Embed Size (px)
Citation preview

Vol.:(0123456789)1 3
Planta (2018) 248:519–544 https://doi.org/10.1007/s00425-018-2911-0
REVIEW
Recent advances in steroidal saponins biosynthesis and in vitro production
Swati Upadhyay1 · Gajendra Singh Jeena1 · Shikha1 · Rakesh Kumar Shukla1
Received: 13 April 2018 / Accepted: 27 April 2018 / Published online: 10 May 2018 © Springer-Verlag GmbH Germany, part of Springer Nature 2018
AbstractMain conclusion Steroidal saponins exhibited numerous pharmacological activities due to the modification of their backbone by different cytochrome P450s (P450) and UDP glycosyltransferases (UGTs). Plant-derived steroidal sapo-nins are not sufficient for utilizing them for commercial purpose so in vitro production of saponin by tissue culture, root culture, embryo culture, etc, is necessary for its large-scale production.
Saponin glycosides are the important class of plant secondary metabolites, which consists of either steroidal or terpenoidal backbone. Due to the existence of a wide range of medicinal properties, saponin glycosides are pharmacologically very important. This review is focused on important medicinal properties of steroidal saponin, its occurrence, and biosynthesis. In addition to this, some recently identified plants containing steroidal saponins in different parts were summarized. The high throughput transcriptome sequencing approach elaborates our understanding related to the secondary metabolic path-way and its regulation even in the absence of adequate genomic information of non-model plants. The aim of this review is to encapsulate the information related to applications of steroidal saponin and its biosynthetic enzymes specially P450s and UGTs that are involved at later stage modifications of saponin backbone. Lastly, we discussed the in vitro production of steroidal saponin as the plant-based production of saponin is time-consuming and yield a limited amount of saponins. A large amount of plant material has been used to increase the production of steroidal saponin by employing in vitro culture technique, which has received a lot of attention in past two decades and provides a way to conserve medicinal plants as well as to escape them for being endangered.
Keywords Metabolites · P450 · UGTs · Transcriptome · Pathway
Introduction
Majority of higher plants and microbes synthesize useful secondary metabolites, which are categorized into alkaloids, terpenoids, polyphenols, polyketides, etc., according to their chemical structures and biosynthetic routes. These com-pounds protect plants from pathogens, competitors and UV light. They also assist in plant adaptation and most of them have valuable medicinal properties that make them useful in
pharmaceutical, agrochemical, flavor and aroma industries. Saponins are important plant secondary metabolites gen-erally known for their multiple pharmacological properties (Williams and Gong 2007).
The word saponin is derived from sapo, i.e., soap, which describes the soapy appearance of saponin when combined with water. Due to their amphiphilic properties and tendency to form the foam, they can be used as surfactant or emulsi-fying agents (Kime et al. 2015). Structurally, saponins are amphiphilic molecules that are composed of one or more hydrophilic sugar residues and hydrophobic steroidal or triterpenoidal part on the basis of which they are called as steroidal saponin or triterpenoidal saponins. The non-sugar water-insoluble part is called as sapogenin, and on the basis of sugar present in saponin molecules they can be either monodesmosidic (contains only one sugar residue), bides-mosidic (contains two sugar residues) or polydesmosidic
* Rakesh Kumar Shukla [email protected]
1 Biotechnology Division (CSIR-CIMAP), Central Institute of Medicinal and Aromatic Plants, (CSIR-CIMAP) P.O. CIMAP (a laboratory under Council of Scientific and Industrial Research, India), Near Kukrail Picnic Spot, Lucknow 226015, India
520 Planta (2018) 248:519–544
1 3
(more than two sugar residues) saponins (Lorent et al. 2014). Structural variants of saponin are found in plants due to the presence of different sugar at different position and orientation. The most common sugars residues that are found in saponins are d-glucose, d-galactose, l-rhamnose, l-arabinose, d-glucuronic acid, d-fucose and d-xylose (Lor-ent et al. 2014).
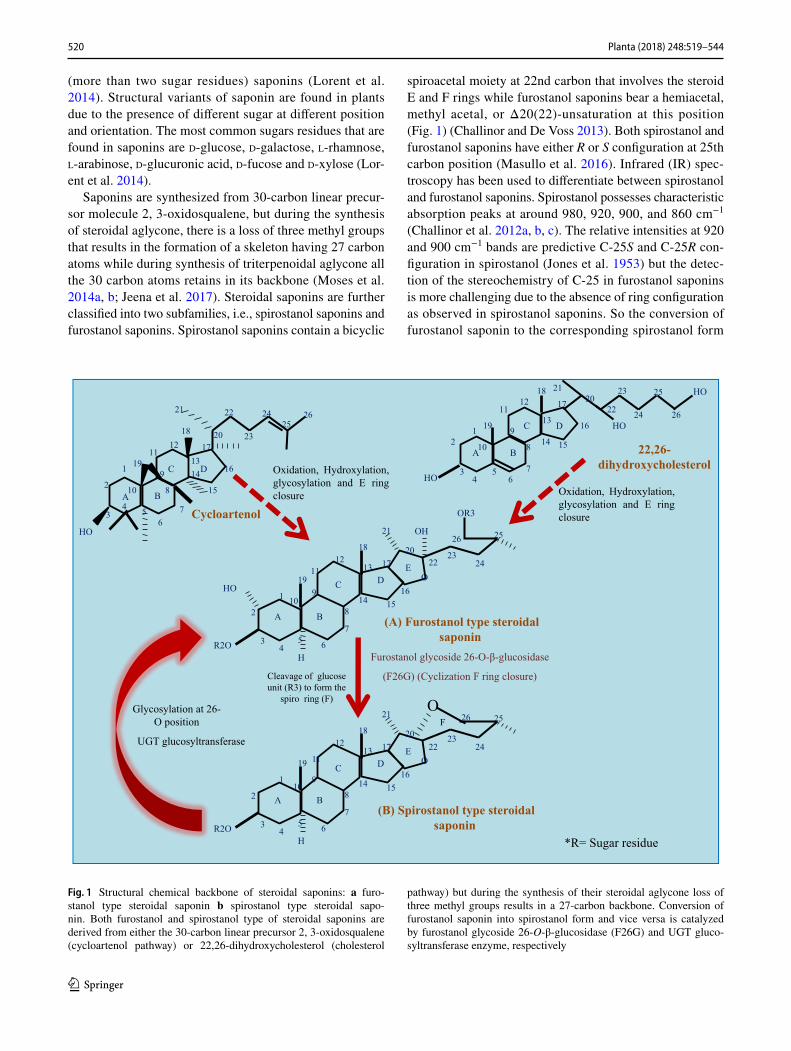
Saponins are synthesized from 30-carbon linear precur-sor molecule 2, 3-oxidosqualene, but during the synthesis of steroidal aglycone, there is a loss of three methyl groups that results in the formation of a skeleton having 27 carbon atoms while during synthesis of triterpenoidal aglycone all the 30 carbon atoms retains in its backbone (Moses et al. 2014a, b; Jeena et al. 2017). Steroidal saponins are further classified into two subfamilies, i.e., spirostanol saponins and furostanol saponins. Spirostanol saponins contain a bicyclic
spiroacetal moiety at 22nd carbon that involves the steroid E and F rings while furostanol saponins bear a hemiacetal, methyl acetal, or Δ20(22)-unsaturation at this position (Fig. 1) (Challinor and De Voss 2013). Both spirostanol and furostanol saponins have either R or S configuration at 25th carbon position (Masullo et al. 2016). Infrared (IR) spec-troscopy has been used to differentiate between spirostanol and furostanol saponins. Spirostanol possesses characteristic absorption peaks at around 980, 920, 900, and 860 cm−1 (Challinor et al. 2012a, b, c). The relative intensities at 920 and 900 cm−1 bands are predictive C-25S and C-25R con-figuration in spirostanol (Jones et al. 1953) but the detec-tion of the stereochemistry of C-25 in furostanol saponins is more challenging due to the absence of ring configuration as observed in spirostanol saponins. So the conversion of furostanol saponin to the corresponding spirostanol form
O
O
R2O
HO
H
OH
OR3
A B
C D E 22
2324
2526
1
2
3 4
5 6
7
8
9 10
1112
13
14 1516
17
18
19
20
21
(A) Furostanol type steroidal saponin
O
R2O H
A B
C D E 22
2324
2526
1
2
3 4
5 6
7
8
9 10
11
1213
14 1516
17
18
19
20
21
(B) Spirostanol type steroidal saponin
Furostanol glycoside 26-O-β-glucosidase
(F26G) (Cyclization F ring closure)Cleavage of glucose unit (R3) to form the
spiro ring (F)
F
*R= Sugar residue
Glycosylation at 26-O position
UGT glucosyltransferase
HO
A B
C D
Cycloartenol
HO
HO
22,26-dihydroxycholesterol
HO
A B
C D
1
1
2
2
3
3
4
4
5
5
6
6
7
7
8
8
9
9
10
10
19
1911
11
12
12
13
1314
14 15
15
16
16
17
17
18
1820
Oxidation, Hydroxylation, glycosylation and E ring closure
Oxidation, Hydroxylation, glycosylation and E ring closure
20
22
23
24
25
26
21
21 22
23
2425
26
Fig. 1 Structural chemical backbone of steroidal saponins: a furo-stanol type steroidal saponin b spirostanol type steroidal sapo-nin. Both furostanol and spirostanol type of steroidal saponins are derived from either the 30-carbon linear precursor 2, 3-oxidosqualene (cycloartenol pathway) or 22,26-dihydroxycholesterol (cholesterol
pathway) but during the synthesis of their steroidal aglycone loss of three methyl groups results in a 27-carbon backbone. Conversion of furostanol saponin into spirostanol form and vice versa is catalyzed by furostanol glycoside 26-O-β-glucosidase (F26G) and UGT gluco-syltransferase enzyme, respectively

521Planta (2018) 248:519–544
1 3
by either hydrolysis or by enzymatic cleavage of glucose moiety at 26th carbon position for ring closure of its side chain is the most authentic method for prediction of C-25 configuration in furostanol saponin (Inoue et al. 1996). In this review, we will focus on the steroidal saponin-producing plants whose transcriptome sequencing has been done along with we encapsulate the information related to diverse appli-cations of steroidal saponin and its biosynthetic enzymes specifically P450s and UGTs that are involved at later step modification of saponin backbone.
The value of steroidal saponins
Steroidal saponins obtained from Dioscorea zingiberensis are widely used for preventing cardiovascular diseases (Qin et al. 2009). In addition to providing cardiovascular protec-tion, steroidal saponin isolated from Ophiopogon japonicus plant also exhibited various other pharmacological activities, such as anticancer, immunomodulation, anti-oxidation, anti-inflammation, cough relief, antimicrobial, and anti-diabetes (Chen et al. 2016). Steroidal saponin constituents from Paris species are used to treat cancer, malignant boil, bleeding and gastritis (Wang et al. 2015a, b, c). They also exhibited anti-fungal activity (Morrissey and Osbourn 1999) and used as an efficient natural sweetener like glycyrrhizin from licorice roots (Kitagawa 2002). In pharmaceutical industry, they are widely used as raw materials for the production of steroid hormones (Guclu Ustundag and Mazza 2007). Steroidal saponin from Anemarrhena asphodeloides, i.e., timosaponin AIII was found to inhibit the growth of tumor cells and can be considered as a significant compound for the develop-ment of the novel anticancerous drug (Wang et al. 2016a, b). Steroidal saponin diosgenin increases bone formation (Fol-warczna et al. 2016), executes anti-thrombotic activity by inhibiting the activity of factor VIII and platelet aggregation (Zhang et al. 2013a, b, c), cardioprotective (Jayachandran et al. 2016), suppresses skin inflammation (Kim et al. 2016), have potential to treat liver fibrosis (Xie et al. 2015), use-ful in the treatment of metabolic disease by regulating cho-lesterol homeostasis, effective in reversing hyperlipidemia (Fuller and Stephens 2015), anti-inflammatory, improves antioxidant status, inhibits lipid peroxidation (Chen et al. 2015) etc. So this compound can be further used for phar-macotherapy of various diseases.
Spicatoside A from Liriope platyphylla possesses anti-inflammatory activities, anti-asthma activities, anti-oste-oclastogenesis activities, memory consolidation activi-ties, neurite outgrowth activities, anti-cancer activities (Ramalingam and Kim 2016) while steroidal saponin from allium species possesses antispasmodic effect, cardiopro-tective activity, antifungal activity and cytotoxic activity (Sobolewska et al. 2016).
The antiproliferative and cytotoxic activity of steroi-dal saponins were mainly explored by researchers. It was observed that Paris polyphylla steroidal saponins (PPSS) induce apoptosis and autophagy in human lung cancer cell lines that indicate the anti-cancerous properties of steroi-dal saponins (He et al. 2015). Additionally, spirostane-type steroidal glycosides from Allium flavum also showed cyto-toxicity against a human cancerous cell line (Rezgui et al. 2014). Trillium tschonoskii steroidal saponins have a poten-tial of prevention and treatment of colorectal cancer (Li et al. 2015a, b, c) and they also have the capability to reverse the multidrug resistance and enhance the drug sensitivity in hepatocellular carcinoma (Wang et al. 2013). Steroidal sapo-nin TTB2 from Trillium tschonoskii exerts anticancer effects against Ewing sarcoma cell line through cell cycle arrest at G2/M and S phase (Huang and Zou 2015). Terrestrosin D, PSVII and diosgenin induce apoptosis in prostate, cervi-cal and breast cancer stem cells, respectively (Zhang et al. 2014a, b; Bhuvanalakshmi et al. 2017). Polyphyllin Ι from Paris polyphylla induces cell cycle arrest and ROS depend-ent autophagy in colorectal cancerous cells.
Anticarcinogenic activity was shown by a variety of ste-roidal saponins from different plants including Digitalis trojana (Kirmizibekmez et al. 2014), Allium schoenopra-sum (Timité et al. 2013), Dioscorea zingiberensis (Tong et al. 2012), Fagonia indica (Waheed et al. 2012), Rhizoma Paridis (Xiao et al. 2012), Solanum violaceum (Yen et al. 2012), Agave sisalana (Chen et al. 2011a, b), Anemarrhena asphodeloides (Kang et al. 2011), Dioscorea bulbifera (Liu et al. 2011), Trigonella foenum-graecum (Kawabata et al. 2011), Paris polyphylla (Zhu et al. 2011), Raphia farinifera (Tapondjou et al. 2015) etc.
Characterized enzymes involved in the biosynthesis of steroidal saponins
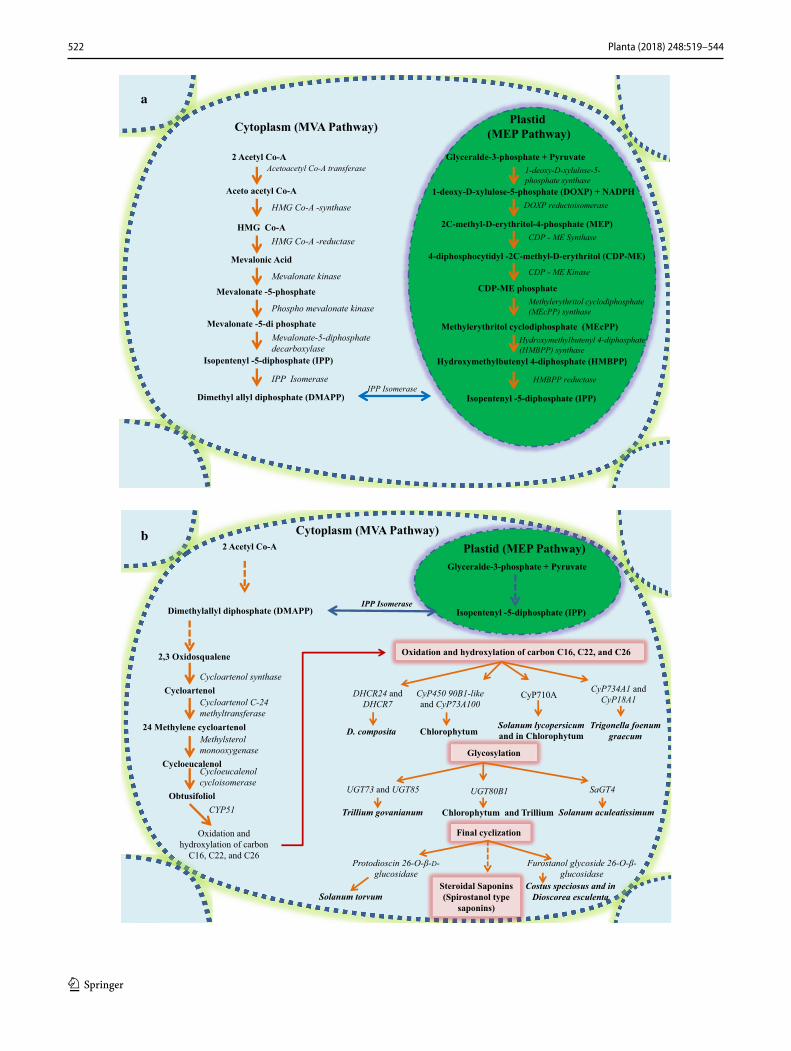
Both the cytosolic mevalonate (MVA) pathway and plas-tidial methylerythritol 4-phosphate (MEP) pathway are involved in the biosynthesis of steroidal saponins. Acetyl Co-A is the precursor of MVA pathway in the cytosol while in the chloroplast, glyceraldehyde-3-phosphate and pyruvate serve as precursor molecules to initiate MEP pathway. Both pathways lead to the formation of dimethylallyl diphos-phate (DMAPP) that is used to synthesize squalene via the sequential action of three enzymes, i.e., geranyl diphosphate synthase (GPS), farnesyl diphosphate synthase (FPS) and squalene synthase (SQS). Squalene is then converted into 2, 3-oxidosqualene under the catalysis of squalene epoxi-dase (SE). The cyclization of 2, 3-oxidosqualene is then carried out by different oxidosqualene cyclases leading to the products that undergo cyclization, hydroxylation and
522 Planta (2018) 248:519–544
1 3
Plastid (MEP Pathway) Cytoplasm (MVA Pathway)
2 Acetyl Co-A
Aceto acetyl Co-A
HMG Co-A
Mevalonic Acid
Mevalonate -5-phosphate
Mevalonate -5-di phosphate
Isopentenyl -5-diphosphate (IPP)
Dimethyl allyl diphosphate (DMAPP)
Acetoacetyl Co-A transferase
HMG Co-A -synthase
HMG Co-A -reductase
Mevalonate kinase
Phospho mevalonate kinase
Mevalonate-5-diphosphate decarboxylase
IPP Isomerase
Glyceralde-3-phosphate + Pyruvate 1-deoxy-D-xylulose-5-phosphate synthase
1-deoxy-D-xylulose-5-phosphate (DOXP) + NADPHDOXP reductoisomerase
2C-methyl-D-erythritol-4-phosphate (MEP) CDP - ME Synthase
4-diphosphocytidyl -2C-methyl-D-erythritol (CDP-ME)
CDP - ME Kinase
CDP-ME phosphate Methylerythritol cyclodiphosphate (MEcPP) synthase
Methylerythritol cyclodiphosphate (MEcPP) Hydroxymethylbutenyl 4-diphosphate (HMBPP) synthase
Hydroxymethylbutenyl 4-diphosphate (HMBPP)
Isopentenyl -5-diphosphate (IPP)
HMBPP reductase IPP Isomerase
Plastid (MEP Pathway) Glyceralde-3-phosphate + Pyruvate
Isopentenyl -5-diphosphate (IPP)
Cytoplasm (MVA Pathway) 2 Acetyl Co-A
Dimethylallyl diphosphate (DMAPP) IPP Isomerase
2,3 Oxidosqualene
Cycloartenol synthase Cycloartenol
Cycloartenol C-24 methyltransferase
24 Methylene cycloartenol
Cycloeucalenol
Obtusifoliol
Cycloeucalenol cycloisomerase
Methylsterol monooxygenase
CYP51
Oxidation and hydroxylation of carbon C16, C22, and C26
Glycosylation
Steroidal Saponins (Spirostanol type
saponins)
Final cyclization
DHCR24 andDHCR7
CyP450 90B1-like and CyP73A100
CyP710A CyP734A1 and
CyP18A1
D. composita Chlorophytum Solanum lycopersicum and in Chlorophytum
Trigonella foenum graecum
UGT73 and UGT85 UGT80B1 SaGT4
Trillium govanianum Chlorophytum and Trillium Solanum aculeatissimum
Protodioscin 26-O-β-D-glucosidase
Furostanol glycoside 26-O-β-glucosidase
Solanum torvum Costus speciosus and in
Dioscorea esculenta
Oxidation and hydroxylation of carbon
C16, C22, and C26
a
b

