Embed Size (px)
Citation preview

FEMS Microbiology Letters 65 (1989) 41-46 41 Published by Elsevier
FEMSLE 03732
Recognition of Fusobacterium nucleaturn subgroups Fn-1, Fn-2 and Fn-3 by ribosomal RNA gene restriction patterns
Paul A. Lawson 1, Saheer E. G h a r b i a 1, H a r o u n N. Shah 1 and D u n c a n R. Cla rk 2
I Department of Oral Microbiology, The London Hospital Medical College, London, and 2 GeneSys Limited, Feltham, Middlesex, U.K.
Received 20 May 1989 Revision received 16 June 1989
Accepted 19 June 1989
Key words" Fusobacterium nucleatum," F. nucleatum subgroups; RNA gene restriction patterns
1. SUMMARY 2. INTRODUCTION
DNA from representative strains of Fusobac- terium nucleatum subgroups Fn-1, Fn-2 and Fn-3 was digested with restriction enzymes E c o R I and TaqI and the electrophoretically separated frag- ments hybridized with a 32p-16S rRNA gene probe from E. coli. The rRNA gene restriction patterns from DNA digested with either enzyme allowed the clustering of strains into the three subgroups. However, TaqI digested DNA yielded a wider distribution of taxonomically useful bands (ca 0.65 + 14.3 kbp) and the pattern produced was characteristic of each subgroup. The present method is a simple and reliable means of identify- ing the three subgroups of F. nucleatum and pro- vides a useful method for further studies of the heterogeneity of F. nucleatum.
Correspondence to: P.A. Lawson, Department of Oral Microbi- ology, The London Hospital Medical College, Turner Street, White.chapel, London E1 2AD, U.K.
Fusobacterium nucleatum forms part of the nor- mal flora of human dental plaque and the mucous membranes of other sites but is increasingly recog- nised as an important component of anaerobic infections [1,2]. Because of its nonfermentative metabolism, carbohydrate fermentation reactions are of little value for its identification [2,3] and several new methods have been introduced [4-7]. In a preliminary study, we reported the value of glutamate dehydrogenase (GDH) as a diagnostic marker for the genus Fusobacterium and showed three G D H electrophoretic patterns within the three human F. nucleatum isolates studies [8]. Another enzyme involved in glutamate catabo- lism, 2-oxoglutarate reductase (OGR) was recently demonstrated in all species of the genus [9]. Fur- thermore, the application of both G D H and O G R electrophoretic analysis to more than 30 human, oral isolates of F. nucleatum showed evidence for three subgroups (designated Fn-1, Fn-2 and Fn-3) within this species [10].
0378-1097/89/$03.50 © 1989 Federation of European Microbiological Societies

42
Recently, patterns of restriction fragments which hybridized with labelled ribosomal-RNA (rRNA) from Escherichia coli have been used to fingerprint bacterial strains [11-13]. Since some rRNA sequences are highly conserved during evolution, a single probe can be used to delineate taxonomically distant bacterial. The present study was therefore undertaken to identify rRNA gene restriction patterns which will recognise the sub- groups of F. nucleatum for subsequent use in studies of human oral ecology.
3. MATERIALS AND METHODS
3.1. Cultures and cultivation Strains used in the present study were initially
characterised by biochemical tests [3] and DNA- DNA hybridization experiments [10]. They were previously assigned to F. nucleatum subgroup Fn-1 (ATCC 25586, 1319, 1336, 28, 11, M287, 33 and 78), Fn-2 (338, 1406, 1445, 1446, 339, 708, and 720) and Fn-3 (L45, F24 and 010) by electro- phoretic patterns of GDH and OGR [10]. Apart from the type strain, all strains were recent human oral isolates. Cultures were maintained by weekly subculture on 2.5% (v/v) blood agar plates at 37°C in an atmosphere of 10% (v/v) CO 2 and 20% (v/v) H 2 in N 2.
