Embed Size (px)
Citation preview

Available online at www.sciencedirect.com
ology 227 (2008) 275–283www.elsevier.com/locate/ytaap
Toxicology and Applied Pharmac
Rats fed soy protein isolate (SPI) have impaired hepatic CYP1A1 inductionby polycyclic aromatic hydrocarbons as a result of interference
with aryl hydrocarbon receptor signaling
Rohit Singhal a, Thomas M. Badger b,c, Martin J. Ronis a,c,⁎
a Department of Pharmacology and Toxicology, University of Arkansas for Medical Sciences, Little Rock, AR 72205, USAb Department of Physiology and Biophysics, University of Arkansas for Medical Sciences, Little Rock, AR 72205, USA
c Arkansas Children's Nutrition Center, Little Rock, AR 72202, USA
Received 17 August 2007; revised 2 November 2007; accepted 12 November 2007Available online 21 November 2007
Abstract
Consumption of soy diets has been found to reduce cancer incidence in animals and is associated with reduced cancer risk in humans. Previously,we have demonstrated that female Sprague–Dawley rats fed purified AIN-93G diets with soy protein isolate (SPI) as the sole protein source hadreduced CYP1A1 induction and basal aryl hydrocarbon receptor (AhR) levels relative to those fed the same diet containing casein (CAS). In thepresent study, the molecular mechanisms underlying reduced AhR expression have been studied. The SPI-effect on AhR was not observed afterfeeding diets containing the purified soy isoflavones genistein or daidzein. Rat hepatoma FGC-4 cells were treated with the serum obtained from ratsfed CAS- or SPI-containing diets. Reduced AhR levels (Pb0.05) were observed after 24 h exposure to SPI-serum without any changes in the overallexpression of chaperone proteins—HSP90 and XAP2. SPI-serum-stimulated AhR degradation was inhibited by treating the cells with theproteasome inhibitor, MG132, and was observed to be preceded by ubiquitination of the receptor. A reduced association of XAP2 with theimmunoprecipitated AhR complex was observed. SPI-serum-mediated AhR degradation was preceded by nuclear translocation of the receptor.However, the translocated receptor was found to be unable to heterodimerize with ARNTor to bind toXRE elements on the CYP1A1 enhancer. Thesedata suggest that feeding SPI-containing diets antagonizes AhR signaling by a novel mechanismwhich differs from those established for knownAhRantagonists.© 2007 Elsevier Inc. All rights reserved.
Keywords: Soy; Aryl hydrocarbon receptor; CYP1A1 inhibition; Proteasome
Introduction
Epidemiological studies suggest lower risk for certaincancers in populations that consume Asian diets rich in soy
Abbreviations: AhR, aryl hydrocarbon receptor; ARNT, aryl hydrocarbonreceptor nuclear translocator; CAS, casein; ChIP, chromatin immunoprecipita-tion assay; CYP1A1, cytochrome P450 1A1; DMBA, 7,12-dimethylbenz [a]anthracene; DAID, daidzein; GEN, genistein; GR, glucocorticoid receptor;HSP90, heat shock protein 90; SPI, soy protein isolate (associated withphytochemicals); PAHs, polycyclic aromatic hydrocarbons; PND, post natalday; XAP2, immunophilin-like-X-associated protein; XRE, xenobiotic responseelement.⁎ Corresponding author. Arkansas Children's Nutrition Center, Slot 512-20B,
