Embed Size (px)
Citation preview

Reconstitution of TCA cycle with DAOCS to engineerEscherichia coli into an efficient whole cell catalystof penicillin GBaixue Lina, Keqiang Fanb, Jian Zhaoa, Junjie Jib, Linjun Wub, Keqian Yangb,1, and Yong Taoa,1
aChinese Academy of Sciences Key Laboratory of Microbial Physiological and Metabolic Engineering, Institute of Microbiology, Chinese Academy ofSciences, Beijing 100101, People’s Republic of China; and bState Key Laboratory of Microbial Resources, Institute of Microbiology, Chinese Academy ofSciences, Beijing 100101, People’s Republic of China
Edited by Arnold L. Demain, Drew University, Madison, NJ, and approved June 25, 2015 (received for review February 10, 2015)
Many medically useful semisynthetic cephalosporins are derivedfrom 7-aminodeacetoxycephalosporanic acid (7-ADCA), which hasbeen traditionally made by the polluting chemical method. Here, awhole-cell biocatalytic process based on an engineered Escherichiacoli strain expressing 2-oxoglutarate–dependent deacetoxycephalo-sporin C synthase (DAOCS) for converting penicillin G to G-7-ADCAis developed. The major engineering strategy is to reconstitute thetricarboxylic acid (TCA) cycle of E. coli to force the metabolic flux togo through DAOCS catalyzed reaction for 2-oxoglutarate to succi-nate conversion. Then the glyoxylate bypass was disrupted to elim-inate metabolic flux that may circumvent the reconstituted TCAcycle. Additional engineering steps were taken to reduce the deg-radation of penicillin G and G-7-ADCA in the bioconversion process.These steps include engineering strategies to reduce acetate accu-mulation in the biocatalytic process and to knock out a host β-lacta-mase involved in the degradation of penicillin G and G-7-ADCA.By combining these manipulations in an engineered strain, the yieldof G-7-ADCA was increased from 2.50 ± 0.79 mM (0.89 ± 0.28 g/L,0.07 ± 0.02 g/gDCW) to 29.01 ± 1.27 mM (10.31 ± 0.46 g/L, 0.77 ±0.03 g/gDCW) with a conversion rate of 29.01 mol%, representingan 11-fold increase compared with the starting strain (2.50 mol%).
DAOCS | TCA cycle | reconstitution | whole-cell catalyst | G-7-ADCA
Most clinically important semisynthetic cephalosporins aremanufactured from 7-aminodeacetoxycephalosporanic acid
(7-ADCA) or 7-aminocephalosporanic acid (7-ACA). The tradi-tional chemical process to produce 7-ADCA by ring expansion ofpenicillin G is expensive and pollutive (1, 2); thus, a bioconversionprocess is highly desirable. To this end, a process based on fer-mentation of Penicillium chrysogenum transformants expressing cefE(encoding deacetoxycephalosporin C synthase from Streptomycesclavuligerus, scDAOCS) in culture medium supplemented withadipate was first developed to produce adipyl 7-ADCA (3), whichcan be further converted to 7-ADCA by glutaryl acylase (3). Asecond process is based on the expression of the cefE gene fromS. clavuligerus in a cefEF-disrupted Acremonium chrysogenum; theengineered strain can convert penicillin N accumulated in the cefEFdisrupted mutant into deacetoxycephalosporin C (DAOC), whichcan be deacylated to 7-ADCA (1). Alternatively, a bioconversionprocess that directly converts penicillin G to G-7-ADCA by DAOCShad also been considered and investigated (4). This process holdspromise to be the most commercially viable route to produce7-ADCA. Imperative to the success of this strategy is the develop-ment of mutant DAOCS enzymes that can efficiently catalyze theconversion of the cheap substrate, penicillin G, to G-7-ADCA.The scDAOCS from S. clavuligerus (also known as penicillin
expandase) is the key enzyme that catalyzes the ring expansion ofpenicillin substrates into the corresponding cephalosporins (5, 6). Itis a Fe (II)- and 2-oxoglutarate (2OG)-dependent oxidase (Fig. 1A).Several structures of DAOCS had been reported (7, 8). Presteady-state kinetics and binding studies of DAOCS (9) suggested thatDAOCS employs a mechanism similar to other 2OG-dependent
