Embed Size (px)
Citation preview

L E T T E RRealistic losses of rare species disproportionately impact higher
trophic levels
Matthew E. S. Bracken1* and
Natalie H. N. Low2
1Marine Science Center,
Northeastern University, 430 Nahant
Road, Nahant, MA 01908, USA2Department of Ecology and
Evolutionary Biology, Brown
University, 80 Waterman Street,
Providence, RI 02912, USA
*Correspondence: E-mail:
AbstractPredicting the consequences of changes in biodiversity requires understanding both species� susceptibility to
extirpation and their functional roles in ecosystems. However, few studies have evaluated the effects of realistic,
non-random biodiversity losses, severely limiting the applicability of biodiversity research to conservation.
Here, we removed sessile species from a rocky shore community in a way that deliberately mimicked natural
patterns of species loss. We found that the rarest species in the system act from the bottom up to
disproportionately impact the diversity and abundance of consumers. Realistic losses of rare species in a diverse
assemblage of seaweeds and sessile invertebrates, collectively comprising <10% of sessile biomass, resulted in a
42–47% decline in consumer biomass. In contrast, removal of an equivalent biomass of dominant sessile
species had no effect on consumers. Our results highlight the �cornerstone� role that rare species can play in
shaping the structure of the community they support.
KeywordsBiodiversity, consumer, cornerstone species, herbivory, rare species, rocky intertidal, seaweed.
Ecology Letters (2012) 15: 461–467
INTRODUCTION
Given global threats to biodiversity (Butchart et al. 2010; Barnosky
et al. 2011), understanding the consequences of biodiversity loss is of
utmost importance. However, the majority of research into the
functional consequences of diversity change is based on biodiversity
gradients constructed of randomly assembled communities of
organisms. Whereas this approach has contributed a great deal to
our understanding of the mechanisms linking biodiversity to
community and ecosystem properties (Hooper et al. 2005; Cardinale
et al. 2006), it is limited in its applicability to natural systems, where
changes in the number and relative abundance of species are not
random (Hutchinson 1959; Menge & Sutherland 1987). Increasing
attention is being paid to realistic species-loss scenarios (e.g. Smith &
Knapp 2003; Solan et al. 2004; Zavaleta & Hulvey 2004), and recent
work has highlighted striking differences in the effects of random vs.
realistic species losses (Bracken et al. 2008).
Progress in understanding how a species� susceptibility to loss is
related to its functional role in a community or ecosystem is essential if
we are to make biodiversity research relevant to conservation (Schwartz
et al. 2000; Srivastava & Vellend 2005). Accomplishing this will require
focusing simultaneously on both the causes and the consequences of
biodiversity change. Here, we link realistic changes in biodiversity to
community structure and species interactions by using natural variation
in diversity at the base of a marine food web to evaluate (1) the
relationships between sessile diversity and consumer diversity and
abundance and (2) the effects of realistic declines in sessile diversity on
higher trophic levels. In our initial surveys of a coastal rocky shore
community, we found that patterns of sessile richness occurred in a
nested order: a few common dominant species were present in all
surveyed plots, whereas several rare species – up to eight additional
species in the most diverse plots, collectively comprising <10% of
sessile biomass – were present only in high-diversity plots.
Both the abundance and diversity of consumers were higher in plots
containing more sessile species (see Results), and we hypothesised that
the rare seaweeds and sessile invertebrates that were uniquely present
in more diverse plots were facilitating those consumers. Species at the
base of a food web often provide both food and habitat for organisms
at higher trophic levels (Pimm 1980; Hunter & Price 1992; Knops
et al. 1999). However, previous work has largely focused on how
sessile species� roles are associated with their abundance (Lindeman
1942; Hutchinson 1959; Oksanen et al. 1981; Hairston & Hairston
1993). For example, dominant, foundation species (sensu Dayton
1972) enhance the diversity and abundance of organisms by providing
physical structure (Ellison et al. 2005; Bracken et al. 2007), and high
biomass of organisms at the base of a food web is typically required to
ensure sufficient energy flow to consumers (Elton 1927; Lindeman
1942; Paine 1980). However, few studies have empirically assessed the
effects of the loss of rare species from the base of food webs (but see
Marsh et al. 2000; Lyons & Schwartz 2001; Zavaleta & Hulvey 2004),
although these rare species often constitute the majority of species
richness in communities (Gotelli & Graves 1996; Novotny & Basset
2000).
We tested our hypothesis that realistic declines in rare species can
impact higher trophic levels by experimentally reducing the number of
rare seaweed and sessile invertebrate species on rocky intertidal reefs
so that richness and composition of experimental removal plots
mimicked values found in naturally occurring low-richness control
plots. We also conducted laboratory mesocosm experiments to
evaluate the preferences of the most common consumer at our study
location for rare vs. abundant seaweed species in our field plots. By
combining field observations and experiments with studies of
herbivore preferences, we document the importance of rare species
in shaping community structure from the bottom-up, mediating the
diversity, and especially the abundance, of organisms at higher trophic
levels.
Ecology Letters, (2012) 15: 461–467 doi: 10.1111/j.1461-0248.2012.01758.x
� 2012 Blackwell Publishing Ltd/CNRS
MATERIALS AND METHODS
Initial surveys
In early July of 2010, we marked 30 permanent square plots
(25 · 25 cm) in the mid-intertidal zone of a moderately wave-exposed
rocky reef at East Point, Nahant, Massachusetts, USA
(42º25¢00.71¢ N, 70º54¢19.23 W). Plots spanned approximately
100 m of shoreline, and community structure did not differ with
distance along the shore, suggesting that no underlying gradients (e.g.
wave energy) were responsible for variation in diversity in our plots.
We initially surveyed plots on 4 July 2010 to estimate per cent cover
of macroalgae and sessile invertebrates and count mobile inverte-
brates. Sessile invertebrates included the barnacle Semibalanus balanoides
L., the mussel Mytilus edulis L., the sea anemone Haliplanella lineata
Verrill, and the hydroid Dynamena pumila L., Macroalgae included the
Heterokontophytes Fucus vesiculosus L. and Ascophyllum nodosum (L.)
