�1;HNiþ1Þ can be obtained. Black and gray circles represent positive and negative intensity peaks respectively.
164 M.K. Rout et al. / Journal of Magnetic Resonance 216 (2012) 161–168
clear form this figure. The change of peak pattern due to glycineresidue at position 35 is evident in Fig. S1C.
Presence of prolines at various positions in the triplet sequencecan be easily identified by the absence of corresponding peaks asdescribed in Table 1.
3.2.1. Assignment protocolThe direct identification of HN and 15N chemical shifts of neigh-
boring residues provided by 3D HNCAN suggests an easy strategyfor unambiguous assignment of resonances along the protein back-bone. While F2 � F1 plane at F3 ¼ HN
i helps the identification ofCa
i ;Cai�1;Ni�1 and Ni+1 chemical shifts (Figs. 3B and 4B), F3 � F2
planes at F1 = Ni�1, Ni and Ni+1 (Fig. 4C–E) assist to identify HNi�1
and HNiþ1 chemical shifts. The sequential peaks in Fig. 4D are at
the same F2 chemical shifts Niþ1 � Cai as that of the self in Fig. 4E
ðHNiþ1;Niþ1 � Ca
i Þ. This types of self-sequential connections are alsopresent between Fig. 4C and D. An experimental illustration ofthe sequential assignment protocol using Ubiquitin is shown inFig. 5. This is divided into two steps.
Step 1: Obtain information from HSQC (HNi and Ni) for the peak
of interest (ith) (V5 in Fig. 5A).
� The F2 � F1 plane of 3D HNCAN at F3 ¼ HNi shows four peaks in a
strip at F1 = Ni (illustrated in Fig. 5B). These four peaks providethe chemical shift values of ðNi � Ca
i Þ and ðNi � Cai�1Þ. This quar-
tet will be modified to doublet in presence of proline at (i) or(i � 1) position of the triplet fragment.� Special peak pattern due to presence of glycine at (i) or (i � 1)
position can be identified from this quartet.� The chemical shifts of Ca
i and Cai�1 can be calculated from
ððNi � Cai Þ;NiÞ and ððNi � Ca
i�1Þ;NiÞ by using the followingequations.
Cai ¼ ðNi þ Ca
i Þ � Ni ð7ÞCa
i�1 ¼ ðNi þ Cai�1Þ � Ni ð8Þ
However these reflect only deviations from the carrier positionsalong the carbon channel. Actual chemical shifts of Ca can be calcu-lated using the following equation:
XCa ¼ jcN15
cC13X Ca�N15ð Þ �X N15ð Þ� �
þX C13offsetð Þ ð9Þ
� Identification of Ni�1 chemical shifts are obtained as shown ear-lier in Fig. 4B.

Fig. 5. Sequential walk through the peaks in the HSQC of Ubiquitin using 3D HNCAN experiment. The backbone assignment protocol is demonstrated using the tripletsequence F4–V5–K6. Here ith residue is V5 and (i + 1)th is K6 using HN
i and Ni chemical shifts of V5 (A) the corresponding Ni � Cai and Ni � Ca
i�1 values in F2 � F1 plane areobtained (B). From these values Ni+1 (K6) chemical shift is also obtained from this plane (B). Self-sequential connections are confirmed between the two F3 � F2 planes atF1 = Ni (V5) and F1 = Ni+1 (K6). From the sequential peaks the HN
iþ1 (K6) chemical shift is obtained (C). Finally the sequential peak in HSQC is identified as K6 (D).
M.K. Rout et al. / Journal of Magnetic Resonance 216 (2012) 161–168 165
Step-2: F3 � F2 planes at F1 = Ni and F1 = Ni±1 are scanned for self-sequential connections as shown before in Fig. 4 for identificationof HN
iþ1 (K6 in Fig. 5C and D). Finally, with the help of ðHNi�1;Ni�1Þ
chemical shifts the corresponding sequential peaks in HSQC areidentified.