oxygenases, where the binding of 2OG occurs first with subsequentformation of a ternary DAOCS·Fe(II)·2OG·penicillin substratecomplex (9). The natural substrate of DAOCS is penicillin N, andits activity toward other penicillin analogs, such as penicillin G, islow (4). To apply DAOCS in a bioconversion process, great effortshad been devoted to improving the catalytic activity of DAOCStoward penicillin G (10). These works led to the discoveries ofmany improved mutants with increased activity toward penicillin G(10). In this study, we chose a scDAOCS mutant (H7) (2) withsignificantly increased activity toward penicillin G (22.48-fold forspecific activity and 80.74-fold for kcat/Km), as the biocatalyst todevelop a bioconversion process of penicillin G in Escherichia coli.Previously, Cho et al. (4) reported successful bioconversion ofpenicillin G to G-7-ADCA using resting cells and extracts ofS. clavuligerus NP1. Further studies revealed that the additionof ethanol or catalase stimulated the bioconversion of penicillinG (11, 12). Heterologous expression of DAOCS in Streptomyceslividans W25 resulted in significantly increased bioconversion ofpenicillin G (0.24 g/L after 24 h at the optimized condition, with10 g/L penicillin G) (13). Whole-cell bioconversion is preferredbecause it does not require the exogenous addition of cosubstrates,such as, 2OG. In this work, we considered E. coli as a whole-cellcatalyst of DAOCS catalyzed reaction because it is a well-characterized host with a clean genetic background.Because DAOCS catalyzed reaction needs 2OG as a cosub-
strate, which is a key intermediate of the tricarboxylic acid (TCA)cycle, the efficiency of this reaction depends on the amount of
Significance
We developed Escherichia coli expressing deacetoxycephalosporinC synthase (DAOCS) as a whole-cell biocatalyst to convert peni-cillin G to G-7-aminodeacetoxycephalosporanic acid (G-7-ADCA).The major strategy used was to reconstitute the tricarboxylic acid(TCA) cycle of E. coli with DAOCS catalyzed reaction; thus themetabolic flux of central metabolism was forced to go throughDAOCS catalyzed reaction to produce G-7-ADCA. This strategywas combined with engineering efforts to reduce the accumula-tion of acetate and the degradation of penicillin G and G-7-ADCAto improve the conversion rate of penicillin G by DAOCS signifi-cantly. Therefore, this work demonstrates the feasibility to re-direct the TCA cycle to drive a desired enzyme reaction, and thisstrategy could be applied to other enzymes that catalyze TCAcycle-coupleable reactions.
Author contributions: B.L., J.Z., K.Y., and Y.T. designed research; B.L., J.Z., and J.J. per-formed research; K.F., J.J., and L.W. contributed new reagents/analytic tools; B.L. and K.F.analyzed data; and B.L., K.F., K.Y., and Y.T. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.1To whom correspondence may be addressed. Email: [email protected] or [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1502866112/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1502866112 PNAS | August 11, 2015 | vol. 112 | no. 32 | 9855–9859
APP
LIED
BIOLO
GICAL
SCIENCE
S
Dow
nloa
ded
by g
uest
on
Apr
il 7,
202
0

2OG that can be diverted from the TCA cycle. In the TCA cycle(Fig. 1B), 2OG is converted to succinyl-CoA and CO2 by 2OGdehydrogenase multienzyme complex (SucAB in E. coli), then tosuccinate by succinyl-CoA synthetase (SucCD). We envision thatif the TCA cycle were disrupted at the 2OG to the succinate step,the TCA cycle can then be forced to go through a DAOCScatalyzed reaction. Such an outcome is a major goal of thiswork: that is, to reconstitute the TCA cycle with a DAOCScatalyzed reaction.
ResultsBioconversion of Penicillin G to Produce G-7-ADCA by E. coli ExpressingscDAOCS. First, the feasibility of in vivo bioconversion of penicillinG by heterologous expressing DAOCS in E. coli without an ex-ogenous supply of 2OG was investigated. An E. coli K12 strainBW25113 containing an araBAD deletion was chosen as the bio-conversion host, in which the scDAOCS gene can be expressedfrom the pBAD promoter after induction with L-arabinose. AscDAOCS mutant (M73T/C155Y/Y184H/T213V/V275I/C281Y/I305M) with improved activity toward penicillin G (2) was used asthe catalytic enzyme. Recombinant E. coli strain BW/H7 was con-structed by transforming plasmid pDB1S-H7 into BW25113 forexpressing scDAOCS-H7. In BW/H7, scDAOCS-H7 was mainlyexpressed as a soluble protein. BW/H7 cells after induction werecollected and suspended in the reaction mixture containing 100 mMpenicillin G. After 5-h incubation at 30 °C, 2.50 ± 0.79 mMG-7-ADCA was formed, with a conversion rate of 2.5 mol%.The feasibility of bioconversion of penicillin G in an E. coliwithout an exogenous supply of 2OG was thus validated.