Le Jolis; the Rhodophytes Hildenbrandia rubra (Sommerfelt) Meneghini,
Mastocarpus stellatus (Stackhouse) Guiry, Chondrus crispus Stackhouse,
Vertebrata lanosa (L.) T. A. Christensen, Dumontia contorta (S. G. Gmelin)
Ruprecht, Porphyra umbilicalis Kutzing, and coralline crusts
[e.g. Clathromorphum circumscriptum (Stromfelt) Foslie]; and the Chloro-
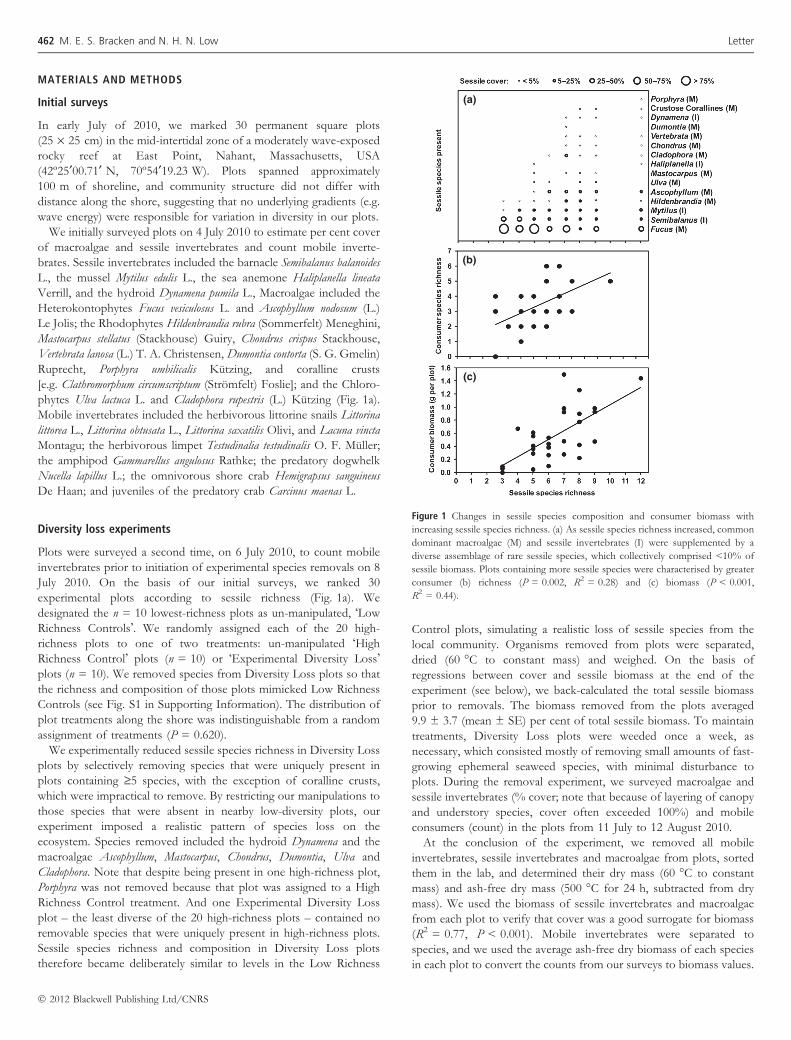
phytes Ulva lactuca L. and Cladophora rupestris (L.) Kutzing (Fig. 1a).
Mobile invertebrates included the herbivorous littorine snails Littorina
littorea L., Littorina obtusata L., Littorina saxatilis Olivi, and Lacuna vincta
Montagu; the herbivorous limpet Testudinalia testudinalis O. F. Muller;
the amphipod Gammarellus angulosus Rathke; the predatory dogwhelk
Nucella lapillus L.; the omnivorous shore crab Hemigrapsus sanguineus
De Haan; and juveniles of the predatory crab Carcinus maenas L.
Diversity loss experiments
Plots were surveyed a second time, on 6 July 2010, to count mobile
invertebrates prior to initiation of experimental species removals on 8
July 2010. On the basis of our initial surveys, we ranked 30
experimental plots according to sessile richness (Fig. 1a). We
designated the n = 10 lowest-richness plots as un-manipulated, �Low
Richness Controls�. We randomly assigned each of the 20 high-
richness plots to one of two treatments: un-manipulated �High
Richness Control� plots (n = 10) or �Experimental Diversity Loss�plots (n = 10). We removed species from Diversity Loss plots so that
the richness and composition of those plots mimicked Low Richness
Controls (see Fig. S1 in Supporting Information). The distribution of
plot treatments along the shore was indistinguishable from a random
assignment of treatments (P = 0.620).
We experimentally reduced sessile species richness in Diversity Loss
plots by selectively removing species that were uniquely present in
plots containing ‡5 species, with the exception of coralline crusts,
which were impractical to remove. By restricting our manipulations to
those species that were absent in nearby low-diversity plots, our
experiment imposed a realistic pattern of species loss on the
ecosystem. Species removed included the hydroid Dynamena and the
macroalgae Ascophyllum, Mastocarpus, Chondrus, Dumontia, Ulva and
Cladophora. Note that despite being present in one high-richness plot,
Porphyra was not removed because that plot was assigned to a High
Richness Control treatment. And one Experimental Diversity Loss
plot – the least diverse of the 20 high-richness plots – contained no
removable species that were uniquely present in high-richness plots.
Sessile species richness and composition in Diversity Loss plots
therefore became deliberately similar to levels in the Low Richness
Control plots, simulating a realistic loss of sessile species from the
local community. Organisms removed from plots were separated,
dried (60 �C to constant mass) and weighed. On the basis of
regressions between cover and sessile biomass at the end of the
experiment (see below), we back-calculated the total sessile biomass
prior to removals. The biomass removed from the plots averaged
9.9 ± 3.7 (mean ± SE) per cent of total sessile biomass. To maintain
treatments, Diversity Loss plots were weeded once a week, as
necessary, which consisted mostly of removing small amounts of fast-
growing ephemeral seaweed species, with minimal disturbance to
plots. During the removal experiment, we surveyed macroalgae and
sessile invertebrates (% cover; note that because of layering of canopy
and understory species, cover often exceeded 100%) and mobile
consumers (count) in the plots from 11 July to 12 August 2010.