3.2.2. Advantages of 3D HNCAN experimentThough the assignment protocol of 3D HNCAN experiment is lit-
tle long compared to that of HNN there are several advantages ofthe new experiment. Firstly, we get information of Ca
i and Cai�1
chemical shifts. Next, in HNN when self and sequential 15N chem-ical shifts are closer, there is a possibility of overlapping of positive(self) and negative (sequential) peaks in the triplet strip present inthe F3 � F1 plane. Then, due to cancellation of some portions, aclear picture of sequential peak cannot be obtained. This type of
problem can be resolved with the help of F2 � F1 plane along withF3 � F2 planes of 3D HNCAN. Fig. 6 shows the sequential connec-tion between the two peaks T52 and S51 of Plasmodium falcipa-rum P2 protein. The standard protocol for HNN connection isshown in Fig. 6A1–A4. F3 � F1 plane in HNN generally providessequential connection between Ni, Ni+1 and Ni�1. Here, the differ-ence between self and sequential 15N chemical shifts is only0.3 ppm. But the connections are clearly visible in 3D HNCAN inFig. 6B1–B3. In Fig. 6B1 the sequential connection is obtained fromtwo peaks Ni þ Ca
i�1 and Ni � Cai�1. The self-sequential connection
(as in Fig. 4C–4E) among B2 and B3 clearly establishes S51–T52connection. Fig. S3 shows another example of similar type of con-nection in P2 (A108–A109).
Fig. 7 illustrates an example of 3D HNCAN application whenboth the sequential 15N chemical shifts (Ni+1 and Ni�1) are close

Fig. 6. Sequential connection between T52 and S51 in Plasmodium falciparum P2 protein is shown using HNN (A1)–(A4) and 3D HNCAN (B1–B3). Red and green peaks arepositive and negative intensity peaks in the figure.
166 M.K. Rout et al. / Journal of Magnetic Resonance 216 (2012) 161–168
or nearly equal; the triplet in F3 � F1 plane of HNN will be a dou-blet. Here the difference of 15N chemical shift between the twosequential peaks (I97 and Q95) is 0.1 ppm. Due to overlapping ofboth the sequential peaks the triplet of F3 � F1 plane in HNN ismodified to doublet (Fig. 7A2). Hence in this case the connectionsbetween F3 � F1 plane and F3 � F2 plane are degenerate (Fig. 7A3).But using 3D HNCAN sequential connection among this tripletstretch can be unambiguously established (Fig. 7B1–B3). All thefour self-peaks are showing sequential connection in B1 and theseconnections are confirmed by self-sequential connection amongFig. 7B2 and B3 as discussed previously.
4. Conclusions
In this paper we have presented an experiment called 3DHNCAN which incorporates CA and N co-evolution in HNN alongwith reduced dimensionality approach. Because of the CA informa-tion it enhances the peak dispersion in the spectra. The scaling fac-
tor k in the reduced dimensionality step can be optimized toseparate overlapping peaks. This modification brings in additionalfeatures in the backbone assignment protocol. This has been dem-onstrated with Ubiquitin and Plasmodium falciparum P2 proteins.We believe the experiment presented here will find application inproteins with different kinds of complexities.
Acknowledgment
We thank the Government of India for providing financial sup-port to the National Facility for High Field NMR at the Tata Instituteof Fundamental Research.
Appendix A
For any triplet sequence having no glycine or proline the rele-vant density operator terms for 3D HNCAN at the end of secondTN in the pulse sequence is:

Fig. 7. Backbone assignment of two sequential peaks Q95 and I97 having the same 15N chemical shifts as that of the self-peak of N96 are shown using HNN (A1)–(A4) and 3DHNCAN (B1–B3). Out of four peaks corresponding to I97 in B3 only two are within the range displayed. The other two are present outside the chemical shift area of the figure.Due to close chemical shifts of Ca
Q95 and CaL94 the quartet corresponding to Q95 in B3 appears as a doublet. Red and green peaks are positive and negative intensity peaks in the
figure.