1212 Marshall Street, Little Rock, AR 72202, USA. Fax: +1 501 364 3161.E-mail address: [email protected] (M.J. Ronis).
0041-008X/$ - see front matter © 2007 Elsevier Inc. All rights reserved.doi:10.1016/j.taap.2007.11.009
(Persky and Van Horn, 1995; Sharp et al., 2005). A number ofstudies have shown that animals fed diets containing soy proteinhave reduced incidence of chemically-induced mammary, pros-tate and colon cancers including those induced by exposure topolycyclic aromatic hydrocarbons (PAHs) (Badger et al., 2005;Simmen et al., 2005; Xiao et al., 2005), but the mechanism(s) isstill unknown. PAHs and halogenated aromatic hydrocarbons,such as dioxins, anthracenes and cholanthrene, are commonlyfound environmental carcinogens. These compounds act asligands to the aryl hydrocarbon receptor (AhR) which is aligand-activated member of the basic helix–loop–helix/Per-ARNT-Sim (bHLH/PAS) family of transcription factors (Kew-ley et al., 2004). It can be activated by numerous structurallydiverse synthetic and natural occurring phytochemicals includ-ing indoles and flavonoids (Denison et al., 2002). Activation of

276 R. Singhal et al. / Toxicology and Applied Pharmacology 227 (2008) 275–283
AhR may initiate a cascade of secondary and tertiary changesin gene expression leading to carcinogenicity, wasting syn-drome, teratogenicity, hepatotoxicity, immunosuppression andenzyme induction (Denison and Heath-Pagliuso, 1998; Polandand Knutson, 1982). The AhR normally remains in the cyto-plasm bound with chaperone proteins—two molecules of heatshock protein 90 (HSP90), immunophillin like X associatedprotein (XAP2) and a co-chaperone to HSP90, 23 kDa protein(p23). These chaperone proteins maintain the stability, con-formation and localization of the AhR protein (Petrulis andPerdew, 2002). Binding of a ligand exposes nuclear localiza-tion signal present in the N-terminal of AhR and leads totranslocation of the complex to nucleus where AhR hetero-dimerizes with aryl hydrocarbon receptor nuclear translocator(ARNT). Together the ligand:AhR/ARNT complex binds to avariety of genes including phase I and II genes such as cyto-chrome P450 1A1 (CYP1A1) and NADPH-quinone reductase(reviewed in Bock, 1994; Bock and Kohle, 2006). Increasingexperimental evidence suggests that both liganded and unli-ganded AhR are involved in cell differentiation, cell cyclecontrol and in maintenance of adequate level of growth factors(Marlowe and Puga, 2005; Huang and Elferink, 2005; Ma andWhitlock, 1996; Puga et al., 2005). Studies performed withtransgenic mice suggest that both constitutive and activatedAhR expression promote hepatocarcinogenesis (Moennikeset al., 2004) and stomach tumors (Andersson et al., 2002).Thus, the AhR has the potential to alter myriad events that haveimportant toxicological end points and actions that lead toreduction in AhR expression might play a differential role inreducing associated toxicities.
Protein degradation through the ubiquitin-26Sproteasomepathway constitutes an important cellular mechanism to regulatethe gene expression and cellular protein levels (reviewed in(DeMartino and Slaughter, 1999; Varshavsky, 1997). Manyproteins, for example, cyclin D1, p53, estrogen receptor-α,NFκB, c-Jun and c-Myc have been known to be degraded by thispathway (Boutillier et al., 1999; Varshavsky, 1997; Hoch-strasser, 1996). Nearly all studies focused on AhR degradationsuggest that ligand activated AhR is degraded by 26S protea-some (Harper et al., 2006; Ma and Baldwin, 2000; Pollenz,2002, 2007); although controversial, a recent report also sug-gests that the calcium-dependent protease calpain may be re-sponsible for the ligand-mediated activation and degradation ofthe AhR (Dale and Eltom, 2006). The mechanism of degradationof unliganded AhR needs further exploration. Previous findingsfrom our lab suggested a reduction in 7,12-dimethylbenz[a]anthracene (DMBA)-induced mammary and hepatic CYP1A1protein levels in female Sprague–Dawley rats (Rowlands et al.,2001) and a reduction in 3-methylcholanthrene-induced hepaticCYP1A1 protein levels in male rats (Ronis et al., 2001) fed soyprotein isolate (SPI)-containing diets compared to diets wherethe protein source was casein (CAS). On subsequent analysis, areduction in the basal hepatic AhR protein level, but not mRNA,was observed which was specific to the consumption of SPI-associated phytochemicals rather than the protein itself. An exvivo model was developed to recapitulate the findings from thein vivo model. A reduction in the AhR protein half life was
observed following treatment with serum from SPI-fed rats (SPI-serum) in the presence of cycloheximide (CHX), a proteinsynthesis inhibitor, suggesting AhR turnover stimulated by an asyet to be identified SPI-associated factor(s) present in the serumof rats fed SPI-containing diets (Singhal et al., 2007).
The earlier studies suggested that AhR degradation might bea mechanism for the chemoprevention following soy consump-tion against DMBA-mediated tumorigenesis (Rowlands et al.,2001; Ronis et al., 2001; Singhal et al., 2007). However, themechanisms that lead to AhR degradation upon soy consump-tion were not investigated. In the present studies, we havedemonstrated that SPI actions differ from those of the soyisoflavones, genistein and daidzein. Also, we have probed theevents preceding AhR degradation upon treatment of FGC-4cells with SPI-serum. We observed that SPI-serum treatmentstimulates AhR translocation to nucleus and a dissociation ofXAP2 from the AhR complex. However, failure of thetranslocated AhR to associate with ARNT prevents DNAbinding and leads to receptor ubiquitination and degradation.Many studies have examined AhR degradation by agonists andantagonists of AhR-mediated signaling by dietary phytochem-icals. Here, we present a novel scenario of how the consumptionof a dietary component might lead to AhR degradation andreduction in associated toxicities.