Reconstitution of the TCA Cycle with DAOCS Catalyzed Reaction. TheDAOCS catalyzed reaction needs 2OG as a cosubstrate (Fig. 1A),which is an important intermediate metabolite in the TCA cycle.Thus, the bioconversion reaction is expected to compete with the
TCA cycle for 2OG. To redirect the metabolic flux toward theDAOCS catalyzed reaction, the TCA cycle was engineeredin vivo by blocking the normal TCA reaction of 2OG to succi-nate, and thus forcing the TCA cycle to compensate by using theDAOCS catalyzed reaction (Fig. 2). In E. coli, the SucAB andSucCD are responsible for the transformation of 2OG to succi-nate, whereas AceA is the enzyme involved in the glyoxylatebypass of an incomplete TCA cycle. To engineer the metabolicflux of the TCA cycle, genes encoding SucA, SucB, SucC, SucD,and AceA were knocked out separately in E. coli BW25113, andpDB1S-H7 was transformed into these engineered strains, re-spectively. In the resulting recombinant strains, DAOCS cata-lyzed reaction was forced to compensate for the transformationof 2OG to succinate; as such, the TCA cycle was reconstructed inthese E. colimutants. The effects of the reconstructed TCA cycleon G-7-ADCA production were analyzed. The yields of G-7-ADCA by PG01/H7 (△sucA), PG02/H7 (△sucB), PG04/H7(△sucC), PG06/H7 (△sucD), and PG08/H7 (△aceA) was 10.04 ±0.77, 4.19 ± 0.73, 3.59 ± 0.75, 1.73 ± 0.73, and 3.79 ± 0.76 mM,respectively, and most strains showed higher production ofG-7-ADCA than that of the starting strain BW/H7 (Fig. 3A).Among these mutants, the △sucA mutant showed the most ob-vious effect of increasing the yield of G-7-ADCA, in which theconversion of 2OG to succinyl-CoA was blocked. Deletion ofaceA, the gene encoding the key enzyme for glyoxylate bypass, alsocaused a moderate increase in G-7-ADCA yield. These resultsrevealed that the TCA cycle and the glyoxylate bypass can beengineered to adopt an alternative route without having significantimpact on cell physiology.
Reduce Acetate Accumulation by △poxB::acs to Improve the Bio-conversion of G-7-ADCA. Penicillin G and G-7-ADCA are suscep-tible to decomposition in an acidic environment. A decrease inacetate production will not only prevent acidification of the re-action medium to maintain a more stable pH environment for theDAOCS bioconversion reaction, but will also avoid the carbon lossfrom the system, increasing the availability of acetyl CoA to drivethe bioconversion of G-7-ADCA. To reduce acetate production, acombination of knocking out the pyruvate oxidase gene (poxB)
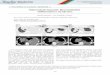
Fig. 1. Reconstitution of TCA cycle using DAOCS catalyzed reaction. (A) DAOCScatalyzed reaction to convert penicillin G to G-7-ADCA, which is coupled with theconversion of 2OG to succinate to compensate the disrupted TCA cycle. (B) Themanipulations to disrupt the TCA cycle and glyoxylate bypass.
Fig. 2. The combined manipulations to convert E. coli BW25113 into anefficient whole-cell biocatalyst for penicillin G transformation.
9856 | www.pnas.org/cgi/doi/10.1073/pnas.1502866112 Lin et al.
Dow
nloa
ded
by g
uest
on
Apr
il 7,
202
0

and overexpressing of acetyl-CoA synthetase (Acs) was applied asreported by Tao et al. (14). E. coli PG03/H7 (△sucA△poxB::acs)was constructed and evaluated for its bioconversion of penicillinG. In this recombinant strain, the accumulation of acetate wasreduced by 38.3% (Fig. 3B), and G-7-ADCA production wasfurther increased by 11% to 11.10 ± 0.70 mM, similar to that ofPG01/H7. It is worth noting that the growth of PG03/H7 wasbetter than that of PG01/H7.
