Embed Size (px)
Citation preview

ISSN - 1135 - 5417
VOLUMEN XXVII
REALACADEMIA GALEGA DE CIENCIAS
Rev i s t a
SANTIAGO DE COMPOSTELA
2008

R E V I S T A
REAL
ACADEMIA GALEGA DE CIENCIAS
VOLUMEN XXVII
SANTIAGO DE COMPOSTELA
2008

ISSN - 1135 - 5417
© REAL ACADEMIA GALEGA DE CIENCIASIMPRIME: GRAFICOLOR MINERVA, S.L. POLÍGONO INDUSTRIAL DEL TAMBRE - GÜTENBERG, 5 - SANTIAGO, 2009DEPÓSITO LEGAL: C - 277 / 84
Subvencionada por: Fundación Pedro Barrié de la Maza Consellería de Innovación e Industria da Xunta de Galicia
Real Academia Galega de Cienciashttp://[email protected]

Pág.
DETERMINACIÓN DE METALES EN ALGAS MARINAS DESTINADAS A LA ALIMENTACIÓN HUMANA PRODUCIDAS Y PROCESADAS EN GALICIA: DESARROLLO DE METODOLOGÍA ANALÍTICA AUTOMÁTICA, por M. Carmen Yebra Biurrun .........................................
EVALUACIÓN DEL EFECTO DE LA SELENOMETIONINA SOBRE LA REPARACIÓN DEL DAÑO EN EL ADN, por Vanessa Valdiglesias, Blanca Laffon, Eduardo Pásaro y Josefina Méndez .................................................
COMPOSICIÓN EN MACRONUTRIENTES, AMINOÁCIDOS Y MINERALES DE ALGUNOS INVERTEBRADOS MARINOS NO UTILIZADOS HABITUALMENTE COMO ALIMENTO, por C. Taboada, R. Millán, I. Miguez, E. Fernández-Pulpeiro ..........................................
DESARROLLO DE BANCOS DE GERMOPLASMA DE CASTAÑO Y ALCORNOQUE MEDIANTE CRIOCONSERVACIÓN DE ÁPICES CAULINARES Y EMBRIONES SOMÁTICOS, por Nieves Vidal, Ana M. Vieitez, M. Rosario Fernández y Beatriz Cuenca ....................................
ESTUDO ANTROPOLÓXICO DOS RESTOS ÓSEOS RECUPERADOS DA NECRÓPOLE DA COVA DO SANTO, PARDOLLÁN, RUBIÁ, OURENSE. UNHA NECRÓPOLE DA IDADE DO BRONCE EN GALICIA, por Olalla López-Costas ........................................................
ESTIMACIÓN DE LA EVAPOTRANSPIRACIÓN DIARIA A PARTIR DE PRONÓSTICOS METEOROLÓGICOS EN GALICIA, por J. A. Rodríguez-Suárez, S. Feijóo, B. Soto ............................................................................
DIFUSIONES DÉMICAS Y COMPONENTES DE LA ESTRUCTURA GENÉTICA DEL NW AFRICANO. PERSPECTIVAS MULTIVARIANTES DE STRS AUTOSÓMICOS, por B. Caeiro, A. Ferreiro, D. Beiroa, J. A. Rodríguez-Pérez, F. Picchi-Figueira, M. M. Regueiro ......................................
LA CORRIENTE DE MALVINAS: ¿UNA VÍA DE DISPERSIÓN PARA CNIDARIOS BENTÓNICOS DE AGUAS FRÍAS?, por M. O. Zamponi ........
VIDA DA REAL ACADEMIA GALEGA DE CIENCIAS .........................
MEMORIA DAS ACTIVIDADES ..............................................................
CURSOS DE CONFERENCIAS .................................................................
PREMIOS DE INVESTIGACIÓN ...............................................................
COMPOSICIÓN DA REAL ACADEMIA ..................................................
PUBLICACIÓNS DA REAL ACADEMIA ................................................
INSTRUCCIÓNS PARA OS AUTORES ..................................................... 231
228
224
222
218
206
205
183
163
145
131
107
95
41
5
ÍNDICE


Revista Real Academia Galega de Ciencias. Vol. XXVII. Págs. 5-40 (2008)
DETERMINACIÓN DE METALES EN ALGAS MARINAS DESTINADAS A LA ALIMENTACIÓN HUMANA
PRODUCIDAS Y PROCESADAS EN GALICIA: DESARROLLO DE METODOLOGÍA ANALÍTICA AUTOMÁTICA
M. CARMEN YEBRA BIURRUN1
Departamento de Química Analítica, Nutrición y Bromatología.Facultad de Química. Universidad de Santiago de Compostela.
15782-Santiago de [email protected]
RESUMEN
Se ha propuesto un método nuevo, simple, rápido y automático basado en la extracción ácida asistida con energía de ultrasonidos para la determinación de metales en algas. Los metales se determinaron en línea por espectrometría de absorción atómica con llama utilizando un sistema de análisis por inyección en flujo. Se estudió la influencia de varias variables en la eficacia del proceso de extracción utilizando diseños experimentales. Además, las metodologías propuestas fueron evaluadas calculando el límite de detección, límite de cuantificación y precisión. La validación se efectuó analizando un material de referencia certificado (NIES-9, ”Sargasso”, National Institute for Environmental Studies, Japan) y mediante estudios de recuperación. Finalmente, la metodología propuesta se ha utilizado para la determinación de Zn, Cu, Fe, Mn, Ca y Mg en algas marinas destinadas a la alimentación humana producidas y procesadas en Galicia.
Palabras clave: metales, extracción ácida continua, ultrasonidos, análisis por inyección en flujo, algas.
1 Premio de Investigación Real Academia Galega de Ciencias-Fundación Caixa Galicia, convocatoria 2007.

6 Revista Real Academia Galega de Ciencias. Vol. XXVII
ABSTRACT
A new, simple, fast and automated method based on continuous ultrasound-assisted acid extraction has been developed for the determination of metals (Zn, Cu, Fe, Mn, Ca and Mg). The target metals have been on-line monitoring by flame atomic absorption spectrometry by using a flow injection analysis manifold. The influence of several extraction parameters on the efficiency of metal leaching has been evaluated by using experimental designs. Limits of detection and quantification and precision of the over all procedure have been assessed. Method validation was performed analysing a seaweed reference material (NIES-9, “Sargasso”, National Institute for Environmental Studies, Japan) and by recovery studies. Finally, the developed continuous extraction methodology has been applied to the determination of Zn, Cu, Fe, Mn, Ca and Mg in edible seaweeds produced and processed in Galicia.
Key words: metals, continuous acid extraction, ultrasounds, flow injection analysis, edible seaweeds.
INTRODUCCIÓN
Las algas se definen como plantas talofitas, unicelulares o pluricelulares que viven de preferencia en el agua, tanto dulce como marina, y que, en general, están provistas de clorofila acompañada a veces de otros pigmentos de colores variados que la enmascaran. El talo de las pluricelulares tiene forma de filamento, de cinta o de lámina y puede ser ramificado. Aunque hay muchas algas de agua dulce, la mayoría vive en el mar. Dentro de éstas, se encuentran las algas unicelulares, microalgas o fitoplancton, las cuales son microscópicas y representan el primer eslabón de la cadena alimenticia de los animales acuáticos. Además, existen algas pluricelulares o macroalgas, visibles al ojo humano y que normalmente se conocen como algas marinas (Pérez-Cirera López-Niño, 2005). Entre las algas marinas macroscópicas se pueden diferenciar cuatro grupos principales: Cyanophyta o algas azules, Chlorophyta o algas verdes, Rhodophyta o algas rojas y Phaeophyceae o algas pardas. Estos grupos se caracterizan no sólo por los pigmentos que poseen, sino también por la complejidad de su talo y sus mecanismos reproductivos (Otero Schmitt et al., 2002). Sin embargo, tienen la característica común de vivir sin raiz, y extraen su alimento y los nutrientes esenciales para su crecimiento del agua del mar que les rodea. Desde hace mucho tiempo, el

7Revista Real Academia Galega de Ciencias. Vol. XXVII
hombre ha usado las algas con distintos fines para el medio ambiente y el ser humano. Estas aplicaciones son:
- agropecuarias. Mejoran la producción vegetal utilizadas como abono y fertilizante, como corrector del pH de la tierra, e incluso como alimento para el ganado.
- alimenticias. Las algas se han incorporado desde hace muchos siglos en la dieta alimenticia, sobre todo en las culturas orientales y especialmente en Japón, Corea y China, y en la “poliacuicultura ecológica”, las algas sirven de complemento dietético para peces o moluscos de granjas de cultivo.
- cosméticas. Los extractos de algas se emplean en productos para el tratamiento de uñas rotas, acné, arrugas, seborrea, e incluso para la caída del cabello, el rejuvenecimiento de la piel, la obesidad o la celulitis. Asimismo, su capacidad fotoprotectora se está utilizando para el desarrollo de cremas solares.
- farmacológicas. Las algas son muy utilizadas en la medicina tradicional oriental.
- industriales. Para la optención de ficocoloides: agarófitos (E-406), alginatos (E-401 al E-405), y fundamentalmente carrageninas (E-407). El agar se emplea en la elaboración de medio de cultivo en laboratorio por tener un gran poder gelificante. El agar también se añade como aditivo gelificante y espesante (E-406) en gran cantidad de alimentos. Asimismo se emplea en la industria fotográfica y en la industria biotecnológica (agarosa para separaciones cromatográficas y electroforéticas). Las carrageninas son polisacáridos sulfatados muy apreciados por diversos tipos de industrias por sus numerosas aplicaciones, normalmente alimentarias.
- medioambientales y energéticas. Como restauradoras de zonas contaminadas, depuradoras de efluentes o como bioindicadores para conocer el estado de un determinado medio (Chaudhuri et al., 2007; Morrison et al., 2007; Kamala-Kannan et al., 2008). Asimismo, su uso como combustible, para generar biogás (metano), hidrógeno o biodiesel es otra línea fructífera de investigación (Castelló Orvay, 1993).
El consumo directo de algas en la alimentación humana está muy extendido en los países orientales (China, Corea, Japón). Sin embargo, en las

8 Revista Real Academia Galega de Ciencias. Vol. XXVII
últimas décadas, la demanda de algas para la alimentación humana esta en alza en países occidentales (González et al., 1998; Fernández Saa, 2002). Las algas más apreciadas para la cocina son:
- wakame (Undaria pinnatifida): pertenece a las algas pardas y es apreciada en la cultura oriental y muy abundante en las costas de Galicia. Rica en calcio, yodo, proteínas y fibra.
- Espagueti de Mar (Himanthalia elongata): es una alga parda, muy sabrosa y frecuente en litorales profundos y aguas agitadas. Rica en fibra y en hierro.
- Kombu (Laminaria ochroleuca, Laminaria hyperborea, Laminaria saccharina): alga de gran tamaño, color pardo y consistencia carnosa. Su principal característica es que es rica en ácido glutámico que potencia el sabor de los demás alimentos y es rica en minerales, especialmente calcio y magnesio. La Laminaria saccharina tiene un sabor ligeramente dulce y es llamada por ello Kombu de azúcar.
- Dulse (Palmaria palmata): es un alga de color rojo rica en vitamina C, proteínas y potasio.
- Nori (Porphyra umbilicalis): es un alga de pequeño tamaño y muchos pliegues. En Galicia se encuentra silvestre en la bajamar. Tiene un sabor intenso y destaca en proteínas, vitamina A y vitamina B12.
- Osmundea (Osmundea pinnatifida): es un alga muy pequeña, pero con un intenso olor a mar. Muy difícil de coger ya que se encuentra bajo las rocas en las que se crían los percebes.
- Codium (Codium tomentosum).
- Lechuga de mar (Ulva rigida): es un alga verde que recuerda mucho en su apariencia a la lechuga.
- Agar-agar Atlántica, Gelatina de algas (Gelidium sesquipedale). Es la fibra soluble extraída del alga Gelidium sesquipedale. Se suele emplear como espesante y gelificante (E-406) y también muy usada en investigación (microbiología).
- Musgo de Irlanda (Chondrus crispus): Se utiliza en su gran mayoría para la producción industrial de carrageninas, pero también puede utilizarse en cocina como espesante (E-407).

9Revista Real Academia Galega de Ciencias. Vol. XXVII
- Verdello (Enteromorpha Intestinalis): Es una alga verde del orden de las Ulvales.
El análisis composicional de las algas desvela que concentran del agua del mar una amplia variedad de nutrientes esenciales: contenido elevado en minerales esenciales (Ca, Na, K, Mg, Mn, P, Cl, Fe, Cu, Zn, I, etc.) (Robledo y Freile Pelegrin, 1997; Rao et al., 2007). Algunas especies son ricas en proteínas. Presentan mayor cantidad en aminoácidos esenciales que las verduras. Contienen antioxidantes y son extremadamente ricas en polifenoles, carotenoides, vitaminas E y C, clorofila, ácidos grasos esenciales, enzimas y fosfolípidos, que neutralizan los radicales libres y ayudan a retrasar el envejecimiento. Además, contienen vitamina A, vitamina B1, vitamina B2, vitamina B9, vitamina B12 y vitamina K. Presentan un bajo contenido calórico en relación con otros alimentos, y como poseen ácidos grasos poliinsaturados de cadena larga (Omega-3 y Omega-6) ayudan a disminuir y a regular el exceso de colesterol. Los glúcidos que contienen se encuentran casi exclusivamente en forma no asimilable (fibra). Por tanto, aportan poco valor energético. Contienen ácido algínico que es un desintoxicante abundante en las algas pardas laminarias (kombu) que ayuda a prevenir la contaminación de elementos radiactivos en el cuerpo.
El agua de mar contiene, de forma natural, pequeñas concentraciones de metales que no sólo no resultan perjudiciales para el ecosistema sino que son necesarios para el desarrollo de los organismos vivos. El problema surge cuando se produce un aumento de concentración y se convierten en sustancias tóxicas para los organismos marinos o se acumulan en la cadena trófica de forma que pueden convertirse en tóxicos para el hombre. Las macroalgas bioconcentran los iones metálicos libres mediante dos procesos físico-químicos. El primer proceso es rápido y reversible e involucra la adsorción del ión metálico sobre la superficie externa de la pared celular. Este proceso puede ser iónico o por formación de complejos con los ligandos de la pared celular. Los polímeros que componen la pared celular son ricos en grupos carboxílicos, fosforílicos, hidroxilos y aromáticos que pueden ligar cationes o producir complejos orgánicos que pueden influir la absorción de metales. El segundo mecanismo de incorporación de metales es más lento, esta regulado por el metabolismo celular y los metales se almacenan en el citoplasma en vacuolas ricas en polifenoles (Granan et al., 1992). Como producto de esta acumulación las algas pueden alcanzar contenidos de elementos traza varios

10 Revista Real Academia Galega de Ciencias. Vol. XXVII
ordenes de magnitud más elevados respecto a las aguas. Además, hay que tener en cuenta que las distintas especies de algas tienen diferentes afinidades por los metales, por lo que su contenido metálico depende de la clase de alga. La concentración de metales puede variar en las diferentes partes del alga, siendo normalmente mayor en las partes más viejas, ya que han estado más tiempo expuestas al medio. Muchas especies muestran variaciones en la concentración de metales dependiendo de sus periodos de inmersión. Para especies que vivan en el mismo hábitat, el contenido metálico depende de la edad del alga. Otros factores que influyen en la concentración metálica de las algas son el área geográfica, temperatura del agua, pH, etc. (Villares et al., 2002; Yamada et al., 2007).
En los procedimientos analíticos en los que se tiene que eliminar la materia orgánica y/o disolver la muestra, la etapa de pretratamiento de la muestra constituye normalmente el talón de Aquiles del proceso analítico debido, sobre todo, al tiempo que consume su realización. Por eso, la automatización de las operaciones previas del proceso analítico es en la actualidad uno de los objetivos principales de la Química Analítica. De esta manera, la preparación de la muestra es una etapa muy importante en la estrategia del análisis. Por lo que en todo procedimiento analítico es deseable que tenga lugar una mínima manipulación de la muestra, utilizando un pretratamiento lo más sencillo posible, lo cual además, evita problemas de pérdida y contaminación de analito(s). La selección del tipo de pretratamiento de la muestra depende de la técnica analitica que se va a utilizar para la determinación de los metales. Como la espectroscopía de absorción atómica con llama es usualmente la técnica de determinación elegida, ésta requiere normalmente que las muestras sólidas estén disueltas para que la muestra pueda ser introducida en la fuente de atomización. Los métodos clásicos utilizados para la preparación de muestras vegetales implican la digestión por vía seca (calcinación) o la digestión por vía húmeda de las muestras (Knapp et al., 1975; Somenath, 2003; Munilla et al., 1995). Los métodos clásicos haciendo uso de una digestión por vía húmeda en recipientes abiertos son procedimientos laboriosos y tediosos, y además, están sujetos a posibles errores por pérdida de analito(s) y/o contaminación de la muestra. Otro inconveniente de este procedimiento es el manejo de ácidos concentrados y la generación en el proceso de gases tóxicos (óxidos de nitrógeno). De la misma manera, los procesos que implican una calcinación de la muestra también presentan inconvenientes, ya que además de la pérdida de analitos volátiles y la posibilidad de contaminación de la

11Revista Real Academia Galega de Ciencias. Vol. XXVII
muestra, se pueden producir reacciones en las paredes del crisol. Tampoco se deben olvidar las dificultades que se pueden producir al disolver el residuo obtenido en la combustión (Meeravali y Sunil, 2000). Debido a estos problemas, se han propuesto otros procedimientos para la preparación de muestras vegetales (Curdova et al., 1998; Zezzi Arruda, 2006). Así, se han utilizado procedimientos de digestión en vía húmeda realizados en recipientes cerrados y bajo la acción de la energía de microondas. Sin embargo, aunque estos procesos son una buena alternativa para llevar a cabo la preparación de la muestra, ya que reducen el tiempo de pretratamiento en comparación con las metodologías clásicas, siguen presentando el inconveniente del uso de ácidos concentrados. Además, como la muestra se disuelve completamente, aumenta la probabilidad de que existan interferencias de matriz. Debido a estos inconvenientes se han propuesto otras estrategias para realizar la preparación de muestras de origen vegetal. Éstas se basan en la calcinación mediante láser, en extracción con líquidos a presión y en procesos de extracción ácida bajo la acción de la energía de ultrasonidos. Sin embargo, las dos primeras propuestas presentan los siguientes inconvenientes: elevado costo del equipo y dificultad para realizar los procesos de manera automática y miniaturizada (Ahlgren et al., 1988; Saeed et al., 1999; Morales-Muñoz, et al, 2007; Moreda-Piñeiro et al., 2007). Como la extracción ácida de metales asistida por la energía de ultrasonidos no presenta estos inconvenientes, es hoy día una de las alternativas más válidas para realizar el pretratamiento de las muestras sólidas, incluídas las de origen vegetal como las algas marinas (Luque Garcia y Luque de Castro, 2002; Luque de Castro y Priego Capote, 2007).
La energía de ultrasonidos facilita y acelera procesos como la disolución y la extracción. Las muestras sometidas a energía de ultrasonidos se dispersan de tal forma que originan una fina suspensión que se ataca fácilmente por los reactivos (ácidos o bases diluídos). Los procesos de extracción en un baño de ultrasonidos proporcionan una mayor eficacia en la disolución de numerosos analitos de muestras complejas y reducen la duración del proceso en comparación con los métodos clásicos de extracción. Esta metodología proporciona recuperaciones entre el 94-100%, con una buena reproducibilidad para la mayoría de los metales. Los metales son extraídos generalmente con ácidos diluidos, lo que hace que no se disuelva la matriz de la muestra, evitándose de esta forma las interferencias que puede producir en la determinación de el (los) analito(s) (Priego Capote y Luque de Castro, 2004; Luque de Castro y Priego Capote, 2007).

12 Revista Real Academia Galega de Ciencias. Vol. XXVII
En lo que se refiere a la determinación de metales en muestras de vegetales y plantas, se han propuesto varias metodologías basadas en la extracción ácida asistida por energía de ultrasonidos. Así, se han determinado Ba, Ca, Cd, Cu, Mg, Ni, Pb, Sr y Zn en diversos tejidos vegetales y plantas (Lavilla et al., 1998; Filgueiras et al., 2002; Nascentes et al., 2001; Borkowska-Burnecka et al., 2003) y As, Ba, Ca, Na, Mg, Cd, Cr, Cu, Fe, K, Mg, Mn, Ni, Pb, V y Zn en algas (Dominguez-Gonzalez et al., 2005; Ladra-Ramos et al., 2005).
La extracción continua de metales en muestras sólidas usando energía de ultrasonidos llevada a cabo en un dispositivo automático basado en el análisis por inyección en flujo (FIA) ha resultado ser una metodología rápida, sencilla y cuantitativa para el pre-tratamiento de muestras sólidas (Ruiz-Jiménez et al., 2003; Moreno-Cid et al., 2004; Yebra et al., 2005). Otras ventajas que presenta esta metodología en comparación con la alternativa no automática comentada en el apartado anterior son las siguientes: reducción de la contaminación de la muestra y de las pérdidas de analito debido a una mínima manipulación de la muestra, bajo consumo de reactivos y de muestra (el sistema es miniaturizado), disminución considerable del tiempo de tratamiento de la muestra, se evitan las etapas de centrifugación y filtración para la separación de la fase líquida del residuo sólido (matriz de la muestra) e incremento de la calidad de la metodolgía analítica.
Este procedimiento combina los beneficios de la extracción asistida con energía de ultrasonidos con los obtenidos por los sistemas FIA (Valcárcel y Luque de Castro, 1987). Estos métodos automáticos todavía no se han propuesto para la extracción ácida asistida por energía de ultrasonidos de metales y su posterior determinación en muestras de algas, siendo por primera vez objeto de estudio y aplicación en este trabajo de investigación.
JUSTIFICACIÓN DEL TRABAJO
El mar es muchas veces el destino final de los contaminantes generados por los seres humanos. Los metales pesados son uno de estos contaminantes. El problema surge cuando se produce un aumento de concentración y se convierten en sustancias tóxicas para los organismos marinos o acumularse en la cadena trófica de forma que pueden convertirse en tóxicos para el hombre. Los productos marinos, entre los que se encuentran las algas, forman parte hoy día de la dieta humana. Por lo que sería interesante conocer su composición en

13Revista Real Academia Galega de Ciencias. Vol. XXVII
lo que se refiere a metales para establecer su valor nutricional y para determinar su seguridad alimentaria. Las algas se utilizan como bioindicadores de la contaminación ambiental, con lo cual también es importante conocer su contenido metálico para establecer el grado de contaminación por metales del medio ambiente marino de donde proceden. Hasta la fecha no se han propuesto métodos analíticos automáticos para la determinación de metales en muestras de algas.
Todo ello justifica la necesidad de proponer metodologías analíticas automáticas para utilizar de manera rutinaria en laboratorios de control de calidad de alimentos y en laboratorios de control de calidad del medio marino. Estos métodos permitirán realizar de manera rápida y fiable un control riguroso del contenido metálico en organismos marinos como son las algas.
PARTE EXPERIMENTAL
En este trabajo de investigación se ha realizado la determinación de metales en algas marinas destinadas a la alimentación humana producidas y procesadas en Galicia. Para llevar a cabo este estudio se han optimizado dispositivos basados en el análisis por inyección en flujo (FIA) que permiten realizar en línea la extracción ácida asistida por energía de ultrasonidos de los metales de las muestras de algas y la determinación continua de los mismos por FAAS.
Reactivos, aparatos e instrumentación analítica
Ácido nítrico (Merck): 65%, d=1,39 g/mL, Pm= 63,01 g/mol, ácido clorhídrico (Merck): 30%, d=1,15 g/mL, Pm=36,46 g/mol, agua ultrapura de 18,2 MΩ cm de resistividad específica, obtenida por el sistema Milli-Q (Millipore Corp.), disolución patrón de cinc (Merck): 1000 mg/L (Zn (NO3)2 en HNO3 0,5 mol/L), disolución patrón de cobre (Merck): 1000 mg/L (Cu(NO3)2 en HNO3 0,5 mol/L), disolución patrón de hierro (Merck): 1000 mg/L (Fe(NO3)3 en HNO3 0,5 mol/L), disolución patrón de manganeso (Merck): 1000 mg/L (Mn(NO3)2 en HNO3 0,5 mol/L), disolución patrón de calcio (Merck): 1000 mg/L (Ca (NO3)2 en HNO3 0,5 mol/L), disolución patrón de magnesio (Merck): 1000 mg/L (Mg (NO3)2 en HNO3 0,5 mol/L). Cloruro de lantano (III) heptahidratado (LaCl3·7 H2O) (Merck). Agua ultrapura de 18,2 MΩ cm de resistividad específica, obtenida por el sistema Milli-Q (Millipore Corp.).

14 Revista Real Academia Galega de Ciencias. Vol. XXVII
Bombas peristálticas GILSON MINIPULS 3, con ocho canales y selector de velocidad. Minicolumna de vidrio borosilicatado químicamente inerte (Omnifit) de 3 mm de diámetro interno y 100 mm de longitud con dos terminales fijos y 0,70 mL de volumen de lecho. En esta minicolumna se introduce la muestra, colocando, en cada uno de sus extremos, un papel de filtro (Whatman 541) para evitar cualquier obturación de sistema por pérdida de muestra durante el proceso (Figura 1).
Figura 1. Minicolumna conteniendo la muestra.
Ultrasonidos Selecta de 550 watios de potencia con selector de temperatura y tiempo programable. Frecuencia de ultrasonidos 40 kHz. Válvulas de selección y de inyección de tipo hexagonal (Rheodyne). Espectrofotómetro de Absorción Atómica Perkin Elmer, modelo 5000, con llama aire-acetileno y sistema nebulizador de bola de impacto y mechero de 10 cm.
Todo el material de vidrio y plástico fue lavado y almacenado en ácido nítrico al 10% (V/V) al menos durante 48 h. Antes de ser utilizado se enjuagó tres veces con agua ultrapura (Milli-Q).
Preparación de las muestras de algas
Para poder llevar a cabo los análisis de las muestras, éstas han sido trituradas y homogeneizadas. Una vez homogeneizadas, se secaron en una estufa a 80 ± 5 ºC hasta tener las muestras a peso constante. Después se tamizaron para conseguir fracciones de partículas menores de 30 µm y partículas entre 30-200 µm.
Procedimiento para la determinación de cinc, cobre, hierro y manganeso
El esquema de trabajo del método optimizado para la determinación de Zn, Cu, Fe y Mn en algas marinas utilizando el dispositivo FIA

15Revista Real Academia Galega de Ciencias. Vol. XXVII
propuesto (Figura 2) es el siguiente: En una primera etapa se realiza la extracción de Zn, Cu, Fe y Mn. Para ello se inserta con cuidado y se pesa
Figura 2. Dispositivo FIA usado para la determinación de Zn, Cu, Fe y Mn en muestras de algas mostrando los valores óptimos. AU, agua ultrapura (portador); B, blanco; BU, baño de ultrasonidos; B1 y B2, bombas peristálticas; D, desecho; DE, disolución extractante; DP, disolución patrón; FAAS, espectrofotómetro de absorción atómica con llama; M, columna conteniendo la muestra; RM, reactor de mezcla; VI, válvula de inyección; VS1, VS2 y VS3, válvulas de selección.
directamente en la minicolumna la muestra de alga marina seca, triturada y homogeneizada (10, 80, 10 y 25 mg, para Zn, Cu, Fe y Mn, respectivamente). Posteriormente, se colocan dos papeles de filtro en los extremos de la columna para evitar pérdidas de muestra a través del sistema. Una vez preparada la minicolumna, se conecta en el sistema de flujo continuo, se llena el sistema de disolución extractante por medio de la VS1 (1 mL de ácido nítrico 0,1M; 3 M; 3M y 0,5 M para Zn, Cu, Fe y Mn, respectivamente), manteniendo la temperatura del baño a 20 ºC (temperatura ambiente). Después, se somete a la minicolumna a la acción de la energía de ultrasonidos durante 0,5 min para el Zn y el Mn, y 1 min para el Cu y el Fe, a la vez que circula a su través la disolución extractante. El sentido del movimiento de la disolución extractante que circula a través de la columna a 6 mL/min se cambia cada 10-15 s para que no se produzcan compactaciones de la muestra en el interior de la columna. Una vez realizada la extracción de los metales de la muestra, se gira la válvula de selección (VS2) pasando el extracto ácido a la segunda parte del sistema, mientras que la disolución blanco va al desecho. En esta parte, el extracto ácido se homogeneiza en un reactor de mezcla (RM). Tras ser homogeneizado, se inyectan 250 µL de extracto ácido en una corriente portadora (agua ultrapura) a través de una válvula de inyección (VI). Esta

16 Revista Real Academia Galega de Ciencias. Vol. XXVII
disolución portadora que circula a un caudal de 3,5 mL/min, transporta el bolo de muestra hasta el nebulizador de un espectrofotómetro de absorción atómica con llama (FAAS), donde se detectan los metales de forma continua. Para realizar las rectas de calibrado se introducen secuencialmente en el sistema FIA disoluciones patrón de distintas concentraciones de cada metal (preparadas en ácido nítrico 0,1 M para el Zn, 3 M para el Cu y el Fe, y 0,5 M para el Mn). El volumen inyectado de cada disolución patrón a través de la válvula de inyección (VI) en la disolución portadora es de 250 mL (igual que el de la muestra). La disolución blanco es ácido nítrico 0,1 M para el Zn, 3 M para el Cu y el Fe, y 0,5 M para el Mn.
Procedimiento para la determinación de calcio y magnesio
El esquema de trabajo del método optimizado para la determinación de Ca y Mg en algas marinas utilizando un dispositivo FIA (Figura 3) es el siguiente: Primero se
Figura 3. Dispositivo FIA usado para la determinación de Ca y Mg en muestras de algas mostrando los valores óptimos. AU, agua ultrapura (portador); B, blanco; BU, baño de ultrasonidos; B1 y B2, bombas peristálticas; D,desecho; DE, disolución extractante; DIL, canal de dilución (disolución de lantano); DP, disolución patrón; FAAS, espectrofotómetro de absorción atómica con llama; M, columna conteniendo la muestra; RM, reactor de mezcla; VI, válvula de inyección; VS1, VS2 y VS3, válvulas de selección.
realiza la extracción de Ca y Mg. Para ello se inserta con cuidado y se pesa directamente en la minicolumna la muestra de alga marina seca, triturada y homogeneizada (30 mg para la determinación de Ca y 5 mg para la determinación de Mg). Después, se colocan dos papeles de filtro en los extremos de la columna para evitar pérdidas de muestra a través del sistema.

17Revista Real Academia Galega de Ciencias. Vol. XXVII
Una vez preparada la minicolumna, se conecta en el sistema de flujo continuo, se llena el sistema de disolución extractante por medio de la VS1 (5 mL de ácido nítrico 3 M para Ca y 0,5 M para Mg), manteniendo la temperatura del baño a 20 ºC (temperatura ambiente). Después, se somete a la minicolumna a la acción de la energía de ultrasonidos durante 0,5 min, a la vez que circula a su través la disolución extractante. El sentido del movimiento de la disolución extractante que circula a través de la columna a 6 mL/min se cambia cada 10-15 s para que no se produzcan compactaciones de la muestra en el interior de la columna. Una vez realizada la extracción de los metales de la muestra, se gira la válvula de selección (VS2) pasando el extracto ácido a la segunda parte del sistema, mientras que la disolución blanco va al desecho. En esta parte, el extracto ácido se homogeneiza en un reactor de mezcla (RM). Tras ser homogeneizado, se inyectan 250 µL de extracto ácido en una corriente portadora (agua ultrapura) a través de una válvula de inyección (VI). Esta disolución portadora que circula a un caudal de 0,2 mL/min converge con un canal de dilución (lantano 2%) que circula a 5,8 mL/min. El canal resultante que contiene el extracto ácido diluido llega directamente hasta el nebulizador de un espectrofotómetro de absorción atómica con llama (FAAS), donde se detectan los metales de forma continua. Para realizar las rectas de calibrado se introducen secuencialmente en el sistema FIA 2 disoluciones patrón de distintas concentraciones de cada metal (preparadas en ácido nítrico 3 M para Ca y 0,5 M para Mg) a través de la válvula de inyección (VI). El volumen inyectado de cada disolución patrón en la disolución portadora es de 250 mL (igual que el de la muestra). Las distintas disoluciones patrón sufren la misma dilución en
Tabla 1. Determinación de Zn, Cu, Fe y Mn. Variables y nivelesconsiderados para el diseño factorial de screening.
línea que las muestras. La disolución blanco es ácido nítrico 3 M para el Ca, y 0,5 M para el Mg.

18 Revista Real Academia Galega de Ciencias. Vol. XXVII
RESULTADOS Y DISCUSIÓN
Determinación de cinc, cobre, hierro y manganeso
Para realizar la optimización de los parámetros que afectan a la extracción de Zn, Cu, Fe y Mn de las muestras de algas, se ha utilizado un diseño factorial debido a que el número de variables implicadas es elevado (6 variables). Así, es necesario aplicar un diseño factorial de screening, para reducir de modo significativo el número de experimentos a realizar en la optimización. Uno de los diseños de screening más utilizados es el diseño de Plackett-Burman. Éste permite fraccionar un diseño factorial completo proporcionando números de combinaciones de factores que son un múltiplo de cuatro. Por ejemplo, si se aplicara un diseño factorial completo a 2 niveles para 6 variables implicaría la realización de 64 experimentos. Sin embargo, aplicando el diseño de Plackett-Burman con un punto central, el número de experimentos a realizar es de trece. Así, las variables a optimizar para el dispositivo FIA 1 son las siguientes: concentración de HNO3 (disolución extractante), concentración de HCl (disolución extractante), tiempo de extracción,
Tabla 2. Determinación de Zn, Cu, Fe y Mn. Concentración certificada y cantidad de muestra analizada del material de referencia “Sargasso”, NIES-9, National Institute for Environmental Studies, Japan.
temperatura de extracción, caudal de extracción y volumen de disolución extractante. Para poder realizar este diseño factorial es necesario considerar para cada variable dos niveles (valor máximo y mínimo). Estos valores fueron seleccionados en función de experimentos realizados en estudios previos (Tabla 1).
El diseño factorial aplicado es el de Plackett-Burman 2^6*3/16 que implica realizar 13 experimentos (Statgraphics Plus v.5.1). En este diseño se ha

19Revista Real Academia Galega de Ciencias. Vol. XXVII
considerado un punto central en el que las variables toman un valor medio entre el límite superior e inferior. Una vez obtenida la matriz del diseño, se realizaron los experimentos fijados por la misma. Para realizar estos experimentos se utilizó un material de referencia certificado (NIES-9,”Sargasso”, National Institute for Environmental Studies, Japan). La concentración certificada para Zn, Cu, Fe y Mn, y la cantidad de material de referencia utilizada (fijada en función de la concentración) para optimizar el proceso se muestran en la Tabla 2.
La variable respuesta considerada ha sido el % eficacia en la extracción (% EF), calculado según la siguiente ecuación:
La matriz del diseño de Plackett-Burman con los valores que toma cada variable en cada experimento y el % eficacia en la extracción
Tabla 3. Matriz del diseño de Plackett-Burman con los valores de % EF para cada variable en cada experimento para la determinación de Zn, Cu, Fe y Mn. A, [HNO3]; B, [HCl]; C, Tiempo de extracción; D, Temperatura de extracción; E, Caudal de extracción; F, Volumen de disolución extractante.

20 Revista Real Academia Galega de Ciencias. Vol. XXVII
obtenido para el Zn, Cu, Fe y Mn, se recogen en la Tabla 3. Los experimentos se han realizado por duplicado, por lo que el % eficacia en la extracción indicada es la media de estos dos experimentos.
Una vez realizados los experimentos, se procede al análisis de los datos. Para conocer que variables son significativas se recurre a las Cartas Pareto estandarizadas (Figura 4).
Figura 4. Cartas Pareto estandarizadas para la determinación de Zn, Cu, Fe y Mn. A, [HNO3]; B, [HCl]; C, Tiempo de extracción; D, Temperatura de extracción; E, Caudal de extracción; F, Volumen de disolución extractante.
A partir de la observación de las cartas Pareto (Figura 4) se pueden establecer las siguientes conclusiones en función de cada variable:
Concentración de ácido nítrico (A): En todos los casos la concentración de HNO3 es una variable estadísticamente significativa con efecto estimado positivo. Esto se debe a su efecto oxidante, el cual facilita el proceso de extracción.

21Revista Real Academia Galega de Ciencias. Vol. XXVII
Concentración de ácido clorhídrico (B): No es una variable estadísticamente significativa. Aunque presenta un efecto estimado positivo para la extracción de Zn, Cu, Fe y Mn.
Tiempo de extracción (C): No es una variable estadísticamente significativa. Aunque presenta un efecto estimado positivo para la extracción de Zn, Cu, Fe y Mn, ya que cuanto más tiempo permanezca la muestra bajo la acción de la energía de ultrasonidos mayor contacto se produce entre la muestra y la disolución extractante, y por tanto se logra una mayor eficacia en la extracción. Sin embargo, tiempos de sonicación elevados suponen elevados tiempos de análisis y por lo tanto una reducción de la frecuencia de muestreo.
Temperatura de extracción (D): No es una variable estadísticamente significativa, y presenta efectos estimados muy pequeños en todos los casos. Esto indica que esta variable prácticamente no influye en el proceso de extracción.
Caudal de extracción (E): Para ninguno de los cuatro metales es una variable estadísticamente significativa. Aunque presenta un efecto estimado positivo.
Volumen de disolución extractante (F): En ninguno de los casos es una variable estadísticamente significativa y siempre presenta un efecto estimado positivo, aunque éste no es muy elevado.
Como el diseño de Plackett-Burman proporciona las tendencias de cada una de las variables hacia el óptimo, y no el valor optimo en sí mismo, se realizaron una serie de experimentos fuera del marco del diseño, de acuerdo con los resultados obtenidos por el mismo, para determinar los valores óptimos de cada una de las variables que afectan a la extracción de Zn, Cu, Fe y Mn. Comenzando por aquellas variables no significativas se comprueba que un volumen de disolución extractante de 1 mL y una temperatura de extracción de 20ºC pueden ser los valores óptimos para estas variables, ya que con ellos se obtienen eficacias en la extracción cuantitativas (experimentos 1 y 3 de la matriz de Plackett-Burman, Tabla 3).
En el estudio de las variables tiempo de extracción y caudal de extracción (Figura 5) se observa que, a pesar de que el tiempo de extracción tiene un efecto estimado bastante positivo, son suficientes 0,5, 1, 1 y 0,5 min a un

22 Revista Real Academia Galega de Ciencias. Vol. XXVII
caudal de 6 mL/min para extraer cuantitativamente el Zn, Cu, Fe y Mn en este tipo de muestras. Así, éstos valores serán elegidos como óptimos para estas variables, ya que con ellos se alcanza la frecuencia de muestreo más elevada
Figura 5. Optimización del tiempo de extracción para Zn, Cu, Fe y Mn para caudales de extracción de 3,5 y 6 mL/min. [HNO3]=3 M; [HCl]=3 M; Temperatura de extracción= 20 ºC; Volumen de disolución extractante= 1 mL.
Con respecto a las concentraciones de los ácidos, se estudió la posibilidad de eliminar el ácido clorhídrico, ya que su concentración resultó ser una variable no estadísticamente significativa en el diseño de screening. Así, los estudios realizados deteminaron que si se elimina de la disolución extractante este ácido (utilizando únicamente HNO3 3M como disolución extractante) se obtienen eficacias en la extracción cuantitativas (95,3-100,5%), lo cual también se puede corroborar con los experimentos 1 y 4 de la matriz de Plackett-Burman, Tabla 3). Una vez comprobado que se podía eliminar el ácido clorhídrico de la disolución extractante, se estudió la posibilidad de disminuir la concentración de HNO3 al máximo. Como se observa en la Figura 6, fue posible disminuir la concentración de este ácido hasta un valor de 0,1 M para la extracción del Zn y 0,5 M para la extracción del Mn, mientras que para la extracción de Cu y Fe la mínima concentración de ácido nítrico necesaria para extraer cuantitativamente estos metales debía ser de 3M.

23Revista Real Academia Galega de Ciencias. Vol. XXVII
Figura 6. Optimización de la concentración de ácido nítrico utilizado como disolución extractante para Zn, Cu, Fe y Mn. [HCl]=0 M; Tiempo de extracción= 0,5 min para Zn, y Mn y 1 min para Cu y Fe; Temperatura de extracción= 20 ºC; Caudal de extracción= 6 mL/min; Volumen de disolución extractante= 1 mL.
Al realizar la optimización de las distintas variables que afectan al proceso de extracción ácida se han utilizado las cantidades de muestra indicadas en la Tabla 2. No obstante, también se ha investigado la posibilidad de analizar cantidades de muestra mayores con el objetivo de incrementar la sensibilidad de la metodología analítica desarrollada. Para realizar este estudio se ha tenido en cuenta la sensibilidad de cada uno de los metales en el espectrofotómetro de absorción atómica con llama y la concentración normalmente existente en las muestras de algas. Así, los resultados obtenidos en este estudio para Zn, Cu, Fe y Mn demostraron que con las condiciones óptimas es posible utilizar cantidades de muestra hasta 60, 100, 25 y 90 mg, para Zn, Cu, Fe y Mn, respectivamente. Cantidades mayores no se probaron porque esto requeriría incorporar un canal de dilución en el dispositivo para no sobrepasar el límite de linealidad del detector.
Los estudios anteriores se realizaron con el material de referencia certificado NIES-9, el cual presenta un diámetro de partícula de 80 mesh (177 µm). Por tanto, con el objetivo de estudiar si esta variable influye en el proceso de extracción de Zn, Cu, Fe y Mn de las muestras de algas se analizaron diferentes porciones de la misma muestra con diferentes tamaños de partícula. Los diámetros de partícula estudiados fueron dos: 1) partículas de muestra con diámetros menores de 30 µm y 2) partículas de muestra con diámetros entre 30-200 µm. Para realizar el estudio de la influencia del tamaño de partícula, se tomó una muestra real de alga, de la que se seleccionaron
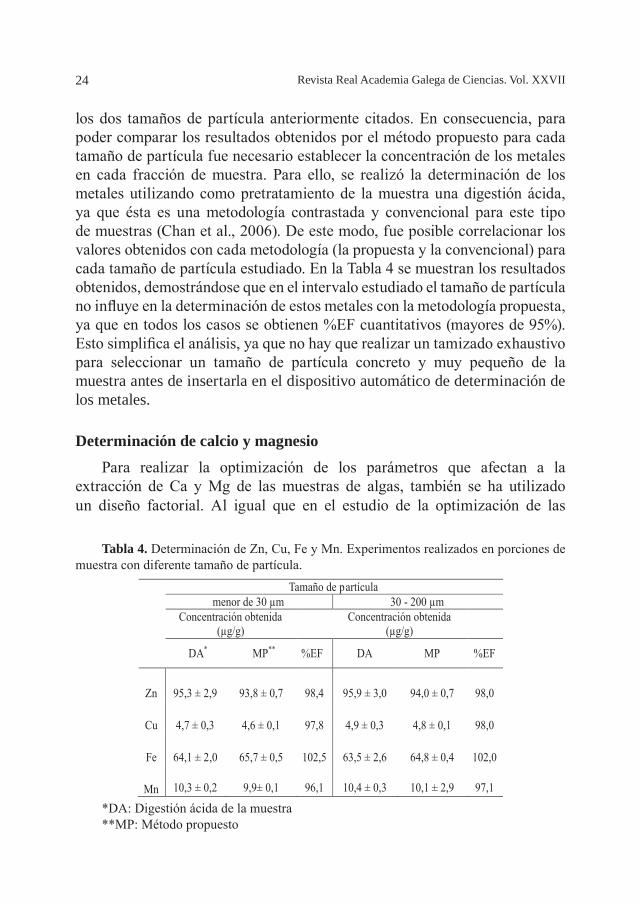
24 Revista Real Academia Galega de Ciencias. Vol. XXVII
los dos tamaños de partícula anteriormente citados. En consecuencia, para poder comparar los resultados obtenidos por el método propuesto para cada tamaño de partícula fue necesario establecer la concentración de los metales en cada fracción de muestra. Para ello, se realizó la determinación de los metales utilizando como pretratamiento de la muestra una digestión ácida, ya que ésta es una metodología contrastada y convencional para este tipo de muestras (Chan et al., 2006). De este modo, fue posible correlacionar los valores obtenidos con cada metodología (la propuesta y la convencional) para cada tamaño de partícula estudiado. En la Tabla 4 se muestran los resultados obtenidos, demostrándose que en el intervalo estudiado el tamaño de partícula no influye en la determinación de estos metales con la metodología propuesta, ya que en todos los casos se obtienen %EF cuantitativos (mayores de 95%). Esto simplifica el análisis, ya que no hay que realizar un tamizado exhaustivo para seleccionar un tamaño de partícula concreto y muy pequeño de la muestra antes de insertarla en el dispositivo automático de determinación de los metales.
Determinación de calcio y magnesioPara realizar la optimización de los parámetros que afectan a la
extracción de Ca y Mg de las muestras de algas, también se ha utilizado un diseño factorial. Al igual que en el estudio de la optimización de las
Tabla 4. Determinación de Zn, Cu, Fe y Mn. Experimentos realizados en porciones de muestra con diferente tamaño de partícula.
*DA: Digestión ácida de la muestra**MP: Método propuesto

25Revista Real Academia Galega de Ciencias. Vol. XXVII
variables implicadas en la extracción de Zn, Cu, Fe y Mn, se ha elegido como diseño de screening el diseño de Plackett-Burman. En este caso, las variables a optimizar para el dispositivo FIA (Figura 3) son las siguientes: concentración de HNO3 (disolución extractante), concentración de HCl (disolución extractante), tiempo de extracción, temperatura de extracción, caudal de extracción, volumen de disolución extractante y concentración de la disolución de lantano. Como se puede observar, se ha incluido como variable la concentración de lantano, a pesar de no ser una variable implicada directamente en el proceso de extracción. Esto se explica porque para poder realizar la determinación tanto del calcio como del magnesio, es necesario eliminar las posibles interferencias químicas que sufren estos dos metales en la celda de atomización. Además, el extracto ácido debe diluirse, ya que las concentraciones de estos dos metales presentes en las muestras objeto de estudio son muy elevadas. De hecho, según el límite de linealidad de estos metales, 5 y 0,5 µg/mL, para Ca y Mg, respectivamente, y teniendo en cuenta las concentraciones a las que se pueden encontrar estos metales en las algas marinas, se escogieron respectivamente, como caudal de portador y caudal de dilución 0,2 y 5,8 mL/min, lo que supone realizar en línea una dilución 1:30. Para realizar el diseño de screening es necesario considerar para cada variable un valor máximo y uno mínimo. Estos valores fueron seleccionados en función de experimentos realizados en estudios previos (Tabla 5).
Tabla 5. Determinación de Ca y Mg. Variables y niveles considerados para el diseño factorial de screening.

26 Revista Real Academia Galega de Ciencias. Vol. XXVII
El diseño factorial aplicado es el de Plackett-Burman 2^7*3/32 que implica realizar 13 experimentos (considerando un punto central) (Statgraphics Plus v.5.1). Para realizar los experimentos establecidos por la matriz del diseño se utilizó el material de referencia certificado (NIES-9,”Sargasso”, National Institute for Environmental Studies, Japan). La concentración certificada es de 1,34 ± 0,05 y 0,65 ± 0,03 %, para Ca y Mg, respectivamente. La cantidad de material de referencia utilizada para optimizar el proceso (fijada en función de la concentración) fue de 30 y 5 mg, para Ca y Mg, respectivamente. La variable respuesta considerada ha sido el % EF. La matriz del diseño de Plackett-Burman con los valores que toma cada variable en cada experimento y el % EF obtenido para el Ca y el Mg, se muestran en la Tabla 6. Los experimentos se han realizado
Tabla 6. Matriz del diseño de Plackett-Burman con los valores de % EF para cada variable en cada experimento para la determinación de Ca y Mg. A, [HNO3]; B, [HCl]; C, Tiempo de extracción; D, Temperatura de extracción; E, Caudal de extracción; F, Volumen de disolución extractante; G, concentración de la disolución de lantano.
por duplicado, por lo que el % EF indicada es la media de estos dos experimentos.
Una vez realizados los experimentos, se analizaron los datos mediante las Cartas Pareto estandarizadas (Figura 7).

27Revista Real Academia Galega de Ciencias. Vol. XXVII
Figura 7. Cartas Pareto estandarizadas para la determinación de Ca y Mg. A, [HNO3]; B, [HCl]; C, Tiempo de extracción; D, Temperatura de extracción; E, Caudal de extracción; F, Volumen de disolución extractante; G, Concentración de la disolución de lantano.
A partir del estudio de las cartas Pareto (Figura 7) se pueden establecer las siguientes conclusiones en función de cada variable:
Concentración de ácido nítrico (A): Para Ca y Mg la concentración de HNO3 es una variable estadísticamente significativa con efecto estimado positivo.
Concentración de ácido clorhídrico (B): No es una variable estadísticamente significativa, aunque presenta un efecto estimado positivo para la extracción de Ca y Mg.
Tiempo de extracción (C): No es una variable estadísticamente significativa. Aunque presenta un efecto estimado positivo para la extracción de Ca y Mg.
Temperatura de extracción (D): No es una variable estadísticamente significativa, y presenta efectos estimados muy pequeños para la extracción de Ca y Mg. Esto indica que esta variable prácticamente no influye en el proceso de extracción.
Caudal de extracción (E): No es una variable estadísticamente significativa para la extracción de estos dos metales. Aunque presenta un efecto estimado positivo.
Volumen de disolución extractante (F): En ninguno de los casos es una variable estadísticamente significativa y siempre presenta un efecto estimado positivo, aunque éste no es muy elevado.

28 Revista Real Academia Galega de Ciencias. Vol. XXVII
Concentración de la disolución de lantano (G): Para Ca y Mg la concentración de la disolución de lantano es una variable estadísticamente significativa con efecto estimado positivo. Esto demuestra que es necesaria esta disolución para eliminar las interferencias producidas por la formación de compuestos estables a la temperatura de llama.
Se realizaron una serie de experimentos fuera del marco del diseño, de acuerdo con los resultados obtenidos por el mismo, para determinar los valores óptimos de cada una de las variables que afectan a la extracción de Ca y Mg. Se comenzó por aquellas variables no significativas y se comprobó que un volumen de disolución extractante de 5 mL y una temperatura de extracción de 20ºC pueden ser los valores óptimos para estas variables, ya que con ellos se obtienen eficacias en la extracción cuantitativas (experimento 1 de la matriz de Plackett-Burman, Tabla 6). Si se tiene en cuenta que el caudal de extracción presenta un efecto estimado positivo y que en los experimentos 5 y 11 de la matriz de Plackett-Burman (Tabla 6) se consiguen eficacias en la extracción cuantitativas, un caudal de extracción de 6 mL/min podía ser considerado como óptimo para la extracción de Ca y Mg.
Al estudiar el tiempo de extracción (Figura 8) se puede ver que aunque esta variable presenta un efecto estimado bastante importante y positivo, son suficientes 0,5 min para extraer cuantitativamente el Ca y el Mg en este tipo de muestras.
Figura 8. Optimización del tiempo de extracción para Ca y Mg. [HNO3]=3 M; [HCl]=3 M; Temperatura de extracción= 20 ºC; Caudal de extracción= 6 mL/min; Volumen de disolución extractante= 1 mL; Concentración de la disolución de lantano= 2%.

29Revista Real Academia Galega de Ciencias. Vol. XXVII
Además, se estudió la posibilidad de eliminar el ácido clorhídrico de la disolución extractante, ya que su concentración resultó ser una variable no estadísticamente significativa en el diseño de screening. Así, los estudios realizados deteminaron que si se elimina se obtienen eficacias en la extracción cuantitativas (98,7-101,7%), lo cual también se puede corroborar con los experimentos 1 y 11 de la matriz de Plackett-Burman, Tabla 6). Una vez comprobado que se podía eliminar el ácido clorhídrico de la disolución extractante, se estudió la posibilidad de disminuir la concentración de ácido nítrico al máximo. Como se observa en la Figura 9, fue posible disminuir la concentración de este ácido hasta un valor de 0,5 M para la extracción del Mg, mientras que para la extracción de Ca la mínima concentración de ácido nítrico necesaria para extraer cuantitativamente este metal debía ser de 3M.
Figura 9. Optimización de la concentración de ácido nítrico utilizado como disolución extractante para Ca y Mg. [HCl]=0 M; Tiempo de extracción= 0,5 min; Temperatura de extracción= 20 ºC; Caudal de extracción= 6 mL/min; Volumen de disolución extractante= 5 mL; Concentración de la disolución de lantano= 2%.
Por último, se estudió la variable concentración de la disolución de lantano. Así, se comprobó que para ambos metales, la concentración de la disolución de lantano, necesaria para eliminar las interferencias, no podía ser menor del 2%.
Al realizar la optimización de las distintas variables que afectan al proceso de extracción ácida se han utilizado las cantidades de muestra antes indicadas.

30 Revista Real Academia Galega de Ciencias. Vol. XXVII
En este caso, también se ha investigado la posibilidad de analizar cantidades de muestra mayores con el objetivo de incrementar la sensibilidad de la metodología analítica desarrollada. Para realizar este estudio se ha tenido en cuenta la sensibilidad de ambos metales en el espectrofotómetro de absorción atómica con llama y la concentración normalmente existente en las muestras de algas. Los resultados obtenidos en este estudio demostraron que con las condiciones óptimas establecidas es posible utilizar cantidades de muestra hasta 50 y 10 mg, para Ca y Mg, respectivamente. Cantidades mayores no se probaron porque esto requeriría aumentar el caudal del canal de dilución del dispositivo FIA para no sobrepasar el límite de linealidad del detector.
Se analizaron diferentes porciones de la misma muestra de alga con diferentes tamaños de partícula (menor de 30 y entre 30 - 200 µm) con el objetivo de estudiar si la variable tamaño de partícula influía en el proceso de extracción de Ca y Mg de las muestras de algas. En la Tabla 7 se muestran los resultados obtenidos, demostrándose que en el intervalo estudiado el tamaño de partícula no influye en la determinación de estos metales con la metodología propuesta, ya que en todos los casos se obtienen %EF cuantitativos.
Tabla 7. Determinación de Ca y Mg. Experimentos realizados en porciones de muestra con diferente tamaño de partícula.
*DA: Digestión ácida de la muestra**MP: Método propuesto
Características analíticas de los métodos propuestos para la determinación de Zn, Cu, Fe y Mn en muestras de algas
Con el objetivo de estudiar las posibles interferencias que puede provocar la matriz de la muestra, es decir la influencia de los demás componentes de

31Revista Real Academia Galega de Ciencias. Vol. XXVII
las algas, se comparó para cada uno de los casos la pendiente de la recta de calibrado con la obtenida para la recta de adición. Los resultados obtenidos aparecen en la Figura 10. Como se puede observar en esta Figura, en todos los casos la recta de calibrado y la recta de adición son paralelas. Dicho paralelismo se corroboró mediante la aplicación de un test estadístico t para un nivel de significación del 95%. Por todo ello, se puede decir que no existe efecto matriz para la determinación de Zn, Cu, Fe y Mn en las muestras de algas por el método propuesto. Lo que se traduce en que se puede utilizar únicamente la recta de calibrado correspondiente para relacionar la señal de la muestra con la concentración existente de cada metal.
Figura 10. Rectas de calibrado y adición para Zn, Cu, Fe y Mn.
Se calculó el límite de detección (LOD), el límite de cuantificación (LOQ) y el intervalo lineal teniendo en cuenta distintas cantidades de muestra para la determinación de Zn, Cu, Fe y Mn en muestras de algas por el método propuesto. Los resultados obtenidos para cada uno de estos parámetros se muestran en la Tabla 8.
La comprobación de la exactitud de la metodología analítica propuesta se realizó de tres maneras: realizando el test de pares de valores (comparación con

32 Revista Real Academia Galega de Ciencias. Vol. XXVII
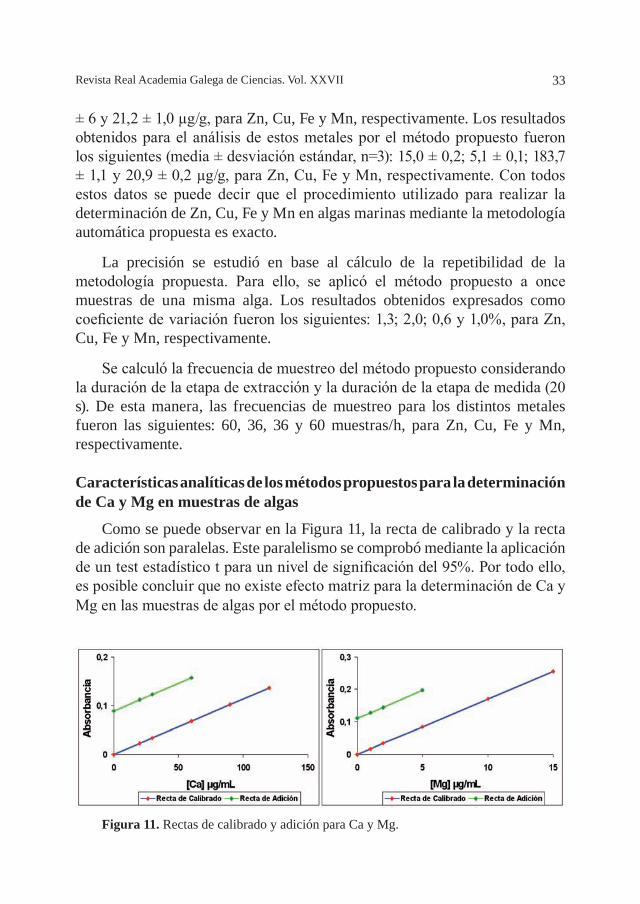
un método convencional de pretratamiento de la muestra realizando la digestión de la misma con ácidos concentrados), estudio de la recuperación analítica y mediante la utilización de un material de referencia. Los valores obtenidos en el test de pares de valores sobre diferentes muestras resultaron comparables estadísticamente al 95% de probabilidad, es decir, no hay diferencias entre los resultados obtenidos por los dos métodos. El estudio de la recuperación del método se realizó analizando una muestra real de alga. En el análisis de esta muestra se utilizaron disoluciones extractantes (HNO3 0,1M para Zn, HNO3 3 M para Cu y Fe, y HNO3 0,5 M para Mn) a las que se le habían adicionado distintas concentraciones de cada metal (0,1; 1,0; 1,0 y 0,5, µg/mL, para Zn, Cu, Fe y Mn, respectivamente). Los resultados obtenidos fueron 98,7, 96,8, 101,3 y 99,3%, para Zn, Cu, Fe y Mn, respectivamente. Se analizó un material de referencia (NIES-9,”Sargasso”, National Institute for Environmental Studies, Japan), de
Tabla 8. LOD, LOQ e intervalo lineal para la determinación de Zn, Cu, Fe y Zn en algas para distintas cantidades de muestra.
composición elemental típica de un alga marina parda. El contenido certificado, para los metales en este material de referencia es: 15,6 ± 1,2; 4,9 ± 0,2; 187
33Revista Real Academia Galega de Ciencias. Vol. XXVII
± 6 y 21,2 ± 1,0 µg/g, para Zn, Cu, Fe y Mn, respectivamente. Los resultados obtenidos para el análisis de estos metales por el método propuesto fueron los siguientes (media ± desviación estándar, n=3): 15,0 ± 0,2; 5,1 ± 0,1; 183,7 ± 1,1 y 20,9 ± 0,2 µg/g, para Zn, Cu, Fe y Mn, respectivamente. Con todos estos datos se puede decir que el procedimiento utilizado para realizar la determinación de Zn, Cu, Fe y Mn en algas marinas mediante la metodología automática propuesta es exacto.
La precisión se estudió en base al cálculo de la repetibilidad de la metodología propuesta. Para ello, se aplicó el método propuesto a once muestras de una misma alga. Los resultados obtenidos expresados como coeficiente de variación fueron los siguientes: 1,3; 2,0; 0,6 y 1,0%, para Zn, Cu, Fe y Mn, respectivamente.
Se calculó la frecuencia de muestreo del método propuesto considerando la duración de la etapa de extracción y la duración de la etapa de medida (20 s). De esta manera, las frecuencias de muestreo para los distintos metales fueron las siguientes: 60, 36, 36 y 60 muestras/h, para Zn, Cu, Fe y Mn, respectivamente.
Características analíticas de los métodos propuestos para la determinación de Ca y Mg en muestras de algas
Como se puede observar en la Figura 11, la recta de calibrado y la recta de adición son paralelas. Este paralelismo se comprobó mediante la aplicación de un test estadístico t para un nivel de significación del 95%. Por todo ello, es posible concluir que no existe efecto matriz para la determinación de Ca y Mg en las muestras de algas por el método propuesto.
Figura 11. Rectas de calibrado y adición para Ca y Mg.