At the conclusion of the experiment, we removed all mobile
invertebrates, sessile invertebrates and macroalgae from plots, sorted
them in the lab, and determined their dry mass (60 �C to constant
mass) and ash-free dry mass (500 �C for 24 h, subtracted from dry
mass). We used the biomass of sessile invertebrates and macroalgae
from each plot to verify that cover was a good surrogate for biomass
(R2 = 0.77, P < 0.001). Mobile invertebrates were separated to
species, and we used the average ash-free dry biomass of each species
in each plot to convert the counts from our surveys to biomass values.
(a)
(b)
(c)
Figure 1 Changes in sessile species composition and consumer biomass with
increasing sessile species richness. (a) As sessile species richness increased, common
dominant macroalgae (M) and sessile invertebrates (I) were supplemented by a
diverse assemblage of rare sessile species, which collectively comprised <10% of
sessile biomass. Plots containing more sessile species were characterised by greater
consumer (b) richness (P = 0.002, R2 = 0.28) and (c) biomass (P < 0.001,
R2 = 0.44).
462 M. E. S. Bracken and N. H. N. Low Letter
� 2012 Blackwell Publishing Ltd/CNRS

Although very little biomass was removed from Diversity Loss
plots, we evaluated the potential effects of biomass removal in two
ways. First, we evaluated relationships between the change in
consumer biomass in removal plots during the summer of 2010
[(final survey on 12 August 2010)–(initial survey on 4 July 2010)] and
both the amount of sessile biomass removed and the number of
sessile species removed.
We also conducted a follow-up experiment to assess whether the
minimal disturbance associated with removals could have affected
consumers. During the summer of 2011, we repeated our field
experiment with an additional treatment, the removal of biomass of
the abundant species present in all plots (i.e. regardless of sessile
richness) without actually reducing sessile richness. The biomass
removed from these Disturbance Controls was equivalent to the
biomass of the less abundant species that was removed in Experi-
mental Diversity Loss treatments. This experiment included 36 plots:
High Richness Controls (n = 9), Disturbance Controls (n = 9),
Experimental Diversity Loss plots (n = 9) and Low Richness Controls
(n = 9). Plots were initially surveyed on 30 July 2011, and removals
were conducted on 2 August 2011. An average of 6.2 ± 2.3
(mean ± SE) g dry biomass was removed from Diversity Loss plots,
and 6.5 ± 0.7 g was removed from Disturbance Control plots; the
amounts removed did not differ between treatments (t-test: P = 0.902)
and represented approximately 6% of total sessile biomass in each
removal plot. Biomass removals for common species involved
thinning the dominant macroalga Fucus and sessile invertebrates
Mytilus and Semibalanus. The experiment was maintained, and plots were
surveyed weekly, until 9 September 2011. Note that the experiment
was preceded by a series of heat waves (e.g. National Climatic Data
Center 2011) that coincided with daytime low tides, which caused
many mobile consumers to seek refuge lower on the shore and led to
low overall consumer richness and abundance in our early surveys.
Consumer preference experiment
We evaluated herbivore growth rates and preferences for abundant vs.
rare seaweeds by measuring consumption and growth rates of the
most common herbivore in our experimental plots, the snail
L. obtusata, using no-choice and multiple-choice grazing experiments.
The experiment was conducted in 0.7-L perforated transparent plastic
mesocosms placed in outdoor tanks plumbed with unfiltered running
seawater. Each mesocosm was divided into two halves by a plastic
mesh screen that separated the experimental snail-addition side from
the no-snail control side. We placed 6 g of seaweed in both sides of
every mesocosm and added 5 g (wet mass, equivalent to approx-
imately 7 individuals) of snails to one, randomly selected side of each
mesocosm. We measured grazing rates in n = 10 replicates each of
seven different treatments: each seaweed species by itself and all six
species (Fucus, Ascophyllum, Cladophora, Mastocarpus, Ulva and Chondrus)
together. Grazing was assessed by comparing, for each species, the
change in algal biomasses on each side of each mesocosm. Values are
reported as the difference in % biomass lost over 4 days in treatments
with and without snails.
Statistical analyses
We analysed our data using general linear models, generalised linear
models (i.e. logistic regression to evaluate whether Cladophora, a
preferred seaweed species, occurred more often in more diverse plots)
and t-tests after verifying that the data met assumptions of normality
and homogeneity of variances. In the multiple-choice grazing
experiment, algal species in each chamber were not independent of
one another, so comparisons between species were made using paired
t-tests. We assessed changes in sessile and mobile consumer
community composition along the shore by conducting non-metric
multidimensional scaling (MDS) analyses based on square-root
transformed data with the Bray-Curtis measure of similarity. Analyses
were conducted using SAS version 9.2 (SAS Institute 2008) and
PRIMER version 6 (Clarke & Gorley 2006).
RESULTS
Initial surveys
We verified that underlying gradients along the shore were not
responsible for variation in diversity in our experimental plots by
evaluating relationships between distance and sessile species cover
(F1,28 = 2.9, P = 0.100), sessile species richness (F1,28 = 0.0,
P = 0.996), sessile species evenness (F1,28 = 2.4, P = 0.125), con-
sumer biomass (F1,28 = 0.0, P = 0.851), consumer richness
(F1,28 = 3.4, P = 0.078) and consumer evenness (F1,28 = 3.4,
P = 0.072). Neither sessile (stress = 0.12, F1,28 = 1.4, P = 0.241)
nor mobile consumer (stress = 0.01, F1,28 = 2.9, P = 0.101) com-
munity composition (i.e. MDS axis 1) changed with distance along the
shore.
In our surveys of 30 plots, we found that as the number of sessile
species increased, the few abundant species found in all plots (e.g. the
rockweed Fucus, the barnacle Semibalanus, and the mussel Mytilus) were
supplemented by a number of rare sessile species (up to eight
additional species in the most diverse plots; Fig. 1a). These rare
species collectively represented an average of <10% of biomass in
plots where they occurred. Increases in sessile species richness were
associated with increases in both the richness (F1,28 = 10.94,
P = 0.002, R2 = 0.28; Fig. 1b) and especially the biomass
(F1,28 = 22.1, P < 0.001, R2 = 0.44; Fig. 1c) of mobile consumer
species, suggesting that rare sessile species could facilitate mobile taxa
from the bottom up.