The Effects of Deregulation of Glyoxylate Pathway and Increased TCAFlux on DAOCS Catalyzed Reaction. IclR is a repressor of theE. coli aceBAK operon, which encodes enzymes of the glyoxylatepathway (Fig. 2). When IclR is inactivated, repression of the ex-pression glyoxylate genes is released, and the glyoxylate pathway isactivated. This strategy could be used to reduce the flux throughthe TCA cycle (15). PG05/H7 (△sucA△poxB::acs△aceA) andPG07/H7 (△sucA△poxB::acs△iclR) were constructed to in-vestigate the effects of increasing glyoxylate pathway on DAOCScatalyzed reactions. The production of G-7-ADCA by PG05/H7and PG07/H7 was 12.19 ± 0.92 and 8.79 ± 0.24 mM, with aconversion rate of 12.19 and 8.79 mol%, respectively. These re-sults were consistent with the prediction that increased fluxthrough the TCA cycle (PG05/H7) is beneficial to G-7-ADCAproduction, because deletion of aceA should reduce the flux of theglyoxylate pathway and increase that of the TCA cycle (16),whereas inactivation of iclR (PG07/H7) should have opposite ef-fects. The small changes observed suggest that glyoxylate bypassonly had a minor contribution to the catabolism of 2OG. Thus, bypushing the flux of the TCA cycle, the DAOCS catalyzed reactioncould be forced to increase (Fig. 3C).To test this hypothesis further, we increased the supply of glu-
cose as a simple mean to increase the flux of the TCA cycle. Whenglucose is taken up by E. coli, it is first broken down into pyruvateby glycolysis; pyruvate is then further converted to acetyl-CoA,which then enters the TCA cycle. Our results showed that dif-ferent concentrations of glucose indeed had a significant impacton G-7-ADCA production in the PG01/H7 (△sucA) strain. As
the concentration of glucose increased from 0.1 to 0.5%, theproduction of G-7-ADCA increased in concert (Fig. 4 A and B).For glucose concentrations higher than 0.5% (such as 1–2%wt/vol), no significant increase of G-7-ADCA production wasobserved (Fig. 4 C and D). Such effects of glucose on G-7-ADCAproduction were not found in BW/H7, in which the TCA cycle isnot coupled with the DAOCS catalyzed reaction. These resultsfurther demonstrated that the flux of the TCA cycle can be cou-pled with the DAOCS catalyzed reaction, and by pushing the TCAcycle, the efficiency of the whole-cell biocatalysis for G-7-ADCAproduction can be improved.
Knocking Out ampC to Reduce the Decomposition of Penicillin G andG-7-ADCA. Decomposition of penicillin G and G-7-ADCA (17)was observed during the bioconversion process in this study.When cell lysis solution (10 OD cell-free extract in 100 mM Trisbuffer) of E. coli strain BW25113 was incubated with penicillinG and G-7-ADCA, both were degraded within several hours(Fig. S1). The gene encoding β-lactamase (AmpC) was knockedout in BW25113 and PG01 to obtain PG12/H7 (△ampC) andPG14/H7 (△sucA△ampC) to avoid this problem. Strain PG12/H7expressing scDAOCS-H7 produced on average 9.63 ± 1.01 mMG-7-ADCA, with a conversion rate of 9.63 mol%, thatwas 3.9-fold higher than that of the starting strain BW/H7(2.50 ± 0.79 mM). The yield of G-7-ADCA of PG14/H7(△sucA△ampC) expressing scDAOCS H7 was about 17.88 ±2.55 mM, with a conversion rate of 17.88 mol%, significantlyhigher than that of the starting strain and PG01/H7 (Fig. 3D).These results clearly indicated that AmpC played an importantrole in the decomposition of penicillin G and G-7-ADCA in thisbioconversion process.
The Effects of Combining the Beneficial Mutations on G-7-ADCAProduction. Finally, the aforementioned beneficial mutations werecombined in an engineered strain. Mutations of sucA and aceAshould totally block succinate supply and result in an increased 2OGaccumulation, whereas mutation of poxB and overexpression of theacetyl-CoA synthetase gene (acs) should reduce acetate accumula-tion, and ampC knockout eventually prevent the decomposition of
Fig. 3. A series of engineering strategies used to reconstitute TCA cycle ofE. coli with DAOCS catalyzed reaction. (A) Effects of blocking the pathway of2OG to succinate in BW on G-7-ADCA production. (B) Effects of engineeringacetate metabolism on G-7-ADCA and acetate production during bio-conversion. (C) Effects of engineering glyoxylate bypass on G-7-ADCA pro-duction. (D) Effects of ampC knockout on G-7-ADCA production. Theparental strain E. coli BW25113 and its mutants transformed with pDB1S-H7were cultured at 30 °C for 16 h and the cells were suspended in reactionmixture containing 100 mM penicillin G as substrate for G-7-ADCA pro-duction. The bioconversion reactions were performed at 30 °C and 200 rpmfor 20 h.