523Planta (2018) 248:519–544
1 3
glycosylation reactions ultimately yielding different types of steroidal saponin glycosides.
Some of the enzymes related to saponin biosynthesis have been characterized and most are yet to be characterized. The exact biosynthetic pathway of saponin biosynthesis is not yet established and now it has become an interesting area of research to find out each enzyme of this biosynthetic path-way and signaling molecules involved in the regulation of saponin biosynthesis. Some characterized enzyme of this pathway in different plant species is reviewed and summa-rized below.
Enzymes of MVA pathway
Enzymes of MVA pathway was characterized in differ-ent plants as 3-hydroxy-3-methylglutaryl CoA reductase (HMGR) is the crucial regulatory and rate-limiting enzyme of mevalonate pathway of saponin biosynthesis that cata-lyzes the irreversible conversion of HMGR to mevalonate. In addition to steroidal saponin, HMGR is also involved in the biosynthesis of phytosterols and sitosterols. By increas-ing HMGR gene expression, diosgenin content significantly increased in in vitro cultures of Dioscorea zingiberensis (Diarra et al. 2013). Similarly in Medicago truncatula, silencing of Mkb1 (E3 ubiquitin ligase), that control the activity of HMGR results in accumulation of monoglyco-sylated saponins (Moses et al. 2014a, b). This enzyme was characterized in different plants including Hevea brasilien-sis (Chye et al. 1991), potato (Bhattacharyya et al. 1995), wheat (Aoyagi et al. 1993), Camptotheca acuminate (Mal-donado-Mendoza et al. 1997), mulberry (Jain et al. 2000), rice (Ha et al. 2001), Capsicum annuum (Ha et al. 2003), Ginkgo biloba (Shen et al. 2006), Corylus avellana (Wang et al. 2007), Euphorbia pekinensi (Cao et al. 2010), Salvia miltiorrhiza (Dai et al. 2011), coffee (Tiski et al. 2011), Gly-cyrrhiza uralensis (Rong et al. 2011), Withania somnifera (Akhtar et al. 2013), Dendrobium officinale (Zhang et al. 2014a, b), Paris fargesii (Liang et al. 2014), Centella asi-atica (Kalita et al. 2015), Arabidopsis (Suzuki et al. 2004), Cyanotis arachnoidea (Wang et al. 2014a, b, c) and Panax ginseng (Kim et al. 2014).
Mevalonate kinase (MK) catalyzes a step in the isopre-noid biosynthetic pathway, which ultimately leads to huge
number of compounds that play important roles in plant growth and development (Tang and Newton 2006). This enzyme was characterized in Catharanthus roseus (Schulte et al. 2000) and Bacopa monniera (Kumari et al. 2015). Mevalonate diphosphate decarboxylase enzyme requires ATP and Mg+2 to convert mevalonate 5-diphosphate (MVAPP) into isopentenyl diphosphate (IPP), has been characterized in Bacopa monniera (Abbassi et al. 2015). Isopentenyl diphosphate isomerase (IPP isomerase) is an important enzyme of isoprenoid biosynthesis that converts IPP into DMAPP has been characterized in Tripterygium wilfordii (Tong et al. 2015), Taxus media (Shen et al. 2015) and Camptotheca acuminate (Pan et al. 2008).
Another important enzyme of isoprenoid biosynthesis is FPS that forms farnesyl diphosphate by a head to tail condensation of two molecules of IPP and one molecule of DMAPP (Fig. 2a). FPS can increase terpenoid accumulation in plants as suggested by gene expression analysis (Zhao et al. 2015). Gene expression studies have demonstrated that FPS expression is positively related to the isoprene content in plants. This gene has been characterized in many plants including Bacopa monniera (Vishwakarma et al. 2012), Tripterygium wilfordii (Zhao et al. 2015) and Arabidopsis thaliana (Keim et al. 2012).
Enzymes of methylerythritol 4‑phosphate (MEP) pathway
Enzymes of MEP pathway was also characterized in dif-ferent plants like 1-deoxy-d-xylulose 5-phosphate synthase (DXS) enzyme that was the first enzyme of MEP pathway in plastid that catalyzes the formation of 1-deoxy-d-xy-lulose 5-phosphate (DXP) by condensation of C1 alde-hyde group of d-glyceraldehyde 3-phosphate (GAP) and pyruvate. Intramolecular rearrangement and reduction of DXP were catalyzed by DXP reductoisomerase (DXR) that results in the formation of 2-C-methyl-d-erythritol 4-phos-phate (MEP). MEP is further converted into 2C-methyl-d-erythritol 2, 4-cyclodiphosphate (MEC) by the activ-ity of MEC synthase enzyme. DXS was characterized in Aquilaria sinensis (Xu et al. 2014), Maize (Cordoba et al. 2011), DXR enzyme was characterized in Ginkgo biloba (Gong et al. 2005), 2C-methyl-d-erythritol 2,4-cyclodi-phosphate (MEC) synthase enzyme of MEP pathway was characterized in Taxus media (Jin et al. 2006) and Ginkgo biloba (Kim et al. 2006). In the last step of MEP pathway both IPP and DMAPP is formed and they also intercon-vertible in another form or isomerized by the activity of IPP isomerase (Fig. 2a). IPP isomerase was characterized in Taxus media (Shen et al. 2015), Tripterygium wilfordii (Tong et al. 2016), Gossypium barbadense (Wang et al.
Fig. 2 Steroidal saponin biosynthetic pathway: a overview of steroi-dal saponin biosynthesis in plants that involve both cytosolic meva-lonate (MVA) and plastidial methylerythritol 4-phosphate (MEP) pathways. b This figure mainly represents known P450 s, UGTs and glucosidases involved in hydroxylation, oxidation, glycosylation and final cyclization of saponin backbone. Enzyme and species names are italicized. Dashed arrows imply multiple steps in the pathway. DHCR24 delta 24-sterol reductase, DHCR7 7-dehydrocholesterol reductase, CyP51 sterol 14α-demethylase
◂

524 Planta (2018) 248:519–544
1 3
2009a, b), Arabidopsis thaliana (Phillips et al. 2008), Camptotheca acuminate (Pan et al. 2008), Cinchona robusta (Ramos-Valdivia et al. 1997) etc.
Both phytosterol and saponins are synthesized from the same precursors that are squalene and 2, 3-oxidosqualene (Haralampidis et al. 2002). SQS is a membrane-bound enzyme that catalyzes the initial enzymatic reaction in the biosynthesis of triterpenoids like plant sterols from MVA isoprenoid pathway (Abe et al. 1993). It catalyzes the con-densation of two FPP molecules into squalene. SQS was characterized in a wide variety of plant species including Panax ginseng (Kim et al. 2011), Chlorophytum borivil-ianum (Kalra et al. 2013), Ornithogalum caudatum (Liu et al. 2017), Panax notoginseng (Jiang et al. 2017), Siraitia grosvenorii (Zhao et al. 2017), Salvia miltiorrhiza (Rong et al. 2016) and Magnolia officinalis (Zha et al. 2016).
SE is another rate-limiting enzyme that catalyzes the epoxidation of the double bond of squalene to produce 2,3 oxidosqualene in saponin biosynthesis. This is the first oxidation step in phytosterol and saponin biosynthesis in plants. It was shown that the expression of SE in Eleuthe-rococcus senticosus was affected by endophytic fungi (Xing et al. 2012). SE is one of the important enzymes for biosynthesis of saponins in vitro using synthetic biol-ogy approach. Major work related to this enzyme was per-formed in Panax species that is supposed to be an impor-tant source of triterpenoidal saponin. Molecular cloning, expression profiling and characterization of SE were done in Panax notoginseng and it was observed that it is highly expressed in root tissues. Methyl jasmonate (MeJA) treat-ment increases the expression level of SE in Panax gin-seng (Choi et al. 2005). Chitosan induces saponin bio-synthesis by increasing the expression level of squalene epoxidase, but it was found that the activity of chitosan and the expression level of SE were inhibited by chitosan inhibitor PD98059.
Two isoforms of SE were identified in Panax ginseng, one is involved in ginsenoside biosynthesis and another one is involved in phytosterol synthesis (Han et al. 2010). Besides Panax species and yeast, SE was also characterized in Euphorbia tirucalli (Uchida et al. 2007), Withania som-nifera (Razdan et al. 2013), Arabidopsis (Laranjeira et al. 2015), Gynostemma pentaphyllum (Guo et al. 2016) etc.
Cycloartenol synthase (CAS) is another membrane-bound enzyme as SQS and SE that is involved in phytosterol bio-synthesis in a plant system. In arabidopsis, CAS1 is reported to be involved in the biogenesis of plastid (Babiychuk et al. 2008). CAS was also characterized in Withania somnifera (Dhar et al. 2014), Nicotiana (Gas-Pascual et al. 2014), Sir-aitia grosvenorii (Zhao et al. 2017) etc. In addition to the above-mentioned enzymes, many P450 and glucosyltrans-ferases that were involved in the biosynthesis of steroidal saponin is discussed below.
Later step modification enzymes (P450 and UGT)
Intermediates of the phytosterol pathway are involved in saponin biosynthesis and primarily enzymes that belong to the multigene families of oxidosqualene cyclases (OSCs), P450 s and family 1 of UGTs are supposed to be involved in saponin biosynthesis (Augustin et al. 2011). Both P450s and UGTs are the member of plant supergene families. In plants, during the biosynthesis of diverse plant secondary metabo-lites like, terpenoids, fatty acids, lignins, hormones, sterols, pigments and defense-related phytoalexins, P450s catalyzes the oxidative reactions (Schuler 1996). P450s and UGTs are considered as a key enzyme, which leads to diversification of natural products in plants. The diverse characteristics of these enzymes make their identification in saponin pathway difficult. Arabidopsis thaliana contains around 246 P450 genes, and it is really difficult to analyze the function of each P450 gene using reverse genetics approach (Nelson 2006). Since these modifications are necessary for making the saponins soluble and active. Researchers are making large-scale experiments and attempts to isolate P450 s and UGTs involved in saponin biosynthetic pathway in plants.
Plant P450 can be divided into two groups that are mainly A-type and non A-type P450. A-type P450 are mainly involved in secondary metabolites biosynthesis while non A-type P450 is a highly diverse group that includes P450 related to lipid or hormone metabolism (Paquette et al. 2000). Animal, microbial and plant P450 differ at their protein level and show sequence homology of 30% or less than 30%. Beside this CyP51 family mem-bers are the most conserved P450 among phyla and share sequence homology more than 30–40% (Bak et al. 2011). In the eukaryotic system, members of CyP51 family cata-lyze 14α-demethylation of sterols.
As reported earlier that cycloartenol is an important intermediate between 2,3 oxidosqualene and obtusifoliol in photosynthetic eukaryotic organisms while in non-photo-synthetic eukaryotic organism cycloartenol is not present and lanosterol is derived directly from oxidosqualene (Fab-ris et al. 2014). Obtusifoliol 14α demethylase gene was characterized from Sorghum bicolor (L.) and showed strict substrate specificity towards obtusifoliol, which belongs to P450 gene family CyP51 (Bak et al. 1997). Wheat sterol 14α demethylase gene (CyP51) was found to complement ERG11 (lanosterol 14-alpha-demethylase) disrupted engi-neered yeast strain (Cabello-Hurtado et al. 1999). Steroidal saponins are mainly synthesized from cholesterol by oxy-genations at C-16, C-22 and C-26 positions (Ohnishi et al. 2009). P450s that were identified in a transcriptome analysis of medicinal plants and supposed to be involved in steroidal saponin biosynthesis is summarized in Fig. 2b.

525Planta (2018) 248:519–544
1 3
Glycosylation of saponins is supposed to be the last step in the biosynthesis of saponins. Glycosylation also regu-lates the biological activities and solubility of these naturally derived saponins (Sawai and Saito 2011). The identification and characterization of these glycosyltransferase enzymes that catalyze the transfer of sugar moiety to these steroidal and terpenoidal backbones will certainly help us to under-stand the mechanism of the diversity of these saponins and also control their biological activities in plants. Despite of their importance only few glucosyltransferases are identified and characterized. We have compiled some of the recently identified and characterized UGTs along with their function (Fig. 2b).
Presence and absence of sugar residue at specific position allow saponin glycoside to achieve their different biological activity. For example presence of C-26 sugar in avenacosides A and B from oat plant inhibited their antimicrobial activity but the removal of sugar residue from same carbon position required to gain antimicrobial activity in this plant (Osbourn 1996). Similarly, 26 desgluco derivatives of tobacco furo-stanol saponin B is required to maintain its hemolytic and antifungal activity while its structural homolog had no such kind of activity (Gruenweller et al. 1990). Number and position of sugar moiety attached with saponin molecule also influence its biological activity as incorporation of α-l-rhamnosyl at C-2 of the glucosyl unit of diosgenin leads to a disaccharide saponin with potent cytotoxicity but when l-rhamnose is attached to C-3 or C-4 of d-glucose, the result-ing diosgenyl saponins do not exhibit significant cytotoxic activity against HL-60 cells (Perez-Labrada et al. 2012).
Biosynthesis of steroidal saponin can be best studied in solanum species. Wide ranges of steroidal saponins are synthesized in this genus. It was reported that SaGT4A glu-cosylate steroidal saponins as well as steroidal alkaloids in Solanum aculeatissimum (Kohara et al. 2005) while StSGT a glycosyltransferase from Solanum tuberosum share 75% sequence homology with SaGT4A only glycosylate steroidal alkaloids (Moehs et al. 1997). It was reported that fructose was added to furostan saponins at the position of C6-OH of 26-O-β-d-glucopyranosyl by β-fructosidase enzyme of a microbial strain of Arthrobacter nitroguajacolicus. This kind of fructosylation is possible only with furostan saponins but not with spirostan saponins due to the stereo-hindrance effect (Liu et al. 2015a, b).
Generally, spirostanol saponins are water-insoluble but the attachment of α-l-rhamnopyranosyl group with the glu-copyranosyl moiety at the C2-OH of the C3-O-glycoside; increase the solubility of saponin as well as its absorption by the human body. Toruzyme 3.0 L, a kind of cyclodextrin glucanotransferase was used to synthesize steroidal sapo-nins with novel sugar chains to increase their solubility as well as their activity. Nine steroidal saponins with different aglycones and sugar chains were used as substrates to be
glucosylated by this cyclodextrin glucanotransferase (Wang et al. 2010). Cantalasaponin I, a spirostanoside from Agave sisalanta was used as substrates to be glucosylated by toru-zyme 3.0 L (CGT), and five new glucosylated products were isolated and identified (Kang et al. 2012a, b, c). Beside gly-cosylation, deglycosylation of steroidal saponin to sapogenin was reported to be carried out by rumen bacteria, especially the cell-associated enzymes fraction of ruminal content (Wang and McAllister 2010). UGTs, as well as glucosidases that were identified in a transcriptome analysis of steroidal saponin-producing plants, were summarized in Fig. 2b.
Newly identified steroidal saponins in last decade
Generally, steroidal saponins were reported in monocot fam-ilies like Dioscoreaceae, Amaryllidaceae, Smilacaceae, Lili-aceae, Agavaceae, Alliaceae, Asparagaceae, Bromeliaceae, Palmae and Scrophulariaceae, etc, and they are accumulated abundantly in crop plants such as yam, alliums, asparagus, fenugreek, yucca and ginseng (Hostettmann and Marston 2005). The presence of steroidal saponin was also reported in certain dicot families like Apocynaceae, Leguminosae, Solanaceae etc. (Moses et al. 2014a, b). Steroidal saponins from different angiosperms were extracted and their struc-ture was elucidated by spectral analysis. Modern spectro-metric methods used for the structural analysis of saponins are mass spectrometry (MS), nuclear magnetic resonancy (NMR) and IR. In different organs of the plant there is a considerable variation in the quantity of saponin. Saponin content depends on age, cultivar and geographical distribu-tion of plant (Vincken et al. 2007). Most commonly steroidal saponins are isolated from underground parts of the plant especially roots and rhizomes, and less often from above ground tissues such as leaves, stems, seeds, and fruits (Chal-linor and De Voss 2013). Cereals and grasses are generally deficient in saponins, with some notable exceptions such as avena species (oats), which accumulate both triterpenoid and steroidal saponins (Osbourn 2003). It is known that over 90 plant families contain steroidal saponins and many new occurrences are still being reported (Hostettmann and Marston 1995). Some recently identified steroidal saponin with their plant part is summarized in Table 1.
Transcriptome analysis of medicinal plants that produce steroidal saponins
The advent of high throughput next-generation sequencing (NGS) has made the transcriptomic analysis of plant possi-ble with increasing speed and affordability. Transcriptome analysis of medicinal plants whose genomic information is

526 Planta (2018) 248:519–544
1 3
Table 1 List of some recently identified, steroidal saponin-producing plants
S. No. Steroidal saponin Plant Plant part References
1. Sarsasapogenin M, Sarsasapogenin N
Asparagus officinalis Roots Huang and Kong (2006)
2. Ornithosaponins A–D Ornithogalum thyrsoides Bulbs Kuroda et al. (2006)3. Three spirostanol and one furo-
stanol saponinCalamus insignis Stem Ohtsuki et al. (2006)
4. Chekiangensosides A and B Cynanchum chekiangense Roots Li et al. (2006)5. Neosibiricosides A–D Polygonatum sibiricum Rhizomes Ahn et al. (2006)6. Two furostane type, two choles-
tane and two spirostane typeTribulus alatus Del Aerial parts Temraz et al. (2006)
7. Spirostane type steroidal saponin Agave shrevei leaves Pereira et al. (2004)8. Two steroidal saponins Allium ursinum L Underground parts Sobolewska et al. (2006)9. Two steroidal saponins Capsicum frutescens Whole Plant De Lucca et al. (2006)10. Timosaponins AI, AII, AIII, and
AIV, and timosaponins BI, and BII
Anemarrhena asphodeloides Rhizome Nian et al. (2006)
11. Three steroidal glycosides Furcraea selloa Leaves El-Sayed et al. (2006)12. Diosgenin Solanum lyratum Whole plant Sun et al. (2006)13. Two new spirostanol saponins Smilax medica Roots Sautour et al. (2006)14. Racemosides A, B and C Asparagus racemosus Fruits Mandal et al. (2006)15. Atropurosides A–G Smilacina atropurpurea Rhizomes Zhang et al. (2006)16. Taccaoside C and D Tacca plantaginea Whole plant Liu et al. (2006)17. Timosaponin N, E1, E2, O and
PurpureagitosidAnemarrhena asphodeloides Rhizomes Kang et al. (2006)
18. Ophiopogonin E Ophiopogon japonicus Tubers Cheng et al. (2006)19. Solanigrosides C–H Solanum nigrum Whole plant Zhou et al. (2006)20. A pair of diastereoisomeric steroi-
dal saponinsTupistra chinensis Rhizomes Zou et al. (2006)
21. Steroidal saponins 1, 2, 3 and 4 Smilax china Whole Plant Shao et al. (2007)22. Ypsilandroside A, B, isoypsilan-
droside A,B and isoypsilandro-genin
Ypsilandra thibetica Whole plant Xie et al. (2006)
23. Polypunctosides A–D Polygonatum punctatum Rhizomes Yang and Yang (2006)24. Dioscin, protodioscin and pro-
togracillinDioscorea septemloba Thunb Rhizomes Tan et al. (2006)
25. Turrillianoside Veronica turrilliana Aerial parts Kostadinova et al. (2007)26. Six new steroidal saponins Asparagus acutifolius Roots Sautour et al. (2007)27. Yuccalan Yucca smalliana Leaves Jin et al. (2007)28. Cynanversicoside A, B, D, G,
glaucoside C and glaucogenin C-3-O-beta-d-thevetopyranoside
Radix Cynanchi Atrati Whole plant Liang et al. (2007)
29. Two new steroidal saponins Dracaena ombet Leaves Moharram and EI-Shenawy (2007)30. Filiasparosides A–D Asparagus filicinus Roots Zhou et al. (2007)31. A pair of stereoisomeric spi-
rostanol saponins and a new cholestane saponin
Paris polyphylla Rhizome Zhao et al. (2007)
32. Elephanoside A, elephanosides B–F, Ys-II, Ys-IV
Yucca elephantipes Stems Zhang et al. (2008)
33. Pseudoprotodioscin, methyl proto-dioscin and dioscin
Dioscorea nipponica Makino Rhizome Lin et al. (2007)
34. Methyl parvifloside, methyl pro-todeltonin, zingiberensis saponin I, deltonin
Dioscorea villosa Roots Hayes et al. (2007)
35. Paridiformoside B Lysimachia Paridiformis Whole plant Xu et al. (2007a, b)