34 Revista Real Academia Galega de Ciencias. Vol. XXVII
Se calculó el límite de detección (LOD), el límite de cuantificación (LOQ) y el intervalo lineal teniendo en cuenta distintas cantidades de muestra para la determinación de Ca y Mg en muestras de algas por el método propuesto. Los resultados obtenidos para cada uno de estos parámetros se muestran en la Tabla 9.
La comprobación de la exactitud de la metodología analítica propuesta se realizó mediante el estudio de la recuperación analítica y analizando un material de referencia certificado. El estudio de la recuperación del método se realizó analizando una muestra real de alga. En el análisis de esta muestra se utilizaron disoluciones extractantes (HNO3 3 M para Ca y HNO3 0,5 M para Mg) a las que se le habían adicionado distintas concentraciones de cada metal (20 y 1 µg/mL, para Ca y Mg, respectivamente). Los resultados obtenidos para la recuperación analítica fueron 98,7 y 101,0%, para Ca y Mg, respectivamente). Lo que significa
Tabla 9. LOD, LOQ e intervalo lineal para la determinación de Ca y Mg en algas para distintas cantidades de muestra.
que se obtiene una recuperación cuantitativa en ambos casos. Además, se analizó un material de referencia (NIES-9,”Sargasso”, National Institute for Environmental Studies, Japan). El contenido certificado, para Ca y Mg en este material de referencia es 1,34 ± 0,05 y 0,65 ± 0,03%, para Ca y Mg, respectivamente. Los resultados obtenidos para la determinación de estos metales en el material de referencia por la metodología propuesta

35Revista Real Academia Galega de Ciencias. Vol. XXVII
(media ± desviación estándar, n=3) fueron 1,33 ± 0,02 y 0,66 ± 0,01%, para Ca y Mg, respectivamente.
La precisión se estudió en base al cálculo de la repetibilidad de la metodología propuesta. Para ello, se aplicó el método propuesto a once muestras de una misma alga. Los resultados obtenidos para el estudio de la precisión expresados como coeficiente de variación estaban en el intervalo 0,9-1,4%, demostrándose la precisión de la metodología analítica propuesta.
Se calculó la frecuencia de muestreo del método propuesto considerando la duración de la etapa de extracción (0,5 min) y la duración de la etapa de medida (25 s). Así, la frecuencia de muestreo para Ca y Mg resultó ser de 55 muestras/h.
APLICACIÓN DE LAS METODOLOGÍAS ANALÍTICAS DESARROLLADAS. DETERMINACIÓN DE ZN, CU, FE, MN, CA Y MG EN ALGAS MARINAS DESTINADAS A LA ALIMENTACIÓN HUMANA PRODUCIDAS Y PROCESADAS EN GALICIA
Los métodos propuestos fueron aplicados para la determinación de de Zn, Cu, Fe, Mn, Ca y Mg en algas marinas destinadas a la alimentación humana producidas y procesadas en Galicia. Las muestras analizadas se adquirieron en supermercados y tiendas de alimentación. Estas algas son las siguientes:
Algas rojas: Agar-agar Atlántica (Gelidium sesquipedale), Dulse (Palmaria palmata), Musgo de Irlanda (Chondrus crispus) y Nori (Porphyra umbilicalis).
Algas pardas: Wakame (Undaria pinnatifida), Espagueti de Mar (Himanthalia elongata), Kombu (Laminaria ochroleuca) y Kombu de azúcar (Laminaria saccharina).
Algas verdes: Lechuga de mar (Ulva rigida).Los resultados obtenidos se muestran en la Tabla 10.
CONCLUSIONES
Las ventajas que presenta la metodología automática propuesta frente a los métodos analíticos utilizados hasta el momento para realizar la determinación de metales en muestras de algas son las siguientes:

36 Revista Real Academia Galega de Ciencias. Vol. XXVII
Tabla 10. Resultados obtenidos para las muestras de algas analizadas: concentraciones de Zn, Cu, Fe, Mn, Ca y Mg. Media ± desviación estándar (n= 3 determinaciones).
* Concentraciones expresadas en % (g metal /g muestra) x 100
- Se utiliza una cantidad de muestra menor: 5-160 mg frente a 0,2-0,5 g.
- El modo dinámico del los sistemas y la reducción del tamaño de muestra hacen que se pueda reducir el tiempo necesario para realizar cuantitativamente la extracción de los metales: 0,5-2 min frente a 30-35 min.
- La extracción tiene lugar con ácidos diluidos (0,1-3 M HNO3) y a temperatura ambiente.
- Debido a la miniaturización, se reduce el volumen de reactivos, lo cual además, en el caso del volumen de la disolución extractante, incrementa la sensibilidad del sistema.
- Se evita la contaminación de la muestra y las pérdidas de analito debido a una mínima manipulación de la muestra.
- Se evitan las etapas de centrifugación y filtración para la separación de la fase líquida del residuo sólido (matriz de la muestra). Este punto con el punto nº 2 hacen que se produzca una disminución considerable del tiempo de tratamiento de la muestra.
- Se produce un incremento en la calidad de la metodolgía analítica inherente a la metodología FIA utilizada.

37Revista Real Academia Galega de Ciencias. Vol. XXVII
Los metales esenciales, también denominados biometales, son los que están presentes en los organismos vivientes y ejercen una función biológica insustituible. Un déficit en el organismo de un metal esencial conduce a serias alteraciones biológicas. Actualmente la Organización Mundial de la Salud considera entre otros metales esenciales para la salud humana a Zn, Cu, Fe, Mn, Ca y Mg. En las algas estudiadas se encuentra un contenido importante de metales, lo que representa un alto valor nutritivo para este alimento. Según los datos obtenidos, las algas más ricas en Zn son Nori y Espagueti de mar, el alga más rica en cobre es la Dulse, las algas más ricas en hierro son Dulse y Lechuga de mar, las algas más ricas en manganeso son Dulse y Lechuga de mar, las algas más ricas en calcio son Espagueti de Mar y Wakame, y el alga más rica en magnesio es la Lechuga de mar. Estas concentraciones suponen una concentración de metales esenciales mayor que la que presentan los vegetales “terrestres” (Mataix Verdú, 1998).

38 Revista Real Academia Galega de Ciencias. Vol. XXVII
BIBLIOGRAFÍA
AHLGREN M., KONTKANEN A., VATTULAINEN K. y VEHVILAINEN H. (1988) Laser ashing: an alternative decomposition method. Part 1. Determination of calcium, magnesium, and potassium in plant material by flame atomic absorption spectrometry. Analyst, 113: 285-287.
BORKOWSKA-BURNECKA J., WISZ J. y ZYRNICKI W. (2003) Applicability of ultrasonic leaching by diluted acids for determination of total metal contents in plant materials. Chemia Analityczna, 48: 115-126.
CASTELLÓ ORVAY F. (1993) Acuicultura Marina. Universitat de Barcelona. Barcelona.
CHAN K.C.,YIP Y. C., CHU, H. S. y SHAM W.C. (2006) High-throughput determination of seven trace elements in food samples by inductively coupled plasma-mass spectrometry. Journal of AOAC International, 89: 469-479.
CHAUDHURI A., MITRA M., HAVRILLA C., WAGUESPACK Y. y SCHWARZ J. (2007) Heavy metal biomonitoring by seaweeds on the Delmarva Peninsula, east coast of the USA. Botanica Marina, 50: 151-158.
CURDOVA E., SZAKOVA J., MIHOLOVA D., MESTEK O. y SUCHANEK M. (1998) Evaluation of various mineralization methods and measurement techniques for trace element analysis of plant materials. Analusis, 26: 116-121.
DOMINGUEZ-GONZALEZ R., MOREDA-PINEIRO A., BERMEJO-BARRERA A. y BERMEJO-BARRERA P. (2005) Application of ultrasound-assisted acid leaching procedures for major and trace elements determination in edible seaweed by inductively coupled plasma-optical emission spectrometry. Talanta, 66: 937-942.
FERNÁNDEZ SAA C. (2002) Las Verduras del Océano Atlántico: Algas de Galicia. Alimento y Salud. Algamar. Pontevedra.
FILGUEIRAS A.V., CAPELO J. L., LAVILLA, I. y BENDICHO, C. (2000) Comparison of ultrasound-assisted extraction and microwave-assisted digestion for determination of magnesium, manganese and zinc in plant samples by flame atomic absorption spectrometry. Talanta, 53: 433-441.
GARNHAM G.W., CODD G.A. y GADD G.M. (1992) Kinetic of uptake and intracellular location of cobalt, manganese and zinc in the estuarine green alga Chlorella salina. Applied Microbiology and Biotechnology, 37: 270-276.
GONZÁLEZ C., GARCÍA O. y MÍGUEZ L. (1998) Algas Marinas de Galicia. Xerais. Vigo.
KAMALA-KANNAN S., PRABHU D.B.B., LEE K.J., KANNAN N., KRISHNAMOORTHY R., SHANTHI K. y JAYAPRAKASH M. (2008) Assessment of heavy metals (Cd, Cr and Pb) in water, sediment and seaweed (Ulva lactuca) in the Pulicat Lake, South East India. Chemosphere, 71: 1233-1240.
KNAPP G., SADJADI B. y SPITZY H. (1975) Behaviour of volatile elements in the incineration of organic matrices with chloric acid. Fresenius’ Zeitschrift fur Analytische Chemie, 274, 275-278. morrison l., baumann h. a., stengel d.b. (2007) An assessment of metal contamination along the Irish coast using the seaweed Ascophyllum nodosum (Fucales, Phaeophyceae). Environmental Pollution, 152: 293-303.
LAVILLA I., PEREZ-CID B. y BENDICHO C. (1998) Leaching of heavy metals from an aquatic plant (Lagarosiphon major) used as environmental biomonitor by ultrasonic extraction. International Journal of Environmental Analytical Chemistry, 72: 47-57.
LADRA-RAMOS N., DOMINGUEZ-GONZALEZ R., MOREDA-PINEIRO, A., BERMEJO-BARRERA A. y BERMEJO-BARRERA P. (2005) Determination of major and trace elements in edible seaweeds by AAS after ultrasound-assisted acid leaching. Atomic Spectroscopy, 26: 59-67.

39Revista Real Academia Galega de Ciencias. Vol. XXVII
LUQUE DE CASTRO M.D. y PRIEGO CAPOTE F. (2007) Analytical Applications of Ultrasound. Elsevier. Amsterdam.
LUQUE GARCÍA, J.L. y LUQUE DE CASTRO M.D. (2002) Acceleration Automation of Solid Sample Treatment, Elsevier, Amsterdam.
MATAIX VERDÚ J. (1998) Tablas de Composición de Alimentos Españoles. Universidad de Granada. Granada.
MEERAVALI N. N. y SUNIL J. K. (2000) Comparison of open microwave digestion and digestion by conventional heating for the determination of Cd, Cr, Cu and Pb in algae using transverse heated electrothermal atomic absorption spectrometry. Fresenius’ Journal of Analytical Chemistry, 366: 313-315.
MORALES-MUÑOZ S., LUQUE-GARCIA J.L. y LUQUE DE CASTRO M.D. (2003) Acidified pressurized hot water for the continuous extraction of cadmium and lead from plant materials prior to ETAAS. Spectrochimica Acta Part B, 58: 159–165.
MOREDA-PIÑEIRO J., ALONSO-RODRÍGUEZ E., LÓPEZ-MAHÍA P., MUNIATEGUI-LORENZO S., PRADA-RODRÍGUEZ D., MOREDA-PIÑEIRO A. Y BERMEJO-BARRERA P. (2007) Development of a new sample pre-treatment procedure based on pressurized liquid extraction for the determination of metals in edible seaweed. Analytica Chimica Acta, 598: 95-102.
MORENO-CID A. y YEBRA M. C. (2004) Continuous ultrasound -assisted extraction coupled to a flow injection -flame atomic absorption spectrometric system for calcium determination in seafood samples. Analytical and Bioanalytical Chemistry, 379: 77-82.
MORRISON L., BAUNANN H.A. y STENGEL D.B. (2007) An assessment of metal contamination along the Irish coast using the seaweed Ascophyllum nodosum (Fucales, Phaeophyceae). Environ. Pollu., 152: 293-309.
MUNILLA M., GOMEZ-PINILLA I., RODENAS S. y LARREA M. T. (1995) Determination of metals in seaweeds used as food by inductively coupled plasma atomic emission spectrometry. Analyst, 23: 463-466.
NASCENTES C. C., KORN M. y ARRUDA M.A.Z. (2001) A fast ultrasound-assisted extraction of Ca, Mg, Mn and Zn from vegetables. Microchemical Journal, 69: 37-43.
OTERO SCHMITT J.J., CASTRO M. y COMESAÑA P. (2002) Guía das Macroalgas de Galicia, Baía, A Coruña.
PÉREZ-CIRERA LÓPEZ-NIÑO J.L. (2005) Algas Mariñas en: A Natureza Ameazada, (2004) Viéitez Cortizo E., Rey Salgado J.M. (Ed.) Consello da Cultura Galega, Sección de Patrimonio Natural. Santiago de Compostela. 145-178.
PRIEGO-CAPOTE, F. y LUQUE DE CASTRO M.D. (2004) Analytical ultrasound. I. Sample preparation. Trends Anal. Chem., 23: 644-653.
RAO P. V. S.; MANTRI V. A. y GANESAN K. (2007) Mineral composition of edible seaweed Porphyra vietnamensis. Food Chemistry, 102: 215-218.
ROBLEDO D. y FREILE PELEGRIN Y. (1997) Chemical and mineral composition of six potentially edible seaweed species of Yucatan. Botanica Marina, 40: 301-306.
RUIZ-JIMENEZ J., LUQUE-GARCIA J. L. y LUQUE DE CASTRO M. D. (2003) Dynamic ultrasound-assisted extraction of cadmium and lead from plants prior to electrothermal atomic absorption spectrometry. Analytica Chimica Acta, 480: 231-237.
SAEED, K. A., AYORINDE, F. O., ERIBO, B. E., GORDON, M. y COLLIER, L. (1999) Characterization of partially transesterified poly(b-hydroxyalkanoate)s using matrix-assisted

40 Revista Real Academia Galega de Ciencias. Vol. XXVII
laser desorption/ionization time-of-flight mass spectrometry. Rapid Communications in Mass Spectrometry. 13: 1951-1957.
SOMENATH M. (2003) Sample Preparation Techniques in Analytical Chemistry. J. Wiley & Sons. New York.
VALCÁRCEL CASES M. y LUQUE DE CASTRO M.D. (1987) Flow-injection Analysis: Principles and Applications. Ellis Horwood Limited. Chichester.
VILLARES R., PUENTE X. y CARBALLEIRA A. (2002) Seasonal variation and background levels of heavy metals in two green seaweeds. Environmental Pollution, 119: 79-90.
YAMADA M., YAMAMOTO K., USHIHARA Y. y KAWAI H. (2007) Variation in metal concentrations in the brown alga Undaria pinnatifida in Osaka Bay, Japan. Phycological Research, 55: 222-230.
YEBRA M.C., CANCELA S. y MORENO-CID A. (2005) Continuous ultrasound-assisted extraction of cadmium from vegetable samples with on-line preconcentration coupled to a flow injection-flame atomic spectrometric system. International Journal of Environmental Analytical Chemistry, 85: 305-313.
ZEZZI ARRUDA, M.A. (2006) Trends in Sample Preparation. Nova Publishers. New York.

Revista Real Academia Galega de Ciencias. Vol. XXVII. Págs. 41-94 (2008)
EVALUACIÓN DEL EFECTO DE LA SELENOMETIONINA SOBRE LA REPARACIÓN DEL DAÑO EN EL ADN
VANESSA VALDIGLESIAS1,2*,1 BLANCA LAFFON2, EDUARDO PÁSARO2 Y JOSEFINA MÉNDEZ1
1Departamento de Biología Celular y Molecular, Universidade da Coruña. Facultad de Ciencias, Campus A Zapateira s/n, 15071 A Coruña, España.
2Unidad de Toxicología. Departamento de Psicobiología. Universidade da Coruña.Edificio de Servicios Centrales de Investigación, Campus Elviña s/n, 15071 A Coruña, España.
E-mail: [email protected]
RESUMEN
En este estudio se han evaluado los efectos de la selenometionina (SeMet), un derivado orgánico de selenio, sobre la inducción, reparación y persistencia del daño en el ADN inducido por la bleomicina (BLM) sobre leucocitos humanos. Se empleó el ensayo del cometa para determinar las roturas de cadena en el ADN, y el tratamiento con la enzima de reparación hOGG1 para el estudio específico del daño oxidativo. La SeMet fue añadida a los leucocitos durante el cultivo (A), la exposición a BLM (B) y/o durante el periodo de reparación (C). El ensayo del cometa se realizó al término de cada una de las fases. Los resultados obtenidos describen un posible papel antigenotóxico de la SeMet sobre el daño en el ADN inducido por la BLM, y también sobre la reparación y la persistencia de este daño, cuando se aplica antes o junto con la BLM.
Palabras clave: selenometionina, bleomicina, ensayo del cometa, daño oxidativo, antigenotoxicidad.
*Premio Academia Galega de Ciencias-Fundación CaixaGalicia a Jóvenes Investigadores, convocatoria año 2007.

42 Revista Real Academia Galega de Ciencias. Vol. XXVII
SUMMARY
The effects of the organic selenium compound selenomethionine (SeMet) on the induction, repair and persistence of DNA damage was evaluated in human leukocytes challenged with bleomycin (BLM). Comet assay was used to determine DNA strand breaks, and incubation with the repair enzyme hOGG1 for the specific recognition of oxidative damage. SeMet was added to the leukocytes during the culture (A), the exposure to BLM (B) and/or during the repair period (C). Our results showed antigenotoxic effect of SeMet on BLM-induced DNA damage and also on repair and persistence of this damage when applied before and simultaneously with BLM.
Key words: selenomethionine, bleomycin, comet assay, oxidative damage, antigenotoxicity.
1. INTRODUCCIÓN1.1. Selenio1.1.1. Descripción
El selenio (Se) es un elemento traza presente en todos los organismos y esencial para el funcionamiento normal de muchos procesos celulares. Desempeña una gran variedad de funciones en los sistemas biológicos, algunas de las cuales residen en su capacidad de actuar como antioxidante y elemento de prevención de enfermedades (Letavoyavá et al., 2006). No obstante, dependiendo de su forma química, el Se también puede actuar como agente tóxico pro-oxidante que puede inducir daño en el ADN y muerte celular (Spallholz, 1994). Dada esta dualidad que presenta el Se, el conocimiento exacto de las funciones que desempeña en el organismo y de su modo de acción a distintos niveles es considerado de gran interés para la salud humana.
El Se fue descubierto en 1817 por el químico sueco Jakob Berzelius y su toxicidad se determinó casi desde ese momento. Sin embargo, no se describió su papel nutricional hasta los años 50 del siglo XX, definido como esencial y aplicado en pasto de ganado y otros organismos. Comienza entonces un gran auge de estudios toxicológicos con Se en microorganismos, plantas, animales y en mucha menor medida en humanos, para esclarecer las causas de la toxicidad, determinar umbrales de ingesta y evaluar sus efectos beneficiosos para el organismo.

43Revista Real Academia Galega de Ciencias. Vol. XXVII
Casi 20 años después comienzan los estudios del posible papel preventivo del Se frente al cáncer. En 1973 J.T. Rotruck y colaboradores publicaron la primera demostración del papel funcional del Se estableciendo las bases bioquímicas y la relación del Se con la enzima glutation-peroxidasa implicada en la defensa antioxidante celular y relacionada, en consecuencia, con la aparición y desarrollo de procesos cancerígenos (Aboul-Fadl, 2005).
Desde entonces, se han multiplicado considerablemente los estudios con Se, haciéndose especialmente importantes los llevados a cabo in vivo en mamíferos. La mayoría de estos trabajos apoyan el hecho de que los posibles efectos beneficiosos o anticancerígenos del Se dependen de su forma química y de su concentración (Ip, 1998). Los primeros estudios con Se empleaban formas inorgánicas como el selenito, el selenato o sus respectivas sales sódicas (Lawson y Birt, 1983; Khalil, 1989; Yu et al., 1991). Aunque varios de estos trabajos señalaban que un suplemento en la dieta con sales de Se resultaba en una menor incidencia de cáncer u otras patologías (Yu et al., 1991 y 1997; Feng et al., 1999), en otros se concluyó que las sales no sólo no disminuían la incidencia, sino que ejercían efectos adversos sobre el organismo, principalmente debidos a la producción de especies reactivas de oxígeno (ROS) resultantes de su metabolismo (Khalil, 1989; Biswas et al., 1999; Wycherly et al., 2004).
Bioquímicamente, el Se es un análogo del azufre por lo que presenta muchas similitudes en sus propiedades físico-químicas (valencias, tamaños atómicos, bandas energéticas, potenciales de ionización, afinidades electrónicas…) aunque difiere en otras pocas (se metaboliza a estados más reducidos, presenta mayor acidez al interaccionar con el hidrógeno) lo que confiere a los compuestos de Se una mayor eficacia como agentes antitumorales frente a sus análogos sulfurados (Briviba et al., 1996; Aboul-Fadl, 2005).
Se han identificado entre 30 y 50 selenoproteínas importantes (25 de ellas en humanos), se han caracterizado completamente 20 por análisis de sus secuencias y se ha asignado función enzimática al menos a 10 de ellas (Hesketh y Villete, 2002). Todas contienen Se en su estructura y realizan funciones celulares importantes, la mayoría relacionadas con la defensa celular y la maquinaria de reparación. De entre las selenoproteínas más importantes y estudiadas destacan la familia de las glutation-transferasas y la de las tiorredoxin-reductasas (Thomson, 2004).
Tanto en la naturaleza como en la dieta, y en consecuencia en el organismo, el Se se puede presentar en distintas formas. El Se orgánico está

44 Revista Real Academia Galega de Ciencias. Vol. XXVII
presente en los alimentos principalmente en forma de selenocisteína (SeCys), selenometionina (SeMet) y metilselenocisteína (MeSeCys), mientras que las formas inorgánicas, como el selenito o el selenato, son mucho menos frecuentes y están en concentraciones mucho más bajas. De las formas orgánicas, la SeMet es la forma predominante en la mayoría de las dietas ricas en Se (Letavayová et al., 2006).
1.1.2. Toxicidad Exposición elevadaA niveles elevados, el Se puede resultar tóxico para el organismo,
conociéndose esta toxicidad desde hace varios siglos (Tinggi, 2003). En humanos, la toxicidad producida por Se se presenta principalmente en individuos expuestos a dosis elevadas o prolongadas en el tiempo.
Los síntomas de la intoxicación por Se, o selenosis, incluyen erupciones, fragilidad de uñas y pelo, paroniquia, decoloración en las uñas, aliento con olor a ajo (debido a la espiración de dimetilseleniuro), formación de vesículas e infecciones secundarias. En exposiciones más severas, el envenenamiento se agudiza llegando a producir caída de pelo, uñas y dientes, lesiones en la piel, alteraciones del sistema nervioso, fatiga, irritabilidad, flacidez muscular, temblores, etc. (Ammar y Couri, 1981; Aboul-Fadl, 2005).
Dos razones fundamentales explican la toxicidad del Se. Por un lado la capacidad de producir ROS durante su metabolismo (Spallholz, 1994 y 1997), y por otro la reducción a otros compuestos mutagénicos que se forman a partir de los compuestos originales de Se (Whiting et al., 1980).
La toxicidad del Se depende no obstante de diversos factores. La forma química y la cantidad suministrada parecen ser los más importantes pero también influyen otros como la edad, el estado fisiológico, interacciones con otros nutrientes o componentes de la dieta, la ruta de administración, etc. (Aboul-Fadl, 2005). En general se piensa que la toxicidad del Se se manifestará de forma aguda o crónica cuando el daño oxidativo que provoca como consecuencia de su metabolismo supere las defensas antioxidantes que posee la célula (Nogueira et al., 2004).
Se estimó que la cantidad más baja de Se suministrado a la cual se producen efectos adversos para el organismo (LOAEL, Lowest Adverse Effect Level) es de 1540±653μg/día (Yang et al., 1989), y la cantidad de Se a la que

45Revista Real Academia Galega de Ciencias. Vol. XXVII
no se espera ningún efecto adverso (NOAEL, No Adverse Effect Level) es de 819±126μg/día (Whanger et al., 1996).
Deficiencia
Además de la toxicidad que presenta el Se per se, se ha relacionado la deficiencia de este elemento en el organismo con la incidencia de varios tipos de cáncer, aparición y progresión de diversas patologías, mayor virulencia de determinadas infecciones y alteraciones de los sistemas nervioso, inmunológico y cardiovascular (revisado en Thomson, 2004; Patrick, 2004). Dos enfermedades, ya descritas, y relacionadas con la deficiencia de Se en la dieta son la enfermedad de Keshan caracterizada por una anomalía del músculo cardíaco, y la enfermedad de Kashin-Beck, que ocasiona alteraciones articulares y óseas (Beck et al., 2003; Thomson, 2004).
Varios estudios describen una relación inversa entre la cantidad de Se en el organismo y la incidencia de cáncer (Alaejos et al., 2000; Surai, 2006). Se vio que individuos con bajos niveles de Se en plasma presentaban una incidencia significativamente mayor de leucemia y de cáncer de intestino, recto, próstata, pulmón, ovario y mama. También se describió una asociación inversa aunque no significativa entre Se ingerido y cáncer de páncreas, piel y vejiga (revisado en Schrauzer et al., 1977).
La deficiencia de Se viene acompañada de un descenso en la respuesta inmune, por lo que está ligada a la aparición, virulencia y progresión de algunas infecciones virales como el SIDA (Beck et al., 2003). Este elemento también parece ejercer un efecto protector en pacientes con hepatitis vírica (B o C) contra la progresión hacia el cáncer de hígado (revisado en Rayman, 2000). El sistema nervioso puede verse también alterado por niveles deficientes de Se. Se ha descrito que una baja concentración de Se puede afectar a los neurotransmisores, pudiendo relacionarse con el desarrollo de la enfermedad de Alzheimer y con una aceleración de procesos seniles. Además una aportación de Se en la dieta reduce los ataques epilépticos en niños (revisado en Rayman, 2000).
En mujeres, bajos niveles de Se incrementan el riesgo de aborto y en hombres se ha relacionado con deficiencias en la movilidad de los espermatozoides y variaciones en la fertilidad (Letavayová et al., 2006). El Se tiene importancia además en la función tiroidea, ya que está relacionado con la correcta actividad de ciertas hormonas tiroideas (Patrick, 2004).

46 Revista Real Academia Galega de Ciencias. Vol. XXVII
1.1.3. Estudios de suplementación con selenio
En 1984, Shamberger publica una recopilación de los principales estudios realizados hasta la fecha con Se destacando la ambigüedad de los resultados. Resume sus efectos genotóxicos, mutagénicos y antimutagénicos en diferentes organismos y ya entonces, concluye que el Se es un agente mutagénico, principalmente a niveles elevados, aunque a niveles nutricionales puede actuar como anticarcinogénico en cáncer de mama, colon, riñón y cáncer de piel. En las últimas décadas, el estudio de la administración de Se en la dieta como posible método de prevención del cáncer o disminución del desarrollo de esta enfermedad se ha hecho más frecuente. Varios son los trabajos desarrollados in vitro, también los realizados con animales y algo menos con humanos (revisados en Whanger, 2004; Rayman, 2005). En ellos los resultados han sido diversos atendiendo a la forma y a la cantidad de Se empleadas.
Efectos beneficiosos
Partiendo de los efectos nocivos que provocan los bajos niveles de Se en el individuo y buscando el resultado contrario comenzó a estudiarse la posibilidad de que un suplemento de Se en el organismo tuviese efectos beneficiosos para el mismo. Se han publicado numerosos trabajos en las últimas décadas donde ciertamente se demuestra un efecto protector o beneficioso de este compuesto (revisado en Schrauzer, 2000a y Patrick, 2004). Algunos estudios indican que el Se confiere estabilidad genómica a células prostáticas cancerígenas (Dong et al., 2003; Karunasinghe et al., 2004; Morris et al., 2006). Además, en los ensayos clínicos realizados con suplementación con Se, se encontró que reduce la incidencia y la mortalidad del cáncer en general (Clark et al., 1996), del cáncer de próstata (Duffield-Lillico et al., 2002), del cáncer de hígado (Yu et al., 1997), del cáncer de estómago (Blot et al., 1993) y del cáncer de pulmón (Reid et al., 2002). En general, las evidencias epidemiológicas en humanos indican una relación significativamente inversa entre los niveles de Se y el riesgo de cáncer (Ganther, 1999).
Los posibles efectos anticancerígenos del Se pueden estar estrechamente relacionados con su papel en la reducción del estrés oxidativo por las selenoproteínas, con su capacidad de incrementar la respuesta inmune o, más probablemente, con su capacidad para producir metabolitos antitumorales (como el metilselenol o sus precursores) que pueden perturbar el metabolismo de las células tumorales, inhibir la angiogénesis e inducir la apoptosis de las

47Revista Real Academia Galega de Ciencias. Vol. XXVII
células cancerígenas (Rayman, 2000 y 2005; Whanger, 2004; Aboul-Fadl, 2005). Además de su relación con los procesos cancerígenos, también se observó que el suministro de Se puede reducir la progresión de la infección por VIH en pacientes portadores (Hurwitz et al., 2007), estimular la reparación del daño en el ADN inducida por distintos carcinógenos (Lawson y Birt, 1983; Kibriya et al., 2007, Luchese et al., 2007), preservar a las células de los efectos de una exposición a radiación UV (Rafferty, 1998 y 2003) y proteger al organismo de la toxicidad de otros compuestos como el plomo (Aykin-Burns y Ercal, 2006), el mercurio (Lemire et al., 2006) o el cadmio (Frisk et al., 2002).
Además, al igual que varios estudios sugerían que la deficiencia en Se está acompañada por una pérdida de inmunocompetencia, también se encontró que una suplementación con Se estimula el sistema inmunológico: linfocitos de voluntarios suplementados con Se en la dieta (en forma de selenito sódico) mostraron un aumento en la respuesta a la estimulación antigénica y una mayor capacidad de convertirse en linfocitos citotóxicos y de destruir células tumorales (Kiremidjian-Schumacher et al., 1994). En general, se cree que las células del sistema inmune pueden tener una necesidad funcional importante de Se (Rayman, 2000; Arthur et al., 2003).
Efectos adversosA pesar de todos los estudios in vitro e in vivo realizados en distintos
organismos que defienden los efectos beneficiosos de los compuestos de Se, otros trabajos pusieron de manifiesto los posibles efectos adversos que se pueden llegar a producir tras la administración. Se encontró que producían aberraciones cromosómicas y alteraciones en la estructura cromosómica en linfocitos humanos (Khalil, 1989; Biswas et al., 2000) y en células de ratones cuya dieta había sido previamente suplementada con Se (Biswas et al., 1999), aumentó la incidencia del cáncer de piel no melanómico en personas suplementadas con Se en dieta (Duffield-Lillico et al., 2003) y produjo estrés oxidativo en células y organismos tratados con este compuesto (Wycherly et al., 2004) llegando en algunos casos a alterar funciones celulares importantes (Brown et al., 2000). Todos los trabajos realizados concluyeron que tanto la toxicidad como el efecto quimiopreventivo del Se vienen determinados por diversos factores, principalmente la forma química y la concentración empleadas en cada caso.

48 Revista Real Academia Galega de Ciencias. Vol. XXVII
1.2. Selenometionina
1.2.1. Introducción histórica
La selenometionina (SeMet) es un aminoácido análogo a la metionina (Met) que contiene Se sustituyendo al átomo original de azufre (Figura 1). La forma isomérica L es la fuente natural común en los alimentos ricos en Se. Este compuesto no puede ser sintetizado por animales superiores, que lo obtienen a través de la dieta, principalmente de las plantas que sí pueden incorporarlo a sus proteínas sustituyendo a la metionina de forma inespecífica (Schrauzer, 2000b).
Figura 1. Estructura química de metionina (a) y selenometionina (b).
En los años 50-60 del siglo XX se identificó la SeMet como una proteína presente en determinadas plantas y se demostró también que era producida por Saccharomyces cerevisiae, Candida albicans, Escherichia coli, bacterias del rumen y algas marinas que crecían en medio que contiene Se (Schrauzer, 2000b). Pero no se supo mucho de la SeMet hasta los años 70 cuando se permitió su uso en alimentación animal. A mediados de esta década salen al mercado las primeras levaduras ricas en Se. Pronto se vio que la SeMet era bien absorbida y retenida por el organismo (Griffiths et al., 1976), por lo que su empleo desde entonces en dietas que necesitan aportación de Se ha ido en aumento. En 1983, levaduras ricas en SeMet fueron escogidas como fuente de Se en algunos tratamientos preventivos contra el cáncer (Schrauzer, 2001).
En Junio de 2000, fue aprobado el uso de levaduras con Se en alimentación animal por la FDA (Food and Drug Administration), a partir de este momento comienza una sustitución paulatina de las sales de Se por levaduras como complemento en la dieta hasta la actualidad, en que prácticamente todo el Se suplementado se administra en forma de levaduras enriquecidas (Schrauzer, 2001). La SeMet es la forma predominante de Se en dietas ricas en este compuesto (Letavayová et al., 2006) y ha sido encontrado como el principal componente de levaduras enriquecidas usadas como suplemento de Se en la dieta (Ayouni et al., 2007).

49Revista Real Academia Galega de Ciencias. Vol. XXVII
Actualmente, y basándose en estudios en humanos, se han establecido unos límites en la dieta, máximo de 400μg/día y mínimo de 40μg/día, dentro de los cuales no se presentan efectos adversos. Una ingesta superior a los 3200-5000μg/día suele implicar la aparición de selenosis y por debajo de los 11μg/día se producen problemas graves de salud por carencia de Se (enfermedad de Keshan). Es por esto que en quimioprevención se utiliza el Se en dosis limitadas (Letavoyavá et al., 2006).
1.2.2. Metabolismo
La SeMet se absorbe con facilidad y, o es inmediatamente metabolizada, o bien es incorporada en la estructura de las proteínas siendo almacenada principalmente en el páncreas (Suzuki et al., 2006a). Las células no distinguen entre Met y SeMet durante la síntesis de proteínas, así que este selenoaminoácido natural se incorpora inespecíficamente en el cuerpo de las proteínas (la albúmina, por ejemplo) en lugar de la Met, tanto si ésta es factor limitante como si no (Letavayová et al., 2006). El reemplazamiento de Met por SeMet no produce, como norma general, alteraciones estructurales en la proteína que lo porta, pero puede influir en la actividad de la enzima si la SeMet sustituye a la Met en una posición próxima al sitio activo. Como el grupo Se-CH3 de la SeMet es más hidrofóbico que el grupo S-CH3 de la Met, el acceso del sustrato puede verse afectado, alterando los parámetros cinéticos de la proteína (Schrauzer, 2000b).
Figura 2. Metabolismo de la SeMet.

50 Revista Real Academia Galega de Ciencias. Vol. XXVII
En la Figura 2 se muestran las posibles rutas metabólicas de la SeMet. Una de ellas la constituye la conversión de SeMet por la vía de trans-selenación en SeCys (similar a la trans-sulfuración del metabolismo de la Met); la SeCys es seguidamente convertida a H2Se siguiendo la ruta metabólica habitual de este compuesto (Esaki et al., 1981). Por otra parte, la SeMet puede generar un importante metabolito secundario: el metilselenol (Suzuki et al., 2006b). Esta ruta está catalizada por la metionina-α,γ-liasa, también conocida como metioninasa (Meuillet, et al., 2004). El metilselenol podrá luego dar lugar a H2Se por acción de una demetilasa o excretarse en la orina tras una serie de reacciones (Suzuki et al., 2006c).
1.2.3. Estudios con selenometionina
Puesto que la historia de la SeMet es reciente, los estudios sobre su caracterización y aplicación se centran en las últimas décadas, intensificándose recientemente debido a las aparentes propiedades beneficiosas que parece tener y a la posibilidad de ser empleado en la prevención de patologías e infecciones severas. Los resultados obtenidos en los diferentes trabajos realizados son bastante heterogéneos incluso en muchos casos contradictorios, por lo que se requieren más estudios que profundicen en sus efectos.
Efectos beneficiosos
Distintos trabajos in vitro defienden los beneficios de la SeMet y las ventajas de su empleo frente a otras fuentes de Se. Varios estudios descartan su citotoxicidad (Stewart et al., 1999; Seo et al., 2002a; El-Sayed et al., 2007), describen su efecto protector frente a metales tóxicos (Frisk et al., 2002), a peroxinitrito (Roussyn et al., 1996), y a radiación UV (Rafferty et al., 1998; Rafferty et al., 2003; Seo et al., 2002a) y destacan su capacidad de inducir una respuesta reparadora en las células tratadas (Russell et al., 1980). Numerosos estudios in vitro defienden también los posibles efectos quimiopreventivos de la SeMet relacionados con su capacidad para incrementar la actividad de las selenoenzimas, principalmente la glutation-peroxidasa (Kajander et al., 1990; Rafferty et al., 2003), retardar o inhibir la síntesis de ADN (Morris et al., 2006) y el crecimiento celular (Kajander et al., 1990; Dong et al., 2003), participar en la activación de la proteína supresora de tumores p53 (Seo et al., 2002b; Smith et al., 2004) o regular la homeostasis celular del zinc (Blessing et al., 2004).

51Revista Real Academia Galega de Ciencias. Vol. XXVII
En diversos trabajos con animales se observó que un suplemento de SeMet en la dieta no produce efectos negativos ni en las ratas alimentadas ni en su descendencia (Taylor et al., 2005), y que la SeMet puede reducir la carcinogénesis de colon inducida por aminas aromáticas (Feng et al., 1999). En humanos, el mayor estudio in vivo fue sin duda el realizado por Clark y colaboradores. Se trata de un estudio a doble ciego donde a una población de más de 1300 individuos se les suplementó con Se/placebo en la dieta durante 10 años. Los resultados muestran un descenso de la incidencia de cáncer en general y puntualmente de pulmón, próstata, colon y recto, y no detecta diferencias significativas entre los tratamientos para cáncer de cabeza y cuello, vejiga, esófago y mama (Clark et al., 1996; Reid et al., 2002, Duffield-Lillico et al., 2003).
Actualmente se está desarrollando otro gran estudio in vivo en donde, desde el año 2001, a más de 32000 personas se les está administrando SeMet y vitamina E en la dieta. Se trata de otro estudio a doble ciego siguiendo el protocolo ya descrito por Clark et al. (1996) y cuyos primeros resultados se obtendrán en el año 2013 (Klein et al., 2001).
Efectos adversos
Los primeros trabajos con SeMet obtuvieron resultados bastante negativos. Los estudios de Ammar y Couri (1981) con ratones concluyeron que una administración de SeMet intravenosa produce diversos efectos neurotóxicos en los animales tales como alteraciones cardiorrespiratorias, parálisis de las patas traseras y, en algunos casos, la muerte del individuo. Si la administración se producía como suplemento en la dieta, también se vio que la SeMet se acumulaba llegando a niveles tóxicos cuando la dieta era previamente rica en Met (Waschulewski y Sunde, 1988). Por otro lado, en un estudio in vitro se observó un incremento significativo de aberraciones cromosómicas en linfocitos humanos tratados con SeMet (Khalil, 1989).
En general, parece que los efectos adversos y la toxicidad de la SeMet dependen de diversos factores como la cantidad suministrada, siendo mayores en cantidades elevadas y mínimas (Kajander et al., 1990; Brown et al., 2000; Waters et al., 2005), el tipo de administración siendo preferible la via oral (Ammar y Couri, 1981), la interacción con otros suplementos comunes como la vitamina C o el sulfato de cobre, ya que se vio que la administración conjunta

52 Revista Real Academia Galega de Ciencias. Vol. XXVII
resulta en un incremento significativo de aductos de ADN (Shen et al., 2001), o los niveles basales de Se en el individuo receptor del suplemento.
En humanos, a partir de los resultados del trabajo de Clark (Clark et al., 1996) se llegó también a la conclusión de que el Se no sólo no disminuye la incidencia de cáncer de piel sino que aumenta, aunque no significativamente, el riesgo de carcinomas celulares escamosos y de cánceres no melanómicos de piel en general (Duffield-Lillico et al., 2003).
1.3. Evaluación de la capacidad de reparación del ADN: el ensayo de competencia de reparación
1.3.1. Reparación del ADN
Los mecanismos de reparación del ADN celular constituyen un sistema de defensa diseñado para proteger la integridad del genoma. Deficiencias en este sistema contribuyen, entre otros, al desarrollo del cáncer. Es por esto que la epidemiología de la capacidad de reparación del ADN y sus efectos sobre la susceptibilidad al cáncer en humanos constituyen una muy importante área de investigación actual (Berwick y Vineis, 2000).
Cuando un agente citotóxico llega a la célula produce una serie de daños que, de no repararse, pueden llevar a esa célula a experimentar distintas alteraciones estructurales o metabólicas, desencadenando finalmente procesos de apoptosis y muerte celular. Los distintos daños que puede sufrir el ADN incluyen apareamientos incorrectos, modificaciones covalentes de las bases nitrogenadas del ADN, roturas de cadena doble o sencilla, entrecruzamientos, etc. La célula posee sin embargo una serie de mecanismos de defensa frente a estos agentes que pueden, en muchos casos, evitar o disminuir el daño que inducirían en ella (Wood, 1996). Una primera línea defensiva la constituyen los sistemas de antioxidación y detoxificación celular, encargados de evitar que el agente ejerza su toxicidad bien por eliminación del tóxico bien mediante su transformación a compuestos bioquímicamente inertes (Luque y Herráez, 2002).
Si esta línea defensiva falla, entra en juego la maquinaria de reparación celular que abarca distintas vías y que está constituida por un gran número de complejos proteicos y enzimas asociadas. La reparación del ADN dañado incluye tres posibilidades: la reversión directa del daño basada fundamentalmente en la acción de distintas enzimas (fotoliasas, alquiltransferasas…); la reparación

53Revista Real Academia Galega de Ciencias. Vol. XXVII
por escisión, que incluye tres vías: la reparación por escisión de bases (Base Excision Repair, BER), la reparación por escisión de nucleótidos (Nucleotide Excision Repair, NER) y la reparación de emparejamientos erróneos (Mismatch Repair, MMR); y finalmente la reparación de roturas de doble cadena que también implica dos vías distintas: la recombinación homóloga (Homologous Recombination, HR) y la unión final no homóloga (Non-Homologous End Joining, NHEJ) (Luque y Herráez, 2002).
Si esta maquinaria no consigue reparar el daño causado por un agente dado, se desarrollará una anomalía en el ADN que puede desembocar finalmente en un proceso cancerígeno o en una patología dada (Buermeyer, et al., 1999; De Boer y Hoeijmakers, 2000). La importancia de estudiar la capacidad de reparar lesiones en el ADN radica por tanto en la observación de que mecanismos deficientes de reparación del genoma se han relacionado con la presencia de una gran variedad de patologías y procesos neoplásicos, y de que alteraciones en estos mecanismos pueden influir en la susceptibilidad de los individuos expuestos a un determinado mutágeno (Au, 1993; Au et al., 1996a; Berwick y Vineis, 2000). No obstante, las técnicas y metodologías para evaluar la capacidad de reparación del ADN están todavía en desarrollo, por lo que su aplicación resulta de rigurosa actualidad. Se conocen diversos métodos para evaluar la capacidad de reparación de las células, que se agrupan en 4 categorías principales, además de las combinaciones entre ellas (Berwick y Vineis, 2000): tests basados en daño inducido en el ADN con agentes químicos o físicos (más conocidos como “ensayos de sensibilidad a mutágenos”), medidas indirectas de reparación del ADN (como la síntesis de ADN no programada y la medida de actividades enzimáticas), tests basados en medidas cinéticas de reparación (entre ellos el host cell reactivation assay) y medidas de variación genética asociada con las diferencias en la reparación del ADN (polimorfismos en genes de reparación).
1.3.2. El ensayo de competencia de reparación del ADN
El ensayo de competencia de reparación del ADN (DNA repair competence assay) es un ensayo citogenético, basado en el challenge assay desarrollado por Au y colaboradores (1991), que permite evaluar la capacidad celular de reparar el daño inducido en el ADN por diversos agentes (Cebulska-Wasilewska, 2003). Partiendo de la observación de que la exposición a agentes mutagénicos puede producir importantes alteraciones

54 Revista Real Academia Galega de Ciencias. Vol. XXVII
en el ADN aumentando así el riesgo a padecer determinadas enfermedades como el cáncer, se plantea como posible causa de esto no sólo los efectos genotóxicos directos inducidos por la exposición, sino también el hecho de que ésta puede originar anomalías en la maquinaria de reparación celular (Au et al., 2001). Estas alteraciones podrían en consecuencia reducir la capacidad de las células de reparar el ADN dañado, lo que disminuiría la defensa del individuo frente a exposiciones posteriores y favorecería la aparición y el desarrollo de los procesos patológicos. El fundamento de este ensayo consiste en que las células expuestas a un determinado agente pueden tener deteriorada su maquinaria de reparación del ADN, disminuyendo así la capacidad de reparación del daño inducido. Cuando esas células son tratadas posteriormente con un mutágeno “in vitro” (irradiadas con rayos X, por ejemplo) presentarán incrementos significativos en el daño generado en el ADN respecto a células no expuestas, y esto puede ser evaluado mediante la aplicación de diversos tests de genotoxicidad (aberraciones cromosómicas, micronúcleos, ensayo del cometa, etc.) (Au, 1993).
Según la hipótesis de partida y el fundamento de la técnica se define por tanto el ensayo de competencia de reparación del ADN como un ensayo de capacidad de reparación, donde las células son forzadas, mediante distintos métodos de inducción, a reparar las lesiones producidas en el ADN (Au, 2003). Lo realmente novedoso de esta técnica, y la ventaja frente a otras técnicas similares, es la posibilidad de cuantificar esa reparación por la evaluación del daño a diferentes tiempos (Schmezer et al., 2001). El desarrollo de la técnica es sencillo. Muestras celulares (generalmente leucocitos periféricos) de poblaciones expuestas a un tóxico (en estudios in vivo) o células cultivadas en presencia de un determinado agente mutagénico (en estudios in vitro), son inducidas a desarrollar lesiones en su ADN mediante algún agente inductor (lo que se denomina challenge: “reto”). Posteriormente, empleando diversos tests de genotoxicidad se evalúa el daño a diferentes tiempos. Lo más habitual es medir a tiempo 0 (inmediatamente después del tratamiento con el agente inductor) y después de un tiempo de reparación (distinto según cada caso), aunque algunos autores establecen también medidas del daño a tiempos intermedios para seguir el proceso de reparación de forma progresiva. El descenso en el daño genotóxico evaluado producido durante la incubación se toma como medida de la capacidad de reparación del ADN (Au, 1993; Au et al., 1995b; Rajaee-Behbahani et al., 2001). Puesto que la incapacidad para reparar el ADN dañado es una de las principales causas del cáncer, se ha propuesto el uso del ensayo de competencia de reparación como un buen

55Revista Real Academia Galega de Ciencias. Vol. XXVII
biomarcador del riesgo a desarrollar esta patología (Au, 1993), basándose en la asociación establecida por numerosos estudios previos entre la mayoría de los cánceres y un elevado índice de anomalías cromosómicas.
Bleomicina como agente inductorLa bleomicina es un antibiótico antineoplásico que se usa en diferentes
terapias (solo o en combinación con otros) para tratar diferentes enfermedades incluidos carcinomas, tumores testiculares, sarcoma Kaposi y linfomas no-Hodgkin (Wozniak et al., 2004).
Figura 3. Estructura química de la bleomicina (análogos).
Bioquímicamente, la bleomicina es un glucopéptido que contiene varios aminoácidos y azúcares y un extremo amino terminal que varía entre los diferentes análogos de la bleomicina (Figura 3). Se han encontrado pocas diferencias en cuanto a la actividad o mecanismos de acción entre esos análogos (Povirk y Austin, 1991). La bleomicina se comporta como radiomimético e induce un amplio espectro de lesiones mutagénicas en células de mamíferos, incluyendo daño en bases del ADN, sitios apurínicos y apirimidínicos, alteraciones en el ARN (Milic y Kopjar, 2004), y la generación de sitios álcali-lábiles (Wozniak et al., 2004) que resultan en roturas de cadena sencilla y doble (Wei et al., 2005). Además, al igual que otros agentes químicos, entre ellos el peróxido de hidrógeno, la bleomicina genera daño oxidativo en el ADN a través de la producción de radicales de oxígeno. En células eucariotas, la genotoxicidad de estos compuestos ha sido investigada concluyéndose que ambos producen aberraciones cromosómicas, mutaciones en genes y roturas de cadena de ADN (Anderson et al., 1994).

56 Revista Real Academia Galega de Ciencias. Vol. XXVII
La bleomicina está clasificada como un agente clastogénico directo, puesto que no necesita activación metabólica para ejercer su actividad, y actúa de forma independiente de la fase S de replicación del material genético en el ciclo celular (Povirk y Austin, 1991). Es por ello frecuente el uso de este compuesto como agente inductor de daño en estudios de medida de la capacidad de reparación celular (Gajecka et al., 2005; Wei et al., 2005).
El ensayo del cometa como test de genotoxicidad
El ensayo de electroforesis en microgel de células aisladas (“single cell gel electrophoresis assay”), también llamado ensayo del cometa, es una técnica sencilla, rápida y sensible para el análisis y cuantificación del daño en el ADN en células individuales (Singh et al., 1988). Esta técnica mide roturas de cadena en el ADN y permite, de este modo, detectar los efectos genotóxicos de diferentes agentes físicos o químicos. El ensayo parte de la lisis de células que se encuentran embebidas en un gel de agarosa preparado sobre un portaobjetos con una solución que contiene detergente, para permeabilizar las células, y alta concentración de sales, para eliminar las proteínas nucleares y permitir el desenrollamiento de la hélice de ADN. De esta manera, se eliminan las membranas, el citoplasma y el nucleoplasma y se obtienen nucleoides. Posteriormente se realiza una incubación en medio neutro o alcalino para permitir el desenrollamiento y relajación de la estructura superenrollada del ADN. A continuación el ADN se somete a electroforesis y se tiñe con un marcador fluorescente. Durante la electroforesis, los fragmentos rotos de ADN y las zonas relajadas de la cromatina migran hacia el ánodo, fuera del núcleo, formándose así una estructura que recuerda a un cometa (Figura 4).
La longitud e intensidad de la cola del cometa son proporcionales al número de roturas en el ADN inducidas por el agente en estudio.
Figura 4. Células sometidas al ensayo del cometa: (a) célula no dañada, (b) célula con daño moderado y (c) célula muy dañada.

57Revista Real Academia Galega de Ciencias. Vol. XXVII
El tipo de daño evaluado depende de las condiciones utilizadas. Si las condiciones son neutras se evalúan las roturas de doble cadena, y si las condiciones son alcalinas, se pueden detectar además las roturas de cadena sencilla, los sitios álcali-lábiles y procesos incompletos de reparación por escisión, ya que las condiciones alcalinas producen la rotura de los puentes de hidrógeno que mantienen unida la estructura de la doble hélice de ADN. Además, este ensayo permite analizar tipos específicos de daño en el ADN mediante la utilización de enzimas de reparación que reconocen y cortan bases nitrogenadas con ciertas modificaciones determinadas (Collins et al., 1993 y 1996).
La enzima hOGG1 para detectar daño oxidativo
La enzima hOGG1 (8-hidroxiguanina ADN-glicosilasa humana) es una glicosilasa que participa en la maquinaria de reparación celular en la vía de reparación por escisión de bases. Cuando un radical hidroxilo entra en contacto con el ADN puede atacar sitios de elevada densidad electrónica llegando a modificar sus bases. De entre todas las bases la guanina es la más propensa a la oxidación, durante la cual se añade un grupo hidroxilo a la posición 8 produciendo como resultado la 8-hidroxi-2’-desoxiguanosina (8-OHdG) (Angerer et al., 2007). La enzima hOGG1 reconoce esta base modificada y corta específicamente el ADN en los puntos donde se encuentra (Smith et al., 2006), iniciando así su reparación.
Para detectar daño oxidativo mediante el ensayo del cometa se pueden emplear diversas enzimas. Las enzimas más comúnmente empleadas son la formamidopirimidina ADN-glicosilasa (Fpg), que reconoce purinas modificadas en el imidazol, o la endonucleasa III (Endo III), que reconoce pirimidinas oxidadas. La enzima hOGG1 sin embargo, es la más eficaz en la determinación específica del daño oxidativo más frecuente (8-OHdG) puesto que, al contrario de las demás, no reconoce bases alquiladas (Smith et al., 2006). Al incubar los leucocitos durante el ensayo del cometa con hOGG1, ésta reconocerá las bases modificadas 8-OHdG, indicativas de daño oxidativo, y las eliminará de la secuencia de ADN generando un sitio abásico, que durante la incubación en medio alcalino dará lugar a una rotura de cadena. Así, el porcentaje de ADN presente en las colas de los cometas generados tras la incubación con esta enzima será mayor y reflejo del daño oxidativo presente en las células.

58 Revista Real Academia Galega de Ciencias. Vol. XXVII
1.4. Interés del estudio y aplicaciones
El trabajo a realizar tiene interés como estudio individual en cuanto a la información que se obtendrá sobre los efectos de la selenometionina en humanos, y sus futuras aplicaciones como suplemento en la dieta de individuos enfermos o susceptibles de padecer una determinada enfermedad. No obstante, el mayor interés que puede presentar este estudio es como parte de un estudio más global sobre el papel del Se en el organismo y sus posibles efectos antioxidantes, ya que aportaría información interesante y relevante sobre su empleo, delimitaría las condiciones para su correcta utilización y podría aportar datos importantes de cara a entender el/los motivos por los que los compuestos de Se son beneficiosos para determinados individuos, por ejemplo pacientes con procesos cancerígenos en desarrollo.
En esta línea, el hecho de emplear la bleomicina (un fármaco empleado en quimioterapia) como agente inductor de daño oxidativo nos permitirá establecer una posible relación del efecto que tendrá la selenometionina, posible antioxidante, en pacientes tratados con este medicamento. Por otro lado, la metodología empleada en la estimación de la capacidad de reparación del ADN, el ensayo de competencia de reparación, tiene interés y aplicación en el campo de la Biomedicina, como técnica citogenética para visualizar problemas en la reparación del ADN en poblaciones de individuos enfermos o susceptibles de estarlo, y en la Toxicología, en estudios tanto in vitro como poblacionales, de exposiciones a cualquier agente cuya posible toxicidad se quiera testar. La incapacidad para reparar el ADN dañado es la principal causa del cáncer.
En base a todo lo descrito anteriormente, las aplicaciones de este estudio podrían ser:
1. Documentar los efectos de la exposición selenometionina a diferentes concentraciones con el fin de descartar posibles alteraciones en el ADN (roturas de cadena o daño oxidativo) producidas por este compuesto y establecer los niveles óptimos para su empleo en la dieta.
2. Evaluar la posible influencia de la selenometionina en la maquinaria de reparación celular y en la capacidad específica de reparar el daño oxidativo, estrechamente relacionado con diversas patologías del sistema nervioso, cardiorrespiratorio y muscular así como la aparición y el desarrollo de procesos neoplásicos.

59Revista Real Academia Galega de Ciencias. Vol. XXVII
3. Avanzar en el campo, actualmente en desarrollo, de la estimación de la capacidad de reparación celular, mediante la aplicación de una técnica cuantitativa recientemente descrita, el ensayo de competencia de reparación del ADN, que puede ser empleada para numerosos estudios de exposición ocupacional, ambiental, o estudios de cohortes en general, siendo muy útil en los trabajos sobre procesos cancerígenos.
2. OBJETIVOS
Dado que la SeMet es un compuesto de Se y, como tal, productor de especies reactivas de oxígeno y otros metabolitos secundarios que pueden alterar el genoma, pero es también un importante elemento traza y un posible agente anticancerígeno empleado ya como suplemento en determinadas dietas, resulta imprescindible caracterizar y delimitar sus efectos sobre el ADN y sobre los mecanismos que intervienen en su reparación. Es por ello que en este estudio nos hemos planteado evaluar el efecto de la SeMet sobre el material genético de leucocitos humanos aislados y su posible papel protector en la reparación del daño inducido experimentalmente en el ADN por bleomicina, estudiando de forma general su implicación sobre las roturas de cadena (simple y doble) y los sitios abásicos, así como específicamente sobre el daño oxidativo. Para esto hemos formulado los siguientes objetivos:
1. Evaluar los efectos genotóxicos de la SeMet utilizando el ensayo del cometa y analizar la posible existencia de una relación dosis-respuesta.
2. Comprobar los efectos genotóxicos de la bleomicina para validar su utilización como agente inductor de daño en el ADN en el ensayo de competencia de reparación.
3. Analizar los efectos de la SeMet sobre la inducción, reparación y persistencia del daño en el ADN en general, y especificamente del daño oxidativo, causados por la bleomicina mediante la utilización del ensayo de competencia de reparación.