Fig. 4. (A–D) Effects of glucose concentrations on G-7-ADCA production.The recombinant strains of BW/H7, PG01/H7, and PG03/H7 were cultured at30 °C for 16 h and the cells were suspended in reaction mixture containing100 mM penicillin G supplemented with 0.1–2% (wt/vol) glucose for G-7-ADCA production. The bioconversion reactions were carried out at 30 °C and200 rpm for 8 h. Samples was withdrawn every 2 h and subjected to analysis.
Lin et al. PNAS | August 11, 2015 | vol. 112 | no. 32 | 9857
APP
LIED
BIOLO
GICAL
SCIENCE
S
Dow
nloa
ded
by g
uest
on
Apr
il 7,
202
0

penicillin G and G-7-ADCA. The engineered strain PG22/H7(△sucA△poxB::acs△ampC△aceA) contains all these beneficialtraits. For biotransformation by most of the mutant strains, 20 hwere long enough for the production levels of G-7-ADCA toreach a plateau (Fig. S2). When we compared all 20 recombi-nant mutants in a bioconversion experiment than was run inparallel (Table S1), PG22/H7 showed the highest yield of G-7-ADCA around 29.01 ± 1.27 mM, with a conversion rate 29.01mol%, followed by PG15/H7 (△sucA△poxB::acs△ampC) around21.48 ± 1.08 mM, with a conversion rate of 21.48 mol% (Fig. 5A–C). Overall, the bioconversion rate increased more than 10-foldfrom 2.50 mol% to 29.01 mol%.
DiscussionDAOCS catalyzes a ring expansion reaction to convert penicillinsinto cephalosporins; the reaction needs 2OG as a cosubstrate,which undergoes oxidative decarboxylation to form succinate. Anin vitro process to carry out such a reaction would require theexogenous supply of 2OG that is a cost burden to the process; inaddition the accumulation of the coproduct succinate could inhibitthe bioconversion rate of the process (9). An in vivo process usingE. coli as a whole-cell catalyst can provide an ideal solution tothese problems because 2OG can be supplied in E. coli cells, andsuccinate will automatically be consumed without accumulation,thus effectively removing its inhibition of the DAOCS catalyzedreaction. However, 2OG is normally metabolized via the TCAcycle; little flux normally goes into the route of producing ofG-7-ADCA. To solve this problem, the TCA cycle of the hostwas forced to use the DAOCS catalyzed reaction: the recon-stituted TCA cycle consumed 2OG and produced succinate andG-7-ADCA, as desired. This restores the normal function of theTCA cycle, in which succinate is further recycled into other TCAintermediates. In the TCA cycle, 2OG is converted to S-succi-nyldihydrolipoamide-E by SucA, the first enzyme catalyzing theconversion from 2OG to succinate. Whereas in sucB, sucC, andsucD single-knockout mutants, S-succinyldihydrolipoamide-Eproduced by SucA can be further metabolized to succinate by
alternative reactions; therefore, only sucA mutation can com-pletely block the conversion of 2OG to succinate; as expected,the sucA mutant showed the highest penicillin G conversionrate compared with that of sucBCD mutants.E. coli K12 is often used as a cell factory for the production
metabolites, because it is a nonpathogenic bacterium with a well-studied genetic background and a powerful genetic tool systemfor metabolic engineering. Therefore, it was chosen as host totest our hypothesis of reconstituting the TCA using a DAOCScatalyzed reaction. The reconstituted TCA cycle indeed workedas predicted, and the flux can be powered by the addition ofglucose. Although there are penicillin-binding proteins in E. coli(18), the amount of penicillin G bound to the cell wall was smallin comparison with the amount supplied and penicillin G coulddiffuse into the cell efficiently (Fig. S3).Penicillin G and G-7-ADCA are susceptible to decomposition
in an acidic environment. Because acetate accumulation oftenoccurs in bioprocesses when high concentration of glucose wasused, reduction of acetate formation can help prevent the de-composition of penicillin G and G-7-ADCA in the penicillinG bioconversion process. On the other hand, the lost carbonresulting from acetate formation reduces the flux of acetyl-CoAentering the TCA cycle. Taking the carbon balance in the systeminto consideration, decrease in acetate production will not onlyprevent acidification of the reaction medium but also reduce thecarbon loss from the system, which may increase the availabilityof acetyl-CoA to drive the G-7-ADCA production. There aretwo major acetate-producing pathways in aerobically grownE. coli: the PoxB and phosphotransacetylase-acetate kinase(Pta-AckA) pathways (19, 20). Elimination of the Pta-AckA(△pta△ackA) pathway is an effective method to reduce acetateformation (19, 20). However, the deletion of ackA or pta alsoimpaired growth (19, 20). ΔpoxB was an alternative way to re-duce acetate formation without affecting growth; and acetate canbe converted by acetyl-CoA synthetase (Acs) to acetyl-CoA.ΔpoxB::acs had been developed as an alternative strategy to re-duce acetate formation. Our recent research revealed that thecombination of inactivation of poxB and overexpression of ace-tyl-CoA synthetase gene (acs) in the E. coli K strain was suc-cessful in controlling acetate accumulation (14).Metabolic engineering can sometimes induce unexpected re-
actions in the host cell, such as fluctuations in protein expressionlevels. In this work, expression levels of DAOCS in different hostsstayed stable; nevertheless, the relative activities of DAOCS indifferent strains were corrected with the corresponding expressionlevels in these strains (Fig. 5D). These results clearly indicated thatthe improvement in G-7-ADCA yields is the direct effect of in-creased metabolic flux via the TCA cycle, and the catalytic effi-ciency of DAOCS was also enhanced with a continuous 2OGsupply powered by glucose.