527Planta (2018) 248:519–544
1 3
Table 1 (continued)
S. No. Steroidal saponin Plant Plant part References
36. Shatavarins VI–X Asparagus racemosus Roots Hayes et al. (2008)37. Aspafiliosides E and F Asparagus filicinus Roots Zhou and Chen (2008)38. Balanitin-6 and balanitin-7 Balanites aegyptiaca Kernels Gnoula et al. (2008)39. Ilwensisaponin A and C Verbascum pterocalycinum var.
mutenseFlowers Akkol et al. (2007)
40. Zingiberenin G Dioscorea zingiberensis Rhizomes Xu et al. (2007a, 2007b)41. Lyconosides Ia, Ib, II, III, and IV Solanum lycocarpum St. Hil. Fruits Nakamura et al. (2008)42. Five steroidal saponins Allium leucanthum C. Koch Flower Mskhiladze et al. (2008a, b)43. Cesdiurins I–III Cestrum diurnum L Leaves Fouad et al. (2008)44. Ophiofurospiside A Ophiopogon japonicus (Thunb.)
Ker-GawlTubers Xu et al. (2008)
45. Mannioside A Dracaena mannii Stem bark Tapondjou et al. (2008)46. Six C-21 steroidal glycosides Cynanchum auriculatum Royle ex
WightRoot tuber Peng et al. (2008)
47. Paris saponin I, Paris saponin V, Paris saponin VI, Paris saponin VII, Paris saponin H
Paris polyphylla Smith var steno-phylla Franch
Whole plant Yin et al. (2008)
48. Stemucronatoside K Stephanotis mucronata Whole plant Ye et al. (2008)49. Three new spirostanol glycosides,
a new furostanol glycoside, and a new cholestane glycoside
Clintonia udensis Rhizomes Matsuo et al. (2008)
50. Three steroidal saponins Paris polyphylla Smith Rhizomes Deng et al. (2008)51. Two new steroidal saponin along
with curillin G, asparagoside E, asparoside A, asparoside B
Smilax aspera subsp. mauritanica Roots Belhouchet et al. (2008)
52. Smilacinoside A, B, C, D and funkioside D, aspidistrin
Smilacina atropurpurea Dried tender aerial parts Yang et al. (2009)
53. Solanolactosides A, B, Torvosides M, N
Solanum torvum Aerial parts Lu et al. (2009)
54. Two new steroidal saponins Tribulus terrestris Fruits Su et al. (2009)55. Polygonoides A and B Polygonatum sibiricum Rhizome Xu et al. (2009a, b, c)56. Five new steroidal saponins Tribulus terrestris Fruits Su et al. (2009)57. Eleven steroidal saponins Trillium erectum Roots Hayes et al. (2009)58. Two new furostanol saponins and
one new spirostanol saponinParis polyphylla Rhizome Zhao et al. (2009)
59. Seven spirostanol type saponins Allium leucanthum Flowers Mskhiladze et al. (2008a, 2008b)60. Two new steroidal saponins Allium macrostemon bunge Dried bulbs Chen et al. (2009)61. Chloragin Chlorophytum nimonii (Grah)
DalzAerial part Lakshmi et al. (2009)
62. Three steroidal saponins Polygonatum odoratum Rhizomes Wang et al. (2009a, 2009b)63. Three new furostanol saponins
together with known protodi-oscin, pseudoprotodioscin and dioscin
Smilax excelsa Rhizomes Ivanova et al. (2009)
64. 15-hydroxy-pseudoprotodioscin and 15-methoxy-pseudoproto-dioscin
Smilax china Tubes Huang et al. (2009)
65. Padelaosides A and B Paris delavayi Rhizomes Zhang et al. (2009a, b)66. Ceparosides C and D Allium cepa L Seeds Yuan et al. (2009)67. Five spirostanol saponins, Three
furostanol saponinsAgave utahensis Whole plant Yokosuka and Mimaki (2009)
68. Zingiberenin F Dioscorea zingiberensis Rhizome Xu et al. (2009a, 2009b, 2009c)69. Ypsilandrosides C–G Ypsilandra thibetica Whole plant Xie et al. (2009)

528 Planta (2018) 248:519–544
1 3
Table 1 (continued)
S. No. Steroidal saponin Plant Plant part References
70. Eight new spirostanol saponins and three new furostanol sapo-nins
Agave utahensis Whole plants Yokosuka et al. (2009)
71. Shatavaroside A and shatavaroside B together with filiasparoside C
Asparagus racemosus Roots Sharma et al. (2009)
72. Yamogenin II Asparagus officinalis L Dried Stems Sun et al. (2010)73. Spirostanol saponin, chantriero-
side ATacca chantrieri Rhizomes Zhang et al. (2009a, b)
74. Three new steroidal saponins Tribulus terrestris L Fruits Liu et al. (2010a, b)75. Two new steroidal saponins Tribulus terrestris L Fruits Liu et al. (2010a, 2010b)76. Two steroidal saponin Hosta sieboldiana Leaf and Leafstalk Yada et al. (2010)77. Monodesmosidic spirostanol
saponiAgave macroacantha Zucc Leaves Eskander et al. (2010)
78. Orchidastrosides A–F, and chloro-maloside D
Chlorophytum orchidastrum Roots Acharya et al. (2010)
79. Timosaponins AIII, BIII, and D Anemarrhena asphodeloides Rhizome Lee et al. (2010)80. Tribufurosides D and E Tribulus terrestris L Fruits Xu et al. (2009a, 2009b, 2009c)81. Three new steroid saponin Tacca integrifolia Rhizome Shwe et al. (2010)82. Parvifloside, protodeltonin, proto-
dioscin, protogracillin, zingibe-rensis saponin, deltonin, dioscin and trillin
Dioscorea zingiberensis C.H. Wright
Rhizome Li et al. (2010)
83. Two new furostanol saponins Lilium longiflorum Thunb Bulbs Munafo et al. (2010)84. Angudracanosides A–F Dracaena angustifolia Fresh stems Xu et al. (2010)85. Esculeoside A Solanum lycopersicum Ripe Fruit Nohara et al. (2010)86. Filiasparosides E, F, filiasparoside
G, asparagusin A filiasparoside A, filiasparoside B, aspafilioside A, aspafilioside B, and filiaspa-roside C
Asparagus filicinus Roots Wu et al. (2010)
87. Fistulosaponins A–F Allium fistulosum Seeds Lai et al. (2010)88. Deistelianosides A, B and arbo-
reasaponins A and BDracaena deisteliana and Dra-
caena arboreaStem and bark Kougan et al. (2010)
89. Ypsilandrosides H–L and a known saponin polyphylloside III
Ypsilandra thibetica Whole plant Lu et al. (2010)
90. Two furostanol saponins Ruscus ponticus Leaves Napolitano et al. (2010)91. Two steroidal saponins Solanum surattense Burm. f. Aerial parts Lu et al. (2011)92. Two new steroidal saponins Agave sisalana Leaves Chen et al. (2011a, 2011b)93. Diosbulbisides D and E Dioscorea bulbifera Rhizomes Liu et al. (2011)94. Lirigramosides A (1) and B Liriope graminifolia (Linn.) Underground parts Wang et al. (2011a, b)95. Five steroidal glycosides, includ-
ing three spirostane, one furo-stane and one cholestane type saponins
Yucca gloriosa L Rhizomes Skhirtladze et al. (2011)
96. Helosides A and B Chamaelirium luteum Roots Challinor et al. (2011)97. A new steroidal saponin Ophiopogon japonicus Dried roots Qu et al. (2011)98. Polyphyllin D Paris polyphylla Rhizome Chan et al. (2011)99. Thirteen steroidal saponins Beaucarnea recurvata Leaves Eskander et al. (2011)100. Esculeoside B-5 Solanum lycopersicum Ripe fruit Ohno et al. (2011)101. DT-13 Liriope muscari (Decne.) Baily Tuber Ma et al. (2011)102. A bidesmosidic steroidal saponins Yucca schidigera Bark Kowalczyk et al. (2011)103. A new aglycone of amplexico-
genin BCynanchum amplexicaule Whole plant Chen et al. (2011a, 2011b)

529Planta (2018) 248:519–544
1 3
Table 1 (continued)
S. No. Steroidal saponin Plant Plant part References
104. Pallidiflosides A, B and C Fritillaria pallidiflora Dry bulbs Shen et al. (2011)105. Desmettianosides A and B Yucca desmettiana Leaves Diab et al. (2012)106. Parvifloside, methyl protodeltonin,
trigofoenoside A-1, zingiberensis saponin I, deltonin, dioscin and prosapogenin A of dioscin
Dioscorea villosa Rhizome Yoon et al. (2012)
107. Seven spirostane and furostane-type glycosides
Cestrum ruizteranianum Fruits Galarraga et al. (2011)
108. Timosaponin J, timosaponin K, (25S)-karatavioside C, timosapo-nin L, and (25S)-officinalisnin-I
Anemarrhena asphodeloides Rhizome Kang et al. (2012a, 2012b, 2012c)
109. Two new furostanol saponins sarsaparilloside B and sarsaparil-loside C
Smilax ornata Lem. Roots Challinor et al. (2012a, 2012b, 2012c)
110. A new steroidal saponin Allium ampeloprasum Bulbs Adão et al. (2012)111. Lycioside A and lycioside B Lycium barbarum Seeds Wang et al. (2011a, 2011b)112. Parisyunnanosides G–I, one new
C(21) steroidal glycoside, paris-yunnanoside J
Paris polyphylla Rhizome Kang et al. (2012a, 2012b, 2012c)
113. Two new C-21 steroidal glycosides Cynanchum amplexicaule Whole plant Chen et al. (2012)114. Dioscins E and F Dioscorea nipponica Rhizomes Zhang et al. (2012a, b)115. Ophiopogonins H–O Ophiopogon japonicus Tuber Zhang et al. (2012a, 2012b)116. Japonicoside A, japonicoside B
and japonicoside CSmilacina japonica Dried rhizomes and roots Liu et al. (2012a, b)
117. Pallidifloside D, pallidifloside E, pallidifloside G, pallidifloside H and pallidifloside I
Fritillaria pallidiflora Schrenk Dry bulbs Shen et al. (2012)
118. Tupisteroide A–C Tupistra chinensis Roots Liu et al. (2012a, 2012b)119. Two new steroidal saponins Tribulus terrestris Fruits Chen et al. (2013)120. Pariposides A–D Paris polyphylla var. yunnanensis Roots Wu et al. (2012a, b)121. 15 steroidal saponins Chamaelirium luteum (false
unicorn)Roots Challinor et al. (2012a, 2012b,
2012c)122. Chonglouosides SL-1-SL-6 Paris polyphylla Stems and Leaves Qin et al. (2012)123. A novel steroidal saponin Fagonia indica Aerial Parts Waheed et al. (2012)124. Eleven steroidal saponins Paris polyphylla Rhizomes Wu et al. (2012a, 2012b)125. Two new furostanol saponins 1
and 2Ruscus aculeatus L Underground parts De Marino et al. (2012)
126. Stauntosides C–K Cynanchum stauntonii Roots Yu et al. (2013)127. Ophiopogonin P–S Ophiopogon japonicus Tuberous roots Li et al. (2013)128. Two new steroidal saponins Smilax microphylla Roots and Rhizome Lin et al. (2012)129. A new isospirostanol-type steroi-
dal saponinSmilax scobinicaulis Roots and Rhizome Zhang et al. (2013a, 2013b, 2013c)
130. Shatavaroside C Asparagus racemosus Roots Sharma et al. (2012)131. Avenacosides A and B Avena sativa L Grains Pecio et al. (2012)132. Seven new steroidal saponins Lilium brownii var. viridulum Bulbs Hong et al. (2012)133. Tupichinin A Tupistra chinensis Rhizomes Pan et al. (2012)134. Pumilum A Lilium pumilum DC Bulbs Zhou et al. (2012)135. Three new steroidal saponins Dracaena marginata Bark Rezgui et al. (2013)136. Nolinospiroside F Ophiopogon japonicus Roots Sun et al. (2013)137. Three new steroidal compounds Hosta longipes Leaves Kim et al. (2013)138. Four spirostane-type glycosides Allium schoenoprasum Whole plant Timité et al. (2013)139. Seven steroidal glycosides Solanum torvum Fruits Colmenares et al. (2013)

530 Planta (2018) 248:519–544
1 3
Table 1 (continued)
S. No. Steroidal saponin Plant Plant part References
140. Torvpregnanosides A and B, two pregnane glycosides, and tor-voside Q, a 23-keto-spirostanol glycoside
Solanum torvum Aerial parts Lee et al. (2013)
141. Solanolactoside C Solanum torvum Swartz Aerial part Shu et al. (2013)142. Anemarnoside A and anemarno-
side BAnemarrhena asphodeloides Whole plant Liu et al. (2013)
143. Three furostanol type saponins Digitalis trojana Aerial parts Kirmizibekmez et al. (2014)144. Two new furostanol saponins Ophiopogon japonicus Tubers Guo et al. (2013)145. Two new steroidal saponins Selaginella uncinata (Desv.) Whole plant Zheng et al. (2013)146. Magueyosides A–E Agave offoyana Flowers Pérez et al. (2013)147. Three novel cholestane-type
steroidal glycosides and two novel spirostane-type steroidal saponins
Polygonatum odoratum Rhizome Bai et al. (2014)
148. Three new steroidal saponins Ophiopogon japonicus (Thunb.) Ker-Gawl
Tubers Ye et al. (2013)
149. Dioscoreanosides A–K Dioscorea bulbifera var. sativa Flowers Tapondjou et al. (2013)150. Drangustosides A–B Dracaena angustifolia Roxb Whole plant Huang et al. (2013)151. Longipetalosides A–C Tribulus longipetalus Whole plant Naveed et al. (2014)152. Cambodianosides A–F Dracaena cambodiana Whole plant Shen et al. (2014)153. Five oleanane-type saponins Ganophyllum giganteum Root bark Montes et al. (2014)154. Three new spirostane-type glyco-
sidesAllium flavum Whole plant Rezgui et al. (2014)
155. Five steroidal saponins Agave offoyana Leaves Pérez et al. (2014)156. Diospreussinosides A–C Dioscorea preussii Rhizomes Tabopda et al. (2014)157. Riparoside B and timosaponin J Smilax riparia Roots and Rhizomes Wu et al. (2014a, b)158. Zingiberenosides A and B Dioscorea zingiberensis Rhizomes Zheng et al. (2014)159. Two new furostanol saponins Asparagus cochinchinensis Roots Zhu et al. (2014)160. Cynanosides A–O Cynanchum atratum Roots Yan et al. (2014)161. Diospreussinosides A–C Dioscorea preussii Rhizomes Tabopda et al. (2014)162. Six pennogenyl saponins Paris quadrifolia L Rhizomes Gajdus et al. (2014)163. Seven steroidal saponins Anemarrhena asphodeloides Rhizomes Guo et al. (2015)164. Polygodosides A–F Polygonatum odoratum Fibrous roots Zhang et al. (2014a, 2014b)165. ASC Ophitopogin japonicas Whole plant Zeng et al. (2015)166. Smilaxchinoside A and smilaxchi-
noside CSmilax riparia Roots and Rhizomes Wu et al. (2014a, 2014b)
167. Sixteen steroidal saponin Tribulus terrestris Whole plant Kang et al. (2014)168. Timosaponin X and timosaponin
YAnemarrhena asphodeloides Rhizomes Yuan et al. (2014)
169. A new steroidal saponin Antigonon leptopus Whole plant Apaya and Chichioco-Hernandez (2014)
170. Spicatoside A and spicatoside D Liriope plathyphylla Whole Plant Choi et al. (2015)171. Ypsilandroside S and ypsilandro-
side TYpsilandra thibetica Whole Plant Si et al. (2014)
172. A new C21 steroidal saponin Azadirachta indica Leaves Liu et al. (2014)173. Four new spirostane steroidal
saponinsBletilla striata Roots Wang and Meng (2015)
174. Parisverticosides A–C Paris verticillata Aerial parts Sun et al. (2014)175. Periplocoside P Periplocae Cortex Whole plant Liu et al. (2015a, 2015b)176. Two new furostanol saponins and
a new spirostanol saponinTupistra chinensis Roots and Rhizomes Li et al. (2015a, 2015b, 2015c)
177. Spirostane-type saponins Dracaena fragrans Bark, Roots and leaves Rezgui et al. (2015)

531Planta (2018) 248:519–544
1 3
not available in public databases helps in the identification of prospective candidate genes involved in the biosynthesis of the secondary metabolic pathway and this will further increase our understanding related to biosynthesis, regula-tion, and diversity of secondary metabolites. Some of the medicinal plants that were explored to understand the ste-roidal saponin biosynthesis are discussed below and sum-marized in Fig. 3.
Dioscorea species
Mainly steroidal saponins are found in the monocot spe-cies such as some species of Dioscoreaceae family. Steroi-dal saponins of this plant species, i.e., diosgenin is used as the raw material for industrial production of steroidal drugs (Lin and Yang 2008). Comparative transcriptome analy-sis of two high saponin-producing Dioscorea esculenta and Dioscorea cayenensis and a low saponin-producing Dioscorea alata was performed using 454 pyrosequencing to investigate steroidal saponin biosynthesis in Dioscorea spe-cies. From transcriptome analysis, it was found that DeF26G
transcript was highly expressed in Dioscorea esculenta but not in Dioscorea alata. The biochemical characteriza-tion of DeF26G revealed that it was a furostanol glycoside 26-O-β-glucosidase that was involved in the conversion of protodioscin (furostanol saponin) to dioscin (spirostanol saponin). The expression of the DeF26G1 gene in the leaves of Dioscorea esculenta was higher than that in the tubers (Nakayasu et al. 2015). Using Illumina deep sequencing Dioscorea composita was used to analyze the transcriptome of 18-month-old Dioscorea composita. This information allows us to understand the biosynthetic pathway of steroi-dal sapogenins (diosgenin) in Dioscorea composita (Wang et al. 2015a, b, c).
Asparagus racemosus
Steroidal saponins in Asparagus racemosus mainly accumu-lates in the root system. The dried root of Asparagus is used as an ayurvedic medicine. The saponin present in the root of Asparagus collectively known as shatavarins. Leaf versus root transcriptome of Asparagus racemosus using Illumina
Table 1 (continued)
S. No. Steroidal saponin Plant Plant part References
178. Two new steroidal saponins Solanum paniculatum L Aerial parts Vieira Júnior et al. (2015)179. Cambodianoside G Dracaena cambodiana Rhizome Luo et al. (2015)180. Polyhydroxy hellebosaponins Helleborus niger L Roots Duckstein et al. (2014)181. Four new furostanol glycosides Hosta plantaginea (Lam.) Aschers Flowers Li et al. (2015a, 2015b, 2015c)182. Trigoneoside XIIIa, parvifloside,
trigoneoside IVa, deltoside, protobioside, lilioglycoside k, zingiberensis newsaponin I, del-tonin, prosapogenin A of dioscin, and trillin
Dioscorea zingiberensis Tubers Wang et al. (2014a, 2014b, 2014c)
183. Eight steroidal saponins Raphia farinifera Mesocarp of the fruits Tapondjou et al. (2015)184. Three new steroidal saponins Helleborus thibetanus Dried roots and rhizomes Zhang et al. (2016a, b)185. Two new steroidal saponins Avena sativa Oat Bran Yang et al. (2016)186. Typaspidosides B–L Aspidistra typica Rhizomes Cui et al. (2016)187. Chlorodeistelianosides A–D Chlorophytum deistelianum Aerial parts Tabopda et al. (2016)188. Esculeosides B-1 and B-2 Solanum lycopersicum Tomato juice Nohara et al. (2015)189. Pratioside G and H Polygonatum prattii Rhizomes Zhang et al. (2016a, 2016b)190. Govanoside A Trillium govanianum Rhizomes Shafiq-ur-Rahman et al. (2015)191. Chonglouosides SL-9-SL-20 Paris polyphylla var. yunnanensis Stems and leaves Qin et al. (2016)192. TTB2 Trillium tschonoskii Maxim Whole plant Huang and Zou (2015)193. Timosaponin B-II Anemarrhena asphodeloides Whole plant Yuan et al. (2015)194. Cynawilfosides A–I Cynanchum wilfordii Roots Li et al. (2016)195. Two new steroidal saponins Cestrum laevigatum L Leaves Ribeiro et al. (2016)196. Taccavietnamosides A–E Tacca vietnamensis Rhizomes Yen et al. (2016)197. Timosaponin AIII Anemarrhena asphodeloides Bge Rhizomes Wang et al. (2016a, 2016b)198. Methyl protodioscin Dioscorea nipponica Roots Chung et al. (2016)199. Padelaosides C–F Paris delavayi Rhizomes Liu et al. (2016a, 2016b)200. Terrestrinin J–T Tribulus terrestris Whole plant Wang et al. (2016a, 2016b)