Diversity loss experiment
Prior to the start of our experiment, Experimental Diversity Loss
plots contained the same number of sessile species as High Richness
Control plots (t = 0.7, d.f. = 18, P = 0.518) and more species than
the Low Richness Control plots (t = 4.9, d.f. = 18, P < 0.001;
Fig. 2a). Removals caused an immediate decline in sessile richness
so that by the next survey date, sessile richness in Diversity Loss plots
matched that in Low Richness Controls (t = 0.37, d.f. = 18,
P = 0.718), but was lower than that of the un-manipulated High
Richness Controls (t = 6.95, d.f. = 18, P < 0.001). This pattern
continued for the duration of our experiment. At the same time,
sessile cover, a good surrogate for biomass in this system, never
differed among the three treatments (P > 0.05; Fig. 2b). Thus, we
experimentally achieved a 40% reduction in sessile species richness,
but the species removed were extremely rare, so removals had no
effect on the total abundance of sessile species.
After experimental removals, the richness of mobile species in
Experimental Diversity Loss plots declined rapidly relative to High
Richness Controls (Fig. 2c). At the end of the experiment, Diversity
Letter Loss of rare �cornerstone� species 463
� 2012 Blackwell Publishing Ltd/CNRS
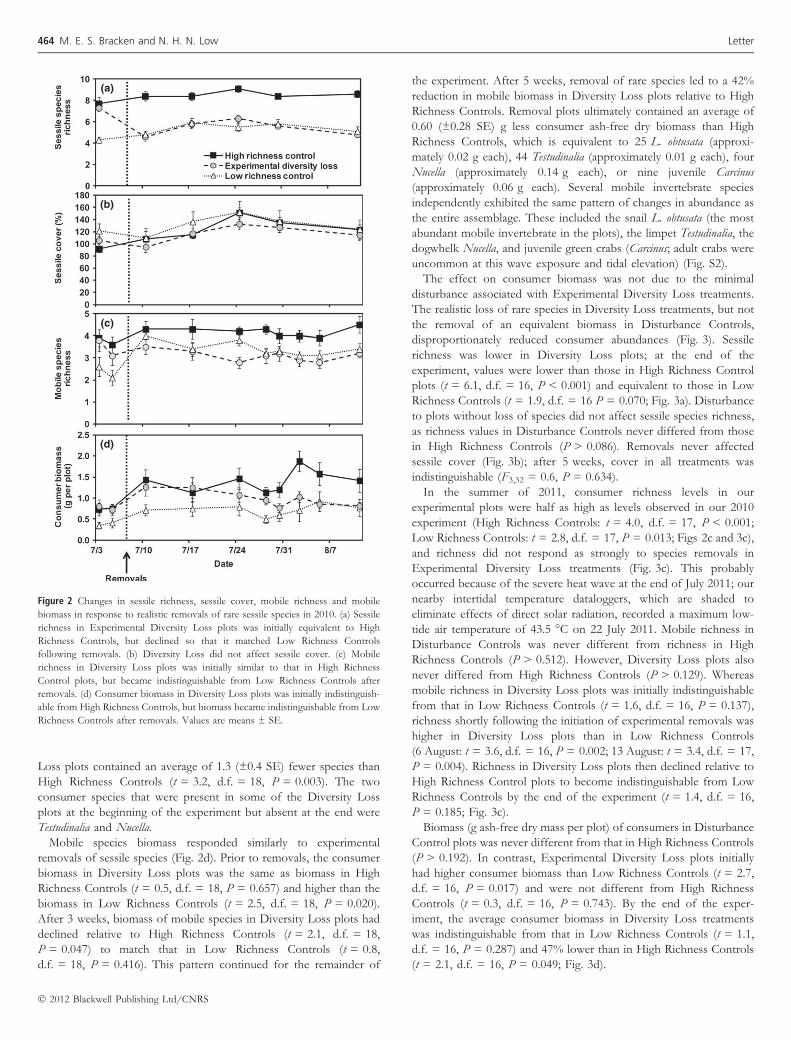
Loss plots contained an average of 1.3 (±0.4 SE) fewer species than
High Richness Controls (t = 3.2, d.f. = 18, P = 0.003). The two
consumer species that were present in some of the Diversity Loss
plots at the beginning of the experiment but absent at the end were
Testudinalia and Nucella.
Mobile species biomass responded similarly to experimental
removals of sessile species (Fig. 2d). Prior to removals, the consumer
biomass in Diversity Loss plots was the same as biomass in High
Richness Controls (t = 0.5, d.f. = 18, P = 0.657) and higher than the
biomass in Low Richness Controls (t = 2.5, d.f. = 18, P = 0.020).
After 3 weeks, biomass of mobile species in Diversity Loss plots had
declined relative to High Richness Controls (t = 2.1, d.f. = 18,
P = 0.047) to match that in Low Richness Controls (t = 0.8,
d.f. = 18, P = 0.416). This pattern continued for the remainder of
the experiment. After 5 weeks, removal of rare species led to a 42%
reduction in mobile biomass in Diversity Loss plots relative to High
Richness Controls. Removal plots ultimately contained an average of
0.60 (±0.28 SE) g less consumer ash-free dry biomass than High
Richness Controls, which is equivalent to 25 L. obtusata (approxi-
mately 0.02 g each), 44 Testudinalia (approximately 0.01 g each), four
Nucella (approximately 0.14 g each), or nine juvenile Carcinus
(approximately 0.06 g each). Several mobile invertebrate species
independently exhibited the same pattern of changes in abundance as
the entire assemblage. These included the snail L. obtusata (the most
abundant mobile invertebrate in the plots), the limpet Testudinalia, the
dogwhelk Nucella, and juvenile green crabs (Carcinus; adult crabs were
uncommon at this wave exposure and tidal elevation) (Fig. S2).