Methods
Animals and diets. The animal studies received prior approval from theInstitutional Animal Care and Use Committee at UAMS. Female Sprague–Dawley rats were obtained from Charles River Laboratories Inc. (Wilmington,MA) and were housed in polycarbonate cages in an environmentally controlledroom with a 12 h light–dark cycle, and were provided with pelleted diets madeof casein (CAS) or SPI. The specialized semi-purified diets were made accord-ing to the AIN-93G diet formula (Reeves et al., 1993) except that corn oilreplaced soybean oil and the protein source was either CAS or SPI. The SPI diethad 430 mg total isoflavones/kg containing 276 mg/kg genistein and 132 mg/kgdaidzein; the stripped SPI-diet contained the same level of SPI protein which hadbeen stripped of phytochemicals by successive ethanol washes (Singhal et al.,2007); the genistein (GEN) diet (CAS+GEN) had 276 mg/kg genistein; and thedaidzein (DAID) diet (CAS+DAID) had 132 mg/kg daidzein. In all the expe-riments, rats were euthanized by injection with 100 mg/kg Nembutol, followedby decapitation, and the serum and liver were collected.
Experiment 1. Female rats (n=12) had ad libitum access to CAS, SPI, GENor DAID-containing diets between postnatal day 21 (PND21) and PND50. Todetermine the effect of the semi-purified diets on DMBA-mediated CYP1A1induction, rats (n=6/from each diet group, PND50) were orally gavaged with50 mg/kg DMBA or sesame oil followed by termination after 24 h. Liver wascollected after termination and kept frozen at −80 °C until used.
Experiment 2 and ex vivo model. Female rats (n=6) were fed ad libitum withCAS, SPI or stripped SPI diets between PND21 and PND46. Serum and liverwere collected after termination. Sera from each group of rats were pooled, filtersterilized and stored at −20 °C until use. Rat FGC-4 cells (obtained from MaryC. Weiss, Pasteur Institute, Paris, France) were cultured in Dulbecco's ModifiedEagle's Medium (DMEM) and fetal bovine serum (FBS, Invitrogen), as de-scribed previously (McGehee et al., 1994). All the experiments were performedat 80% confluency from cell passages 9 to 14. After 24 h incubation in serum-free DMEM, cells were treated with 5% serum from rats fed CAS, SPI orstripped SPI-containing diets. In the experiments, where indicated, cells weretreated with protein synthesis inhibitor, CHX (Sigma, St. Louis, MO), andproteasome inhibitor, MG132 (EMD Chemicals Inc., San Diego, CA), at thefinal concentration of 5 mg/L and 5 μM 1 h prior to serum treatment.
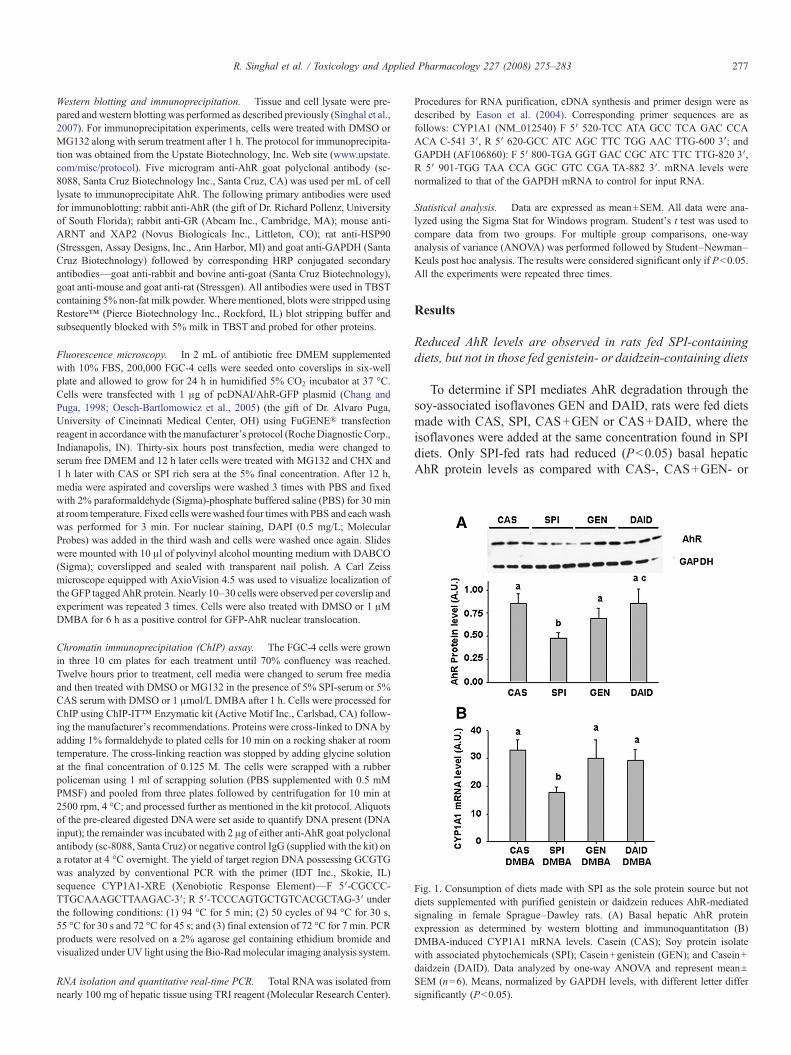
Fig. 1. Consumption of diets made with SPI as the sole protein source but notdiets supplemented with purified genistein or daidzein reduces AhR-mediatedsignaling in female Sprague–Dawley rats. (A) Basal hepatic AhR proteinexpression as determined by western blotting and immunoquantitation (B)DMBA-induced CYP1A1 mRNA levels. Casein (CAS); Soy protein isolatewith associated phytochemicals (SPI); Casein+genistein (GEN); and Casein+daidzein (DAID). Data analyzed by one-way ANOVA and represent mean±SEM (n=6). Means, normalized by GAPDH levels, with different letter differsignificantly (Pb0.05).