532 Planta (2018) 248:519–544
1 3
sequencing approach revealed that majority of transcripts related to steroidal saponin biosynthesis including P450 and UDP-glucosyltransferase was found to be upregulated in root tissue as compared to leaf. This will lead to the hypothesis that steroidal saponin accumulated in the roots of Asparagus racemosus, which is mainly responsible for its pharmaco-logical properties (Upadhyay et al. 2014).
Chlorophytum borivilianum
Safed musli has high saponin and polysaccharide content but the main active compound known as borivilianosides is found in roots. Steroidal saponins, borivilianosides (spi-rostanol type), are the major type of saponins present in Chlorophytum borivilianum. Earlier phytochemical stud-ies indicated that initial reactions of saponins biosynthetic pathway occur in leaves, while later step modifications and
storage occurs in roots (Kalra et al. 2013). Illumina HiSeq 2000 sequencing analysis of root tissue of this plant helps in the identification of candidate genes that encode enzymes controlling later stages of the saponin biosynthetic pathway and modifications (Kumar et al. 2016; Kalra et al. 2013).
Paris polyphylla
The rhizome as well as leaves and stem of Paris polyphylla is an important source of steroidal saponins. The root tissue is the main site of steroid saponin biosynthesis in Paris poly-phylla so comparative transcriptome analysis was performed between 8-year-old and 4-year-old roots of Paris polyphylla. Comparative saponin biosynthesis in different aged roots showed that genes including P450 and UGTs involved in saponin biosynthesis were upregulated in the 8-year-old
More F26G
enzyme
A. fistulosum Monosomic addition lines
(MALs)
A.cepa aggregatum group (AA)
Transcriptome analysis of
steroidal saponin producing
medicinal plants
Dioscorea Species
Parispolyphylla
Trigonella foenum graecum
Allium Species
Trillium govanianum
Asparagus racemosus
Chlorophytum borivilianum
D. Composita and
D. zingiberensis
D.alata
D.esculenta
D.cayenensis
Leaf Root
Leaf Root
Leaf Fruit
Stem Rhizome
4 year old root
8 year old root
More Sterol
Squalene treatment
Cholesterol treatment MeJA
treatment
Leaf Stem
Tuber
More steroidal saponin and more accumulation of transcripts related to saponin biosynthesis
Fig. 3 Representation of steroidal saponin-producing plant with tran-scriptome data: diagrammatic representation of steroidal saponin-pro-ducing non-model plants with their plant part whose comparative or non-comparative transcriptome sequencing has been performed using
NGS platform. Purple star indicate more steroidal saponin accumula-tion and more number of upregulated transcripts related to steroidal saponin biosynthesis

533Planta (2018) 248:519–544
1 3
root, an understandable finding considering that the amount of saponin is higher in older roots (Liu et al. 2016a, b).
Trigonella foenum‑graecum
The seeds and leaves of Trigonella foenum-graecum (fenu-greek) are used as ayurvedic medicine in the Indian sub-continent to treat gastrointestinal ailments, high cholesterol, diabetes, wounds and inflammation. Leaves and seeds of this plant are edible and can be used as condiments (Chevallier 2000). Representational difference analysis (RDA) of cDNA of was performed using transcriptome user-friendly analysis (TRUFA, a webserver platform dedicated to RNA-seq analy-sis) was used to identify up-regulated genes in fenugreek in response to MeJA, cholesterol and squalene (Kornobis et al. 2015). MeJA mainly act as elicitor in plant defense under biotic and abiotic stresses. Majority of the transcripts involved in diosgenin biosynthesis including various P450 was found to be upregulated after MeJA treatment (Ciura et al. 2017) that further confirms the involvement of MeJA in the upregulation of secondary biosynthetic pathway related gene.
Trillium govanianum
Diosgenin that accumulates in rhizome as “Trillarin” is con-sidered as the main bioactive constituent of Trillium gova-nianum. In addition to diosgenin govanoside, borassoside, and pennogenin were also reported to be present in the rhi-zome of Trillium govanianum (Rahman et al. 2015). Spatial differential expression of genes associated with steroidal saponin biosynthesis in rhizome, stem, leaf and fruit tissues of Trillium govanianum were studied using Illumina GAIIx sequencing platform. The expression analysis revealed that maximum numbers of genes are involved in saponin bio-synthesis and found to be highly expressed in fruit and leaf tissues, indicating active biosynthesis of steroidal sapo-nin takes place in fruit and leaf tissues. The downstream genes of steroidal saponin biosynthesis in leaf (SQS, CPI1, CyP5G1, FK, HDY1 and SMO2) and fruit (SQLE, SMT1, SMO1, DWF5, UGT80B1 and β-glucosidase) also showed higher expression level. These results indicate that steroidal saponins biosynthesis mainly occur in leaf and fruit tissues (Singh et al. 2017).
Allium species
Steroidal saponins were reported in more than 40 differ-ent species of allium. Tigogenin, diosgenin, alliogenin, gitogenin, agigenin, and β-chlorogenin are among the most common spirostanol sapogenins identified in the plants of allium species (Sobolewska et al. 2016). High-through-put RNA-Seq of the root, bulb and leaf tissues of Allium
fistulosum with extra chromosome 2A from shallot (FF2A), monosomic addition lines (MALs) and Allium cepa aggreg-atum group (AA) was carried out using Illumina’s HiSeq 2500 platform to gain molecular insight into the Allium sap-onin biosynthesis pathway. It was observed that in FF2A line acetyl-CoA-acetyltransferase and SQS are highly expressed transcripts that are related to saponin biosynthesis. Further oxidation, hydroxylation and glycosylation steps of the sapo-nin backbone via P450 and UGT family transcripts, respec-tively, were also found to be remarkably up-regulated in the FF2A line (Abdelrahman et al. 2017).
Production of steroidal saponin in culture media
Due to the limited availability of medicinal plants and slow production of medicinally important secondary metabo-lites, it is necessary to develop in vitro techniques for the production of bioactive compounds that can be useful for pharmacological purposes. Advances in functional genom-ics and application of multidisciplinary approaches help to discover new plant saponins that have different pharmaco-logical properties. Plant cell and tissue culture methods have been explored as potentially more efficient alternatives for the production of saponin and other plant derivatives. These methods were started many years ago but the optimization of culture condition was still going on to increase the saponin production for commercial use.
In vitro culture methods to obtain steroidal saponin can be possible by optimizing plant growth regulator/plant hor-mone concentration in culture media, light treatment, growth conditions, precursor feeding and other parameters to obtain better results. For example, plant hormone auxin was used to induce the production of steroidal saponin solanine, solaso-dine and solanidine in the tissue and suspension culture of Solanum lyratum (Kuo et al. 2012). Similarly, steroidal sapo-nin (aculeatiside A, aculeatiside B) production was observed in considerable amount in the hairy root culture of Solanum aculeatissimum under continuous light treatment for 8 weeks (Ikenaga et al. 1995). Epicotyls of Solanum aculeatissimum were used to induce callus culture and it was observed that it was able to produce steroidal saponins aculeatiside A and B (Ikenaga et al. 2000). Attempts were also made for the production of steroidal saponins from the roots of Asparagus racemosus and its callus and suspension cultures were estab-lished to produce high levels of saponins both intracellularly and extracellularly. Being an endangered species it is neces-sary to propagate this plant through tissue culture because traditional method of propagation through seeds is not an efficient method and it takes a lot of time. Using in vitro cul-ture methods 20 times higher levels of saponins (shatavarins)

534 Planta (2018) 248:519–544
1 3
were produced that has found to stimulate immunological response (Pise et al. 2015).
Root cultures are considered as reliable and alternative sources for the production of valuable secondary metabo-lites. It is easy to culture root tissues of dicot plant but in the case of monocot plants where root forms modified struc-tures such as bulb, corm or tubers the root culture seems to be difficult but the tuberous root culture of Chlorophy-tum borivilianum was developed and considerable amount of saponin was quantified (Basu and Jha 2015). Effect of growth regulators was determined in the adventitious root culture of Dioscorea nipponica and it was observed that dioscin and prosapogenin C content in indole-3-butyric acid (IBA) treated roots were significantly higher as compared to 1-naphthaleneacetic acid (NAA) treated roots (Ahn et al. 2005). In in vitro shoot culture of Bacopa monnieri it was observed that application of abiotic elicitors (MeJA, salicylic acid, and CuSO4) enhances bacoside production and among these elicitors, CuSO4 was found to accumulate bacoside in a large amount (Sharma et al. 2015).
Somatic embryogenesis is a valuable technique in medici-nal plant improvement programs, such as for propagation and genetic transformation. Embryogenic callus culture derived from the leaf explants of Tribulus terrestris L. accu-mulated steroidal saponin, i.e., diosgenin. Saponin content in embryogenic callus culture of Tribulus terrestris was found to be similar to that of seeds of naturally growing plants after the application of casein hydrolysate (CH) in growth media (Nikam et al. 2009). Fagonia indica burm was cultured in vitro on MS medium supplemented with 2,4-d for rapid regeneration of plant. Somatic embryogenesis was induced by the elimination of 2,4-dichlorophenoxyacetic acid (2,4-d) and a slight increase of 6-benzylaminopurine (BA) concen-tration (Ebrahimi and Payan 2013). Shoot organ culture of Yucca schidigera accumulated all kinds of sapogenins as in plant shoot but in different concentrations and their glyco-side pattern was similar to that of saponins present in mature Yucca schidigera rhizome (Kaneda et al. 1987).
Steroidal saponin production in cell and tissue culture of certain plant species can also be increased by the addition of hormone or biotic and abiotic growth regulators like the production of diosgenin was stimulated by light or fungal elicitation in the cell suspension cultures of Dioscorea gale-ottiana when cells were grown in the dark condition. Appli-cation of low phosphate and sucrose in the growth medium results in highest production of diosgenin (Rojas et al. 1999). Diosgenin accumulation in the tissue culture of Dioscorea deltoidea was enhanced in the presence of 2,4-d (Marshall and Staba 1976).
Microbial transformation of saponin present in Dioscorea zingiberensis tubers was difficult due to the association of saponin with starch so it cannot be able to make con-tact with microorganisms so a combined technology of
enzymatic saccharification for excluding starch from raw herb and the microbial transformation was established to prepare diosgenin from the tubers of Dioscorea zingiber-ensis. Using these approach saponins from Dioscorea zin-giberensis was converted into diosgenin using the fungal culture of Trichoderma reesei (Zhu et al. 2010). A strain of Gibberella intermedia was also used for conversion of saponins into diosgenin by optimizing culture conditions, and using this method three times higher diosgenin yield was observed as compared to original culture media (Zhang et al. 2013a, b, c). In addition to this, nine steroidal saponins were obtained by the fermentation of total furostanol sapo-nins from Dioscorea zingiberensis tubers incubated with the culture of a fungus Absidia coerulea. Two compounds among them show induced platelet aggregation activity due to change in their chemical structure (Pang et al. 2015). Bio-transformation of total steroidal saponins of Agave sisalana into tigogenin was achieved using a rod-shaped bacterium obtained from soil samples of karst area of Guilin, China. The microbial transformation is an eco-friendly approach in the production of tigogenin (Wang et al. 2014a, b, c).
Rhizomes and roots of Ruscus aculeatus contain steroidal saponins (ruscogenins) that have several pharmacological properties. Due to its slow growth rate, complicated cultiva-tion methods and pollination failure make this plant a vul-nerable species. Ruscogenin biosynthesis in Ruscus aculea-tus is greatly influenced by its clonal origin and culture type that forms the basis of its ex situ conservation (Ivanova et al. 2015). In vitro culture of Ruscus aculeatus has a potential capacity to biosynthesize steroidal saponin. Rhizome seg-ments of this plant are used to generate calli. It was observed that young regenerated plants of about 6 weeks old have a greater capacity to biosynthesize and accumulate saponins than calli cultures and the neoruscogenin contents were found to be greater than those of ruscogenin in all samples, i.e.,plantlets and calli (Palazón et al. 2006).
It is challenging to produce saponins in the heterologous system, i.e., in microbes, and only limited successful path-way engineering attempts have been reported. The effects of several previously unexplored gene knockout targets of Sac-charomyces cerevisiae were assessed for the heterologous production and to improve the production capabilities of this saponin production platform. Dramatic expansion of the endoplasmic reticulum stimulates the production of recom-binant triterpene biosynthetic enzymes through disruption of the phosphatidic acid phosphatase-encoding PAH1 of Sac-charomyces cerevisiae through CRISPR/Cas9 which results in the accumulation of triterpenoids and triterpene sapo-nin (Arendt et al. 2017). Transgenic yeast strains express-ing β-amyrin synthase (bAS), P450 reductase, CyP93E2 and CyP72A61v2 were able to produce soyasapogenol B, and yeast strain over expressing bAS, P450 reductase, CyP716A12 and CyP72A68v2 produce gypsogenic acid.

535Planta (2018) 248:519–544
1 3
Additionally, P450 s that seemed not to work together in planta were combinatorially expressed in transgenic yeast for the production of saponins (Fukushima et al. 2013). Com-binatorial synthetic biology program was also successful for the synthesis of monoglycosylated saponins in yeast by combining CyP716Y1 with oxidosqualene cyclase, P450, and glycosyltransferase genes from different plant species (Moses et al. 2014a, b). Another transgenic yeast strain expressing dammarenediol-II synthase and protopanaxadiol synthase genes of Panax ginseng, together with a NADPH-cytochrome P450 reductase gene of Arabidopsis thaliana, were able to produce protopanaxadiol (Dai et al. 2013) and bioactive ginsenosides Rh2 or Rg3 were produced from sim-ple sugars by microbial fermentation (Wang et al. 2015a, b, c). These platforms will provide an alternative approach to replace the traditional method of ginsenosides extraction from Panax plants. Elicitation process is another one of the most effective strategies to enhance metabolites biosynthesis and accumulation in biotechnological systems. By adopting this elicitation strategy, yeast extract can be used to elicit ginsenoside production in hairy root cultures of Panax quinquefolium (Kochan et al. 2017). Reports were avail-able, that employ synthetic biology approach for heterolo-gous production of triterpenoidal saponin but this technique was not reported in context of steroidal saponin production. Efforts will be made for the development of engineered yeast system that will provide an efficient system for large-scale production of steroidal saponins in near future.
Future prospects
Steroidal saponins possess a wide range of medicinal and biological properties due to the presence of structural diver-sity in them. To find out new plant sources of steroidal sapo-nins and to identify the presence of new steroidal saponins in the known medicinal plants will be the area of interest of many researchers. The anti-cancerous properties of most of the steroidal saponins were already being explored widely and is still going on but the other pharmacological proper-ties associated with this compound is yet to be investigated.
The emergence of NGS platform has revolutionized our understanding related to saponin biosynthesis especially the modification of saponin backbone. Most of the P450 s and UGTs that were involved in terpenoidal saponin biosynthesis have been characterized but identification and characteriza-tion of these two enzymes that will convert obtusifoliol to furostanol and spirostanol type of steroidal saponin are yet to be needed as they are responsible for providing different biological properties to steroidal saponins. Transcriptome data of non-model plant will be helpful in discovering new candidates of P450 s and UGTs that may be involved in the
saponin backbone modification but they need to be charac-terized further.
The advancement in the synthetic biotechnology tools like CRISPR/Cas9-based approaches will be beneficial to engineer medicinal plants regarding secondary metabolites production and to functionally characterize the biosynthetic (backbone modifying) and regulatory enzymes related to steroidal saponin biosynthesis. Due to the lack of availabil-ity of functional mutants in most of the medicinal plants, the genome editing technology based on type II CRISPR/Cas system can be utilized to establish mutants for gene function analysis. This technique relies on Agrobacterium tumefaciens mediated transformation technology and can also be combined with Agrobacterium rhizogenes-mediated transformation. Agrobacterium rhizogenes-based Ri trans-formed hairy roots show the characteristics of rapid growth, reduced apical dominance, high branching, and enhanced stable production of secondary metabolites, making them a promising system for investigating biosynthesis of vari-ous secondary metabolites, especially in medicinal plants. CRISPR/Cas9 system has a wide range of applications, with more accuracy and precision as compared to other genome editing tools. This approach can be applied for converting saponin-producing plants into biofactories for mass pro-duction of saponin by diverting the flux towards steroidal biosynthetic pathway. For example, by blocking the bAS gene of triterpenoidal saponin biosynthetic pathway, phy-toene synthase gene of carotenoid biosynthetic pathway or geraniol synthase gene of alkaloid biosynthesis, metabolic flux can be shifted towards steroidal saponin production. Additionally this technique will be beneficial for elucidat-ing the function of novel P450 and UGTs. As the life cycle of saponin metabolite producing plants is generally high so Cas9 based system can further be employed to control the spatiotemporal patterns of gene expression in plants and modulating life cycles of various economically useful plants to obtain useful metabolites in short duration of time.
Saponin molecules are synthesized in plants in very trace amount that will not satisfy the needs of its commercial use and the plant sources are also very limited. So there will be the need for the production of saponins using in vitro cul-ture methods. Tissue culture-based techniques were already being developed and need to be further optimized. Combi-natorial biosynthesis of steroidal saponins in yeast system will be a part of future research and research is going on to generate semi-synthetic steroidal saponin.
Author contribution statement SU has compiled and written the manuscript. GSJ and S have edited the manuscript. RKS has conceptualized, supervised and edited the manuscript.
Funding Swati, Gajendra, and Shikha acknowledge CSIR-UGC for fellowship. Shukla RK would like to acknowledge