Materials and MethodsMaterials. The bacterial strains and plasmids used in this study are listed inTable S2. E. coli DH5α was used for DNA manipulation. E. coli K12 (BW25113)and its derivative strains were used for scDAOCS-H7 expression andG-7-ADCA production. E.coli knockout strains were obtained from the Keiocollection (National BioResource Project, NIG) (21). Vector pDB1S (Table S2)was used for gene expression. Plasmid pCP20 was used to remove the FLPrecognition target (FRT) -flanked resistant marker in E. coli K12 (Table S2).
Penicillin G was purchased from North China Pharmaceutical. G-7-ADCAwas provided byGuan-Zhu Xu (Institute ofMicrobiology, Chinese Academy ofSciences, Beijing, People’s Republic of China).
Culture Conditions. E. coli strains were grown at 37 °C on a shaker at 250 rpmin Luria-Bertani (LB) medium (1% tryptone, 0.5% yeast extract and 1% NaCl,pH 7.0) with ampicillin (50 μg/mL) or kanamycin (25 μg/mL), added as re-quired. For scDAOCS-H7 expression, overnight cultures were inoculated intoan autoinduction medium, ZYM medium (22), with 1% inoculums and in-cubated with shaking at 30 °C for 16 h. ZYM contains 1% tryptone; 0.5%
Fig. 5. The cumulative effects of different combinations of△sucA, △aceA,△poxB::acs, and△ampCmutations on G-7-ADCA production. (A) G-7-ADCAyields of double-mutation strains. (B) G-7-ADCA yields of triple mutationstrains. (C) G-7-ADCA yields of selected engineered strains. (D) Correctedrelative activities of DAOCS of selected engineered strains. The recombinantstrains were cultured at 30 °C for 16 h and the cells were suspended in re-action mixture containing 100 mM penicillin G as substrate for G-7-ADCAproduction. The bioconversion reactions were carried out at 30 °C and200 rpm for 20 h.
9858 | www.pnas.org/cgi/doi/10.1073/pnas.1502866112 Lin et al.
Dow
nloa
ded
by g
uest
on
Apr
il 7,
202
0

yeast extract; 25 mM Na2HPO4; 25 mM KH2PO4; 50 mM NH4Cl; 5 mM Na2SO4;0.5% glycerol; 0.05% glucose; 0.2% L-arabinose; 2 mM MgSO4, and tracemetal mix containing 0.05 mM FeCl3, 0.02 mM CaCl2, 0.01 mM MnCl2, 0.01 mMZnSO4, and 2 mM each of CoCl2, NiCl2, Na2MoO4, Na2SeO3, and H3BO3.
Construction of Recombinant Expression Vectors. S. clavuligerus scDAOCS H7was amplified from pET30a-H7 (2) with primer pairs (P1: AACATGCCATGG-ACACGACGGTGCCCACCTTCA and P2: CCGGAATTCTTACTATGCCTTGG-ATGTGCGGCGCA; the restriction sites were underlined in primers.) and insertedinto the pDB1S at NcoI and EcoRI sites to produce pDB1S-H7. This plasmid wastransformed into E. coli strain BW25113 to obtain recombinant strain BW/H7for the expression of scDAOCS-H7. The expressed scDAOCS-H7 was analyzedby SDS/PAGE. Gel was analyzed by BrandScan 5.0. Protein concentration wasdetermined by the method of Bradford (23) using BSA as a standard.