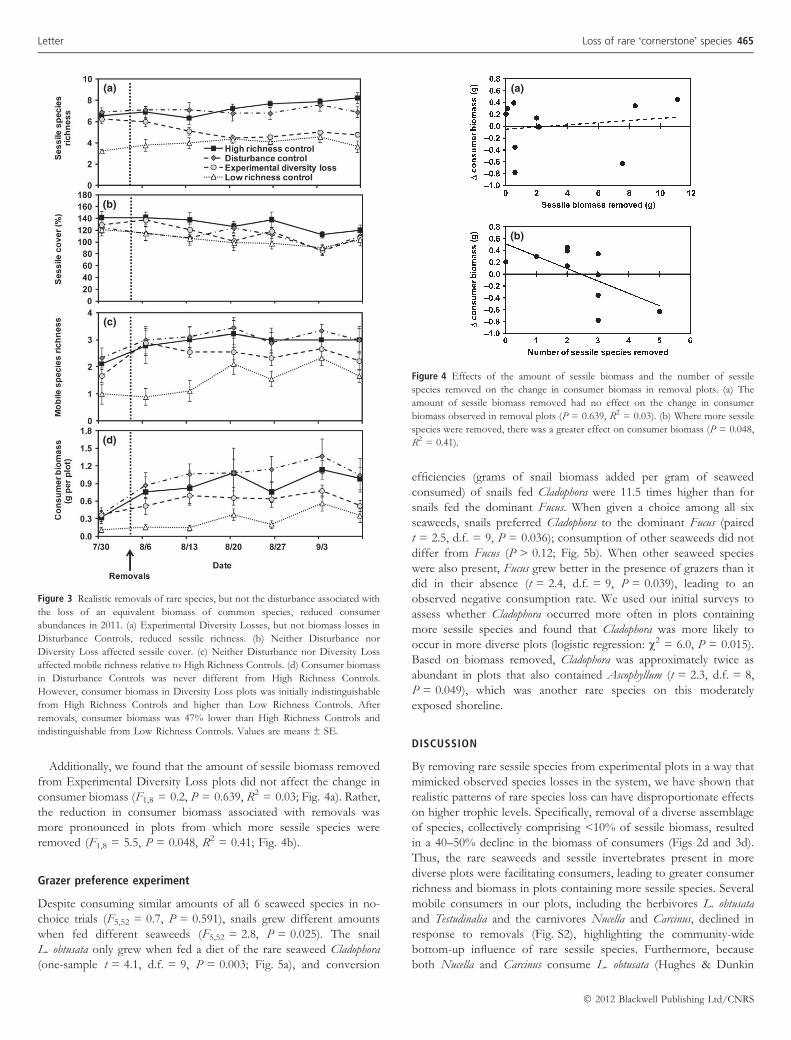
The effect on consumer biomass was not due to the minimal
disturbance associated with Experimental Diversity Loss treatments.
The realistic loss of rare species in Diversity Loss treatments, but not
the removal of an equivalent biomass in Disturbance Controls,
disproportionately reduced consumer abundances (Fig. 3). Sessile
richness was lower in Diversity Loss plots; at the end of the
experiment, values were lower than those in High Richness Control
plots (t = 6.1, d.f. = 16, P < 0.001) and equivalent to those in Low
Richness Controls (t = 1.9, d.f. = 16 P = 0.070; Fig. 3a). Disturbance
to plots without loss of species did not affect sessile species richness,
as richness values in Disturbance Controls never differed from those
in High Richness Controls (P > 0.086). Removals never affected
sessile cover (Fig. 3b); after 5 weeks, cover in all treatments was
indistinguishable (F3,32 = 0.6, P = 0.634).
In the summer of 2011, consumer richness levels in our
experimental plots were half as high as levels observed in our 2010
experiment (High Richness Controls: t = 4.0, d.f. = 17, P < 0.001;
Low Richness Controls: t = 2.8, d.f. = 17, P = 0.013; Figs 2c and 3c),
and richness did not respond as strongly to species removals in
Experimental Diversity Loss treatments (Fig. 3c). This probably
occurred because of the severe heat wave at the end of July 2011; our
nearby intertidal temperature dataloggers, which are shaded to
eliminate effects of direct solar radiation, recorded a maximum low-
tide air temperature of 43.5 �C on 22 July 2011. Mobile richness in
Disturbance Controls was never different from richness in High
Richness Controls (P > 0.512). However, Diversity Loss plots also
never differed from High Richness Controls (P > 0.129). Whereas
mobile richness in Diversity Loss plots was initially indistinguishable
from that in Low Richness Controls (t = 1.6, d.f. = 16, P = 0.137),
richness shortly following the initiation of experimental removals was
higher in Diversity Loss plots than in Low Richness Controls
(6 August: t = 3.6, d.f. = 16, P = 0.002; 13 August: t = 3.4, d.f. = 17,
P = 0.004). Richness in Diversity Loss plots then declined relative to
High Richness Control plots to become indistinguishable from Low
Richness Controls by the end of the experiment (t = 1.4, d.f. = 16,
P = 0.185; Fig. 3c).
Biomass (g ash-free dry mass per plot) of consumers in Disturbance
Control plots was never different from that in High Richness Controls
(P > 0.192). In contrast, Experimental Diversity Loss plots initially
had higher consumer biomass than Low Richness Controls (t = 2.7,
d.f. = 16, P = 0.017) and were not different from High Richness
Controls (t = 0.3, d.f. = 16, P = 0.743). By the end of the exper-
iment, the average consumer biomass in Diversity Loss treatments
was indistinguishable from that in Low Richness Controls (t = 1.1,
d.f. = 16, P = 0.287) and 47% lower than in High Richness Controls
(t = 2.1, d.f. = 16, P = 0.049; Fig. 3d).
(a)
(b)
(c)
(d)
Figure 2 Changes in sessile richness, sessile cover, mobile richness and mobile
biomass in response to realistic removals of rare sessile species in 2010. (a) Sessile
richness in Experimental Diversity Loss plots was initially equivalent to High
Richness Controls, but declined so that it matched Low Richness Controls
following removals. (b) Diversity Loss did not affect sessile cover. (c) Mobile
richness in Diversity Loss plots was initially similar to that in High Richness
Control plots, but became indistinguishable from Low Richness Controls after
removals. (d) Consumer biomass in Diversity Loss plots was initially indistinguish-
able from High Richness Controls, but biomass became indistinguishable from Low
Richness Controls after removals. Values are means ± SE.
464 M. E. S. Bracken and N. H. N. Low Letter
� 2012 Blackwell Publishing Ltd/CNRS
Additionally, we found that the amount of sessile biomass removed
from Experimental Diversity Loss plots did not affect the change in
consumer biomass (F1,8 = 0.2, P = 0.639, R2 = 0.03; Fig. 4a). Rather,
the reduction in consumer biomass associated with removals was
more pronounced in plots from which more sessile species were
removed (F1,8 = 5.5, P = 0.048, R2 = 0.41; Fig. 4b).