536 Planta (2018) 248:519–544
1 3
SCIENCE & ENGINEERING RESEARCH BOARD EMR/2016/005256 (GAP397) for funding.
References
Abbassi SJ, Vishwakarma RK, Patel P, Kumari U, Khan BM (2015) Bacopa monniera recombinant mevalonate diphosphate decar-boxylase: biochemical characterization. Int J Biol Macromol 79:661–668
Abdelrahman M, El-Sayed M, Sato S, Hirakawa H, Ito SI, Tanaka K, Mine Y, Sugiyama N, Suzuki M, Yamauchi N, Shigyo M (2017) RNA-sequencing-based transcriptome and biochemi-cal analyses of steroidal saponin pathway in a complete set of Allium fistulosum—A. cepa monosomic addition lines. PLoS ONE 12:e0181784
Abe I, Rohmer M, Prestwich GD (1993) Enzymatic cyclization of squalene and oxidosqualene to sterols and triterpenes. Chem Rev 93:2189–2206
Acharya D, Mitaine-Offer AC, Kaushik N, Miyamoto T, Paululat T, Mirjolet JF, Duchamp O, Lacaille-Dubois MA (2010) Steroidal saponins from Chlorophytum orchidastrum. J Nat Prod 73:7–11
Adão CR, Pereira da Silva B, Tinoco LW, Parente JP (2012) Haemo-lytic activity and immunological adjuvant effect of a new ste-roidal saponin from Allium ampeloprasum var. porrum. Chem Biodivers 9:58–67
Ahn JH, Son KH, Sohn HY, Kwon ST (2005) In vitro culture of adven-titious roots from Dioscorea nipponica Makino for the produc-tion of steroidal saponins. Kor J Plant Biotechnol 32:217–223
Ahn MJ, Kim CY, Yoon KD, Ryu MY, Cheong JH, Chin YW, Kim J (2006) Steroidal saponins from the rhizomes of Polygonatum sibiricum. J Nat Prod 69:360–364
Akhtar N, Gupta P, Sangwan NS, Sangwan RS, Trivedi PK (2013) Cloning and functional characterization of 3-hydroxy-3-methyl-glutaryl coenzyme A reductase gene from Withania somnifera: an important medicinal plant. Protoplasma 250:613–622
Akkol EK, Tatli II, Akdemir ZS (2007) Antinociceptive and anti-inflammatory effects of saponin and iridoid glycosides from Verbascum pterocalycinum var. mutense. Hub Mor Z Naturforsch C 62:813–820
Aoyagi K, Beyou A, Moon K, Fang L, Ulrich T (1993) Isolation and characterization of cDNAs encoding wheat 3-hydroxy-3-meth-ylglutaryl coenzyme A reductase. Plant Physiol 102:623–628
Apaya MK, Chichioco-Hernandez CL (2014) New steroidal saponin from Antigonon leptopus Hook. and Arn. Pharmacogn Mag 10:S501–S505
Arendt P, Miettinen K, Pollier J, De Rycke R, Callewaert N, Goossens A (2017) An endoplasmic reticulum-engineered yeast platform for overproduction of triterpenoids. Metab Eng 40:165–175
Augustin JM, Kuzina V, Andersen SB, Bak S (2011) Molecular activi-ties, biosynthesis and evolution of triterpenoid saponins. Phyto-chemistry 72:435–457
Babiychuk E, Bouvier-Navé P, Compagnon V, Suzuki M, Muranaka T, Van Montagu M, Kushnir S, Schaller H (2008) Albinism and cell viability in cycloartenol synthase deficient Arabidopsis. Plant Signal Behav 3:978–980
Bai H, Li W, Zhao H, Anzai Y, Li H, Guo H, Kato F, Koike K (2014) Isolation and structural elucidation of novel cholestane glyco-sides and spirostane saponins from Polygonatum odoratum. Steroids 80:7–14
Bak S, Kahn RA, Olsen CE, Halkier BA (1997) Cloning and expres-sion in Escherichia coli of the obtusifoliol 14α-demethylase of Sorghum bicolor (L.) Moench, a cytochrome P450 orthologous
to the sterol 14α-demethylases (CYP51) from fungi and mam-mals. Plant J 11:191–201
Bak S, Beisson F, Bishop G, Hamberger B, Höfer R, Paquette S, Werck-Reichhart D (2011) Cytochromes p450. Arabidopsis Book 9:e0144
Basu S, Jha TB (2015) In vitro root culture: an alternative source of bioactives in the rare aphrodisiac herb Chlorophytum bori-vilianum Sant et Fern. Plant Tissue Cult Biotech 23:133–146
Belhouchet Z, Sautour M, Miyamoto T, Lacaille-Dubois MA (2008) Steroidal saponins from the roots of Smilax aspera subsp. mau-ritanica. Chem Pharm Bull (Tokyo) 56:1324–1327
Bhattacharyya MK, Paiva NL, Dixon RA, Korth KL, Stermer BA (1995) Features of the hmg 1 subfamily of genes encoding HMG-CoA reductase in potato. Plant Mol Biol 28:1–15
Bhuvanalakshmi G, Basappa Rangappa KS, Dharmarajan A, Sethi G, Kumar AP, Warrier S (2017) Breast cancer stem-like cells are inhibited by diosgenin, a steroidal saponin, by the attenuation of the Wnt β-catenin signaling via the Wnt antagonist secreted frizzled related protein-4. Front Pharmacol 8:124
Cabello-Hurtado F, Taton M, Forthoffer N, Kahn R, Bak S, Rahier A, Werck-Reichhart D (1999) Optimized expression and catalytic properties of a wheat obtusifoliol 14α-demethylase (CYP51) expressed in yeast. Complementation of erg11Delta yeast mutants by plant CYP51. Eur J Biochem 262:435–446
Cao X, Zong Z, Ju X, Sun Y, Dai C, Liu Q, Jiang J (2010) Molecu-lar cloning, characterization and function analysis of the gene encoding HMG-CoA reductase from Euphorbia Pekinensis Rupr. Mol Biol Rep 37:1559–1567
Challinor VL, De Voss JJ (2013) Open-chain steroidal glycosides, a diverse class of plant saponins. Nat Prod Rep 30:429–454
Challinor VL, Stuthe JM, Bernhardt PV, Lehmann RP, Kitching W, De Voss JJ (2011) Structure and absolute configuration of helo-sides A and B, new saponins from Chamaelirium luteum. J Nat Prod 74:1557–1560
Challinor VL, Parsons PG, Chap S, White EF, Blanchfield JT, Lehmann RP, De Voss JJ (2012a) Steroidal saponins from the roots of Smilax sp.: structure and bioactivity. Steroids 77:504–511
Challinor VL, Piacente S, De Voss JJ (2012b) NMR assignment of the absolute configuration of C-25 in furostanol steroidal saponins. Steroids 77:602–608
Challinor VL, Stuthe JM, Parsons PG, Lambert LK, Lehmann RP, Kitching W, De Voss JJ (2012c) Structure and bioactivity of ste-roidal saponins isolated from the roots of Chamaelirium luteum (false unicorn). J Nat Prod 75:1469–1479
Chan JY, Koon JC, Liu X, Detmar M, Yu B, Kong SK, Fung KP (2011) Polyphyllin D, a steroidal saponin from Paris polyphylla, inhibits endothelial cell functions in vitro and angiogenesis in zebrafish embryos in vivo. J Ethnopharmacol 137:64–69
Chen HF, Wang GH, Luo Q, Wang NL, Yao XS (2009) Two new ste-roidal saponins from Allium macrostemon bunge and their cyto-toxity on different cancer cell lines. Molecules 14:2246–2253
Chen G, Chen H, Li W, Pei YH (2011a) Steroidal glycosides from Cynanchum amplexicaule. J Asian Nat Prod Res 13:756–760
Chen PY, Chen CH, Kuo CC, Lee TH, Kuo YH, Lee CK (2011b) Cytotoxic steroidal saponins from Agave sisalana. Planta Med 77:929–933
Chen G, Yi S, Hua HM, Lu X, Pei YH (2012) Two new steroidal gly-cosides from Cynanchum amplexicaule. J Asian Nat Prod Res 14:559–563
Chen G, Su L, Feng SG, Lu X, Wang H, Pei YH (2013) Furostanol saponins from the fruits of Tribulus terrestris. Nat Prod Res 27:1186–1190
Chen Y, Tang YM, Yu SL, Han YW, Kou JP, Liu BL, Yu BY (2015) Advances in the pharmacological activities and mechanisms of diosgenin. Chin J Nat Med 13:578–587

537Planta (2018) 248:519–544
1 3
Chen MH, Chen XJ, Wang M, Lin LG, Wang YT (2016) Ophiopogon japonicus—A phytochemical, ethnomedicinal and pharmacologi-cal review. J Ethnopharmacol 181:193–213
Cheng ZH, Wu T, Yu BY (2006) Steroidal glycosides from tubers of Ophiopogon japonicus. J Asian Nat Prod Res 8:555–559
Chevallier A (2000) Encyclopedia of herbal medicine. Dorling Kinder-sley Publishing, New York, p 271
Choi DW, Jung J, Ha YI, Park HW, In DS, Chung HJ, Liu JR (2005) Analysis of transcripts in methyl jasmonate-treated ginseng hairy roots to identify genes involved in the biosynthesis of ginsenosides and other secondary metabolites. Plant Cell Rep 23:557–566
Choi SJ, Choi J, Jeon H, Bae SK, Ko J, Kim J, Yoon KD (2015) Appli-cation of high-performance countercurrent chromatography for the isolation of steroidal saponins from Liriope plathyphylla. J Sep Sci 38:18–24
Chung YL, Pan CH, Wang CC, Hsu KC, Sheu MJ, Chen HF, Wu CH (2016) Methyl protodioscin, a steroidal saponin, inhibits neoin-tima formation in vitro and in vivo. J Nat Prod 79:1635–1644
Chye ML, Kush A, Tan CT, Chua NH (1991) Characterization of cDNA and genomic clones encoding 3-hydroxy-3-methylglu-taryl-coenzyme A reductase from Hevea brasiliensis. Plant Mol Biol 16:567–577
Ciura J, Szeliga M, Grzesik M, Tyrka M (2017) Next-generation sequencing of representational difference analysis products for identification of genes involved in diosgenin biosynthesis in fenu-greek (Trigonella foenum-graecum). Planta 245:977–991
Colmenares AP, Rojas LB, Mitaine-Offer AC, Pouységu L, Quideau S, Miyamoto T, Tanaka C, Paululat T, Usubillaga A, Lacaille-Dubois MA (2013) Steroidal saponins from the fruits of Solanum torvum. Phytochemistry 86:137–143
Cordoba E, Porta H, Arroyo A, San Román C, Medina L, Rodriguez-Concepcion M, León P (2011) Functional characterization of the three genes encoding 1-deoxy-d-xylulose 5-phosphate synthase in maize. J Exp Bot 62:2023–2038
Cui JM, Kang LP, Zhao Y, Zhao JY, Zhang J, Pang X, Yu HS, Jia DX, Liu C, Yu LY, Ma BP (2016) Steroidal saponins from the rhi-zomes of Aspidistra typica. PLoS ONE 11:e0150595
Dai Z, Cui G, Zhou SF, Zhang X, Huang L (2011) Cloning and char-acterization of a novel 3-hydroxy-3-methylglutaryl coenzyme A reductase gene from Salvia miltiorrhiza involved in diterpenoid tanshinone accumulation. J Plant Physiol 168:148–157
Dai Z, Liu Y, Zhang X, Shi M, Wang B, Wang D, Huang L, Zhang X (2013) Metabolic engineering of Saccharomyces cerevisiae for production of ginsenosides. Metab Eng 20:146–156
De Lucca AJ, Boue S, Palmgren MS, Maskos K, Cleveland TE (2006) Fungicidal properties of two saponins from Capsicum frutescens and the relationship of structure and fungicidal activity. Can J Microbiol 52:336–342
De Marino S, Festa C, Zollo F, Iorizzi M (2012) Novel steroidal com-ponents from the underground parts of Ruscus aculeatus L. Mol-ecules 17:14002–14014
Deng D, Lauren DR, Cooney JM, Jensen DJ, Wurms KV, Upritchard JE, Cannon RD, Wang MZ, Li MZ (2008) Antifungal saponins from Paris polyphylla Smith. Planta Med 74:1397–1402
Dhar N, Rana S, Razdan S, Bhat WW, Hussain A, Dhar RS, Vaishnavi S, Hamid A, Vishwakarma R, Lattoo SK (2014) Cloning and functional characterization of three branch point oxidosqualene cyclases from Withania somnifera (L.) dunal. J Biol Chem 289:17249–17267
Diab Y, Ioannou E, Emam A, Vagias C, Roussis V (2012) Desmetti-anosides A and B, bisdesmosidic furostanol saponins with mol-luscicidal activity from Yucca desmettiana. Steroids 77:686–690
Diarra ST, He J, Wang J, Li J (2013) Ethylene treatment improves diosgenin accumulation in in vitro cultures of Dioscorea
zingiberensis via up-regulation of CAS and HMGR gene expres-sion. Elect J Biotech 16:1–10
Duckstein SM, Lorenz P, Conrad J, Stintzing FC (2014) Tandem mass spectrometric characterization of acetylated polyhydroxy helle-bosaponins, the principal steroid saponins in Helleborus niger L. roots. Rapid Commun Mass Spectrom 28:1801–1812
Ebrahimi MA, Payan A (2013) Induction of callus and somatic embry-ogenesis from cotyledon explants of Fagonia indica Burm. J Med Plants By Prod 2:209–214
El-Sayed MM, Abdel-Hameed ES, El-Nahas HA, El-Wakil EA (2006) Isolation and identification of some steroidal glycosides of Fur-craea selloa. Pharmazie 61:478–482
Eskander J, Lavaud C, Harakat D (2010) Steroidal saponins from the leaves of Agave macroacantha. Fitoterapia 81:371–374
Eskander J, Lavaud C, Harakat D (2011) Steroidal saponins from the leaves of Beaucarnea recurvata. Phytochemistry 72:946–951
Fabris M, Matthijs M, Carbonelle S, Moses T, Pollier J, Dasseville R, Baart GJ, Vyverman W, Goossens A (2014) Tracking the sterol biosynthesis pathway of the diatom Phaeodactylum tricornutum. New Phytol 204:521–535
Folwarczna J, Zych M, Nowińska B, Pytlik M, Bialik M, Jagusiak A, Lipecka-Karcz M, Matysiak M (2016) Effect of diosgenin, a steroidal sapogenin, on the rat skeletal system. Acta Biochim Pol 63:287–295
Fouad MA, Mohamed KM, Kamel MS, Matsunami K, Otsuka H (2008) Cesdiurins I-III, steroidal saponins from Cestrum diurnum L. J Nat Med 62:168–173
Fukushima EO, Seki H, Sawai S, Suzuki M, Ohyama K, Saito K, Muranaka T (2013) Combinatorial biosynthesis of legume natu-ral and rare triterpenoids in engineered yeast. Plant Cell Physiol 54(5):740–749
Fuller S, Stephens JM (2015) Diosgenin, 4-hydroxyisoleucine, and fiber from fenugreek: mechanisms of actions and potential effects on metabolic syndrome. Adv Nutr 6:189–197
Gajdus J, Kaczyński Z, Kawiak A, Lojkowska E, Stefanowicz-Hajduk J, Ochocka JR, Stepnowski P (2014) Isolation and identification of cytotoxic compounds from the rhizomes of Paris quadrifolia L. Pharmacogn Mag 10:S324–S333
Galarraga E, Mitaine-Offer AC, Amaro-Luis JM, Miyamoto T, Tan-aka C, Pouységu L, Quideau S, Rojas LB, Lacaille-Dubois MA (2011) Steroidal saponins from the fruits of Cestrum ruizterani-anum. Nat Prod Commun 6:1825–1826
Gas-Pascual E, Berna A, Bach TJ, Schaller H (2014) Plant oxidos-qualene metabolism: cycloartenol synthase-dependent sterol biosynthesis in Nicotiana benthamiana. PLoS ONE 9:e109156
Gnoula C, Mégalizzi V, De Nève N, Sauvage S, Ribaucour F, Guissou P, Duez P, Dubois J, Ingrassia L, Lefranc F, Kiss R, Mijatovic T (2008) Balanitin-6 and -7: diosgenyl saponins isolated from Balanites aegyptiaca Del. display significant anti-tumor activity in vitro and in vivo. Int J Oncol 32:5–15
Gong Y, Liao Z, Chen M, Zuo K, Guo L, Tan Q, Huang Z, Kai G, Sun X, Tan F, Tang K (2005) Molecular cloning and characteriza-tion of a 1-deoxy-d-xylulose 5-phosphate reductoisomerase gene from Ginkgo biloba. DNA Seq 16:111–120
Gruenweller S, Schroeder E, Kesselmeier J (1990) Biological activities of furostanol saponins from Nicotiana tabacum. Phytochemistry 29:2485–2490
Guclu Ustundag O, Mazza G (2007) Saponins: properties, applications and processing. Crit Rev Food Sci Nutr 47:231–258
Guo Y, Liu YX, Kang LP, Zhang T, Yu HS, Zhao Y, Xiong CQ, Ma BP (2013) Two novel furostanol saponins from the tubers of Ophiopogon japonicus. J Asian Nat Prod Res 15:459–465
Guo J, Xu C, Xue R, Jiang W, Wu B, Huang C (2015) Cytotoxic activ-ities of chemical constituents from rhizomes of Anemarrhena asphodeloides and their analogues. Arch Pharm Res 38:598–603