3.2. Isolation and purification of DNA Cells from two, 3-day-old plates were harvested,
washed and suspended in 2 ml TES buffer (0.05 M Tris-HCl, 0.005 M EDTA, 0.05 M NaCI, pH 8.0) and 100 #1 lysozyme (Sigma, Poole, U.K.) (10 mg/ml) added. After leaving on ice for 15 rain, 1 /~1 diethylpyrocarbonate was added, followed by Proteinase K (BDH, Essex, U.K.) and RNase (Sigma) to a final concentration of 50/~g/ml each and the mixture incubated at 65 °C for 1 h. Lysis was achieved by the addition of SDS (20% w/v) to a final concentration of 2% (v/v) and the incubation continued for a further 10 min. Pro- teins were denatured and removed by three extrac- tions times with an equal volume of phenol/ chloroform/isoamyl alcohol (25 : 24 : 1, v/v), once with chloroform/isoamyl alcohol (24:1, v /v and finally once with chloroform. DNA was precipi-
tated by the addition of ice-cold absolute ethanol, spooled out, washed with 70% (v/v) ethanol, air dried then redissolved in TE buffer (0.01 M Tris- HCI, 0.001 M EDTA, pH 8.0) and stored at - 20°C .
3. 3. Probe isolation and labelling The 1.5 kilobase pair (kbp) BclI-BstE II frag-
ment from the plasmid pKK3535 [14] which con- tained the majority of the gene coding for an E. coli 16S rRNA gene was used as DNA probe. It was separated on low melting point agarose and labelled using the oligonucleotide primer exten- sion method of Feinberg and Vogelstein [15] in- corporating 32p-dCTP. The labelled nucleic acid was separated from unincorporated low molecular weight material by spun column chromatography [161.
3.4. Restriction endonuclease digestion and agarose gel electrophoresis
Chromosomal DNA (3-4 /zg) was digested to completion with the restriction endonucleases EcoRI (New England Biolabs, Massachusetts, U.S.A.) and TaqI (Northumbria Biologicals Limi- ted, Northumberland, U.K.) according to the manufacturers instructions. The restriction frag- ments were separated by electrophoresis through 1% (w/v) agarose gels at 1 V/cm. for 18 h in Tris-borate buffer containing 0.5/~g/ml ethidium bromide [16]. Lambda DNA digested with HindIII was used as a molecular weight marker. The gel was photographed using the Polaroid MP4 system with 665 film.
3.5. Southern blotting and hybridization Gels were sequentially soaked in 1.5 M NaC1,
0.5 M NaOH for 40 min followed by 1.5 M NaCI, 0.5 M Tris-HC1, 1 mM EDTA for 40 min and blotted [17] on Hybond-N nylon membrane (Amersham, Amersham, U.K.), the blotted DNA was covalently attached by baking at 80 °C for 2 h. A prehybridization step was carried out for 1-3 at 65 ° C in a solution containing [5 x SSC(1 x SSC is 0.1 M NaCI, 0.015 M trisodium citrate), 5 x Denhardts's solution (1 x Denhardt's is 0.02% (w/v) BSA, 0.02% (w/v) Ficoll and 0.02% (w/v) polyvinyl-polyppyrollidone), 0.5% (w/v) SDS and
20 t tg/ml boiled salmon sperm DNA]. Dextran sulphate was added to 10% (w/v) and probe DNA added to 5 x 105 c.p.m./ml (denatured by boiling for 3 rain) and hybridization continued for a further 18 h at 65°C. Following incubation the membranes were washed twice at 65 ° C in 2 x SSC, 0.1% (w/v) SDS followed by two washes at 65°C in 0.2 x SSC and 0.1% (w/v) SDS, each wash being 15 min duration. Membranes were wrapped in Saran wrap and exposed to photographic film (Kodak XAR5) at - 7 0 °C for 1-3 days.
3.6. DNA fragment size estimation The molecular size (in kbp) of restriction frag-
ments was calculated using a modified program of Duggleby et al. [18] using HindIII cleaved DNA as a marker.
4. RESULTS AND DISCUSSION
The three subgroups of F. nucleatum desig- nated Fn-1, Fn-2 and Fn-3 were recently shown to contain high DNA-DNA homology (70-90%) using the type strain ATCC 25586 as the reference DNA probe under stringent conditions of hybridi- zation [10]. However, reciprocal hybridization ex- periments with reference DNA from subgroup Fn-3 showed a lower affinity with both subgroups Fn-I (69±80%) and Fn-2 (60-83%). This subgroup was the least frequently isolated from clinical sam- ples and studies are now in progress to determine its site specificity. Each subgroup was readily re- cognised by its characteristic electrophoretic pat- tern of GDH and OGR [10] but was otherwise indistinguishable by conventional bacteriological tests.