Grazer preference experiment
Despite consuming similar amounts of all 6 seaweed species in no-
choice trials (F5,52 = 0.7, P = 0.591), snails grew different amounts
when fed different seaweeds (F5,52 = 2.8, P = 0.025). The snail
L. obtusata only grew when fed a diet of the rare seaweed Cladophora
(one-sample t = 4.1, d.f. = 9, P = 0.003; Fig. 5a), and conversion
efficiencies (grams of snail biomass added per gram of seaweed
consumed) of snails fed Cladophora were 11.5 times higher than for
snails fed the dominant Fucus. When given a choice among all six
seaweeds, snails preferred Cladophora to the dominant Fucus (paired
t = 2.5, d.f. = 9, P = 0.036); consumption of other seaweeds did not
differ from Fucus (P > 0.12; Fig. 5b). When other seaweed species
were also present, Fucus grew better in the presence of grazers than it
did in their absence (t = 2.4, d.f. = 9, P = 0.039), leading to an
observed negative consumption rate. We used our initial surveys to
assess whether Cladophora occurred more often in plots containing
more sessile species and found that Cladophora was more likely to
occur in more diverse plots (logistic regression: v2 = 6.0, P = 0.015).
Based on biomass removed, Cladophora was approximately twice as
abundant in plots that also contained Ascophyllum (t = 2.3, d.f. = 8,
P = 0.049), which was another rare species on this moderately
exposed shoreline.
DISCUSSION
By removing rare sessile species from experimental plots in a way that
mimicked observed species losses in the system, we have shown that
realistic patterns of rare species loss can have disproportionate effects
on higher trophic levels. Specifically, removal of a diverse assemblage
of species, collectively comprising <10% of sessile biomass, resulted
in a 40–50% decline in the biomass of consumers (Figs 2d and 3d).
Thus, the rare seaweeds and sessile invertebrates present in more
diverse plots were facilitating consumers, leading to greater consumer
richness and biomass in plots containing more sessile species. Several
mobile consumers in our plots, including the herbivores L. obtusata
and Testudinalia and the carnivores Nucella and Carcinus, declined in
response to removals (Fig. S2), highlighting the community-wide
bottom-up influence of rare sessile species. Furthermore, because
both Nucella and Carcinus consume L. obtusata (Hughes & Dunkin
(a)
(b)
(c)
(d)
Figure 3 Realistic removals of rare species, but not the disturbance associated with
the loss of an equivalent biomass of common species, reduced consumer
abundances in 2011. (a) Experimental Diversity Losses, but not biomass losses in
Disturbance Controls, reduced sessile richness. (b) Neither Disturbance nor
Diversity Loss affected sessile cover. (c) Neither Disturbance nor Diversity Loss
affected mobile richness relative to High Richness Controls. (d) Consumer biomass
in Disturbance Controls was never different from High Richness Controls.
However, consumer biomass in Diversity Loss plots was initially indistinguishable
from High Richness Controls and higher than Low Richness Controls. After
removals, consumer biomass was 47% lower than High Richness Controls and
indistinguishable from Low Richness Controls. Values are means ± SE.
(a)
(b)
Figure 4 Effects of the amount of sessile biomass and the number of sessile
species removed on the change in consumer biomass in removal plots. (a) The
amount of sessile biomass removed had no effect on the change in consumer
biomass observed in removal plots (P = 0.639, R2 = 0.03). (b) Where more sessile
species were removed, there was a greater effect on consumer biomass (P = 0.048,
R2 = 0.41).
Letter Loss of rare �cornerstone� species 465
� 2012 Blackwell Publishing Ltd/CNRS

1984; Rangeley & Thomas 1987), and Carcinus consumes Nucella
(Rangeley & Thomas 1987), the effects of rare species removals
probably propagate up the food web.
The rare species removed from our plots play essential roles in
providing food (Lubchenco & Menge 1978; Long & Trussell 2007)
and ⁄ or habitat (Lubchenco 1978; Bertness et al. 1999) for these
consumers, and the fact that the effect of removals on consumer
biomass increased with the number of sessile species removed
(Fig. 4b) suggests that sessile species may act together to influence
mobile consumers. For example, the canopy of the seaweed
Ascophyllum, one of the rare species removed from our plots, reduces
temperature and desiccation stress, facilitating a variety of taxa
(Bertness et al. 1999), including another rare seaweed, Cladophora
(Ingolfsson & Hawkins 2008). Cladophora biomass was twice as high in
our removal plots that also contained Ascophyllum. Our grazer
preference experiment highlights the importance of Cladophora to
consumers; snails only grew when fed Cladophora, and they consumed
it more readily than the dominant Fucus (Fig. 5). Cladophora was more
likely to occur in plots containing more sessile species, and it was
present in most (7 ⁄ 10) of the removal plots in our 2010 experiment.
In fact, when alternative, more palatable seaweeds (e.g. Cladophora)
were available, Fucus actually grew more when grazers were present
than when absent (Fig. 5b), highlighting a potential role of herbivores
in enhancing the dominant seaweed species in a diverse assemblage.
The rapid response of consumers to removals of rare sessile species
was particularly striking. In contrast to removals of rocky intertidal
consumers, where effects on the diversity and biomass of sessile
species can take months to become apparent (Paine 1974; Castilla &
Duran 1985), we observed negative effects of the removal of sessile
species on consumer biomass and diversity within 3 weeks. Effects of
consumer removals are mediated by recruitment of sessile species
(Paine 1974; Castilla & Duran 1985; Menge et al. 1994), whereas
mobile consumers can rapidly emigrate in search of higher quality
food (Root 1973; Levinton & Kelaher 2004). Especially at small spatial
scales such as the plot sizes in our experiment, the movement of
mobile organisms away from suboptimal habitats, demonstrated both
here and in prior work (Parker & Stuart 1976; Levinton & Kelaher
2004), adds an under-appreciated behavioural dimension to our
understanding of the bottom-up consequences of species loss. At
larger spatial scales, rarity of palatable food sources is probably
mediated, at least in part, by consumers.
The rare species we removed had large and disproportionate effects
on higher trophic levels, and therefore acted similarly to keystone
species in their effect on community structure (Power et al. 1996).