538 Planta (2018) 248:519–544
1 3
Guo H, Li R, Liu S, Zhao N, Han S, Lu M, Liu X, Xia X (2016) Molec-ular characterization, expression, and regulation of Gynostemma pentaphyllum squalene epoxidase gene 1. Plant Physiol Biochem 109:230–239
Ha SH, Lee SW, Kim YM, Hwang YS (2001) Molecular characteriza-tion of Hmg2 gene encoding a 3-hydroxy-methylglutaryl-CoA reductase in rice. Mol Cells 11:295–302
Ha SH, Kim JB, Hwang YS, Lee SW (2003) Molecular characteriza-tion of three 3-hydroxy-3-methylglutaryl-CoA reductase genes including pathogen-induced Hmg2 from pepper (Capsicum annuum). Biochim Biophys Acta 1625:253–260
Han JY, In JG, Kwon YS, Choi YE (2010) Regulation of ginsenoside and phytosterol biosynthesis by RNA interferences of squalene epoxidase gene in Panax ginseng. Phytochemistry 71:36–46
Haralampidis K, Trojanowska M, Osbourn AE (2002) Biosynthesis of triterpenoid saponins in plants. Adv Biochem Eng Biotechnol 75:31–49
Hayes PY, Lambert LK, Lehmann R, Penman K, Kitching W, De Voss JJ (2007) Complete (1)H and (13)C assignments of the four major saponins from Dioscorea villosa (wild yam). Magn Reson Chem 45:1001–1005
Hayes PY, Jahidin AH, Lehmann R, Penman K, Kitching W, De Voss JJ (2008) Steroidal saponins from the roots of Asparagus rac-emosus. Phytochemistry 69:796–804
Hayes PY, Lehmann R, Penman K, Kitching W, De Voss JJ (2009) Steroidal saponins from the roots of Trillium erectum (Beth root). Phytochemistry 70:105–113
He H, Sun YP, Zheng L, Yue ZG (2015) Steroidal saponins from Paris polyphylla induce apoptotic cell death and autophagy in A549 human lung cancer cells. Asian Pac J Cancer Prev 16:1169–1173
Hong XX, Luo JG, Guo C, Kong LY (2012) New steroidal saponins from the bulbs of Lilium brownii var. viridulum. Carbohydr Res 361:19–26
Hostettmann K, Marston A (1995) Saponins. Cambridge University Press, Cambridge
Hostettmann K, Marston A (2005) Saponins, chemistry and pharmacol-ogy of natural products. Cambridge University Press, Cambridge
Huang X, Kong L (2006) Steroidal saponins from roots of Asparagus officinalis. Steroids 71:171–176
Huang W, Zou K (2015) Cytotoxicity of the saponin TTB2 on Ewing sarcoma cells. Exp Ther Med 10:625–628
Huang HL, Liu RH, Shao F (2009) Structural determination of two new steroidal saponins from Smilax china. Magn Reson Chem 47:741–745
Huang HC, Lin MK, Hwang SY, Hwang TL, Kuo YH, Chang CI, Ou CY, Kuo YH (2013) Two anti-inflammatory steroidal saponins from Dracaena angustifolia Roxb. Molecules 18:8752–8763
Ikenaga T, Oyama T, Muranaka T (1995) Growth and steroidal saponin production in hairy root cultures of Solanum aculeatissimum. Plant Cell Rep 14:413–417
Ikenaga T, Handayani R, Oyama T (2000) Steroidal saponin production in callus cultures of Solanum aculeatissimum Jacq. Plant Cell Rep 19:1240–1244
Inoue K, Shimomura K, Kobayashi S, Sankawa U, Ebizuka Y (1996) Conversion of furostanol glycoside to spirostanol glycoside by β-glucosidase in Costus speciosus. Phytochemistry 41:725–727
Ivanova A, Mikhova B, Klaiber I, Dinchev D, Kostova I (2009) Ste-roidal saponins from Smilax excelsa rhizomes. Nat Prod Res 23:916–924
Ivanova T, Dimitrova D, Gussev C, Bosseva Y, Stoeva T (2015) Ex situ conservation of Ruscus aculeatus L.—ruscogenin biosynthesis, genome-size stability and propagation traits of tissue-cultured clones. Biotechnol Biotechnol Equip 29:27–32
Jain AK, Vincent RM, Nessler CL (2000) Molecular characterization of a hydroxymethylglutaryl-CoA reductase gene from mulberry (Morus alba L.). Plant Mol Biol 42:559–569
Jayachandran KS, Vasanthi AH, Gurusamy N (2016) Steroidal sapo-nin diosgenin from Dioscorea bulbifera protects cardiac cells from hypoxia-reoxygenation Injury through modulation of pro-survival and pro-death molecules. Pharmacogn Mag 12:14–20
Jeena GS, Fatima S, Tripathi P, Upadhyay S, Shukla RK (2017) Comparative transcriptome analysis of shoot and root tissue of Bacopa monnieri identifies potential genes related to triter-penoid saponin biosynthesis. BMC Genom 18:490
Jiang D, Rong Q, Chen Y, Yuan Q, Shen Y, Guo J, Yang Y, Zha L, Wu H, Huang L, Liu C (2017) Molecular cloning and func-tional analysis of squalene synthase (SS) in Panax notogin-seng. Int J Biol Macromol 95:658–666
Jin H, Gong Y, Guo B, Qiu C, Liu D, Miao Z, Sun X, Tang K (2006) Isolation and characterization of a 2C-methyl-d-erythritol 2, 4-cyclodiphosphate synthase gene from Taxus media. Mol Biol (Mosk) 40:1013–1020
Jin YL, Kuk JH, Oh KT, Kim YJ, Piao XL, Park RD (2007) A new steroidal saponin, yuccalan, from the leaves of Yucca smal-liana. Arch Pharm Res 30:543–546
Jones RN, Katzenellenbogen E, Dobriner K (1953) Steroid metabo-lism. XVII. The infrared absorption spectra of the steroid sapo-genins. J Am Chem Soc 75:158–166
Kalita R, Patar L, Shasany AK, Modi MK, Sen P (2015) Molecular cloning, characterization and expression analysis of 3-hydroxy-3-methylglutaryl coenzyme A reductase gene from Centella asiatica L. Mol Biol Rep 42:1431–1439
Kalra S, Kumar S, Lakhanpal N, Kaur J, Singh K (2013) Charac-terization of Squalene synthase gene from Chlorophytum bori-vilianum (Sant. and Fernand.). Mol Biotechnol 54:944–953
Kaneda N, Nakanishi H, Staba EJ (1987) Steroidal constituents of Yucca shidigera plants and tissue cultures. Phytochemistry 26:1425–1429
Kang LP, Ma BP, Shi TJ, Zhang J, Xiong CQ (2006) Two new furo-stanol saponins from the rhizomes of Anemarrhena asphode-loides. Yao Xue Xue Bao 41:527–532
Kang YJ, Chung HJ, Nam JW, Park HJ, Seo EK, Kim YS, Lee D, Lee SK (2011) Cytotoxic and antineoplastic activity of timosaponin A-III for human colon cancer cells. J Nat Prod 74:701–706
Kang LP, Liu YX, Eichhorn T, Dapat E, Yu HS, Zhao Y, Xiong CQ, Liu C, Efferth T, Ma BP (2012a) Polyhydroxylated steroidal glycosides from Paris polyphylla. J Nat Prod 75:1201–1205
Kang LP, Wang YZ, Feng B, Huang HZ, Zhou WB, Zhao Y, Xiong CQ, Tan DW, Song XB, Ma BP (2012b) Structure elucida-tion and complete NMR spectral assignments of glucosylated saponins of cantalasaponin I. Magn Reson Chem 50:79–83
Kang LP, Zhang J, Cong Y, Li B, Xiong CQ, Zhao Y, Tan DW, Yu HS, Yu ZY, Cong YW, Liu C, Ma BP (2012c) Steroidal glyco-sides from the rhizomes of Anemarrhena asphodeloides and their antiplatelet aggregation activity. Planta Med 78:611–616
Kang LP, Wu KL, Yu HS, Pang X, Liu J, Han LF, Zhang J, Zhao Y, Xiong CQ, Song XB, Liu C, Cong YW, Ma BP (2014) Steroidal saponins from Tribulus terrestris. Phytochemistry 107:182–189
Kawabata T, Cui MY, Hasegawa T, Takano F, Ohta T (2011) Anti-inflammatory and anti-melanogenic steroidal saponin glycosides from Fenugreek (Trigonella foenum-graecum L.) seeds. Planta Med 77:705–710
Keim V, Manzano D, Fernández FJ, Closa M, Andrade P, Caudepón D, Bortolotti C, Vega MC, Arró M, Ferrer A (2012) Characteri-zation of Arabidopsis FPS isozymes and FPS gene expression analysis provide insight into the biosynthesis of isoprenoid pre-cursors in seeds. PLoS ONE 7:e49109
Kim SM, Kuzuyama T, Chang YJ, Kim SU (2006) Cloning and char-acterization of 2-C-methyl-d-erythritol 2,4-cyclodiphosphate synthase (MECS) gene from Ginkgo biloba. Plant Cell Rep 25:829–835

539Planta (2018) 248:519–544
1 3
Kim TD, Han JY, Huh GH, Choi YE (2011) Expression and func-tional characterization of three squalene synthase genes asso-ciated with saponin biosynthesis in Panax ginseng. Plant Cell Physiol 52:125–137
Kim CS, Kim SY, Moon E, Lee MK, Lee KR (2013) Steroidal con-stituents from the leaves of Hosta longipes and their inhibi-tory effects on nitric oxide production. Bioorg Med Chem Lett 15:1771–1775
Kim YJ, Lee OR, Oh JY, Jang MG, Yang DC (2014) Functional analy-sis of 3-hydroxy-3-methylglutaryl coenzyme A reductase encod-ing genes in triterpene saponin-producing ginseng. Plant Physiol 165:373–387
Kim JE, Go J, Koh EK, Song SH, Sung JE, Lee HA, Kim DS, Son HJ, Lee HS, Lee CY, Hong JT, Hwang DY (2016) Diosgenin effectively suppresses skin inflammation induced by phthalic anhydride in IL-4/Luc/CNS-1 transgenic mice. Biosci Biotech-nol Biochem 80:891–901
Kime B, Barminas JT, Nkafamiya II, Akinterinwa A (2015) Extrac-tion and evaluation of a saponin-based surfactant from Balanites aegyptiaca plant as an emulsifying agent. Int J Innov Sci Eng Technol 2:733–737
Kirmizibekmez H, Masullo M, Festa M, Capasso A, Piacente S (2014) Steroidal glycosides with antiproliferative activities from Digi-talis trojana. Phytother Res 28:534–538
Kitagawa I (2002) Licorice root. A natural sweetener and an important ingredient in Chinese medicine. Pure Appl Chem 74:1189–1198
Kochan E, Szymczyk P, Kuźma Ł, Lipert A, Szymańska G (2017) Yeast extract stimulates ginsenoside production in hairy root cul-tures of American ginseng cultivated in shake flasks and nutrient sprinkle bioreactors. Molecules 22(6):880
Kohara A, Nakajima C, Hashimoto K, Ikenaga T, Tanaka H, Shoyama Y, Yoshida S, Muranaka T (2005) A novel glucosyltransferase involved in steroid saponin biosynthesis in Solanum aculeatis-simum. Plant Mol Biol 57:225–239
Kornobis E, Cabellos L, Aguilar F, Frias-Lopez C, Rozas J, Marco J, Zardoya R (2015) TRUFA: a user-friendly web server for de novo RNA-seq analysis using cluster computing. Evol Bioinform 11:97–104
Kostadinova EP, Alipieva KI, Kokubun T, Taskova RM, Handjieva NV (2007) Phenylethanoids, iridoids and a spirostanol saponin from Veronica turrilliana. Phytochemistry 68:1321–1326
Kougan GB, Miyamoto T, Tanaka C, Paululat T, Mirjolet JF, Duchamp O, Sondengam BL, Lacaille-Dubois MA (2010) Steroidal sapo-nins from two species of Dracaena. J Nat Prod 73:1266–1270
Kowalczyk M, Pecio Ł, Stochmal A, Oleszek W (2011) Qualitative and quantitative analysis of steroidal saponins in crude extract and bark powder of Yucca schidigera Roezl. J Agric Food Chem 59:8058–8064
Kumar S, Kalra S, Singh B, Kumar A, Kaur J, Singh K (2016) RNA-Seq mediated root transcriptome analysis of Chlorophytum bori-vilianum for identification of genes involved in saponin biosyn-thesis. Funct Integr Genom 16:37–55
Kumari U, Vishwakarma RK, Sonawane P, Abbassi S, Khan BM (2015) Biochemical characterization of recombinant mevalonate kinase from Bacopa monniera. Int J Biol Macromol 72:776–783
Kuo CI, Chao CH, Lu MK (2012) Effects of auxins on the production of steroidal alkaloids in rapidly proliferating tissue and cell cul-tures of Solanum lyratum. Phytochem Anal 23:400–404
Kuroda M, Ori K, Mimaki Y (2006) Ornithosaponins A–D, four new polyoxygenated steroidal glycosides from the bulbs of Orni-thogalum thyrsoides. Steroids 71:199–205
Lai W, Wu Z, Lin H, Li T, Sun L, Chai Y, Chen W (2010) Anti-ischemia steroidal saponins from the seeds of Allium fistulosum. J Nat Prod 73:1053–1057
Lakshmi V, Kumar R, Pandey K, Joshi BS, Roy R, Madhusudanan KP, Tiwari P, Srivastava AK (2009) Structure and activities of
a steroidal saponin from Chlorophytum nimonii (Grah) Dalz. Nat Prod Res 23:963–972
Laranjeira S, Amorim-Silva V, Esteban A, Arró M, Ferrer A, Tavares RM, Botella MA, Rosado A, Azevedo H (2015) Arabidop-sis squalene epoxidase 3 (SQE3) complements SQE1 and is important for embryo development and bulk squalene epoxi-dase activity. Mol Plant 8:1090–1102
Lee B, Trinh HT, Jung K, Han SJ, Kim DH (2010) Inhibitory effects of steroidal timosaponins isolated from the rhizomes of Anemarrhena asphodeloides against passive cutaneous ana-phylaxis reaction and pruritus. Immunopharmacol Immuno-toxicol 32:357–363
Lee CL, Hwang TL, He WJ, Tsai YH, Yen CT, Yen HF, Chen CJ, Chang WY, Wu YC (2013) Anti-neutrophilic inflammatory steroidal glycosides from Solanum torvum. Phytochemistry 95:315–321
Li X, Sun H, Ye Y, Chen F, Pan Y (2006) C-21 steroidal glycosides from the roots of Cynanchum chekiangense and their immuno-suppressive activities. Steroids 71:61–66
Li H, Huang W, Wen Y, Gong G, Zhao Q, Yu G (2010) Anti-throm-botic activity and chemical characterization of steroidal sap-onins from Dioscorea zingiberensis CH Wright. Fitoterapia 81:1147–1156
Li N, Zhang L, Zeng KW, Zhou Y, Zhang JY, Che YY, Tu PF (2013) Cytotoxic steroidal saponins from Ophiopogon japonicus. Ster-oids 78:1–7
Li XJ, Wang L, Xue PF, Xie HX, Wei H, Wang J (2015a) New steroidal glycosides from Hosta plantaginea (Lam.) Aschers. J Asian Nat Prod Res 17:224–231
Li Y, Liu C, Xiao D, Han J, Yue Z, Sun Y, Fan L, Zhang F, Meng J, Zhang R, Wang Z, Mei Q, Wen A (2015b) Trillium tschonoskii steroidal saponins suppress the growth of colorectal cancer cells in vitro and in vivo. J Ethnopharmacol 168:136–145
Li Y, Wang X, He H, Zhang D, Jiang Y, Yang X, Wang F, Tang Z, Song X, Yue Z (2015c) Steroidal saponins from the roots and rhizomes of Tupistra chinensis. Molecules 20:13659–13669
Li JL, Gao ZB, Zhao WM (2016) Identification and evaluation of antie-pileptic activity of C21 steroidal glycosides from the roots of Cynanchum wilfordii. J Nat Prod 79:89–97
Liang M, Zheng Z, Yuan Y, Kong L, Shen Y, Liu R, Zhang C, Zhang W (2007) Identification and quantification of C21 steroidal saponins from Radix Cynanchi Atrati by high-performance liquid chroma-tography with evaporative light scattering detection and electro-spray mass spectrometric detection. Phytochem Anal 18:428–435
Liang Y, Jiang X, Hu Q, Li X, Yin H, Li D, Zhang Y, Liu X (2014) Cloning and characterization of 3-hydroxy-3-methylglutaryl-CoA reductase (HMGR) gene from Paris fargesii Franch. Indian J Biochem Biophys 51:201–206
Lin JT, Yang DJ (2008) Determination of steroidal saponins in differ-ent organs of yam (Dioscorea pseudojaponica Yamamoto). Food Chem 108:1068–1074
Lin S, Wang D, Yang D, Yao J, Tong Y, Chen J (2007) Characterization of steroidal saponins in crude extract from Dioscorea nipponica Makino by liquid chromatography tandem multi-stage mass spec-trometry. Anal Chim Acta 599:98–106
Lin T, Huang HL, Liu RH, Shu JC, Ren G, Shao F, Liu LS (2012) Steroidal saponins and pregnane glycosides from Smilax micro-phylla. Magn Reson Chem 50:813–817
Liu HY, Ni W, Hao XJ, Chen CX (2006) Steroidal saponins from Tacca plantaginea. J Asian Nat Prod Res 8:293–298
Liu T, Chen G, Yi GQ, Xu JK, Zhang TL, Hua HM, Pei YH (2010a) New pregnane and steroidal glycosides from Tribulus terrestris L. J Asian Nat Prod Res 12:209–214
Liu T, Lu X, Wu B, Chen G, Hua HM, Pei YH (2010b) Two new steroidal saponins from Tribulus terrestris L. J Asian Nat Prod Res 12:30–35

540 Planta (2018) 248:519–544
1 3
Liu H, Chou GX, Wang JM, Ji LL, Wang ZT (2011) Steroidal sapo-nins from the rhizomes of Dioscorea bulbifera and their cyto-toxic activity. Planta Med 77:845–848
Liu CX, Guo ZY, Xue YH, Zhang HY, Zhang HQ, Zou K, Huang NY (2012a) Tupisteroide A–C, three new polyhydroxylated ste-roidal constituents from the roots of Tupistra chinensis. Magn Reson Chem 50:320–324
Liu X, Zhang H, Niu XF, Xin W, Qi L (2012b) Steroidal saponins from Smilacina japonica. Fitoterapia 83:812–816
Liu QB, Peng Y, Li LZ, Gao PY, Sun Y, Yu LH, Song SJ (2013) Steroidal saponins from Anemarrhena asphodeloides. J Asian Nat Prod Res 15:891–898
Liu L, Zhao YL, Cheng GG, Chen YY, Qin XJ, Song CW, Yang XW, Liu YP, Luo XD (2014) Limonoid and steroidal saponin from Azadirachta indica. Nat Prod Bioprospect 4:335–340
Liu JY, Lu L, Kang LP, Liu YX, Zhao Y, Xiong CQ, Zhang YQ, Yu LY, Ma BP (2015a) Selective glycosylation of steroidal saponins by Arthrobacter nitroguajacolicus. Carbohydr Res 402:71–76
Liu Y, Ouyang Y, Wang ZQ, Qiao L, Li S, Zhao SH, Liu MY (2015b) A new C21 steroidal saponins from Periplocae Cortex. Zhong-guo Zhong Yao Za Zhi 40:455–457
Liu T, Li X, Xie S, Wang L, Yang S (2016a) RNA-seq analysis of Paris polyphylla var. yunnanensis roots identified candidate genes for saponin synthesis. Plant Divers 38:163–170
Liu Y, Tian X, Hua D, Cheng G, Wang K, Zhang L, Tang H, Wang M (2016b) New steroidal saponins from the rhizomes of Paris delavayi and their cytotoxicity. Fitoterapia 111:130–137
Liu M, Li LN, Pan YT, Kong JQ (2017) cDNA isolation and functional characterization of squalene synthase gene from Ornithogalum caudatum. Protein Expr Purif 130:63–72
Lorent JH, Quetin-Leclercq J, Mingeot-Leclercq MP (2014) The amphiphilic nature of saponins and their effects on artificial and biological membranes and potential consequences for red blood and cancer cells. Org Biomol Chem 12:8803–8822
Lu Y, Luo J, Huang X, Kong L (2009) Four new steroidal glycosides from Solanum torvum and their cytotoxic activities. Steroids 74:95–101
Lu Y, Chen CX, Ni W, Hua Y, Liu HY (2010) Spirostanol tetraglyco-sides from Ypsilandra thibetica. Steroids 75:982–987
Lu Y, Luo J, Kong L (2011) Steroidal alkaloid saponins and steroidal saponins from Solanum surattense. Phytochemistry 72:668–673
Luo Y, Shen HY, Zuo WJ, Wang H, Mei WL, Dai HF (2015) A new steroidal saponin from dragon’s blood of Dracaena cambodiana. J Asian Nat Prod Res 17:409–414
Ma S, Kou J, Yu B (2011) Safety evaluation of steroidal saponin DT-13 isolated from the tuber of Liriope muscari (Decne.) Baily. Food Chem Toxicol 49:2243–2251
Maldonado-Mendoza IE, Vincent RM, Nessler CL (1997) Molecular characterization of three differentially expressed members of the Camptotheca acuminata 3-hydroxy-3-methylglutaryl CoA reduc-tase (HMGR) gene family. Plant Mol Biol 34:781–790
Mandal D, Banerjee S, Mondal NB, Chakravarty AK, Sahu NP (2006) Steroidal saponins from the fruits of Asparagus racemosus. Phy-tochemistry 67:1316–1321
Marshall JG, Staba EJ (1976) Hormonal effects on diosgenin biosyn-thesis and growth in Dioscorea deltoidea tissue cultures. Phyto-chemistry 15:53–55
Masullo M, Pizza C, Piacente S (2016) Ruscus genus: a rich source of bioactive steroidal saponins. Planta Med 82:1513–1524
Matsuo Y, Watanabe K, Mimaki Y (2008) New steroidal glycosides from rhizomes of Clintonia udensis. Biosci Biotechnol Biochem 72:1714–1721
Moehs CP, Allen PV, Friedman M, Belknap WR (1997) Cloning and expression of solanidine UDP-glucose glucosyltransferase from potato. Plant J 11:227–236
Moharram FA, EI-Shenawy SM (2007) Antinociceptive and anti-inflammatory steroidal saponins from Dracaena ombet. Planta Med 73:1101–1106
Montes EG, Mitaine-Offer AC, Amaro-Luis JM, Paululat T, Delaude C, Pouységu L, Quideau S, Rojas LB, Delemasure S, Dutartre P, Lacaille-Dubois MA (2014) Acylated oleanane-type saponins from Ganophyllum giganteum. Phytochemistry 98:236–242
Morrissey JP, Osbourn AE (1999) Fungal resistance to plant antibiot-ics as a mechanism of pathogenesis. Microbiol Mol Biol Rev 63:708–724
Moses T, Papadopoulou KK, Osbourn A (2014a) Metabolic and func-tional diversity of saponins, biosynthetic intermediates and semi-synthetic derivatives. Crit Rev Biochem Mol Biol 49:439–462
Moses T, Pollier J, Almagro L, Buyst D, Van Montagu M, Pedreño MA, Martins JC, Thevelein JM, Goossens A (2014b) Combinato-rial biosynthesis of sapogenins and saponins in Saccharomyces cerevisiae using a C-16α hydroxylase from Bupleurum falcatum. Proc Natl Acad Sci USA 111(4):1634–1639
Mskhiladze L, Kutchukhidze J, Chincharadze D, Delmas F, Elias R, Favel A (2008a) In vitro antifungal and antileishmanial activi-ties of steroidal saponins from Allium leucanthum C. Koch—a Caucasian endemic species. Georgian Med News 154:39–43
Mskhiladze L, Legault J, Lavoie S, Mshvildadze V, Kuchukhidze J, Elias R, Pichette A (2008b) Cytotoxic steroidal saponins from the flowers of Allium leucanthum. Molecules 13:2925–2934
Munafo JP, Ramanathan A, Jimenez LS, Gianfagna TJ (2010) Isola-tion and structural determination of steroidal glycosides from the bulbs of Easter lily (Lilium longiflorum Thunb.). J Agric Food Chem 58:8806–8813
Nakamura S, Hongo M, Sugimoto S, Matsuda H, Yoshikawa M (2008) Steroidal saponins and pseudoalkaloid oligoglycoside from Bra-zilian natural medicine, “fruta do lobo” (fruit of Solanum lyco-carpum). Phytochemistry 69:1565–1572
Nakayasu M, Kawasaki T, Lee HJ, Sugimoto Y, Onjo M, Muranaka T, Mizutani M (2015) Identification of furostanol glycoside 26-O-β-glucosidase involved in steroidal saponin biosynthesis from Dioscorea esculenta. Plant Biotechnol 32:299–308
Napolitano A, Muzashvili T, Perrone A, Pizza C, Kemertelidze E, Piacente S (2010) Steroidal glycosides from Ruscus ponticus. Phytochemistry 72:651–661
Naveed MA, Riaz N, Saleem M, Jabeen B, Ashraf M, Ismail T, Jabbar A (2014) Longipetalosides A–C, new steroidal saponins from Tribulus longipetalus. Steroids 83:45–51
Nelson D (2006) Plant cytochrome P450 s from moss to poplar. Phy-tochem Rev 5:193–204
Nian H, Qin LP, Chen WS, Zhang QY, Zheng HC, Wang Y (2006) Protective effect of steroidal saponins from rhizome of Anemar-rhena asphodeloides on ovariectomy-induced bone loss in rats. Acta Pharmacol Sin 27:728–734
Nikam TD, Ebrahimi MA, Patil VA (2009) Embryogenic callus culture of Tribulus terrestris L. a potential source of harmaline, harmine and diosgenin. Plant Biotechnol Rep 3:243–250
Nohara T, Ono M, Ikeda T, Fujiwara Y, El-Aasr M (2010) The tomato saponin, esculeoside A. J Nat Prod 73:1734–1741
Nohara T, Fujiwara Y, Zhou JR, Urata J, Ikeda T, Murakami K, El-Aasr M, Ono M (2015) Saponins, esculeosides B-1 and B-2, in tomato juice and sapogenol, esculeogenin B1. Chem Pharm Bull (Tokyo) 63:848–850
Ohnishi T, Yokota T, Mizutani M (2009) Insights into the function and evolution of P450 s in plant steroid metabolism. Phytochemistry 70:1918–1929
Ohno M, Ono M, Nohara T (2011) New solanocapsine-type tomato glycoside from ripe fruit of Solanum lycopersicum. Chem Pharm Bull (Tokyo) 59:1403–1405
Ohtsuki T, Sato M, Koyano T, Kowithayakorn T, Kawahara N, Goda Y, Ishibashi M (2006) Steroidal saponins from Calamus