60 Revista Real Academia Galega de Ciencias. Vol. XXVII
3. MATERIAL Y MÉTODOS
3.1. Diseño experimental
Este estudio se ha dividido en dos partes experimentales. En la primera parte se evaluó el potencial genotóxico de la SeMet. Se cultivaron leucocitos humanos en presencia de distintas concentraciones de este compuesto (1, 5, 10, 50 y 100μM), junto con un control negativo, y se realizó el ensayo del cometa según el protocolo estándar para evaluar daño en el ADN, y la incubación con la enzima hOGG1 para detectar daño oxidativo. En base a los resultados obtenidos se seleccionó la concentración más adecuada para llevar a cabo el ensayo de competencia de reparación del ADN en el resto del trabajo.
En la segunda parte se utilizó la técnica del ensayo de competencia de reparación del ADN, empleando bleomicina como agente inductor y el ensayo del cometa (con y sin incubación con hOGG1) como test de genotoxicidad, para analizar el efecto de la SeMet sobre la capacidad de reparación celular del daño causado por la bleomicina. Para ello se cultivaron leucocitos en presencia de SeMet durante distintas fases: fase de cultivo (A), fase de inducción de daño (B), fase de reparación (C), fases AB y fases ABC, incorporando en cada caso los controles correspondientes.
3.2. Obtención de muestras
Para la realización de este estudio in vitro se han empleado leucocitos mononucleares (linfocitos y monocitos) extraídos de sangre periférica humana. Las muestras de sangre se obtuvieron de 3 donantes voluntarios varones no fumadores (D1-D3) de edades comprendidas entre 20 y 30 años que firmaron un consentimiento informado previo a la realización del estudio. El Comité de Ética de la Investigación de la Universidade da Coruña (CEI-UDC) aprobó el protocolo experimental empleado en la realización de este trabajo (informe Nº PI 1/2007).
Las muestras de sangre se recogieron en tubos con heparina de litio. El aislamiento de los leucocitos mononucleares se realizó mediante centrifugación en un gradiente de densidad de Ficoll. Para ello un volumen de 10 ml de sangre se diluyó en proporción 1:1 en tampón fosfato pH 7.4 (PBS) (Phosphate Buffered Saline Tablets, Sigma) y fue depositado sobre un volumen equivalente de solución de Ficoll (Lymphocyte Isolation Solution, Rafer). Tras someterlo a centrifugación durante 30 min a 2100 rpm, se recogió la nube linfocítica y

61Revista Real Academia Galega de Ciencias. Vol. XXVII
se realizaron dos lavados con 10 ml de PBS. El botón celular resultante se resuspendió en medio de congelación (50% suero bovino fetal, 40% RPMI 1640, 10% DMSO) y se congeló a -80ºC utilizando para ello un contenedor que garantiza el descenso gradual de la temperatura a razón de 1ºC/min (NALGENE® Cryo 1ºC Freezing Container, Nalgene Nunc International), minimizando así el posible daño celular durante la criopreservación.
3.3. Ensayo del cometa
Parar evaluar la posible genotoxicidad de la SeMet sobre el ADN celular, se empleó ensayo del cometa siguiendo el protocolo general descrito por Singh et al. (1988), con algunas modificaciones posteriores (Laffon et al., 2005). Además, se ha utilizado también la variante de este ensayo que introduce una incubación con la enzima de reparación hOGG1, que genera roturas de cadena a partir de los lugares donde hay guaninas oxidadas (8-OHdG), lo que permite evaluar específicamente el daño de carácter oxidativo (Collins, 2004).
3.3.1. Descongelación de leucocitos y tratamiento con SeMet
El día anterior a realizar el ensayo del cometa, los leucocitos fueron retirados de la cámara de -80ºC y descongelados en un baño a 37ºC. Se colocaron en un medio de cultivo compuesto por 4.5 ml de RPMI 1640 suplementado con 15% suero bovino fetal, 1% fitohemaglutinina, 1% L-glutamina (200mM) y 1% penicilina (5000U/ml)/estreptomicina (5000µg/ml) (todo de Invitrogen). Se añadió la SeMet (1, 5, 10, 50 y 100μM) de forma que los tratamientos constituyesen el 1% del volumen final del cultivo. El control negativo se estableció sin la adición de ningún tratamiento.Todos los tubos se mantuvieron en estufa a 37ºC durante 20h para garantizar que los leucocitos, que circulan en sangre periférica en fase G0, se encontrasen en fase G1 gracias a la estimulación del crecimiento mitótico propiciada por la fitohemaglutinina.
3.3.2. Preparación de los microgeles y lisis
Transcurrido el tiempo de cultivo los tubos se retiraron de la estufa, se centrifugaron a 1500 rpm durante 10 min y se resuspendieron en medio RPMI 1640. Se evaluó la viabilidad celular mediante la técnica de exclusión del

62 Revista Real Academia Galega de Ciencias. Vol. XXVII
colorante azul de tripán, resultando en todos los casos superior al 80%. Las células fueron luego nuevamente centrifugadas y resuspendidas primero en PBS y luego en 150 μl de agarosa de bajo punto de fusión al 0.5% en tampón fosfato, y depositadas en dos puntos sobre portaobjetos ya cubiertos con una primera capa de agarosa normal al 0.5% en agua destilada y deshidratados por calentamiento a 65ºC durante 15min. Sobre cada gota se depositó un cubreobjetos y se colocaron sobre hielo durante 15min. Una vez solidificada la agarosa se retiraron los cubreobjetos. Posteriormente se procedió a la lisis celular, para lo que se sumergieron los portaobjetos en una solución de lisis a pH 10 compuesta por NaCl 2.5M, Na2EDTA 100mM, Tris-HCl 10mM, NaOH 250mM y Triton X-100 1%, permaneciendo 1h en oscuridad a 4ºC. Durante todos los pasos posteriores se mantuvieron estrictas condiciones de oscuridad para evitar daños adicionales sobre el material genético. Los portaobjetos en los que se realizó el ensayo del cometa tradicional pasan directamente a la electroforesis después de la lisis; aquellos portaobjetos en los que se evaluó el daño oxidativo son sometidos al tratamiento con la enzima de reparación antes de la electroforesis.
3.3.3. Tratamiento con hOGG1
Tras la lisis se llevó a cabo el tratamiento con la enzima hOGG1 en aquellos portaobjetos donde se evaluaría el daño oxidativo, siguiendo el protocolo propuesto por Smith et al. (2006). Para ello, los portaobjetos fueron previamente lavados 3 veces durante 5 min con un tampón compuesto por EDTA 0.5 mM, BSA 0.2 mg/ml, KCl 0.1M y Hepes 40 mM pH 8, se añadieron 50 μl de hOGG1 (0.0016 U/μl) preparada en tampón sobre cada gota, se depositó un cubreobjetos sobre ellas y se incubaron en la estufa a 37ºC durante 10min. Los controles siguieron el mismo protocolo que los portaobjetos con enzima, pero empleando 50 μl de tampón sin enzima en el tratamiento.
3.3.4. Electroforesis, neutralización y tinción
Los portaobjetos, retirándoles previamente el cubreobjetos, se sumergieron entonces en una solución alcalina en el tanque de electroforesis, ubicado sobre una cubeta con hielo, para facilitar el desenrollamiento del ADN y expresión de los sitios álcali-lábiles. La solución, de pH>13, estuvo compuesta por Na2EDTA 1 mM y NaOH 300 mM. Tras 20min de incubación se inició el proceso electroforético de 20min de duración a 300 mA y 25 V (0.83V/cm).

63Revista Real Academia Galega de Ciencias. Vol. XXVII
A continuación se procedió a la neutralización de las muestras mediante 3 lavados de 5min con una solución de pH 7.5 elaborada con Tris-HCl 0.4 M. Todos los portaobjetos fueron codificados para asegurar la realización de un estudio “ciego” y teñidos con 60 μl de 4,6-diamidino-2-fenilindol (DAPI) 5 μg/ml.
3.3.5. Captura y análisis de las imágenes
La captura de las imágenes fue realizada mediante el programa Leica QWIN (Leica Imaging Systems), evaluando como parámetro estimador del daño en el ADN el porcentaje de ADN en la cola del cometa (%TDNA), medido éste desde el centro estimado de la célula. Se analizaron un total de 100 células por tratamiento y donante, 50 procedentes de cada una de las réplicas.
3.4. El ensayo de competencia de reparación del ADN
Ya que en el ensayo de competencia de reparación se ha utilizado el ensayo del cometa como test de genotoxicidad, ambos ensayos comparten elementos comunes en sus protocolos, coincidiendo en los pasos de descongelación de leucocitos, preparación de los microgeles, tratamiento con la enzima hOGG1, lisis, electroforesis, neutralización, tinción y captura de imágenes, que ya se han explicado previamente. La diferencia fundamental radica en el tratamiento con el agente inductor de daño, la bleomicina, y en el tiempo durante el cual se permite la reparación del daño inducido, lo que determina las distintas fases del protocolo: cultivo (A), inducción de daño (B) y reparación (C).
Tras la descongelación y transcurridas las 20h de cultivo, los leucocitos se retiraron de la estufa, se centrifugaron a 1500 rpm durante 10min y se resuspendieron en medio RPMI 1640. Entonces se añadió la bleomicina (20 μg/ml) se incubaron 30 min a 37ºC. Posteriormente las células fueron centrifugadas en PBS 3 min a 9000 rpm, para eliminar los posibles restos de bleomicina, y nuevamente centrifugadas en las mismas condiciones y resuspendidas en RPMI 1640. Para los tratamientos sin reparación, las células se centrifugaron de nuevo directamente, y para los tratamientos con reparación se incubaron durante 15 min a 37ºC antes de realizar la tercera centrifugación. El botón celular resultante se resuspendió en agarosa y se prepararon los microgeles, continuando con el protocolo anteriormente descrito para el ensayo del cometa.

64 Revista Real Academia Galega de Ciencias. Vol. XXVII
3.5. Análisis estadístico
La normalidad de la variable obtenida en el estudio (%TDNA) se evaluó mediante el test de bondad de ajuste de Kolmogorov-Smirnov. Dado que se alejaba significativamente de la distribución normal, se asumieron los tests no paramétricos como los más adecuados para su utilización en el análisis de los datos. Se empleó por tanto el análisis de Kruskal-Wallis para varias muestras independientes que nos permitió, complementado con el test U de Mann-Whitney, establecer la existencia de diferencias significativas entre los distintos tratamientos realizados. La existencia de una posible relación dosis-respuesta para la SeMet se evaluó mediante el coeficiente de correlación de Pearson. El nivel de significación fue fijado en 5%. Todos los análisis estadísticos se llevaron a cabo mediante el paquete estadístico SPSS para Windows, versión 15.0 (2007) (Illinois, USA).
4. RESULTADOS
4.1. Genotoxicidad de la selenometionina
Figura 5. Resultados del ensayo del cometa en leucocitos humanos tratados con SeMet. aP≤0.05, bP≤0.01, diferencia significativa respecto al control.
Con el fin de evaluar la posible genotoxicidad de la SeMet sobre leucocitos humanos y determinar en base a ello la concentración óptima a emplear en el estudio de su influencia sobre la capacidad de reparación del ADN, hemos realizado el ensayo del cometa utilizando 5 concentraciones distintas (1, 5, 10, 50 y 100µM), además de un control negativo. Los resultados obtenidos se muestran en la Figura 5, tomando como parámetro cuantitativo el porcentaje promedio de ADN en la cola del cometa (%TDNA) determinado a partir del
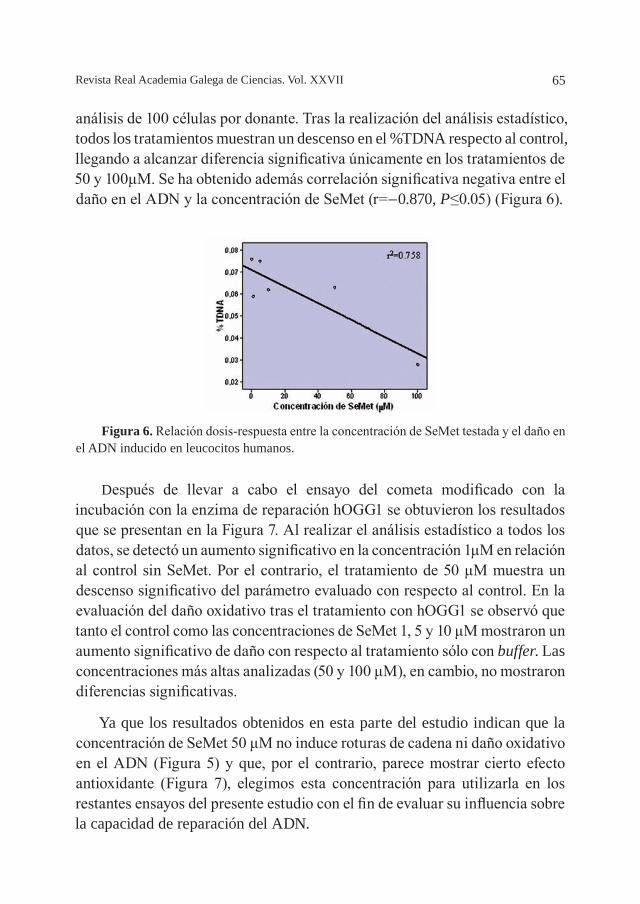
65Revista Real Academia Galega de Ciencias. Vol. XXVII
análisis de 100 células por donante. Tras la realización del análisis estadístico, todos los tratamientos muestran un descenso en el %TDNA respecto al control, llegando a alcanzar diferencia significativa únicamente en los tratamientos de 50 y 100μM. Se ha obtenido además correlación significativa negativa entre el daño en el ADN y la concentración de SeMet (r=−0.870, P≤0.05) (Figura 6).
Figura 6. Relación dosis-respuesta entre la concentración de SeMet testada y el daño en el ADN inducido en leucocitos humanos.
Después de llevar a cabo el ensayo del cometa modificado con la incubación con la enzima de reparación hOGG1 se obtuvieron los resultados que se presentan en la Figura 7. Al realizar el análisis estadístico a todos los datos, se detectó un aumento significativo en la concentración 1μM en relación al control sin SeMet. Por el contrario, el tratamiento de 50 μM muestra un descenso significativo del parámetro evaluado con respecto al control. En la evaluación del daño oxidativo tras el tratamiento con hOGG1 se observó que tanto el control como las concentraciones de SeMet 1, 5 y 10 μM mostraron un aumento significativo de daño con respecto al tratamiento sólo con buffer. Las concentraciones más altas analizadas (50 y 100 μM), en cambio, no mostraron diferencias significativas.
Ya que los resultados obtenidos en esta parte del estudio indican que la concentración de SeMet 50 μM no induce roturas de cadena ni daño oxidativo en el ADN (Figura 5) y que, por el contrario, parece mostrar cierto efecto antioxidante (Figura 7), elegimos esta concentración para utilizarla en los restantes ensayos del presente estudio con el fin de evaluar su influencia sobre la capacidad de reparación del ADN.

66 Revista Real Academia Galega de Ciencias. Vol. XXVII
Figura 7. Resultados del ensayo del cometa tras los tratamientos con SeMet empleando la enzima hOGG1. aP≤0.05, bP≤0.01, diferencia significativa respecto al control; cP≤0.05, dP≤0.01, diferencia significativa respecto a la misma concentración sin emplear enzima.
4.2. Genotoxicidad de la bleomicina
Previo al estudio de la interacción de la SeMet con la capacidad de reparación del ADN evaluamos los efectos genotóxicos de la bleomicina, compuesto que se utilizaría como agente inductor de daño en el ensayo de competencia de reparación del ADN, sobre leucocitos humanos realizando nuevamente el ensayo del cometa, con posterior tratamiento con enzima hOGG1 para estudiar por separado la inducción específica de daño oxidativo. El daño en el ADN presente tras el tratamiento con bleomicina resultó significativamente mayor que su correspondiente control, tanto antes como después del tiempo de reparación (Figura 8). No obstante, tras la reparación, el %TDNA disminuyó significativamente con respecto al detectado inmediatamente después de la inducción con bleomicina.
Figura 8. Resultados del ensayo del cometa tras el tratamiento con bleomicina. bP≤0.01, diferencia significativa respecto al control; dP≤0.01, diferencia significativa respecto al tratamiento sin reparación.

67Revista Real Academia Galega de Ciencias. Vol. XXVII
Al aplicar el tratamiento con la enzima hOGG1 se pone de manifiesto un aumento significativo del daño en el ADN respecto al control, tanto antes como después de la reparación (Figura 9). En relación al propio tratamiento con hOGG1, antes de la reparación se obtiene un aumento significativo del %TDNA respecto al tratamiento sólo con tampón, que desaparece tras el tiempo de reparación testado. Además, después de la reparación se obtiene un descenso significativo del daño evaluado con respecto al detectado antes de ese periodo.
Figura 9. Resultados del ensayo del cometa tras el tratamiento con bleomicina empleando la enzima hOGG1 (bP≤0.01, diferencia significativa respecto al control; dP≤0.01, diferencia significativa respecto al mismo tratamiento sin emplear enzima; eP≤0.01, diferencia significativa respecto al tratamiento sin reparación).
4.3. Efecto de la selenometionina sobre la inducción de daño
Con el objeto de evaluar de qué manera influye la SeMet en la inducción de daño por bleomicina, tratamos los leucocitos con SeMet en la fase A, durante el tiempo inicial de cultivo, en la fase B, simultáneamente al tratamiento con bleomicina, y durante ambas fases AB. El daño fue posteriormente evaluado mediante el ensayo del cometa y los datos obtenidos se recogen en la Figura 10. Al tratar con SeMet durante la fase B de inducción de daño y conjuntamente en las fases AB de cultivo y de inducción, el daño disminuye con respecto al detectado al emplear sólo bleomicina, llegando a resultar este descenso significativo en el último caso. Sin embargo, el tratamiento durante la fase A de cultivo resultó en un incremento significativo de daño.
Después de realizar la incubación con la enzima hOGG1 los tratamientos con SeMet en las fases A y B experimentaron un ligero descenso de daño

68 Revista Real Academia Galega de Ciencias. Vol. XXVII
no significativo (Figura 11). Sin embargo, el tratamiento durante ambas fases conjuntamente produjo un aumento significativo en el %TDNA. La incubación con la enzima generó incremento en el daño encontrado respecto a la incubación sólo con buffer en el tratamiento sólo con bleomicina y con este compuesto junto con SeMet durante las fases B y AB.
Figura 10. Efecto de la SeMet sobre la inducción de daño por bleomicina (BLM). bP≤0.01, diferencia significativa respecto al tratamiento sólo con bleomicina.
Figura 11. Efecto de la SeMet sobre la inducción de daño por bleomicina (BLM) tras incubación con hOGG1. aP≤0.05, bP≤0.01, diferencia significativa respecto al tratamiento sólo con bleomicina; dP≤0.01, diferencia significativa respecto al mismo tratamiento sin emplear enzima.
4.4. Efecto de la selenometionina sobre la reparación del daño Para evaluar la influencia de la SeMet sobre la capacidad de reparación
de los leucocitos, evaluamos el daño existente tras el periodo de reparación

69Revista Real Academia Galega de Ciencias. Vol. XXVII
habiendo tratado con SeMet de nuevo en las fases A, B, AB y ahora también en la fase C, durante la reparación, y en la combinación de las tres fases ABC. Los resultados obtenidos se muestran en la Figura 12.Tras la reparación, los tratamientos con bleomicina y SeMet en las fases C, AB y ABC muestran un descenso del daño evaluado en relación con el tratamiento sólo con bleomicina, resultando significativo únicamente en el segundo caso.
Figura 12. Efecto de la SeMet sobre la reparación del daño inducido por bleomicina (BLM). bP≤0.01, diferencia significativa respecto al tratamiento sólo con bleomicina.
Los datos recogidos tras la incubación con la enzima hOGG1 nos aportan información específica acerca la influencia de la SeMet sobre la reparación del daño oxidativo inducido previamente por la bleomicina. Los resultados pueden observarse en la Figura 13.
Figura 13. Efecto de la SeMet sobre la reparación del daño inducido por bleomicina (BLM) tras incubación con hOGG1. aP≤0.05, bP≤0.01, diferencia significativa respecto al tratamiento sólo con bleomicina; cP≤0.05, dP≤0.01, diferencia significativa respecto al mismo tratamiento sin emplear enzima.
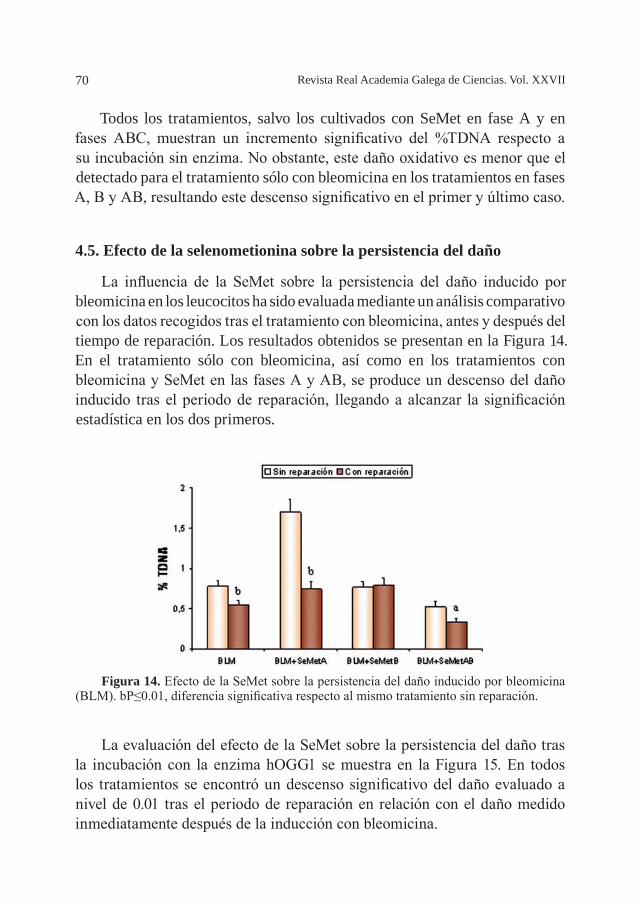
70 Revista Real Academia Galega de Ciencias. Vol. XXVII
Todos los tratamientos, salvo los cultivados con SeMet en fase A y en fases ABC, muestran un incremento significativo del %TDNA respecto a su incubación sin enzima. No obstante, este daño oxidativo es menor que el detectado para el tratamiento sólo con bleomicina en los tratamientos en fases A, B y AB, resultando este descenso significativo en el primer y último caso.
4.5. Efecto de la selenometionina sobre la persistencia del daño
La influencia de la SeMet sobre la persistencia del daño inducido por bleomicina en los leucocitos ha sido evaluada mediante un análisis comparativo con los datos recogidos tras el tratamiento con bleomicina, antes y después del tiempo de reparación. Los resultados obtenidos se presentan en la Figura 14. En el tratamiento sólo con bleomicina, así como en los tratamientos con bleomicina y SeMet en las fases A y AB, se produce un descenso del daño inducido tras el periodo de reparación, llegando a alcanzar la significación estadística en los dos primeros.
Figura 14. Efecto de la SeMet sobre la persistencia del daño inducido por bleomicina (BLM). bP≤0.01, diferencia significativa respecto al mismo tratamiento sin reparación.
La evaluación del efecto de la SeMet sobre la persistencia del daño tras la incubación con la enzima hOGG1 se muestra en la Figura 15. En todos los tratamientos se encontró un descenso significativo del daño evaluado a nivel de 0.01 tras el periodo de reparación en relación con el daño medido inmediatamente después de la inducción con bleomicina.

71Revista Real Academia Galega de Ciencias. Vol. XXVII
Figura 15. Efecto de la SeMet sobre la persistencia del daño inducido por bleomicina (BLM) tras incubación con hOGG1. bP≤0.01, diferencia significativa respecto al mismo tratamiento sin reparación.
4.6. Eficacia de reparación
Los resultados obtenidos pueden también analizarse utilizando el índice Eficacia de Reparación (RE), que viene determinado por la relación entre la diferencia entre el daño inducido y reparado y el daño inducido (Cebulska-Wasilewska et al.,2003). Se calcula según la fórmula:
en donde %TDNATto0 indica el daño en el ADN detectado tras el tratamiento correspondiente sin reparación, y %TDNATto15 indica el daño en el ADN para el mismo tratamiento después de los 15min de reparación.
En la Figura 16 se muestran los resultados obtenidos para la RE, tanto respecto al daño general en el ADN como respecto al daño específicamente oxidativo, evaluado tras la incubación con la enzima hOGG1. Todos los tratamientos con SeMet incrementan la RE del daño en el ADN respecto al tratamiento sólo con bleomicina, excepto el realizado en fase B, aunque no significativamente. De igual forma la RE del daño oxidativo aumentó en todos los tratamientos, excepto durante la fase C, llegando a alcanzar la significación estadística en la fase AB.

72 Revista Real Academia Galega de Ciencias. Vol. XXVII
Figura 16. Influencia de la SeMet sobre la eficacia de reparación (%RE) del daño inducido por bleomicina. aP≤0.05, diferencia significativa respecto al tratamiento sólo con bleomicina.
5. DISCUSIÓN
5.1. Utilización de selenometionina
El Se es un elemento traza imprescindible para el correcto desarrollo de muchos procesos celulares. Entre las principales funciones que desempeña en el organismo está la de formar parte de las selenoproteínas, muchas de las cuales son enzimas de la maquinaria antioxidante y por tanto indispensables en los procesos de defensa celular (Letavoyavá et al., 2006). No obstante, según en la forma y concentración en la que se encuentre, el Se también puede resultar tóxico para las células. Aunque son varios los factores que influyen en la toxicidad del Se (edad, estado fisiológico del organismo, ruta de administración, etc.), en general se piensa que ésta se fundamenta en dos razones principales: por un lado la capacidad de producir especies reactivas de oxígeno (ROS) durante su metabolismo (Spallholz, 1994 y 1997), y por otro la reducción a otros compuestos mutagénicos que se forman a partir de los compuestos originales de Se (Whiting et al., 1980).
A pesar de esta toxicidad en determinadas condiciones, se ha descrito un posible papel anticancerígeno del Se (Clark et al., 1996; Yu et al., 1997; Feng et al., 1999; Finley y Davis, 2001; Reid et al., 2002). Los mecanismos sugeridos para la prevención de cáncer por el Se incluyen los efectos sobre la muerte celular programada, la reparación del ADN, su papel en las selenoenzimas,

73Revista Real Academia Galega de Ciencias. Vol. XXVII
los efectos sobre el sistema inmune, actividad antiangiogénica y la inhibición específica del crecimiento de células tumorales por algunos metabolitos del Se (Rayman, 2000 y 2005; El-Bayoumy, 2001; Whanger, 2004).
Se tiene constancia por muchos estudios de que el Se es tóxico en cantidades algo más altas que las recomendadas. Sin embargo está bien establecido que la administración de Se más efectiva para la protección contra el cáncer es a niveles elevados, a menudo llamados niveles supra-nutricionales o farmacológicos (Aboul-Fadl, 2005). Dada esta enorme paradoja resulta de vital importancia delimitar las condiciones de administración y realizar más estudios in vitro para definirlas, así como realizar estudios más exhaustivos que evalúen los posibles efectos genotóxicos a distintos niveles y la influencia sobre la maquinaria de reparación celular. Incrementar el número de investigaciones en este sentido podría evitar los resultados adversos que están dando algunos primeros estudios de incidencia de cáncer en humanos utilizando suplementos de Se (Duffield-Lillico et al., 2003).
Varios son los motivos por los que se ha escogido la SeMet frente a otras formas químicas del Se para llevar a cabo este estudio. En primer lugar los alimentos ricos en Se suelen presentar mayoritariamente esta forma química por lo que, de entre todas las variantes del Se, la SeMet resulta la forma más adecuada para llevar a cabo una aportación de este elemento en la dieta (Schrauzer, 2000a; Letavayová et al., 2006). Otras características, principalmente metabólicas, avalan esta afirmación: se incorpora de forma inespecífica a la estructura de las proteínas, lo que permite el almacenamiento celular y tisular de Se para su empleo posterior, no genera tantas especies reactivas de oxígeno como otros compuestos de Se que siguen rutas metabólicas distintas, es incorporado al interior de las células más rápidamente que otros compuestos de Se como el selenito o la selenocisteína, su metabolismo específico hace que se requieran unos niveles más altos de SeMet que de otras formas de Se para dar lugar a los mismos efectos tóxicos (Beilstein y Whanger, 1987; Smith et al., 2004; Letavoyavá et al., 2006) y es hidrolizada a metilselenol, un metabolito activo del Se presuntamente responsable de su actividad quimiopreventiva (Ip, 1998; Ip et al., 1999; Wang et al., 2002; Cho et al., 2004). Además, la SeMet se administra vía oral (Ip, 1999) y los resultados obtenidos de ensayos clínicos en fase I indican que la SeMet puede ser administrada en dosis bastante elevadas sin llegar a producir toxicidad significativa (Reid et al., 2004; Fakih et al., 2006).

74 Revista Real Academia Galega de Ciencias. Vol. XXVII
En humanos, uno de los primeros estudios con SeMet como suplemento de la dieta (Griffiths et al., 1976) ya puso de manifiesto los beneficios que podía presentar este compuesto al concluir que prácticamente la totalidad de la SeMet era retenida por el organismo, siendo la eliminación de Se en la orina y en las heces muy pequeña. Observaron también que, al comparar estos resultados con los obtenidos en un estudio previo siguiendo el mismo protocolo (Thomson y Stewart, 1974), la asimilación de la SeMet era mucho mayor que la del selenito. Los posteriores resultados del trabajo de Thomson et al. (1993) en mujeres también apoyan el uso de SeMet como suplemento en humanos por ser más efectiva que otros compuestos de Se, debido a que su asimilación y biodisponibilidad para las células era mayor. Otro estudio posterior realizado nuevamente en mujeres (Robinson et al., 1997) añadió a estas observaciones previas, la capacidad de la SeMet de contribuir a mantener los niveles de Se en el organismo sin efectos adversos para el mismo.
Algunos estudios con SeMet in vitro e in vivo ya apuntan a un posible efecto beneficioso de este compuesto (Rafferty et al., 1998; Seo et al., 2002a y b), sin embargo debido a la gran controversia que genera y a los múltiples resultados que da el empleo del Se, es necesario realizar estudios más exhaustivos que evalúen los posibles efectos genotóxicos a distintos niveles y la influencia sobre la maquinaria de reparación celular.
A lo largo de este estudio se han evaluado los efectos de la exposición a la SeMet sobre el material genético y sobre la maquinaria de reparación celular de leucocitos humanos. Se ha estudiado la relación de este compuesto con el daño inducido en el ADN en general y también con el daño específicamente oxidativo. También se evaluó la influencia de la SeMet sobre la capacidad de los leucocitos para reparar estas lesiones inducidas previamente por un agente radiomimético: la bleomicina.
5.2. Genotoxicidad de la selenometionina
Para evaluar el efecto de la SeMet sobre el ADN y determinar la concentración óptima para su empleo posterior se ha seleccionado el ensayo del cometa como test de genotoxicidad. Este ensayo en su versión alcalina se considera como un estimador muy sensible del daño en el ADN inducido por una gran cantidad de compuestos genotóxicos, ya que detecta de manera inespecífica las roturas de cadena sencilla y doble y los sitios apurínicos o apirimidínicos sensibles al álcali que se generan a partir de aductos

75Revista Real Academia Galega de Ciencias. Vol. XXVII
(Singh et al., 1988; Olive et al., 1992). Pero además este ensayo permite también, mediante pequeñas modificaciones en su protocolo, evaluar de forma inespecífica distintos tipos de lesiones en el ADN como: entrecruzamientos entre ambas cadenas, presencia de bases modificadas, daño oxidativo, etc. (Collins et al., 2001; Olive y Banáth, 2006). Esta versatilidad, junto con la sencillez y sensibilidad del método, son las que han hecho que se haya considerado este ensayo como el idóneo para realizar el presente estudio.
El parámetro cuantitativo que se ha empleado como estimador del daño ocasionado sobre el ADN ha sido el porcentaje de ADN en la cola del cometa (%TDNA), relacionado de forma directa con la cuantía de roturas de cadena sencilla que se producen por efecto de un agente genotóxico (Singh et al., 1988). El %TDNA es el parámetro más útil, pues conlleva una relación lineal con la frecuencia de roturas, no se ve afectado relativamente por los ajustes del umbral de fluorescencia, y permite la discriminación del daño en el rango más amplio posible (en teoría de 0 a 100%), además aporta una indicación muy clara de la apariencia real de los cometas (Collins, 2004). Algunos autores han sugerido además que en determinados casos donde la longitud de la cola del cometa no distingue claramente entre las distintas dosis, entonces la cuantificación del %TDNA puede representar una variación más clara del daño. Por otro lado, las lesiones inducidas en el ADN muestran colas distintas en el ensayo del cometa según el agente que las produzca. Los radicales de oxígeno por ejemplo generan una cola de longitud similar a la de otros agentes pero de diferente intensidad, por lo que en el caso de estudiar daño oxidativo es preferible escoger el %TDNA, que tiene en cuenta esa intensidad al evaluar el daño (Olive et al., 1992; Mckeley-Martin et al., 1993; Anderson et al., 1994).
Algunos estudios con Se han dado como resultado un daño oxidativo en las células u organismos estudiados como consecuencia de la exposición a este compuesto. Se ha llegado a la conclusión de que la toxicidad del Se se manifestará de forma aguda o crónica cuando el daño oxidativo que provoca supere las defensas antioxidantes que posee la célula (Nogueira et al., 2004). El balance entre prooxidantes y antioxidantes, no sólo en el caso del Se sino de cualquier compuesto, es crucial para la supervivencia y la funcionalidad en organismos aerobios. Un desequilibrio favoreciendo a los prooxidantes y/o desfavoreciendo antioxidantes es lo que se conoce como estrés oxidativo (Arteel y Sies, 2001). Con el fin de determinar si la SeMet induce la producción ROS provocando con ello la consecuente aparición del

76 Revista Real Academia Galega de Ciencias. Vol. XXVII
daño oxidativo, introducimos el empleo de la enzima hOGG1 en el ensayo del cometa original para evaluar de forma específica este tipo de daño.
La enzima hOGG1 es una glicosilasa de la maquinaria de reparación de células de mamíferos que participa en la vía de reparación por escisión de bases. Esta enzima es capaz de reconocer específicamente las guaninas modificadas por la adición de un grupo hidroxilo a la posición 8, originando 8-hidroxi-2’-desoxiguanosina (8-OHdG). Cuando el balance oxidativo celular se desestabiliza, bien por causas endógenas bien por agentes exógenos, se generan un elevado número de ROS que pueden interaccionar con el ADN desestabilizándolo y dañándolo. La guanina es la base más propensa a la oxidación y, de los 20 aductos de ADN producto del estrés oxidativo conocidos, la formación de 8-OHdG es el daño más frecuente (Angerer et al., 2007). La importancia de reconocer específicamente las bases 8-OHdG, mediante del empleo de la enzima hOGG1, radica por tanto en el hecho de que se trata de elementos potencialmente mutagénicos y de que su presencia en el ADN es consecuencia directa de la exposición a ROS por lo que son consideradas como un buen biomarcador de estrés oxidativo (Boiteux y Radicella, 1999; Kuo et al., 2007).
Las enzimas más frecuentemente utilizadas, hasta la fecha, para la detección de daño oxidativo mediante el ensayo del cometa son la endonucleasa III (endoIII), que reconoce pirimidinas oxidadas (Collins et al., 1993), y la formamidopirimidina ADN-glicosilasa (fpg), que es específica para purinas oxidadas, incluyendo la 8-OHdG (Collins et al., 1996). Sin embargo, en este estudio se ha utilizado hOGG1 debido a que recientemente se ha demostrado su mayor especificidad frente al daño oxidativo, ya que tanto la endoIII como la fpg también reconocen daño provocado por alquilación (Smith et al., 2006).
Durante el ensayo del cometa se incubaron los leucocitos con hOGG1 con el fin que esta enzima reconociese las bases 8-OHdG indicativas de daño oxidativo y las eliminase de la secuencia de ADN. Tras su actuación se generaron sitios abásicos que dieron lugar, debido a las condiciones fuertemente alcalinas en las que se desarrolla el ensayo, a roturas de cadena. Así, la cantidad de ADN presente en las colas de los cometas generados tras la incubación con esta enzima será mayor y representará el daño oxidativo presente en las células.

77Revista Real Academia Galega de Ciencias. Vol. XXVII
Los resultados obtenidos en el ensayo del cometa sin incubación con enzima (Figuras 5 y 6) muestran la existencia de una correlación negativa entre la dosis de SeMet empleada y el daño detectado, resultando este descenso significativo en las concentraciones más altas. Estos datos obtenidos sugieren por tanto que la SeMet no sólo no ejerce efectos genotóxicos sobre el ADN sino que por el contrario reduce el daño basal presente en las células estudiadas. Estos resultados apoyarían las hipótesis de otros autores que sugieren que una pequeña cantidad de SeMet en la dieta (inferior a 200 μg/día) puede proteger a la célula del daño endógeno (Schrauzer, 2001; Letavoyavá et al., 2006).
Tras el tratamiento con hOGG1 se encontró un incremento significativo en el promedio de %TDNA en las células del control y en las concentraciones de SeMet 1, 5 y 10μM con respecto a las mismas condiciones sin enzima (Figura 7), lo que apoya el uso de esta enzima junto con el ensayo del cometa como buen indicador del daño oxidativo en el ADN (Smith et al., 2006). Sin embargo, no se encontraron diferencias en los tratamientos 50 y 100 μM con respecto a la incubación con buffer, lo que sugiere un posible papel antioxidante de la SeMet en las concentraciones más altas evaluadas. Además, la concentración de SeMet 50 μM experimenta un descenso significativo respecto al daño oxidativo basal mostrado por el control, dato que señala nuevamente a un efecto protector de este compuesto frente al daño oxidativo. Los datos recogidos en la bibliografía muestran que los niveles de Se en los fluidos corporales (suero, plasma y sangre total) son más bajos en los pacientes con cáncer que en los sujetos control (revisado en Alaejos et al., 2000) indicando además que, frecuentemente, la diferencia entre las poblaciones no es mayor que las concentraciones de Se correspondientes a 50 y 100 µm de SeMet (4 y 8 mg/l). Nuestros resultados apoyan por tanto que la administración de un suplemento de SeMet para llegar a alcanzar estos niveles orgánicos puede ser beneficiosa para la prevención del cáncer.
Al contrario de estos resultados positivos encontrados en las concentraciones más altas testadas, tras la incubación con la enzima se encontró que la concentración 1μM mostraba un incremento significativo del daño oxidativo respecto al control sin SeMet. Trabajos anteriores ya describen efectos adversos de cantidades limitadas de SeMet en el organismo (Waters et al., 2005; Vega et al., 2007). Estos estudios concluyeron que a concentraciones demasiado bajas, al igual que ocurre en cantidades demasiado elevadas, la SeMet podía inducir daño en el ADN, disminuir la actividad de la

78 Revista Real Academia Galega de Ciencias. Vol. XXVII
glutation-peroxidasa produciendo un incremento en el daño oxidativo, causar alteraciones del sistema inmune y, en consecuencia de todo ello, aumentar el riesgo a desarrollar procesos neoplásicos.
Confirmando estudios anteriores que ya ponían de manifiesto la relevancia de la concentración y la forma química de Se empleadas (Shamberger, 1984; Ip, 1998; Schrauzer, 2001), vemos que las distintas concentraciones de SeMet testadas en este trabajo influyen en el efecto que ejerce este compuesto sobre el ADN. Al estudiar la genotoxicidad de la SeMet observamos que el daño evaluado disminuye conforme aumenta la concentración de SeMet, mientras que en el caso específico del estudio del daño oxidativo los efectos que ejercerá la SeMet son beneficiosos o perjudiciales según sea la concentración empleada. En base a los resultados obtenidos y debido a todas estas observaciones decidimos emplear la concentración de 50 μM durante los restantes ensayos del presente estudio, puesto que a esta concentración la SeMet no es genotóxica ni induce daño oxidativo en el ADN y, por el contrario, parece mostrar cierto efecto antioxidante.
5.3. El ensayo de competencia de reparación del ADN
El organismo está constantemente expuesto a numerosos agentes que pueden alterar su material genético favoreciendo la aparición y desarrollo de numerosas patologías, entre ellas el cáncer. Para protegerse de ese daño, las células poseen una serie de mecanismos de defensa y de reparación encargados de disminuir en la medida de lo posible las lesiones que causan estos agentes y de esta forma minimizar los efectos finales resultantes (Wood, 1996). Estos procesos de reparación del ADN representan mecanismos que permiten la supervivencia de la célula ya que, además de corregir los errores propios de la replicación, juegan un papel decisivo frente a los agentes tóxicos tanto exógenos (radiación, compuesto químicos, virus, etc.) como endógenos (radicales libres, metabolitos secundarios, etc.) que pueden afectar al genoma. Errores o deficiencias en estos procesos de reparación producen una cascada de acumulación de aberraciones en el genoma, afectando también a las propias proteínas que intervienen en la reparación y culminando en la llamada inestabilidad genética, punto de partida de multitud de procesos neoplásicos (Buermeyer et al., 1999; De Boer y Hoeijmakers, 2000).
La importancia de estudiar la capacidad individual de reparar lesiones en el ADN radica, por tanto, en la observación de que mecanismos deficientes de

79Revista Real Academia Galega de Ciencias. Vol. XXVII
reparación del genoma se han relacionado con la presencia de gran cantidad de patologías y procesos neoplásicos, y de que alteraciones en estos mecanismos pueden variar la susceptibilidad de los individuos expuestos a un determinado mutágeno. La incapacidad para reparar el ADN dañado es además la principal causa del cáncer (Au, 1993; Au et al., 1996a; Berwick y Vineis, 2000).
La relación entre los distintos compuestos de Se y la capacidad de reparación celular ha sido ya descrita y establecida (Stewart et al., 1999; El-Bayoumy, 2001; Hartwig et al., 2003). Muchos autores defienden que el papel protector que puede desempeñar el Se frente al cáncer y a otras patologías viene determinado por su presencia en las selenoenzimas, muchas de las cuales son importantes proteínas de la maquinaria de reparación celular y participan activamente en procesos de reparación de daño oxidativo (Combs Jr. y Gray, 1998; Ganther, 1999; Rayman 2005). Es por esto que en varios estudios de administración de Se en la dieta se han encontrado incrementos en la capacidad de reparación celular (Seo et al., 2002a; Abul-Hassan et al., 2004). No obstante, en contra de esta hipótesis está el trabajo de Gladyshev et al. (1998), que evaluaron la expresión de varias selenoproteínas antioxidantes en tres tipos de células cancerígenas y las compararon con células normales. La glutation-peroxidasa fue disminuyendo mientras que la tiorredoxín-reductasa fue aumentando en todas las células cancerígenas comparadas con todas las líneas celulares (o tejidos) control. Si las funciones antioxidantes de estas proteínas fueran de alguna forma la base de la actividad anticancerígena del Se, se podría esperar que hubiera una expresión en paralelo de ambas enzimas.
Para evaluar cómo afecta una exposición a SeMet a la maquinaria de reparación de leucocitos humanos hemos utilizado el ensayo de competencia de reparación del ADN in vitro. El ensayo de competencia de reparación del ADN es un ensayo citogenético en el que las células son forzadas, mediante distintos métodos de inducción, a reparar las lesiones producidas en el ADN. Evaluando estas lesiones y comparándolas con un control tendremos una estimación directa de la capacidad de reparación que poseen esas células (Au et al., 1991). Este ensayo puede ser utilizado para determinar una respuesta anormal de reparación del ADN debida, por ejemplo, a patologías como síndrome de Down; también es capaz de responder a exposiciones excesivas a tóxicos como indicador de deficiencia en la reparación del ADN (en el caso concreto del butadieno, por ejemplo, se llegó a establecer una relación causa-efecto (Au et al., 1995a)) y puede ser empleado en estudios in vivo e

80 Revista Real Academia Galega de Ciencias. Vol. XXVII
in vitro (Au et al., 1996b). Es además más sensible para detectar alteraciones biológicas por exposición a tóxicos que otros ensayos citogenéticos estándar (Au et al., 2001).
Aunque los mecanismos por los que se producen anomalías en la reparación son desconocidos, se cree que estas anomalías pueden estar causadas por un bloqueo de los procesos de reparación del ADN o por mutaciones en genes que codifican para enzimas de reparación (Au, 1993). Puesto que la reparación del ADN es un proceso que implica un gran número de enzimas y vías distintas, defectos en algún punto de la maquinaria, por pequeños que sean, afectarán a la capacidad final de reparar el daño, por lo que podrán ser detectados mediante el ensayo de competencia de reparación del ADN. Esta elevada sensibilidad de este ensayo permite también poner de manifiesto alteraciones en la capacidad de reparación muy tenues y difícilmente detectables por otros medios. Las exposiciones ambientales a bajas concentraciones de mutágenos pueden producir aberraciones cromosómicas que no se manifiestan de forma inmediata, pero pueden también causar alteraciones en los mecanismos de reparación. Esto implica que muchas células que tienen defectos en la reparación en principio muy sutiles, tras el ensayo de competencia de reparación se amplifican y pueden ser visualizados (Au, 2001). Por lo tanto, la extensa y multiplicada respuesta al ensayo de competencia de reparación hace de éste un método más sensible que los ensayos de genotoxicidad tradicionales (Au, 2007).
El empleo del ensayo del cometa como test de genotoxicidad en la realización del el ensayo de competencia de reparación del ADN constituye una poderosa herramienta para la determinación de la capacidad de reparación y presenta importantes ventajas frente a otros métodos. Además se estableció que la aplicación de este ensayo era un buen biomarcador de daño genotóxico y de capacidad de reparación de daño en el ADN (Cebulska-Wasilewska et al., 2005a). Lo realmente novedoso de esta combinación de técnicas y la ventaja frente a otras técnicas similares es la posibilidad de cuantificar la reparación por la evolución del daño a diferentes tiempos, realizando el ensayo inmediatamente después de tratar las células con el mutágeno y nuevamente tras el correspondiente periodo de reparación, durante el cual las células permanecen en cultivo (Rajaee-Behbahani et al., 2001; Schmezer et al., 2001). Se pueden realizar también otras evaluaciones de daño a tiempos intermedios para estudiar la dinámica de la reparación. Esta determinación del daño a diferentes tiempos nos permite establecer una relación entre ADN dañado y

81Revista Real Academia Galega de Ciencias. Vol. XXVII
ADN reparado, obteniendo así el porcentaje de reparación en cada población de estudio. Lo que significa que al combinar el ensayo de competencia de reparación con el ensayo del cometa obtenemos una medida directa y cuantitativa de la capacidad de reparación, que puede reflejarse mediante el empleo de distintos índices: de eficacia de reparación, de daño residual tras la reparación y de sensibilidad al mutágeno empleado como inductor (Cebulska-Wasilewska et al., 2005a y 2005b).
Para realizar el estudio del efecto de la SeMet sobre la capacidad de reparación celular mediante el ensayo de competencia de reparación hemos cultivado leucocitos con esta forma de Se en tres fases distintas del estudio (fase de cultivo A, fase de inducción de daño B y fase de reparación C) y combinaciones de éstas (fases AB y fases ABC), y posteriormente se evaluó el daño existente en las células para cada uno de los tratamientos, antes y después de reparar, y se comparó con un control. Para llevar a cabo el ensayo decidimos emplear la bleomicina como agente inductor de daño, el ensayo del cometa como test de genotoxicidad y el tratamiento con la enzima hOGG1 como medida específica del daño oxidativo.
5.3.1. Genotoxicidad de la bleomicina
Empleamos la bleomicina como agente inductor de daño en el ensayo de competencia de reparación porque es un conocido agente radiomimético que causa roturas de cadena sencilla y doble, así como daño oxidativo en el ADN por liberación de radicales libres (Milic y Kopjar, 2004). Las condiciones utilizadas en el ensayo, con y sin incubación con hOGG1, permiten determinar y estudiar por separado la dinámica de ambas lesiones. Por otro lado, la bleomicina es un medicamento empleado en quimioterapia por lo que, teniendo en cuenta que el Se es un conocido agente quimiopreventivo, se pueden relacionar los resultados obtenidos en este estudio con los efectos anticancerígenos del Se empleado como suplemento en la dieta de pacientes con cáncer y establecer una posible interacción entre ambos compuestos. Varios estudios clínicos empleando Se mostraron que éste influye sobre los efectos de los agentes quimioterapeúticos usados en tratamientos contra el cáncer (Aboul-Fadl, 2005). Duffiel-Lillico et al. (2003) realizaron un amplio estudio in vivo sobre 1312 hombres y mujeres con un historial de dos o más carcinomas de células basales o uno o más carcinomas de células escamosas de piel a los que durante 10 años se suplementó en la dieta para estudiar si

82 Revista Real Academia Galega de Ciencias. Vol. XXVII
este suplemento podía prevenir el cáncer de piel no melanómico. Tras este tiempo observaron que no sólo no había diferencias significativas entre los suplementados con Se y los tratados con placebo, sino que el suplemento aumentaba significativamente el riesgo de carcinoma de células escamosas y de cáncer de piel no melanómico, y apuntaron como posible causa de esto la interacción con los medicamentos empleados para tratar estas patologías. Concluyeron que era posible que esos fármacos anularan o incluso revertieran los efectos del Se administrado.
Los resultados del tratamiento con bleomicina (Figura 8) muestran un incremento significativo del daño en el ADN con respecto al control tanto antes como después del periodo de reparación. No obstante, tras la reparación, el porcentaje de ADN en la cola ha disminuido significativamente con respecto al detectado inmediatamente después de la inducción con bleomicina. Tras la incubación con hOGG1 (Figura 9), el daño presente también es significativamente mayor antes y después de reparar con respecto al control sin bleomicina. Pero, nuevamente, tras la reparación se reduce significativamente el daño medido. Estos resultados coinciden con las aportaciones de Povirk y Austin (1991) y Milic y Kopjar (2004), que defienden que la bleomicina es un potente agente genotóxico que induce roturas de cadena y daño oxidativo en el ADN celular, y con los trabajos de Schmezer et al. (2001) y Glei y Pool-Zobel (2006), que concluyen que la mayor parte del daño inducido por la bleomicina se repara en los primeros 15 min tras el tratamiento.
5.3.2. Efecto de la selenometionina sobre la inducción de daño
Los datos obtenidos para cada uno de los tratamientos con SeMet en las diferentes fases del estudio (Figura 10) muestran que este compuesto influye en la inducción del daño que la bleomicina ejerce sobre el ADN de los leucocitos estudiados. En los tratamientos con SeMet en fase de cultivo A, este daño es significativamente mayor mientras que disminuye en los tratamientos con SeMet en la fase de inducción de daño B y en ambas juntas AB respecto al tratamiento sólo con bleomicina, significativamente en el último caso.
Tras el tratamiento con la enzima (Figura 11) ocurre lo contrario, en el tratamiento con SeMet en fase A el daño oxidativo detectado es menor que el que presentan las células del tratamiento sólo con bleomicina mientras que en las fases B y AB, este daño aumenta. Estos resultados indican que la SeMet podría actuar como elemento protector frente a las roturas de cadena

83Revista Real Academia Galega de Ciencias. Vol. XXVII
inducidas si el cultivo con SeMet se realiza durante la fase B o conjuntamente en las fases A y B y como antioxidante si los leucocitos son cultivados con este compuesto previamente al tratamiento con bleomicina. Estos resultados coinciden con otros previos en donde se describe un efecto protector de la SeMet frente al daño inducido por varios agentes, químicos o físicos, en el ADN (Roussyn et al., 1996; Rafferty et al., 2003). Concuerdan también con otro estudio que muestra las propiedades antigenotóxicas de este compuesto de Se sobre el daño inducido en el ADN por la doxorrubicina, un fármaco anticancerígeno, tanto en ratas (Santos et al., 2007) como en linfocitos humanos (Santos y Takahashi, 2008). En relación a otros fármacos quimioterapéuticos, Rao et al. (1998) demostraron que la SeMet protege contra la nefrotoxicidad inducida por el cisplatino sin interferir en su actividad antitumoral, y Cao et al. (2004) concluyeron en su estudio que la SeMet protege selectivamente contra la toxicidad inducida por varios agentes quimioterapéuticos al tiempo que aumenta la actividad antitumoral y la tasa de curación.
5.3.3. Efecto de la selenometionina sobre la reparación del daño
El tiempo permitido para la reparación del daño inducido por bleomicina se fijó en 15~min puesto que éste es el tiempo establecido por Schmezer et al. (2001) como el óptimo para analizar la reparación del daño inducido por la exposición a bleomicina. Además, estudios previos realizados en nuestro laboratorio también confirman este hecho, ya que no se encontraron diferencias significativas entre el daño evaluado tras un periodo de reparación de 15 min frente al obtenido tras un periodo de reparación de 30 min.
Los resultados obtenidos después del periodo de reparación (Figuras 12 y 13) nos indican que la SeMet favorece la reparación del daño causado por la bleomicina, ya sean roturas de cadena o daño oxidativo, especialmente cuando se aplica durante la fase de cultivo previa al tratamiento conjuntamente con la fase de inducción de daño. Si el tratamiento con SeMet ocurre durante la fase de reparación sucede que aumenta la reparación de daño en general pero disminuye la reparación específica de daño oxidativo, llegando a alcanzar un incremento significativo de este daño con respecto al tratamiento sólo con bleomicina.
El papel de Se en la reparación del ADN fue puesto de manifiesto por primera vez cuando se demostró que un tratamiento con Se mejoraba la capacidad de reparación de células previamente dañadas con radiación UV

84 Revista Real Academia Galega de Ciencias. Vol. XXVII
debido al aumento en la actividad de los complejos proteicos encargados de dicha reparación (Seo et al., 2002a). Otro estudio demostró recientemente que el tratamiento previo con SeMet induce una respuesta de reparación del ADN dependiente de p53, elevando la expresión de las proteínas responsables del reconocimiento del ADN dañado e incrementando la tasa de reparación celular, protegiendo de esta manera al ADN frente a las exposiciones posteriores a agentes inductores de daño (Fischer et al., 2007). Inducir la reparación del ADN podría ser un mecanismo de quimioprevención que muy pocos compuestos han demostrado tener (Collins et al., 2003). Zhang et al. (2008) indicaron además que el Se sólo aumenta la reparación del ADN en los tejidos normales como consecuencia de la modulación selectiva del Se sobre el Nrf2 de tejidos tumorales y normales (Kim et al., 2007), lo que apoya el empleo de un suplemento de SeMet en los pacientes de cáncer tratados con fármacos quimioterapéuticos.
No obstante, los resultados de esta parte del estudio ponen también de manifiesto que, según la fase en la que se administre, la SeMet puede actuar reduciendo o incrementando el daño oxidativo en las células. Esto coincide con lo descrito en otros trabajos que señalan el doble papel biológico que puede representar el Se (Miller et al., 2001; Drake, 2006). Este compuesto puede ejercer un efecto prooxidante o antioxidante, llegando en consecuencia a resultar nocivo o beneficioso para el organismo receptor en base a las características que presente el compuesto y al propio organismo. El Se actúa como antioxidante formando parte de las selenoproteínas, muchas de las cuales son importantes enzimas de la maquinaria de reparación celular como la glutation-peroxidasa y la tiorredoxin-reductasa. Y actúa como prooxidante generando ROS y promoviendo en consecuencia la oxidación del ADN in vivo (Letavayová et al., 2006).
5.3.4. Efecto de la selenometionina sobre la persistencia de daño
Al comparar los datos recogidos en nuestro ensayo en los distintos tratamientos antes y después de reparar vemos que, tras la reparación el daño originalmente inducido por la bleomicina se reduce en todos los tratamientos salvo en el cultivo con SeMet en fase B (Figura 14). Al estudiar específicamente el daño oxidativo reparado vemos que de nuevo en todas las fases del estudio sin excepción se reduce significativamente el daño oxidativo medido tras el periodo de reparación con respecto al existente inmediatamente después del

85Revista Real Academia Galega de Ciencias. Vol. XXVII
tratamiento con bleomicina (Figura 15). Se verifica así que el tiempo permitido en el estudio para la reparación es generalmente suficiente para reparar las roturas de cadena simple y doble, los sitios álcali-lábiles y el daño oxidativo inducido por la bleomicina.
La influencia de la SeMet sobre la persistencia del daño inducido por la bleomicina en cada uno de los tratamientos es diferente. En la evaluación del daño global en el ADN, en el tratamiento con SeMet en fase A el descenso del %TDNA detectado tras la reparación es mayor que el evaluado en el tratamiento sólo con bleomicina por lo que la persistencia del daño es considerablemente menor, en la fase AB apenas se aprecia variación y en la fase B aumenta la persistencia con respecto al tratamiento sólo con bleomicina. Tras la incubación con la enzima hOGG1, en la fase A aumenta la persistencia de daño, en la fase B se mantiene y en la fase AB disminuye con respecto a los valores obtenidos en el tratamiento sólo con bleomicina. Estas observaciones indican que el cultivo con SeMet en fase A favorece el descenso del daño general inducido con bleomicina mientras que el tratamiento con este compuesto en las fases A y B conjuntamente favorecería una reparación específica del daño oxidativo. El cultivo en fase B, en cambio, no sería beneficioso en ninguno de los dos tipos de daño. Estos resultados confirmar la idoneidad del tratamiento con SeMet en las fases A y B conjuntamente para obtener efectos protectores contra el daño inducido por la bleomicina.
5.3.5. Eficacia de reparación
Los resultados obtenidos en este estudio se pueden exponer también mediante el empleo de un índice llamado “eficacia de reparación” (RE, repair efficence) (Cebulska-Wasilewska et al., 2003). Mediante la representación gráfica de los índices RE calculados podemos ver que la influencia de la SeMet en la capacidad de reparación celular para cada uno de los tratamientos del estudio (Figura 16).
En relación a la eficacia de la reparación del daño en general, los índices indican que la SeMet aumenta la eficacia de reparación en todos los tratamientos, salvo en el cultivo en fase B, aunque este incremento no llega en ningún caso a alcanzar significación estadística. En cuanto al daño específicamente oxidativo, el cultivo con SeMet en cualquiera de las fases o combinaciones de éstas aumenta la eficacia de reparación de las células, especialmente en el caso concreto del tratamiento conjunto en las fases A y

86 Revista Real Academia Galega de Ciencias. Vol. XXVII
B. Estos resultados ponen de manifiesto el efecto positivo que ejerce la SeMet sobre la capacidad de reparación del daño inducido en los leucocitos del estudio, especialmente cuando se aplica antes o simultáneamente con la bleomicina, y apoyan los mecanismos de quimioprevención de la SeMet basados tanto en sus propiedades antioxidantes como su capacidad de interferir en las vías de reparación del ADN (Santos y Takahashi, 2008).
6. CONCLUSIONES
En resumen, en el presente trabajo se han obtenido las siguientes conclusiones:
1. La SeMet no ejerce actividad genotóxica sobre leucocitos humanos en el rango de dosis evaluado, presentando una relación dosis-respuesta negativa que determina la existencia de cierto efecto protector a las dosis de 50 y 100µM, tanto sobre el daño en el ADN en general como sobre el daño oxidativo.
2. Se ha confirmado la validez de la bleomicina como agente inductor de daño en el ADN y de daño oxidativo, y del periodo de 15 min como el más adecuado para que se produzca la reparación de la mayor parte del daño inducido por este compuesto.
3. La presencia de SeMet ejerce un efecto protector frente a la inducción, reparación y persistencia del daño en el ADN y del daño oxidativo inducidos por la bleomicina, especialmente si el tratamiento se realiza durante las fases de cultivo e inducción de daño conjuntamente.
AGRADECIMIENTOS
Este trabajo ha sido financiado por un proyecto de la Xunta de Galicia (INCITE08PXIB106155PR). Durante su realización, V. Valdiglesias ha disfrutado de una beca predoctoral de la Universidade da Coruña.