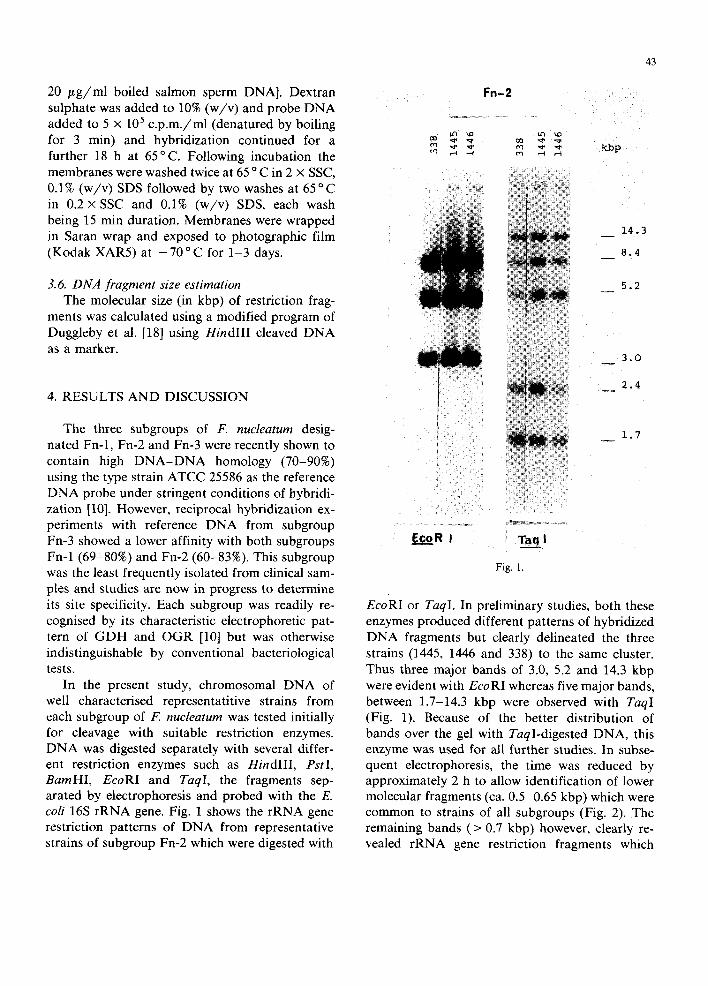
In the present study, chromosomal DNA of well characterised representatitive strains from each subgroup of F. nucleatum was tested initially for cleavage with suitable restriction enzymes. DNA was digested separately with several differ- ent restriction enzymes such as HindIII, PstI, BamHI, EcoRI and TaqI, the fragments sep- arated by electrophoresis and probed with the E. coli 16S rRNA gene. Fig. 1 shows the rRNA gene restriction patterns of DNA from representative strains of subgroup Fn-2 which were digested with
Fn-2
43
"~ ~ ~ ~ kbp
14.3
8.4
5 . 2
3 . 0
2 . 4
1 .7
F~oR I Taq l
Fig. 1.