Gene Knockout. P1 transduction was used to transfer selectable mutation fromone E. coli strain to another. P1 transduction was performed according toThomason et al. (24). The resistance genes were eliminated by using a helperplasmid pCP20 encoding the FLP recombinase. The mutant strains were ana-lyzed by PCR and further confirmed by sequencing (25).
Bioconversion Conditions. Cells cultured at 30 °C for 16 h were collected bycentrifugation at 8,000 × g for 10 min, and washed with 0.85% NaCl solutiontwice. For the biotransformation, cells were suspended in the reaction mixture(2 mL) contained 100mM penicillin G, 0.5% glucose, 3.6 mMFeSO4 and 100 mMMops buffer to form cell suspension (OD600 nm = 40). The bioconversion re-actions were performed at 30 °C and 200 rpm on a shaker. Samples werecentrifuged at 10,000 × g for 5 min, filtrated with a 0.22-μm filter membraneand the resulting supernatants were analyzed for penicillin G and G-7-ADCA byHPLC. At least three independent experiments were performed for each strain.
Decomposition Assay of Penicillin G and G-7-ADCA by Host Cell Lysis Solution.After 12 h of incubation in LB medium at 37 °C and 250 rpm, culture broths
from E. coli BW25113 and E. coli ΔampC were centrifuged at 8,000 × g for10 min at 4 °C. Cells were washed, suspended in 0.1 M Tris·HCl buffer (pH 7.5),and treated by ultrasonication on ice and then centrifuged at 12,000 × g for5 min at 4 °C. The resulting supernatant was designated as the host cell lysissolution. 10 mM penicillin G and 1 mM G-7-ADCA solution were prepared in0.1 M Tris·HCl buffer (pH 7.5). Substrate solutions (100 μL) were mixed with100 μL of host cell lysis solution (the final concentration 10 OD/mL) in anEppendorf centrifuge tube, and incubated in a 30 °C water bath for 0–5 h.A natural decomposition control of substrate was prepared by incubatingpenicillin G and G-7-ADCA with 0.1 M Tris·HCl buffer (pH 7.5) under thesame conditions.
Analytical Methods. Cell density was estimated bymeasuring the optical densityat 600 nm. The expression of recombinant enzymes was analyzed by 12% SDS/PAGE analysis.
The concentrations of penicillin G and G-7-ADCA were measured by HPLC(Agilent 1260 series, Hewlett–Packard), equipped with a ZORBAX EclipseXDB-C18 column (4.6 × 100 mm; Agilent) and a diode-array detector.Analysis was performed at 25 °C at a flow rate of 0.5 mL/min with a mobilephase of 30% (vol/vol) acetonitrile and 70% (vol/vol) water with 0.1% tri-fluoroacetic acid. Penicillin G was measured at 220 nm and G-7-ADCA wasmeasured at 260 nm. Retention times for G-7-ADCA and penicillin G were 8.6and 11.2 min, respectively.
The concentration of acetate in the bioconversion was measure by HPLC(Agilent 1260 series, Hewlett–Packard) equipped with a Bio-Rad AminexHPX-87H column (7.8 × 300 mm) and a diode-array detector. Analysis wasperformed at 65 °C with a mobile phase of 6 mM H2SO4 at a flow rate of0.55 mL/min and measurement at 210 nm.
ACKNOWLEDGMENTS. This work was supported by Ministry of Science andTechnology of China Grant 2013CB734000, Key Research Program of theChinese Academy of Sciences Grant KGZD-EW-606, and National NaturalScience Foundation of China Grant 31170038.
1. Velasco J, et al. (2000) Environmentally safe production of 7-aminodeacetoxycephalos-poranic acid (7-ADCA) using recombinant strains of Acremonium chrysogenum. NatBiotechnol 18(8):857–861.
2. Ji J, et al. (2012) Iterative combinatorial mutagenesis as an effective strategy forgeneration of deacetoxycephalosporin C synthase with improved activity towardpenicillin G. Appl Environ Microbiol 78(21):7809–7812.
3. Crawford L, et al. (1995) Production of cephalosporin intermediates by feeding adipicacid to recombinant Penicillium chrysogenum strains expressing ring expansion ac-tivity. Biotechnology (N Y) 13(1):58–62.
4. Cho H, et al. (1998) Elucidation of conditions allowing conversion of penicillin G andother penicillins to deacetoxycephalosporins by resting cells and extracts of Strepto-myces clavuligerus NP1. Proc Natl Acad Sci USA 95(20):11544–11548.