87Revista Real Academia Galega de Ciencias. Vol. XXVII
7. BIBLIOGRAFIA
ABOUL-FADL, T. (2005) Selenium derivatives as cancer preventive agents. Curr. Med. Chem. 5, 637-652.
ABUL-HASSAN, K.S., LEHNERT, B.E., GUANT, L. Y WALMSLEY, R. (2004) Abnormal DNA repair in selenium-treated human cells. Mutat. Res. 565, 45-51.
ALAEJOS, M.S., DIAZ ROMERO, F.J. Y DIAZ ROMERO, C. (2000) Selenium and cancer: some nutritional aspects. Nutrition 16, 376-383.
AMMAR, E.M. Y COURI, D. (1981) Acute toxicity of sodium selenite and selenomethionine in mice after ICV or IV administration. Neurotoxicology 2, 383-386.
ANDERSON, D. YU, T.-W., PHILLIPS, B.J. Y SCHMEZER, P. (1994) The effect of various antioxidants and other modifying agents on oxygen-radical-generated DNA damage in human lymphocytes in the comet assay. Mutat. Res. 307, 261-271.
ANGERER, J., EWERS, U. Y WILHELM, M. (2007) Human biomonitoring: stage of the art. Int. J. Hyg. Environ. Health 210, 201-228.
ARTEEL, G.E. Y SIES, H. (2001) The biochemistry of selenium and the glutathione system. Environ. Toxicol. Pharmacol. 10, 153-158.
ARTHUR, J.R., MCKENZIE, R.C. Y BECKETT, G.J. (2003) Selenium in the inmune system. J. Nutr. 133, 1457-1459.
AU, W.W. (2001) Life style factors and acquired susceptibility to environmental disease. Int. J. Hyg. Environ. Health 204, 17-22.
AU, W.W. (1993) Abnormal chromosome repair and risk of developing cancer. Environ. Health Perspect. 101 (Suppl.3), 303-308.
AU, W.W. (2003) Mutagen sensitivity assays in population studies. Mutat. Res. 544, 273-277.
AU, W.W. (2007) Usefulness of biomarkers in population studies: from exposure to susceptibility and to prediction of cancer. Int. J. Hyg. Environ. Health 210, 239-246.
AU, W.W., WALKER, D.M., WARD JR., J.B., WHORTON, E., LEGATOR, M.S. Y SINGH, V. (1991) Factors contributing to chromosome damage in lymphocytes of cigarette smokers. Mutat. Res. 260, 137-144.
AU, W.W., BECHTOLD, W.E., WHORTON, E.B. Y LEGATOR, M.S. (1995a) Chromosome aberrations and response to gamma-ray challenge in lymphocytes of workers exposed to 1,3-butadiene. Mutat. Res. 334, 125-130.
AU, W.W., LANE, R.G., LEGATOR, M.S., WHORTON, W.B., WILKINSON, G.S. Y GABEHART, G.J. (1995b) Biomarker monitoring of a population residing near uranium mining activities. Environ. Health Perspect. 103, 466-470.
AU, W.W., WILKINSON, G.S., TYRING, S.K., LEGATOR, M.S., EL ZEIN, R., HALLBERG, L. Y HEO, M.Y. (1996a) Monitoring populations for DNA repair deficiency and for cancer susceptibility. Environ. Health Perspect. 104 (Suppl. 3), 579-584.
AU, W.W., RODRIGUEZ, G., ROCCO, C., LEGATOR, M.S. Y WILKINSON, G.S. (1996b)Chromosome damage and DNA repair response in lymphocytes of women who had children with neural tube defects. Mutat. Res. 361, 17-21.
AU, W.W., OH, H.Y., GRADY, J., SALAMA, S.A. Y HEO, M.Y. (2001) Usefulness of genetic susceptibility and biomarkers for evaluation of environmental health risk. Environ. Mol. Mutagen. 37, 215-225.

88 Revista Real Academia Galega de Ciencias. Vol. XXVII
AYKIN-BURNS, N. Y ERCAL, N. (2006). Effects of selenocystine on lead-exposed Chinese hamster ovary (CHO) and PC-12 cells. Toxicol. Appl. Pharmacol. 4, 136-143.
AYOUNI, L., BARBIER, F., IMBERT, J.L., LANTÉRI, P. Y GRENIER-LOUSTALOT, M.F.. (2007) Speciation of selenium in a commercial dietary supplement by liquid chromatography coupled with inductively coupled plasma-mass spectrometry (ICP-MS). J. Toxicol. Environ. Health 70, 735-741.
BECK, M.A., LEVANDER, O.A. Y HANDY, J. 2003. Selenium deficiency and viral infection. J. Nutr. 133, 1463-1467.
BEILSTEIN, M.A. Y WHANGER, P.D. (1987) Metabolism of selenomethionine and effects of interacting compounds by mammalian cells in culture. J. Inorg. Biochem. 29, 137-52.
BERWICK, M. Y VINEIS, P. (2000) Markers of DNA repair and susceptibility to cancer in humans: an epidemiologic review. J. Natl. Cancer Inst. 92, 874-897.
BISWAS, S., TALUKDER, G. Y SHARMA, A. (1999) Comparison of clastogenic effects of inorganic selenium salts in mice in vivo as related to concentrations and duration of exposure. Biotechnol. Tech. 12, 361-368.
BISWAS, S., TALUKDER, G. Y SHARMA, A. (2000) Chromosome damage induced by selenium salt in human peripheral lymphocytes. Toxicol. in Vitro 14, 405-408.
BLESSING, H., KRAUS, S., HEINDL, P., BAL, W. Y HARTWIG, A. (2004) Interaction of selenium compounds with zinc finger proteins involved in DNA repair. Eur. J. Biochem. 271, 3190-3199.
BLOT, W.J., LI, J.Y., TAYLOR, P.R., GUO, W., DAWSEY, S., WANG, G.Q., YANG, C.S., ZHENG, S.F., GAIL, M., LI, G.Y., YU, Y., LIU, B.Q., TANGREA, J., SUN, Y.H., LIU, F., FRAUMENI, J.F., ZHANG, Y.H. Y LI, B. (1993) Nutrition intervention trials in Linxian, China: supplementation with specific vitamin/mineral combinations, cancer incidence, and disease-specific mortality in the general population. J. Natl. Cancer Inst. 85, 1483-1492.
BOITEUX, S. Y RADICELLA, P. (1999) Base excision repair of 8-hydroxyguanine protects DNA from endogenous oxidative stress. Biochimie. 81, 59-67.
BRIVIBA, K., ROUSSYN, I., SHAROV, V.S. Y SIES, H. (1996) Attenuation of oxidation and nitration reactions of peroxynitrite by selenomethionine, selenocystine and ebselen. Biochem. J. 319, 13-15.
BROWN, K.M., PICKARD, K., NICOL, F., BECKETT, G.J., DUTHIE, G.G. Y ARTHUR, J.R. (2000) Effects of organic and inorganic selenium supplementation on selenoenzyme activity in blood lymphocytes, granulocytes, platelets and erythrocytes. Clin. Sci. 98, 593-599.
BUERMEYER, A.B., DESCHENES, S.M., BAKER, S.M. Y LISKAY, R.M. (1999) Mammalian DNA mismatch repair. Annu. Rev. Genet. 33, 533-564.
CÃO, S., DURRANI, F.A. Y RUSTUM, Y.M. (2004) Selective modulation of the therapeutic efficacy of anticancer drugs by selenium containing compounds against human tumor xenografts. Clin. Cancer Res. 10, 2561-2569.
CEBULSKA-WASILEWSKA, A. (2003) Response to challenging dose of X-rays as a predictive assay for molecular epidemiology. Mutat. Res. 544, 289-297.
CEBULSKA-WASILEWSKA, A. PANEK, A., ZABINSKI, Z., MOSZCZYNSKI, P. Y AU, W.W. (2005b) Occupational exposure to mercury vapour on genotoxicity and DNA repair. Mutat. Res. 586, 102-114.
CEBULSKA-WASILEWSKA, A., WIECHEC, A., PANEK, A., BINKOVÁ, B., SRÁM, R. J. Y FARMER, P.B. (2005a) Influence of enviromental exposure to PAHs on the susceptibility of lymphocytes to DNA-damage induction and on their repair capacity. Mutat. Res. 588, 73-81.
CHO, S.D., JIANG, C., MALEWICZ, B., DONG, Y., YOUNG, C.Y., KANG, K.S., LEE, Y.S., IP, C. Y LU, J. (2004) Methyl selenium metabolites decrease prostate-specific antigen expression by inducing protein degradation and suppressing androgen-stimulated transcription. Mol. Cancer Ther. 3, 605-611.

89Revista Real Academia Galega de Ciencias. Vol. XXVII
CLARK, L.C., COMBS JR, G.F., TURNBULL, B.W., SLATE, E.H., CHALKER, D.K., CHOW, J., DAVIS, L.S., GLOVER, R.A., GRAHAM, G.F., GROSS, E.G., KRONGRAD, A., LESHER JR, J.L., PARK, H.K., SANDERS JR, B.B., SMITH, C.L. Y TAYLOR, J.R. (1996) Effects of selenium supplementation for cancer prevention in patients with carcinoma of the skin: a randomized controlled trial. J. Am. Med. Assoc. 276, 1957-1963.
COLLINS, A.R. (2004) The comet assay for DNA damage and repair. Mol. Biotechnol. 26, 249-261.
COLLINS, A.R., DUSINSKÁ, M., GEDIK, C.M. Y STETINA, R. (1996) Oxidative DNA damage to DNA: do we have a reliable biomarker? Environ. Health Perspect. 104 (Suppl 3), 465-469.
COLLINS, A.R., DUSINSKÁ, M., HORVÁTHOVÁ, E., MUNRO, E., SAVIO, M. Y STETINA, R. (2001) Inter-individual differences in repair of DNA base oxidation, measured in vitro with the comet assay. Mutagenesis 16, 297-301.
COLLINS, A.R., DUTHIE, S.J. Y DOBSON, V.L. (1993) Direct enzymic detection of endogenous oxidative base damage in human lmphocyte DNA. Carcinogenesis 14, 1733-1735.
COLLINS, A.R., HARRINGTON, V., DREW, J. Y MELVIN, R. (2003) Nutritional modulation of DNA repair in a human intervention study. Carcinogenesis 24, 511-515.
COMBS JR, G.F. Y GRAY, W.P. (1998) Chemopreventive agents: selenium. Pharmacol. Ther. 79, 179-192.
DE BOER, J. Y HOEIJMAKERS, J.H. (2000) Nucleotide excision repair and human syndromes. Carcinogenesis. 21, 453-460
DONG, Y., ZHANG, H., HAWTHORN, L., GANTHER, H.E. Y IP, C. (2003) Delineation of the molecular basis for selenium-induced growth arrest in human prostate cancer cells by oligonucleotide array. Cancer Res. 63, 52-59.
DRAKE, E.N. (2006) Cancer chemoprevention: selenium as a prooxidant, not an oxidant. Med. Hypotheses 67, 318-322.
DUFFIELD-LILLICO, A.J., REID, M.E., TURNBULL, B.W., COMBS JR, G.F., SLATE, E.H., FISCHBACH, L. A., MARSHALL, J.R. Y CLARK, L.C. (2002) Baseline characteristics and the effect of selenium supplementation on cancer incidence in a randomized clinical trial: a summary report of the nutritional prevention of cancer trial. Cancer Epidemiol. Biomarkers Prev. 11, 630-639.
DUFFIELD-LILLICO, A.J., SLATE, E.H., REID, M.E., TURNBULL, B.W., WILKINS, P.A., COMBS JR, G.F., PARK, H.K., GROSS, E.G., GRAHAM, G.F., STRATTON, M.S., MARSHALL, J.R. Y CLARK, L.C. (2003) Selenium supplementation and secondary prevention of nonmelanoma skin cancer in randomized trial. J. Natl. Cancer Inst. 95, 1477-1481.
EL-BAYOUMY, K. (2001) The protective role of selenium on genetic damage and on cancer. Mutat. Res. 475, 123-139.
EL-SAYED, W.M., ABOUL-FADL, T., ROBERTS, J.C., LAMB, J.G. Y FRANKLIN, M.R. (2007) Murine hepatoma (Hepa1c1c7) cells: A responsive in vitro system for chemoprotective enzyme induction by organoselenium compounds. Toxicol. in Vitro 21, 157-164.
ESAKI, N., NAKAMURA, T., TANAKA, H., SUZUKI, T., MORINO, Y. Y SODA, K. (1981) Enzymatic synthesis of selenocysteine in rat liver. Biochemistry 20, 4492-4496.
FAKIH, M.G., PENDYALA, L., CREAVEN, P.J., SMITH, P., ROSS, M.E. Y RUSTUM, Y. (2006) A phase I dose escalation study of selenomethionine (SLM) in combination with fixed dose irinotecan (CPT-11) in patients with advanced solid tumors. Proc. Am. Assoc. Cancer Res. 47, 686.
FENG, Y., FINLEY, J.W., DAVIS, C.D., BECKER, W.K., FRETLAND, A.J. Y HEIN, D.W. (1999) Dietary selenium reduces the formation of aberrant crypts in rats administered 3,2´-dimethyl-4-aminobiphenyl. Toxicol. Appl. Pharmacol. 157, 36-42.

90 Revista Real Academia Galega de Ciencias. Vol. XXVII
FINLEY, J.W. Y DAVIS C.D. (2001) Selenium (Se) from high-broccoli is utilized differently than selenite selenate and selenomethionine, but is more effective in inhibiting colon carcinogenesis. Biofactors 14, 191-196.
FISCHER, J.L., MIHELC, E.M., POLLOK, K.E. Y SMITH, M.L. (2007) Chemotherapeutic selectivity conferred by selenium: a role for p53-dependent DNA repair. Mol. Cancer Ther. 6, 355-361.
FRISK, P., YAQOB, A. Y LINDH, U. (2002) Indications of selenium protection against cadmium toxicity in cultured K-562 cells. Sci. Total Environ. 296, 189-197.
GAJECKA, M., RYDZANICZ, M., JASKULA-SZTUL, R., WIERZBICKA, M., SZYFTER, W. Y SZYFTER, K. (2005) Reduced DNA repair capacity in laryngeal cancer subjects. Curr. Res. Head Neck Cancer 62, 25-37.
GANTHER, H.E. (1999) Selenium metabolism, selenoproteins and mechanisms of cancer prevention: complexities with thioredoxin reductase. Carcinogenesis 20, 1657-1666.
GLADYSHEV, V.N., FACTOR, V.M., HOUSSEAU, F. Y HATFIELD, D.L. (1998) Contrasting patterns of regulation of the antioxidant selenoproteins, thioredoxin reductase, and glutathione peroxidase, in cancer cells. Biochem. Biophys. Res. Commun. 251, 488-93.
GLEI, M. Y POOL-ZOBEL, B.L. (2006) The main catechin of green tea, (-)-epigallocatechin-3-gallate (EGCG), reduces bleomycin-induced DNA damage in human leucocytes. Toxicol. in Vitro 20, 295-300.
GRIFFITHS, N.M., STEWART, R.D. Y ROBINSON, M.F. (1976) The metabolism of u in four women. Br. J. Nutr. 35, 373-382.
HARTWIG, A., BLESSING, H., SCHWERDTLE, T. Y WALTER, I. (2003) Modulation of DNA repair processes by arsenic and selenium compounds. Toxicology. 193, 161-169.
HESKETH, J.E. Y VILLETE, S. (2002) Intracellular trafficking of micronutrients: from gene regulation to nutrient requeriments. Proc. Nutr. Soc. 61, 405-414.
HURWITZ, B.E., KLAUS, J.R., LLABRE, M.M., GONZALEZ, A., LAWRENCE, P.J., MAHER, K.J., GREESON, J.M., BAUM, M.K., SHOR-POSNER, G., SKYLER, J.S. Y SCHNEIDERMAN, N. 2007. Suppression of human immunodeficiency virus type 1 viral load with selenium supplementation. A randomized controlled trial. Arch. Intern. Med. 167, 148-154.
IP, C. (1998) Lessons from basic research in selenium and cancer prevention. J. Nutr. 128, 1845-1854.
IP, C., SHU, Z., THOMPSON, H.J., LISK, D. Y GANTHER, H.E. (1999) Chemoprevention in mammary cancer with Se-allylselenocysteine and other selenoamino acids in the rat. Anticancer Res. 19, 2875-2880.
KAJANDER, E.O., HARVIMA, R.J., KAUPPINEN, L., AKERMAN, K.K., MARTIKAINEN, H., PAJULA, R. Y KARENLAMPI, S.O. (1990) Effects of selenomethionine on cell growth and on S-adenosylmethionine metabolism in cultured malignant cells. Biochem. J. 267, 767-774.
KARUNASINGHE, N., RYAN, J., TUCKEY, J., MASTERS, J., JAMIESON, M., CLARKE, L.C., MARSHALL, J.R. Y FERGUSON, L.R. (2004) DNA stability and serum selenium levels in a high-risk group for prostate cancer. Cancer Epidemiol. Biomarkers Prev. 13, 391-397.
KHALIL, A.M. (1989) The induction of chromosome aberrations in human purified peripheral blood lymphocytes following in vitro exposure to selenium. Mutat. Res. 224, 503-506.
KIBRIYA, M.G., JASMINE, F., ARGOS, M., VERRET, W.J., RAKIBUZ-ZAMAN, M., AHMED, A., PARVEZ, F. Y AHSAN, H. (2007) Changes in gene expression profiles in response to selenium supplementation among individuals with arsenic-induced pre-malignant skin lesions. Toxicol. Lett. 169, 162-176.

91Revista Real Academia Galega de Ciencias. Vol. XXVII
KIM, Y-J., BAEK, S-H., BOGNER, P.N., IP, C., RUSTUM, Y.M., FAKIH, M.G., RAMNATH, N. Y PARK, Y-M. (2007) Targeting the Nrf2-Prx1 pathway with selenium to enhance the efficacy and selectivity of cancer therapy. J. Cancer Mol. 3, 37-43.
KIREMIDJIAN-SCHUMACHER, L., ROY, M., WISHE, H.I., COHEN, M.W. Y STOTZKY, G. (1994) Supplementation with selenium and human immune cell functions. II. Effect on cytotoxic lymphocytes and natural killer cells. Biol. Trace Elem. Res. 41, 115-127.
KLEIN, E.A., THOMPSON, I.M. LIPPMAN, S.M., GOODMAN, P.J., ALBANES, D., TAYLOR, P.R. Y COLTMAN, C. (2001) SELECT: the next prostate cancer prevention trial. J. Urol. 166, 1311-1315.
KUO, H.W., CHOU, S.Y., HU, T.W., WU, F.Y. Y CHEN, D.J. (2007) Urinary 8-hydroxy-2´-deoxyguanosine (8-OHdG) and genetic polymorphisms in breast cancer patients. Mutat. Res. 631, 62-68.
LAFFON, B., TEIXEIRA, J.P., SILVA, S., LOUREIRO, J., TORRES, J., PÁSARO, E., MÉNDEZ, J. Y MAYAN, O. (2005) Genotoxic effects in a population of nurses handling antineoplastic drugs, and relationship with genetic polymorphisms in DNA repair enzymes. Am. J. Ind. Med. 48, 128-136.
LAWSON, T Y BIRT, D.F. (1983) Enhancement of the repair of carcinogen-induced DNA damage in the hamster pancreas by dietary selenium. Chem. Biol. Interact. 45, 95-104.
LEMIRE, M., MERGLER, D., FILLION, M., PASSOS, C.J., GUIMARAES, J.R., DAVIDSON, R. Y LUCOTTE, M. (2006) Elevated blood selenium levels in the Brazilian Amazon. Sci. Total Environ. 366, 101-111.
LETAVAYOVÁ, L., VLCKOVÁ, V. Y BROZMANOVÁ, J. (2006) Selenium: from cancer prevention to DNA damage. Toxicology. 227, 1-14.
LUCHESE, C., STANGHERLIN, E.C., ARDAIS, A.P., NOGUEIRA, C.W. Y SANTOS, F.W. (2007) Diphenyl diselenide prevents oxidative damage induced by cigarette smoke exposure in lung of rat pups. Toxicology. 230, 189-196.
LUQUE, J. Y HERRÁEZ, A. (2002) Texto Ilustrado de Biología Molecular e Ingeniería Genética. Conceptos, Técnicas y Aplicaciones en Ciencias de la Salud. Pp Harcourt. Madrid.
MCKELVEY-MARTIN, V.J., GREEN, M.H.L., SCHMEZER, P., POOL-ZOBEL, B.L., DE MÉO, M.P. Y COLLINS, A. (1993) The single cell gel electrophoresis assay (comet assay): A European review. Mutat. Res. 288, 47-63.
MEUILLET, E., STRATTON, S., PRASAD, C.D., GOULET, A.C., KAGEY, J., PORTERFIELD, B. Y NELSON, M.A. (2004) Chemoprevention of prostate cancer with selenium: an update on current clinical trials and preclinical findings. J. Cell. Biochem. 91, 443-458.
MILIC, M. Y KOPJAR, N. (2004) Evaluation of in vitro genotoxic activity of bleomycin and mitomycin C in human lymphocytes using the alkaline comet assay. Arh. Hig. Rada. Toksikol. 55, 249-259.
MILLER, S., WALKER, W.W., ARTHUR, J.R., NICOL, F., PICKARD, K., LEWIN, M.H., HOWIE, A.F. Y BECKETT, G.J. (2001) Selenite protects human endothelial cells from oxidative damage and induces thioredoxin reductase. Clin. Sci. 100, 543-550.
MORRIS, J.D.H., PRAMANIK, R., ZHANG, X., CAREY, A.M., RAGAVAN, N., MARTIN, F.L. Y MUIR, G.H. (2006) Selenium- or quercetin-induced retardation of DNA synthesis in primary prostate cells occurs in the presence of a concomitant reduction in androgen-receptor activity. Cancer Lett. 239, 111-122.
NOGUEIRA, C.W., ZENI, G. Y ROCHA, J.B.T. (2004) Organoselenium and organotellurium compounds: toxicology and pharmacology. Chem. Rev. 104, 6255-6285.
OLIVE, P.L. Y BANÁTH, J.P. (2006) The comet assay: a method to measure DNA damage in individual cells. Nat. Protoc. 1, 23-29.

92 Revista Real Academia Galega de Ciencias. Vol. XXVII
OLIVE, P.L., WLODEK, D., DURAND, R.E. Y BANÁTH, J.P. (1992) Factors influencing DNA migration from individual cells subjected to gel electrophoresis. Exp. Cell Res. 198, 259-267.
PATRICK, L.N.D. (2004) Selenium biochemistry and cancer: a review of the literature. Altern. Med. Rev. 9, 239-258.
POVIRK, L.F. Y AUSTIN, M.J. (1991) Genotoxicity of bleomycin. Mutat. Res. 257, 127-143.
RAFFERTY, T.S., GREEN, M.H.L., LOWE, J.E., ARLETT, C., HUNTER, J.A.A., BECKETT, G.J. Y MCKENZIE, R.C. (2003) Effects of selenium compounds on induction of DNA damage by broadband ultraviolet radiation in human keratinocytes. Br. J. Dermatol. 148, 1001-1009.
RAFFERTY, T.S., MCKENZIE, R.C., HUNTER, J.A.A., HOWIE, A.F., ARTHUR, J.R., NICOL, F. Y BECKETT, G.J. (1998) Differential expression of selenoproteins by human skin cells and protection by selenium from UVB-radiation-induced cell death. Biochem. J. 332, 231-236.
RAJAEE-BEHBAHANI, N., SCHMEZER, P., RISCH, A., RITTGEN, W., KAYSER, K.W., DIENEMANN, H., SCHULZ, V., DRINGS, P., THIEL, S. Y BARTSCH, H. (2001) Altered DNA repair capacity and bleomycin sensitivity as risk markers for non-small cell lung cancer. Int. J. Cancer (Pred. Oncol.) 95, 86-91.
RAO, M., KAMATH, R. Y RAO, M.N.A. (1998) Protective effect of selenomethionine against cisplatin-induced nephrotoxicity in C57BL76J mice bearing B16F1 melanoma without reducing antitumour activity. Pharm. Pharmacol. Commun. 4, 549-552.
RAYMAN, M.P. (2000) The importance of selenium to human health. Lancet 356, 233-241.
RAYMAN, M.P. (2005) Selenium in cancer prevention: a review of the evidence and mechanism of action. Proc. Nutr. Soc. 64, 527-542.
REID, M.E., DUFFIELD-LILLICO, A.J., GARLAND, L., TURNBULL, B.W., CLARK, L.C. Y MARSHALL, J.R. (2002) Selenium supplementation and lung cancer incidence: an update of the nutritional prevention of cancer trial. Cancer Epidemiol. Biomarkers Prev. 11, 1285-1291.
REID, M.E., STRATTON, M.S., LILLICO, A.J., FAKIH, M., NATARAJAN, R., CLARK, L.C. Y MARSHALL, J.R. (2004) A report of high-dose selenium supplementation: response and toxicities. J. Trace Elem. Med. Biol. 18, 69-74.
ROBINSON, M.F., THOMSON, C.D., JENKINSON, C.P., LUZHEN, G. Y WHANGER, P.D. (1997) Long-term supplementation with selenate and selenomethionine: urinary excretion by New Zealand women. Br. J. Nutr. 77, 551-563.
ROTRUCK J.T., POPE, A.L., GANTHER, H.E., SWANSON, A.B., HAFEMAN, D.G. Y HOEKSTRA, W.G. (1973) Selenium: biochemical role as a component of glutathione peroxidase. Science. 179, 588-590.
ROUSSYN, I., BRIVIBA, K., MASUMOTO, H. Y SIES, H. (1996) Selenium-containing compounds protect DNA from single-strand breaks caused by peroxynitrite. Arch. Bioquim. Biophys. 330, 216-218.
RUSSELL, G.R., NADER, C.J. Y PARTICK, E.J. (1980) Induction of DNA repair by some selenium compounds. Cancer Lett. 10, 75-81.
SANTOS, R.A. Y TAKAHASHI, C.S. (2008) Anticlastogenic and antigenotoxic effects of selenomethionine on doxorubicin-induced damage in vitro in human lymphocytes. Food Chem. Toxicol. 46, 671-677.
SANTOS, R.A., JORDAO JR, A.A., VANNUCCHI, H. Y TAKAHASHI C.S. (2007) Protection of doxorubicin-induced DNA damage by sodium selenite and selenomethionine in Wistar rats. Nutr. Res. 27, 343-348.
SCHMEZER, P., RAJAEE-BEHBAHANI, N., RISCH, A., THIEL, S., RITTGEN, W., DRINGS, P., DIENEMANN, H., KAYSER, K.W., SCHULZ, V. Y BARTSCH, H. (2001) Rapid screening assay for

93Revista Real Academia Galega de Ciencias. Vol. XXVII
mutagen sensitivity and DNA repair capacity in human peripheral blood lymphocytes. Mutagenesis 16, 25-30.
SCHRAUZER, G.N. (2000a) Anticarcinogenic effects of selenium. CMLS Cell. Mol. Life Sci. 57, 1864-1873.
SCHRAUZER, G.N. (2000b). Selenomethionine: A review of its nutritional significance, metabolism and toxicity. J. Nutr. 130, 1653-1656.
SCHRAUZER, G.N. (2001) Nutritional selenium supplements: product types, quality, and safety. J. Am. Coll. Nutr. 20, 1-4.
SCHRAUZER, G.N., WHITE, D.A. Y SCHNEIDER, C.J. (1977) Cancer mortality correlation studies. III. Statistical association with dietary selenium intakes. Bioinorg. Chem. 7, 23-31.
SEO, Y.R., CHRISTOPHER, S. Y SMITH, M.L. (2002a) Selenomethionine induction of DNA repair response in human fibroblasts. Oncogene 21, 3663-3669.
SEO, Y.R., KELLEY, M.R. Y SMITH, M.L. (2002b) Selenomethionine regulation of p53 by a ref1-dependent redox mechanism. PNAS 99, 14548-14553.
SHAMBERGER, R.J. 1984. The genotoxicity of selenium. Mutat. Res. 154, 29-48.
SHEN, C.L., SONG, W. Y PENCE, B.C. (2001) Interactions of selenium compounds with other antioxidants in DNA damage and apoptosis in human normal keratinocytes. Cancer Epidemiol. Biomarkers Prev. 10, 385-390.
SINGH, N.P., MCCOY, M.T., TICE, R.R. Y SCHNEIDER, E.L. (1988) A simple technique for quantitation of low levels of DNA damage in individual cells. Exp. Cell Res. 175, 184-191.
SMITH, C.C., O’DONOVAN, M.R. Y MARTIN, E.A. (2006) hOGG1 recognizes oxidative damage using the comet assay with greater specificity than FPG or ENDOIII. Mutagenesis 21, 185-190.
SMITH, M.L., LANCIA, J.K., MERCER, T.I. E IP, C. (2004). Selenium compounds regulate p53 by common and distinctive mechanisms. Anticancer res. 24, 1401-1408.
SPALLHOLZ, J.E. 1994. On the nature of selenium toxicity and carcinostatic activity. Free Radic. Biol. Med. 17, 45-64.
SPALLHOLZ, J.E. (1997) Free radical generation by selenium compounds and their prooxidant toxicity. Biomed. Environ. Sci. 10, 260-70.
STEWART, M.S., SPALLHOLZ, J.E., NELDNER, K.H. Y PENCE, B.C. (1999) Selenium compounds have disparate abilities to impose oxidative stress and induce apoptosis. Free Radic. Biol. Med. 26, 42-48.
SURAI, P. (2006) Selenium and human health. In: Selenium in nutrition and health. Nottingham University Press, Nottingham, p. 643-808.
SUZUKI, K.T., DOI, C. Y SUZUKI, N. (2006b) Metabolism of 76Se-methylselenocysteine compared with that of 77Se-selenomethionine and 82Se-selenite. Toxicol. Appl. Pharmacol. 217, 185-195.
SUZUKI, K.T., KURASAKI, K., OGAWA, S. Y SUZUKI, N. (2006c) Metabolic transformation of methylseleninic acid through key selenium intermediate selenide. Toxicol. Appl. Pharmacol. 215, 189-197.
SUZUKI, K.T., SOMEKAWA, L., KURASAKI, K. Y SUZUKI, N. (2006a) Simultaneous tracing of 76Se-selenite and 77Se-selenomethionine by absolute labeling and speciation. Toxicol. Appl. Pharmacol. 217, 43-50.
TAYLOR, J.B., FINLEY, J.W. Y CATON, J.S. (2005) Effect of the chemical form of supranutritional selenium on selenium load and selenoprotein activities in virgin, pregnant and lacting rats. J. Anim. Sci. 83, 422-429.

94 Revista Real Academia Galega de Ciencias. Vol. XXVII
THOMSON, C.D. (2004) Assessment of requirements for selenium and adequacy of selenium status: a review. Eur. J. Clin. Nutr. 58, 391-402.
THOMSON, C.D. Y STEWART, R.D. (1974) The metabolism of 75Se-selenite in young women. Br. J. Nutr. 32, 47-57.
THOMSON, C.D., ROBINSON, M.F., BUTLER, J.A. Y WHANGER, P.D. (1993) Long-term supplementation with selenate and selenomethionine: selenium and glutathione peroxidase (EC 1.11.1.9) in blood components of New Zealand women. Br. J. Nutr. 69, 577-588.
TINGGI, U. (2003) Essentially and toxicity of selenium and its status in Australia: a review. Toxicol. Lett. 137, 103-110.
VEGA, L., RODRIGUEZ-SOSA, M., GARCÍA-MONTALVO, E.A., DEL RAZO, L.M. Y ELIZONDO, G. (2007) Non-optimal levels of dietary selenomethionine alter splenocyte response and modify oxidative stress markers in female mice. Food Chem. Toxicol. 45, 1147-1153.
WANG, Z., JIANG, C. Y LU J. (2002) Induction of caspase-mediated apoptosis and cell cycle G1 arrest by selenium metabolite methylselenol. Mol. Carcinog. 34, 113-120.
WASCHULEWSKI, I.H. Y SUNDE, R.A. (1988) Effect of dietary methionine on tissue selenium and glutathione peroxidase (EC 1.11.1.9) activity in rats given selenomethionine. Br. J. Nutr. 60, 57-68.
WATERS, D.J., SHEN, S., GLICKMAN, L.T., COOLEY, D.M., BOSTWICK, D.G., QIAN, J., COMBS JR, G.F. Y MORRIS, J.S. (2005) Prostate cancer risk and DNA damage: translational significance of selenium supplementation in a canine model. Carcinogenesis 26, 1256-1262.
WEI, Z., LIFEN, J., JILIANG, H., JIANLIN, L., BAOHONG, W. Y HONGPING, D. (2005) Detecting DNA repair capacity of peripheral lymphocytes from cancer patients with UVC challenge test and bleomycin challenge test. Mutagenesis 20, 271-277.
WHANGER, P., VENDELAND, S., PARK Y.C. Y XIA, Y. (1996) Metabolism of subtoxic levels of selenium in animals and humans. Ann. Clin. Lab. Sci. 26, 99-113.
WHANGER, P.D. (2004) Selenium and its relationship to cancer: an update. Br. J. Nutr. 91, 11-28.
WHITING, R.F., WEI, L. Y STICH, H.F. (1980) Unscheduled DNA synthesis and chromosome aberrations induced by inorganic and organic selenium compounds in the presence of glutathione. Mutat. Res. 78, 159-169.
WOOD, R.D. (1996) DNA repair in eukaryotes. Annu. Rev. Biochem. 65, 135-167.
WOZNIAK, K., ARABSKI, M., MALECKA-PANAS, E., DRZEWOSKI, J. Y BLASIAK, J. (2004)DNA damage in human colonic mucosa cells induced by bleomycin and the protective action of vitamin E. Cell. Mol. Biol. Lett. 9, 31-45.
WYCHERLY, B.J., MOAK, M.A. Y CHRISTENSEN, M.J. (2004) High dietary intake of sodium selenite induces oxidative DNA damage in rat liver. Nutr. Cancer 48, 78-83.
YANG, G., YIN, S., ZHOU, R. GU, L., YAN, B., LIU, Y. Y LIU, Y. (1989) Studies of safe maximal daily dietary Se-intake in a seleniferous area in China. J. Trace Elem. Electrolytes Health Dis. 3, 123-130.
YU, S.Y., ZHU, Y.J. Y LI WG. (1997) Protective role of selenium against hepatitis B virus and primary liver cancer in Qidong. Biol. Trace Elem. Res. 56, 117-24
YU, S.Y., ZHU, Y.J., LI, W.G., HUANG, Q.S., HUANG, C.Z., ZHANG, Q.N. Y HOU, C. (1991) A preliminary report on the intervention trials of primary liver cancer in high-risk populations with nutritional supplementation of selenium in China. Biol. Trace Elem. Res. 29, 289-94
ZHANG, J., PENG, D., LU, H. Y LIU Q. (2008) Attenuating the toxicity of cisplatin by using selenosulfate with reduced risk of selenium toxicity as compared with selenite. Toxicol. Appl. Pharmacol. 226, 251-259.

Revista Real Academia Galega de Ciencias. Vol. XXVII. Págs. 95-106 (2008)
COMPOSICIÓN EN MACRONUTRIENTES, AMINOÁCIDOS Y MINERALES DE ALGUNOS INVERTEBRADOS MARINOS
NO UTILIZADOS HABITUALMENTE COMO ALIMENTO
C. TABOADAa*, R. MILLÁNa, I. MIGUEZa, E. FERNÁNDEZ-PULPEIROb
a Departamento de Fisiología. Facultad de Farmacia. Universidad de Santiago de Compostela. 15782 Santiago de Compostela. España.
b Departamento de Zología y Antropología Física. Facultad de Biología. Universidad de Santiago de Compostela. 15782 Santiago de Compostela.España
*E-mail: [email protected]
RESUMEN
Se analizó la humedad, proteína, grasa, minerales y contenido en aminoácidos de los invertebrados marinos Paracentrotus lividus (Lamarck, 1816), Marthasteria glaciales (Linnaeus, 1758), Monodonta lineada (Da Costa, 1778), Holothuria forskali Delle Chiaje, 1823, Patella vulgata Linneus, 1758, Actinia equina (Linneus, 1758), Anemonia viridis (Forskal, 1775) y Mytilus galloprovincialis Lamarck, 1718, especies que habitualmente no se consumen, con el fin de deducir su posible utilidad como alimentos. El contenido en proteína osciló entre un 9 y un 13,5% y el de grasa entre un 0,22 y un 5,48% de materia seca. En todas las especies los amino ácidos más abundantes fueron el ácido aspártico, ácido glutámico, glicina, arginina, treonina y alanina mientras que el contenido en minerales varía ampliamente entren las distintas especies. Tanto por la composición como por la calidad de la proteína, las especies objeto de estudio parecen adecuadas para consumo humano y explotación comercial.
Palabras clave: Amino ácidos, grasa, invertebrados marinos, minerales, proteína.

96 Revista Real Academia Galega de Ciencias. Vol. XXVII
ABSTRACT
The moisture, protein, total lipid, mineral contents and amino acid compositions of the marine invertebrates Paracentrotus lividus (Lamarck, 1816), Marthasteria glaciales (Linnaeus, 1758), Monodonta lineada (Da Costa, 1778), Holothuria forskali Delle Chiaje, 1823, Patella vulgata Linneus, 1758, Actinia equina (Linneus, 1758), Anemonia viridis (Forskal, 1775) and Mytilus galloprovincialis Lamarck, 1718, species not traditionally used as foodstuffs, were determined in order to deduce their potential usefulness. Protein content ranged from 9 to 13.5% dry weight and total lipid content ranged from 0.22 to 5.48% dry weight. Aspartic acid, glutamic acid, glycine, arginine, threonine and alanine were the predominant amino acids in the above mentioned species. Mineral content has been shown to vary according to the species. The composition of the marine invertebrates under study and the good quality of their proteins make them suitable for consumption and commercial exploitation.
Key words: Amino acids, marine invertebrates, minerals, lipids, protein.
INTRODUCCIÓN
El creciente número de publicaciones de los últimos años relacionando la dieta con diversas enfermedades crónicas, despierta gran interés por alimentos que resultan beneficiosos para la salud y reduzcan la incidencia de enfermedades (Sloan, 1999; Roche, 2006). En este sentido, los invertebrados marinos son una buena fuente de proteínas de alta calidad ya que su composición en aminoácidos está bien equilibrada, son además ricos en minerales y vitaminas y tienen un contenido en grasa relativamente bajo. Muchos se utilizan como alimentos o como suplementos y cada vez se reconoce más su papel destacado en nutrición (Soriguer y col., 1997). Pero precisamente este reconocimiento de los efectos beneficiosos de los productos marinos en el consumidor, tiene como consecuencia que los stocks de aquellos que se usan tradicionalmente como alimentos hayan declinado por sobreexplotación (Venugopal y Shahidi, 1995) y de ahí el interés de encontrar especies nuevas, desconocidas o no utilizadas que puedan tener valor como alimento, bien sea para su utilización directa para consumo humano o como surimí, hidrolizados, concentrados de proteína, etc. (Wijkstrom, 2004; Rossano y col., 2005).

97Revista Real Academia Galega de Ciencias. Vol. XXVII
Los alimentos de origen marino son una buena fuente de ácidos omega-3, con un destacado potencial en nutrición y beneficiosos para la salud y sus proteínas poseen una composición en amino ácidos bien equilibrada, por lo que podrían ser utilizados directamente como fuente de aminoácidos o en preparaciones como dispersiones o hidrolizados (Shahidi, 2004).
Pero antes de que estas especies que tradicionalmente no se explotan se utilicen como alimentos o en la industria alimentaria, se necesita información sobre su composición nutritiva para compararla con la de aquellas habitualmente utilizadas y poder así deducir su posible utilidad.
Por ello, en este trabajo analizamos el contenido en humedad, grasa y proteína de los siguientes invertebrados marinos recogidos en las costas de Galicia: Paracentrotus lividus (erizo), Marthasteria glaciales (estrella de mar), Monodonta lineata (caramujo), Holothuria forskali (pepino de mar), Patella vulgata (lapa), Actinia equina (actinia) y Anemonia viridis (anemona) así como la de Mytilus galloprovincialis (mejillón) con la cual se compararon. Desde el punto de vista nutricional, no sólo es importante la cantidad de proteína sino también su calidad, que depende de la composición en amino ácidos, razón por la cual se determinó este parámetro en las especies anteriormente mencionadas. Por otra parte, en los organismos vivos, el daño oxidativo a macromoléculas puede controlarse mediante dos tipos de sistemas antioxidantes, uno de ellos está representado por enzimas capaces de eliminar especies reactivas de oxígeno, como la superóxido dismutasa, peroxidasa y catalasa, dependientes de cobre, selenio, hierro, manganeso y cinc (Passi y col., 2002), lo que justifica el interés de averiguar también su contenido en minerales.
2. MATERIAL Y MÉTODOS
2.1. Material utilizado
Todas las especies estudiadas se recogieron en las costas de Galicia y se trasladaron al laboratorio, lo antes posible, en recipientes refrigerados con agua de mar. Las determinaciones se realizaron en la vianda de mejillón, caramujo, pepino de mar, lapa, actinia y anémona y en las gónadas de erizo y estrella de mar. Todas las determinaciones se realizaron por triplicado.

98 Revista Real Academia Galega de Ciencias. Vol. XXVII
2.2. Composición en macronutrientes
La determinación de humedad se realizó por desecación a 105º C hasta peso constante utilizando un analizador Scaltec SMO 01 (AOAC, 2001).
La cantidad total de proteína se determinó en cada muestra por conversión del contenido en nitrógeno determinado por el método Kjeldahl (AOAC, 2001).
Para el cálculo del contenido en grasa se utilizó el procedimiento de extracción con éter etílico usando el sistema Shoxlet system (AOAC, 2001).
Con Anemonia viridis y Actinia equina, las determinaciones se realizaron tanto con el material crudo como después de someterlo a fritura en aceite de oliva para eliminar los polipéptidos tóxicos termolábiles que contienen.
2.4. Análisis de aminoácidos
El contenido en aminoácidos de las proteínas se determinó mediante HPLC. Para ello se situaron las muestras en recipientes con ClH 6M, se hidrolizaron a 110º y cada solución se secó en rotavapor. Los residuos disueltos en ClH 20 mM y filtrados se derivatizaron mediante un sistema AccQ Tag (Waters, Milford, MA). El sistema HPLC consistió en una bomba binaria, un módulo desgasificador y un detector de fluorescencia, todos ellos Waters, usando una longitud de onda de excitación y emisión de 250 y 395nm respectivamente. Los cromatogramas se registraron con un software Breeze. Las soluciones patrón de aminoácidos se obtuvieron de Waters, excepto para la norleucina, que se preparó en agua MilliQ a partir de un estándar individual (Sigma Chemicals Co, USA, Ref. N-8513).
2.5. Contenido en minerales
El contenido en minerales se determinó mediante la técnica de espectrometría de emisión óptica ICP-OES en un espectrofotómetro Parkin Elmer 4300DV. Para la preparación de los blancos y de las muestras de las especies marinas ensayadas, se siguió el método de Förstenr y Wittmann (1983) modificado. Para ello se situaron las muestras en recipientes con H2O2 al 30% (Merck Suprapur) y se calentaron progresivamente hasta que se volvieron

99Revista Real Academia Galega de Ciencias. Vol. XXVII
transparentes. Los residuos obtenidos tras la evaporación se disolvieron en HNO3-HCl (1:3) (Merck Supapur), se digirieron a 110º C y se filtraron.
3. RESULTADOS Y DISCUSIÓN
Estudiamos la composición del músculo de cinco invertebrados marinos y las gónadas de dos, todos ellos de escaso valor en el mercado, y se compararon con la del mejillón, molusco que se considera un buen alimento, con objeto de conocer las posibilidades que presentan estas posibles nuevas fuentes de alimento.
Tabla 1. Composición en macronutrientes (%) de invertebrados marinos.
Además, el contenido en aminoácidos de todas ellas, se comparó con el de la caseína puesto que, junto con la albúmina, se considera una proteína patrón.
3.1. Humedad, proteína y grasa
El contenido en agua, proteína y grasa de las especies estudiadas se recoge en la Tabla 1. El porcentaje de humedad osciló entre un 78,4 %, en el caso del caramujo, y el 92,1% para actinia, siendo 85% el valor obtenido

100 Revista Real Academia Galega de Ciencias. Vol. XXVII
con el mejillón. En general, la humedad de pescados y mariscos utilizados para consumo humano se sitúa entre el 60 y el 85% (Sikorski y col., 1990). El menor contenido en grasa correspondió a la holoturia con sólo un 0,22% y el valor más elevado se registró en las gónadas de la estrella de mar con un 5,48%, siendo 1,9 el porcentaje de grasa que contiene el mejillón. Como cabía esperar, las muestras frescas de huevas son las que presentan niveles más altos de grasa, así como las sometidas a proceso de fritura. Comparando estos datos con los que se pueden recoger en la bibliografía para invertebrados marinos, el contenido lipídico de la oreja de mar oscila entre un 0,70% según datos de González y col. (2001) y un 0,80% en el trabajo publicado por Dunstan y col. (1996). King y col. (1990) señalan valores de grasa en marisco de consumo habitual, que se sitúan entre el 0,7% de las vieras y el 3,1% en mejillones, mientras que Gökoolu y Yerlikaya (2003) consideran que el contenido lipídico del cangrejo de mar es prácticamente irrelevante en su composición. En un reciente trabajo realizado por Sirot y col. (2008) sobre la composición lipídica de mariscos y pescados consumidos en Francia, indican un 0,4% en berberechos, el 1,1% en mejillones, un 0,55% para las ostras y un 1,8% para el caramujo, cifra esta última algo superior a la que obtuvimos en nuestro estudio. De todos modos, hay que tener presente que el contenido en agua y humedad depende del tamaño de las especies y de la época entre otros factores, siendo mayores las variaciones en moluscos que en crustáceos. En relación con la proteína, se sitúa en un rango que va desde un 9% en el caso de actinias hasta un 13,5% para lapas, correspondiéndoles valores intermedios a éstos al resto de las especies. Sikorski y col. (1990), encontraron niveles proteicos entre el 10 y el 24% en pescados e invertebrados marinos. De todos modos, si consideramos solamente los datos publicados para invertebrados que actualmente están en el mercado, como es el caso de la almeja, la ostra o el berberecho, podemos ver que las especies objeto de este trabajo tienen una cantidad de proteína similar o incluso superior.
En resumen, en lo que respecta a macronutrientes, podemos afirmar que la composición encontrada en los invertebrados marinos objeto de este estudio concuerda con los datos publicados para otras especies habituales en la dieta humana.

101Revista Real Academia Galega de Ciencias. Vol. XXVII
Tabla 2. Composición en aminoácidos (mg amino acido/g proteína) de los invertebrados marinos estudiados.
3.2. Composición en aminoácidos
La composición en aminoácido, expresada en mg de amino ácido/g de proteína, se indica en la Tabla 2. Todas las especies analizadas contienen todos los amino ácidos esenciales, y el ácido aspártico, glicina, arginina, treonina, alanina y ácido glutámico fueron los amino ácidos
Tabla 2 (Continuación).

102 Revista Real Academia Galega de Ciencias. Vol. XXVII
predominantes en la fracción proteica a pesar de que este último se encontró en todas las muestras en valores inferiores a los de la caseína. El glutámico fue también muy abundante en cangrejo de mar, seguido de aspártico, arginina, lisina y leucina, según los datos publicados por Nack y col. (2004) así como en muchos cefalópodos (Zlatanos y col. 2006). El ácido glutámico es también el predominante en holoturias, seguido de glicina y aspártico, con leucina y lisina presentes en cantidades elevadas (Zhong y col., 2007), mientras que en ostras los más abundantes son también el ácido glutámico pero en este caso junto con la taurina (Tapiero y Tew, 1996). Glutámico, treonina y alanina, junto con otros compuestos, son responsables del olor y sabor característicos de los productos marinos en general, y además, la arginina intensifica estas propiedades organolépticas, aminoácido, por otra parte, muy adecuado para el consumo humano por ser precursor del óxido nítrico, importante vasodilatador. En caso del glutámico, aunque se considera un aminoácido no esencial, hay que tener presente que pasa a ser condicionalmente esencial en casos de traumatismos, cirugías, sepsis, quimioterapia y radioterapia, situaciones en las que la utilización excede a la síntesis. Además, la sal sódica del glutámico se añade a diversos alimentos para mejorar e intensificar el sabor (Tapiero y col., 2002). Según los resultados obtenidos, las huevas de estrella de mar contienen en general valores más altos de aminoácidos esenciales que el mejillón, lo que también ocurre con el caramujo con excepción de fenilalanina, lisina e isoleucina. El contenido en treonina en caramujos y estrella de mar es incluso superior al de la
Tabla 3. Composición mineral de los invertebrados marinos estudiados. El contenido en Mn, Fe, Zn, Cu y Se se expresa en μg/g materia seca y el de K y Ca en mg/g materia seca.
caseína, proteína considerada de referencia por la FAO.

103Revista Real Academia Galega de Ciencias. Vol. XXVII
El aminoácido limitante es la metionina excepto en el caso del caramujo, en el que aunque es baja la cifra para este aminoácido, es la histidina el que está presente en menor concentración.
3.3. Composición mineral
Los datos correspondientes a macrominerales y elementos traza analizados se recogen en la Tabla 3, en la que se puede ver que los resultados varían en función de la especie considerada. Manganeso, hierro, zinc, cobre y selenio son minerales que están implicados en los mecanismos de actividad antioxidante en humanos, por lo que su inclusión en la dieta puede ser beneficioso. Generalmente el marisco es una buena fuente de zinc y ostras, almejas y mejillones lo son también de hierro (King y col., 1990). Análisis de camarones realizados por Tsanev y col. (1996), revelaron un alto contenido en hierro, cobre, manganeso y zinc. También encontramos que el caramujo es más rico en minerales antioxidantes, salvo en selenio, que el mejillón, siendo especialmente alta la cantidad de cobre que contienen. Además, caramujo y lapa son ricos en hierro, mientras que a las huevas de estrella de mar les corresponden los valores más altos de zinc, elemento de función estructural e imprescindible para el normal crecimiento y maduración. Por su parte el selenio es un elemento traza esencial que actúa a través de su presencia en diversas proteínas biológicamente activas (Young, 1981; Holben y Smith, 1999). Actúa como antioxidante inhibiendo la oxidación de lípidos ya que entre las selenoproteínas, la glutatión peroxidada, que se encuentra en células de diferentes tejidos así como en plasma, es la enzima antioxidante mejor caracterizada (Burk, 1997). La concentración de selenio en alimentos marinos incrementa en el orden siguiente: moluscos < crustáceos < pescados (Chien, y col., 2003).
Con respecto al calcio, aunque el marisco es rico en este mineral, la cifra más alta se encontró en holoturias, 139.5 mg/g frente a 80 mg/g en mejillón, valores ambos superiores a 30 mg/g citado por Heu y col. (2003) en camarones.
El potasio es fundamental para el buen funcionamiento del sistema nervioso y para el ritmo normal del corazón, y su deficiencia generalmente causa trastornos cognoscitivos, intolerancia a la glucosa y alteraciones en el ritmo cardíaco. Teniendo en cuenta que los alimentos de origen marino son

104 Revista Real Academia Galega de Ciencias. Vol. XXVII
ricos en sodio, su contenido en potasio es importante a la hora de mantener una ratio sodio/potasio equilibrada.
Ni cobalto ni plomo se encontraron en las partes comestibles de las especies analizadas.
En conclusión, teniendo en cuenta la composición de los invertebrados marinos objeto de este trabajo, y la buena calidad de su proteína que se puede deducir de su contenido en aminoácidos, parecen adecuados para su explotación comercial y podrían ser útiles para consumo humano. El elevado contenido en minerales antioxidantes junto con el papel que se le atribuye a la administración oral de arginina para mejorar la función cardiovascular, los hace susceptibles de la necesidad de profundizar en el estudio de los efectos de la ingesta de estas especies, que incluso podrían llegar a considerarse alimentos funcionales.
AGRADECIMIENTOS
Este trabajo fue financiado por la Xunta de Galicia. Proyecto número PGIDT01PX120310PR.

105Revista Real Academia Galega de Ciencias. Vol. XXVII
BIBLIOGRAFIA
AOAC (2001). Official methods of analyses. Association of Official Analytical Chemists. Washington DC.
BURK, R.F. (1997) Selenium-dependent glutathione peroxidises. In F.P. Guengerich, Comprehensive toxicology, Biotransformation, vol. 3, 229-242. Oxford: Pergamon.
CHIEN, L.C., YEH, C.Y., HUANG, S.Y., SHIEH, M.J., HAN, B.C. (2003). Pharmacokinetic model of daily selenium intake from contaminated seafood in Taiwan. Sci. Total Environ., 311: 57-64.
DUNSTAN, G.A., BAILLIE, H.J., BARRET, S.M., VOLKMAN, J.K. (1996). Effect of diet on the lipid composition of wild and cultured abalone. Aquaculture, 140: 115-127.
FÖRSTNER, U., WITTMANN, G.T.W. (1983). Metal pollution in the aquatic environment. Berlin: Springer-Verlag.
GONZALEZ, M., CARIDE, B., LAMAS, A., TABOADA, C. (2001). Dietary effects of marine food intake on intestinal and hepatic enzyme activities in rats. Int. J. Vitamin and Nutrition Res., 71:128-132.
GÖKOOLU, N., YERLIKAYA, P. (2003). Determination of proximate composition and mineral contents of blue crab (Callinectes sapidus) and swin crab (Portunus pelagicus) caught off the Gulf of Antalya. Food Chemistry, 80: 495-498.
HEU, M.S., KIM, J.S., SHAHIDI, F. (2003). Components and nutritional quality of shrimp processing by-products. Food Chemistry, 82:235-242.
HOLBEN, D.H., SMITH, A.M. (1999). The diverse role of selenium with seleno proteins: A review. J. Amer. Dietetic Asso., 99: 836-843.
KING, I., CHILDS, M.T., DORSET, C., OSTRANDER, J.G., MONSEN, E.R. (1990). Shellfish: proximate composition, minerals, fatty acids and sterols. J. Amer. Dietetic Asso., 90: 677-685.
NACZK, M., WILLIAMS, J., BRENNAN, K., LIYANAPATHIRANA, C., SHAHIDI, F. (2004). Compositional characteristics of green crab (Carcinus maenas). Food Chemistry, 88: 429-434.
PASSI, S., CATAUDELLA, S., DI MARCO, P., DE SIMONA, F., RASTRELLI, L. (2002). Fatty acid composition and antioxidant levels in muscle tissue of different Mediterranean marine species of fish and shellfish. J. Agric. Food Chem., 50:7314-7322.
ROCHE, H.M. (2006). Nutrigenomics-new approaches for human nutrition research. J. Sci. Food Agric., 86: 1156-1163.
ROSSANO, R., CAGGIANO, M.A., MASTRANGELO, L., DI LAURO, R., HÚNGARO, N., ETTORRE, M., RICCIO, P. (2005). Proteins, fatty acids and nutritional value in the muscle of the fish species Mora moro (Risso, 1810). Molecular Nutrition & Food Research, 49: 926-931.
SHAHIDI, F. (2004). Functional seafood products. ACS Symposium Series, 702: 29-49.
SIKORSKI, Z.E., KOLAKOWSKA, A., AND SUN PAN, B. (1990). Nutritional composition of the main groups of marine foodstuffs organisms. In Z.E. Sikorski, Seafood: Resources, Nutritional Composition and Preservation, 41-72. Florida: CRC Press, Inc.
SIROT, V., OSEREDCZUK, M., BEMRAR-AOUACHIRIA, N., VOLATIER, J.L., LEBLANC, J. C. (2008)2008. Lipid and fatty acid composición of fish and seafood consumed in France: CALIPSO study. J. Food Compo. Analalysis,1: 8-16.
SLOAN, E. (1999). The new market: foods for the not-so-healthy. Food Tech., 53: 54-60.

106 Revista Real Academia Galega de Ciencias. Vol. XXVII
SORIGUER, F., SERNA, S., VALVERDE, E., HERNANDO, J., MARTIN-REYES, A., SORIGUER, M., PAREJA, A., TINAHONES, F., ESTEVA, I. (1997). Lipid, protein and calorie content of different Atlantic and Mediterranean fish, shellfish, and molluscs commonly eaten in the south of Spain. Eur. J. Epidem., 13: 451-463.
TAPIERO, H., TEW, K.D. (1996). Increased glutathione expression in cells induced by Crassostera gigas extract (JCOE). Biom. Pharmacother., 50: 149-153.
TAPIERO, H., MATHE, G., COUVREUR, P., TEW, K.D. (2002). Glutamine and glutamate. Biom. Pharmacother., 56: 446-457.
TSANEV, R., RIBAROVA, F., MECHKUEVA, L., RUSEVA, A., BAKARDZHIEVA, M., RIZOV, N., SHISHKOV, S. (1996). Black sea shrimps nutritive value determination. Khigiena i Zdraveopazvane, 39: 40-42.
VENUGOPAL, V., SHAHIDI, F. (1995). Value-added products from underutilized fish species. Critic. Reviews Food Sci. Nutrition, 35: 431-453.
WIJKSTROM, U. (2004). More efficient utilization of fish and fisheries products. En, M. M. Sakaguchi (ed.), Future demand for, and supply, fish and shellfish as food. 9-23. Oxford: Elsevier.
YOUNG, V.R. (1981). Selenium: a case for its essentiality in man. New England J. Medicine, 304: 1228-1230.
ZHONG, Y., KHAN, M.A., SHAHIDI, F. (2007). Compositional characteristics and antioxidant properties of fresh and processed sea cucumber (Cucumaria frondosa). J. Agric. Food Chem., 55: 1188-1192.
ZLATANOS, S., LASKARIDIS, K., FEIST, C., SAGREDOS, A. (2006). Proximate composition, fatty acid analysis and protein digestibility-corrected amino acid score of three Mediterranean cephalopods. Mol. Nutri. Food Res. 50: 967-970.