277R. Singhal et al. / Toxicology and Applied Pharmacology 227 (2008) 275–283
Western blotting and immunoprecipitation. Tissue and cell lysate were pre-pared and western blotting was performed as described previously (Singhal et al.,2007). For immunoprecipitation experiments, cells were treated with DMSO orMG132 along with serum treatment after 1 h. The protocol for immunoprecipita-tion was obtained from the Upstate Biotechnology, Inc. Web site (www.upstate.com/misc/protocol). Five microgram anti-AhR goat polyclonal antibody (sc-8088, Santa Cruz Biotechnology Inc., Santa Cruz, CA) was used per mL of celllysate to immunoprecipitate AhR. The following primary antibodies were usedfor immunoblotting: rabbit anti-AhR (the gift of Dr. Richard Pollenz, Universityof South Florida); rabbit anti-GR (Abcam Inc., Cambridge, MA); mouse anti-ARNT and XAP2 (Novus Biologicals Inc., Littleton, CO); rat anti-HSP90(Stressgen, Assay Designs, Inc., Ann Harbor, MI) and goat anti-GAPDH (SantaCruz Biotechnology) followed by corresponding HRP conjugated secondaryantibodies—goat anti-rabbit and bovine anti-goat (Santa Cruz Biotechnology),goat anti-mouse and goat anti-rat (Stressgen). All antibodies were used in TBSTcontaining 5% non-fat milk powder. Where mentioned, blots were stripped usingRestore™ (Pierce Biotechnology Inc., Rockford, IL) blot stripping buffer andsubsequently blocked with 5% milk in TBST and probed for other proteins.
Fluorescence microscopy. In 2 mL of antibiotic free DMEM supplementedwith 10% FBS, 200,000 FGC-4 cells were seeded onto coverslips in six-wellplate and allowed to grow for 24 h in humidified 5% CO2 incubator at 37 °C.Cells were transfected with 1 μg of pcDNAI/AhR-GFP plasmid (Chang andPuga, 1998; Oesch-Bartlomowicz et al., 2005) (the gift of Dr. Alvaro Puga,University of Cincinnati Medical Center, OH) using FuGENE® transfectionreagent in accordance with themanufacturer's protocol (RocheDiagnostic Corp.,Indianapolis, IN). Thirty-six hours post transfection, media were changed toserum free DMEM and 12 h later cells were treated with MG132 and CHX and1 h later with CAS or SPI rich sera at the 5% final concentration. After 12 h,media were aspirated and coverslips were washed 3 times with PBS and fixedwith 2% paraformaldehyde (Sigma)-phosphate buffered saline (PBS) for 30 minat room temperature. Fixed cells werewashed four timeswith PBS and eachwashwas performed for 3 min. For nuclear staining, DAPI (0.5 mg/L; MolecularProbes) was added in the third wash and cells were washed once again. Slideswere mounted with 10 μl of polyvinyl alcohol mounting medium with DABCO(Sigma); coverslipped and sealed with transparent nail polish. A Carl Zeissmicroscope equipped with AxioVision 4.5 was used to visualize localization ofthe GFP taggedAhR protein. Nearly 10–30 cells were observed per coverslip andexperiment was repeated 3 times. Cells were also treated with DMSO or 1 μMDMBA for 6 h as a positive control for GFP-AhR nuclear translocation.