There is no theoretical reason why keystone species cannot operate
from anywhere in a food web (Pimm 1982). However, the term, as
originally defined by Paine (1969), generally refers to predators. We
therefore call rare species that have large and disproportionate
bottom-up effects on consumers �cornerstone species� to differentiate
them from both keystone predators (Paine 1969, 1974) and
foundation species (Dayton 1972; Ellison et al. 2005). Architecturally,
a keystone represents a relatively small portion of an arch, but
determines its stability and function from the top-down, and a
foundation supports a structure from the bottom-up, but by virtue of
its mass. A cornerstone represents a relatively small portion of a
foundation, but defines the shape of the structure above it.
We removed species in a way that mimicked realistic species losses in
the system – our removal treatments were specifically designed to match
naturally occurring low-richness plots – demonstrating the importance
of rare species as mediators of community structure and dynamics. We
show that large and disproportionate effects of rare species can act not
only from the top-down, as several previous studies have demonstrated
(Power et al. 1996), but also from the bottom-up, influencing the
richness and especially the abundance of other organisms in a
community (see also Nason et al. 1998). With respect to conservation
priorities in an era of global biodiversity change (Butchart et al. 2010),
understanding the factors that influence biodiversity is critical. Here,
a suite of inconspicuous species maintains rocky intertidal community
structure, and this effect increases with their diversity. It is not just
common, dominant species that mediate abundance and diversity at
higher trophic levels (e.g. Ellison et al. 2005; Gaston & Fuller 2008). Rare
species act as the cornerstones of this community, comprising relatively
little biomass, but dictating the shape of the community they support.
ACKNOWLEDGEMENTS
We thank C. Bracken-Sorte, A. Drouin, V. Perini, S. Trieweiler, and
students in the Three Seas Program for assistance in setting up and
maintaining experiments. B. Menge, E. Sanford, C. Sorte, J.
Stachowicz, G. Trussell, T. Wootton and anonymous referees
provided helpful comments on the manuscript. This work was
funded by the National Science Foundation (OCE-0961364 to M. E.
S. B. and G. Trussell). This is contribution number 280 of the
Northeastern University Marine Science Center.
AUTHOR CONTRIBUTION
MESB and NHNL designed and performed the research; MESB
analysed the data and wrote the paper; and both MESB and NHNL
contributed substantially to revisions.
(a)
(b)
Figure 5 Grazing and growth rates of snails feeding on different seaweeds. (a) When
snails were offered each seaweed by itself, they consumed an equivalent amount of
the tissue of each species (ANOVA: P = 0.591). However, their growth depended on
the species provided (ANOVA: P = 0.025). Specifically, snail growth only occurred
when snails were fed Cladophora. (b) Snails that were given all six seaweed species at
the same time consumed more Cladophora than they did Fucus. Asterisks (*) indicate
rare species (a) on which snails grew or (b) which were consumed at higher rates than
the dominant Fucus (P < 0.05). Values are means ± SE.
466 M. E. S. Bracken and N. H. N. Low Letter
� 2012 Blackwell Publishing Ltd/CNRS

REFERENCES
Barnosky, A.D., Matzke, N., Tomiya, S., Wogan, G.O.U., Swartz, B., Quental, T.B.
et al. (2011). Has the Earth�s sixth mass extinction already arrived? Nature, 471,
51–57.
Bertness, M.D., Leonard, G.H., Levine, J.M., Schmidt, P.R. & Ingraham, A.O.
(1999). Testing the relative contribution of positive and negative interactions in
intertidal communities. Ecology, 80, 2711–2726.
Bracken, M.E.S., Bracken, B.E. & Rogers-Bennett, L. (2007). Species diversity and
foundation species: potential indicators of fisheries yields and marine ecosystem
functioning. Calif. Coop. Oceanic Fish. Invest. Rep., 48, 82–91.
Bracken, M.E.S., Friberg, S.E., Gonzalez-Dorantes, C.A. & Williams, S.L. (2008).
Functional consequences of realistic biodiversity changes in a marine ecosystem.
Proc. Natl. Acad. Sci. USA, 105, 924–928.
Butchart, S.H.M., Walpole, M., Collen, B., van Strien, A., Scharlemann, J.P.W.,
Almond, R.E.A. et al. (2010). Global biodiversity: indicators of recent declines.
Science, 328, 1164–1168.
Cardinale, B.J., Srivastava, D.S., Duffy, J.E., Wright, J.P., Downing, A.L., Sankaran, M.
et al. (2006). Effects of biodiversity on the functioning of trophic groups and
ecosystems. Nature, 443, 989–992.
Castilla, J.C. & Duran, L.R. (1985). Human exclusion from the rocky intertidal zone
of central Chile: the effects on Concholepas concholepas (Gastropoda). Oikos, 45,
391–399.
Clarke, K.R.Gorley, R.N. (2006). PRIMER v6: User Manual ⁄ Tutorial. PRIMER-E,
Ltd., Plymouth.
Dayton, P.K. (1972). Toward an understanding of community resilience and the
potential effects of enrichment to the benthos at McMurdo Sound, Antarctica.
In: Proceedings of the Colloquium on Conservation Problems in Antarctica (ed. Parker,
B.C.). Allen Press, Lawrence, pp. 81–96.
Ellison, A.M., Bank, M.S., Clinton, B.D., Colburn, E.A., Elliott, K., Ford, C.R. et al.
(2005). Loss of foundation species: consequences for the structure and dynamics
of forested ecosystems. Front. Ecol. Environ., 3, 479–486.
Elton, C.S. (1927). Animal Ecology. University of Chicago Press, Chicago.
Gaston, K.J. & Fuller, R.A. (2008). Commonness, population depletion and con-
servation biology. Trends Ecol. Evol., 23, 14–19.
Gotelli, N.J. & Graves, G.R. (1996) Null Models in Ecology. Smithsonian Institution
Press, Washington.
Hairston, N.G. Jr & Hairston, N.G. Sr (1993). Cause-effect relationships in energy
flow, trophic structure, and interspecific interactions. Am. Nat., ?????, 379–411.
Hooper, D.U., Chapin, F.S. III, , Ewel, J.J., Hector, A., Inchausti, P., Lavorel, S.
et al. (2005). Effects of biodiversity on ecosystem functioning: a consensus of
current knowledge. Ecol. Monogr., 75, 3–35.
Hughes, R.N. & Dunkin, S.B. (1984). Behavioural components of prey selection by
dogwhelks, Nucella lapillus (L.), feeding on mussels, Mytilus edulis L., in the lab-
oratory. J. Exper. Mar. Biol. Ecol., 77, 45–68.