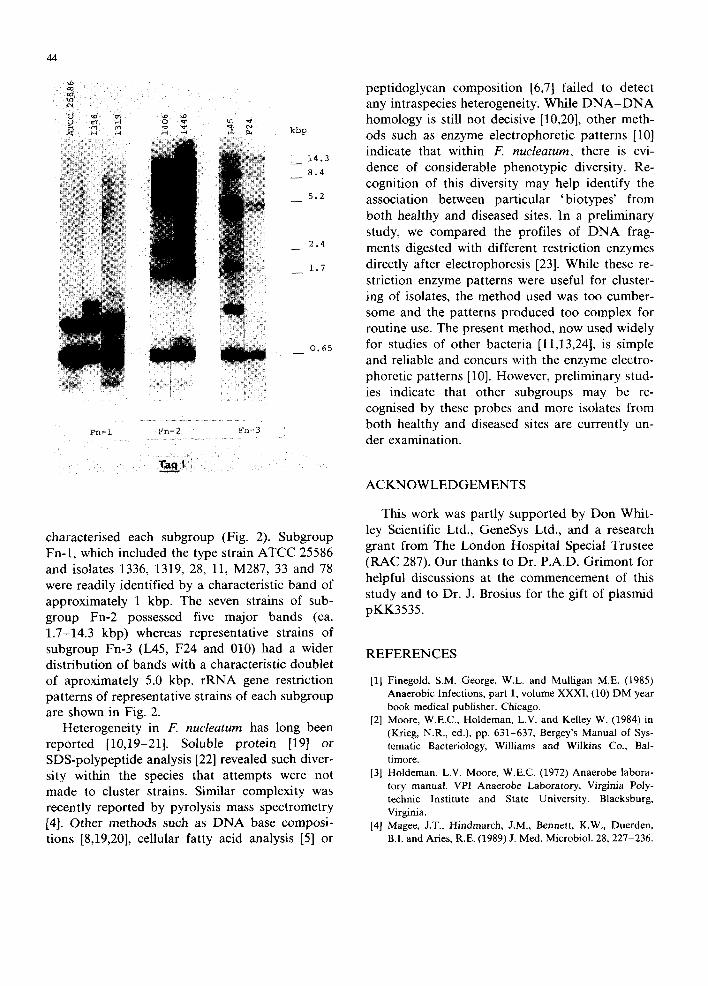
EcoRI or TaqI. In preliminary studies, both these enzymes produced different patterns of hybridized DNA fragments but clearly delineated the three strains (1445, 1446 and 338) to the same cluster. Thus three major bands of 3.0, 5.2 and 14.3 kbp were evident with EcoRI whereas five major bands~ between 1.7-14.3 kbp were observed with TaqI (Fig. 1). Because of the better distribution of bands over the gel with TaqI-digested DNA, this enzyme was used for all further studies. In subse- quent electrophoresis, the time was reduced by approximately 2 h to allow identification of lower molecular fragments (ca. 0.5-0.65 kbp) which were common to strains of all subgroups (Fig. 2). The remaining bands (> 0.7 kbp) however, clearly re- vealed rRNA gene restriction fragments which
44
~m
tt~ c~
u ~o
kbp
14.3
8.4
5.2
2.4
1.7
O.65
Fn-i Fn-2 Fn-3
Taq I
characterised each subgroup (Fig. 2). Subgroup Fn-1, which included the type strain ATCC 25586 and isolates 1336, 1319, 28, 11, M287, 33 and 78 were readily identified by a characteristic band of approximately 1 kbp. The seven strains of sub- group Fn-2 possessed five major bands (ca. 1.7-14.3 kbp) whereas representative strains of subgroup Fn-3 (L45, F24 and 010) had a wider distribution of bands with a characteristic doublet of aproximately 5.0 kbp. rRNA gene restriction patterns of representative strains of each subgroup are shown in Fig. 2.
Heterogeneity in F. nucleatum has long been reported [10,19-21]. Soluble protein [19] or SDS-polypeptide analysis [22] revealed such diver- sity within the species that attempts were not made to cluster strains. Similar complexity was recently reported by pyrolysis mass spectrometry [4]. Other methods such as DNA base composi- tions [8,19,20], cellular fatty acid analysis [5] or
peptidoglycan composition [6,7] failed to detect any intraspecies heterogeneity. While DNA-DNA homology is still not decisive [10,20], other meth- ods such as enzyme electrophoretic patterns [10] indicate that within F. nucleatum, there is evi- dence of considerable phenotypic diversity. Re- cognition of this diversity may help identify the association between particular 'biotypes' from both healthy and diseased sites. In a preliminary study, we compared the profiles of DNA frag- ments digested with different restriction enzymes directly after electrophoresis [23]. While these re- striction enzyme patterns were useful for cluster- ing of isolates, the method used was too cumber- some and the patterns produced too complex for routine use. The present method, now used widely for studies of other bacteria [11,13,24], is simple and reliable and concurs with the enzyme electro- phoretic patterns [10]. However, preliminary stud- ies indicate that other subgroups may be re- cognised by these probes and more isolates from both healthy and diseased sites are currently un- der examination.