Revista Real Academia Galega de Ciencias. Vol. XXVII. Págs. 107-129 (2008)
DESARROLLO DE BANCOS DE GERMOPLASMA DE CASTAÑO Y ALCORNOqUE MEDIANTE
CRIOCONSERVACIÓN DE ÁPICES CAULINARES Y EMBRIONES SOMÁTICOS
NIEVES VIDAL1, ANA M. VIEITEZ1, M. ROSARIO FERNÁNDEZ2 Y BEATRIZ CUENCA2
1Instituto de Investigaciones Agrobiológicas de Galicia, CSIC, Apartado 122, 15780 Santiago de Compostela
2 TRAGSA, Departamento de Mejora Agroforestal, Crta. Maceda-Valdrey, Km. 2, 32700 Maceda, Ourense
Correspondencia: [email protected], [email protected]
RESUMEN
Los experimentos descritos en este trabajo demuestran la posibilidad de conservar, a largo plazo, germoplasma de castaño y alcornoque mediante la crioconservación de ápices caulinares y embriones somáticos, respectivamente. En ambos casos los explantos se han precultivado en un medio con alto contenido en sacarosa y posteriormente se han sometido al tratamiento de vitrificación mediante la solución PSV2 antes de la inmersión rápida en nitrógeno líquido (NL). Los 23 clones de castaño crioconservados sobreviven a la acción del NL, pero la capacidad de recuperación de la formación de brotes (con porcentajes entre el 4 y el 53%) sólo fue posible en 16 de los clones estudiados. Cuando el compuesto Supercool al 0.1% se añadió a la solución de vitrificación PVS2, los porcentajes de supervivencia y recuperación de brotes aumentaron en los 8 clones estudiados.
Los embriones somáticos de los 17 clones de alcornoque sometidos a crioconservación recuperan su capacidad embriogénica, con tasas de

108 Revista Real Academia Galega de Ciencias. Vol. XXVII
recuperación que varían entre el 25 y el 100%. El protocolo de crioconservación para esta especie se ha simplificado en relación a los publicados previamente.
Los resultados obtenidos permiten, por vez primera, el desarrollo de un banco de germoplasma con fines aplicados de castaño y alcornoque.
Palabras clave: Ápices caulinares, banco de germoplasma, Castanea sativa, crioconservación, embriones somáticos, nitrógeno líquido, Quercus suber.
ABSTRACT
This work describes experiments demonstrating the feasibility of long-term conservation of both shoot tips of European chestnut and somatic embryos of cork oak by cryopreservation. In both cases, the explants have been pre-cultured on high-sucrose medium followed by the application of PVS2 vitrification solution prior cryostorage. Shoot recovery was obtained in 16 of the 23 chestnut clones tested, and the rates increased with the use of the compound Supercool during the vitrification procedure.
The somatic embryos of the all cork oak genotypes assayed (17) were successfully cryopreserved. The protocol for cryopreservation of cork oak has been simplified in relation to those previously published.
The results obtained show the possibility, for the first time, of the development of an applied gene bank for chestnut and cork oak.
Key words: Castanea sativa, chestnut, cork oak, cryopreservation, germplasm bank, liquid nitrogen, Quercus suber, shoot tips, somatic embryos.
INTRODUCCIÓN
Se estima que el valor económico derivado de los beneficios que proporciona la biodiversidad al ser humano está comprendido entre los 13.000 y 54.000 millones de dólares por año. Sin embargo, sería enormemente limitante evaluar los beneficios de la biodiversidad solamente en términos económicos ya que permite al hombre disfrutar de otro tipo de beneficios entre los que se encuentran los científicos, estéticos, humanísticos, morales,

109Revista Real Academia Galega de Ciencias. Vol. XXVII
naturalísticos e incluso negativos, como puede ser la ansiedad derivada del estudio de su degradación (Kellert, 2005).
Muchos científicos están convencidos que la Tierra está inmersa en un episodio de extinción masiva de las especies (Wilson, 1993). En el reino vegetal, más de 33.000 especies de plantas vasculares están amenazadas, lo que supone el 12.5% del total, que se calcula en 270.000 especies. Aunque se están dando pasos para relativizar la crisis de la biodiversidad su perspectiva es incierta más allá del siglo XXI, a no ser que los éxitos en conservación se aceleren muchísimo (Avise y col., 2008).
Las colecciones de semillas, colecciones en campo, huertos semilleros y los bancos clonales (bien in situ o ex situ) son las formas tradicionales de almacenamiento y conservación de los recursos genéticos vegetales. En el primer caso, sólo especies con semillas “ortodoxas” pueden conservarse por largos períodos de tiempo sin pérdida de su capacidad germinativa; sin embargo, las especies con semillas “heterodoxas” o recalcitrantes tienen una viabilidad muy limitada y su conservación en colecciones convencionales es problemática. Por otra parte, las colecciones en bancos clonales tienen algunas limitaciones: es necesaria una elevada superficie de terreno y una mano de obra especializada para su mantenimiento y conservación y estas colecciones están siempre expuestas tanto a incidencias abióticas (tormentas, fuego, inundaciones) como al ataque de agentes bióticos (plagas, enfermedades) que pueden poner en peligro, en cualquier momento, la propia supervivencia de la colección.
Las tecnologías del cultivo in vitro de tejidos vegetales ofrecen la posibilidad de desarrollar sistemas de almacenamiento y conservación de los recursos genéticos altamente eficaces que pueden ser complementarios a los bancos convencionales de germoplasma. Para que el sistema sea viable, es un requisito imprescindible que la especie a almacenar disponga de un protocolo de regeneración bien definido, para poder obtener plantas completas una vez pasado el período de almacenamiento. De los sistemas disponibles, el más sencillo es el almacenamiento del germoplasma producido in vitro a baja temperatura (2-4º C). Como ventajas en el uso de este método se pueden citar las escasas necesidades de espacio físico y los bajos costes laborales (los subcultivos del material pueden alargarse desde las 4 semanas en el mantenimiento rutinario de los cultivos hasta los 2 años), la eliminación de problemas relacionados con patógenos y la reducción de la erosión genética

110 Revista Real Academia Galega de Ciencias. Vol. XXVII
si se consiguen condiciones óptimas del almacenamiento (Engelmann, 1991). Diferentes factores parecen regular el éxito del almacenamiento en frío de los cultivos in vitro: el estado fisiológico de los brotes, el tipo de explanto, el medio de cultivo, el tipo de contenedor, la temperatura y las condiciones de luz (Orlikowska, 1992). Utilizando este sistema, han podido almacenarse durante 12 meses a 2-4º C cultivos de diferentes clones de castaño, roble y cerezo (Janeiro y col., 1995). También se han obtenido frecuencias cercanas al 100% de viabilidad en 7 de los 8 clones o cultivares de Camellia japonica L. y Camellia reticulata Lindley almacenados en frío durante 12 meses (Ballester y col., 1997) o en cultivos de Fagus sylvatica durante 18 meses. En el trabajo de camelia se comprobó también que la encapsulación de los ápices en perlas de alginato es menos efectiva que el almacenamiento in vitro, ya que después de 75 días de almacenamiento sólo sobrevivieron el 10% de los ápices encapsulados.
A pesar de lo expuesto anteriormente, cuando es posible, la crioconservación (conservación del material vegetal en nitrógeno líquido, NL) es el método más seguro y más económico para la conservación a largo plazo de las especies que son propagadas vegetativamente o que tienen semillas que son recalcitrantes (heterodoxas) al almacenamiento (Pence, 1995). La condición indispensable para iniciar un proceso de crioconservación es disponer de un sistema de regeneración de plantas factible y repetitivo (por ejemplo, a partir de ápices caulinares o embriogénesis somática), que permita la recuperación de plantas completas después de que las células, órganos o tejidos hayan estado inmersos en NL. Aunque inicialmente los métodos de crioconservación se llevaron a cabo mediante un acondicionamiento de las células de los tejidos con crioprotectores y posterior enfriamiento lento, hoy día es más común utilizar metodologías basadas en la vitrificación de los tejidos, es decir, la transición del agua celular directamente de la fase líquida a una fase amorfa o vítrea, evitando la formación de cristales de hielo intra e intercelular (Sakai, 1995). El método de vitrificación conlleva el tratamiento de las muestras con sustancias crio-protectoras, deshidratación de los tejidos con soluciones de vitrificación con elevado potencial osmótico y enfriamiento inmediato por inmersión directa de las muestras en NL.
Es nuestra intención aplicar los procesos de crioconservación a dos especies forestales de enorme interés ecológico, ambiental, social, económico, etc.: castaño y alcornoque. Ambas especies están amenazadas bien por el ataque de patógenos o por una explotación irracional.

111Revista Real Academia Galega de Ciencias. Vol. XXVII
La Estrategia Forestal Española (MIMAM, 1999) del Ministerio de Medio Ambiente, asume una postura activa para la mejora y conservación de la biodiversidad, estableciendo la necesidad de crear una serie de herramientas entre las que cabe señalar la Red de Mejora y Conservación de Recursos Genéticos Forestales. Por otra parte la mejora genética forestal se encuadra en el llamado Eje A.3 del Plan Forestal Español (2002) donde, entre otras, se consideran medidas como la determinación de la variabilidad genética de las especies forestales, establecimiento de las plantaciones de mejora, conservación, etc. La Ley de Montes (2003) establece en su artículo 53 que “el Ministerio de Medio Ambiente en colaboración con las Comunidades Autónomas elaborará y desarrollará programas de ámbito nacional que promuevan la mejora genética y la conservación de recursos genéticos forestales”. Por otra parte uno de los objetivos específicos de la Estrategia Española para la Conservación y Uso Sostenible de los Recursos Genéticos Forestales (MIMAM, 2006) es apoyar los trabajos de mejora genética y conservación de recursos forestales. En el marco de todas estas políticas y medidas nacionales de conservación, que a su vez derivan de medidas europeas previas, se encuadra la trasposición de la Directiva 1999/105/CE a través del Real Decreto 289/2003 de 7 de marzo sobre Comercialización de Materiales Forestales de Reproducción (MFR) que se aplica a aquellos materiales que vayan a ser empleados en plantaciones con fines forestales, bien sea con carácter productor, protector o conservador de biodiversidad. La aplicación del RD establece que los materiales forestales comercializados para ser empleados en las plantaciones, correspondientes a 68 especies forestales y 3 géneros con híbridos artificiales, deben de estar inscritos en el Catálogo Nacional de Materiales de Base. En el anexo 1 del RD figura la especie Castanea sativa y entre los híbridos artificiales se recogen también los del género Castanea. El castaño es una especie susceptible del ataque de enfermedades fúngicas como son la tinta y el chancro.
Entre el año 2000 y 2005, el Departamento de Mejora Agroforestal (DMA) de TRAGSA en Maceda (Ourense) realizó una selección exhaustiva de ejemplares de castaño, empleando los datos del III Inventario Forestal Nacional, superpuestos con el Mapa Forestal, localizando en zonas de fuerte afección de la enfermedad, parcelas donde el castaño fuese primera o segunda especie. Se seleccionaron un total de 206 pies candidatos muestreando un total de 18.808,84 ha contenidas en 513 teselas del Mapa Forestal con presencia de castaño, en las zonas 1 y 2 de regresión del castaño según Bohuier (1979)

112 Revista Real Academia Galega de Ciencias. Vol. XXVII
(Rodriguez y col, 2005). Siguiendo metodologías ya definidas (Vieitez y col.,1986; Sánchez y col., 1997), estos 206 clones se han establecido in vitro para iniciar su propagación (Cuenca y col, 2009) y se han comenzado las tareas de testaje de resistencia y caracterización molecular que permita concluir el grado de resistencia que presentan los materiales con certeza. Durante el tiempo que duren los ensayos en campo, será necesario asegurar la conservación del germoplasma seleccionado, muy valioso no sólo como posibles genotipos a reproducir e incluir en el Catálogo Nacional de Materiales de Base (CNMB) sino también como base genética amplia de un posible plan de mejora genética del castaño.
Por otra parte, esta misma empresa realizó entre 2002 y 2007 una selección de alcornoques plus escogidos por su calidad y producción de corcho, en diferentes rodales selectos de cuatro Regiones de Procedencia extremeñas. Se seleccionaron en total 105 árboles con criterios productivos además de sanitarios y al mismo tiempo, cuidando de mantener una adecuada representación de los diferentes rodales que asegurara una elevada diversidad genética. Estos árboles han sido clonados mediante embriogénesis somática siguiendo la metodología definida por Hernández y col. (2003) en el caso de clonación de alcornoques adultos, y la metodología definida por Bueno y col. (1992) en el caso de clonación de progenies. De este modo, en la actualidad disponemos de líneas embriogénicas correspondientes a 33 genotipos adultos, y se pretende clonar el máximo de árboles plus seleccionados. Se dispone además, de varios cientos de líneas embriogénicas correspondientes a las progenies de 73 de estos árboles, que fueron inducidas a partir de materiales embrionarios. Se han establecido ya tres parcelas con este tipo de materiales, con el objetivo de validar la superioridad productiva de los genotipos adultos, así como la heredabilidad del carácter a través de las progenies. El objetivo último, es de nuevo, proponer al Catálogo Nacional de Materiales de Base (CNMB) los clones seleccionados de alcornoque o bien huertos semilleros establecidos con estos genotipos, como materiales cualificados. La crioconservación sería la herramienta que asegurara la conformidad genética de los materiales durante el período de ensayo, así como la conservación a largo plazo de los materiales seleccionados para futuras actuaciones de mejora en esta especie.
En el presente estudio se pretende investigar la posibilidad de crioconservar el material de castaño y alcornoque previamente seleccionado en el DMA de TRAGSA y definir las condiciones de desarrollo de un banco de germoplasma

113Revista Real Academia Galega de Ciencias. Vol. XXVII
a nivel aplicado. Puesto que en castaño el material es mantenido in vitro y usado para micropropagación a través del desarrollo de yemas axilares, se utilizará el proceso de crioconservación de ápices caulinares (Vidal y col., 2005) mientras que, en el caso del alcornoque, la colección del material vegetal se encuentra en forma de líneas embriogénicas mantenidas mediante embriogénesis repetitiva o secundaria, por lo que se utilizará el sistema de crioconservación de embriones somáticos (Martínez y col., 2003; Valladares y col., 2004). En ambas especies se partirá de protocolos pre-establecidos y se adaptarán, según necesidades, a cada uno de los clones y especies en estudio. Este es el primer intento de desarrollar bancos de germoplasma estables para el mantenimiento, a largo plazo, de estas dos especies.
MATERIAL Y MÉTODOS
Material vegetal y condiciones de cultivo
Se han utilizado cultivos de brotes de castaño (Castanea sativa Mill) de 23 clones seleccionados de origen adulto (Tabla 1) y establecidos in vitro en el DMA (Maceda, Ourense). El material vegetal fue seleccionado en campo por sus características de aparente resistencia a la enfermedad de la tinta y en el momento presente están llevándose a cabo los ensayos de resistencia frente a Phytophthora cinnamomi y P. cambivora, causantes de la enfermedad de la tinta.
Por otra parte, se han utilizado 17 líneas embriogénicas de alcornoque (Quercus suber L) establecidas a partir de árboles seleccionados por su forma, tamaño, condición fitosanitaria y calidad del corcho localizados en 15 rodales selectos de 4 regiones de procedencia de Extremadura (Tabla 5).
Los cultivos de castaño se mantienen rutinariamente mediante proliferación de brotes axilares en medio basal GD (Gresshoff y Doy, 1972) suplementado con 0.4 µM de N6-benzyladenina (BA), 0.09 M de sacarosa y 0.7% (w/v) Bacto agar a pH 5.6 y los subcultivos se llevan a cabo cada 5 semanas. Por su parte, las líneas embriogénicas de alcornoque, mantenidas mediante embriogénesis secundaria, se subcultivan mensualmente en medio basal consistente en los macronutrientes de Schenk y Hildebrant (1972) (medio SH), micronutrientes, vitaminas y Fe-EDTA de Murashige y Skoog (1962), 0.09 M de sacarosa, 0.6% agar y sin reguladores de crecimiento. Los cultivos de las dos especies se incuban en cámaras de crecimiento bajo un

114 Revista Real Academia Galega de Ciencias. Vol. XXVII
fotoperíodo de 16 h suministrado por lámparas fluorescentes de luz blanca y fría (50-60 µmol m-2 s-1) con un régimen de 25ºC en el período de luz y 20ºC en el de oscuridad.
Crioconservación de ápices caulinares de castañoSe ha seguido el método descrito por Vidal y col. (2005) y Jorquera y
col. (2005) modificado y adaptado al material vegetal disponible. A partir de cultivos de 3 a 5 semanas (Fig. 2A), se aislaron brotes de 10 mm de longitud que incluyen las yemas apical y axilares y se transfirieron a placas Petri con medio GD suplementado con 0.2 µM BA, almacenándose en frío a 3-4ºC durante 2 - 3 semanas bajo una luz tenue. Después de esta etapa de endurecimiento, se aislaron de la yema apical (bajo microscopio estereoscópico y en la cabina de flujo estéril, Fig. 2B) ápices caulinares de 0,5-1 mm de tamaño que comprenden el meristemo apical y entre 3 y 6 primordios foliares (Fig. 2C). Estos ápices se precultivaron en medio basal con 0.2 M de sacarosa durante 48 horas a 4ºC. Posteriormente, los ápices se dispusieron en crioviales y se sumergieron durante 20 minutos y a temperatura ambiente en una mezcla crioprotectora (“loading solution”, Matsumoto y col., 1994) compuesta por 2 M glicerol + 0.4 M sacarosa y a continuación se deshidrataron en una solución de vitrificación PVS2 modificada (Sakai y col., 1990) consistente en 30% glicerol, 15% etilenglicol y 15% de dimetilsulfóxido (DMSO) en medio de cultivo GD con 0.4 M de sacarosa. El tiempo de exposición a la solución de PSV2 fue de 120 minutos a 0ºC. Finalmente los ápices se suspendieron en crioviales conteniendo 0.6 ml de PVS2 y se sumergieron en NL (Fig. 2D).
En algunos ensayos, se ha estudiado la adición del compuesto Supercool (polyvinyl alcohol/vinyl acetate copolímero) al 0.1% a la disolución crioprotectora sobre la supervivencia y recuperación de brotes.
Para determinar la viabilidad del material crioconservado, parte de los crioviales que contienen los ápices se retiraron del tanque de NL al menos 24-48 h más tarde de iniciado el proceso, se calentaron en un baño de agua a 40ºC durante 2 minutos, y se eliminó la solución PVS2 que se reemplazó por medio basal GD líquido suplementado con 1.2 M sacarosa durante 20 minutos. El medio de recuperación consistió en el medio basal GD suplementado con 2.2 µM BA, 2.9 µM ácido indolacético (AIA) y 0.9 µM zeatina. Los explantos se colocaron inicialmente sobre papel de filtro dispuesto sobre el medio de recuperación con agar reducido (0.6%) durante 24 h y luego fueron transferidos a medio de recuperación fresco con agar normal (0.7%). Las transferencias se repitieron a las 2 y 4 semanas. Después de 8 semanas de

115Revista Real Academia Galega de Ciencias. Vol. XXVII
cultivo, se determinó la supervivencia de los cultivos como el porcentaje de explantos que han sobrevivido al efecto del NL (% supervivencia). Estos explantos fueron transferidos a tubos que contienen medio GD y 0.4 µM BA y a las 12 semanas se calculó el porcentaje de explantos que muestran desarrollo de brotes (% recuperación), es decir, entre los explantos vivos, aquéllos que forman nuevos brotes mayores de 1 mm. Se utilizaron tres repeticiones por tratamiento y clon, con 12 ápices por repetición y cada experimento se repitió 2 veces.
Crioconservación de líneas embriogénicas de alcornoqueSe ha seguido el método descrito por Valladares y col. (2004) con
modificaciones. Las muestras consistieron en grupos de embriones somáticos (2-4 mg) en estado globular-torpedo aislados de los cultivos embriogénicos 3 semanas después del último subcultivo (Fig. 3A). Estos grupos de embriones (Fig. 3B) se precultivaron durante 3 días en medio SH basal que contiene 0.3 M sacarosa, se transfirieron posteriormente a crioviales y se sumergieron en la misma solución de vitrificación PVS2 utilizada en el caso del castaño preparada en medio GD con 0.4 M sacarosa. Después de 60 minutos a temperatura ambiente, los grupos de embriones se resuspendieron en 0.6 ml de PVS2 y los crioviales se sumergieron directamente en NL (Fig. 3C). Para determinar la eficacia del procedimiento, parte de los embriones se retiraron de los tanques de NL, 24-48 horas después de la inmersión, se sumergieron en un baño de agua a 40ºC y se eliminó la solución PVS2. Se lavaron con medio que contiene sacarosa 1.2 M y se transfirieron a placas con medio SH solidificado con la concentración de agar reducido (0,5%) sobre papel de filtro, siendo transferidos a las 24 horas a jarras de cultivo con medio SH solidificado con agar en la concentración normal (0,6%). La capacidad de recuperación de los cultivos (% recuperación) se determinó como el porcentaje de explantos que mostraron recuperación de la capacidad embriogénica 8 semanas después de aislarlos del NL.
Con el objetivo de simplificar la metodología de crioconservación y adaptarla a las posibilidades prácticas de la empresa, se realizó una serie de experimentos para determinar la necesidad o no del uso de placas con papel de filtro durante las 24 primeras horas tras la descongelación. Asimismo se determinó la influencia de la duración del periodo de precultivo en sacarosa 0,3 M sobre los porcentajes de recuperación de la capacidad embriogénica.
En la Figura 1, se presenta, de forma esquematizada, la metodología seguida para la crioconservación del castaño y del alcornoque.

116 Revista Real Academia Galega de Ciencias. Vol. XXVII
Figura 1. Esquema de las etapas necesarias para la crioconservación de ápices caulinares de castaño (izquierda, 9 etapas) y embriones somáticos de alcornoque (derecha, 6 etapas).
RESULTADOS
El método de crioconservación basado en el proceso de vitrificación es muy similar en el caso de los ápices caulinares de castaño y en el de los embriones somáticos de alcornoque (Fig. 1). Debido a la propia naturaleza del tejido utilizado (Fig. 2A, B, C), el procedimiento de la conservación de los ápices de castaño requiere de 3 etapas más que en el caso del alcornoque (Fig. 3A), como son el endurecimiento en frío de los cultivos, la utilización de una disolución protectora y la transferencia de los cultivos a las 24 h, 2 y 4 semanas después de retirarlos del NL.
Crioconservación de ápices caulinares de castañoEn los 23 genotipos de castaño crioconservados, los ápices que fueron
retirados del contenedor de nitrógeno líquido para evaluar su capacidad de recuperación presentaban inicialmente un color verde, pero a las pocas horas de su transferencia al medio de recuperación todos los explantos adquirieron un aspecto necrosado. Una vez transcurridas dos semanas

117Revista Real Academia Galega de Ciencias. Vol. XXVII
desde la transferencia, comenzaron a aparecer zonas verdes que indicaban la reactivación del crecimiento en los explantos aparentemente necrosados. En algunos casos este nuevo crecimiento dio lugar a la formación de nuevos brotes (Fig. 2F) que pudieron observarse bajo microscopio estereoscópico a partir de las 4 semanas desde la transferencia, mientras que en otros casos sólo se obtuvo tejido desorganizado (callo) o a lo sumo la expansión de alguna hoja que terminó encalleciéndose (Fig. 2E). El hecho de que no todos los explantos que sobreviven formen brotes viables explica que los porcentajes de
Figura 2. Proceso de crioconservación de ápices caulinares de castaño. A, cultivo de brotes en proliferación; B, yema terminal; C, ápice caulinar utilizado como explanto en crioconservación; D, tanque de NL; E, cultivo en el que sólo se ha formado callo después de la recuperación del NL; F, formación de nuevos brotes en cultivos recuperados del NL; G, proliferación de brotes derivados de un ápice crioconservado; H, plantas completas regeneradas.
supervivencia sean sustancialmente mayores a los de recuperación de brotes. Los brotes formados pueden multiplicarse (Fig. 2G) y enraizarse (Fig. 2H), regenerando, de estas forma, plantas completas.

118 Revista Real Academia Galega de Ciencias. Vol. XXVII
En la Tabla 1, se muestran los resultados obtenidos después de la crioconservación de 23 clones de castaño. Al cabo de 8 semanas se constata la supervivencia del 100% de los clones estudiados pero en 7 clones no se logró la recuperación de los cultivos. Esto significa, como se apuntó anteriormente, que en todos los clones de castaño crioconservados en NL hay explantos que sobrevivieron pero en los citados 7 clones sólo se obtuvo formación de callo y no de desarrollo de brotes. El porcentaje de supervivencia no está relacionado con la capacidad de recuperación de los cultivos ya que hay clones con valores del 89% (clon CO31) y del 96% (clon PO39) que finalmente no consiguen recuperarse. El porcentaje de recuperación de brotes está relacionado, claramente, con el genotipo y varía desde valores tan bajos como el 2-4% (en los clones CO30 y PO20, respectivamente) hasta valores del 53% en el clon P034.
En otra serie de experimentos se trató de estudiar el posible efecto beneficioso de la adición del compuesto Supercool sobre la supervivencia y recuperación de los cultivos. Se comprobó (Tabla 2) el efecto positivo de la
Tabla 1. Efecto de la inmersión en nitrógeno líquido (NL) sobre la supervivencia y recuperación de la capacidad de multiplicación de ápices caulinares de 23 clones de castaño. Datos tomados a las 8 (supervivencia) y 12 semanas (recuperación) después del tratamiento con NL.

119Revista Real Academia Galega de Ciencias. Vol. XXVII
adición de este compuesto a la supervivencia de los clones estudiados con porcentajes que, en la mayoría de clones, alcanza el 100%. La excepción es el clon C030 en el que se observa un ligero aumento de la supervivencia con respecto a la acción de la solución crioprotectora PVS2 sin Supercool. En los clones que muestran recuperación de brotes con la solución PSV2, la adición de Supercool duplica el porcentaje de recuperación, incluso en el clon P029, la recuperación es 3 veces mayor que sólo con PVS2. Sin embargo, no se obtuvo efecto positivo alguno en aquellos clones en que la recuperación es ya nula o muy baja (clones P039, C030, C017), excepto en el clon PO20 en donde la recuperación alcanza el 8% en presencia de Supercool.
Tabla 2: Influencia de la adición de Supercool 0,1% a la solución crioprotectora en la supervivencia y recuperación de brotes de 8 clones de castaño.
Crioconservación de líneas embriogénicas de alcornoqueAl igual que en el caso de los ápices de castaño, en el caso de los embriones
somáticos de alcornoque una vez que fueron retirados del tanque de nitrógeno líquido para evaluar su capacidad de recuperación, adquirieron un aspecto necrosado a las pocas horas de su transferencia al medio de recuperación. Transcurridos unos 10 días de la descongelación y transferencia a medio de recuperación, comenzó a hacerse evidente la reactivación del crecimiento con la aparición de nuevas estructuras embriogénicas que surgían de los explantos iniciales mediante un proceso de embriogénesis secundaria (Fig. 3D, E). Los

120 Revista Real Academia Galega de Ciencias. Vol. XXVII
Figura 3. Proceso de crioconservación de embriones somáticos de alcornoque. A, cultivo embriogénico proliferando mediante embriogénesis repetitiva de donde se aíslan los grupos de embriones (B) que serán sometidos a crioconservación en el tanque de NL (C). D y E, nuevos embriones somáticos regenerados a partir de los explantos crioconservados en NL.
embriones, una vez aislados, pueden somaterse a los procesos de maduración y germinación y regenerar plantas completas.
El protocolo inicial de crioconservación de alcornoque (Valladares y col, 2004) incluye la inoculación de los embriones somáticos en placa sobre papel de filtro estéril y con la concentración de agar reducida durante las primeras 24 horas tras la descongelación y el lavado. Uno de los objetivos de nuestro trabajo consistió en simplificar y flexibilizar el protocolo de crioconservación para adaptarlo al manejo de un gran número de clones tratando de eliminar la etapa de inoculación sobre papel de filtro. En los 3 clones de Q. suber estudiados (Tabla 3) la eliminación de la inoculación o siembra sobre papel de filtro de los explantos no sólo no tiene efectos contraproducentes sino que parece favorecer ligeramente la recuperación de los cultivos (si bien las diferencias no son estadísticamente significativas). Por este motivo en los experimentos siguientes con los 14 clones restantes se eliminó este paso del protocolo de crioconservación.

121Revista Real Academia Galega de Ciencias. Vol. XXVII
Tabla 3. Tasas de recuperación embriogénica (% + error estándar) de embriones somáticos de 3 clones de alcornoque crioconservados y sembrados en medio de recuperación, bien directamente sobre el agar o tras un paso previo de 24 horas sobre papel de filtro. Los datos corresponden a medias de 3 repeticiones con 10 explantos cada una, y han sido tomados a las 8 semanas desde la aplicación de los tratamientos.
Por otra parte, también se evaluó el efecto de la duración del precultivo en sacarosa 0,3 M sobre la recuperación de la capacidad embriogénica de los explantos crioconservados, que en el protocolo original estaba fijada en 3 días. Los resultados obtenidos se presentan en la Tabla 4. En uno de los genotipos (clon DII3A (597/04)) la tasa de recuperación embriogénica es del 100%, independientemente de los días que hayan permanecido en el medio de precultivo. En los otros dos genotipos no se observan diferencias significativas en los porcentajes de recuperación, lo que indica que cualquiera de los periodos ensayados es igualmente eficiente para lograr la recuperación de la capacidad embriogénica de los cultivos de alcornoque.
Tabla 4. Tasas de recuperación embriogénica (% + error estándar) de embriones somáticos de 3 clones de alcornoque precultivados durante 1, 2 o 3 días en medio basal suplementado con sacarosa 0,3 M y crioconservados en nitrógeno líquido. Los datos corresponden a medias de 3 repeticiones con 10 explantos cada una, y han sido tomados a las 8 semanas de la descongelación e inoculación en el medio de recuperación.

122 Revista Real Academia Galega de Ciencias. Vol. XXVII
Una vez adaptado el protocolo original de crioconservación de embriones somáticos de alcornoque, se procedió al inicio del desarrollo del banco de germoplasma con las entrada de 17 clones. A diferencia de lo observado en el caso del castaño, en todos los explantos que mostraron reactivación del crecimiento se obtuvieron nuevos embriones somáticos (de 6 a 10 nuevos embriones por explanto original reactivo, por término medio) que permitieron la recuperación del cultivo. En la Tabla 5 se muestran los resultados obtenidos en los ensayos de crioconservación de las líneas embriogénicas de alcornoque. El porcentaje de recuperación claramente depende del clon, pero todos los clones ensayados sobreviven al efecto del NL. El clon Rozo 1A(66/02) es el que presenta una reactividad más baja (25%) mientras que los clones DII3A (597/04), DII3A/02 y Chap A(752/04) alcanzan el 100% de explantos reactivos.
Teniendo en cuenta tanto el número de explantos que todavía están crioconservados como los porcentajes de recuperación calculados, puede decirse que la supervivencia de los clones ensayados está garantizada mientras se efectúa la evaluación en campo de los mismos.
Tabla 5. Efecto de la inmersión en nitrógeno líquido (NL) sobre la reactividad de los embriones somáticos de 17 líneas embriogénicas de alcornoque. Datos tomados 8 semanas después del tratamiento con NL.

123Revista Real Academia Galega de Ciencias. Vol. XXVII
DISCUSIÓN
Los resultados obtenidos demuestran claramente que es posible el desarrollo de bancos de germoplasma aplicados mediante el uso de ápices caulinares de castaño o embriones somáticos de alcornoque almacenados por tiempo indefinido en nitrógeno líquido. El banco, localizado en el DMA de TRAGSA en Maceda (Ourense), dispone en la actualidad de 16 genotipos de castaño y 17 de alcornoque perfectamente viables una vez recuperados del NL. El banco continuará completándose con nuevas entradas a medida que se conozcan los resultados de los ensayos que se llevan a cabo en la actualidad, permitiendo, de esta forma, la evaluación en campo de los genotipos crioconservados durante el período de tiempo que fuese necesario.
Cuando se analizan los resultados obtenidos en el proceso de crioconservación de castaño y alcornoque, el tipo de explanto utilizado en cada especie puede ser el hecho diferencial. El ápice caulinar de castaño utilizado en los ensayos contiene el meristemo apical y entre 3 y 6 primordios foliares que pueden contener células ya vacuoladas y muy diferenciadas y, por tanto, con más dificultades para superar el proceso de vitrificación. La formación de hielo intracelular durante el proceso de crioconservación lleva a la ruptura celular y consecuentemente a la muerte de los tejidos. En un estudio previo, Vidal y col. (2005) estudiaron el efecto del tamaño del ápice caulinar en la recuperación de los cultivos de castaño después del tratamiento con NL y encontraron que 0.5-1 mm era el tamaño más adecuado para permitir la recuperación de los cultivos. En términos generales se acepta que cuanto menor es el tamaño del ápice mayor y más homogéneo es el número de células de tipo meristemático pequeñas y poco vacuoladas, requisito importante en criogénesis, mayor es el éxito de la crioconservación. El tamaño del ápice ha tenido también una gran importancia en diferentes especies (Tagaki y col., 1997; Escobar y col., 1997; Niino y col., 1997). Además del tamaño como factor limitante, al aislar el ápice de la yema terminal del explanto se produce un daño físico debido al corte de los tejidos, daño que puede extenderse a todo el explanto y condicionar el proceso de crioconservación.
El estado fisiológico de la planta madre de la que se aísla el ápice en el momento de su aislamiento (Reed, 2000), el propio genotipo o la adición de distintos agentes crioprotectores pueden también afectar el éxito de la crioconservación del castaño. En el primer caso, aquéllos clones de castaño que no superaron el proceso de crioconservación (Tabla 1), podrían

124 Revista Real Academia Galega de Ciencias. Vol. XXVII
someterse a condiciones de crecimiento diferentes (contenido de reguladores de crecimiento en el medio de cultivo, tratamientos de luz/oscuridad, etc.) que pudiesen dar lugar al aislamiento de ápices caulinares más proclives a la crioconservación. No es descartable que, a pesar de las mejoras potenciales que puedan introducirse, el propio genotipo sea el causante de la falta de recuperación en parte de los clones estudiados ya que no todos los genotipos responden igual a los mismos tratamientos (Vidal y col., 2005). Por otra parte, un hecho a resaltar de los resultados obtenidos en nuestro trabajo es el efecto positivo del polímero Supercool cuando se adiciona a la solución de vitrificación PVS2. La utilización de este crioprotector tuvo su reflejo en el aumento de la supervivencia y de la recuperación de los cultivos. El Supercool inhibe la formación de hielo en soluciones acuosas o crioprotectoras (Wowk y col., 2000) y ha mejorado la crioconservación de ápices caulinares de patata (Zhao y col., 2005). Además del Supercool, existen otros tipos de compuestos crioprotectores, entre los que se encuentran proteínas anticoagulantes que fueron utilizadas con éxito (Xia y col., 2000), aunque su elevado precio limita su aplicación a gran escala.
La capacidad de recuperación de los embriones somáticos de alcornoque tras la crioconservación es mayor que la de los ápices caulinares de castaño. En su conjunto, el embrión somático está formado por células de naturaleza embriogénica, de pequeño tamaño, poco vacuoladas y en división, lo que representa una mayor capacidad para el almacenamiento en NL que las estructuras más complejas como son los ápices caulinares. El estado de desarrollo de los embriones tiene una influencia notable en la posterior recuperación (Valladares et al. 2004). Por eso, hemos utilizado embriones en los estados globular y torpedo que responden mejor al almacenamiento en NL que los embriones más diferenciados como son los que están en estado cotiledonar. Además, los embriones somáticos se separan unos de otros con una gran facilidad, no se produce un daño físico por escisión del tejido a crioconservar, ya que pueden individualizarse facilmente. Por otra parte, con que una célula de carácter embriogénico quede viva, puede regenerar un embrión, mientras que con los ápices caulinares es necesario que sobreviva el conjunto de la estructura organizada que constituye el meristemo, ya que, si sólo sobreviven unas pocas células, lo que se origina es un callo.
Cuando embriones somáticos de castaño se someten a crioconservación (Corredoira y col., 2004) se obtienen también mejores tasas de supervivencia

125Revista Real Academia Galega de Ciencias. Vol. XXVII
y recuperación que con los ápices caulinares. Utilizando esta metodología ha sido posible también la crioconservación de líneas transgénicas de castaño transformadas con genes marcadores (Corredoira y col., 2007). Teniendo en cuenta estos resultados, parecería lógico que los bancos criogénicos de castaño se conformasen mediante cultivos embriogénicos (que también permiten la propagación clonal) en vez de ápices caulinares. Sin embargo, a día de hoy, es más factible el establecimiento in vitro de cultivos de castaño obtenidos mediante el desarrollo de yemas axilares (de cuyos brotes se aíslan los ápices caulinares) que la inducción de embriogénesis somática a partir de explantos de individuos adultos.
En nuestro estudio se ha simplificado el protocolo inicialmente propuesto para la crioconservación de Q. suber (Valladares y col., 2004). Se ha determinado que no es necesario inocular los explantos crioconservados sobre papel de filtro durante las primeras 24 horas tras la descongelación y lavado. Este paso había sido concebido inicialmente para eliminar los exudados que se producen en las primeras horas tras la siembra, pero los resultados obtenidos sugieren que o bien los exudados carecen de efecto tóxico o que la ausencia de manipulación de los explantos durante este periodo es beneficioso. Otra simplificación del protocolo está relacionada con el tiempo de permanencia de los embriones en el medio de precultivo con elevadas concentraciones de sacarosa. No se encontraron diferencias significativas entre 1 y 3 días de precultivo, lo que es una ventaja a la hora de programar y llevar a cabo todo el proceso de crioconservación.
Con el aumento de la tecnología de la crioconservación se ha hecho evidente la necesidad de garantizar la fidelidad del germoplasma almacenado en NL. La información disponible hasta el momento sobre los estudios moleculares en especies leñosas recuperadas del NL confirman la estabilidad genética de los cultivos, corroborando, de esta forma, la fiabilidad de las técnicas criogénicas en la conservación de especies leñosas (Lambardi y col., 2002). Recientemente, este aspecto se ha confirmado en ensayos de crioconservación de embriones somáticos de roble (Sánchez y col., 2008).
La mayoría de la investigación reciente sobre la crioconservación de cultivos embriogénicos en especies forestales está relacionado con las coníferas, en cuya explotación comercial esta técnica es rutinariamente aplicada a las líneas embriogénicas que están siendo evaluadas en campo (Cyr, 2000). En especies caducifolias, la aplicación práctica de esta tecnología está

126 Revista Real Academia Galega de Ciencias. Vol. XXVII
menos avanzada; sin embargo, los resultados obtenidos en el presente trabajo demuestran que la crioconservación a largo plazo de genotipos seleccionados de castaño y alcornoque es posible y, por tanto, el desarrollo aplicado de bancos de germoplasma para la conservación de forma indefinida de estas dos especies es factible.
AGRADECIMIENTOS
El trabajo ha sido parcialmente financiado a través de los proyectos RECATI, código PGDIT07MRU003E (Plan INCITE, Xunta de Galicia) y SEFEAL, código CIT-010000-2007-5 (proyecto del Plan Nacional de Investigación Científica, Desarrollo e Inovación Tecnológica (I+D+i 2004-2007) en la parte dedicada al fomento de la investigación técnica del Ministerio de Educación y Ciencia).
Se agradece al equipo del Dr. Mariano Toribio el haber suministrado las líneas embriogénicas de alcornoques adultos inducidas en el Instituto Madrileño de Investigación y Desarrollo Rural, Agrario y Alimentario y al equipo de la Dra. Mª Ángeles Bueno el suministro de las líneas embriogénicas de progenies utilizadas en este trabajo e inducidas en el Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria (en ambos casos, a través del proyecto SEFEAL).

127Revista Real Academia Galega de Ciencias. Vol. XXVII
BIBLIOGRAFÍA
AVISE, J.C., HUBBELL, S.P., AYALA, F.J. (2008) In the light of evolution II: Biodiversity and extinction. Proc. Nat. Acad. Sci., 105 suppl.1: 11453-11457.
BALLESTER, A., JANEIRO, L.V., VIEITEZ, A.M. (1997) Cold storage of shoot cultures and alginate encapsulation of shoot tips of Camellia japonica L. and Camellia reticulata Lindley. Sci. Hortic., 71: 67-78.
BUENO, M.A., ASTORGA, R., MANZANERA, J.A. (1992) Plant regeneration through somatic embryogenesis in Quercus suber L. Physiol. Plant., 85: 30-34.
CORREDOIRA, E., SAN-JOSÉ, M.C., BALLESTER, A., VIEITEZ, A.M. (2004) Cryopreservation of zygotic embryo axes and somatic embryos of European chestnut. CryoLetters, 25: 33-42.
CORREDOIRA, E., SAN-JOSÉ, M.C., VIEITEZ, A.M., BALLESTER, A. (2007) Improving genetic transformation of European chestnut and cryopreservation of transgenic lines. Plant Cell Tissue Organ Cult., 91: 281-288.
CUENCA, B., FERNÁNDEZ, M.R., OCAÑA, L., SALINERO, C., PINTOS, C., MANSILLA, J.P., RIAL, C. (2009) Selection of Castanea sativa Mill. for resistance to Phytophthora cinnamomi: testing of selected clones. Acta Hortic. (en prensa).
CYR, D. R. (2000) Cryopreservation: roles in clonal propagation and germplasm conservation of conifers. En Cryopreservation of Tropical Plant Germplasm. Current Research Progress and Application. F. Engelmann y H. Takagi (eds.). IPGRI, Rome, Italy, 261-268.
DIRECTIVA 1999/105/CE DEL CONSEJO, de 22 de diciembre de 1999, sobre la comercialización de materiales forestales de reproducción. DO L 11 de 15.1.2000, p. 17/40.
ENGELMANN, F. (1991) In vitro conservation of tropical plant germplasm – a review. Euphytica 57: 227-243.
ESCOBAR, R.H., MAFLA, G., ROCA, W.W. (1997) A methodology for recovering cassava plants from shoot tips maintained in liquid nitrogen. Plant Cell Rep., 16: 474-478.
GRESSHOFF, P.P., DOY, C.H. (1972) Development and differentiation of haploid Lycopersicon esculentum. Planta, 107: 167-170.
HERNÁNDEZ, I., CELESTINO, C., ALEGRE, J., TORIBIO, M. (2003) Vegetative propagation of Quercus suber L. by somatic embryogenesis: II. Plant regeneration from selected cork oak trees. Plant Cell Rep. 21: 765-770.
JANEIRO, L.V., VIEITEZ, A.M., BALLESTER, A. (1995) Cold storage of in vitro cultures of wild cherry, chestnut and oak. Ann. Sci. For., 52: 287-293.
JORQUERA, L., VIDAL, N., SÁNCHEZ, C., VIEITEZ, A.M. (2005) Optimizing conditions for successful plant regeneration from cryopreserved Castanea sativa shoot tips. Acta Hortic. 693: 511-518.
KELLER, S.R. (2005) Perspectives on an ethic toward the sea. Benthic Habitats and the Effects of Fishing, eds. Bames, P.W., Thomas, J.P. (American Fisheries Soc., Symposium 41, Bethesda, MD), 703-712.
LAMBARDI, M., LYNCH, P.T., BENELLI, C., MEHRA, A. (2002) Towards the cryopreservation of olive germplasm. Adv. Hortic. Sci., 16: 165-174.
LEY 43/2003, de 21 de noviembre, de Montes. www.boe.es/boe/dias/2003/11/22/pdfs/A41422-41442.pdf

128 Revista Real Academia Galega de Ciencias. Vol. XXVII
MARTÍNEZ, M.T., BALLESTER, A., VIEITEZ, A.M. (2003) Cryopreservation of embryogenic cultures of Quecus robur using desiccation and vitrification procedures. Cryobiology, 46: 182-189.
MATSUMOTO, T., SAKAI, A., YAMADA, K. (1994) Cryopreservation of in vitro-grown apical meristems of wasabi (Wasabia japonica) by vitrification and sub sequent high plant regeneration. Plant Cell Rep., 13: 442-446.
MIMAM (1999) Estrategia Forestal Española. Dirección General de Conservación de la Naturaleza. Madrid. 297 pp.
MIMAM (2006) Estrategia de Conservación y uso sostenible de los recursos genéticos forestales. Dirección General de Biodiversidad. Madrid. 81 pp. MURASHIGE, T., SKOOG, F. (1962) A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiol. Plant., 15: 473-497.
NIINO, T., TASHIRO, K., SUZUKI, M., OHUCHI, S., MAGOSHI, J., AKIHAMA, T. (1997) Cryopreservation of in vitro grown shoot tips of cherry and sweet cherry by one-step vitrification. Sci. Hort., 70: 155-163.
ORLIKOWSKA, T. (1992) Effect of in vitro storage at 4º C on surviving and proliferation of two apple rootstocks. Plant Cell Tissue Organ Cult., 31: 1-7.
PLAN FORESTAL ESPAÑOL (2002-2032) Aprobado por el Consejo de Ministros el 5 de julio de 2002. www.la-moncloa.es/ConsejodeMinistros/Referencias/_2002/c0507020.htm#PlanForestal
PENCE, V.C. (1995) Cryopreservation of recalcitrant seeds. En: Biotechnology in Agriculture and Forestry, vol. 32 Cryopreservation of Plant Germplasm I. Y.P.S. Bajaj (ed.), Springer, Berlin, Heidelberg, 29-50.
REAL DECRETO 289/2003, de 7 de marzo, sobre comercialización de los materiales forestales de reproducción. (BOE nº 58 del 8 de marzo de 2003. pp. 9262-9299).
REED, B.M. (2000) Genotype considerations in temperate fruit crop cryopreservation. En: Engelmann, F., Takagi, H. (eds.) Cryopreservation of tropical plant germplasm. IPGRI, Roma, 200-204.
RODRÍGUEZ, L., CUENCA, B., LÓPEZ C.A., LARIO, F.J., OCAÑA, L. (2005) Selection of Castanea sativa Mill. genotypes resistant to ink disease in Galicia (Spain) Acta Hort., 693: 645-651.
SAKAI, A., KOBAYASHI, S., OIYAMA, I. (1990) Cryopreservation of nucellar cells of navel orange (Citrus sinensis Osb. var. brasiliensis Tanaka) by vitrification. Plant Cell Rep., 9: 30-33.
SAKAI, A. (1995) Cryopreservation of germplasm of woody plants. En: Biotechnology in Agriculture and Forestry, vol. 32 Cryopreservation of Plant Germplasm I. Y.P.S. Bajaj (ed.), Springer, Berlin, Heidelberg, 53-69.
SÁNCHEZ, M.C., SAN-JOSÉ, M.C., FERRO, E., BALLESTER, A., VIEITEZ, A.M. (1997) Improving micropropagation conditions for adult-phase shoots of chestnut. J Hort Sci 72:433-443.
SÁNCHEZ C, MARTÍNEZ, M.T., VIDAL, N., SAN-JOSÉ, M.C., VALLADARES, S., VIEITEZ, A.M. (2008) Preservation of Quercus robur germplasm by cryostorage of embryogenic cultures derived from mature trees and RAPD analysis of genetic stability. CryoLetters, 29: 493-504.
SCHENK, R.U., HILDEBRANDT, A.C. (1972) Medium and techniques for induction and growth of monocotyledonous and dicotyledonous plant cell culture. Can. J. Bot., 50: 199-204.
TAGAKI, H., THINH, N.T., ISLAM, O.M., SENBOKU, T., SAKAI, A. (1997) Cryopreservation of in vitro grown shoot tips of taro (Colocasia esculenta (L.) Schott) by vitrification. I. Investigation of basic conditions of the vitrification procedure. Plant Cell Rep., 16: 594-599.
VALLADARES, S., TORIBIO, M., CELESTINO, C., VIEITEZ, A.M. (2004) Cryopreservation of embryogenic cultures from mature Quercus suber trees using vitrification. CryoLetters, 25: 177-186.

129Revista Real Academia Galega de Ciencias. Vol. XXVII
VIEITEZ, A.M., VIEITEZ, M.L., VIEITEZ, E. (1986) Chestnut (Castanea spp). In: Bajaj YPS (ed) Biotechnology in agriculture and forestry vol 1. Springer-Verlag, Berlin, Heidelberg, 393-414.
VIDAL, N., SÁNCHEZ, C., JORQUERA, L., BALLESTER, A., VIEITEZ, A.M. (2005) Cryopreservation of chestnut by vitrification of in vitro-grown shoot tips. In Vitro Cell. Dev. Biol. Plant, 41: 63-68.
WILSON, E.O (1993) The Diversity of Life. Harvard University Press, Cambridge, MA.
WOWK, B., LEITL, E., RASCH, C.M., MESBAH-KARIMI, N., HARRIS, S.B., FAHY, G.M. (2000) Vitrification enhancement by synthetic ice blocking agents. Cryobiology, 40: 228-236.
XIA, H.Y., HARVEY, K., LABARRERS, C.A., KOVACS, R. (2000) Plantlet cryopreservation using a combination of epinephrine and dimethylsulphoxide as cryoprotectans. Cryobiology, 41: 97-105.
ZHAO, M.A., XHU, Y.Z., DHITAL, S.P., KHU, D.M., SONG, Y.S., WANG, M.Y., LIM, H.T. (2005) An efficient cryopreservation procedure for potato (Solanum tuberosum L.) utilizing the new ice blocking agent, Supercool X1000. Plant Cell Rep., 24: 477-481.


Revista Real Academia Galega de Ciencias. Vol. XXVII. Págs. 131-144 (2008)
ESTUDO ANTROPOLÓXICO DOS RESTOS ÓSEOS RECUPERADOS DA NECRÓPOLE DA COVA DO SANTO,
PARDOLLÁN, RUBIÁ, OURENSE. UNHA NECRÓPOLE DA IDADE DO BRONCE EN GALICIA.
OLALLA LÓPEZ-COSTAS1
1Área de Antropoloxía Física, Dpto. de Zooloxía e Antropoloxía Física, Facultade de Bioloxía, Universidade de Santiago de Compostela. Avda. Lope Gómez de Marzoa s/n
Campus Sur. 15782 Santiago de Compostela, A Coruña.Correspondencia: e-mail [email protected]
RESUMO
No presente estudo analizamos os primeiros restos óseos humanos recollidos na Cova do Santo en Ourense, datada na idade do Bronce. Este xacemento é único en Galicia, pola súa cronoloxía e a boa conservación do material óseo atopado no interior. As pezas estudadas non se atopáton en contexto anatómico e pertencen a individuos adultos novos ou subadultos, homes e mulleres, destacando unha importante representación de ósos infantís. Identificáronse restos de un mínimo de 9 esqueletos. Estes individuos de morfoloxía mediterránea en sentido amplo presentan unha talla supramedia a outras series peninsulares coetáneas. A porcentaxe de traumatismos e entesoexóstoses é destacada influída probablemente polo médio montañoso que rodea ao xacemento. As patoloxías dexenerativas e infecciosas non están presentes ou mostran un grao leve. Trátase dunha poboación nova cunha actividade física dura.
Palabras clave: Antropoloxía Física, Galicia, idade do Bronce, necrópole.

132 Revista Real Academia Galega de Ciencias. Vol. XXVII
ABSTRAC
An anthropological study was undertaken on the first human remains recovered from the necropolis of Cova do Santo, Ourense. The site is located inside a cave and date to the Bronze Age. The bones studied were found disarticulated and represent subadults and young adults, males and females in the sample. A minimal number of 9 individuals were identified. They were of middle to tall height and the typology is Mediterraneous in senso lapto. The prevalence of trauma and musculoskeletal stress markers is noticeably high, perhaps influenced by the mountainous region. There were no degenerative joint lesions or if present, levels were low. It seems a young population with a hard physical activity.
Key words: Bronze age, Galicia, necropolis, physical anthropology.
INTRODUCIÓN
Unha necrópole de época prehistórica é sempre un achado importante, máis aínda sé se trata de Galicia, onde as alteracións tafonómicas son moi graves chegando a descompoñer case a totalidade da materia orgánica. A Cova do Santo situase no lugar denominado Pardollán, na parroquia de San Estevo do mesmo nome, próximo ao linde da provincia de Ourense coa de León. É unha zona de xeoloxía calcárea, moi diferente da común en Galicia. Esta área caracterízase por posuír terreos básicos onde é usual a presenza de covas e abrigos; ambos feitos permiten unha boa preservación do material arqueolóxico. Debido a que as covas son un refuxio natural para a fauna e un hábitat ocupacional humano é relativamente normal atopar nelas restos de outras épocas. Pero o caso da Cova do Santo ten un especial interese xa que no seu interior acháronse un numero elevado de ósos, tanto humanos como animais, que semellan ser depositados nun mesmo período cronolóxico (López Costas, 2008).
A entrada á gruta encóntrase nun lugar elevado dende o que se divisa a o val do Sil. O seu interior divídese en tres cámaras; unha de primeiro acceso ou “A”, e outras dúas que parten desta última; a chamada“B”(Figura 1) situada cara o suroeste e a “C”, ao sur, cuio acceso atópase impracticable. Segundo parece, a actual entrada dende o exterior sería secundaria a un derrubamento que colapsasou a entrada orixinal situada a máis dun metro

133Revista Real Academia Galega de Ciencias. Vol. XXVII
en vertical baixo a agora presente (información proporcionada polos arqueólogos). Nas tres cámaras da Cova do Santo atopáronse numerosos ósos humanos e animais en superficie, mesturados coas pedras e coa terra do chan. Non hai certeza de que os achados continúen en profundidade pero é moi probable que así sexa. Así mesmo, existen numerosos ósos introducidos nas fendas das paredes laterais (Figura 2). Polo tanto, o escenario semella un
Figura 1. Vista xeral da cámara B.
Figura 2. Restos óseos humanos e animais no interior dunha fenda, nunha das paredes laterais da cámara B.
caos aleatorio de osos, pedras e terra, sendo a cámara “A” a que máis reflicte esta situación.

134 Revista Real Academia Galega de Ciencias. Vol. XXVII
MATERIAL E MÉTODOS
Nesta primeira fase de estudo recolléronse os ósos das cámaras A e B que pola súa situación, nas vías naturais de paso ou en zonas con alta probabilidade de desprendemento, puideran resultar danados nas futuras intervencións. Ningunha mostra foi tomada da cámara C pola dificultade do acceso. Todos os materiais, un total de 61 pezas óseas, foron colectados en Febreiro do ano 2008. Tres pezas foron empregadas para a datación radiocarbónica, situándoas, e polo tanto tamén ao resto do xacemento, na Idade do Bronce.
Os restos estudados presentan un estado de conservación moi bo, aínda que algúns teñen roturas recentes. Neste estudo exporánse os resultados obtidos nunha mostra do material depositado na cova. Preténdese analizar aspectos como os grupos de idade que estan representados, se hai individuos de ambos sexos, ou achegarnos aos estados de saude e enfermidade dos mesmos. Para elo identificouse cada peza diferenciando os ósos de procedencia humana, e procedeuse á determinación do sexo, idade e a un coidadoso estudo antropométrico e paleopatolóxico. A metodoloxía específica de observación consignarase a cada un dos apartados.
RESULTADOS
A distribución dos ósos dentro da cova non parece seguir ningún patrón.Non se atopou correspondencias sobre a presenza dos restos nunha ou noutra cámara. Ningún dos ósos está en contexto anatómico, polo que estes puideron ser trasladados á cova unha vez esqueletizados ou enterrados nela e movidos despois coetáneamente ou no transcurso dos séculos.
Distribución por Idades
A paleodemografía proporciónanos unha aproximación ao perfil da poboación. Para a clasificación por idades identificáronse as pezas pertencentes a adultos e subadultos, precisándose posteriormente os intervalos de anos (infantís I: 0 a 6 anos, infantís II: 7 a 12, xuvenís: 13 a 19, adultos propiamente ditos: 21 a 40, maduros: 41 a 60, e senís: maiores de 61 anos).
Na determinación nos individuos adultos usouse o método de evaluación do grado de cerre das suturas craniais e a sincondrose esfeno-basilar Broca,

135Revista Real Academia Galega de Ciencias. Vol. XXVII
ref. Olivier e Demoulin (1976), Masset (1982), Meindl e Lovejoy (1985), o cerre da liña epifisaria, Brothwell (1987), o inicio do desgaste dental de Brothwell (1987), a superficie auricular do ilion, Lovejoy e col. (1985). A pesar de que se coñecen outros métodos estes non se empregaron debido a ausencia do material sobre o que están desenvolvidos.
Pertencentes a adultos e atopados na cámara A estudáronse, un cranio, unha mandíbula, catro vértebras e cinco costelas, unha escápula, tres coxais, un úmero e unha tibia, tres radios e tres fémures, cinco peronés e un óso animal probablemente ovicáprido. Da cámara B recuperáronse fragmentos de outro cranio, unha escápula, catro vértebras e unha costela, un coxal, un cúbito e un peroné, dúas tibias, seis fémures, un astrágalo e dous ósos de ovicáprido.
Puidose determinar a idade en catro dos coxais, dous pertencen a adultos novos entre 20 e 30 anos, e outros dous a individuos de entre 35 e 40 anos. Ningún coxal atópase nas fases dexenerativas típicas da madurez.
O cálculo da idade da morte nos individuos subadultos realizouse mediante o grado de peche das metáfises, Pyle e Hoerr (1955), McKern e Stewart (1957), Brothwell (1987), a lonxitude dos osos longos sen as súas epífises Stloukal e Hanakova (1978) e a erupción dentaria, Ubelaker (1989). Nos individuos perinatais emprégaronse as táboas de Moorrees e col. (1963) e os criterios de Fazekas e Kósa (1978) para casos forenses.
Na cámara A atopáronse dúas mandíbulas pertencentes a individuos de entre 4 e 6 anos; un ilio, un fémur e unha tibia, todos eles dereitos, pertencentes a infantes dende 5 a 8 anos; e unha clavícula, unha vértebra dorsal, un cúbito e dúas tibias xuvenís. E na cámara B achouse un fragmento de parietal dereito infantil. Polo tanto, hai presenza de, polo menos, dous individuos infantís, un entre 4 e 6 anos e outro entre 6 e 8 anos, e un individuo xuvenil de entre 17 e 20 anos.
Distribución por sexos
A determinación sexual fíxose nos individuos adultos e xuvenís coa fin de detectar a presenza de ambos sexos na cova. A distinción sexual realizase en base á morfoloxía pélvica, Stewart (1968), Phenice (1969), Krogman e Iscan (1986), á morfoloxía craneal e mandibular, Ferembach e col. (1974, 1978), Keen

136 Revista Real Academia Galega de Ciencias. Vol. XXVII
(1950), e as dimensións do esqueleto postcraneal Alemán e col. (1997). Tendo en conta as características propias da poboación galega, López e col. (2007).
Os catro coxais foron clasificados coma masculinos, e dous fémures dereitos e un esquerdo coma femininos. Concluíuse que, ó menos, catro individuos son homes e dúas mulleres.
N.M.I.
Cos restos desarticulados calculouse o número mínimo de individuos ou N.M.I., tendo en conta o número de pezas óseas repetidas, o maior número de articulacións individualizadas, as unidades patolóxicas, e algúns criterios tafonómicos. O cálculo do número mínimo de individuos da mostra recollida fíxose sen ter en conta en que cámara se atoparon.
Entre o material recollido hai un mínimo de cinco adultos e tres nenos. Nos adultos, atopáronse catro coxais esquerdos masculinos e cinco fémures esquerdos, dos cales dous foron clasificados coma femininos. Polo tanto o número de individuos adultos elévase a seis se temos en conta o sexo. Sumándolle os subadultos e tendo en conta a determinación sexual o N.M.I. é de nove, aínda que pola morfoloxía e características probablemente o número de adultos sexa maior e con seguridade o de subadultos.
Táboa 1. Estaturas calculadas segundo os diferentes autores. Os fémures identificados como Rex4-a e Rex4-b poderían pertencer a un mesmo individuo.

137Revista Real Academia Galega de Ciencias. Vol. XXVII
EstaturaA estatura é un marcador que proporciona información sobre a herencia e
orixe dos individuos, xa que ten un alto compoñente xenético, pero tamén sobre outros factores como a nutrición e o estado de saúde da poboación. A talla estimouse coas fórmulas de Pearson (1899), Trotter e Glesser (1958), Olivier e Demoulin (1976) e Mendonça (2000) (Táboa 1). As estaturas calculadas coas pezas estudadas están na media das outras necrópoles peninsulares próximas no tempo (Táboa 2), aínda que lixeiramente superiores. Este feito podería ter a súa explicación no compoñente xenético, xa que a maioría das necrópolis estudadas en Galicia presentan valores de estatura altos, López e col. (2007).
Táboa 2. Comparación dos valores máximo e mínimo da poboación estudada cos promedios de outras peninsulares; estaturas calculadas mediante o método de Pearson.
PatoloxíaO estudo dos estados de saúde e enfermidade danos información sobre
os modos de vida das poboacións antigas e a súa adaptación ao entorno. A avaliación patolóxica fíxose de maneira visual e macroscópica, seguindo os criterios de Aufderheide e Rodríguez-Martín (1998), Campillo (2001), Ortner (2003).
So se puido avaliar a patoloxía maxilodentaria en tres casos, dous deles pertencen a individuos infantís. A saude oral destas pezas é boa, sen marcas de hipoplasia de esmalte ou periodontite. Na mandíbula adulta destacamos un desgaste moderado sen presenza de caries e coa pérdida dun molar.
As patoloxías dexenerativas fálannos das condicións de vida dos individuos e axudannos na determinación da idade. Dúas das 6 vértebras

138 Revista Real Academia Galega de Ciencias. Vol. XXVII
dorsais e as dúas lumbares estudadas presentan modificación diagnosticadas como osteoartrose, aínda que en grao moi leve. Tamén atopánse afectadas o estremo vertebral de 1 das 6 costelas presentes e a articulación distal de dous fémures, ambas en grao leve. Estes resultados coinciden coa idea de poboación nova obtida na determinación da idade.
Tres pezas teñen alteracións traumáticas, os traumatismos están moi influidos polo habitat, nun medio con numerosas colinas e barrancos coma este cabe agardar un procentaxe de fracturas alto. A mais grave e unha fractura consolidada nun ilio esquerdo, Rex8k (Figura 3) a nivel da liña semicircular posterior. É probable que o trauma comprometera a articulación sacro ilíaca e repercutira no movemento do glúteo maior, este individiuo desenrolou unha artrose secundaria na rexión sacroilíaca. Así
Figura 3. Fractura consolidada nun coxal esquerdo.
Figura 4. Periostite inactiva en diáfise de tibia dereita.