Hunter, M.D. & Price, P.W. (1992). Playing chutes and ladders: heterogeneity and
the relative roles of bottom-up and top-down forces in natural communities.
Ecology, 73, 723–732.
Hutchinson, G.E. (1959). Homage to Santa Rosalia, or why are there so many kinds
of animals? Am. Nat., 93, 145–159.
Ingolfsson, A. & Hawkins, S.J. (2008). Slow recovery from disturbance: a 20 year
study of Ascophyllum canopy clearances. J. Mar. Biol. Assoc. UK, 88, 689–691.
Knops, J.M.H., Tilman, D., Haddad, N.M., Naeem, S., Mitchell, C.E., Haarstad, J.
et al. (1999). Effects of plant species richness on invasion dynamics, disease
outbreaks, insect abundances and diversity. Ecol. Lett., 2, 286–293.
Levinton, J. & Kelaher, B. (2004). Opposing organizing forces of deposit-feeding
marine communities. J. Exp. Mar. Biol. Ecol., 300, 65–82.
Lindeman, R.L. (1942). The trophic-dynamic aspect of ecology. Ecology, 23, 399–
417.
Long, J.D. & Trussell, G.C. (2007). Geographic variation in seaweed induced
responses to herbivory. Mar. Ecol. Prog. Ser., 333, 75–80.
Lubchenco, J. (1978). Plant species diversity in a marine intertidal community:
importance of herbivore food preference and algal competitive abilities.
Am. Nat., 112, 23–39.
Lubchenco, J. & Menge, B.A. (1978). Community development and persistence in a
low rocky intertidal zone. Ecol. Monogr., 48, 67–94.
Lyons, K.G. & Schwartz, M.W. (2001). Rare species loss alters ecosystem function
– invasion resistance. Ecol. Lett., 4, 358–365.
Marsh, A.S., Arnone, J.A. III, Bormann, B.T. & Gordon, J.C. (2000). The role of
Equisetum in nutrient cycling in an Alaskan shrub wetland. J. Ecol., 88, 999–1011.
Menge, B.A. & Sutherland, J.P. (1987). Community regulation: variation in dis-
turbance, competition, and predation in relation to environmental stress and
recruitment. Am. Nat., 130, 730–757.
Menge, B.A., Berlow, E.L., Blanchette, C.A., Navarrete, S.A. & Yamada, S.B.
(1994). The keystone species concept: variation in interaction strength in a rocky
intertidal habitat. Ecol. Monogr., 64, 249–286.
Nason, J.D., Herre, E.A. & Hamrick, J.L. (1998). The breeding structure of a
tropical keystone plant resource. Nature, 391, 685–687.
National Climatic Data Center (2011). State of the Climate: National Overview for July
2011. National Oceanic and Atmospheric Administration, Asheville.
Novotny, V. & Basset, Y. (2000). Rare species in communities of tropical insect
herbivores: pondering the mystery of singletons. Oikos, 89, 564–572.
Oksanen, L., Fretwell, S.D., Arruda, J. & Niemela, P. (1981). Exploitation eco-
systems in gradients of primary productivity. Am. Nat., 118, 240–261.
Paine, R.T. (1969). A note on trophic complexity and community stability.
Am. Nat., 103, 91–93.
Paine, R.T. (1974). Intertidal community structure: experimental studies on the
relationship between a dominant competitor and Its principal predator. Oecologia,
15, 93–120.
Paine, R.T. (1980). Food webs: linkage, interaction strength and community
infrastructure. J. Anim. Ecol., 49, 667–685.
Parker, G.A. & Stuart, R.A. (1976). Animal behavior as a strategy optimizer:
evolution of resource assessment strategies and optimal emigration thresholds.
Am. Nat., 110, 1055–1076.
Pimm, S.L. (1980). Properties of food webs. Ecology, 61, 219–225.
Pimm, S.L. (1982). Food Webs. Chapman and Hall, London.
Power, M.E., Tilman, D., Estes, J.A., Menge, B.A., Bond, W.J., Mills, L.S. et al.
(1996). Challenges in the quest for keystones. Bioscience, 46, 609–620.
Rangeley, R.W. & Thomas, M.L.H. (1987). Predatory behaviour of juvenile shore
crab Carcinus maenas (L.). J. Exper. Mar. Biol. Ecol., 108, 191–197.
Root, R.B. (1973). Organization of a plant-arthropod association in simple and
diverse habitats: the fauna of collards (Brassica oleracea). Ecol. Monogr., 43, 95–124.
SAS Institute. (2008). SAS Version 9.2. SAS Institute, Cary, NC.
Schwartz, M.W., Brigham, C.A., Hoeksema, J.D., Lyons, K.G., Mills, M.H. & van
Mantgem, P.J. (2000). Linking biodiversity to ecosystem function: implications
for conservation ecology. Oecologia, 122, 297–305.
Smith, M.D. & Knapp, A.K. (2003). Dominant species maintain ecosystem function
with non-random species loss. Ecol. Lett., 6, 509–517.
Solan, M., Cardinale, B.J., Downing, A.L., Engelhardt, K.A.M., Ruesink, J.L. &
Srivastava, D.S. (2004). Extinction and ecosystem function in the marine ben-
thos. Science, 306, 1177–1180.
Srivastava, D.S. & Vellend, M. (2005). Biodiversity-ecosystem function research: is
it relevant to conservation? Annu. Rev. Ecol. Evol. Syst., 36, 267–294.
Zavaleta, E.S. & Hulvey, K.B. (2004). Realistic species losses disproportionately
reduce grassland resistance to biological invaders. Science, 306, 1175–1177.
SUPPORTING INFORMATION
Additional Supporting Information may be downloaded via the online
version of this article at Wiley Online Library (www.ecologyletters.com).
As a service to our authors and readers, this journal provides
supporting information supplied by the authors. Such materials are
peer-reviewed and may be re-organised for online delivery, but are not
copy-edited or typeset. Technical support issues arising from
supporting information (other than missing files) should be addressed
to the authors.
Editor, Tim Wootton
Manuscript received 20 December 2011
First decision made 24 January 2012
Second decision made 7 February 2012
Manuscript accepted 8 February 2012
Letter Loss of rare �cornerstone� species 467
� 2012 Blackwell Publishing Ltd/CNRS





