Chromatin immunoprecipitation (ChIP) assay. The FGC-4 cells were grownin three 10 cm plates for each treatment until 70% confluency was reached.Twelve hours prior to treatment, cell media were changed to serum free mediaand then treated with DMSO or MG132 in the presence of 5% SPI-serum or 5%CAS serum with DMSO or 1 μmol/L DMBA after 1 h. Cells were processed forChIP using ChIP-IT™ Enzymatic kit (Active Motif Inc., Carlsbad, CA) follow-ing the manufacturer's recommendations. Proteins were cross-linked to DNA byadding 1% formaldehyde to plated cells for 10 min on a rocking shaker at roomtemperature. The cross-linking reaction was stopped by adding glycine solutionat the final concentration of 0.125 M. The cells were scrapped with a rubberpoliceman using 1 ml of scrapping solution (PBS supplemented with 0.5 mMPMSF) and pooled from three plates followed by centrifugation for 10 min at2500 rpm, 4 °C; and processed further as mentioned in the kit protocol. Aliquotsof the pre-cleared digested DNAwere set aside to quantify DNA present (DNAinput); the remainder was incubated with 2 μg of either anti-AhR goat polyclonalantibody (sc-8088, Santa Cruz) or negative control IgG (supplied with the kit) ona rotator at 4 °C overnight. The yield of target region DNA possessing GCGTGwas analyzed by conventional PCR with the primer (IDT Inc., Skokie, IL)sequence CYP1A1-XRE (Xenobiotic Response Element)—F 5′-CGCCC-TTGCAAAGCTTAAGAC-3′; R 5′-TCCCAGTGCTGTCACGCTAG-3′ underthe following conditions: (1) 94 °C for 5 min; (2) 50 cycles of 94 °C for 30 s,55 °C for 30 s and 72 °C for 45 s; and (3) final extension of 72 °C for 7 min. PCRproducts were resolved on a 2% agarose gel containing ethidium bromide andvisualized under UV light using the Bio-Radmolecular imaging analysis system.
RNA isolation and quantitative real-time PCR. Total RNAwas isolated fromnearly 100 mg of hepatic tissue using TRI reagent (Molecular Research Center).
Procedures for RNA purification, cDNA synthesis and primer design were asdescribed by Eason et al. (2004). Corresponding primer sequences are asfollows: CYP1A1 (NM_012540) F 5′ 520-TCC ATA GCC TCA GAC CCAACA C-541 3′, R 5′ 620-GCC ATC AGC TTC TGG AAC TTG-600 3′; andGAPDH (AF106860): F 5′ 800-TGA GGT GAC CGC ATC TTC TTG-820 3′,R 5′ 901-TGG TAA CCA GGC GTC CGA TA-882 3′. mRNA levels werenormalized to that of the GAPDH mRNA to control for input RNA.
Statistical analysis. Data are expressed as mean±SEM. All data were ana-lyzed using the Sigma Stat for Windows program. Student's t test was used tocompare data from two groups. For multiple group comparisons, one-wayanalysis of variance (ANOVA) was performed followed by Student–Newman–Keuls post hoc analysis. The results were considered significant only if Pb0.05.All the experiments were repeated three times.
Results
Reduced AhR levels are observed in rats fed SPI-containingdiets, but not in those fed genistein- or daidzein-containing diets
To determine if SPI mediates AhR degradation through thesoy-associated isoflavones GEN and DAID, rats were fed dietsmade with CAS, SPI, CAS+GEN or CAS+DAID, where theisoflavones were added at the same concentration found in SPIdiets. Only SPI-fed rats had reduced (Pb0.05) basal hepaticAhR protein levels as compared with CAS-, CAS+GEN- or

Fig. 3. SPI-serummediates AhR degradation by ubiquitin-proteasomemachineryin FGC-4. Cells were treated with MG132 or Calpain Inhibitor II followed byserum from SPI- or CAS-fed rats after 1 h for 24 h. Cells were lysed and (A) AhRprotein expression (top panel) and levels (bottom panel) were analyzed. GAPDH
278 R. Singhal et al. / Toxicology and Applied Pharmacology 227 (2008) 275–283
CAS+DAID-fed rats. SPI consumption in the rats also led topartial blockade (Pb0.05) in DMBA-mediated hepatic CYP1A1mRNA induction compared with CAS-, GEN- or DAID-fed rats(Fig. 1).