139Revista Real Academia Galega de Ciencias. Vol. XXVII
mesmo, achouse unha hernia discal na carilla distal dunha vértebra dorsal (Rex1b), con 9mm. de lonxitude. Por último, a presenza dunha fractura consolidada no terzo proximal da diáfise dun peroné (Rex8t), producíndose unha leve torsión froito da consolidación e un principio de artrose na articulación coa tibia, de orixe secundario á fractura. As entesoexóstosis, como as fracturas, falannos dos patróns de actividade física das persoas. A existencia de entesofitos nos osos longos das extremidades inferiores é frecuente, non así nas superiores ou outros ósos.
A presenza de enfermidades infecciosas é sempre dificil de interpretar, éstas indícannos que o individuo está afectado pero tamén a supervivencia do mesmo un tempo prolongado. Atopamos marcas de periostite inactiva na diáfise dunha tibia dereita, Rex8q (Figura 4). Ademais o maxilar dun cranio (Rex6a) presenta unha osteolise infecciosa que se encontra baixo estudo na actualidade.
Morfoloxía e antropometría
O exame morfolóxico e antropométrico fíxose macroscópicamente, segundo Martin e Saller (1957), Olivier (1960), Olivier e Demoulin (1960), Ferembach (1974), Knussmann (1988). Tamén se documentou a presenza de carácteres morfolóxicos discretos ou epixenéticos no cranio Berry e Berry (1967), e no esqueleto postcraneal, Finnegan e Faust (1974), Finnegan (1978). Os individuos clasifícanse como mediterráneos en sentido amplo, segundo os criterios de Lopez e col. (2000), Pons(1949). Destaca a presenza de numerosos ósos wormianos nos cranios, e da chamada fosa de Allen nun fémur esquerdo (Rex8k).
Tafonomía
Coa fin de comprender mellor como foi a deposición dos restos na cova e, en xeral que sucedeu dende a morte dos individuos ata hoxe, estudáronse as alteracións tafonómicas. Para o exame visual seguíronse as recomendación de Botella e col. (1999). O estado de conservación dos restos é moi bo e só aqueles que se atopaban na cámara A próximos ao acceso dende o exterior, tiñan alteración por fungos e raíces. Moitos ósos están fracturados recentemente

140 Revista Real Academia Galega de Ciencias. Vol. XXVII
por atoparse en zonas de tránsito onde foron pisados ou presionados baixo as pedras. Algúns ósos presentan na súa superficie unha película de carbonato cálcico. Todos estes feitos levannos a concluír que o material leva depositado no interior da cova un periodo de tempo moi prolongado.
CONCLUSIÓNS
No enterramento da Idade do Bronce na Cova do Santo atópanse representadas a maioría dos grupos de idade e ambos sexos, habendo unha ampla presencia de pezas infantís e xuvenís. O número de individuos alí depositado semella ser bastante amplo.
As pezas estudadas falannos dunha poboación nova cunha inusual presenza de traumatismos. As fracturas aparecen consolidadas correctamente polo que tiveron que ser tratadas ou o paciente cumplir repouso. Os marcadores musculo-esqueléticos indican que a actividade física dos individuos podería ser bastante dura. O número mostral non nos permite estudar diferentes patróns de actividade entre hombes e mulleres. Os marcadores relacionados co estrés medioambiental estan pouco presentes e as patoloxías infecciosas crónicas observadas atópanse inactivas, aínda que as infeccións probablemente constituíron a primeira causa de morte.
A morfoloxía das pezas analizadas é semellante as outras necrópoles galegas estudadas; a tipoloxía clasifícase como mediterranea en sentido amplo e os individuos teñen tallas dentro da media das poboacións peninsulares pero son lixeiramente máis altos. Por último a preservación dos restos e moi boa e as alteracións tafonómicas débense a fracturas recentes consecuencia da situación das pezas nos lugares de tránsito. A tafonomía indica que o material leva depositado no interior da cova moito tempo.
Como conclusión final dicir que somos conscientes de que é moi dificil obter unha idea do estado de saude da poboacion con tan poucos restos e estando estes desarticulados. Neste estudo tratamos de obter unha idea preliminar sobre esta necrópole, única na nosa comunidade. A pesar de que é posible que as caracteristicas que aquí se expoñen non sexan aplicables a todolos individuos depositados na Cova do Santo, si están presentes nas persoas ás cales as pezas óseas estudadas pertenceron.

141Revista Real Academia Galega de Ciencias. Vol. XXVII
AGRADECEMENTOS
O presente traballo inclúese nas actividades da bolsa FPU da autora.
Este traballo forma parte do proxecto de investigación: “Antropoloxía dos restos óseos humanos de Galicia. Estudio paleobiodemográfico, morfolóxico, paleopatolóxico, paleoepidemiolóxico e de etnoxénesis da poboación galega”, financiado pola Consellería de Cultura e Deporte, Patrimonio Cultural, da Xunta de Galicia.
Grazas á Dirección Xeral de Patrimonio Cultural e a o Servizo de Arqueoloxía, da Xunta de Galicia polo seu apoio e financiación. Agradecer ao arqueólogo Fidel Méndez Fernández, director da intervención, a súa confianza e axuda. Unha especial mención ao Catedrático de Antropoloxía da Universidade de Santiago de Compostela Prof. Tito A. Varela pola súa axuda e apoio e aos veciños de Pardollán.

142 Revista Real Academia Galega de Ciencias. Vol. XXVII
BIBLIOGRAFÍA
ALEMÁN, I., BOTELLA, M.C., RUIZ, L. (1997) Determinación del sexo en el esqueleto postcraneal. Estudio de una población mediterránea actual. Archivo Español de Morfología, 2: 7-17.
AUFDERHEIDE, A. C., RODRÍGUEZ-MARTÍN, C. (1998) The Cambridge encyclopedia of human paleopathology. Cambridge University Press.
BERRY, A.C., BERRY R.J. (1967) Epigenetic variation in the human cranium. Journal of Anatomy, 101: 361-379.
BOTELLA, M.C., JIMENEZ, S.A., ALEMÁN, I. (1999) Los huesos humanos, manipulación y alteraciones. Ed. Bellaterra.
BROTHWELL, D.R. (1987) Desenterrando huesos. Fondo de cultura económica. Mèxico.
CAMPILLO, D. (2001) Introducción a la paleopatología. Edicions Bellaterra.
FAZEKAS, I.G., KOSA, F. (1978) Forensic Fetal. Osteology. Akademiai Kaido. Budapest, Hungary.
FEREMBACH, D. (1974) Techniques anthropologiques, 1. Craniologie. París, Ecole Pratique des Hautes Etudes, Laboratoire d´Anthropologie Biologique, Laboratoire Broca.
FEREMBACH, D., SCHWIDETZKY, I., STLOUKAL, M. (1978) Raccomandazioni per la determinazione dell´etá e del sesso sullo scheletro. Rivista di Antropologia. Istituto Italiano di Antropologia. Roma, LX, 5-45.
FINNEGAN M. (1978) Non-metric variation of the infracranial skeleton. Journal of Anatomy, 125: 23–37.
FINNEGAN M., FAUST, M. (1974) Bibliography of Human and Nonhuman Non-metric Variation of the Infra-cranial Skeleton, Research Reports 14, Department of Anthropology, University of Massachusetts, Amherst.
GARRALDA, M.D. (1974) Estudio antropológico de la población del Neolítico y Bronce I en la Península Iberica. Tesis Doctoral, Universidad Complutense de Madrid.
JIMENEZ BROBEIL, S.A. (1988) Estudio antropologogico de las poblaciones neolíticas y de la edad del cobre en la Alta Andalucia. Tesis Doctoral, Universidad de Granada.
JIMENEZ BROBEIL, S.A., AL-OUMAOUI, I., NÁJERA, T., MOLINA, F. ( 2008) Salud y enfermedad en Motilla de Azuer; una población de la Edad del Bronce de La Mancha. Revista Española de Antropología Física, 28: 57-70.
KEEN, J. A. (1950) A study of the difference between male and female skulls. American Journal of Physical Anthropology, 8: 65-79.
KNUSSMANN, R, (1988) Anthropologie, I. Stuttgart y Nueva York. Gustav Fischer.
KROGMAN, W.M., ISCAN, Y.M. (1986) The Human Skeleton in Forensic Medicine. Ch. C. Thomas Ed. Springfield, Illinois.
LOVEJOY, C.O., MEINDL, R.S., PRYZBECK, T.R.,MEENSFORTH, R. (1985) Chronological metamorphosis of the Auricular surface of the ilium: a new method for the determination of adult skeletal age at death. American Journal of Physical Anthropology, 68: 1-14.

143Revista Real Academia Galega de Ciencias. Vol. XXVII
LÓPEZ COSTAS, O. (2008) A Cova do Santo: primeiros achados antropolóxicos, Pardollán, Rubiá, Ourense. Inédito. Orixinal entregado na Dirección Xeral de Patrimonio Cultural da Xunta de Galicia.
LÓPEZ COSTAS, O., CASTRO, J. C., BOTELLA, M. C., NÚÑEZ, M. (2007) Estudio de la población medieval gallega. En Martinez-Almagro Andrés(ed.). Diversidad Biológica y Salud Humana. Universidad Católica de San Antonio. Murcia, pp. 535-539.
LÓPEZ, M., RUÍZ, L., SOUICH, P. Du. (2000) Algunas características craneales del tipo mediterráneo en restos antropologicos medievales y modernos. En L. Caro, H. Rodriguez, E. Sánchez, B. López, and M. J. Blanco, eds., Tendencias actuales de investigación en la Antropología Física Española, Universidad de León, pp. 65-69.
MARTIN, R., SALLER, K. (1957) Lehrbuch der Anthropologie in systematischer darstelung mit besonderer Berücksinchtigung der Anthropologischen Methoden. Gustav Fischer Verlag. Stuttgart.
MASSET, C. (1982) Estimation de l´âge an décès par les sutures crâniennes. Thése de Doctorat d´Etat. Lab. Anthropologie Biologique, Université Paris VII.
MCKERN, T. W., STEWART, T. D. (1957) Skeletal age changes in young American males. Headquarters, Quartermaster Research and Development Command, Technical Report EP-45. Natick, Mass.
MEINDL, R.S., LOVEJOY, C.O. (1985) Ectocranial suture closure:a revised method for the determination of skeletal age at death base on the lateral-anterior sutures. American Journal of Physical Anthropology, 68: 57-66.
MENDONÇA, M. C. (2000) Estimation of heigh from the length of long bones in a Portuguese adult population. American Journal of Physical Anthropology, 112: 39-48.
MOORREES, C. F. A., FANNING, E. A., HUNT, Jr. E. E. (1963) Age variation of formation stages for ten permanent. teeth. Journal of Dental Research, 42: 1490-1502.
OLIVIER, G. (1960) Pratique anthropologique. Paris. Ed. Vigot Frères.
OLIVIER, G., DEMOULIN, F. (1976) Pratique anthropologique à l´usage des étudiants. París. Université París 7.
ORTNER, D.J. (2003) Identification of Pathological Conditions in Human Skeletal Remains, 2nd. ed. Academic Press.
PEARSON, K. (1899) Mathematical contribution to the theory of evolution. V. On the reconstruction of the stature of prehistoric races. Philosophical Transactions of the Royal Society, 192: 169-244.
PHENICE, T.W. (1969) A newly developed visual method of sexing the os pubis. American Journal of Physical Anthropology, 30: 297-302.
PONS, J. (1949) Restos humanos procedentes de las necrópolis de época romana de Tarragona y Ampurias (Gerona). Trabajos del Instituto de Antropología y Etnología de Bernardino de Sahagun, VII, 19-206.
PYLE, S. I., HOERR, N. L. (1955) Radiographic atlas os skeletal devolpment of the knee. Charles C. Thomas. Springfield.
STEWART, T.D. (1968) Identification by the skeletal structures. En Gradwohl´s Legal Medicine. Ed. Francis E.Cmaps. 2ªedición, 123-154.
STLOUKAL, M., HANAKOVA, H. (1978) Die länge der Längsknochem altslavischer Bevölkerungen unter besonderer Berücksichtiguns von Wachstumstragen. Homo, 29: 53-69.

144 Revista Real Academia Galega de Ciencias. Vol. XXVII
TROTTER, M., GLESSER, G.C. (1958) A pre-evaluation of estimation of stature based on measurements of stature taken during life and of long bones after death. American Journal of Physical Anthropology, 16: 79-123.
TURBÓN, D. (1981) Antropología de Catalunya en el II milenio a. C. Tesis Doctoral. Universidad de Barcelona, Barcelona.
UBELAKER, D.H. (1989) Human skeletal remains. Excavation, analysis, interpretation. Manuals on Archeology 2. Taraxacum. Washington.

Revista Real Academia Galega de Ciencias. Vol. XXVII. Págs. 145-162 (2008)
ESTIMACIÓN DE LA EVAPOTRANSPIRACIÓN DIARIA A PARTIR DE PRONÓSTICOS METEOROLÓGICOS
EN GALICIA
J.A. RODRÍGUEZ-SUÁREZ1,2*, S. FEIJÓO1, B. SOTO1
1Departamento de Bioloxia Vexetal e Ciencias do Solo, Facultade de Ciencias, Universidade de Vigo, 32004 Ourense, Spain
2Departamento de Ciencias da Navegación e da Terra, Facultade de Ciencias. Universidade da Coruña, 15008 A Coruña, Spain*Correspondencia e-mail : [email protected]
RESUMEN
Se ha determinado la evapotranspiración de referencia (ETo) a partir de los pronósticos meteorológicos realizados por MeteoGalicia (Conselleria de Medio Ambiente, Xunta de Galicia) en 3 estaciones climatológicas. En cada estación se ha estimado la ETo mediante los métodos FAO-Penman Monteith (FAO-PM) y Hargreaves (MHar) a partir de los pronósticos realizados con 1, 2 y 3 días de antelación. Las estimaciones de ETo se han comparado con la ETo calculada mediante el método FAO-PM a partir de los valores observados. Las estaciones climatológicas seleccionadas están ubicadas en zonas con diferentes características climáticas: Ribadeo, Ourense y Pontevedra.
El método de MHar proporciona las mejores estimaciones en Ourense y Pontevedra, mientras que el método FAO-PM tiende a sobrestimar la ETo debido a la baja correlación entre la velocidad del viento pronosticada y observada. Sin embargo, en Ribadeo las estimaciones mediante el método FAO-PM son mejores que las obtenidas mediante el método MHar.
Palabras clave: Evapotranspiración de referencia, FAO-Penman Monteith, pronósticos meteorológicos.

146 Revista Real Academia Galega de Ciencias. Vol. XXVII
ABSTRACT
Reference evapotranspiration (ETo) was determined on 3 weather stations from weather forecasts realized by MeteoGalicia (Conselleria de Medio Ambiente, Xunta de Galicia). In each station, ETo was calculated by means of FAO-Penman Monteith (FAO-PM) and Hargreaves methods from weather forecasts made 1, 2 and 3 days in advance. The predicted ETo was compared with an Eto obtained with FAO-Penman Monteith method applied to measured values of meteorological parameters. The weather stations selected for this study were in Ribadeo, Ourense and Pontevedra.
In Ourense and Pontevedra, best predicted ETo values were obtained with Hargreaves’ method, and FAO-PM overestimated ETo due to low quality of wind velocity prediction. Nevertheless, the FAO-PM method was the most accurate for ETo predictions in Ribadeo.
Keywords: FAO-Penman Monteith, reference evapotranspiration, weather forecast messages
INTRODUCCIÓN
La evapotranspiración (ET) es la combinación de dos procesos separados por los que el agua se pierde por evaporación desde la superficie del suelo y mediante la transpiración de los vegetales. Este proceso conjunto constituye uno de los componentes fundamentales del ciclo hidrológico. Depende fundamentalmente de factores climáticos: radiación solar, temperatura, humedad relativa, presión de vapor, presión atmosférica y viento; y de factores de cultivo: tipo de cultivo, variedad o etapa de desarrollo.
La medida de la ET no es sencilla. Su determinación directa necesita de equipos específicos de alto coste y medida compleja, con el inconveniente de que estos procedimientos, en general, no son adecuados para mediciones rutinarias (Jensen y col., 1990; Al-Ghobari, 2000). Una alternativa más sencilla consiste en su estimación a partir de datos meteorológicos, desarrollándose a lo largo de los años multitud de ecuaciones teóricas, empíricas o semiempíricas, para su cálculo. De todas ellas la FAO ha establecido como método de referencia internacionalmente recomendado el FAO-Penman Monteith (FAO-PM) (Allen y col., 1998), por ser el más preciso y obtener los mejores resultados al ser comparado con medidas experimentales en gran

147Revista Real Academia Galega de Ciencias. Vol. XXVII
variedad de climas y regiones del mundo (Chiew y col., 1995; García y col., 2004; Gavilan y col., 2006; Popova y col., 2006; Jabloun y Sahli, 2008). El método FAO-PM permite calcular una ET de referencia, ETo, definida como “la tasa de evapotranspiración de una superficie de pasto verde que cubre totalmente el suelo y que no presenta restricciones de agua”, la cual permite comparar la ET entre diferentes regiones y épocas del año, además de entre diferentes cultivos.
En el caso de Galicia, la evapotranspiración potencial presenta unos valores medios anuales en torno a 700 mm, aunque puede variar según la zona entre 500 y 800 mm. Esta variación se debe a que, aún considerando el clima gallego como un clima suave de influencia oceánica muy lluvioso, su irregular orografía propicia la existencia de múltiples microclimas con fuertes variaciones en un área menor de 200 Km2 (Castillo Rodríguez y col., 2006). A lo largo del año los mayores valores de ET se dan en primavera y verano (Soto y Díaz-Fierros, 1996) coincidiendo con los meses en los que las precipitaciones son menores y en los que la demanda de agua aumenta, sobre todo para regadío. En consecuencia, en esta época del año es indispensable una gestión adecuada de los recursos hídricos que garantice un suministro suficiente de agua para todas las demandas existentes. En este sentido, es necesario conocer con exactitud el valor de la ET ya que es un parámetro fundamental en estudios hidrológicos y agrícolas tales como diseño y programación de sistemas de riego, prevención de incendios, regulación de embalses, producción agrícola y forestal, o planificación de los recursos hídricos y usos del territorio, entre otros (Castillo y Castelví, 2001; Mardikis y col., 2005; Chauhan y Shrivastava, 2009).
Dada la importancia de la determinación adecuada de la ET, disponer de herramientas que permitan conocer su valor con antelación y precisión resulta de gran ayuda en la mejora de la gestión de los recursos hídricos. En esta línea, la determinación de la ETo con varios días de antelación podría realizarse a partir de los pronósticos realizados por los servicios de meteorología (Jiabing y col., 2007). En Galicia, MeteoGalicia (Conselleria de Medio Ambiente e Desenvolvemento Sostible, Xunta de Galicia) realiza predicciones de las condiciones meteorológicas con una antelación de varios días y estos datos son publicados diariamente y están a disposición de los usuarios.
En este trabajo se ha evaluado la calidad de la ETo predicha con 1, 2 y 3 días de atelación en diferentes zonas de Galicia a partir de los pronósticos

148 Revista Real Academia Galega de Ciencias. Vol. XXVII
meteorológicos realizados por MeteoGalicia, disponibles vía Web (www.meteogalicia.es). Los objetivos planteados en este estudio son: (1) estimar la ETo por diferentes métodos a partir de los pronósticos meteorológicos; 2) analizar la calidad de estas estimaciones comparándolas con la ETo calculada por el método de referencia FAO-PM a partir de datos observados y 3) determinar la influencia del tiempo de antelación, 1, 2 ó 3 días, en la calidad de las predicciones de la ETo.
MATERIAL Y METODOS
Datos meteorológicos
Estaciones Meteorológicas
Para realizar este estudio se han seleccionado tres estaciones meteorológicas pertenecientes a la red de Estaciones de MeteoGalicia. Concretamente se trata de la estación Ourense-Ciencias (Ourense), Lourizán (Pontevedra), y Pedro Murias (Ribadeo), cuyas características y localización se muestran en la Tabla 1 y Figura 1.
Tabla 1. Localización de las estaciones meteorológicas empleadas en este estudio.
Las 3 estaciones se han seleccionado por su situación en zonas de características climáticas diferentes: oceánico húmedo de temperaturas suaves (Pedro Murias, Ribadeo), oceánico húmedo de temperaturas moderadas con riesgo de sequía estival (Lourizán, Pontevedra) y clima oceánico continental (Ourense-Ciencias, Ourense).
Los datos de temperatura máxima (Tmax) y mínima (Tmin), humedad relativa (HR) y radiación solar (Rs) registrados en estas estaciones se han empleado para el cálculo de la ETo mediante FAO-PM y los resultados se han

149Revista Real Academia Galega de Ciencias. Vol. XXVII
Figura 1. Localización de las estaciones meteorológicas empleadas en este estudio.
comparado con los obtenidos empleando los pronósticos meteorológicos con 1, 2 o 3 días de antelación mediante los métodos FAO-PM y MHar.
Modelos de predicción meteorológica
MeteoGalicia durante el año 2008 ofreció a través de su web (http://www.meteogalicia.es) pronósticos meteorológicos realizados con diferentes modelos: ARPS con resolución de 6, 18 y 54 km2, GFS, y MM5 con resolución de 10 y 50 km2. Este servicio da la posibilidad de descargar series temporales de pronósticos de temperatura, módulo y dirección del viento, humedad relativa y precipitación con hasta tres días de antelación para diferentes ciudades gallegas.
Para este estudio se ha utilizado las series temporales del modelo ARPS. Se recogieron para cada día los pronósticos realizados con 1, 2 y 3 días de antelación para las ciudades de Ourense, Pontevedra y Ribadeo entre el 19 de febrero de 2008 y el 1 de septiembre de 2008 con el fin de emplear esta información para el cálculo de la ETo. Se ha elegido este periodo por ser la época del año en la que se dan las mayores demandas hídricas por los cultivos y las menores precipitaciones, y por tanto mayores necesidades de riego que es necesario conocer con anterioridad con el fin de hacer una gestión y uso adecuado del agua.

150 Revista Real Academia Galega de Ciencias. Vol. XXVII
Cálculo de la ETo
El método FAO-Penman Monteith
El método FAO-PM es el método de referencia recomendado internacionalmente para el cálculo de la ETo diaria (mm día-1). La expresión empleada para su cálculo es la siguiente:
[1]
donde Rn es la radiación neta en la superficie del cultivo (MJ m-2 día-1), G el flujo de calor del suelo (MJ m-2 día-1), T la temperatura media del aire a 2 metros de altura (ºC), u2 la velocidad media del viento a 2 metros de altura (m s-1), es la presión de vapor de saturación (kPa), ea la presión real de vapor (kPa), Δ la pendiente de la curva de presión de vapor (kPa ºC-1) y γ es la constante psicrométrica (kPa ºC-1). Complementariamente Allen y col. (1998, 2006) propusieron una serie de ecuaciones para poder calcular los parámetros de la ecuación [1] a partir de datos meteorológicos de T, HR, u2 y Rs.
Los pronósticos de Rs del modelo ARPS no son ofrecidos en la web de MeteoGalicia, por lo que este parámetro tuvo que estimarse a partir de Tmax y Tmin diarias por el método descrito por Allen y col. (1998, 2006) según el cual:
[2]
donde Rse es la radiación solar estimada (MJ m-2 día-1), Ra es la radiación extraterrestre (MJ m-2 día-1), Tmax es la temperatura máxima del aire (ºC), Tmin es la temperatura mínima del aire (ºC) y kRs un coeficiente de ajuste (ºC-0.5).
La radiación extraterrestre, Ra, se calcula mediante la expresión:
[3]
donde Gsc es la constante solar (0.082 MJ m-2 min-1), dr distancia relativa inversa Tierra-Sol, ωs el ángulo de radiación a la puesta del sol [rad], φ es la latitud [rad] y δ la declinación solar [rad].

151Revista Real Academia Galega de Ciencias. Vol. XXVII
El parámetro kRs de la ecuación [2] es un coeficiente de ajuste empírico que toma diferentes valores en zonas costeras y zonas de interior. Aunque el método recomienda valores de 0.16 para zonas de interior y 0.19 para zonas costeras, en este trabajo se ha calculado su valor para cada estación a partir de los registros históricos de Rs, Tmax y Tmin de cada una de ellas. Los valores obtenidos son de 0.13 (n = 1130, R2 = 0.85) para la estación de Ourense, 0.17 (n = 2532, R2 = 0.82) para la estación de Pontevedra, y 0.21 (n = 2357, R2 = 0.77) para la estación de Ribadeo, los cuales difieren ligeramente de los valores recomendados.
La ETo calculada por el método FAO-PM a partir de los pronósticos (ETpro) fue comparada con la ETo calculada por el método FAO-PM con los valores registrados en cada estación (ETPM).
El método Hargreaves
El método de Hargreaves (MHar) (Hargreaves y Samani, 1985) es un método alternativo para el cálculo de la ETo que cuenta con la ventaja de requerir menos parámetros para su determinación:
[4]
donde ETHar es la ETo (mm día-1), Tmed es la temperatura media diaria (ºC), Tmax es la temperatura máxima diaria (ºC), Tmin la temperatura mínima diaria (ºC), y Ra la radiación extraterrestre (mm día-1) que se calcula de acuerdo a la ecuación [3].
Antes de aplicar este método en una región deben compararse sus resultados con los obtenidos mediante el método de referencia (FAO-PM). Si es necesario, el método debe ser ajustado determinando los coeficientes empíricos de correlación de la forma:
[5]
donde b y a son los coeficientes de ajuste.
En este trabajo se ha ajustado previamente este método para cada estación empleando los registros históricos de Rs, u2, HR, Tmed, Tmax y Tmin. A partir de ellos se ha calculado la ETo por ambos métodos obteniéndose los parámetros de calibrado a y b que se recogen en la Tabla 2.

152 Revista Real Academia Galega de Ciencias. Vol. XXVII
Tabla 2. Coeficientes de ajuste (b y a) obtenidos entre el método de Hargreaves y el método de referencia (FAO-PM) para cada estación de estudio. R2 es el coeficiente de correlación y n es el número de datos empleados en el ajuste.
En general, existe una buena correlación entre la ETo obtenida por ambos métodos, sobre todo en las estaciones de Ourense-Ciencias y Lourizan, R2 de 0.91 y 0.93 respectivamente, mientras que en la estación de Ribadeo se ha obtenido un R2 de 0.81. Al igual que con la ETpro, la ETHar se ha comparado con la ETPM para comprobar la validez y precisión del método propuesto.
Análisis de los resultadosPara comprobar la bondad de las estimaciones realizadas se ha recurrido
a los siguientes índices estadísticos:
(a) Coeficientes de regresión lineal b y R2, en los que b es el coeficiente de regresión poblacional o pendiente de la línea recta y R2 el coeficiente de correlación lineal. Cuanto más próximos sean los dos a 1 mejor es la estimación.
(b) El error relativo (RE):
donde RMSE es el error cuadrático medio, N es el número de observaciones, O los valores observados, P los valores estimados, y Ō la media de los observados. Cuanto más próximo a 0 sea el valor de este índice menor es el error cometido en la estimación.
(c) El índice de Willmott (1982) (d):
donde Pi´ = Pi – Ō y Oi´ = Oi – Ō. Un valor de d igual a 1 índica una estimación perfecta.

153Revista Real Academia Galega de Ciencias. Vol. XXVII
RESULTADOS Y DISCUSIÓNAnálisis de los parámetros meteorológicos pronosticadosTemperatura máxima y mínima
En la Tabla 3 se recogen los resultados obtenidos al comparar los valores pronosticados con diferentes días de antelación (1, 2 y 3) con los valores de los parámetros meteorológicos observados. Tmax y Tmin son los parámetros mejor estimados por el ARPS en las tres estaciones, obteniendo en prácticamente la totalidad de los casos valores de b dentro del rango 1±0.1, R2 entre 0.63 y 0.79 y un RE por debajo de 0.15. En consecuencia los valores de d son también buenos y prácticamente en su totalidad superiores a 0.9. Los pronósticos realizados con 1, 2 y 3 días de antelación son muy semejantes entre sí, si bien todos los resultados mejoran ligeramente cuanto menor es el período de antelación del pronóstico.
Tabla 3. Índices estadísticos obtenidos al comparar los valores pronosticados y observados de los parámetros empleados en el cálculo de la ETo en cada estación y día de antelación del pronóstico.

154 Revista Real Academia Galega de Ciencias. Vol. XXVII
Humedad relativa
La HR es un parámetro globalmente bien pronosticado, valores de b entre 0.90-0.96 con un RE bajo. Sin embargo los R2 obtenidos, especialmente en la estación de Pedro Murias, indican poca precisión en su estimación. En consecuencia, los valores del índice d son peores que los obtenidos con la temperatura, aunque en general pueden considerarse aceptables. De nuevo, las estimaciones a 1 2 y 3 días son muy similares en las tres estaciones mejorando el resultado cuanto menor es el periodo de antelación del pronóstico.
Velocidad del viento
El parámetro peor estimado por el modelo ARPS es u2. Todos los parámetros estadísticos indican que no existe relación entre los valores pronosticados y observados, sobre todo en el caso de las estaciones de Ourense-Ciencias y Lourizán, donde los valores de b indican que los pronósticos están sobrestimados, superando los valores pronosticados en más del doble a los valores observados.
Radiación solar
La Rs estimada a partir de los pronósticos de Tmax y Tmin se ajusta adecuadamente a la observada con prácticamente todos los valores de b dentro del intervalo 1.0 ± 0.1. Al igual que la HR, presenta cierta dispersión que explica los bajos valores de R2 y un RE no tan bueno como el obtenido con la HR o la T. A pesar de esto, el valor d está razonablemente cercano a 1, siempre por encima de 0.7, pudiendo considerarse que la Rs esta adecuadamente estimada.
Tabla 4. Índices estadísticos obtenidos al comparar ETopro (pronostico de ETo calculado mediante el método de FAO-PM) con EToPM (ETo calculada mediante el método FAO-PM a partir de los datos registrados) para cada estación y día de antelación del pronóstico.

155Revista Real Academia Galega de Ciencias. Vol. XXVII
Estimación de la ETo a partir de los pronósticos meteorológicos
Método FAO-Penman Monteith
Los resultados obtenidos en el cálculo de la ETpro se recogen en la Tabla 4 y Figura 2, donde se representa la ETpro con 3 2 y 1 días de antelación frente la ETPM para cada estación de estudio.
Figura 2. ETo diaria calculada por el método FAO-PM para cada estación a partir de los pronósticos meteorológicos realizados con 1 2 y 3 días de antelación, frente a la ETo diaria calculada a partir de los datos meteorológicos registrados en cada estación por el método FAO-PM.
Existe una buena correlación entre ETpro y ETPM en las tres estaciones ya que el valor del índice b es muy próximo a 1 y d está siempre por encima

156 Revista Real Academia Galega de Ciencias. Vol. XXVII
de 0.8. En Ourense-Ciencias y Lourizán se obtiene un buen R2, entre 0.67 y 0.75, pero en Pedro Murias no supera un valor de 0.52, consecuencia muy probablemente de la dispersión que ya presentaban las estimaciones de HR y Rs para esta zona. A pesar de los aceptables valores de b, d y R2 obtenidos en Ourense-Ciencias y Lourizán, la ETpro está ligeramente sobrestimada en estas estaciones como se puede observar claramente en la Figura 2, donde la nube de puntos aparece desplazada por encima de la línea 1:1.
Para identificar la causa que provoca esta sobrestimación se ha separado la ETPM y la ETpro en los dos factores que conforman el cálculo de la ETo mediante el método FAO-PM: el factor aerodinámico dependiente del viento y factor radiactivo dependiente de la radiación y temperatura. En la Figura 3 se muestra la ETPM y la ETpro calculada con un día de antelación para Ourense-Ciencias, separadas ambas en su factor aerodinámico y radiactivo.
Figura 3. ETo calculada por el método FAO-PM separada en sus factores aerodinámico (a) y radiactivo (b) calculada a partir de los pronósticos meteorológicos con un día de antelación (línea continua) y de los datos registrados (línea discontinua) en la estación Ourense-Ciencias.
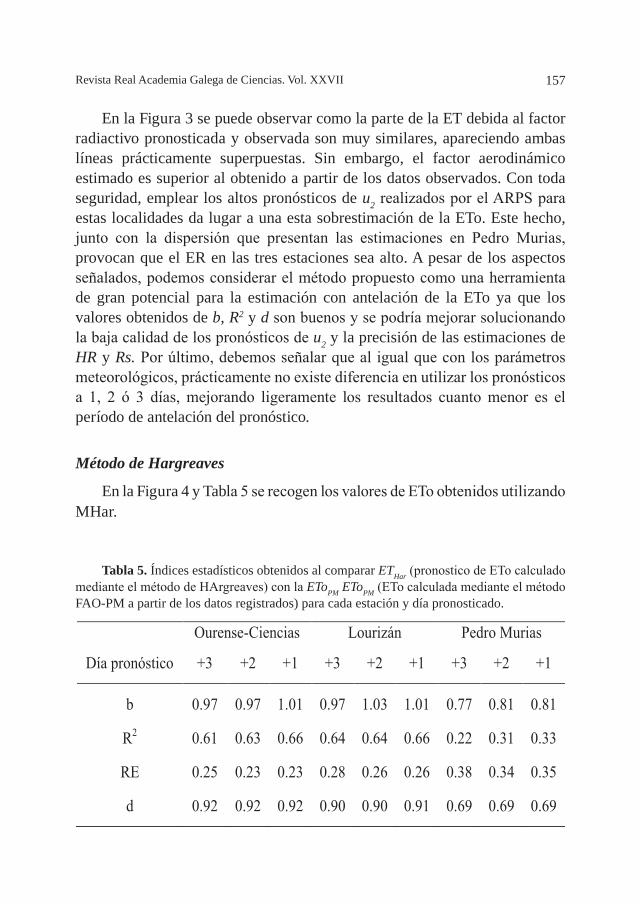
157Revista Real Academia Galega de Ciencias. Vol. XXVII
En la Figura 3 se puede observar como la parte de la ET debida al factor radiactivo pronosticada y observada son muy similares, apareciendo ambas líneas prácticamente superpuestas. Sin embargo, el factor aerodinámico estimado es superior al obtenido a partir de los datos observados. Con toda seguridad, emplear los altos pronósticos de u2 realizados por el ARPS para estas localidades da lugar a una esta sobrestimación de la ETo. Este hecho, junto con la dispersión que presentan las estimaciones en Pedro Murias, provocan que el ER en las tres estaciones sea alto. A pesar de los aspectos señalados, podemos considerar el método propuesto como una herramienta de gran potencial para la estimación con antelación de la ETo ya que los valores obtenidos de b, R2 y d son buenos y se podría mejorar solucionando la baja calidad de los pronósticos de u2 y la precisión de las estimaciones de HR y Rs. Por último, debemos señalar que al igual que con los parámetros meteorológicos, prácticamente no existe diferencia en utilizar los pronósticos a 1, 2 ó 3 días, mejorando ligeramente los resultados cuanto menor es el período de antelación del pronóstico.
Método de Hargreaves
En la Figura 4 y Tabla 5 se recogen los valores de ETo obtenidos utilizando MHar.
Tabla 5. Índices estadísticos obtenidos al comparar ETHar (pronostico de ETo calculado mediante el método de HArgreaves) con la EToPM EToPM (ETo calculada mediante el método FAO-PM a partir de los datos registrados) para cada estación y día pronosticado.
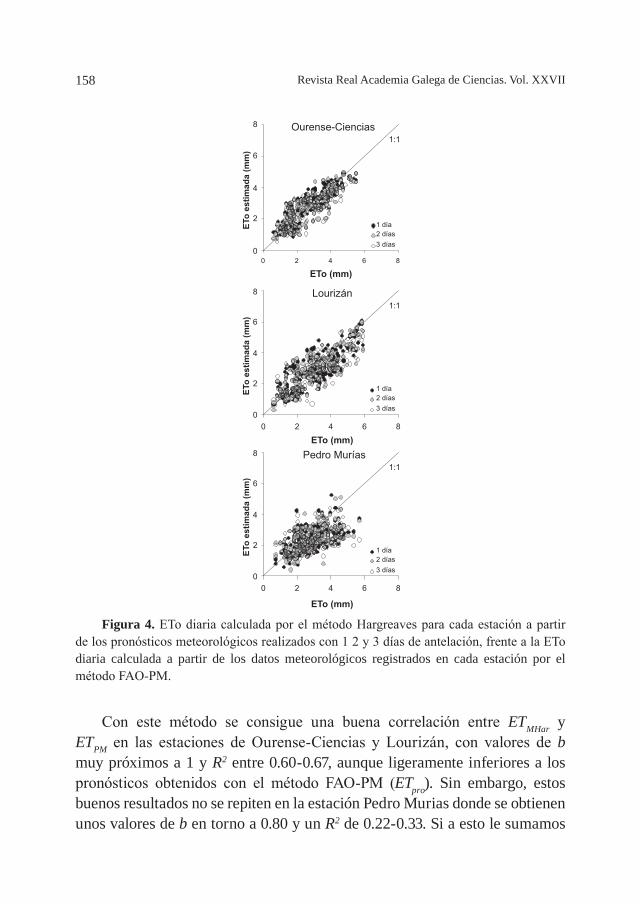
158 Revista Real Academia Galega de Ciencias. Vol. XXVII
Figura 4. ETo diaria calculada por el método Hargreaves para cada estación a partir de los pronósticos meteorológicos realizados con 1 2 y 3 días de antelación, frente a la ETo diaria calculada a partir de los datos meteorológicos registrados en cada estación por el método FAO-PM.
Con este método se consigue una buena correlación entre ETMHar y ETPM en las estaciones de Ourense-Ciencias y Lourizán, con valores de b muy próximos a 1 y R2 entre 0.60-0.67, aunque ligeramente inferiores a los pronósticos obtenidos con el método FAO-PM (ETpro). Sin embargo, estos buenos resultados no se repiten en la estación Pedro Murias donde se obtienen unos valores de b en torno a 0.80 y un R2 de 0.22-0.33. Si a esto le sumamos

159Revista Real Academia Galega de Ciencias. Vol. XXVII
un RE por encima de 0.35 y un d inferior a 0.7, podemos concluir que este método no es adecuado en esta estación.
El problema derivado del uso de los pronósticos de u2 encontrado con FAO-PM en las estaciones de Ourense-Ciencias y Lourizán no se repite con MHar ya que este método no requiere este parámetro. En consecuencia, el RE mejora notablemente, sobre todo en Ourense-Ciencias en donde se reduce prácticamente a la mitad, y también el índice d que sube a valores por encima de 0.9.
Una vez más, los resultados obtenidos con los diferentes períodos de antelación en el pronóstico son muy similares. MHar se muestra como un método válido para las estaciones de Ourense-Ciencias y Lourizán, pero no en la de Pedro Murias donde no se consigue una estimación adecuada de la ETo.
Comparación método FAO-PM y Hargreaves
A la vista de los resultados obtenidos, ninguno de los dos métodos evaluados, FAO-PM y Hargreaves se ha mostrado como adecuado para el pronóstico de la ETo en el conjunto de las estaciones. Sin embargo, en función de los resultados obtenidos, podemos separar por un lado las estaciones de Ourense-Ciencias y Lourizán y por otro Pedro Murias. En las primeras, los dos métodos han funcionado de manera muy similar obteniéndose unos resultados que ofrecen valores de a, b y R2 muy similares. FAO-PM obtiene R2 ligeramente mejores, pero el error que se produce al emplear los pronósticos de u2 del ARPS, como ya se comentó en apartados anteriores, provoca que la ETpro esté sobrestimada y en consecuencia los RE obtenidos sean mayores. MHar al no emplear este parámetro en sus cálculos presenta un RE mucho menor y un índice d por encima de 0.9, siendo por tanto el método recomendado para estas estaciones. En la estación de Pedro Murias, MHar no ha funcionado adecuadamente, obteniéndose valores de b y R2 claramente inferiores a 1. FAO-PM funciona mucho mejor, valores de b en torno a 1 pero con R2 bajos indicando baja precisión en la estimación. Probablemente, las características climáticas de esta zona sean la causa de esta mayor imprecisión. La temperatura es uno de los parámetros con más peso en la ETo y es el mejor pronosticado por el ARPS. El clima de Ribadeo, caracterizado por temperaturas suaves, hace que este parámetro tenga menor peso en el global de la ETo, aumentando por tanto el peso del resto de variables, peor

160 Revista Real Academia Galega de Ciencias. Vol. XXVII
pronosticadas, y por tanto aumentando el error de la estimación. Este mismo motivo explicaría también los malos resultados obtenidos con MHar.
Por otra parte, si atendemos a la ETo acumulada durante el período estudiado (Tabla 6) podemos ver como no existe prácticamente diferencia en usar los pronósticos con 1, 2 ó 3 días de antelación en el valor de ETo obtenido.
Tabla 6. ETo acumulada (en mm) durante el período de estudio calculada según el método FAO-PM y ETo acumulada según los pronósticos meteorológicos realizados con 1, 2 y 3 días de antelación y calculada mediante Hargreaves y FAO-PM. Entre paréntesis se muestra la diferencia en porcentaje respecto a la ETo obtenida mediante FAO-PM.
De nuevo podemos ver como el método FAO-PM se muestra el más adecuado en la estación de Pedro Murias, con un error en la ETo acumulada durante todo el periodo inferior al 5%, mientras que en las estaciones de Ourense-Ciencias y Lourizán es el método MHar el que mejor funciona, con un error inferior al 5% en el caso de Ourense-Ciencias y del 10% en Lourizán.
CONCLUSIONES
En este trabajo se ha evaluado la calidad de las estimaciones de la ETo con varios días de antelación (1, 2 o 3) en 3 localizaciones de Galicia a partir de los pronósticos meteorológicos realizados por MeteoGalicia. Para ello se han empleado los métodos FAO-PM y MHar y los pronósticos meteorológicos realizados por el modelo ARPS con 1 2 y 3 días de antelación.

161Revista Real Academia Galega de Ciencias. Vol. XXVII
Los valores de ETo obtenidos mediante ambos métodos se han comparado con los calculados por el método FAO-PM a partir los valores registrados en cada estación. Los resultados muestran que la ETo pronosticada presenta valores satisfactorios en todos los casos, con excepción del método de MHar en la estación de Pedro Murias. En las estaciones de Ourense-Ciencias y Pontevedra, a pesar de que con ambos métodos se obtiene un buen ajuste con la ETo observada, valores de b en torno a 1 y R2 por encima de 0.65, es recomendable el uso del método de MHar debido a que FAO-PM sobrestima la ETo como consecuencia de emplear los inadecuados pronósticos de u2 realizados por el modelo ARPS. Probablemente, con unos pronósticos de u2 adecuados el método FAO-PM sería el que obtendría unos mejores resultados.
En la estación de Pedro Murias es preferible el método FAO-PM ya que MHar no consigue estimaciones adecuadas, como indican los índices estadísticos empleados. De las tres estaciones también es en esta donde peores resultados se obtienen con FAO-PM, ya que a pesar de que b está dentro del rango 1±0.1, el valor de R2 es bajo, entre 0.38 y 0.51, indicando que la precisión de las estimaciones es baja. El clima de esta zona caracterizado por temperaturas más suaves provocan que este parámetro, el cual es el mejor estimado por el ARPS, tenga menos peso en el global de la ETo aumentando el del resto de parámetros, peor estimados, induciendo más error en su cálculo. Es destacable que prácticamente en todos los casos estudiados son mínimas las diferencias obtenidas en el cálculo de la ETo empleando pronósticos meteorológicos con 1, 2 o 3 días de antelación, si bien, cuanto más corto es el periodo de antelación mejor es la precisión en la estimación de la ETo.
En definitiva, podemos considerar la metodología propuesta como una herramienta adecuada para estimar con antelación la ETo, siendo de gran interés su aplicación en la gestión y diseño de regadíos, gestión de recursos hídricos, o en la prevención de incendios.

162 Revista Real Academia Galega de Ciencias. Vol. XXVII
BILIOGRAFÍA
AL-GHOBARI, H.M. (2000) Estimation of reference evapotranspiration for southern regions of Saudi Arabia. Irrig. Sci. 19:81–86.
ALLEN, R.G., PEREIRA, L.S., RAES, D., SMITH, M. (1998) Crop evapotranspiration: guidelines for computing crop requirements. FAO Irrigation and Drainage Paper No. 56. FAO, Rome, Italy.
ALLEN, R.G., PEREIRA, L.S., RAES, D., SMITH, M. (2006) Evapotranspiración de cultivo. Guía para la determinación de los requerimientos de agua de los cultivos. Estudio FAO Riego y Drenaje 56. FAO, Roma, Italia.
CASTILLO, E.F., CASTELLVÍ, F (2001) Agrometeorología. Ediciones Mundi-Prensa. Madrid.
CASTILLO RODRÍGUEZ F., MARTÍNEZ CORTIZAS A., BLANCO CHAO R. (2006) A variabilidade natural do clima de Galicia. Ed. Consellería de Medio Ambiente e Desenvolvemento Sostible, Xunta de Galicia. Santiago de Compostela, Galicia.
CHAUHAN, S., SHRIVASTAVA R.K. (2009). Performance Evaluation of Reference Evapotranspiration Estimation Using Climate Based Methods and Artificial Neural Networks. Water Resour. Manage. DOI 10.1007/s11269-008-9301-5.
CHIEW, F.H.S., KAMALADASA, N.N., MALANO, H.M., McMAHON, T.A. (1995) Penman-Monteith, FAO-24 reference crop evapotranspiration and class-A pan data in Australia. Agric. Water Manage., 28: 9–21.
GARCIA, M., RAES, D., ALLEN, R., HERBAS, C. (2004) Dynamics of reference evapotranspiration in the Bolivian highlands (Altiplano). Agric. Forest Meteorol., 125: 67–82.
GAVILÁN, P., LORITE, I.J., TORNERO, S., BERENGENA, J. (2006) Regional calibration of Hargreaves equation for estimating reference ET in a semiarid environment. Agric. Water Manage., 81: 257–281.
HARGRAVES, G.H. SAMANI, Z.A. (1985) Reference crop evapotranspiration from temperature. Applied Eng. Agri., 1: 6-99.
JABLOUN, M., SAHLI, A. (2008) Evaluation of FAO-56 methodology for estimating reference evapotranspiration using limited climatic data Application to Tunisia. Agric. Water Manage., 95: 707–715.
JIABING, C., YU, L., TINGWU, L., PEREIRA, L.S. (2007) Estimating reference evapotranspiration with the FAO Penman-Monteith equation using daily weather forecast messages. Agric. For. Met., 145: 22-35.
JENSEN, M.E., BURMAN, R.D., ALLEN R.G. (1990) Evapotranspiration and irrigation water requirements. (ASCE manuals and reports on engineering practices no. 70) American Society of Civil Engineers, New York.
MARDIKIS, M. G., KALIVAS, D. P., KOLLIAS, V. J. (2005) Comparison of Interpolation Methods for the Prediction of Reference Evapotranspiration—An Application in Greece. Water Resour. Manage., 19: 251-278.
POPOVA, Z., KERCHEVA, M., PEREIRA, L.S. (2006) Validation of the FAO methodology for computing ETo with missing climatic data application to South Bulgaria. Irrig. Drain., 55: 201–215.
SOTO, B., DÍAZ-FIERROS, F. (1996) As augas de Galicia. Balance hídrico. In: F. Díaz-Fierros (ed.), As Augas de Galicia. Consello da Cultura Galega, Santiago de Compostela.
WILLMOTT, C.J. (1982) Some comments on the evaluation of model performance. Bull. Am. Meteorol. Soc., 63: 1309–1313.

Revista Real Academia Galega de Ciencias. Vol. XXVII. Págs. 163-181 (2008)
DIFUSIONES DÉMICAS Y COMPONENTES DE LA ESTRUCTURA GENÉTICA DEL NW AFRICANO.
PERSPECTIVAS MULTIVARIANTES DE STRS AUTOSÓMICOS
B. CAEIRO*, A. FERREIRO, D. BEIROA, J. A. RODRÍGUEZ-PÉREZ, F. PICCHI-FIGUEIRA, M. M. REGUEIRO
Área de Antropoloxía Física, Facultade de Bioloxía, Universidade de Santiago de Compostela, Campus Sur, 15782 Santiago de Compostela
*Correspondencia: [email protected]
RESUMEN
Se aborda el estudio de una batería de STRs autosómicos de DNA en una muestra de 160 individuos de la población general del Norte de Marruecos. El análisis de los modelos mutacionales apunta al SMM como el mecanismo principal en la génesis de la variabilidad alélica.
Los Análisis Cladístico y Multivariante evidencian que las poblaciones del NW africano presentan una composición genética rica y compleja, resultante de contribuciones genéticas muy diversas. El aporte Negroide establecido desde le África subsahariana queda registrada como un componente minoritario en el perfil de los actuales habitantes del NW africano que, mayoritariamente, se emparentan biológicamente con las poblaciones del grupo Caucasoide, debido fundamentalmente a expansiones procedentes de Oriente Medio (Neolítico e Islamización). No obstante, las evidencias genéticas apoyan la hipótesis de una presencia temprana del componente caucasoide en la región, estimada entre 40.000-25.000 años a.C., de la que los actuales bereberes son herederos directos.
Palabras clave: ADN, estructura genética, marcadores genético-moleculares, norte de África, poblaciones africanas, polimorfismo genético, STR.

164 Revista Real Academia Galega de Ciencias. Vol. XXVII
ABSTRACT
The analysis of the genetic polymorphism of autosomal STRs among 160 unrelated individuals from Northern Morocco has been accomplished. The distribution of gene frequencies points out to SMM as the uppermost source of allele diversity.
Cladistic and Multivariant Analyses call attention to the intrincate and rich genetic composition displayed by the NW African populations. The analyses disclose a minor Subsaharian contribution into the genetic structure of NW Africa, which in actual fact are genetically close related to the Caucasoid main group. This consideration is mainly attributed to demic expansions arose from the Near East region (essentially the Neolithic and Islam expansions). Even though, early Caucasoid contributions can still be recorded at the present time among the Berber populations, as the direct descendants of these first modern humans inhabiting the region 40,000-25,000 years b.p.
Key words: African populations, DNA, genetic polymorphism, genetic structure, northern Africa, short tandem repeats.
INTRODUCCIÓN
La difusión démica y el subsiguiente flujo génico constituye un mecanismo de capital relevancia para comprender el proceso y naturaleza biológica de la expansión de los humanos modernos. El NW de Africa es sin duda un ejemplo ilustrativo, dado que se ubica en un emplazamiento geográfico que aglutina aportaciones Europeas desde la fachada nordmediterránea, así como aportes subsaharianos, y finalmente un tercer componente que procedente de Oriente Medio se expansiona a lo largo de la franja norteafricana. Las distintas poblaciones que actualmente habitan el Norte de África, no proceden de un único evento migratorio que posteriormente se ha diferenciado en la región, sino que un elevado número de poblaciones y civilizaciones han dejado su huella tanto genética como cultural en las poblaciones actuales.
El registro paleontológico aporta evidencias de restos correspondientes a los humanos modernos en el NW africano, con una antigüedad estimada en 40.000 años (Alimen, 1987), de una tipología cro-magnoide, semejante a la de sus homólogos europeos de la época. Tales poblaciones se relacionan estrechamente con la industria lítica Ateriense, ampliamente distribuida

165Revista Real Academia Galega de Ciencias. Vol. XXVII
por la región y que se vería reemplazada, sin una clara continuidad, por la industria Iberomauritana, desarrollada en el periodo comprendido entre 22.000 y 9.5000 años atrás (Newmann, 1995). Ésta, a su vez se propaga por la orilla mediterránea, a lo largo de la costa hasta alcanzar el NW del continente, concluyendo en la vertiente atlántica, al W, y Mauritania, hacia el Sur (Camps, 1974). Tal expansión se produce en un contexto de condiciones climáticas sensiblemente más favorables que las actuales, dado que por entonces el Sahara alcanzaba su máximo grado de humedad, previa a su progresiva desertificación.
Poblaciones de substrato caucasoide y subsahariano confluyeron, desde el N y S, respectivamente, en la franja Sahariana, de modo que se desarrollaron diversas culturas Pre y Neolíticas, como lo acredita el registro arqueológico (Doutour y col., 1988). La expansión Neolítica procedente de Oriente Medio alcanza el NW africano hace 5.500 años estableciendo, así, contacto con la cultura capsiense, que, desarrollada en el Mesolítico significaba una continuación evolutiva de la cultura Iberomauritana (Ferembach, 1985; Camps-Fabrer, 1989). Finalmente, ya en etapas históricas, diferentes invasiones alcanzan la zona, siendo la expansión islámica (siglo VII A.D.) la que, irradiando desde Oriente Medio, ocasiona mayor repercusión y alcance.
Los estudios de las poblaciones actuales fueron inicialmente llevados a cabo con marcadores genéticos “clásicos” (antígenos eritrocitarios y leucocitarios, proteínas plasmáticas y alozimas) (Mahmnoud y col., 1987; Cavalli-Sforza y col., 1994; Pinto y col., 1994; Arnáiz-Villena y col., 1995; Kandil y col., 1999). Tales estudios se desarrollaron a partir de una información genético-poblacional dispersa e incompleta, que limita severamente tanto la relevancia como la consistencia de las conclusiones. Más recientemente, otras aproximaciones llevadas a cabo con marcadores genéticos de ADN (básicamente mt-DNA y Y-SNPs) (Pinto y col., 1994; Rando y col., 1998; Lucotte y col., 2001; Bosch y col., , 2001; Maca-Meyer y col., 2003) mediante el análisis filogeográfico de matri y patrilinajes, han permitido avanzar notablemente en el conocimiento de los substratos genéticos pretéritos de las poblaciones actuales. No obstante tales estudios presentan un enfoque geográficamente local, destinados básicamente a rastrear e identificar diversos legados genéticos reflejados en la actual composición biológica de poblaciones puntuales.

166 Revista Real Academia Galega de Ciencias. Vol. XXVII
En el intrincado marco de componentes genéticos que han configurado el paisaje genético del NW de África, el presente trabajo aborda el análisis de una batería de marcadores de ADN, del grupo de los STRs (“Short Tandem Repeats”) autosómicos. Tal estudio se apoya asimismo en el amplio numero de datos genético-poblacionales, disponibles en la actualidad, tanto para el N. de Africa, como para las poblaciones de las áreas geográficamente concurrentes (Europa circunmediterránea, Oriente Medio, África subsahariana), que serán clave para perfilar y aportar solidez a nuestros análisis, al proporcionar un marco poblacional diverso, superando asimismo las limitaciones y sesgos inherentes a marcadores uniparentales.
MATERIAL Y METODOS
Muestra
La muestra estadística se obtuvo a partir de individuos de ambos sexos, no emparentados biológicamente, seleccionados al azar a partir de la población general árabe del Norte de Marruecos. Cada individuo seleccionado responde a los requerimientos de autoctonía, al menos dos generaciones previas, y sus procedencias geográficas corresponden a las regiones de Titt Aoouen, Fes, Al Hoseima, Taza, Nadar y Oudjda (Fig.1). Las muestras fueron obtenidas tras consentimiento de los donantes, y respetando los requerimientos éticos y de confidencialidad de la convención de Helsinki.
Figura 1. Norte de Marruecos. Agrupaciones regionales. 1: Titt’aouen; 2: Al Hoseima; 3: Taza; 4: Fes, 5: Nador; 6: Oudjda

167Revista Real Academia Galega de Ciencias. Vol. XXVII
De cada individuo se procedió a la extracción de 2 mL de sangre venosa periférica, tratada con EDTA-Na2 (10%, w/v) (10 µL/mL sangre). El paquete celular se obtuvo tras una inicial centrifugación a 3.000 r.p.m., 10 min. El DNA fue obtenido mediante extracción con solventes orgánicos (fenol/cloroformo/isoamílico) (Maniatis y col., 1982), con modificaciones propias. Tras su extracción y cuantificación, el DNA fue almacenado a -20ºC hasta su análisis.
La amplificación de los STRS se llevó a cabo mediante PCR en un GeneAmp PCR System 9700 Thermal Cycler (Applied Biosystems). Los monoplexes se desarrollaron en un volumen total de reacción de 12,5 uL (10-50 ng DNA molde, 200 uM cada d-NTP, Tris-ClH 20 mM, pH 8.4), de acuerdo con las siguientes secuencias de primers:
HUMTPOX (Anker y col., 1992)PRIMER1:5 -CAC TAG CAC CCA GAA CCG TC-3´PRIMER2:5 -CCT TGT CAG CGT TTA TTT GCC-3´HUMTH01 (Gill y col., 1992)PRIMER1:5 -GTG ATT CCC ATT GGC CTG TTC CTC-3´PRIMER2:5 -GTG GGC TGA AAA GCT CCC GAT TAT-3´HUMVWA31/A (Kimpton y col., 1992)PRIMER1:5 -CCC TAG TGA ATG ATA AGA ATA ATC-3´PRIMER2:5 -GGA CAG ATG ATA AAT ACA TAG GAT GGA TGG-3´CSF1PO (Hammond y col., 1994)PRIMER1:5 -’AAC CTG AGT CTG CCA AGG ACT AGC-3’PRIMER2:5 -TTC CAC ACA CCA CTG GCC ATC TTC-3 ’FGA (Mills y col., 1992)PRIMER1: 5 -CCA TAG GTT TTG AAC TCA CAG-3´PRIMER2: 5 -CTT CTC AGA TCC TCT GAC AC-3´
Electroforesis
La separación molecular de los fragmentos amplificados se efectuó mediante electroforesis horizontal en geles ultrafinos de poliacrilamida (PAGE), (120 mm x 220 mm x 0,4 mm) con valores de C: 5% y T entre 5% - 10%, dependiendo del tamaño molecular del STR. En cada uno de los STRs, la electroforesis se llevó a cabo con tampón Tris-ClH 0,375 M, pH 8,8 para el gel, y Tris-Glicina 0,125 M, pH 8,8, para el puente, aplicando un voltaje

168 Revista Real Academia Galega de Ciencias. Vol. XXVII
constante de 7,8 V/cm, y condiciones de refrigeración a 4º C. Completada la separación electroforética, el revelado de las bandas se desarrolló mediante tinción por Silver Staining de acuerdo con las condiciones descritas previamente (Budowle y col., 1991), con modificaciones propias (Luis y Caeiro, 1996).
Análisis estadístico
El análisis del equilibrio Hardy-Weinberg (HWE) se llevó a cabo mediante el test exacto basado en las cadenas de Markov (Guo y Thompson, 1992) (valor de dememorización 1.000; 1.000 interacciones /batch), según “GENEPOP Statistical Package” (Raymond y Rousset, 1995). La corrección de Bonferroni se aplicó mediante SISA (Simple Iteractive Statistical Analysis) (Uitenbroeck, 1997). Los modelos mutacionales para la estimación del número de alelos/locus se desarrollaron mediante IAM (Ewens, 1972) y SMM (Kimura y Crow, 1964; Kimura y Ohta, 1978).
El Análisis de Escalamiento Multidimensional (“Multidimensional Scaling”) (MDS) se desarrolló a partir del paquete estadístico SPSS v.15.0 1. El Análisis de Componentes Principales (PCA) se efectuó mediante la aplicación del NTSYS vs 1.79 (Rolf y Slice, 1992). Los árboles genéticos se elaboraron mediante estrategias de Neighbor Joining (Saitou y Nei, 1987) a partir de la matriz de Distancias Genéticas de Reynolds (Reynolds y col., 1983) y DA de Nei (Nei y col., 1983). La estimación de los valores de bootstraping de la consistencia estadística de las ramas de los árboles, se efectuó mediante PHYLIP Software Statistical Package version 3.63 (Felsenstein, 2004). Los Filogramas se efectuaron mediante el Tree Wiew version 1.6.6 (Win 32) R.D.M. PAGE, 2001.
RESULTADOS Y DISCUSION
En la Tabla 1 se relaciona la distribución de frecuencias alélicas y los parámetros biostadísticos correspondientes a la batería de los STRs investigados en el presente estudio. Se han encontrado hasta un total de 38 variantes alelicas (promedio: 7, 6 ± 0.1172 alelos/locus). Los valores insesgados de Heterozigosidad (He) (Nei y Roychoudhury, 1974) oscilan entre 0.6951 para el TPOX y 0.8557 para el locus FGA, determinando así un valor promedio de He = 0.7698.