5. Cooper RD (1993) The enzymes involved in biosynthesis of penicillin and cephalo-sporin; Their structure and function. Bioorg Med Chem 1(1):1–17.
6. Kovacevic S, Weigel BJ, Tobin MB, Ingolia TD, Miller JR (1989) Cloning, characteriza-tion, and expression in Escherichia coli of the Streptomyces clavuligerus gene en-coding deacetoxycephalosporin C synthetase. J Bacteriol 171(2):754–760.
7. Valegård K, et al. (1998) Structure of a cephalosporin synthase. Nature 394(6695):805–809.
8. Valegård K, et al. (2004) The structural basis of cephalosporin formation in a mono-nuclear ferrous enzyme. Nat Struct Mol Biol 11(1):95–101.
9. Tarhonskaya H, et al. (2014) Studies on deacetoxycephalosporin C synthase support aconsensus mechanism for 2-oxoglutarate dependent oxygenases. Biochemistry 53(15):2483–2493.
10. Goo K-S, Chua C-S, Sim T-S (2009) Directed evolution and rational approaches toimproving Streptomyces clavuligerus deacetoxycephalosporin C synthase for cepha-losporin production. J Ind Microbiol Biotechnol 36(5):619–633.
11. Fernández M-J, et al. (1999) Stimulatory effect of growth in the presence of alcoholson biotransformation of penicillin G into cephalosporin-type antibiotics by restingcells of Streptomyces clavuligerus NP1. Appl Microbiol Biotechnol 52(4):484–488.
12. Gao Q, Demain AL (2002) Improvement in the resting-cell bioconversion of penicillinG to deacetoxycephalosporin G by addition of catalase. Lett Appl Microbiol 34(4):290–292.
13. Gao Q, Piret JM, Adrio JL, Demain AL (2003) Performance of a recombinant strain ofStreptomyces lividans for bioconversion of penicillin G to deacetoxycephalosporin G.J Ind Microbiol Biotechnol 30(3):190–194.
14. Tao Y, Cheng Q, Kopatsis AD (2012) Metabolic engineering for acetate control inlarge scale fermentation. Methods Mol Biol 834:283–303.
15. Waegeman H, et al. (2011) Effect of iclR and arcA knockouts on biomass formationand metabolic fluxes in Escherichia coli K12 and its implications on understanding themetabolism of Escherichia coli BL21 (DE3). BMC Microbiol 11:70.
16. Nimmo HG, et al. (1987) Regulation of the enzymes at the branchpoint between thecitric acid cycle and the glyoxylate bypass in Escherichia coli. Biochem Soc Symp 54:93–101.
17. Wang Y, Yu H, Zhang J, Luo H, Shen Z (2012) Double knockout of β-lactamase andcephalosporin acetyl esterase genes from Escherichia coli reduces cephalosporin Cdecomposition. J Biosci Bioeng 113(6):737–741.
18. Sauvage E, Kerff F, Terrak M, Ayala JA, Charlier P (2008) The penicillin-binding pro-teins: Structure and role in peptidoglycan biosynthesis. FEMS Microbiol Rev 32(2):234–258.
19. Dittrich CR, Bennett GN, San KY (2005) Characterization of the acetate-producingpathways in Escherichia coli. Biotechnol Prog 21(4):1062–1067.
20. Phue JN, Lee SJ, Kaufman JB, Negrete A, Shiloach J (2010) Acetate accumulationthrough alternative metabolic pathways in ackA− pta− poxB− triple mutant in E. coli B(BL21). Biotechnol Lett 32(12):1897–1903.
21. Baba T, et al. (2006) Construction of Escherichia coli K-12 in-frame, single-geneknockout mutants: the Keio collection. Mol Syst Biol 2(1):2006.0008.
22. Studier FW (2005) Protein production by auto-induction in high density shaking cul-tures. Protein Expr Purif 41(1):207–234.
23. Bradford MM (1976) A rapid and sensitive method for the quantitation of microgramquantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254.
24. Thomason LC, Costantino N, Court DL (2007) E. coli genome manipulation by P1transduction. Curr Protoc Mol Biol 79(1):1.17.1–1.17.8.
25. Datsenko KA, Wanner BL (2000) One-step inactivation of chromosomal genes in Es-cherichia coli K-12 using PCR products. Proc Natl Acad Sci USA 97(12):6640–6645.
Lin et al. PNAS | August 11, 2015 | vol. 112 | no. 32 | 9859
APP
LIED
BIOLO
GICAL
SCIENCE
S
Dow
nloa
ded
by g
uest
on
Apr
il 7,
202
0