Exposure to SPI-serum results in AhR degradation byubiquitin-proteasome machinery in FGC-4 cells
Previously published data from our laboratory suggested that24 h treatment of FGC-4 cells with serum from rats fed SPIresulted in AhR degradation and a lower DMBA-mediatedCYP1A1 induction (Singhal et al., 2007). One of the reasons forthis degradation could be that a phytochemical bound to SPItargets AhR chaperone protein expression levels. Serum fromrats fed stripped SPI-diets (SPI−), where the protein was strippedof phytochemicals, had no effect on AhR or CYP1A1 expressionlevels (data not shown). These data are consistent with ourpreviously performed in vivo studies demonstrating a lack ofeffect of stripped SPI-diets on hepatic CYP1A1-induction(Singhal et al., 2007). The effect of SPI-serum treatment on thetotal cellular levels of XAP2 and HSP90 was evaluated. Asshown in Fig. 2, no effect of SPI-serum was observed on eitherXAP2 or HSP90 protein expression in the total cell lysate;however, AhR levels were reduced (Pb0.05) by SPI-serumtreatment comparedwith CAS serum-treated groups, as observedin our previous study (Singhal et al., 2007). No significant effectof SPI-serum was observed on ARNTor glucocorticoid receptor(GR) expression and protein levels (Fig. 2).
Fig. 2. Exposure to SPI-serum results in AhR degradation in FGC-4 cells. TheFGC-4 cells were treated with sera from SPI- or CAS-fed rats for 24 h.(A) Protein expression and (B) immune levels of AhR, HSP90, XAP2, ARNTand GR were determined by western blot analysis and immunoquantitation,respectively. GAPDH was used as a loading control. Data are represented aspercent of CAS control (100%), n=3 experiments. ⁎ represents difference fromCAS, Pb0.05.
was used as an internal loading control. Data are means±SEM, n=3 experiments.⁎ represents difference from CAS control, Pb0.05. (B) FGC-4 cells were treatedwith MG132 or DMSO followed by serum treatment for 24 h. AhR wasimmunoprecipitated and probed for ubiquitin (top panel) or AhR (bottom panel)proteins by western blotting.
Most studies performed with exogenous ligands suggest 26Sproteasome-mediated AhR degradation. However, recent re-ports also suggest roles of calpain proteases in ligand-mediatedAhR degradation (Dale and Eltom, 2006). How the unligandedAhR degrades is still not clearly understood. Therefore, todelineate the mechanism of degradation triggered by SPI, cellswere treated either withMG132 or Calpain Inhibitor II (5 μM) orDMSO. Twenty-four hour treatment with MG132 resulted in thecomplete blockage of AhR degradation while Calpain Inhibitor-II or DMSO (negative control) had no affect on degradation(Fig. 3A). Proteins which are targeted for proteasomal degrada-tion are generally tagged with ubiquitin molecules in order to beidentified by the proteasomes (Ciechanover, 1998). Immuno-precipitation and subsequent immunoblotting of AhR proteinwith ubiquitin antibody from the cells treated with MG132 orDMSO revealed that SPI serum treatment resulted in AhRubiquitination prior to degradation, as shown in Fig. 3B (upperpanel). The immunoblot used for probing ubiquitin protein onimmunoprecipitated AhR was stripped and reprobed for AhRprotein. Once again, the immunoblot revealed that treatmentwith MG132 blocked the SPI serum-mediated degradation ofAhR (bottom panel).

Fig. 4. SPI-mediated proteasomal degradation of AhR is preceded by reducedassociation with XAP2 in FGC-4 cells. The FGC-4 cells were treated withMG132 followed by serum from either CAS- or SPI-fed rats after 1 h for 24 h.The AhR was immunoprecipitated and immunoblotted for XAP2, HSP90 andAhR proteins. The experiments were repeated three times.