ACKNOWLEDGEMENTS
This work was partly supported by Don Whit- ley Scientific Ltd., GeneSys Ltd., and a research grant from The London Hospital Special Trustee (RAC 287). Our thanks to Dr. P.A.D. Grimont for helpful discussions at the commencement of this study and to Dr. J. Brosius for the gift of plasmid pKK3535.
REFERENCES
[1] Finegold, S.M. George, W.L. and Mulligan M.E. (1985) Anaerobic Infections, part 1, volume XXXI, (10) DM year book medical publisher. Chicago.
[2] Moore, W.E.C., Holdeman, L.V. and Kelley W. (1984) in (Krieg, N.R., ed.), pp. 631-637, Bergey's Manual of Sys- tematic Bacteriology, Williams and Wilkins Co., Bal- timore.
[3] Holdeman, L.V. Moore, W.E.C. (1972) Anaerobe labora- tory manual. VPI Anaerobe Laboratory, Virginia Poly- technic Institute and State University. Blacksburg, Virginia.
[4] Magee, J.T., Hindmarch, J.M., Bennett, K.W., Duerden, B.I. and Aries, R.E. (1989) J. Med. Microbiol. 28, 227-236.

[5] Jantzen, E. and Hofstad, T. (1981) J. Gen. Microbiol. 123, 163-171.
[6] Miyagawa, E., Azuma, R. and Suto, T. (1981) J. Gen. Appl. Microbiol. 27, 199-208.
[7] Vasstrand, E.N., Jansen, H.B., Miron, T. and Hofstad, T. (1981) Infect. Immun. 36, 114-122.
[8] Gharbia, S.E. and Shah, H.N. (1988) J. Gen. Microbiol. 134, 327-332.
[9] Gharbia, S.E. and Shah, H.N. (1989) J. Clin. Microbiol. (in press).
[10] Gharbia, S.E. and Shah, H.N. (1989) Int. J. Syst. Bacteriol. (in press).
[11] Grimont. F., Grimont, P.A.D. (1986) Annales de l'Institut Pasteur/Microbiologie 137B, 165-175.
[12] Irino, K., Grimont, F., Casin, I., Grimont, P.A.D. and The Brazilian Purpuric Fever Study Group. (1988) J. Clin. Microbiol. 26, 1535-1538.
[13] Saunders, N.A., Harrison, T.G., Kachwalla, N. and Taylor, A.G. (1988) J. Gen. Microbiol. 134, 2363--374.
[14] Brosius, J., Ullrich, A., Raker, M.A. Gray, A., Dull, T.J., Gutrell, R.R. and Noller, H.F. (1981) Plasmid 6, 112-118.
45
[15] Vogelstein, B. and Feinberg, A.P. (1984) Anal. Biochem. 137, 266-267.
[16] Maniatis, T., Fritisch, E.F. and Sambrook, J. (1982) Molecular Cloning: a Laboratory Manual. Cold Spring Harbor laboratory, Cold Spring Harbor, New York.
[17] Southern E.M. (1975) J. Mol. Biol. 98, 503-517. [18] Duggleby, R.G., Kinns, H. and Rood, R.I. (1981) Anal.
Biochem. 110, 49-55. [19] Calhoon, D.A., Mayberry, W.R. and Slots, J. (1983) J.
Dent. Res. 62, 1181-1185. [20] Love, D.N., Cato, E.P., Johnson, J.L., Jones, R.F. and
Bailey, M. (1987) Int. J. Syst. Bacteriol. 37, 23-26. [21] Hofstad, T. (1979) Microbiol. Rev. 43, 103-115. [22] Cato, E.P., Moore, L.V. and Moore W.E. (1983) Int. J.
Syst. Bacteriol. 35, 475-477. [23] Lawson, P.A., Shah, H.N. and Clark, D.R. (1988) J. Den.
Res. 110. [24] De Buyser, M.L., Morvan, A., Grimont, F. and E1 Solh,
N. (1989) J. Gen. Microbiol. 136, 989-999.





