541Planta (2018) 248:519–544
1 3
insignis, and their cell growth and cell cycle inhibitory activi-ties. Bioorg Med Chem 14:659–665
Osbourn AE (1996) Preformed antimicrobial compounds and plant defense against fungal attack. Plant Cell 8:1821–1831
Osbourn AE (2003) Saponins in cereals. Phytochemistry 62:1–4Palazón J, Moyano E, Bonfill M, Osuna LT, Cusidó RM, Pinol
MT (2006) Effect of organogenesis on steroidal saponin bio-synthesis in calli cultures of Ruscus aculeatus. Fitoterapia 77:216–220
Pan X, Chen M, Liu Y, Wang Q, Zeng L, Li L, Liao Z (2008) A new isopentenyl diphosphate isomerase gene from Camptotheca acuminata: cloning, characterization and functional expression in Escherichia coli. DNA Seq 19:98–105
Pan ZH, Li Y, Liu JL, Ning DS, Li DP, Wu XD, Wen YX (2012) A cytotoxic cardenolide and a saponin from the rhizomes of Tupis-tra chinensis. Fitoterapia 83:1489–1493
Pang X, Wen D, Zhao Y, Xiong CQ, Wang XQ, Yu LY, Ma BP (2015) Steroidal saponins obtained by biotransformation of total furo-stanol glycosides from Dioscorea zingiberensis with Absidia coerulea. Carbohydr Res 402:236–240
Paquette SM, Bak S, Feyereisen R (2000) Intron–exon organization and phylogeny in a large superfamily, the paralogous cytochrome P450 genes of Arabidopsis thaliana. DNA Cell Biol 19:307–317
Pecio Ł, Jędrejek D, Masullo M, Piacente S, Oleszek W, Stochmal A (2012) Revised structures of avenacosides A and B and a new sulfated saponin from Avena sativa L. Magn Reson Chem 50:755–758
Peng YR, Li YB, Liu XD, Zhang JF, Duan JA (2008) Antitumor activ-ity of C-21 steroidal glycosides from Cynanchum auriculatum Royle ex Wight. Phytomedicine 15:1016–1020
Pereira BD, Valente AP, Parente JP (2004) A new steroidal saponin from Agave shrevei. Nat Prod Res 20:385–390
Pérez AJ, Calle JM, Simonet AM, Guerra JO, Stochmal A, Macías FA (2013) Bioactive steroidal saponins from Agave offoyana flowers. Phytochemistry 95:298–307
Pérez AJ, Simonet AM, Calle JM, Pecio Ł, Guerra JO, Stochmal A, Macías FA (2014) Phytotoxic steroidal saponins from Agave offoyana leaves. Phytochemistry 105:92–100
Perez-Labrada K, Brouard I, Estévez S, Marrero MT, Estévez F, Ber-mejo J, Rivera DG (2012) New insights into the structure–cyto-toxicity relationship of spirostan saponins and related glycosides. Bioorg Med Chem 20:2690–2700
Phillips MA, D’Auria JC, Gershenzon J, Pichersky E (2008) The Arabi-dopsis thaliana type I isopentenyl diphosphate isomerases are targeted to multiple subcellular compartments and have overlap-ping functions in isoprenoid biosynthesis. Plant Cell 20:677–696
Pise MV, Rudra JA, Upadhyay A (2015) Immunomodulatory potential of shatavarins produced from Asparagus racemosus tissue cul-tures. J Nat Sci Biol Med 6:415–420
Qin Y, Wu X, Huang W, Gong G, Li D, He Y, Zhao Y (2009) Acute toxicity and sub-chronic toxicity of steroidal saponins from Dioscorea zingiberensis C.H.Wright in rodents. J Ethnophar-macol 126:543–550
Qin XJ, Sun DJ, Ni W, Chen CX, Hua Y, He L, Liu HY (2012) Steroi-dal saponins with antimicrobial activity from stems and leaves of Paris polyphylla var. yunnanensis. Steroids 77:1242–1248
Qin XJ, Yu MY, Ni W, Yan H, Chen CX, Cheng YC, He L, Liu HY (2016) Steroidal saponins from stems and leaves of Paris poly-phylla var. yunnanensis. Phytochemistry 121:20–29
Qu Y, Zhang Y, Pei L, Wang Y, Gao L, Huang Q, Ojika M, Sak-agami Y, Qi J (2011) New neuritogenic steroidal saponin from Ophiopogon japonicus (Thunb.) Ker-Gawl. Biosci Biotechnol Biochem 75:1201–1204
Rahman SU, Ismail M, Khurram M, Haq IU (2015) Pharmacognostic and ethnomedicinal studies on Trillium govanianum. Pak J Bot 47:187–192
Ramalingam M, Kim SJ (2016) Pharmacological activities and appli-cations of spicatoside A. Biomol Ther (Seoul) 24:469–474
Ramos-Valdivia AC, van der Heijden R, Verpoorte R, Camara B (1997) Purification and characterization of two isoforms of isopentenyl-diphosphate isomerase from elicitor-treated Cin-chona robusta cells. Eur J Biochem 249:161–170
Razdan S, Bhat WW, Rana S, Dhar N, Lattoo SK, Dhar RS, Vish-wakarma RA (2013) Molecular characterization and promoter analysis of squalene epoxidase gene from Withania somnifera (L.) Dunal. Mol Biol Rep 40:905–916
Rezgui A, Mitaine-Offer AC, Pertuit D, Miyamoto T, Tanaka C, Delemasure S, Dutartre P, Lacaille-Dubois MA (2013) Steroi-dal saponins from Dracaena marginata. Nat Prods Commun 8:157–160
Rezgui A, Mitaine-Offer AC, Paululat T, Delemasure S, Dutartre P, Lacaille-Dubois MA (2014) Cytotoxic steroidal glycosides from Allium flavum. Fitoterapia 93:121–125
Rezgui A, Mitaine-Offer AC, Miyamoto T, Tanaka C, Lacaille-Dubois MA (2015) Spirostane-type saponins from Dracaena fragrans “Yellow Coast”. Nat Prod Commun 10:37–38
Ribeiro PR, Araújo AJ, Costa-Lotufo LV, Braz-Filho R, Nobre Jun-ior HV, da Silva CR, de Andrade Neto JB, Silveira ER, Lima MA (2016) Spirostanol glucosides from the leaves of Cestrum laevigatum L. Steroids 106:35–40
Rojas R, Alba J, Magaña-Plaza I, Cruz F, Ramos-Valdivia AC (1999) Stimulated production of diosgenin in Dioscorea galeottiana cell suspension cultures by abiotic and biotic factors. Biotech-nol Lett 21:907–911
Rong Q, Xu Q, Liu C, Huang L (2011) Cloning and characterization of 3-hydroxy-3-methylglutary CoA reductase cDNA of Glycyr-rhiza uralensis. Zhongguo Zhong Yao Za Zhi 36:1275–1279
Rong Q, Jiang D, Chen Y, Shen Y, Yuan Q, Lin H, Zha L, Zhang Y, Huang L (2016) Molecular cloning and functional analysis of squalene synthase 2 (SQS2) in Salvia miltiorrhiza Bunge. Front Plant Sci 7:1274
Sautour M, Miyamoto T, Lacaille-Dubois MA (2006) Bioactive ste-roidal saponins from Smilax medica. Planta Med 72:667–670
Sautour M, Miyamoto T, Lacaille-Dubois MA (2007) Steroi-dal saponins from Asparagus acutifolius. Phytochemistry 68:2554–2562
Sawai S, Saito K (2011) Triterpenoid biosynthesis and engineering in plants. Front Plant Sci 2:25
Schuler M (1996) Plant cytochrome P450 monooxygenases. Crit Rev Plant Sci 15:235–284
Schulte AE, van der Heijden R, Verpoorte R (2000) Purification and characterization of mevalonate kinase from suspension-cultured cells of Catharanthus roseus (L) G Don. Arch Biochem Biophys 378:287–298
Shao B, Guo H, Cui Y, Ye M, Han J, Guo D (2007) Steroidal saponins from Smilax china and their anti-inflammatory activities. Phyto-chemistry 68:623–630
Sharma U, Saini R, Kumar N, Singh B (2009) Steroidal saponins from Asparagus racemosus. Chem Pharm Bull (Tokyo) 57:890–893
Sharma U, Kumar N, Singh B (2012) Furostanol saponin and diphe-nylpentendiol from the roots of Asparagus racemosus. Nat Prod Commun 7:995–998
Sharma M, Ahuja A, Gupta R, Mallubhotla S (2015) Enhanced baco-side production in shoot cultures of Bacopa monnieri under the influence of abiotic elicitors. Nat Prod Res 29:745–749
Shen G, Pang Y, Wu W, Liao Z, Zhao L, Sun X, Tang K (2006) Cloning and characterization of a root-specific expressing gene encoding 3-hydroxy-3-methylglutaryl coenzyme A reductase from Ginkgo biloba. Mol Biol Rep 33:117–127
Shen S, Chen CJ, Bu R, Ga L, Li GY, Tan Y, Li X, Wang JH (2011) Three new steroidal saponins from Fritillaria pallidiflora. J Asian Nat Prod Res 13:1014–1022

542 Planta (2018) 248:519–544
1 3
Shen S, Li G, Huang J, Chen C, Ren B, Lu G, Tan Y, Zhang J, Li X, Wang J (2012) Steroidal saponins from Fritillaria pallidiflora Schrenk. Fitoterapia 83:785–794
Shen HY, Zuo WJ, Wang H, Zhao YX, Guo ZK, Luo Y, Li XN, Dai HF, Mei WL (2014) Steroidal saponins from dragon’s blood of Dracaena cambodiana. Fitoterapia 94:94–101
Shen T, Qiu F, Chen M, Lan XZ, Liao ZH (2015) Cloning and func-tional characterization of a cDNA encoding isopentenyl diphos-phate isomerase involved in taxols biosynthesis in Taxus media. Yao Xue Xue Bao 50:621–626
Shu W, Wu C, Zhang Y, Ye WC, Zhou G (2013) Two new steroidal gly-cosides isolated from the aerial part of Solanum torvum Swartz. Nat Prod Res 27:1982–1986
Shwe HH, Aye M, Sein MM, Htay KT, Kreitmeier P, Gertsch J, Reiser O, Heilmann J (2010) Cytotoxic steroidal saponins from the rhi-zomes of Tacca integrifolia. Chem Biodivers 7:610–622
Si YA, Yan H, Ni W, Liu ZH, Lu TX, Cheng CX, Liu HY (2014) Two new steroidal saponins from Ypsilandra thibetica. Nat Prod Bio-prospect 4:315–318
Singh P, Singh G, Bhandawat A, Singh G, Parmar R, Seth R, Sharma RK (2017) Spatial transcriptome analysis provides insights of key gene(s) involved in steroidal saponin biosynthesis in medici-nally important herb Trillium govanianum. Sci Rep 7:45295
Skhirtladze A, Perrone A, Montoro P, Benidze M, Kemertelidze E, Pizza C, Piacente S (2011) Steroidal saponins from Yucca glo-riosa L. rhizomes: LC–MS profiling, isolation and quantitative determination. Phytochemistry 72:126–135
Sobolewska D, Janeczko Z, Kisiel W, Podolak I, Galanty A, Trojanow-ska D (2006) Steroidal glycosides from the underground parts of Allium ursinum L. and their cytostatic and antimicrobial activity. Acta Pol Pharm 63:219–223
Sobolewska D, Michalska K, Podolak I, Grabowska K (2016) Steroidal saponins from the genus Allium. Phytochem Rev 15:1–35
Su L, Chen G, Feng SG, Wang W, Li ZF, Chen H, Liu YX, Pei YH (2009) Steroidal saponins from Tribulus terrestris. Steroids 74:399–403
Sun LX, Fu WW, Li W, Bi KS, Wang MW (2006) Diosgenin glucuro-nides from Solanum lyratum and their cytotoxicity against tumor cell lines. Z Naturforsch C 61:171–176
Sun Z, Huang X, Kong L (2010) A new steroidal saponin from the dried stems of Asparagus officinalis L. Fitoterapia 81:210–213
Sun K, Cao S, Pei L, Matsuura A, Xiang L, Qi J (2013) A steroidal saponin from Ophiopogon japonicus extends the lifespan of yeast via the pathway involved in SOD and UTH1. Int J Mol Sci 14:4461–4475
Sun CL, Ni W, Yan H, Liu ZH, Yang L, Si YA, Hua Y, Chen CX, He L, Zhao JH, Liu HY (2014) Steroidal saponins with induced platelet aggregation activity from the aerial parts of Paris verticillata. Steroids 92:90–95
Suzuki M, Kamide Y, Nagata N, Seki H, Ohyama K, Kato H, Masuda K, Sato S, Kato T, Tabata S, Yoshida S, Muranaka T (2004) Loss of function of 3-hydroxy-3-methylglutaryl coenzyme A reductase 1 (HMG1) in Arabidopsis leads to dwarfing, early senescence and male sterility, and reduced sterol levels. Plant J 37:750–761
Tabopda TK, Mitaine-Offer AC, Tanaka C, Miyamoto T, Mirjolet JF, Duchamp O, Ngadjui BT, Lacaille-Dubois MA (2014) Steroidal saponins from Dioscorea preussii. Fitoterapia 97:198–203
Tabopda TK, Mitaine-Offer AC, Paululat T, Delemasure S, Dutartre P, Ngadjui BT, Lacaille-Dubois MA (2016) Steroidal saponins from Chlorophytum deistelianum. Phytochemistry 126:34–40
Tan DW, Kang LP, Lü N, Zhang J, Ma BP (2006) Study on steroi-dal saponins of Dioscorea septemloba thumb. Zhong Yao Cai 29:1176–1179
Tang W, Newton RJ (2006) Mevalonate kinase activity during different stages of plant regeneration from nodular callus cultures in white pine (Pinus strobus). Tree Physiol 26:195–200
Tapondjou LA, Ponou KB, Teponno RB, Mbiantcha M, Djoukeng JD, Nguelefack TB, Watcho P, Cadenas AG, Park HJ (2008) In vivo anti-inflammatory effect of a new steroidal saponin, mannioside A, and its derivatives isolated from Dracaena mannii. Arch Pharm Res 31:653–658
Tapondjou LA, Jenett-Siems K, Böttger S, Melzig MF (2013) Steroidal saponins from the flowers of Dioscorea bulbifera var. sativa. Phytochemistry 95:341–350
Tapondjou LA, Siems KJ, Böttger S, Melzig MF (2015) Steroidal saponins from the mesocarp of the fruits of Raphia farinifera (Arecaceae) and their cytotoxic activity. Nat Prod Commun 10:1941–1944
Temraz AE, Gindi OD, Kadry HA, De Tommasi N, Braca A (2006) Steroidal saponins from the aerial parts of Tribulus alatus Del. Phytochemistry 67:1011–1018
Timité G, Mitaine-Offer AC, Miyamoto T, Tanaka C, Mirjolet JF, Duchamp O, Lacaille-Dubois MA (2013) Structure and cyto-toxicity of steroidal glycosides from Allium schoenoprasum. Phytochemistry 88:61–66
Tiski I, Marraccini P, Pot D, Vieira LG, Pereira LF (2011) Characteri-zation and expression of two cDNA encoding 3-hydroxy-3-meth-ylglutaryl coenzyme A reductase isoforms in coffee (Coffea ara-bica L.). OMICS 15:719–727
Tong QY, He Y, Zhao QB, Qing Y, Huang W, Wu XH (2012) Cyto-toxicity and apoptosis-inducing effect of steroidal saponins from Dioscorea zingiberensis Wright against cancer cells. Steroids 77:1219–1227
Tong YR, Su P, Zhao YJ, Zhang M, Wang XJ, Hu TY, Gao W, Huang LQ (2015) Cloning and expression analysis of 4- (cytidine-5-di-phospho) -2-C-methyl-D-erythritol kinase gene in Tripterygium wilfordii. Zhongguo Zhong Yao Za Zhi 40:4165–4170
Tong Y, Zhang M, Su P, Zhao Y, Wang X, Zhang X, Gao W, Huang L (2016) Cloning and functional characterization of an isopen-tenyl diphosphate isomerase gene from Tripterygium wilfordii. Biotechnol Appl Biochem 63:863–869
Uchida H, Sugiyama R, Nakayachi O, Takemura M, Ohyama K (2007) Expression of the gene for sterol-biosynthesis enzyme squalene epoxidase in parenchyma cells of the oil plant, Euphorbia tiru-calli. Planta 226:1109–1115
Upadhyay S, Phukan UJ, Mishra S, Shukla RK (2014) De novo leaf and root transcriptome analysis identified novel genes involved in steroidal sapogenin biosynthesis in Asparagus racemosus. BMC Genom 15:746
Vieira Júnior GM, da Rocha CQ, de Souza Rodrigues T, Hiruma-Lima CA, Vilegas W (2015) New steroidal saponins and antiulcer activity from Solanum paniculatum L. Food Chem 186:160–167
Vincken JP, Heng L, de Groot A, Gruppen H (2007) Saponins, clas-sification and occurrence in the plant kingdom. Phytochemistry 68:275–297
Vishwakarma RK, Patel KA, Sonawane P, Singh S, Ruby Kumari U, Agrawal DC, Khan BM (2012) Molecular characterization of farnesyl pyrophosphate synthase from Bacopa monniera by comparative modeling and docking studies. Bioinformation 8:1075–1081
Waheed A, Barker J, Barton SJ, Owen CP, Ahmed S, Carew MA (2012) A novel steroidal saponin glycoside from Fagonia indica induces cell-selective apoptosis or necrosis in cancer cells. Eur J Pharm Sci 47:464–473
Wang Y, McAllister TA (2010) A modified spectrophotometric assay to estimate deglycosylation of steroidal saponin to sapogenin by mixed ruminal microbes. J Sci Food Agric 90:1811–1818
Wang W, Meng H (2015) Cytotoxic, anti-inflammatory and hemostatic spirostane-steroidal saponins from the ethanol extract of the roots of Bletilla striata. Fitoterapia 101:12–18
Wang Y, Guo B, Zhang F, Yao H, Miao Z, Tang K (2007) Molec-ular cloning and functional analysis of the gene encoding