M.K. Rout et al. / Journal of Magnetic Resonance 216 (2012) 161–168 167
rAf ¼
��2HizNiyC
21AC3AC9A cosðXNi�Ca
i�1t2Þ
� 2HizNiyC22AC5AC10A cosðXNi�Ca
ðiÞt2Þ
þ2Hði�1ÞzNði�1ÞyC1AC4AC7AC9A cosðXNði�1Þ�Cai�1
t2Þ
þ2Hðiþ1ÞzNðiþ1ÞyC2AC6AC8AC10A cosðXNðiþ1Þ�Cait2Þ�
cosðXNit1Þ
ðA1ÞWhere
C1A ¼ cosð2p1JCai Ni
TNÞ sinð2p2JCai�1Ni
TNÞ ðA2ÞC2A ¼ sinð2p1JCa
i NiTNÞ cosð2p2JCa
i�1NiTNÞ ðA3Þ
C3A ¼ cosð2p1JCai�1Ni�1
sCNÞ cosð2p2JCai�1Ni
sCNÞ ðA4ÞC4A ¼ sinð2p1JCa
i�1Ni�1sCNÞ sinð2p2JCa
i�1NisCNÞ ðA5Þ
C5A ¼ cosð2p1JCai Ni
sCNÞ cosð2p2JCai Niþ1
sCNÞ ðA6ÞC6A ¼ sinð2p1JCa
i NisCNÞ sinð2p2JCa
i Niþ1sCNÞ ðA7Þ
C7A ¼ sinð2p1JCai�1Ni�1
TNÞ cosð2p2JCai�2Ni�1
TNÞ ðA8Þ
C8A ¼ cosð2p1JCaiþ1Niþ1
TNÞ sinð2p2JCai Niþ1
TNÞ ðA9ÞC9A ¼ cosð2p1JCa
i�1Cbi�1
sCNÞ ðA10ÞC10A ¼ cosð2p1JCa
i CbisCNÞ ðA11Þ
and sCN, 2TN = A + B + C = D + E + F are the delays as shown in Fig. 1.The 1JCaN
and 2JCaNin Eqs. (A2)–(A11) are one-bond and two-bond
coupling constants respectively. The first two terms in Eq. (A1) pro-vide four peaks in a strip centered at F3 ¼ HN
i and the third andfourth terms gives four peaks corresponding to the sequential HN
values i.e. two for each F3 ¼ HNi�1 and F3 ¼ HN
iþ1 respectively.For the standard values of 1JCaN and 2JCaN (7–11 Hz), the JCaCb
coupling constant (35–40 Hz), and the commonly used delays(12–16 ms), the functions C1A to C8A are positive while C9A andC10A are negative.
Peak intensities for the different special triplets of residues con-taining glycines and/or prolines in the different planes of the 3DHNCAN can be derived by simple modification of Eq. (A1) and thereare six cases that need to be considered explicitly: (i) GYZ; (ii) XGZ;(iii) XYG; (iv) GGZ; (v) XGG and (vi) GGG. In all the above six casesith position is the middle of the triplet sequence. In case of glycine

168 M.K. Rout et al. / Journal of Magnetic Resonance 216 (2012) 161–168
the 1JCai Cb
icoupling constant will be zero, which in turn makes Eqs.
(A10) and (A11) positive depending upon the position of the gly-cine in the triplet. Thus for all the above cases Eq. (A1) will be mod-ified as follows:
(i)
rAf ¼
�� 2HizNiyC
21AC3A cosðXNi�Ca
i�1t2Þ � 2HizNiyC
22AC5AC10A
cosðXNi�CaðiÞ
t2Þ þ 2Hði�1ÞzNði�1ÞyC1AC4AC7A cosðXNði�1Þ�Cai�1
t2Þ
þ 2Hðiþ1ÞzNðiþ1ÞyC2AC6AC8AC10A cosðXNðiþ1Þ�Cait2Þ�
cosðXNit1Þ ðA12Þ
(ii)
rAf ¼
��2HizNiyC
21AC3AC9A cosðXNi�Ca
i�1t2Þ�
2HizNiyC22AC5A cosðXNi�Ca
ðiÞt2Þ þ 2Hði�1ÞzNði�1ÞyC1AC4AC7AC9A
� cosðXNði�1Þ�Cai�1
t2Þ þ 2Hðiþ1ÞzNðiþ1ÞyC2AC6AC8A
� cosðXNðiþ1Þ�Cait2Þ�
cosðXNit1Þ ðA13Þ
(iii)
rAf ¼��2HizNiyC
21AC3AC9A cosðXNi�Ca
i�1t2Þ�
2HizNiyC22AC5AC10A cosðXNi�Ca
ðiÞt2Þ
þ2Hði�1ÞzNði�1ÞyC1AC4AC7AC9A cosðXNði�1Þ�Cai�1
t2Þ
þ2Hðiþ1ÞzNðiþ1ÞyC2AC6AC8AC10A cosðXNðiþ1Þ�Cait2Þ�
cosðXNit1Þ ðA14Þ
(iv)
rAf ¼
��2HizNiyC
21AC3A cosðXNi�Ca
i�1t2Þ � 2HizNiyC
22AC5A cos
ðXNi�CaðiÞ
t2Þ þ 2Hði�1ÞzNði�1ÞyC1AC4AC7A cosðXNði�1Þ�Cai�1
t2Þ
þ 2Hðiþ1ÞzNðiþ1ÞyC2AC6AC8A cosðXNðiþ1Þ�Cait2Þ�
cosðXNit1Þ ðA15Þ
(v)
rAf ¼
��2HizNiyC
21AC3AC9A cosðXNi�Ca
i�1t2Þ � 2HizNiyC
22AC5A cos
ðXNi�CaðiÞ
t2Þ þ 2Hði�1ÞzNði�1ÞyC1AC4AC7AC9A cosðXNði�1Þ�Cai�1
t2Þ
þ 2Hðiþ1ÞzNðiþ1ÞyC2AC6AC8A cosðXNðiþ1Þ�Cait2Þ�
cosðXNit1Þ ðA16Þ
(vi)
rAf ¼
��2HizNiyC
21AC3A cosðXNi�Ca
i�1t2Þ � 2HizNiyC
22AC5A cos
ðXNi�CaðiÞ
t2Þ þ 2Hði�1ÞzNði�1ÞyC1AC4AC7A cosðXNði�1Þ�Cai�1
t2Þ
þ 2Hðiþ1ÞzNðiþ1ÞyC2AC6AC8A cosðXNðiþ1Þ�Cait2Þ�
cosðXNit1Þ ðA17Þ
Appendix B. Supplementary material
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.jmr.2012.01.022.