279R. Singhal et al. / Toxicology and Applied Pharmacology 227 (2008) 275–283
SPI-mediated proteasomal degradation of AhR is preceded byreduced association with XAP2 in FGC-4 cells
Our next objective was to determine what promotes AhRubiquitination following SPI-serum treatment. It is well knownthat AhR chaperones determine the stability, conformation,
Fig. 5. SPI-mediated proteasomal degradation of AhR is preceded by nuclear transloAhR-GFP expression vector and fixed for fluorescence microscopy as described in Mfor nuclear translocation (left two panels) or treated with CHX and MG132 followedwere treated with MG132 followed by either CAS or SPI sera or DMBA in the preseCells were scraped and subjected to AhR immunoprecipitation followed by immuno
localization and degradation of the AhR protein (Petrulis andPerdew, 2002). It could be that the treatment with SPI-serumtriggers dissociation of chaperone proteins or reduces associa-tion of the chaperone proteins with the AhR. To test this hypo-thesis, the AhR was immunoprecipitated from the cells treatedwith serum in the presence ofMG132, and probed for XAP2 andHSP90 proteins. A reduced association of AhR protein withXAP2 was observed in the group treated with SPI-serum ascompared with a CAS-serum treated group. No affect was ob-served on the association of AhR protein with HSP90 (Fig. 4).
Exposure to SPI-serum results in nuclear translocation of AhRprotein in FGC-4 cells
To determine if the degradation is preceded by translocationof AhR to the nucleus upon SPI-serum treatment, we visualizedthe cellular localization of AhR in FGC-4 cells using a plasmidexpressing a GFP-tagged AhR protein. In non-stimulated cells(no treatment), GFP-tagged AhR was diffusely distributed in thecytoplasm in great majority of transfected cells, while DMBA(1 μM) treatment for 6 h resulted in the complete nucleartranslocation of AhR-GFP as shown by the co-localization of
cation. (A). The FGC-4 cells were transiently transfected with 1 μg of pcDNAI/ethods. Transfected cells were treated with DMBA or DMSO for 6 h as controlby serum from CAS- or SPI-fed rats for 12 h (right two panels). (B). FGC-4 cellsnce of CAS serum (lanes 2, 3 and 4) or left without treatment (lane 1) for 24 h.blotting for ARNT protein.

280 R. Singhal et al. / Toxicology and Applied Pharmacology 227 (2008) 275–283
green colored GFP and blue colored nuclear DAPI stain. Thisdemonstrates the fusion protein obtained from transfection ofFGC-4 cells with AhR-GFP plasmid responses to the exoge-nous ligand treatment. Although MG132 had been suggested tocause AhR nuclear translocation (Santiago-Josefat and Fernan-dez-Salguero, 2003), we observed that cells treated with CASserum in the presence of CHX and MG132 had cytosolic loca-lization of AhR-GFP at 24 h. However, SPI serum treatmentcaused the AhR-GFP fusion protein to translocate to nucleus(Fig. 5A).
Next we evaluated if the SPI-mediated nuclear translocatedAhR heterodimerizes with ARNT before degradation. As ex-pected, DMBA treatment in the presence of MG132 resulted inAhR heterodimerization with ARNT, as observed by immuno-precipitating AhR and immunoblotting with ARNT antibody.SPI serum treatment did not cause AhR-ARNT heterodimeriza-tion. This suggests that although SPI serum treatment trans-locates AhR to nucleus, it does not result in a conformationrequired for AhR to heterodimerize with ARNT and instead istargeted for degradation (Fig. 5B).
It is well known that AhR requires heterodimerization withARNT to bind at XREs in the enhancer region of CYP1A1gene. Therefore, we predicted that nuclear translocated AhRafter exposure to SPI serum would not bind to the CYP1A1transcription regulatory sites. To asses this, cells were treatedwith CAS or SPI serum or DMBAwith CAS serum as a positivecontrol in the presence of MG132 and were subjected to ChIPassay as described in Methods. As expected, DMBA treatmentled to binding of AhR at the XRE while reduced constitutivebinding of AhR was observed upon SPI-serum treatment com-pared to CAS-serum treatment (Fig. 6).
Fig. 6. Nuclear translocated AhR does not bind to XRE in CYP1A1 enhancerregion. FGC-4 cells were treated with MG132 followed by sera from eitherCAS- or SPI-fed rats or DMBA in the presence of CAS serum (lanes 2, 3 and 4)or left without treatment (lane 1) for 24 h. ChIP assays were performed withchromatin prepared from the treated cells using anti-AhR antibodies asdescribed in Methods. (A). Schematic representation of putative XREs (circle)and TATA box (rectangle) upstream of CYP1A1 transcription start site. Arrowsdisplay the primer binding sites. (B) PCRs of DNA from ChIP assay for input orimmunoprecipitated fractions CYP1A1 or negative IgG. The abbreviations Cand S stand for CAS and SPI, respectively.