169Revista Real Academia Galega de Ciencias. Vol. XXVII
Tabla 1. Frecuencias alélicas de los loci TPOX, VWA, TH01; FGA, CSF1PO en la población del Norte de Marruecos. He: Heterozigosidad esperada; s.e.: Error típico; HW(P): Test exacto de probabilidad de equilibrio Hardy-Weinberg; N: número total de alelos examinados.

170 Revista Real Academia Galega de Ciencias. Vol. XXVII
El análisis de la distribución de las frecuencias fenotípicas teóricas y empíricas indica que no existen evidencias estadísticas que cuestionen el carácter panmíctico de la población para los loci examinados. Los valores de HWE configuran valores de p que se enmarcan en el intervalo comprendido entre 0.9142 (TPOX) y 0.3328 (CSF1PO) (Tabla 1). Tal resultado se ve reforzada tras la aplicación de la corrección de Bonferroni ( p corregido=0.001). Los análisis de los modelos mutacionales (Tabla 2) apuntan al SMM como el modelo que mejor explica la diversidad alélica reflejada, y al slipage como el mecanismo molecular básico en la génesis de tal variabilidad alélica (Chakraborty y Daiger, 1991; Kimura y Ohta, 1978). La consideración conjunta del número de alelos/locus, He y valores de SMM, sugiere que no existen evidencia de subestructura poblacional o de efectos de “cuello de botella” (“bottlenecks”) que hayan incidido de manera relevante en el reciente devenir evolutivo de la población (Deka y col., 1992).
Al objeto de dotar de un significado biológico, en un marco poblacional suficientemente amplio, los valores de las frecuencias alélicas obtenidas en nuestro estudio, fueron integrados junto con los datos disponibles en la literatura científica. Establecemos así cuatro grandes áreas: Norte y NW
Tabla 2. Nº de alelos/locus observados y esperados por los modelos mutacionales IAM/SMM.
africano, África subsahariana, Continente Europeo y Oriente Medio, en estrechamente relacionada con nuestra población, tanto démica como geográficamente. Con ello, se fundamentan las bases requeridas para la interpretación de la estructura e historia evolutiva de nuestra población el marco del NW africano, desde una perspectiva genético-molecular.

171Revista Real Academia Galega de Ciencias. Vol. XXVII
El Substrato Caucasoide
Los valores de Distancia de Genética de Reynolds (Reynolds y col., 1983) se elaboraron a partir de la distribución de frecuencias de un total de 84 variantes alélicas diferentes. A partir de la matriz de distancias se diseñó el dendrograma sin raíz, mediante la aplicación de estrategias de Neigbor Joining (Saitou and Nei, 1987) (Fig. 2). Destaca, por una parte, la coherencia biológica de las agrupaciones resultantes, así como la robustez y consistencia estadística de las ramas, como lo reflejan los elevados valores de bootstrap. Las poblaciones norteafricanas y de Oriente Medio, ocupan posiciones centrales, derivando respectivamente de puntos nodales que en la topografía del árbol se sitúan muy próximos entre si, y que en la figura se señalan con sendas flechas. Tal proximidad sugiere una estrecha relación genética en origen, que tras la progresiva diferenciación de las ramas, desemboca en los perfiles biológicos de las actuales poblaciones.
Los análisis estadísticos llevados a cabo desde perspectivas radicalmente diferentes, proporcionan conclusiones que son congruentes con las anteriores, a la vez que amplían el horizonte de exploración de los resultados. La Proyección en el plano I/II del Análisis de Multidimensional
Figura 2. Distancias Genéticas de Reynolds: Árbol sin raíz y valores de Bootstraping

172 Revista Real Academia Galega de Ciencias. Vol. XXVII
Scaling (MDS) pone de manifiesto la relevancia de la Dimension I (70.82% de la varianza), a lo largo de la cual se produce la distribución, en áreas netamente diferenciadas, de los grandes grupos poblacionales objeto de estudio (Figura 3). Ello se refleja en una gradación de Norte a Sur de la distribución los clusters, desde las poblaciones nórdicas y centroeuropeas, que ocupan las posiciones mas positivas de la Dimensión, hasta las poblaciones subsaharianas (proyecciones más negativas) que se ubican en las posiciones opuestas, pasando por circunmediterráneos (europeos y norteafricanos).
Las poblaciones de substrato caucasoide, tanto norteafricanas como europeas si bien se esparcen en áreas diferentes, todas ellas comparten valores positivos en sus proyecciones a la Dimensión I. En otras palabras,
Figura 3. Multidimensional Scaling. Poblaciones. Subsaharianas: ANG (Angola); CAV (Cabo Verde); GUI1 (Guinea-Bissau); GUI2 (Guinea ecuatorial); HUT (Hutus); KEN (Kenia); MOZ (Mozambique); NAM (Namibia); RWA (Ruanda); TAN (Tanzania); TUT (Tutsi); USN (Afroamericanos USA); XHO (Xhosa,Sudáfrica). Norteafricanas: BER1 (Bereberes Marruecos Central); BER2( Bereberes E Marruecos); BER3 (Bereberes Egipto); BER4 (Bereberes Túnez); EGI (Musulmanes Egipto); MAR1(Musulmanes Marruecos); MAR2(Musulmanes N Marruecos), TUN(Túnez). Europeas: ALE (Alemania); AND (Andalucía); AUS (Austria); BEL (Bélgica); CHI (Chipre), CLM (Castilla-La Mancha); FIN (Finlandia); GRE (Grecia); MAL (Malta); POL (Polonia); POR (Portugal); SIC (Sicilia).
esto indica que en la actualidad se ha producido una diferenciación démica que, en la Figura 3 se expresa en la delimitación de clusters bien definidos. Tal modelo sugiere la existencia de un substrato ancestral común, previo a la diferenciación de las poblaciones actuales.

173Revista Real Academia Galega de Ciencias. Vol. XXVII
El origen del componente caucasoide de las poblaciones norteafricanas es en la actualidad objeto de debate. El registro osteológico atestigua la presencia de poblaciones de humanos modernos en la región, con una antigüedad estimada en 40.000 años. Presentaban una tipología arcaica del tipo Cromagnoide, estrechamente asociados a la expansión de la industria Ateriense, que se expandía por todo el NW desde el Maghreb hasta el Sahara. Consiguientemente, la tipología de tales ocupantes se relacionaba estrechamente con la de sus homólogos europeos nordmediterráneos de la época ( Camps, 1974; Alimen, 1987; Doutour y col., 1988). Otros autores
Figura 4. Análisis de Componentes Principales. Valores de proyección al Eje II.
postulan una introducción tardía del componente caucasoide en el NW africano, como consecuencia de la expansión Neolítica, procedente de Oriente Medio, a lo largo de la costa Norte del continente. No obstante, esta hipótesis no encaja con las evidencias paleontológicas que atestiguan la presencia de componentes caucasoides diferenciados, en algunas poblaciones del NW africano (v.g, bereberes), decenas de miles de años antes de la expansión Neolítica, y como lo corroboran asimismo los análisis de los marcadores genéticos. El análisis detallado de la información extraída a partir de los marcadores autosómicos de ADN aporta claves esclarecedoras. En la Fig. 4, se reflejan los valores de proyección al eje II de los eigenvectores, tras el Análisis de Componentes Principales. Dicho eje II es especialmente relevante en la diferenciación de los clusters caucasoides. Se aprecia que las muestras bereberes presentan sistemáticamente proyecciones positivas, claramente alejadas de las restantes poblaciones, (árabes norteafricanos, Oriente Medio y europeas), cuyas posiciones se solapan entre si. Esto parece indicar que en tanto entre los bereberes ya existía un substrato caucasoide diferenciado tal como lo indican sus proyecciones al Eje II, las restantes poblaciones aun no habían diferido entre si. De modo que estas ultimas parecen descender de un antecesor común, también caucasoide, pero de naturaleza genética diferente al bereber, y cuyo origen es comparativamente mas reciente, lo que explicaría que, consecuentemente, sus poyecciones al Eje II se solapen entre si.

174 Revista Real Academia Galega de Ciencias. Vol. XXVII
Los alelos TPOX*9, D8S1179 ( alelos 13 y 14), D3S1358 (alelos 16 y 17) juegan un papel decisivo.
A partir de la información proporcionada por los patrones filogeográficos de los subclados del Haplogrupo I, derivados de la mutación M170, se estima que el inicio de la divergencia entre las poblaciones de Oriente Medio y el continente europeo se originaría hace 25.000 años aproximadamente (Torroni y col., 1998; Semino y col., 2000; Cinnioglu y col., 2004). La integración de este dato en lo referente al poblamiento del NW africano con la información discutida en párrafos anteriores, respalda la idea de la presencia temprana de los primeros componentes caucasoides en el NW Africano en los predecesores directos de los actuales bereberes, y cuyo origen surge en el intervalo cronológico comprendido entre los 40.000 y 25.000 años.
La Expansión Islámica
La contribución genética resultante de la expansión islámica producida a lo largo de la costa Norte del continente africano, constituye un segundo pilar fundamental para explicar la incardinación de las poblaciones norteafricanas dentro del grupo caucasoide. Tal expansión, de Este a Oeste originada desde Oriente Medio, no es sólo de carácter socio-cultural sino también de índole genética, y como tal ha dejado un profundo impacto en la actual composición del panorama genético norteafricano. A diferencia de otras contribuciones démicas acaecidas en la región, en el caso de la expansión islámica se dispone de una información precisa tanto en lo referente al período cronológico como en la identificación de las poblaciones paternas implicadas, así como de la ruta seguida en tal difusión.
En la Figura 5 se presentan los resultados de los análisis de la contribución genética debida a la islamización a partir de los patrones de distribución de las frecuencias alélicas, mediante Análisis de Componentes Principales. Al objeto de centrar la atención en este aspecto, se han considerado preferentemente poblaciones norteafricanas y de Oriente Medio, tomando las europeas, como un referente vinculado. Por una parte, la proyección I/II (86.74 %de la varianza total) pone de manifiesto una gradación de las proyecciones poblacionales a lo largo del Eje I como consecuencia del flujo génico E-W. Las poblaciones genéticamente más próximas a las del Oriente Medio tienden a ser las del Este norteafricano. Por el contrario, la población de Marruecos aquí estudiada (MAR1) ocupa

175Revista Real Academia Galega de Ciencias. Vol. XXVII
Figura 5. Análisis de componentes principales. Proyección I/II. ÁREA I: BER1 (Bereberes Marruecos Central); BER2 (Bereberes E Marruecos); BER3 (Bereberes Egipto); BER4 (Bereberes Túnez); COP (Coptos, Egipto); EGI (Musulmanes Egipto); MAR1 (Musulmanes Marruecos); MAR2 (Musulmanes N Marruecos); SAH (Sáhara occidental); TUN (Túnez); TUN2 (Zriba, Túnez). ÁREA II: ARA (Arabia Saudí); GEO (Georgia); IRK (Irak); IRN (Irán); JOR (Jordania); QAT (Qatar); SIR (Siria), TUR (Turquía), YEM (Yemen). ÁREA III: ALE (Alemania); AND (Andalucía); BEL (Bélgica); BIE (Bielorrusia); CAL (Calabria, Italia); CHI (Chipre); CLM (Castilla-La Mancha); GRE (Grecia); HUN (Hungría); MAL (Malta); POL (Polonia); POR (Portugal); SER (Serbia); SIC (Sicilia).
en el plano posiciones distanciadas, agrupándose con otras muestras del NW africano (MOR2, ARG TUN, SAH, MZBT). Tal gradación debida a los fenómenos de flujo génico se plasma en diversas aleloclinas, particularmente los alelos TPOX*8, CSF1PO (alelos 10 y 12), D5S818*13, D13S317*9; TH01 (alelos 9 y 10), VWA*15, D21SS11*28. Es, asimismo, interesante notar la influencia de las poblaciones de Oriente Medio, en las muestras europeas circunmediterráneas, especialmente las más orientales (Chipre, Grecia, Malta) las que presentan una mayor cercanía, en el plano, al área en la que se distribuyen las muestras de Oriente Medio, en contraposición a las mediterráneas occidentales (Península Ibérica: Andalucía, Portugal; Castilla-La Mancha). Tal patrón de distribución reproduce en el mediterráneo europeo la gradación Este a Oeste referida en la costa norteafricana.
Los patrones filogeográficos sugieren que la contribución genética impresa en las actuales poblaciones del Norte Africano no obedece tanto a un mero aporte puntual y masivo, por una invasión territorial, sino más

176 Revista Real Academia Galega de Ciencias. Vol. XXVII
bien, a una difusión gradual y progresiva de Este a Oeste. Una vez unificados en el ámbito cultural (lingüístico, religioso, etc), como consecuencia de la expansión islámica, en los territorios conquistados, en los últimos trece siglos aproximadamente, se desarrollan condiciones favorables para una mayor permeabilidad y difusión genética, tal como lo demuestran los patrones clinales mostrados en los Análisis Multivariantes. Tal interpretación se ve asimismo apoyada por los resultados derivados del análisis del mt-DNA, cuya herencia materna (contribución femenina) define modelos de gradación de los matrilinajes a lo largo de la costa norteafricana (Pinto y col., 1994; Maca-Meyer y col., 2003). En consecuencia, tales resultados parecen descartar que la contribución biológica acarreada por la islamización en el Africa nordsahariana se haya limitado a una mera contribución puntual por una invasión (masculina) masiva.
Tanto los análisis de PCA como los MDS proponen que la difusión démica que acompañó a la expansión islámica se produjo preferentemente a lo largo de la franja costera y regiones adyacentes. A tal efecto, los alelos TPOX*9, D3S1358 (alelos 16 y 17), D8S1179 (alelos 13 y 14), VWa *12 son especialmente informativos. Tanto por la información derivada de la Figura 4 como de la Figura 5, se aprecia que las poblaciones Bereberes, fragmentadas y diseminadas en el interior geográfico norteafricano, experimentaron, ya desde el Neolítico y etapas inmediatamente anteriores, la diferenciación biológica subsiguiente a la deriva genética originada por el aislamiento geográfico antes referido. Eso explica que en los Análisis de Componentes Principales se distribuyan de manera dispersa y ocupen en el plano I/II posiciones periféricas, claramente distanciadas de las otras poblaciones costeras norteafricanas. En tales poblaciones bereberes, la expansión islámica parece haber tenido más bien un significado cultural, en tanto que, comparativamente, el impacto de la contribución genética parece haber sido escaso. En consecuencia, los Bereberes parecen haber preservado hasta la actualidad una parte significativa de la memoria genética de los primeros grupos paleolíticos de Homo sapiens que habitaron la región.
La Contribución Subsahariana
El desierto del Sahara constituye una frontera natural entre el Norte Africano y el África subsahariana. El Análisis de MDS de la distribución de frecuencias alélicas, circunscrito a ambos contextos poblacionales

177Revista Real Academia Galega de Ciencias. Vol. XXVII
(Figura 3), es especialmente aclaratorio. La proyección I/II presenta a las poblaciones norteafricanas agrupadas conjuntamente con las europeas del ámbito circundemiterráneo, en un área específica y diferenciada, netamente alejada de las proyecciones que agrupan al Africa subsahariana (Cuadrantes II y III, respectivamente), lo que parecería indicar que el legado genético de éstas últimas en la composición démica del NW africano no ha sido particularmente relevante. Una parte substancial de los patrones de agrupamiento es desempeñada por los alelos D13S317*9, D18S*13, TPOX*8, TPOX*9, CSF1PO*13; FGA*21, TH01*9.3.
Si bien en la actualidad el Sahara constituye una barrera natural que limita notablemente el flujo génico en ambas direcciones, los resultados arriba señalados no parecen ser acordes con la historia demográfica de ambas regiones. Esto es, durante miles de años el Sahara, lejos de las condiciones climáticas actuales, estuvo habitado por poblaciones metamórficas resultantes de la mezcla démica de componentes Caucasoides norteafricanos, y por contribuciones Negroides, al Sur, tal como lo refleja el registro arqueológico de la época (Camps, 1974; Newmann, 1995). Tal etapa alcanza su apogeo de máxima humedad hace 9.000 a. C., el denominado “Sahara Fértil”. A partir de entonces sobreviene una progresiva aridificación, que en etapas sucesivas conduce a las condiciones desérticas actuales. Tal proceso climático desencadenó la redistribución de los pobladores saharianos en dos direcciones opuestas, hacia el Sur (Sahel) y hacia el Norte (Maghreb) y NW africano (Alimen, 1987; Barry Cox y Moore, 1993). Teniendo en cuenta tales eventos démicos, cabría esperar que en la composición genética actual del NW africano, se registrase un significativo aporte genético del substrato Negroide, notablemente superior al que se observa en la actualidad, a tenor de los datos aportados por los STRs autosómicos. Pero tal no es el caso.
Por otra parte, de acuerdo con la información histórica disponible, conocemos que las poblaciones de los Afroamericanos (USN) y Cabo Verde (CAV) constituyen claros ejemplos actuales de poblaciones metamórficas originadas como consecuencia de un repetido aporte caucasoide sobre un substrato negroide inicial, de naturaleza bantú. (Parra y col., 1995, 1998). Se advierte que las posiciones en que se ubican Afroamericanos USA (USN) y Cabo Verde (CAV), claramente se incardinan en el área en la que se distribuyen las poblaciones subsaharianas, no obstante, en ambas poblaciones se aprecia un claro acercamiento a las regiones del plano propias de las poblaciones

178 Revista Real Academia Galega de Ciencias. Vol. XXVII
caucasoides, demostrando su carácter de poblaciones híbridas. Por el contrario, las poblaciones norteafricanas no reflejan tales pautas, lo que parece sugerir una historia evolutiva netamente diferente.
Cabría preguntarse si tales resultados indican que en las poblaciones del NW africano ha habido un escaso aporte genético subsahariano. Un análisis más detenido nos indica que tal apreciación no es suficientemente sólida; es importante tener en cuenta que las muestras subsaharianas que relacionan en la Figura 3 corresponden a poblaciones de substrato y cultura bantú, cuyo perfil genético es de naturaleza diferente al componente negroide de las primigenias poblaciones autóctonas saharianas. Por consiguiente, el alejamiento de las posiciones que se advierte en la proyección, entre poblaciones subsaharianas y NW africano no impugna una significativa contribución genética subsahariana, acaecida en etapas pretéritas, en el transcurso del período de convivencia en el Sahara Fértil. La expansión bantú que introdujo y expandió el Neolítico en el África subsahariana desde hace 5.000 años aproximadamente (Cavalli-Sforza y col., 1994; Newmann, 1995) de manera progresiva reemplazaría y desbancaría a otras poblaciones negroides autóctonas, tecnológicamente menos desarrolladas, hasta su confinamiento y extinción final. Ello explicaría la ausencia de estas últimas en el actual paisaje genético del África subsahariana, habiendo así experimentado un proceso parejo al que todavía se puede observar, en la actualidad, en las poblaciones de Pigmeos y Khoisánidos.

179Revista Real Academia Galega de Ciencias. Vol. XXVII
BIBLIOGRAFÍA
ALIMEN, H. (1987) Evolution du climat et des civilisations depuis de 40 000 ans du nord au sud du Sahara occidental (Premières conceptions confrontées aux donnés récentes). Bull. LAssoc. Franç. L Étude Quaternaire, 4: 25-227.
ANKER, R., STEINBRUECK, T., DONNIS-KELLER, B. (1992) Tetranucleotide repeat polymorphism at the human thyroid peroxidase (hTPO) locus. Hum. Mol. Genet, 1: 137.
ARNAIZ-VILLENA, A., BENMAMAR, D., ALVAREZ, M., DIAZ-CAMPOS, N., VARELA, P., GOMEZ-CASADO, E., MARTINEZ-LASO, J. (1995) HLA alleles and haplotype frequencies in Algerians. Relatedness to Spaniards and Basques. Hum. Immunol., 43: 259-268.
BARRY COX, C., MOORE, P.D. (1993) Biogeography. An Ecological and Evolutionary Approach. Blackwell, Publ. N.York.
BOSCH, E., CALAFELL, F., COMAS, D., OEFNER, P.J., UNDERHILL, P.A., BERTRANPETIT J. (2001) High-resolution shows a sharp discontinuity and limited gene flow between northeastern Africa and Iberian Peninsula. Am. J. Hum. Genet. 68: 1019-1029.
BUDOWLE, B., CHAKRABORTY, R., GIUSTI, A.M., EISENBERG, A.J., ALLEN, R.C. (1991) Analysis of the VNTR locus D1S80 by the PCR followed by high-resolution PAGE. Am. J. Hum. Genet., 48: 137-144.
CAMPS, G. (1974) Les Civilisations Préhistoriques de l Afrique du Nord et du Sahara. Doin Eds., Paris.
CAMPS-FABRER, H. (1989) Capsien et Natoufien au Proche-Orient. Colloque “L Homme maghrebin et son environnement depuis 100000 Ans”. Maghina (Algerie) Trav. Du LAMPO, 71-104.
CAVALLI-SFORZA, L.L., MENOZZI, P., PIAZZA, A. (1994) The History and Geography of human genes. Princeton, New Jersey.
CHAKRABORTY, R., DAIGER, S. (1991) Polymorphisms at VNTR loci suggest homogeneity of a white population of Utah. Hum. Biol, 63: 571-587.
CINNIOGLU, C., KING, R., KIVISILD, E., KALFOGLU, E., ATASOV, S., CAVALLERI, G.R., LILIIE, A.S., ROSEMAN, C.C., LIN, A.A., PRINCE, K., OEFNER, P.J., SHEN, P., SEMINO, O., CAVALLI-SFORZA, L.L., UNDERHILL, P.A. (2004) Excavating Y-chromosome haplotype strata in Anatolia. Hum. Genet., 114: 127-148.
DEKA, R., DECROO, S., YU, L.M., FERRELL, R.E. (1992) Variable Number of Tandem repeats (VNTR) polymorphism at locus D17S5 (YNZ22) in four ethnically defined human populations. Hum. Genet., 90: 86-90.
DOUTOUR, O., VERNET, R., AUMASSIP. G. (1988) Le peuplement préhistorique du Sahara. In “Milieux, hommes et techniques du Sahara préhistorique. Problèmes actuelles.” Aumassip, G. ed. Editions L Harmattan, Paris.
EWENS, W.J. (1972) The sampling theory of selectively neutral alleles. Theor. Pop. Biol., 3: 87-112.
FELSENSTEIN J. (2004) PHYLIP (Phylogeny Interference Package) version 3.63. http://evolution.gs.washington.edu/phylip.html
FEREMBACH, D. (1985)On the Origin of the Iberomaurisians (Upper Paleolithic: North Africa). A New Hypothesis. J. Hum. Evol., 14: 393-397.
GILL, P., KIMPTON, C.P., SULLIVAN, K. (1992) A rapid polymerase chain reaction method for indentifying fixed specimens. Electrophoresis, 13: 173-175.

180 Revista Real Academia Galega de Ciencias. Vol. XXVII
GUO, S. W., THOMPSON, E.A. (1992) Performing the exact test of Hardy-Weinberg proportion for multiple alleles. Biometrics, 48: 361-372.
HAMMOND, H.A., JIN, L., ZHONG, C.T., CASKEY, T., CHAKRABORTY, R. (1994) Evaluation of thirteen STR loci for use personal identification applications. Am. J. Hum. Genet., 55: 175-189.
KANDIL, M.; MORAL, P.; ESTEBAN, E., AUTORI, L., MAMELI, G.E., ZAOUI, D., CALO, C., LUNA, F., VACCA, L., VONA, G. (1999) Red cell enzyme polymorphisms in Moroccans and Southern Spaniards: new data from the genetic history of Western Mediterranean. Hum. Biol., 71: 791-802.
KIMURA, K., CROW, J.F. (1964) The number of alleles that can be maintained in a finite population. Genetics, 49: 725-738.
KIMURA, M., OHTA, T. (1978) Stepwise Mutation Model and distribution of alleles frequencies in a finite population. Proc. Natl. Acad. Sci. USA, 75: 2868-2872.
LUCOTTE G., GÉRARD, N., MERCIER, G. (2001) North African Genes in Iberia Studied by Y-Chromosome DNA Haplotype 5. Hum. Biol., 73: 763-769.
LUIS, J.R., CAEIRO, B. (1996) Further Genetic Variability of the VNTR D1S80 (pMCT118): Correspondence Analysis Studies. Am. J. Hum. Biol., 8: 81-87.
MACA-MEYER, N., GONZÁLEZ, A.M., PESTANA, J., FLORES, C., LARRUGA, J.M., CABRERA, V.M. (2003) Mitochondrial DNA transit between West Asia and North Africa inferred from U6 phylogeography. BMC Genetics, 4: 15-26.
MANIATIS, T., FRITSCH, E.F., SAMBROCK, J. (1982) Molecular Cloning: a Laboratory Manual. Cold Spring Harbor, NY.
MAHMOUD, L. A., IBRAHIM, A.A., GHONEM, H.R., JOUVENCEAUX, A. (1987) Human Blood Groups in Dakhlya, Egypt. Ann. Hum. Biol., 14: 487-493.
MILLS, K.A., EVEN, D., MURRAY, J.C. (1992). Tetranucleotide repeat polymorphism at the alpha fibrinogen locus (FGA). Hum. Mol. Genet., 779.
NEI, M., TAJIMA, F., TATENO, Y. (1983) Accuracy and estimated phylogenetic trees from molecular data. J. Mol. Evol., 19: 153-170.
NEI, M., ROYCHOUDHURY, A.K. (1974) Sampling variants of heterozygosity and genetic distance. Genetics, 76: 379-390.
NEWMANN, J.L. (1995) The peopling of Africa: a geographic interpretation. New Haven, Yale University Press.
PARRA, E.J., TEIXEIRA-RIBEIRO, J.C., CAEIRO, B. (1995) Genetic structure of the population of Cabo Verde (West Africa): Evidence of substantial European admixture. Am. J. Phys. Anthropol., 97: 381-389.
PARRA, E.J., MARCINI, J.A., AKEY, J. , MARTINSON, J., BATZER, M.A., COOPER, R., FORRESTER, T., ALLISON, D.B., DEKA, R., FERRELL, R.E., SHRIVER, M.D. (1998) Estimating African-American admixture proportions by use of population-specific alleles. Am. J. Hum. Genet., 63: 1839-1851.
PINTO, F., GONZÁLEZ, A.M., HERNÁNDEZ, M., LARRUGA, J.M., CABRERA, V.M. (1994) Genetic relationship between the Canary islanders and their African and Spanish ancestors inferred from mitochondrial DNA sequences. Ann. Hum. Genet., 60: 321-330.
RANDO, J.C., PINTO, F., GONZÁLEZ, A.M., HERNÁNDEZ, M., LARRUGA, J.M., CABRERA, V.M., BANDELT, H.J.(1998) Mitochondrial DNA analysis of north-west African populations reveals genetic exchange with European, near-eastern and sub-Saharan populations. Ann. Hum. Genet., 62: 531-550.

181Revista Real Academia Galega de Ciencias. Vol. XXVII
RAYMOND, M., ROUSSET, F. (1995) GENEPOP (version 1.2): a population genetics software for exact tests and ecumenicism. J. Hered. 86: 248-249.
REYNOLDS, J., WEIR, B.S., COCKERHAM, C.C. (1983) Estimation of the coancestry coeficient: basis for a short term genetic distance. Genetics, 105: 767-779.
ROHLF, F.J., SLICE, D.E. (1992) NTSYS-pc 1.70. Stony Brook, N.Y. Applied Biostatistics, Inc.
SAITOU, N., NEI, M. (1987) The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol. Biol. Evol., 4: 406-425.
SEMINO, O., PASSARINO, G., OEFNER, P., LIN A. A., ARBUZOVA, S., , BECKMAN, L.E., DE BENEDICTIS, G., FRACALACCI, P., KOUVATS, A., LIMBORSKA S., MARCIKIAE, M., MIRA, A., NIKA, B., PRIMORAC, D., SANTACCHIARA-BENERECETTI, A. S., CAVALLI_SFORZA, L.L., UNDERHILL, P.A. (2000) The Genetic Legacy of Paleolithic Homo sapiens in Extant Europeans: A Y chromosome Perespective. Science, 29: 1155-1159.
TORRONI, A., BANDELT, H.J., D´URBANO, L., LAHERMO, P., MORAL, P., SELLITO, D., RENGO, G., FORSTER, P., SAVONTAUS, M.L., BONNÉ-TAMIR, B., SCOZZARI, R. (1998) mt-DNA analysis reveals a major late Paleolithic expansion from southwestern to Northeastern Europe. Am. J. Hum. Genet., 62: 1137-1152.
UITENBROECK, D.G. (1997) SISA (Simple Interactive Statistical Analysis).http: //home.clara.net/sisa/


Revista Real Academia Galega de Ciencias. Vol. XXVII. Págs. 183-203 (2008)
LA CORRIENTE DE MALVINAS: ¿UNA VÍA DE DISPERSIÓN PARA CNIDARIOS
BENTÓNICOS DE AGUAS FRÍAS?
M. O. ZAMPONI
CONICET. Laboratorio de Biología de Cnidarios (LABIC)http://www.labic.8m.com
Facultad de Ciencias Exactas y Naturales. Departamento de Ciencias Marinas.Universidad Nacional de Mar del Plata. Funes 3250. B760 2AYL Mar del Plata. Argentina.
e-mail: [email protected]: 0054 (00223) 475-3554
RESUMEN
La Corriente de Malvinas constituye una vía de dispersión para aquellas especies de cnidarios bentónicos que tienen la mayor concentración en la subregión Templada Fría (= subregión Patagónica o Magallánica).Los cnidarios bentónicos como organismos suspensívoros pasivos encuentran en la Corriente Circumpolar Antártica y en su continuidad la Corriente de Malvinas, zonas de alta productividad biológica que contribuye al asentamiento de poblaciones constituidas por especies poseedoras de un sistema esqueletal rígido ( calcáreo o espicular).
Palabras clave: Corriente de Malvinas, cnidaria, organismos bentónicos.
ABSTRACT
The Malvinas s Current is a dispersion way for species like cnidarians that they have big concentration Polar Front. To be benthic cnidarians species

184 Revista Real Academia Galega de Ciencias. Vol. XXVII
feeders passive organism find in the Antarctic Circumpolar Current and her branch Malvinas´ Current, zones of high biologic productivity. This zones let the settlement of species with a squeletal system (calcareous or spicule).
Key words: Cnidaria, Malvinas s Current, benthic organism.
INTRODUCCIÓN
El área del presente estudio está circunscripta a la región Subantártica o Notal (Boschi, 1976), también denominada Antiboreal por Ekman (1953) y Kramp (1959) y se extiende por el océano Pacífico hasta el sur de Chile. Esta región está conformada por subregiones o provincias conocidas como subregión Templada Cálida y Templada Fría, también denominadas respectivamente subregión Argentina o Bonaerense y Patagónica o Magallánica. La primera de ellas se extiende hacia el norte hasta Cabo Frío a una latitud de 23°S y hacia el sur hasta los 43°/44°S; la subregión Templada Fría comprende el litoral de Tierra del Fuego, Islas Malvinas y la plataforma patagónica y su límite norte se establece aproximadamente entre los 34° y 35°S.
Figura 1. Aspecto general de la Región Antartica y Región Subantartica.
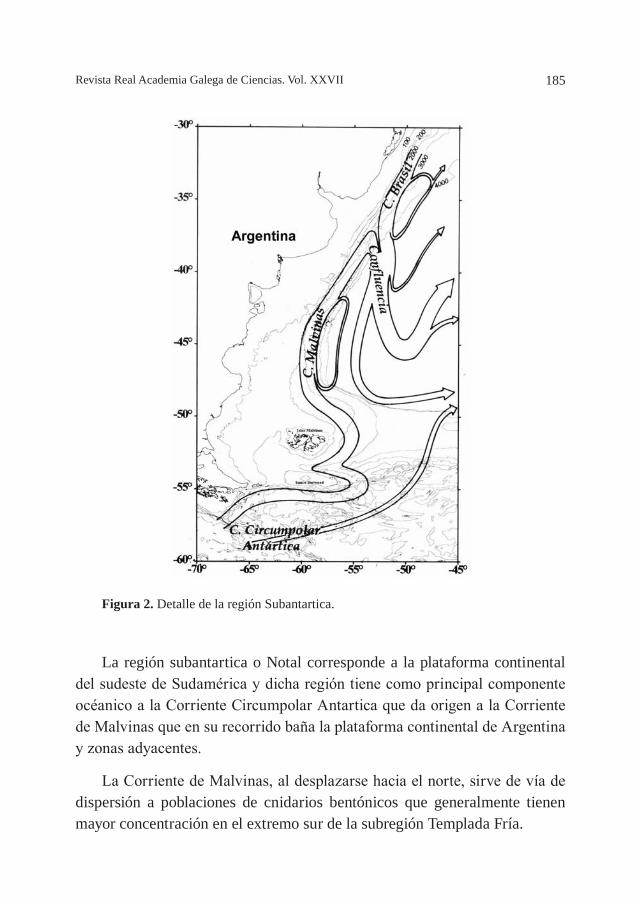
185Revista Real Academia Galega de Ciencias. Vol. XXVII
Figura 2. Detalle de la región Subantartica.
La región subantartica o Notal corresponde a la plataforma continental del sudeste de Sudamérica y dicha región tiene como principal componente océanico a la Corriente Circumpolar Antartica que da origen a la Corriente de Malvinas que en su recorrido baña la plataforma continental de Argentina y zonas adyacentes.
La Corriente de Malvinas, al desplazarse hacia el norte, sirve de vía de dispersión a poblaciones de cnidarios bentónicos que generalmente tienen mayor concentración en el extremo sur de la subregión Templada Fría.

186 Revista Real Academia Galega de Ciencias. Vol. XXVII
MATERIAL Y METODOS
El material utilizado en este análisis fueron diversas especies de cnidarios bentónicos pertenecientes a la subclase Leptothecatae y a los órdenes Stylasterina, Gorgonacea, Pennatulacea, Alcyonacea, Actiniaria y Scleractinia.
El área de distribución de las especies corresponde a la denominada región Subantártica o Notal situada entre 35° y 55°LS. Dicha área fue dividida en una cuadrícula de 5° de LS y 5° de LW para precisar lo más aproximado posible los puntos de encuentro de las diversas coordenadas que indican la posición de hallazgo de las epecies indicadas.
Dada la diversidad de cnidarios bentónicos existentes en el área de estudio se seleccionaron aquellas especies que tienen distribución en la Corriente Circumpolar Antártica y que también se localizan en la subprovincia Patagónica o Magallánica, para luego ser halladas a través de la Corriente de Malvinas en sectores de la subprovincia Argentina o Bonaerense.
Las fuentes de consulta bibliográfica para el actual estudio fueron los trabajos de Blanco y col. (1994, 2000a, b, 2003), Cairns (1982, 1983), Pérez (1999) y Riemann-Zurneck (1973, 1978, 1980, 1986).
RESULTADOS
La Corriente de Malvinas y zona de influencia.
El agua de la plataforma continental argentina es de origen subantartico y la misma ingresa a la plataforma entre las Islas Malvinas y Tierra del Fuego, modificada por el aporte de aguas diluidas del Estrecho de Magallanes (Piola y Rivas, 1997). Las aguas del extremo norte de la Corriente Circumpolar Antártica da origen a la Corriente de Malvinas, aunque debido al elevado transporte de esta última, el impacto de la descarga continental no es significativa y la salinidad de esta agua es en general del orden de 33,75 UPS o superior. En la boca del Estrecho de Magallanes se observa un mínimo de salinidad (S < 32,5 UPS) que se extiende hacia el NNE próximo a la costa hasta 47°S, para luego separarse de ésta y ubicarse en la plataforma central.

187Revista Real Academia Galega de Ciencias. Vol. XXVII
El mínimo de salinidad (S <33,6 UPS) se detecta frente a Mar del Plata (38°S) entre 140 y 180 km de la costa.
Entre Tierra del Fuego y las Islas Malvinas existen regiones relativamente profundas y la circulación parece seguir los complejos contornos de la batimetría. El flujo en regiones profundas y cerca del talud es relativamente rápido comparado con el que se produce sobre la plataforma; la densidad de esta agua es superior a la de la plataforma ubicada al norte y al oeste. Esta masa de agua fluye hacia el sur de las Islas Malvinas para reencontrarse con el flujo principal de la Corriente de Malvinas cerca de 53°S 56°W, mientras que la influencia de esta agua sobre la plataforma continental es escasa.
En los golfos San Jorge, Nuevo, San José y San Matías se restringe el intercambio con el mar abierto y las aguas resultan influenciadas por la fuerza atmosférica; estas características del flujo atmosférico y la limitada renovación de las aguas producen una amplitud térmica anual en los golfos que en el mar abierto. Las aguas cálidas alcanzarían el litoral bonaerense (41°S) en octubre, la península Valdés en enero y el golfo San Jorge en febrero; este flujo estaría compuesto por aguas de origen subantártico y las aguas cálidas del sur del litoral bonaerense estarían asociadas a intrusiones de la Corriente de Brasil. Boltovskoy (1970 y 1981) sugiere que la presencia de aguas cálidas en el litoral bonaerense se restringe al norte de los 40°S y que la misma se debe principalmente al calentamiento estacional.
Las provincias zoogeográficas Argentina y Magallánica han sido tradicionalmente reconocidas para este sector del Atlántico Sudoccidental, a partir de información hidrográfica y biológica, de manera tal que la distribución de especies se halla relacionada con la dinámica de la Corriente Circumpolar Antártica y su prolongación en la conocida Corriente de Malvinas.
Los resultados de campañas oceanográficas a través de las asociaciones bentónicas corroboran la clásica división zoogeográfica y sugieren la posibilidad de subdividir el sector Atlántico de la provincia Magallánica en dos distritos: uno interno asociado a la Corriente Patagónica (Brandhorst y Castello, 1971) y otro externo relacionado a la Corriente de Malvinas (Bastida y col., 1992 en Bastida y col., 2007). Según Bastida y col. (1992), estos distritos podrían ser denominados Patagónico y Malvinense respectivamente.

188 Revista Real Academia Galega de Ciencias. Vol. XXVII
Listado de las especies halladas
Subclase Leptothecatae
Abietinella operculata (Jaderholm, 1903)
Acryptolaria conferta (Allman, 1877)
Hebella striata Allman, 1888
Lafoea fruticosa (M. Sars, 1851)
Synthecium robustum Nuttin, 1904
Phialella chilensis (Hartlaub, 1905)
Amphisbetia operculata (Linnaeus, 1758)
Sertularella conica Allman, 1877
Sertularella gayi gayi (Lamouroux, 1821)
Sertularella robusta Coughtrey, 1876
Symplectoscyphus magellanicus (Marktanner-Turneretscher, 1890)
Symplectoscyphus subdichotomus (Kirchenpauer, 1884)
Symplectoscyphus filiformis (Allman, 1888)
Symplectoscyphus modestus (Hartlaub, 1900)
Orden Stylasterina
Sporadopora dichotoma (Moseley, 1876)
Errina echinata (Moseley, 1879)
Errina labiata (Moseley, 1879)
Errina lowei Cairns, 1983
Stylaster densicaulis Moseley, 1879
Orden Gorgonacea
Primnoella biserialis Wright y Studer, 1889
Primnoella compressa Kukenthal, 1908

189Revista Real Academia Galega de Ciencias. Vol. XXVII
Convexella magelhaenica (Studer, 1879)
Thouarella acanthina (Wright y Studer, 1889)
Thouarella antarctica Valenciennes, 1846
Orden Pennatulacea
Renilla reniformes (Pallas, 1766)
Orden Alcyonacea
Alcyonium paessleri May, 1899
Alcyonium patagonicum (May, 1899)
Orden Actiniaria
Actinostola crassicornis (Hertwig, 1882)
Bolocera tuediae subsp. occidua (McMurrich, 1893)
Hormathia pectinata (Hertwig, 1882)
Amphianthus aff. lacteus (McMurrich, 1893)
Isosicyonis alba (Studer, 1879)
Isotealia antarctica Carlgren, 1899
Antholoba achates Dana, 1849
Orden Scleractinia
Fungiacyathus marenzelleri (Vaughan, 1906)
Desmophyllum cristagalli Milne Edwards y Haime, 1848
Bathelia candida Moseley, 1881
Flabellum thouarsii Milne Edwards y Haime, 1849
Flabellum areum Cairns, 1982
Flabellum curvatum Moseley, 1881
Javania cailleti (Duchassaing y Michelotti, 1864)

190 Revista Real Academia Galega de Ciencias. Vol. XXVII
Distribución latitudinal de cnidarios bentónicos de aguas frías.
Se han determinado 41 especies de cnidarios pertenecientes a los siguientes taxa:
Leptothecatae: 14 especiesStylasterina: 5 especiesGorgonacea: 5 especiesPennatulacea: 1 especieAlcyonacea: 2 especiesActiniaria: 7 especiesScleractinia: 7 especies
La distribución de las mismas puede observarse en las Figs. 3-8.
Del análisis de las cartas de distribución pareciera indicar que en general hay mayor concentración de especies en la subregión Patagónica o Magallánica, especialmente para los taxa Leptothecatae, Stylasterina, Actiniaria y Scleractinia.
Esta mayor concentración podría ser explicada zoogeograficamente como que la subregión Patagónica podría ser considerada el centro de dispersión de las especies estudiadas. A través de la Corriente Circumpolar Antártica y su originaria Corriente de Malvinas, se distribuyen dichas especies a lo largo de la plataforma continental hasta los 30°LS (Fig. 7) y 35° LS (Fig. 8) respectivamente.
Asi como la subregión Patagónica es considerada centro de dispersión, también constituye el genocentro del género Renilla (Pennatulacea) (Zamponi, 2001) y de la especie Bolocera tuediae subs. occidua (Actiniaria) (Riemann-Zurnek, 1980). Esta subespecie tiene su especie vicariante en Bolocera tuediae subs tuediae de distribución en el océano Atlántico Norte.
Es posible que las características oceanográficas de la región sean favorables para el desarrollo y poblamiento de especies que se hallan formadas por un sistema esqueletal calcáreo y de espículas y/o escleritas. La formación de estos tipos de esqueletos requieren de un habitat más o menos estable y lo menos vulnerable posible.
La relación entre este tipo de ambiente y la formación de un sistema esqueletal rígido es tratado en la sección siguiente del artículo.

191Revista Real Academia Galega de Ciencias. Vol. XXVII
Figura 3. Distribución de especies de Leptothecatae.

192 Revista Real Academia Galega de Ciencias. Vol. XXVII
Figura 4. Distribución de especies de Stylasterina.

193Revista Real Academia Galega de Ciencias. Vol. XXVII
Figura 5. Distribución de especies de Gorgonacea.

194 Revista Real Academia Galega de Ciencias. Vol. XXVII
Figura 6. Distribución de especies de Pennatulacea y Alcyonacea.
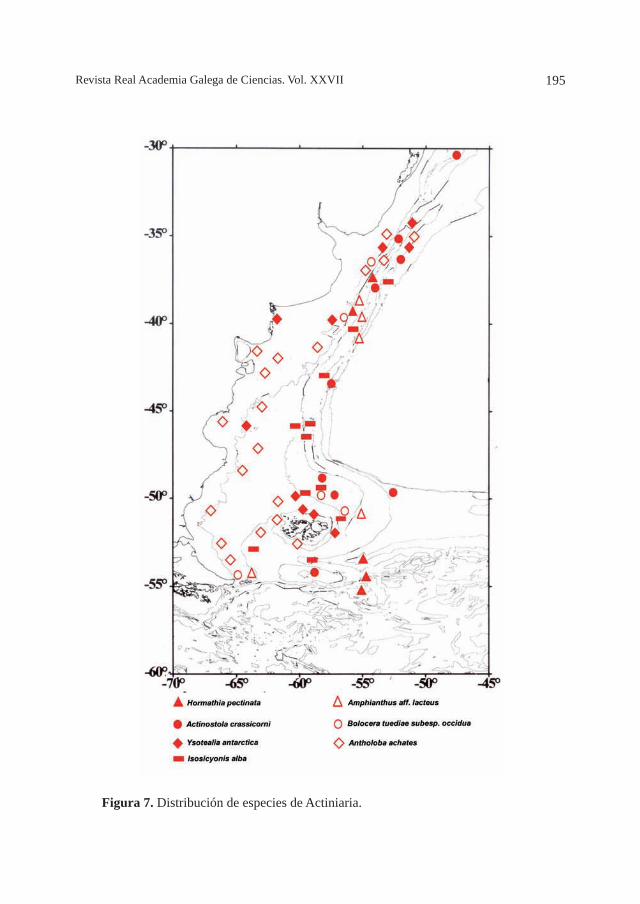
195Revista Real Academia Galega de Ciencias. Vol. XXVII
Figura 7. Distribución de especies de Actiniaria.

196 Revista Real Academia Galega de Ciencias. Vol. XXVII
Figura 8. Distribución de especies de Scleractinia.

197Revista Real Academia Galega de Ciencias. Vol. XXVII
Estabilidad Ambiental y Formación Esqueletal
Los taxa mencionados merecen ser analizados desde una perspectiva morfológica y ecológica, ya que, los mismos incluyen:
1. especies que carecen de un sistema esqueletal rígido.
2. especies con sistema esqueletal calcáreo. (Figs. 9 y 10)
3. especies con sistema esqueletal de espículas y/o escleritas. (Fig. 11)
El primer grupo pertenece a los taxa Leptothecatae y Actiniaria, mientras que al segundo conjunto pertenece los órdenes Stylasterina y Scleractinia y al último grupo son los Gorgonacea, Pennatulacea y Alcyonacea.
Aquellas especies con sistema esqueletal calcáreo y con sistema esqueletal espicular requieren de un habitat generalmente estable y lo menos perturbable posible, ya que los procesos de formación de esqueleto calcáreo como espicular exigen de cierto equilibrio ambiental.
El proceso de calcificación es complejo y comienza en el extremo basal del pólipo, donde la epidermis está constituida por un epitelio escamoso, profundo y lateralmente interdigitado. El organismo captura los iones Ca+ del medio externo para depositarlo extracelularmente y formar los septos y travéculas, mientras que los iones CO3= también tomados del exterior se depositan en el interior del citoplasma celular. Si el ambiente es estable y en equilibrio los iones Ca+ se depositan bajo la forma de aragonita, pero si el ambiente presenta alteraciones producidas por causas diversas, se interrumpe la deposición de Ca+ bajo la forma aragonita, y los mismos se disponen irregularmente formando un esqueleto débil que se rompe o quiebra por la acción de corrientes y olas, originando en el tiempo organismos débiles que traerá como consecuencia la extinción de la especie.
En las especies con sistema esqueletal espicular, el proceso de formación de las espículas es menos exigente que el de la calcificación, no obstante también aquí se requiere de cierta estabilidad ambiental. Las espículas que son intracelulares se originan a partir de células especiales denominadas escleroblastos; éstas se sitúan en la base de las células ectodérmicas y pasan a la mesogloea donde aumentan de volumen y el citoplasma se llena de

198 Revista Real Academia Galega de Ciencias. Vol. XXVII
granulaciones calcáreas amorfas que se agrupan en una concreción central que se van engrosando poco a poco hasta tener el tamaño de un núcleo. La concreción central va adquiriendo forma irregular y el centro se separa en dos mitades o núcleos mediante un eje protoplasmático. La espícula ahora está formada y continúa la acumulación de calcita y poco a poco la cobertura de protoplasma se estira y el esclerito termina totalmente recubierto de una capa citoplasmática ligeramente más importante en la vecindad de los núcleos. Las espículas se hallan formadas por dos componentes esenciales, uno orgánico y el otro mineral, internamente relacionados entre ellos; en este complejo, el elemento mineral está constituido por una mezcla de microcristales de calcita asociados a una trama orgánica.
Los dos procesos son mecanismos lentos que toman tiempo de realización, con lo cual se debe tratar que los ambientes donde se desarrollan estas especies sean protegidos y tenidos en cuenta en las políticas ambientalistas, para asegurar que dichas especies se perpetúen en el tiempo y constituyan patrimonios faunísticos de alto valor biotecnológico.
Figura 9. Especies de Stylasterina con sistema esqueletal calcáreo.1.Sporadopora dichotoma.(x 0.84); 2. Errina echinata. (x 0.38); 3. E. labiata. (x 0.90) 4.; E. lowei. (x 1.84); 5. Stylaster densicaulis. (x 0.63). (según Cairns, 1983).

199Revista Real Academia Galega de Ciencias. Vol. XXVII
Figura10. Especies de Scleractinia con sistema esqueletal calcáreo.1. Bathelia candida. (x 1.2); 2. Flabellum thouarsii. (x 24.9); 3. F. areum.(x 25.8); 4. F. curvatum. (x 40.7); 5. Javania cailleti. (x 10.2); 6. Desmophyllum cristagalli. (x 66.4). (según Cairns, 1982).
Figura 11. Especies de Alcyonacea, Gorgonacea y Pennatulacea con sistema esqueletal de espículas y/o escleritas.1. Alcyonium paessleri.( tamaño natural); 2. A. patagonicum. (tamaño natural); 3. Thouarella acanthina. (tamaño natural); 4. Convexella magelhaenica (tamaño natural);. 5. Primnoella biserialis. (tamaño natural); 6. P. compressa. (tamaño natural); 7. Renilla reniformes. (tamaño natural).

200 Revista Real Academia Galega de Ciencias. Vol. XXVII
CONCLUSION
Las zonas de alta productividad biológica están generalmente asociadas a sistemas frontales, que en zonas someras con mareas de gran amplitud hace que la fricción con el fondo genere frentes, que separan la región estratificada más rica en nutrientes, de la verticalmente homogénea; en consecuencia la existencia de sistemas frontales en las proximidades de la península de Valdés, en el golfo San Jorge, a lo largo de casi toda la costa comprendida entre 50°S y la Isla de los Estados, alrededor de las Islas Malvinas y el Banco Burdwood, estaría explicando de la alta concentración de especies de cnidarios hallados en dichas zonas que tendría relación directa con el rol de organismos suspensivoros (filtradores y carnívoros) (Zamponi, 2006) que tienen los cnidarios en los ecosistemas.
White y Petersen (1996) han demostrado que perturbaciones de la presión atmosférica, el viento, la cobertura del hielo marino austral y la temperatura superficial del océano se propagan con la Corriente Circumpolar Antártica y afectaría la totalidad de la plataforma continental. Este conjunto de factores influirían a lo largo del año, sobre la mayor o menor concentración de especies lo cual hace suponer que aquellas especies que poseen un esqueleto rígido se concentren en zonas donde los factores mencionados tengan una influencia menor, para de esta manera el ambiente sea lo más propicio posible para el asentamiento y desarrollo de la especie.
El asentamiento de las diferentes poblaciones también estaría favorecido por los fondos duros, que son afloramientos de material litificado con diferente grado de consolidación. Los afloramientos no ocupan más del 2% de la superficie de la plataforma y se distribuyen dispersamente uniendo el borde oriental de la subregión Argentina o Bonaerense con el extremo nororiental de la subregión Patagónica o Magallánica. Dichos afloramientos constituyen elevaciones sobre la plataforma ubicados a 85-100 m de profundidad; otros afloramiento pero aislados se encuentran en la inmediaciones de Islas de los Estados e Islas Malvinas.
Los afloramientos indicados y las corrientes oceánicas, tienen gran influencia sobre las estrategias de alimentación de estos organismos suspensivoros que favorecidos por los sistemas frontales crean zonas de alta productividad. El resultado de estos fenómenos permite un movimiento continuo de aguas profundas que puede contribuir al proceso de resuspensión,

201Revista Real Academia Galega de Ciencias. Vol. XXVII
que es esencial para el desarrollo y posterior asentamiento de comunidades bentónicas como lo constituyen este grupo de cnidarios.
La conjunción de factores oceanográficos y topográficos sumados a la riqueza de nutrientes y oxigenación de la Corriente de Malvinas, hace que la mayor concentración de especies se establezca en la subregión Templada Fría (= subregión Patagónica o Magallánica), ya que es la zona de influencia de esta corriente. Dicha corriente al continuar su recorrido, sigue aportando riqueza en nutrientes y abundante oxigenación constituyendo una vía de dispersión para las especies, haciendo que muchas de ellas puedan distribuirse hasta los límites norte de la subregión Templada Cálida (= subregión Argentina o Bonaerense).
AGRADECIMIENTOS
El autor agradece a la Sra. Graciela Testa (CONICET) la asistencia técnica en la compaginación del texto y las ilustraciones, sin la cual hubiese sido difícil ordenar. El trabajo ha sido realizado a través del PIP n 5071 (CONICET) y EXA n 362/06 (UNMdP).

202 Revista Real Academia Galega de Ciencias. Vol. XXVII
BIBLIOGRAFÍA
BASTIDA, R., ROUX, A., MARTINEZ, D. (1992) Benthic communities of the Argentine continental shelf. Oceanol. Acta, 15: 687-698.BASTIDA, R., ZAMPONI, M.O., BREMEC, C., ROUXA,A., GENZANO, G., ELÍAS, Y.R. (2007) Las comunidades bentónicas. J. I. Carreto y C. Bremec (eds.). E. E. Boschi (ed. general). En: El Mar Argentino y sus Recursos Pesqueros. T. 5. El Ecosistema Marino. Instituto Nacional de Investigación y Desarrollo Pesquero. Mar del Plata. pp. 91-125.
BLANCO, O. M., ZAMPONI, M. O., GENZANO, G. N. (1994) Lafoeidae de la Argentina (Coelenterata, Hydrozoa, Hydroida). Naturalia patagónica, Ciencias Biológicas, 2: 1 – 31.
BLANCO, O. M., ZAMPONI, M. O., GENZANO, G. N. (2000a) Campanulinidae de la Argentina (Coelenterata, Hydrozoa, Hydroida). Rev. Mus. La Plata. (Nueva Serie), XIV ( 163, 164, 165): 267 – 278.
BLANCO, O. M., ZAMPONI, M. O., GENZANO, G. N. (2000b) Syntheciidae de la Argentina (Coelenterata, Hydrozoa, Hydroida). Rev. Mus. La Plata. (Nueva Serie), XIV (163, 164, 165): 285– 295.
BLANCO, O. M., ZAMPONI, M. O., GENZANO, G. N. (2003) Sertulariidae de la Argentina (Coelenterata, Hydrozoa). www.labic.8m.com 53 p.
BOLTOVSKOY, E. (1970) Masas de Agua (característica, distribución, movimientos) en la superficie del Atlántico Sudoccidental, según indicadores biológicos. Ser. Hidrog. Naval, Buenos Aires, H-643, 99 pp.
BOLTOVSKOY, E. (1981) Masas de agua en el Atlántico Sudoccidental. D. Boltvskoy (ed.). En: Atlas del Zooplancton del Atlántico Sudoccidental. Instituto Nacional de Investigación y Desarrollo Pesquero. Mar del Plata. pp. 227-237.
BOSCHI, E. E. (1976) Nuevos aportes al conocimiento de la distribución geográfica de los crustáceos decápodos del Mar Argentino. Physis (Buenos Aires), Secc A., 35: 59-68.
BRANDHORST, W., CASTELLO, J.P. (1971) Evaluación de los recursos de anchoita (Engraulis anchoita) frente a la Argentina y Uruguay. I. Las condiciones oceanográficas, sinopsis del conocimiento actual sobre la anchoita y el plan para su evaluación. Proyecto Desarrollo Pesquero. FAO. Publicación n° 29, 63 pp.
CAIRNS, S. D. (1982). Antaqrctic and Subantarctic Scleractinia. Biology of the Antarctic Seas, XI, 34: 1-74.
CAIRNS, S. D. (1983) Antarctic and Subantarctic Stylasterina (Coelenterata: Hydrozoa). Biology of the Antarctic Seas, XIII, 38 (2): 61 – 164.
EKMAN, S. (1953) Zoogeography of the Sea. Sidgwick and Jackson Limited. 422 pp. London.
KRAMP, P. L. (1959) The Hydromedusae of the Atlantic Ocean and adjacent watyers. Dana Report n° 46.
PEREZ, C. D. (1999) Taxonomía, distribución y diversidad de los Pennatulacea, Gorgonacea y Alcyonacea del Mar Epicontinental Argentino y zonas de influencia. Tésis Doctoral. UNMdP. ii + xii;254 p.
PIOLA, A. R., RIVAS, A.L. (1997) Corrientes en la plataforma continental. E. E. Boschi (ed.). En: El Mar Argentino y sus Recursos Pesqueros. T. 1. Instituto Nacional de Investigación y Desarrollo Pesquero. Mar del Plata. pp. 119-132.

203Revista Real Academia Galega de Ciencias. Vol. XXVII
RIEMANN-ZURNECK, K. (1973) Actiniaria des Sudwestatlantik. I. Hormathiidae. Helgolander wiss Meeresunters, 25: 273 – 325.
RIEMANN- ZURNECK, K. (1978) Actiniaria des Suvestatlantik. IV. Actinostola crassicornis (Hertwig, 1882) mit einer Diskussion verwandter Arten. Veroff. Inst. Meeresforsch. Bremerh. 17: 65–85.
RIEMANN-ZURNECK, K. (1980) Actiniaria des Sudwestatlantik. V. Bolocera, Isotealia, Isocyonis (Actiniidae). Mitt. Hamb. Zool. Mus. Inst., 77: 19–33.
RIEMANN-ZURNECK, K. (1986) Zur Biogeographie des Sudwestatlantik mit besonderer Berucksichtigung der Seeanemonen (Coelenterata: Actiniaria). Helgolander Meeresunters, 40: 91–149.
RIEMANN-ZURNECK, K. (1980) Actiniaria des Sudwestatlantik. V. Bolocera, Isotealia, Isosicyonis (Actiniidae). Mitt. hamb. Zool. Mus. Inst. Band. 77: 19-33.
WHITE, W. B., PETERSON, R.G. (1996) An antarctic circumpolar wave in surface pressure, wind, temperature and sea ice extent. Nature, 380: 699-702.
ZAMPONI, M. O. (2001) La distribución bioceánica y continua del género Renilla Lamarck, 1816 (Cnidaria: Pennatulacea) y su endemismo en el continente americano. Physis (Buenos Aires), Sec. A., 58 (134-135): 7-9.
ZAMPONI, M. O. (2006) Estrategias morfológicas adaptativas y de distribución en organismos suspensivoros pasivos: filtradores y carnívoros. Rev. Real Acad. Galega Cienc., 25: 59-74.