References
[1] L.E. Kay, M. Ikura, R. Tschudin, A. Bax, 3-Dimensional triple-resonance NMR-spectroscopy of isotopically enriched proteins, J. Magn. Reson. 89 (1990) 496–514.
[2] M. Ikura, L.E. Kay, A. Bax, Improved three-dimensional 1H 13C-1H correlationspectroscopy of a 13C-labeled protein using constant-time evolution, J. Biomol.NMR 1 (1991) 299–304.
[3] S. Grzesiek, A. Bax, An efficient experiment for sequential backboneassignment of medium-sized isotopically enriched proteins, J. Magn. Reson.99 (1992) 201–207.
[4] S. Grzesiek, A. Bax, Correlating backbone amide and side-chain resonances inlarger proteins by multiple relayed triple resonance NMR, J. Am. Chem. Soc.114 (1992) 6291–6293.
[5] N. Morelle, J.P. Simorre, M. Caffrey, T. Meyer, M. Cusanovich, D. Marion, 1H and13C NMR assignment and secondary structure of Chlorobium limicola f.thiosulfatophilum ferrocytochrome c555, FEBS Lett. 365 (1995) 172–178.
[6] M. Caffrey, J.P. Simorre, B. Brutscher, M. Cusanovich, D. Marion, NMRassignment of Rhodobacter capsulatus ferricytochrome c’, a 28 kDaparamagnetic heme protein, Biochemistry 34 (1995) 5904–5912.
[7] M.L. Remerowski, T. Domke, A. Groenewegen, H.A.M. Pepermans, C.W. Hilbers,F.J.M. Vandeven, H-1, C-13 and N-15 NMR backbone assignments andsecondary structure of the 269-residue protease subtilisin-309 from Bacilluslentus, J. Biomol. NMR 4 (1994) 257–278.
[8] M.F. Leopold, J.L. Urbauer, A.J. Wand, Resonance assignment strategies for theanalysis of NMR spectra of proteins, Mol. Biotechnol. 2 (1994) 61–93.
[9] K. Pervushin, R. Riek, G. Wider, K. Wuthrich, Transverse relaxation-optimizedspectroscopy (TROSY) for NMR studies of aromatic spin systems in C-13-labeled proteins, J. Am. Chem. Soc. 120 (1998) 6394–6400.
[10] B.R. Donald, J. Martin, Automated NMR assignment and protein structuredetermination using sparse dipolar coupling constraints, Prog. Nucl. Magn.Reson. Spectrosc. 55 (2009) 101–127.
[11] P. Permi, A. Annila, Coherence transfer in proteins, Prog. Nucl. Magn. Reson.Spectrosc. 44 (2004) 97–137.
[12] P. Schanda, H. Van Melckebeke, B. Brutscher, Speeding up three-dimensionalprotein NMR experiments to a few minutes, J. Am. Chem. Soc. 128 (2006)9042–9043.
[13] D. Rovnyak, D.P. Frueh, M. Sastry, Z.Y.J. Sun, A.S. Stern, J.C. Hoch, G. Wagner,Accelerated acquisition of high resolution triple-resonance spectra using non-uniform sampling and maximum entropy reconstruction, J. Magn. Reson. 170(2004) 15–21.
[14] S.C. Panchal, N.S. Bhavesh, R.V. Hosur, Improved 3D triple resonanceexperiments, HNN and HN(C)N, for HN and15N sequential correlations in(13C,15N) labeled proteins: application to unfolded proteins, J. Biomol. NMR 20(2001) 135–147.
[15] N.S. Bhavesh, S.C. Panchal, R.V. Hosur, An efficient high-throughput resonanceassignment procedure for structural genomics and protein folding research byNMR, Biochemistry 40 (2001) 14727–14735.