543Planta (2018) 248:519–544
1 3
3-hydroxy-3-methylglutaryl coenzyme A reductase from hazel (Corylus avellana L. Gasaway). J Biochem Mol Biol 40:861–869
Wang D, Li D, Zhu W, Zhang J, Peng P (2009a) Steroidal saponins from the rhizomes of Polygonatum odoratum. Nat Prod Res 23:940–947
Wang Y, Qiu C, Zhang F, Guo B, Miao Z, Sun X, Tang K (2009b) Molecular cloning, expression profiling and functional analyses of a cDNA encoding isopentenyl diphosphate isomerase from Gossypium barbadense. Biosci Rep 29:111–119
Wang YZ, Feng B, Huang HZ, Kang LP, Cong Y, Zhou WB, Zou P, Cong YW, Song XB, Ma BP (2010) Glucosylation of ste-roidal saponins by cyclodextrin glucanotransferase. Planta Med 76:1724–1731
Wang K, Sasaki T, Li W, Li Q, Wang Y, Asada Y, Kato H, Koike K (2011a) Two novel steroidal alkaloid glycosides from the seeds of Lycium barbarum. Chem Biodivers 8:2277–2284
Wang KW, Zhang H, Shen LQ, Wang W (2011b) Novel steroidal sapo-nins from Liriope graminifolia (Linn.) Baker with anti-tumor activities. Carbohydr Res 346:253–258
Wang H, Zhai Z, Li N, Jin H, Chen J, Yuan S, Wang L, Zhang J, Li Y, Yun J, Fan J, Yi J, Ling R (2013) Steroidal saponin of Tril-lium tschonoskii reverses multidrug resistance of hepatocellular carcinoma. Phytomedicine 20:985–991
Wang JJ, Liu YX, Wen D, Yu HS, Kang LP, Pang X, Ma BP, Chen YD (2014a) Study on steroidal saponins from Dioscorea zingiber-ensis and their platelet aggregation activities. Zhongguo Zhong Yao Za Zhi 39:3782–3787
Wang QJ, Zheng LP, Zhao PF, Zhao YL, Wang JW (2014b) Clon-ing and characterization of an elicitor-responsive gene encod-ing 3-hydroxy-3-methylglutaryl coenzyme A reductase involved in 20-hydroxyecdysone production in cell cultures of Cyanotis arachnoidea. Plant Physiol Biochem 84:1–9
Wang Y, Li X, Sun H, Yi K, Zheng J, Zhang J, Hao Z (2014c) Biotrans-formation of steroidal saponins in sisal (Agave sisalana Perrine) to tigogenin by a newly isolated strain from a karst area of Guilin, China. Biotechnol Biotechnol Equip 28:1024–1033
Wang P, Wei Y, Fan Y, Liu Q, Wei W, Yang C, Zhang L, Zhao G, Yue J, Yan X, Zhou Z (2015a) Production of bioactive ginsenosides Rh2 and Rg3 by metabolically engineered yeasts. Metab Eng 29:97–105
Wang X, Chen D, Wang Y, Xie J (2015b) De novo transcriptome assembly and the putative biosynthetic pathway of steroidal sapogenins of Dioscorea composita. PLoS ONE 10:e0124560
Wang YH, Niu HM, Zhang ZY, Hu XY, Li H (2015c) Medicinal values and their chemical bases of Paris. Zhongguo Zhong Yao Za Zhi 40:833–839
Wang Y, Xu L, Lou LL, Song SJ, Yao GD, Ge MY, Hayashi T, Tashiro SI, Onodera S, Ikejima T (2016a) Timosaponin AIII induces apoptosis and autophagy in human melanoma A375-S2 cells. Arch Pharm Res 40:69–78
Wang ZF, Wang BB, Zhao Y, Wang FX, Sun Y, Guo RJ, Song XB, Xin HL, Sun XG (2016b) Furostanol and spirostanol saponins from Tribulus terrestris. Molecules 21:E429
Williams JR, Gong H (2007) Biological activities and syntheses of ste-roidal saponins: the shark-repelling pavoninins. Lipids 42:77–86
Wu JJ, Cheng KW, Zuo XF, Wang MF, Li P, Zhang LY, Wang H, Ye WC (2010) Steroidal saponins and ecdysterone from Asparagus filicinus and their cytotoxic activities. Steroids 75:734–739
Wu X, Wang L, Wang GC, Wang H, Dai Y, Ye WC, Li YL (2012a) New steroidal saponins and sterol glycosides from Paris poly-phylla var. yunnanensis. Planta Med 78:1667–1675
Wu X, Wang L, Wang H, Dai Y, Ye WC, Li YL (2012b) Steroidal saponins from Paris polyphylla var. yunnanensis. Phytochem-istry 81:133–143
Wu XH, Wang CZ, Zhang J, Wang SQ, Han L, Zhang YW, Yuan CS (2014a) Effects of smilaxchinoside A and smilaxchinoside C,
two steroidal glycosides from Smilax riparia, on hyperurice-mia in a mouse model. Phytother Res 28:1822–1828
Wu XH, Zhang J, Wang SQ, Yang VC, Anderson S, Zhang YW (2014b) Riparoside B and timosaponin J, two steroidal gly-cosides from Smilax riparia, resist to hyperuricemia based on URAT1 in hyperuricemic mice. Phytomedicine 21:1196–1201
Xiao X, Zou J, Bui-Nguyen TM, Bai P, Gao L, Liu J, Liu S, Xiao J, Chen X, Zhang X, Wang H (2012) Paris saponin II of Rhizoma Paridis—a novel inducer of apoptosis in human ovarian cancer cells. Biosci Trends 6:201–211
Xie BB, Liu HY, Ni W, Chen CX, Lü Y, Wu L, Zheng QT (2006) Five new steroidal compounds from Ypsilandra thibetica. Chem Biodivers 3:1211–1218
Xie BB, Liu HY, Ni W, Chen CX (2009) Ypsilandrosides C–G, five new spirostanol saponins from Ypsilandra thibetica. Steroids 74:950–955
Xie WL, Jiang R, Shen XL, Chen ZY, Deng XM (2015) Diosgenin attenuates hepatic stellate cell activation through transforming growth factor-β/Smad signaling pathway. Int J Clin Exp Med 8:20323–20329
Xing Z, Long Y, Lao F, He S, Liang N, Li B (2012) Effect of endo-phytic fungi on expression amount of key enzyme genes in saponins biosynthesis and Eleutherococcus senticosus sapo-nins content. Zhongguo Zhong Yao Za Zhi 37:2041–2045
Xu DP, Hu CY, Wei L, Pang ZJ (2007a) Isolation and structure deter-mination of steroidal saponin from Dioscorea zingiberensis. Yao Xue Xue Bao 42:1162–1165
Xu FQ, Zhong HM, Liu HY, Liu HQ, Chen CX (2007b) Steroidal saponins from Lysimachia Paridiformis. J Asian Nat Prod Res 9:493–497
Xu TH, Xu YJ, Xie SX, Zhao HF, Han D, Li Y, Niu JZ, Xu DM (2008) A novel steroidal glycoside, ophiofurospiside A from Ophiopogon japonicus (Thunb.) Ker-Gawl. J Asian Nat Prod Res 10:415–418
Xu DP, Hu CY, Wang L, Wang XC, Pang ZJ (2009a) Isolation and structure determination of steroidal saponin from Dioscorea zingiberensis. Yao Xue Xue Bao 44:56–59
Xu DP, Hu CY, Zhang Y (2009b) Two new steroidal saponins from the rhizome of Polygonatum sibiricum. J Asian Nat Prod Res 11:1–6
Xu YJ, Xu TH, Liu Y, Xie SX, Si YS, Xu DM (2009c) Two new ste-roidal glucosides from Tribulus terrestris L. J Asian Nat Prod Res 11:548–553
Xu M, Zhang YJ, Li XC, Jacob MR, Yang CR (2010) Steroidal sap-onins from fresh stems of Dracaena angustifolia. J Nat Prod 73:1524–1528
Xu Y, Liu J, Liang L, Yang X, Zhang Z, Gao Z, Sui C, Wei J (2014) Molecular cloning and characterization of three cDNAs encoding 1-deoxy-d-xylulose-5-phosphate synthase in Aquilaria sinensis (Lour.) Gilg. Plant Physiol Biochem 82:133–141
Yada H, Kimura T, Suzuki M, Ohnishi-Kameyama M, Shinmoto H (2010) New steroidal saponin from Hosta sieboldiana. Biosci Biotechnol Biochem 74:861–864
Yan Y, Zhang JX, Liu KX, Huang T, Yan C, Huang LJ, Liu S, Mu SZ, Hao XJ (2014) Seco-pregnane steroidal glycosides from the roots of Cynanchum atratum and their anti-TMV activity. Fitoterapia 97:50–63
Yang QX, Yang CR (2006) Cytotoxic steroidal saponins from Polygo-natum punctatum. Chem Biodivers 3:1349–1355
Yang SL, Liu XK, Wu H, Wang HB, Qing C (2009) Steroidal saponins and cytoxicity of the wild edible vegetable—Smilacina atropur-purea. Steroids 74:7–12
Yang J, Wang P, Wu W, Zhao Y, Idehen E, Sang S (2016) Steroidal saponins in oat bran. J Agric Food Chem 64:1549–1556
Ye Y, Chen F, Sun H, Li X, Xu S (2008) Stemucronatoside K, a novel C(21) steroidal glycoside from Stephanotis mucronata, inhibited

544 Planta (2018) 248:519–544
1 3
the cellular and humoral immune response in mice. Int Immu-nopharmacol 8:1231–1238
Ye Y, Qu Y, Tang R, Cao S, Yang W, Xiang L, Qi J (2013) Three new neuritogenic steroidal saponins from Ophiopogon japonicus (Thunb.) Ker-Gawl. Steroids 78:1171–1176
Yen CT, Lee CL, Chang FR, Hwang TL, Yen HF, Chen CJ, Chen SL, Wu YC (2012) Indiosides G–K: steroidal glycosides with cyto-toxic and anti-inflammatory activities from Solanum violaceum. J Nat Prod 75:636–643
Yen PH, Chi VT, Kiem PV, Tai BH, Quang TH, Nhiem NX, Anh HL, Ban NK, Thanh BV, Minh CV, Park S, Kim SH (2016) Spiro-stanol saponins from Tacca vietnamensis and their anti-inflamma-tory activity. Bioorg Med Chem Lett S0960–894:30533–30539
Yin HX, Xue D, Bai N, Chen C, Chen Y, Zhang H (2008) Isolation and identification of major steroidal saponins of Paris polyphylla Smith var stenophylla Franch. Sichuan Da Xue Xue Bao Yi Xue Ban 39:485–488
Yokosuka A, Mimaki Y (2009) Steroidal saponins from the whole plants of Agave utahensis and their cytotoxic activity. Phyto-chemistry 70:807–815
Yokosuka A, Jitsuno M, Yui S, Yamazaki M, Mimaki Y (2009) Steroi-dal glycosides from Agave utahensis and their cytotoxic activity. J Nat Prod 72:1399–1404
Yoon KD, Chin YW, Yang MH, Choi J, Kim J (2012) Application of high-speed countercurrent chromatography–evaporative light scattering detection for the separation of seven steroidal saponins from Dioscorea villosa. Phytochem Anal 23:462–468
Yu JQ, Deng AJ, Qin HL (2013) Nine new steroidal glycosides from the roots of Cynanchum stauntonii. Steroids 78:79–90
Yuan L, Ji TF, Li CJ, Wang AG, Yang JB, Su YL (2009) Two new steroidal saponins from the seeds of Allium cepa L. J Asian Nat Prod Res 11:213–218
Yuan JC, Zhang J, Wang FX, Pang X, Zhao Y, Xiong CQ, Ma BP (2014) New steroidal glycosides from the rhizome of Anemar-rhena asphodeloides. J Asian Nat Prod Res 16:901–909
Yuan YL, Guo CR, Cui LL, Ruan SX, Zhang CF, Ji D, Yang ZL, Li F (2015) Timosaponin B-II ameliorates diabetic nephropathy via TXNIP, mTOR, and NF-κB signaling pathways in alloxan-induced mice. Drug Des Devel Ther 9:6247–6258
Zeng KW, Song FJ, Li N, Dong X, Jiang Y, Tu PF (2015) ASC, a bioac-tive steroidal saponin from Ophitopogin japonicas, inhibits angi-ogenesis through interruption of Src tyrosine kinase-dependent matrix metalloproteinase pathway. Basic Clin Pharmacol Toxicol 116:115–123
Zha L, Liu S, Su P, Yuan Y, Huang L (2016) Cloning, prokaryotic expression and functional analysis of squalene synthase (SQS) in Magnolia officinalis. Protein Expr Purif 120:28–34
Zhang Y, Li HZ, Zhang YJ, Jacob MR, Khan SI, Li XC, Yang CR (2006) Atropurosides A–G, new steroidal saponins from Smi-lacina atropurpurea. Steroids 71:712–719
Zhang Y, Zhang YJ, Jacob MR, Li XC, Yang CR (2008) Steroidal saponins from the stem of Yucca elephantipes. Phytochemistry 69:264–270
Zhang L, Liu JY, Xu LZ, Yang SL (2009a) Chantriolide C, a new withanolide glucoside and a new spirostanol saponin from the rhizomes of Tacca chantrieri. Chem Pharm Bull (Tokyo) 57:1126–1128
Zhang T, Liu H, Liu XT, Chen XQ, Wang Q (2009b) Steroidal saponins from the rhizomes of Paris delavayi. Steroids 74:809–813
Zhang LJ, Yu HS, Kang LP, Feng B, Quan B, Song XB, Ma BP, Kang TG (2012a) Two new steroidal saponins from the biotransforma-tion product of the rhizomes of Dioscorea nipponica. J Asian Nat Prod Res 14:640–646
Zhang T, Kang LP, Yu HS, Liu YX, Zhao Y, Xiong CQ, Zhang J, Zou P, Song XB, Liu C, Ma BP (2012b) Steroidal saponins from the tuber of Ophiopogon japonicus. Steroids 77:1298–1305
Zhang C, Feng S, Zhang L, Ren Z (2013a) A new cytotoxic steroidal saponin from the rhizomes and roots of Smilax scobinicaulis. Nat Prod Res 27:1255–1260
Zhang J, Li H, Li H, Lu Z, Shi J, Xu Z (2013b) Screening and condi-tion optimization of a strain for efficiently biotransformation of saponins in Dioscorea zingiberensis into diosgenin. Sheng Wu Gong Cheng Xue Bao 29:848–852
Zhang R, Huang B, Du D, Guo X, Xin G, Xing Z, Liang Y, Chen Y, Chen Q, He Y, Huang W (2013c) Anti-thrombosis effect of diosgenyl saponins in vitro and in vivo. Steroids 78:1064–1070
Zhang H, Chen L, Kou JP, Zhu DN, Qi J, Yu BY (2014a) Steroidal sapogenins and glycosides from the fibrous roots of Polygonatum odoratum with inhibitory effect on tissue factor (TF) procoagu-lant activity. Steroids 89:1–10
Zhang L, Wang JT, Zhang DW, Zhang G, Guo SX (2014b) Molecular characterization of a HMG-CoA reductase gene from a rare and endangered medicinal plant, Dendrobium officinale. Yao Xue Xue Bao 49:411–418
Zhang H, Su YF, Yang FY (2016a) Three new steroidal saponins from Helleborus thibetanus. Nat Prod Res 30:1724–1730
Zhang Y, Yang CR, Zhang YJ (2016b) Steroidal saponins from the rhi-zomes of Polygonatum prattii. J Asian Nat Prod Res 18:268–273
Zhao Y, Kang LP, Liu YX, Zhao Y, Xiong CQ, Ma BP, Dong FT (2007) Three new steroidal saponins from the rhizome of Paris polyphylla. Magn Reson Chem 45:739–744
Zhao Y, Kang LP, Liu YX, Liang YG, Tan DW, Yu ZY, Cong YW, Ma BP (2009) Steroidal saponins from the rhizome of Paris polyphylla and their cytotoxic activities. Planta Med 75:356–363
Zhao YJ, Chen X, Zhang M, Su P, Liu YJ, Tong YR, Wang XJ, Huang LQ, Gao W (2015) Molecular cloning and characterisation of farnesyl diphosphate synthase from Tripterygium wilfordii. PLoS ONE 10:e0125415
Zhao H, Tang Q, Mo C, Bai L, Tu D, Ma X (2017) Cloning and char-acterization of squalene synthase and cycloartenol synthase from Siraitia grosvenorii. Acta Pharm Sin B 7:215–222
Zheng J, Zheng Y, Zhi H, Dai Y, Wang N, Wu L, Fan M, Fang Y, Zhao S, Zhang K (2013) Two new steroidal saponins from Selaginella uncinata (Desv.) spring and their protective effect against anoxia. Fitoterapia 88:25–30
Zheng L, Zhou Y, Zhang JY, Song M, Yuan Y, Xiao YJ, Xiang T (2014) Two new steroidal saponins from the rhizomes of Dioscorea zingiberensis. Chin J Nat Med 12:142–147
Zhou LB, Chen DF (2008) Steroidal saponins from the roots of Aspara-gus filicinus. Steroids 73:83–87
Zhou X, He X, Wang G, Gao H, Zhou G, Ye W, Yao X (2006) Steroidal saponins from Solanum nigrum. J Nat Prod 69:1158–1163
Zhou LB, Chen TH, Bastow KF, Shibano M, Lee KH, Chen DF (2007) Filiasparosides A–D, cytotoxic steroidal saponins from the roots of Asparagus filicinus. J Nat Prod 70:1263–1267
Zhou ZL, Feng ZC, Fu CY, Zhang HL, Xia JM (2012) Steroidal and phenolic glycosides from the bulbs of Lilium pumilum DC and their potential Na +/K + ATPase inhibitory activity. Molecules 17:10494–10502
Zhu YL, Huang W, Ni JR, Liu W, Li H (2010) Production of dios-genin from Dioscorea zingiberensis tubers through enzymatic saccharification and microbial transformation. Appl Microbiol Biotechnol 85:1409–1416
Zhu L, Tan J, Wang B, Guan L, Liu Y, Zheng C (2011) In-vitro anti-tumor activity and antifungal activity of pennogenin steroidal saponins from Paris Polyphylla var. yunnanensis. Iran J Pharm Res 10:279–286
Zhu GL, Hao Q, Li RT, Li HZ (2014) Steroidal saponins from the roots of Asparagus cochinchinensis. Chin J Nat Med 12:213–217
Zou K, Wang J, Du M, Li Q, Tu G (2006) A pair of diastereoisomeric steroidal saponins from cytotoxic extracts of Tupistra chinensis rhizomes. Chem Pharm Bull (Tokyo) 54:1440–1442

























![? HN]ä[N5gT[ ²IO[IO weÁvb wefvM (evsjv)](https://img.pdfslide.us/doc/110x75/6217f5ba2a2121694f6271e0/-hnn5gt-ioio-wevb-wefvm-evsjv.jpg)