VIDA DA
REAL ACADEMIA GALEGA DE CIENCIAS

206 Revista Real Academia Galega de Ciencias. Vol. XXVII
MEMORIA DAS ACTIVIDADES DA REAL ACADEMIA GALEGA DE CIENCIAS
2008
INSTAURACIÓNDÍA DO CIENTÍFICO GALEGO
O Presidente dá Real Academia Galega de Ciencias (RAGC), Profesor Ernesto Vieitez, no último Pleno da RAGC do ano 2006, celebrado nas instalacións do Real Club do Mar de Aguete, Pontevedra, propón a instauración dunha homenaxe de recoñecemento a aqueles científicos que, ben nacidos en Galicia ou non, pero que realizasen a súa carreira científica en Galicia, contribúan ao desenvolvemento do coñecemento e da innovación de maneira notable e significativa. É dicir, propón a posibilidade de instaurar o Día do Científico Galego, como recoñecemento e homenaxe a todas as persoalidades relevantes que traballaron no campo da investigación científica. Entende que a sociedade galega está, dalgunha forma, en débeda cos científicos que, directa ou indirectamente contribuíron de forma relevante ao progreso da rexión.
Así como hai un día adicado ás letras galegas e a un escritor concreto, parece lóxico pensar que o mesmo poida suceder con aquelas persoas que dedicaron, en Galicia, a súa vida profesional ao engrandecimiento das ciencias experimentais.
O Pleno discute a iniciativa, que é acollida con gran satisfacción por todos os Académicos, propoñéndose desde a denominación da xornada ata diseñar unha lista de posibles candidatos.
O Pleno entende que, unha iniciativa desta envergadura, require unha planificación moi seria e precisa, polo menos nos seus primeiros anos, a fin de que poida ir calando na sociedade galega para que un día, esta poida asumir a celebración como algo propio. Non se entende unha iniciativa desta índole sen o apoio incondicional do Goberno Autonómico, autoridades académicas, autoridades provinciais e locais e incluso o sector industrial. La RAGC toma a iniciativa no evento e ofrécese para desevolver esta andaina. Pero non pretende ser o único actor patrimonial do evento xa que é consciente que a base do éxito da proposta é a colaboración de todos os estamentos involucrados.

207Revista Real Academia Galega de Ciencias. Vol. XXVII
O primeiro que se pretende conseguir é dar a coñecer ás achegas científicas dos nomeados, que avances na ciencia e na tecnoloxía achegaron e cales foron as repercusións dos seus descubrimentos. Coa instauración do Día do Científico Galego preténdese, así mesmo, sensibilizar aos poderes públicos sobre a necesidade de apoiar de forma continua e ao maior nivel posible a investigación científica na rexión.
Poderase demostrar a través desta conmemoración que, ainda con poucos medios materiais, insignes científicos galegos fixeron achegas importantísimas non só no ámbito rexional senón tamén no nacional e internacional. A sociedade galega terá coñecemento dos logros dos seus científicos máis renomeados e poderá chegar a comprender que o investimento que se faga en ciencia dará uns froitos que beneficien a toda a sociedade. É evidente que, ao amparo dos exemplos dos mellores científicos, aparecen cada vez máis vocacións en novos que algún día poidan emular e superar, os éxitos dos seus predecesores.
Poden resumirse estes argumentos no feito de que o obxectivo final do Día do Científico Galego está en sensibilizar aos poderes públicos e á sociedade galega en xeral da importancia do apoio unánime á ciencia e á tecnoloxía, base para o desenvolvemento dunha sociedade avanzada e libre.
Dende o primeiro momento que se expuso a iniciativa no Pleno do 6 de Xullo de 2006, tívose claro que a data idónea era a do 23 de Abril, día na que se publicou no BOE a constitución oficial da RAGC (23 Abril 1977).
Houbo coincidencia no Pleno da RAGC da existencia de investigadores ilustres que poderían formar a lista de candidatos: desde o Padre Sarmiento e Isidro Parga Pondal ata Cruz Gallástegui ou Luis Iglesias. Con todo, pronto xurdiu o nome de D. Enrique Vidal Abascal como o candidato máis firme para ser considerado como o primeiro homenaxeado. Coinciden en D. Enrique Vidal dous feitos incuestionables, ademais dos seus propios méritos científicos: cumprirse en 2008 o centenario do seu nacemento (12 de Outubro de 1908), e ser un dos fundadores e primeiro Presidente da Real Academia Galega de Ciencias.
Ernesto Viéitez CortizoPresidente

208 Revista Real Academia Galega de Ciencias. Vol. XXVII
Prof. Dr. Enrique Vidal Abascal con orbígrafo deseñado por él para o cálculo de órbitas planetarias.
ENRIqUE VIDAL ABASCAL: O FACTOR HUMANO E A INTELIXENCIA COLECTIVA.
SEMBLANZA, POR FÉLIX VIDAL COSTA
Excelentísima Sra. Conselleira de Educación e Señor Director Xeral, Excelentísimo Señor Presidente da Academia Galega de Ciencias e membros da Academia, Excelentísimo Sr. Presidente do Consello da Cultura Galega, Magnífico Señor Reitor da Universidade de Santiago, Magnífico Señor Reitor da Universidade da Coruña, Excelentísimo Señor ex-Presidente da Xunta de Galicia, Ilustrísimos Señores Decanos, en particular da Facultade de Matemáticas da Universidade de Santiago, Señor Director do Observatorio Astronómico da Universidade de Santiago, Ilustrísimo Señor Alcalde de Lalín, Señora representante da Alcaldía de Santiago de Compostela, outras autoridades e representantes doutras Institucións, benqueridos familiares e amigos, miñas donas, meus señores.
Permítanme que, en primeiro lugar, exprese ao Señor Presidente da Academia Galega de Ciencias e aos meus colegas académicos a nosa máis profunda gratitude, a miña e a dos meus familiares, por decidir dedicar a Primeira Edición do Día do Científico Galego ao noso pai, Enrique Vidal Abascal. También quisiera decir ya ahora el gran honor y satisfacción que es

209Revista Real Academia Galega de Ciencias. Vol. XXVII
para nosotros contar con la compañía de todos ustedes en este acto, muy especialmente con la compañía de nuestra querida amiga la profesora Doña Antonia Ferrín Moreiras, que fue una estrecha colaboradora de nuestro padre antes y durante el periodo de gestación de la Sección de Matemáticas de la Universidad de Santiago, de cuya creación se celebra precisamente este año su cincuentenario.
Mis colegas académicos me han hecho un gran honor al pedirme que haga en este acto una semblanza de mi padre. Pero al mismo tiempo me han puesto en una delicada situación, precisamente por la proximidad afectiva, que podría incidir en la siempre necesaria objetividad de cualquier acto académico. Sin embargo, fueron sensibles a mis aprensiones y aceptaron que mi intervención fuese corta, y que estuviese esencialmente centrada en lo que podríamos denominar “el factor humano” de sus actividades científicas. Así pues, no comentaré nada sobre sus contribuciones personales en varios aspectos de la Astronomía primero y de la Geometría Diferencial después, aspectos que abordarán posteriormente en este acto, naturalmente con mucha más autoridad que yo, dos de sus más brillantes discípulos, los profesores Luís Cordero y Manuel de León. Otros aspectos acaban de ser evocados, con mucho afecto y pertinencia, por la Señora Conselleira de Educación, la profesora Laura Sánchez Piñón.
La familia de mi padre tenía una larga tradición de servidores del Estado: su padre fue, en los años 20 del siglo pasado, Director General de Hacienda siendo ministro Don José Calvo Sotelo, y fue posteriormente Presidente del Consejo de Cuentas del Estado. Pero antes había estado en la Delegación de Hacienda de Oviedo, lo que explica el lugar de nacimiento de mi padre, y había sido Delegado de Hacienda en Ourense y posteriormente en A Coruña, lo que a su vez explica que una parte de los estudios de bachillerato de mi padre los realizase en esa última ciudad. Su abuelo y bisabuelo paternos habían sido jueces en Galicia. Su abuelo materno, de Cantabria, fue también juez y magistrado.
Esta larga tradición familiar de servidores de lo público influyó probablemente para que mi padre se decantase por ser profesor de matemáticas en vez de arquitecto, como dudaba al empezar a estudiar “Ciencias Exactas” en Madrid (los dos primeros años de esa carrera siendo muy comunes a las dos opciones, y el dibujo y la pintura siendo su otra gran vocación). Fue su bisabuelo paterno el que hizo construir a principios del siglo XIX la que mi familia llamaba “la casa de Lalín”, que colindaría con la que posteriormente construiría la familia Aller. Esa cercanía sería uno de los orígenes de la gran amistad entre las dos familias, en particular la de mi abuelo primero y después la de mi padre con el gran astrónomo Don Ramón María Aller Ulloa, amistad que tendría tan profundas consecuencias en la vocación por las matemáticas de mi padre y en una buena parte de su vida profesional. Además de santiagués, mi padre siempre se sintió lalinense: Fue allí donde se inscribiría en su juventud en el Partido Galleguista, y más tarde sería nombrado hijo adoptivo de Lalín, en 1984, siendo su alcalde Don José Cuiña Crespo.
También se sintió, desde su regreso a Santiago de Compostela en 1941, ya como Catedrático de Instituto y profesor de la Universidad, fuertemente comprometido con esa ciudad y con su Universidad, compromisos que serían ampliamente reconocidos siendo, en 1995, descubierta por el alcalde Gerardo Estévez una placa conmemorativa en su casa de la

210 Revista Real Academia Galega de Ciencias. Vol. XXVII
calle de la Rosa. Unos meses más tarde, el Ayuntamiento de Santiago puso su nombre a una calle de la ciudad. Ese mismo año se celebró en la Universidad de Santiago de Compostela un solemne “Acto Académico In Memoriam do Profesor Enrique Vidal Abascal”, en el cual hicieron sendas semblanzas, con gran afecto y conocimiento de causa, los Profesores D. Xosé Masa Vázquez, del Departamento de Geometría y Topología de la Universidad de Santiago, D. Andres Lichnerowicz, del College de France y el entonces Rector de la Universidad de Santiago, Profesor D. Darío Villanueva Prieto.
Dos documentos esenciales para entender su profundo compromiso con Galicia son su Discurso de Ingreso en la Real Academia Galega das Letras, en 1971, y el Discurso Inaugural, leído en la Solemne Apertura del Curso Académico 1973-74 de la Universidad de Santiago de Compostela, compromiso que sería reconocido por la concesión de la Medalla Castelao de 1989 (creo que entonces se concedía una sola medalla anual), siendo Presidente de la Xunta de Galicia Don Gerardo Fernández Albor, “a toda unha vida dedicada a Galicia”. Anteriormente había sido nombrado, en 1974, “Officier dans l’Ordre des Palmes Académiques” por el Gobierno Francés, en reconocimiento de su compromiso con la cultura y la ciencia de ese país. Esos compromisos han tenido una gran influencia en sus planteamientos y actividades científicas. Como ya he indicado antes, tales actividades así como las numerosas distinciones y premios que le fueron concedidos por ellas, serán evocadas posteriormente por los Profesores Cordero y de León.
Otra experiencia sin duda decisiva para la formación intelectual de mi padre fue su estancia en Ginebra en su juventud, en el año 1934, en el Instituto de Investigación que dirigía Jean Piaget, que en aquel momento era el más importante e influyente estudioso de la fenomenología del aprendizaje. Pienso que esa experiencia suiza, que años más tarde complementaría con su estancia en Lausanne, en el Departamento del celebre matemático George de Rahm, contribuyó a conformar dos de los rasgos esenciales de su personalidad como profesor y como matemático: El primero, de carácter pedagógico, se puede resumir diciendo que tanto en sus clases y seminarios como en sus libros y publicaciones, siempre se proponía explicar con claridad y sencillez incluso las ideas y los resultados matemáticos más complejos: Siempre insistía en que ser ininteligible no es sinónimo, ni mucho menos, de ser profundo.
Otro aspecto de su personalidad científica en el que también influyeron sus tempranas experiencias suizas, y que creo que es crucial para entender una buena parte de sus planteamientos profesionales, es el que él mismo denominó “inteligencia colectiva”. Para aclarar a lo que se refería, me voy a permitir reproducir un párrafo de la semblanza que en el año 1962 le solicitó la Revista Matemática Hispano Americana sobre el matemático español Julio Rey Pastor, fallecido ese mismo año. Pero antes decir brevemente que Rey Pastor ha sido sin duda el más influyente matemático español de la primera mitad del siglo XX, cuyas inteligencia y brillantez mi padre las comparaba a las de Don José Ortega y Gasset. Pero como el mismo Rey Pastor reconocería en su discurso de entrada en la Academia de Ciencias de Madrid en 1956, gran parte de sus trabajos (los de Rey Pastor) y de los de sus discípulos estaban, decía literalmente Don Julio, “fuera de cauce”.

211Revista Real Academia Galega de Ciencias. Vol. XXVII
Pues bien, en su semblanza de Rey Pastor mi padre escribía: “Ese fuera de cauce, hay que decirlo con dura sinceridad para hacer honor al legado del egregio maestro, desfasó a la matemática española por otros cincuenta años. Todavía pesan hoy (les recuerdo que mi padre está escribiendo eso en el año 1962) como un farragoso lastre en algunos de nuestros más significados matemáticos esos años, aún recientes, durante los que una de las asignaturas más importantes de la carrera era la Geometría descriptiva, y los métodos gráficos y sintéticos eran estudiados en España con el ardor que nuestros antepasados ponían, por ejemplo, en atravesar los Andes. No es pues ánimo ni deseo lo que nos falta, ni siquiera inteligencia individual, ni insignes maestros, pero sí, en cambio, eso que pudiéramos denominar “inteligencia colectiva”, que acierte a ponernos en la ruta fecunda y a la que solo se llega por un meditado plan discutido conjuntamente. Para las empresas de uno solo basta con que uno acierte. Para las empresas colectivas es preciso buscar la manera de sumar los saberes y las voluntades. En la labor científica, como en muchas otras labores, se aplica la bien conocida regla, la recordaba Jean Cocteau hace poco en un congreso en Niza, que dice que uno y uno son once”.
La promoción en Galicia de la “inteligencia colectiva” fue sin duda la principal motivación profesional de mi padre, mucho mayor que cualquier ambición científica de carácter personal. Este planteamiento fue el que motivó sus afanes para conseguir que se reconociese en España y en la propia Universidad de Santiago la enorme valía de su maestro, el gran astrónomo Don Ramón María Aller Ulloa y para que se le construyese en el campus de nuestra Universidad un modesto observatorio astronómico.
Fue también ese planteamiento el que le llevó a promover primero, y a conseguir después poner en marcha, a partir de octubre de 1957, la Sección de Matemáticas de la Universidad de Santiago, arranque para el que contó en ese primer curso crucial con la ayuda del propio Don Ramón Aller (que ya tenía entonces 79 años, pero al ser Catedrático Extraordinario no se jubilaba…) y con la de la profesora Antonia Ferrín. Esa Sección de Matemáticas, a la que se incorporarían años más tarde otros profesores, era la cuarta creada en España, después de las ya tradicionales de Madrid, Barcelona y Zaragoza.
Es evidente que incluso sin ese afán de mi padre por promover la “inteligencia colectiva” en Galicia, se habría terminado por crear una Facultad de Matemáticas en la Universidad de Santiago. Pero en ese caso, este año probablemente se estaría celebrando el 25 aniversario de su creación, como ha sido recientemente el caso de la Facultad de Física, en vez del 50 aniversario. Y Galicia, con la que mi padre se sentía tan comprometido, se quedaría una vez más en el furgón de cola. Como solía decir el gran físico Niels Bohr, “es muy difícil hacer predicciones, sobre todo del futuro”, pero quizás sin ese adelanto de 25 años, de una generación, hoy no se hablaría de hacer en Santiago una de las sedes, incluso la principal, del Centro Estatal de Matemáticas. Además, se trataba de la primera creación, desde hacía muchos años, de una nueva Sección (antecedente de una nueva Facultad) en la Universidad de Santiago, y su incidencia en algunos planteamientos “conceptuales” del conjunto de la Universidad fue probablemente apreciable.
En cualquier caso, y esto ya no es una especulación, una vez creada la Sección de Matemáticas, y siguiendo con su afán de promover la “inteligencia colectiva”, mi padre impulsó

212 Revista Real Academia Galega de Ciencias. Vol. XXVII
una potente, y en aquel entonces pionera, política de relaciones con los más importantes centros internacionales de matemáticas, logrando que visitasen Santiago no pocos de los más prestigiosos matemáticos de aquellos años, enviando a sus discípulos a hacer largas estancias en los mejores Institutos de Investigación y en las mejores Universidades, y organizando, en 1963, lo que sería el primer Congreso Internacional de Matemáticas celebrado en España.
Todo ello, y algo más (un importante “algo más”), hizo que la Sección de Matemáticas de la Universidad de Santiago en general, y el Departamento de Geometría y Topología muy en particular, fuese uno de los protagonistas de lo que, por contraste con el “fuera de cauce” que denunciaba en 1956 Don Julio Rey Pastor, podríamos denominar el “encauzamiento” de la matemática española, a partir de los años 60 y 70 de siglo pasado. Y ese “algo más” es en realidad otro crucial “factor humano”, en palabras de mi padre “la inteligencia, la vocación y el esfuerzo del grupo de jóvenes licenciados (ahora ya no tan jóvenes...., este comentario es mio) que hicieron sus tesis de doctorado en Matemáticas en Santiago durante aquellos años”. El éxito de la joven Sección de Matemáticas, y especialmente el éxito de sus discípulos era para mi padre la mejor constatación de su acierto como “sembrador de inteligencia colectiva” y la mejor recompensa a sus esfuerzos. En su último artículo periodístico, escrito a los 85 años, todavía decía, con la generosidad del verdadero maestro, “e un orgullo pra min verme superado polos que foron meus discípulos. Ese é o mais grande honor para un profesor”.
Sei que me estou alongando máis aló do tempo que eu mesmo me impuxera, pero non quixese rematar esta semblanza sen referirme brevemente á propia Real Academia Galega de Ciencias, organizadora deste acto, a pesar de que xa o fixo con moita máis autoridade o seu Presidente, o Profesor Ernesto Viéitez. Non me resisto a nomear ás seis persoas, naquel entón todos membros da Real Academia Galega das Letras, que o 26 de xaneiro de 1973 presentaron a solicitude de constitución da Academia Galega de Ciencias a o Ministerio de Educación e Ciencia. A copia da solicitude deuma, así como unha boa parte dos documentos que consultei para preparar esta semblanza, o meu irmán Enrique. Citándoos na mesma orde coa que as súas sinaturas aparecen no documento de solicitude, eran: Enrique Vidal Abascal, Isidro Parga Pondal, Luis Iglesias Iglesias, Domingo García Sabel, Antonio Fraguas y Fraguas e Eugenio Torre Enciso. Creo que a motivación central do meu pai para promover e participar, de xeito particularmente activo, nesa aventura intelectual era, unha vez máis, a promoción da “intelixencia colectiva” en Galicia. Esta vez mediante a creación dunha institución que xa existía noutras partes de España (nesos anos a Academia de Ciencias de Madrid era, como decía con certa ironía meu pai, “casi municipal”), que complementaba a outras institucións científicas galegas, moi especialmente a Universidade. Como cada vez que recibía algunha distinción, seguramente o meu pai hoxe tamén diría “sentirse abrumado” pola vosa xenerosa decisión, benqueridos Presidente e colegas académicos, de que lle dedicaseis a primeira edición do “Día do Científico Galego”. E tamén se sentiría orgulloso, benquerido Presidente, da evolución desta Academia e estaría esperanzado de que entre todos, académicos, institucións de tutela, Universidades e centros de investigación, seremos capaces de facela aínda máis útil para o progreso de Galicia, para o cal será cada vez máis crucial o progreso da “intelixencia colectiva” galega.

213Revista Real Academia Galega de Ciencias. Vol. XXVII
Y termino con una cita que pretende resumir otros aspectos de la personalidad de mi padre, y no solo de su “factor humano” profesional, aspectos que obviamente no he podido evocar aquí. La cita es del Narciss y Goldmund de Hermann Hesse y dice: “¡Crear sin tener que pagar por ello el precio de vivir! ¡Vivir sin tener que renunciar a la nobleza de crear!” Pienso que mi padre fue en buena medida capaz, sin duda gracias a la ayuda de nuestra madre y casi siempre disfrutando con todas las tareas que se proponía, de vivir y de crear, de ser realista pero al mismo tiempo pionero y visionario, de ser buena persona y luchador por la libertad y contra la injusticia y el sufrimiento humano, y de estar comprometido con su familia, con sus amigos, con sus alumnos y discípulos, con su Universidad y con su país. Y por todo ello, y quizás ese ha sido su mayor éxito, fue muy querido, como lo pone en evidencia la concesión de tan apreciada distinción y la presencia de todos ustedes en este acto.
Moitas grazas
Santiago de Compostela, 23 de Abril de 2008,Día do Científico Galego e
31 Aniversario da Creación da Real Academia Galega de Ciencias.

214 Revista Real Academia Galega de Ciencias. Vol. XXVII
Presidencia do Día do Científico Galego;, o Prof. Dr. Senén Barro Ameneiro, Reitor da USC, Dona Laura Sánchez Piñón, Conselleira de Educación e Ordenación Universitaria o Prof. Dr. Ernesto Viéitez Cortizo, Presidente da RAGC, o Prof. Dr. Ramón Villares Paz, Presidente do Consello da Cultura Galega e o Prof. D. José María Barja Pérez, Reitor da UDC.
DÍA DO CIENTÍFICO GALEGO, 2008
No ano 2008, instaurose o Día do Científico Galego. Con esta celebración preténdese facer unha homenaxe de recoñecemento a aqueles científicos que, bien nados en Galicia ou non, desenrrolaron a súa carreira científica en Galicia, e que contribuíron ao desenvolvemento do coñecemento e da innovación de maneira notable e significativa.
Na súa primeira edición contouse coa presencia da Excma. Sra. Dña. Laura Sánchez Piñón, Conselleira de Educación e Ordenación Universitaria, co Presidente do Consello da Cultura Galega, Excmo. Sr. D. Ramón Villares Paz, e cos reitores das Univeresidades de Santiago de Compostela e da Coruña, os Magfcos. Sres., D. Senén Barro Ameneiro e D. José María Barja Pérez, así como con numerosas personalidades.
Adicado na súa primeira edición ao Prof. Dr. Enrique Vidal Abascal, con motivo do centenario do seu nacemento. A celebración contóu coas seguintes intervencións:
Apertura do acto por parte do Prof. Dr. Ernesto Viéitez Cortizo, Presidente da -Real Academia Galega de Ciencias.
Intervención da Conselleira de Educación e Ordenación Universitaria, a Excma. -Sra. Dona Laura Sánchez Piñón.

215Revista Real Academia Galega de Ciencias. Vol. XXVII
Semblanza da familia, a cargo dun fillo do homenaxeado, e Membro de Número -da Real Academia Galega de Ciencia, o Profesor Dr. Félix Vidal Costa, co título:“Enrique Vidal Abascal: o factor humano”.
Conferencia do Prof. Dr. Manuel de León Rodríguez, ex-alumno do homenaxeado e -Profesor de Investigación del Instituto de Matemáticas y Física Fundamental, CSIC en Madrid, baixo o título:“Enrique Vidal Abascal, el sembrador de geometría”.
Conferencia do Profesor Dr. Luís A. Cordero Rego, ex-alumno do homenaxeado, -Catedrático de Xeometría e Topoloxía da Universidade de Santiago de Compostela e Membro de Número da Real Academia Galega de Ciencias, co titulo:“Enrique Vidal Abascal, o meu mestre”.

216 Revista Real Academia Galega de Ciencias. Vol. XXVII
Presidencia da Apertura do Curso 2008.O CN. D. José Luís Urcelay Verdugo o Prof. Dr. Ernesto Viéitez Cortizo, o Prof. Dr. Franco Fernández González e o Prof. Inv. Dr. Antonio Ballester Álvarez-Pardiñas.
APERTURA DO CURSO ACADÉMICO
No mes de febreiro tivo lugar a apertura do curso do ano 2008. A lección maxistral correu a cargo do Prof. Dr. Tito A. Varela López, baixo o título “ADN móbil e secuencias Alu”
CONVENIOS
Este ano a Academia asinou os seguintes convenios de colaboración:
Coa Excma. Deputación Provincial Da Coruña, para o desenvolvemento dos XI •Avances en Ciencia e Tecnoloxía.
Coa Fundación Pedro Barrié de la Maza a Real Academia asinou un convenio de •colaboración destinado a publicacións.
Coa Fundación CaixaGalicia asinouse un convenio de colaboración para os •Premios de Investigación desta Real Academia, ós que concorren un gran número de investigadores tanto na modalidade de Traballo de Investigación coma na modalidade de Novos Investigadores, dotados con 15.000 e 9.000 euros respectivamente, e para o Premio a un Traballo publicado (o en vías de publicación) na Revista da Real Academia Galega de Ciencias, dotado con 3.000 euros.
Coa Consellería de Cultura e Deporte, en relación coa organización das XVII •Xornadas Luso – Galaicas de Ciencias e Desenvolvemento.

217Revista Real Academia Galega de Ciencias. Vol. XXVII
Coa Excma. Deputación de Lugo para a celebración do Curso de Conferencias de •Impactos Ambientais.
Co Colexio Oficial de Farmacéuticos da Provincia de Pontevedra para á •organización do ciclo de Conferencias Maxistrais sobre Avances en Ciencias Farmacéuticas.
PUBLICACIÓNS
No ano 2008 a Real Academia Galega de Ciencias editou a Revista, financiada por entidades públicas e privadas.
Publicouse o volume XXVI da Revista da Real Academia Galega de Ciencias, na •que se recollen traballos científicos dun amplo espectro de temas.
Xunto coa Revista, publicou o Volume XXVIa, volume especial adicado ao XXV •Aniversario da fundación da Facultade de Físicas da Universidade de Santiago de Compostela.

218 Revista Real Academia Galega de Ciencias. Vol. XXVII
CURSOS DE CONFERENCIAS
A Real Academia celebrou os seguintes cursos de conferencias:
XVII XORNADAS LUSO-GALAICAS DE CIENCIAS E DESENVOLVEMENTO
A Real Academia ven celebrando ininterrompidamente dende 1990 ás Xornadas Luso-Galaicas de Ciencias e Desenvolvemento, nas que se tratan moi diversos temas de actualidade.
Celebradas os días 21, 22 e 23 de Outubro, no Salón de Actos do Pazo de San Roque, en Santiago de Compostela, nestas xornadas contaron coa participación dos seguintes conferenciantes:
Día 21 de outubroMa • gfco e Excmo. Prof. Dr. Senén Barro Ameneiro. Reitor da Universidade de Santiago de Compostela“A responsabilidade social nas Universidades”
Prof. Dr. Nuno Filipe da Cruz Batista Mateus. Profesor do Dpto. de química •da Facultade de Ciências da Universidade do Porto“A química dos sabores do vinho”
Profa. Dra. Ilda da Conceiçâo Abreu de Noronha. Doutora do Departamento •de Botânica na Facultade de Ciencias da Universidade do Porto“O pólen de milho transgénico. Implicaçao no desequilibrio ecológico das colmeias”
Día 22 de outubroProf. Dr. Alexandre Lobo da Cunha. Profesor de Ciencias do Meio Aquático •da Universidade do Porto“O peroxissoma: a investigaçao sobre um organelo celular”
Prof. Dr. Helder M. Crespo. Profesor do Dpto. de Física da Universidade •do Porto“Lasers ultra-curtos estabilizados em fase absoluta: A luz controlada”
Prof. Dr. Félix Vidal Costa. Catedrático do Dpto. de Física da Materia •Condensada da Universidade de Santiago de Compostela. Membro de número da Real Academia Galega de Ciencias“Nanocousas versus cousas pequenas: algúns exemplos con superconductores”
Día 23 de outubroProf. Dr. Vicente Pérez Villar. Catedrático do Dpto. de Física da Materia •Condensada da Universidade de Santiago de Compostela. Membro de número da Real Academia Galega de Ciencias“Circulación abisal y cambio climático”

219Revista Real Academia Galega de Ciencias. Vol. XXVII
Clausura •Prof. Dr. Ramón Villares Paz. Presidente do Consello da Cultura“A Primeira República Portuguesa e a súa influencia en Galicia”
Nestas Xornadas contouse coa participación de un centenar de asistentes, maioritariamente universitarios, que seguiron atentamente ás exposicións dos conferenciantes, rematando con pequenos debates e preguntas.
XI AVANCES EN CIENCIA E TECNOLOXÍA BIOCOMBUSTIBLES: PRODUCION, USO E IMPACTO
Celebrados nos días 3, 4, 5, 6, 10, 11 e 12 do mes de novembro no Salón de Actos no Pazo de San Roque, en Santiago de Compostela, coa participación dos seguintes conferenciantes:
Día 3 de novembroProf. Dr. Miguel Ángel Ríos Fernández. Catedrático de química Física. •Profesor Emérito da Universidade de Santiago de Compostela. Académico Numerario da Real Academia Galega de Ciencias“A encrucillada enerxética. Un reto para a ciencia e a innovación tecnolóxica”
Día 4 de novembroDr. Luis Ortiz Torres. Catedrático de Enxeñería Agroforestal da Universidade •de Vigo. Director da Escola Universitaria de Enxeñería Técnica Forestal da Universidade de Vigo“O impacto dos biocombustibles sólidos: Un reto e unha oportunidade histórica para Galicia”
Día 5 de novembroD. Juan José Losada López. Jefe de Seguridad Industrial de Bioetanol •Galicia, S.A.“Presente e futuro da produción de Bioetanol. Estratexia do Grupo Abengoa”
Día 6 de novembroProf. Dr. Juan M. Lema Rodicio. Catedrático de Enxeñería química da •Universidade de Santiago de Compostela. Director da Escola Técnica Superior de Enxeñería da Universidade de Santiago de Compostela“Oportunidades e retos da codixestión anaerobia de residuos agroindustriais”
Día 10 de novembroDr. Gregorio Antolín Giraldo. Director da División químico-Alimentaria e •da Área de Biocombustibles da Fundación CARTIF“Revalorización de biomasa residual mediante procesos integrados de gasificación e extracción de compoñentes de alto valor agregado”

220 Revista Real Academia Galega de Ciencias. Vol. XXVII
Día 11 de novembroD • r. Manuel Bao Iglesias. Catedrático de Enxeñería química da Universidade de Santiago de Compostela e Académico Numerario da Real Academia Galega de Ciencias“Procesos productivos del Biodiesel: Presente e Futuro”
Día 12 de novembroClausura •D. Salustiano Mato de la Iglesia. Director Xeral de Investigación, Desenvolvemento e Innovación da Xunta de Galicia “A aposta galega en biocombustibles”
Este ciclo de conferencias ven celebrándose dende 1998, baixo o patrocinio da Deputación Provincial Da Coruña. Nesta nova edición o interés surxido por estas conferencias levou a que o aforo completárase ao cabo de dous días do inicio da fecha de inscripción.
XI Avances en Ciencia e Tecnología. O Excmo. Prof. Dr. Ernesto Viéitez Cortizo, Presidente da RAGC, acompañado de D. Salustiano Mato de la Iglesia, Director Xeral de I+D+i da Xunta de Galicia e do Prof. Dr. Tito A. Varela López, Membro Numerario da RAGC, durante a conferencia que impartiu o Director Xeral.

221Revista Real Academia Galega de Ciencias. Vol. XXVII
CURSO DE CONFERENCIAS DE IMPACTOS AMBIENTAIS: ENERXÍAS ALTERNATIVAS
Este ciclo de conferencias celebrouse os días 3, 4, 5, 10, 11, 12 e 13 de novembro no Salón de Actos da Excma. Deputación Provincial de Lugo, participando os seguintes conferenciantes:
Día 3 de novembroD. Juan Casares Long. • Catedrático de Enxeñería química da universidade de Santiago de Compostela
“A enerxía na sociedade tecnolóxica: cara un novo equilibrio”
Día 4 de novembroD. Francisco Dans del Valle. Director da Asociación Forestal de Galicia •“Transformación da biomasa en enerxía térmica e eléctrica: experiencias galegas”
Día 5 de novembroD. Antonio Rigueiro Rodríguez. Catedrático do Departamentode Producción •Vexetal da Universidade de Santiago de Compostela. Membro de Número da Real Academia Galega de Ciencias“Residuos e cultivos agroforestáis para produción enerxética”
Día 10 de novembroD. Manuel Bao Iglesias. Catedrático do Departamento de Enxeñería química •da Universidade de Santiago de Compostela. Membro de Número da Real Academia Galega de Ciencias“Biocombustibles”
Día 11 de novembroD. Ramón Velo Sabín • . Profesor Titular do Departamento de Enxeñería Agroforestal da Universidade de Santiago de Compostela“Enerxía solar en Galicia”
Día 12 de novembroD. J. Gregorio Iglesias Rodríguez • . Profesor do Departamento de Enxeñería Agroforestal da Universidade de Santiago de Compostela“Enerxía mareomotriz en Galicia”
Día 13 de novembro Clausura •Xerente de SOTAVENTO, S.A. “Sotavento, centro de referencia en Galicia en investigación e divulgación en Enerxías Renovables”
O ciclo de conferencias contou coa participación de máis de un centenar de asistentes, aumentando así o número de inscriciones de anos pasados.

222 Revista Real Academia Galega de Ciencias. Vol. XXVII
CICLO DE CONFERENCIAS MAGISTRALES SOBRE AVANCES EN CIENCIAS FARMACÉUTICAS
A Real Academia reanudou o Ciclo de Avances en Ciencias Farmacéuticas que viña organizando conxuntamente co Colexio Oficial de Farmacéuticos da Provincia de Pontevedra.
Celebráronse en Vigo os días 16 de xuño e 13, 16 e 30 outubro no Salón de Actos do Colexio Oficial de Farmacéuticos, e contaron coa participación dos seguintes conferenciantes:
Día 16 de xuñoDr. Antonio Fontdevila Vivanco. Catedrático de Xenética. Universidade •Autónoma de Barcelona“Medicina evolutiva: un enfoque darvinista da enfermidade”
Día 13 de outubroD. Santiago Cuellar Rodríguez. Director do Dpto. Técnico do Consello Xeral •de Colexios Oficiais de Farmacéuticos“Innovación científica dos novos medicamentos registrados en España”
Día 16 de outubroDr. Mª Isabel Loza García. Profa. Titular do Dpto. de Farmacoloxía. •Universidade de Santiago de Compostela“Farmacoxenética e mellores resultados da farmacoterapia”
Día 30 de outubroDr. Alfonso Domínguez-Gil Hurlé. Catedrático de Farmacia. Universidade de •Salamanca. Xefe de Servicio de Farmacia. Hospital Clínico de Salamanca“¿Son os medicamentos costo-efectivos?”
PREMIOS DE INVESTIGACIÓN, ANO 2007
No ano 2007, o Tribunal otorgou os seguintes premios:
Premio na modalidade de Traballos de Investigación, dotado con 15.000€: Título: “Determinación de metales en algas marinas destinadas a la alimentación humana producidas y procesadas en Galicia: desarrollo de metodología analítica automática.”Autor: Dona María del Carmen Yebra Biurrun. Universidade de Santiago de Compostela.
Premio Promoción de Novos Investigadores menores de 28 anos, dotado con 9.000€:Título: “Evaluación del efecto de la selenometionina sobre la reparación del daño en el ADN”Autor: Dona Vanessa Valdiglesias García. Universidade da Coruña

223Revista Real Academia Galega de Ciencias. Vol. XXVII
Premio a un Traballo publicado (o en vías de publicación) na Revista da Real Academia Galega de Ciencias, dotado con 3.000€:Titulo: “Respuesta de plantas de álamo a elevadas concentraciones de plomo”Autores: Don José Luís Couselo Bandín. Instituto de Investigacións Agrobiolóxicas de Galicia, CSIC.
Premios de Investigación 2007. Membros da Real Academia Galega de Ciencias e da Fundación Caixa Galicia, cos premiados.

224 Revista Real Academia Galega de Ciencias. Vol. XXVII
COMPOSICIÓN DA REAL ACADEMIA
XUNTA DE GOBERNO
Ernesto Viéitez Cortizo, PresidenteLuis Asorey García, VicepresidenteAntonio Ballester Álvarez-Pardiñas, SecretarioTito A. Varela López, TesoreroFrancisco Río Barja, BibliotecarioPresidentes das Seccións
SECCIÓNS
MATEMÁTICAS, FÍSICA E FÍSICA DO COSMOSLuis Cordero Rego, Gerardo Rodríguez López, Félix Vidal Costa, Vicente Pérez-Villar.
QUÍMICA E XEOLOXÍAFernando Fraga Rodríguez, Francisco Guitián Ojea, Manuel Bao Iglesias, Franco Fernández González, Miguel Angel Ríos Fernández, Antonio Ballester Álvarez-Pardiñas.
FARMACIA E BIOLOXÍAManuel Pereiro Miguens, Ernesto Viéitez Cortizo, Jesús Méndez Sánchez, Rafael Tojo Sierra, Luis Asorey García
CIENCIAS TÉCNICASOdón Abad Flores, Valeriano Yepes Hernández de Madrid, Ramón de Vicente Vázquez, Antonio Rigueiro Rodríguez.
CIENCIAS SOCIAIS E ECONÓMICASLuis Suárez-Llanos Gómez, Juan Quintáns Seoane, Xosé Manuel Beiras Torrado, Francisco Río Barja, Tito A. Varela López.
CONSELLO DE PUBLICACIÓNS
Ernesto Viéitez Cortizo, PresidenteAntonio Ballester Álvarez-Pardiñas, SecretarioFrancisco Río Barja, BibliotecarioLuis Cordero Rego, Vocal

225Revista Real Academia Galega de Ciencias. Vol. XXVII
ACADÉMICOS NUMERARIOS
Dr. Ing. D. Odón Abad FloresCantalarrana, Ctra. de Samoedo, s/n. 15160 SADA (A Coruña)
Dr. D. Luis Asorey GarcíaPlaza do Ferrol, 6-10 - 3º - 27001 LUGO
Prof. Dr. D. Manuel Bao IglesiasAvda. de las Ciencias, 4A - 1ºD - 15706 SANTIAGO DE COMPOSTELA
Prof. Dr. D. José Manuel Beiras TorradoPlaza de Mazarelos, 1 - 15703 SANTIAGO DE COMPOSTELA
Prof. Dr. D. Luis Cordero RegoRepública del Salvador, 19 - 1ºA - 15701 SANTIAGO DE COMPOSTELA
Excmo. Prof. Dr. D. Ramón de Vicente VázquezAvda. del Mar. Edificio San Bernardo, 21 - 6º Izqda. - 15406 FERROL
Prof. Dr. D. Franco Fernández GonzálezFray Rosendo Salvado, 24 - 2º A - 15701 SANTIAGO DE COMPOSTELA
Prof. Inv. Dr. D. Fernando Fraga RodríguezLa Coruña, 35 - 7º B - 36208 VIGO
Prof. Dr. D. Manuel Freire Ramac/ Monte dos Postes, 6 - 4º A - SANTIAGO DE COMPOSTELA
Prof. Dr. D. Francisco Guitián OjeaSenra, 16 - 15702 SANTIAGO DE COMPOSTELA
Prof. Inv. Dr. D. Jesús Méndez SánchezPza. Dr. Puente Castro, 5 - 6º Dcha. - 15702 SANTIAGO DE COMPOSTELA
Dr. D. Manuel Pereiro MiguensRepública del Salvador, 3 - 6º B - 15702 SANTIAGO DE COMPOSTELA
Prof. Dr. D. Vicente Pérez VillarAvda. de las Ciencias, 4 A - 2º B - 15706 SANTIAGO DE COMPOSTELA
Prof. Dr. D. Juan Quintás SeoaneAlcalá, 27 - 28014 MADRID
Prof. Dr. D. Antonio Rigueiro RodríguezAvda. Ramón Ferreiro, 32 - 7º B - 27002 LUGO
Prof. Dr. D. Francisco Río BarjaSto. Domingo de la Calzada, 2 - 6º C - 15701 SANTIAGO DE COMPOSTELA
Prof. Dr. D. Gerardo Rodríguez LópezFernando III El Santo, 33 - 3º B - 15702 SANTIAGO DE COMPOSTELA
Prof. Dr. D. Luis Suárez-Llanos GómezLa Rosa, 16 - 6º B - 15701 SANTIAGO DE COMPOSTELA
Prof. Dr. D. Rafael Tojo SierraOrense, 9 - 7º - 15701 SANTIAGO DE COMPOSTELA

226 Revista Real Academia Galega de Ciencias. Vol. XXVII
Prof. Dr. D. Tito A. Varela LópezFray Rosendo Salvado, 15 - Esc. 3, 7º L - 15701 SANTIAGO DE COMPOSTELA
Prof. Dr. D. Félix Vidal CostaBugallido, Buceleiras - 15886 AMES (A Coruña)
Excmo. Prof. Dr. D. Ernesto Viéitez CortizoGeneral Pardiñas, 28 - 2º Dcha. - 15701 SANTIAGO DE COMPOSTELA
Dr. Ing. D. Valeriano Yepes Hernández de MadridFonte do Espiño, 3 - 15719 Liáns - OLEIROS (A Coruña)
Prof. Dr. D. Miguel Angel Ríos FernándezFacultad de Química. Universidad de SantiagoSANTIAGO DE COMPOSTELA
Prof. Invest. Dr. D. Antonio Ballester Álvarez-PardiñasInstituto de Investigaciones Agrobiológicas de Galicia - CSICSANTIAGO DE COMPOSTELA
ACADÉMICOS DE HONRA
Excma. Sra. Dña. Carmela Arias y Díaz de Rábago. Condesa de Fenosa.Cantón Grande, 9. A Coruña.
Excmo. Prof. Dr. Federico Mayor Zaragoza.Mar Caribe, 15. Majadahonda - 28220 Madrid.
Excmo. Prof. Dr. Carl A. Leopold.1203 East Shore Drive. Ithaca, NY 14850. Estados Unidos.

227Revista Real Academia Galega de Ciencias. Vol. XXVII
ACADÉMICOS CORRESPONDENTES
Prof. Dr. Emil Den Tex. Geologische en Mineralogische. Institut der Rijksuniversiteit. Univ. Leiden. Holanda
Prof. Dr. Benito Sánchez Rodríguez. Conde de Gondomar, 7. Pontevedra.
Prof. Dr. Luis Castedo Expósito. Química Orgánica. Facultad de Química. Santiago.
Prof. Dra. Mª Pilar Fernández Otero. Depto. de Fisiología Animal. Universidad de Na-varra. Pamplona.
Prof. Dr. José L. Blanco González. Gran de Gracia, 7. 08012 Barcelona.
Prof. Dr. Jaime García Lombardero. Comisión de las Comunidades Europeas. Rue de la Loi 200. B1049, Bruselas. Bélgica.
Prof. Dr. Lieven Vanhecke. Department of Mathematis. Katholieke Universiteit Leuven, Departament of Mathematics, Celestijnenlaan 200 B, B-300. Leuven (Bélgica).
Prof. Dr. Jochen Kleinschmit. Niedersächsische Forstliche Versuchsanstalt. Abteilung Forstpflanzenzüchtung. NFV-Abt. C-W-3513 Staufenberg-Escheroche, Alemania.
Prof. Dr. Roberto Salema. Instituto de Biología Molecular y Celular. Rua do Campo Alegre, 823. 4100 Porto. Portugal.
Excmo. Prof. Dr. Santiago Grisolía. Fundación Valenciana de Estudios Avanzados. Pintor López, 7. 46003 Valencia.
Prof. Dr. Fernando Noronha. Centro de Geología da Universidade do Porto. Praça Gomez Teixeira. 4000 Porto. Portugal.
Prof. Dr. Gary J. Griffin. Departament of Plant Pathology, Physiology and Weed Science. Virginia Polytechnic Institute and State University. Blacksburg, Virginia 24061-0331. Estados Unidos.

228 Revista Real Academia Galega de Ciencias. Vol. XXVII
PUBLICACIÓNS DA REAL ACADEMIADISCURSOS DE RECEPCIÓN DE NOVOS ACADÉMICOSA determinación da arquitectura. Rafael Baltar Tojo, 1980.As ciencias matemáticas, esas descoñecidas. Luis A. Cordero Rego, 1982.Concepto de metodoloxía da antropoloxía biolóxica. Primeiras aplicacións na poboación galega. Tito A. Varela, 1982.Home e técnica. Odón Abad Flores, 1982.Ideas sobre la investigación agraria. Valeriano Yepes Hernández de Madrid, 1984.Evolución del crecimiento, maduración y desarrollo humano en Galicia, 1900-1980. Rafael Tojo Sierra, 1984.Ecuacións diferenciais e ciencia. Gerardo Rodríguez López, 1985.O home dentro do sabio: Pedro Joseph de Bermés (1770-1824). Valentín Paz Andrade, 1985.La historia de la micología médica. Manuel Pereiro Miguens, 1986.La industrialización de la sardina hace 200 años. La pesca en Galicia en la época de Cornide Saavedra. Francisco López Capont, 1986.Oceanografía de la plataforma gallega. Fernando Fraga Rodríguez, 1987.Cartografía Xurisdicional de Galicia no século XVIII. Provincia de Santiago. Francisco Río Barja, 1988.El matorral como recurso renovable. Manuel Bao Iglesias, 1989.Polímeros de alta estabilidad térmica. Santiago González-Babé Ozores, 1991.Sobre algúns aspectos experimentais das fluctuacións do parámetro de orde nos óxidos de cobre super conductores. Félix Vidal Costa, 1992.Historia y objetivos del diseño cuantitativo de fármacos. Franco Fernández González, 1993.Acto de ingreso del Excmo. Prof. Dr. D. Camilo José Cela en la Rea1 Academia Galega de Ciencias bajo la presidencia de S.A.R. el Príncipe de Asturias D. Felipe de Borbón y Grecia, 1996.Estructuras Dinámicas. Vicente Pérez Villar, 1998.Las timosinas α en la Biología de las células de mamíferos. Manuel Freire Rama, 1998.Génesis de la Ciencia Nuclear. La Radioactividad y el descubrimiento del Radio. Ramón de Vicente Vázquez, 2000.Antropología y transformismo. El pensamiento biológico y la posición crítica del P. Agustino Zacarías Martínez Núñez (†). Arzobispo que fue de Santiago de Compostela. Luis Asorey García, 2002.Bosques e paisaxe en Galicia. Antonio Rigueiro Rodríguez, 2002.Reflexiones sobre la Ciencia y la Técnica. Miguel Angel Ríos Fernández, 2004.La biotecnología vegetal aplicada a la producción forestal. Antonio Ballester Álvarez-Pardiñas, 2004.

229Revista Real Academia Galega de Ciencias. Vol. XXVII
OUTRAS PUBLICACIÓNS
Boletín da Academia Galega de Ciencias, volume I, 1982.Boletín da Academia Galega de Ciencias, volume II, 1983.Boletín da Academia Galega de Ciencias, volume III, 1984.Boletín da Academia Galega de Ciencias,volume IV, 1985.Boletín da Academia Galega de Ciencias, volume V, 1986.Boletín da Academia Galega de Ciencias, volume VI, 1987.Boletín da Academia Galega de Ciencias, volume VII, 1988.Boletín da Academia Galega de Ciencias, volume VIII, 1989.Boletín da Academia Galega de Ciencias, volume IX, 1990.Revista Academia Galega de Ciencias, volume X, 1991.Revista Academia Galega de Ciencias, volume XI, 1992.Revista Academia Galega de Ciencias, volume XII, 1993.Revista Real Academia Galega de Ciencias, volume XIII, 1994.Revista Real Academia Galega de Ciencias, volume XIV, 1995.Revista Real Academia Galega de Ciencias, volume XV, 1996.Revista Real Academia Galega de Ciencias, volume XVI, 1997.Reglamento de la Real Academia Galega de Ciencias (modificado R.D. 70/1998) (B.O.E. 18-2-1998).Guía de las babosas ibéricas. José Castillejo Murillo, 1998, 156 pp.Revista Real Academia Galega de Ciencias, volume XVII, 1998.El desarrollo industrial pesquero en el siglo XVIII. Los salazoneros catalanes llegan a Galicia. Francisco López Capont, 1999.Revista Real Academia Galega de Ciencias, volume XVIII, 1999.Revista Real Academia Galega de Ciencias, volume XIX, 2000.Revista Real Academia Galega de Ciencias, volume XX, 2001.Revista Real Academia Galega de Ciencias, volume XXI, 2002.Revista Real Academia Galega de Ciencias, volume XXII, 2003.Revista Real Academia Galega de Ciencias, volume XXIII, 2004.Revista Real Academia Galega de Ciencias, volume XXIV, 2005.Revista Real Academia Galega de Ciencias, volume XXV, 2006.Revista Real Academia Galega de Ciencias, volume XXVI, 2007.Revista Real Academia Galega de Ciencias, volume XXVII, 2008.

230 Revista Real Academia Galega de Ciencias. Vol. XXVII
CURSOS DE CONFERENCIAS PUBLICADOS
– “Homenaje a Cruz Gallástegui”, 1985, 191 pp.– “La lluvia ácida”, 1987, 60 pp.– “España y el Mercado Común”, 1988, 98 pp.– “Especies frondosas en la repoblación de Galicia”, 1989, 67 pp.– “Conferencias sobre el Río Miño”, 1989. l l2 pp.– “Homenaje a Valentín Paz Andrade”, 1991, 95 pp.– “Drogodependencias”, 1991, 132 ps.– “El hombre y su entorno”, 1992, 120 pp.– “Las especies frondosas en la repoblación de Galicia”, 1992, 125 pp.– “La conservación del bosque”, 1992, 131 pp.– “El cáncer, prevención y futuro”, 1993, 206 pp.– “Xornadas Luso-Galaicas de Ciencias, 1990-1992”, 1993, 249 pp.– “Biotecnologías”, 1994, 155 pp.– “Las Rías Bajas y el Arco Atlántico”, 1995, 253 pp.– “La pesca en Galicia: Presente y futuro”, 1996, 129 pp.– “Biodiversidad forestal”, 1997, 148 pp.– “La conservación del medio ambiente”, 1998, 221 pp.– “La conservación del entorno humano”, 1999, 87 pp.– “Biotecnologías: Biotecnología alimentaria”, 1999, 262 pp.– “Recursos agroalimentarios de Galicia”, 1999, 182 pp.– “Las aguas de la provincia de Ourense”, 1999, 126 pp.– “Recursos naturales de Galicia”, 1999, 105 pp.– “Energía y Futuro”, 2000, 246 pp.– “Aspectos actuales del cáncer”, 2001, 143 pp.– “IV y V Avances de Ciencia y Tecnología”, 2003, 403 pp.– “XI Jornadas Luso-Galaicas de Ciencia y Desarrollo”, 2004, 125 pp.

231Revista Real Academia Galega de Ciencias. Vol. XXVII
INSTRUCCIÓNS PARA OS AUTORES
1. A Revista da Real Academia Galega de Ciencias é unha publicación de difusión internacional que publica artigos orixinais, de revisión ou notas relacionadas coa Química, Xeoloxía, Bioloxía, Farmacia, Matemáticas, Física do Cosmos, Ciencias Técnicas, Ciencias Sociais e Económicas.
2. Presentación dos Artigos. O manuscrito orixinal xunto con dúas copias deberán ser enviados ó Sr. Secretario da Real Academia Galega de Ciencias, San Roque, 2, 15704 Santiago de Com-postela. España.
3. Preparación dos Manuscritos. Os traballos presentaranse a dobre espacio, incluíndo tablas e lendas, en papel DIN-A4 (297 x 215 mm), con unha marxe de 2,5 cms. As páxinas irán numera-das consecutivamente. As posicións das táboas e ilustracións (debuxos ou fotografías) deberán ser indicadas no texto; as táboas e lendas para as ilustracións incluiranse ó final en páxinas separadas. Deberá incluirse un título reducido de non máis de 40 caracteres (incluindo espacios), para o en-cabezamento das páxinas, sempre que o título completo sexa máis largo que aquél. Os manuscritos presentaranse terminados, en forma completa, e coa dirección do autor ou autores. Para a súa aceptación e publicación, serán sometidos ó xuízo de tres censores.
Co fin de facilita-la composición da Revista da Real Academia Galega de Ciencias, os autores deberán entregar xunto co traballo un CD que conteña os archivos de texto (preferentemente en Microsoft Word 97 para Windows) e gráficos. Os gráficos almacenaranse en formato CorelDraw para Windows (.CDR) ou en formato .EPS, e as fotografías deberán grabarse en formatos PhotoShop, .TIFF ou .JPG (este último coa menor compresión posible). Para conseguir unha mellor calidade de reproducción, é necesario que os archivos EPS, TIFF e JPG incluídos no diskette fosen gravados directamente desde o programa de gráficos. Na etiqueta do diskette figurará o título do traballo e o nome e iniciais do primeiro asinante.
(a) Resumo. Ó comenzo do traballo incluirase un resumo, que non debe exceder das 150 palabras, no que se deberá reflectir concisamente o contido do traballo sen facer referencia ó texto principal. Virá escrito no mesmo idioma do traballo e máis nun segundo idioma de uso común en publica-cións científicas.
(b) Verbas claves. Adxuntarase unha selección ad hoc de verbas claves que se imprimirá co resumo. Deberán presentarse coa súa traducción ó mesmo idioma utilizado no resumo.
(c) Bibliografía. As referencias bibliográficas ordenaranse alfabeticamente, ó final do traballo, dispostas como nos exemplos seguintes:
Clutton-Brok, T.H., Harvey, P.H. (1977). Primate ecology and social organization. Journal of Zoology, 183, 1-39.
Kawase, M. (1969). Root promoting substances in Salix alba. Physiol. Plant, 23, 159-170.
As citas no texto escribiranse como segue: Smith e Robinson (1984). Se a cita é de máis de dous autores, pode usarse Smith et al (1979).
Corrección de probas. Os autores recibirán as probas do prelo para a súa corrección, debendo evitarse as modificacións substanciais do manuscrito que, en caso de se producir, serán cargadas ós autores. As probas deberán ser devoltas ó editor, debidamente corrixidas, dentro dos dez días seguintes ó da súa recepción polo autor.
Separatas. Entregaranse 25 separatas gratuítas de cada traballo. As copias adicionais deberán ser encargadas separadamente, realizando o pedido ó devolver as probas corrixidas.

232 Revista Real Academia Galega de Ciencias. Vol. XXVII
INSTRUCCIONES PARA LOS AUTORES
1. La Revista de la Real Academia Gallega de Ciencias es una publicación de difusión internacional que publica artículos originales, de revisión o notas relacionadas con la Química, Geología, Biología, Farmacia, Matemáticas, Física del Cosmos, Ciencias Técnicas, Ciencias Sociales y Económicas.
2. Presentación de los Artículos. El manuscrito original junto con dos copias deberán ser enviados al Sr. Secretario de la Real Academia Gallega de Ciencias, San Roque, 2, 15704 Santiago de Compostela. España.
3. Preparación de los Manuscritos. Los trabajos se presentarán a doble espacio, incluyendo tablas y leyendas, en papel DIN-A4 (297 x 215 mm), con un margen de 2,5 cms. Las páginas irán numeradas consecutivamente. Las posiciones de las tablas e ilustraciones (dibujos o fotografías) deberán ser indicadas en el texto; las tablas y leyendas para las ilustraciones se incluirán al final en páginas separadas. Deberá incluirse un título reducido de no más de 40 caracteres (incluyendo espacios), para el encabezamiento de las páginas, siempre que el título completo sea más largo que aquél. Los manuscritos se presentarán terminados, en forma completa, y con la dirección del autor o autores. Para su aceptación y publicación, serán sometidos al juicio de tres censores.
Con el fin de facilitar la composición de la Revista de la Real Academia Gallega de Ciencias, los autores deberán entregar junto con el trabajo un CD que contenga los archivos de texto (preferen-temente en Microsoft Word 97 para Windows) y gráficos. Los gráficos se almacenarán en formato CorelDraw para Windows (.CDR) o en formato .EPS, y las fotografías deberán grabarse en formatos PhotoShop, .TIFF o .JPG (este último con la menor compresión posible). Para conseguir una mejor calidad de reproducción, es necesario que los archivos EPS, TIFF y JPG incluídos en el diskette fuesen grabados directamente desde el programa de gráficos. En la etiqueta del diskette figurará el título del trabajo y el nombre e iniciales del primer firmante.
(a) Resumen. Al comienzo del trabajo se incluirá un resumen, que no debe exceder de las 150 palabras, en el que se deberá reflejar concisamente el contenido del trabajo sin hacer referencia al texto principal. Vendrá escrito en el mismo idioma del trabajo y en un segundo idioma de uso común en publicaciones científicas.
(b) Palabras claves. Se adjuntará unha selección ad hoc de palabras claves que se imprimirá con el resumen. Se deberán presentar con su traducción al mismo idioma utilizado en el resumen.
(c) Bibliografía. Las referencias bibliográficas se ordenarán alfabeticamente, al final del trabajo, dispuestas como en los ejemplos siguientes:
Clutton-Brok, T.H., Harvey, P.H. (1977). Primate ecology and social organization. Journal of Zoology, 183, 1-39.
Kawase, M. (1969). Root promoting substances in Salix alba. Physiol. Plant, 23, 159-170.
Las citas en el texto se escribirán como sigue: Smith e Robinson (1984). Si la cita es de más de dos autores, puede usarse Smith et al (1979).
Corrección de pruebas. Los autores recibirán las pruebas de imprenta para su corrección, debiendo evitarse las modificaciones substanciales del manuscrito que, en caso de producirse, serán cargadas a los autores. Las pruebas deberán ser devueltas al editor, debidamente corregidas, dentro de los diez días siguientes al de su recepción por el autor.
Separatas. Se entregarán 25 separatas gratuitas de cada trabajo. Las copias adicionales deberán ser encargadas separadamente, realizando el pedido al devolver las pruebas corregidas.

233Revista Real Academia Galega de Ciencias. Vol. XXVII
INSTRUCTIONS TO AUTHORS
1. The Revista de la Real Academia Galega de Ciencias is a Journal with international distribution which publishes research papers and notes in various areas of Chemistry, Geology, Biology, Phar-macy, Mathematics, Physic, Technology, and Social and Economical Sciences.
2. Submission of Manuscripts. The original typescript and two copies should be sent to the Sr. Secretario de la Real Academia Gallega de Ciencias, San Roque, 2, 15704 Santiago de Com-postela. Spain.
3. Rules for the Manuscript. Papers should be written in double spacing throughout (including tables, legends and footnotes), on one side of DIN-A4 (297 x 215 mm) paper with a margin of 2.5 cm all round. The position of tables and illustrations (pictures or photographs) should be indicated in the text: tables and legends for illustrations should be typed separately at the end of the manuscript. A short running title of not more that 40 characters (including spaces), suitable for page headings, should be suplied if the full title is longer than this. Manuscripts should be submitted in a complete and finished form and with the author’s complete address. Before aceptation for publication, they will be submitted to three referees.
In order to help with the composition of the Revista de la Real Academia Galega de Ciencias, authors should hand in together with their work a CD containing text files (Microsoft Word 97 for Windows is the prefered program) and graphics. Graphics should be saved in CorelDraw for Windows (.CDR) or as .EPS, and photos should be saved in PhotoShop, .TIFF o .JPG formats (JPG’s with as little compression as possible). In order to obtain the best possible print quality, the EPS, TIFF and JPG files included on the disk should be saved directly from the graphics programme. The disk label should feature the title of the work and the surname and initials of the main author.
(a) Abstract. An abstract will be printed at the head of all papers; abstracts should not exceed 150 words, and should be intelligible to the general reader without reference to the main text. Abstracts should be written in the same language as the paper and in a second language of common use in the scientific literature.
(b) Key words. An ad hoc selection of key words should be supplied; this will be printed with the abstract.
(c) References. References should be listed alphabetically at the end of the paper, arranged as in the following examples:
Clutton-Brok, T.H., Harvey, P.H. (1977). Primate ecology and social organization. Journal of Zoology, 183, 1-39.
Kawase, M. (1969) Root promoting substances in Salix alba. Physiol. Plant, 23, 159-170.
Citation in the text should read thus: Smith and Robinson (1984). When a citation has more than two authors, Smith et al. (1979) may be used.
Corrections. The publisher provides proofs for checking. Corrections must be restricted to typo-graphical errors; those that represent substantial alterations from the submitted manuscript may be carged to the author. Corrections should be returned to the editor within at most ten days.
Reprints. For each published contribution, 25 reprints will be sent free of charge. Additional copies may be purchased; the order form shoul be returned with the proofs.