[16] J. Chugh, D. Kumar, R.V. Hosur, Tuning the HNN experiment: generation ofserine–threonine check points, J. Biomol. NMR 40 (2008) 145–152.
[17] A. Chatterjee, A. Kumar, R.V. Hosur, Alanine check points in HNN and HN(C)Nspectra, J. Magn. Reson. 181 (2006) 21–28.
[18] D. Kumar, S. Paul, R.V. Hosur, BEST-HNN and 2D-(HN)NH experiments forrapid backbone assignment in proteins, J. Magn. Reson. 204 (2010) 111–117.
[19] D. Kumar, J.G. Reddy, R.V. Hosur, hnCOcaNH and hncoCANH pulse sequencesfor rapid and unambiguous backbone assignment in ((13)C, (15)N) labeledproteins, J. Magn. Reson. 206 (2010) 134–138.
[20] A. Borkar, D. Kumar, R.V. Hosur, AUTOBA: automation of backbone assignmentfrom HN(C)N suite of experiments, J. Biomol. NMR 50 (2011) 285–297.
[21] H.S. Atreya, T. Szyperski, Rapid NMR data collection, Methods Enzymol. (2005)78–108.
[22] T. Szyperski, D.C. Yeh, D.K. Sukumaran, H.N.B. Moseley, G.T. Montelione,Reduced-dimensionality NMR spectroscopy for high-throughput proteinresonance assignment, Proc. Natl Acad. Sci. USA 99 (2002) 8009–8014.
[23] D. Pantoja-Uceda, J. Santoro, Aliasing in reduced dimensionality NMR spectra:(3,2)D HNHA and (4,2)D HN(COCA)NH experiments as examples, J. Biomol.NMR 45 (2009) 351–356.
[24] F. Fiorito, S. Hiller, G. Wider, K. Wuthrich, Automated resonance assignment ofproteins: 6D APSY-NMR, J. Biomol. NMR 35 (2006) 27–37.
[25] R.L. Narayanan, U.H.N. Durr, S. Bibow, J. Biernat, E. Mandelkow, M.Zweckstetter, Automatic assignment of the intrinsically disordered proteintau with 441-residues, J. Am. Chem. Soc. 132 (2010) 11906–11907.
[26] T. Szyperski, D. Braun, C. Fernandez, C. Bartels, K. Wuthrich, A novel reduced-dimensionality triple-resonance experiment for efficient polypeptide backboneassignment, 3D-COHNNCA, J. Magn. Reson. Ser. B 108 (1995) 197–203.
[27] T. Szyperski, M. Pellecchia, K. Wuthrich, 3d Ha/bCa/b (Co)NHN, a projected 4dNMR experiment for sequential correlation of polypeptide 1Ha/b, 13Ca/b andbackbone 15N and 1HN chemical-shifts, J. Magn. Reson. B 105 (1994) 188–191.
[28] T. Szyperski, G. Wider, J.H. Bushweller, K. Wuthrich, Reduced dimensionality intriple-resonance NMR experiments, J. Am. Chem. Soc. 115 (1993) 9307–9308.
[29] F. Delaglio, S. Grzesiek, G.W. Vuister, G. Zhu, J. Pfeifer, A. Bax, Nmrpipe – amultidimensional spectral processing system based on unix pipes, J. Biomol.NMR 6 (1995) 277–293.
[30] R. Keller, The Computer Aided Resonance Assignment Tutorial, 2004.[31] T.D. Goddard, D.G. Kneller, SPARKY 3.[32] D. Marion, M. Ikura, R. Tschudin, A. Bax, Rapid recording of 2D NMR-spectra
without phase cycling – application to the study of hydrogen-exchange inproteins, J. Magn. Reson. 85 (1989) 393–399.
[33] A.J. Shaka, J. Keeler, R. Freeman, Evaluation of a new broad-band decouplingsequence – Waltz-16, J. Magn. Reson. 53 (1983) 313–340.
[34] A.J. Shaka, J. Keeler, T. Frenkiel, R. Freeman, An improved sequence for broad-band decoupling – Waltz-16, J. Magn. Reson. 52 (1983) 335–338.
[35] A.J. Shaka, Composite pulses for ultra-broadband spin inversion, Chem. Phys.Lett. 120 (1985) 201–205.
[36] M. Piotto, V. Saudek, V. Sklenar, Gradient-tailored excitation for single-quantum NMR-spectroscopy of aqueous-solutions, J. Biomol. NMR 2 (1992)661–665.