Discussion
In the present study, we demonstrate the following: (1) Soyconsumption results in the reduction in basal AhR protein levels,but feeding the SPI-associated isoflavones, GEN and DAID, atthe same concentration as found in SPI does not have any affecton AhR-mediated signaling. (2) Treatment of FGC-4 cells withserum obtained from rats fed SPI-containing diets results inproteasome-mediated degradation of AhR. Interestingly, wefound that one of the chaperone proteins, XAP2, had reducedassociation with AhR upon SPI serum treatment. Although AhRtranslocates to nucleus, it does not heterodimerize with ARNTorbind to XRE present in the CYP1A1 enhancer region.
Previously, we demonstrated that soy consumption results inreduction in AhR protein levels (Rowlands et al., 2001; Singhalet al., 2007; Ronis et al., 2001). However, the mechanisms werenot studied. Soy contains a number of protein-associated iso-flavone glycosides and other phytochemical conjugates that arehydrolyzed by the gut bacteria during digestion (Tokunaga,2004). Therefore, an in vitromodel using phytochemical extractsfrom SPI cannot mimic the in vivo SPI feeding model, since thesoy extracts require appropriate digestion. To overcome thisdrawback, an ex vivomodel was developed, using the FGC-4 rathepatoma cell line. The FGC-4 cell line is derived fromH4IIEC3hepatoma and expresses a wide spectrum of differentiated hepa-tocyte-specific proteins (Angrand et al., 1990), especially albu-min and cytochrome P450 enzymes (McGehee et al., 1994).
Many dietary components such as epigallocatechin galate(EGCG) in green tea, resveratrol in red wine, curcumin andphytochemicals such as kaempferol have been shown to anta-gonize AhR-mediated signaling (Casper et al., 1999; Ciolinoet al., 1998; Palermo et al., 2005; Puppala et al., 2007) and thusreduce activated AhR-mediated toxicity, particularly carcino-genicity. It is conceivable that the AhR-mediated signaling canbe targeted at number of steps involved in signaling, for ex-ample, by (1) transcriptional modification of AhR mRNA andpost-translational modification of AhR protein; (2) influencingthe stability of the AhR complex; (3) competition with theligand for binding AhR; (4) modifying conformation of AhR;(5) preventing the association of the AhR with ARNT; and/or(6) preventing the ability of binding of ligand AhR:ARNTcomplex with the DNA binding elements. So far, most of thephytochemicals have been shown to antagonize the AhR-mediated pathway by competitively binding with AhR and thusreducing AhR transformation upon exogenous ligand exposure.EGCG, however, was demonstrated to inhibit the AhR tran-scriptional activity through a direct binding to the C-terminus ofthe AhR chaperone protein HSP90 which resulted in the main-tenance of the interaction of HSP90 with XAP2. The treatmentwith EGCG thereby stabilizes the AhR–HSP90–XAP2 com-plex and transforms AhR to a nuclear form unable to bind withARNT and transcription regulatory sites (Palermo et al., 2005).
Ours is the first group to demonstrate that soy consumptionreduces AhR signaling by mediating its degradation. GEN andDAID, isoflavones associated with soy, have been suggestedto be responsible for some of the chemopreventive actionsexhibited by soy consumption such as reduction in epidermal

281R. Singhal et al. / Toxicology and Applied Pharmacology 227 (2008) 275–283
growth factor receptor and mammary carcinogenesis (Brownet al., 1998; Constantinou et al., 1996). However, our datasuggest that an as yet to be identified SPI-associated phyto-chemical(s) is responsible for AhR degradation and is not GENor DAID, since feeding rats with diet made with either GEN orDAID or soy protein stripped of phytochemicals had no affect onAhR degradation or DMBA-mediated CYP1A1 induction.There have been 136 phytochemicals identified as bound toSPI (Fang et al., 2004) and it is thus not surprising that not alleffects of soy are mediated via isoflavones. It is likely that soy-associated factor(s) directly destabilize the AhR by binding thatleads to its degradation. However, current lack of identificationof the SPI-associated bioactive component is a limitation incarrying out AhR binding experiments. An additional possibilityis that the soy-feeding alters some endogenous serum factorwhich modulates AhR signaling and thus acts indirectly.
It has been demonstrated both in vivo and in vitro that ligandbinding leads to rapid degradation of AhR (Pollenz, 2002).Studies performed to investigate the ligand-mediated AhR de-gradation suggest that the degradation is mediated via 26Sproteasome pathway following ubiquitination of the receptor(Davarinos and Pollenz, 1999; Ma and Baldwin, 2000; Song andPollenz, 2002). Although controversial, it has been recentlyreported that calpain proteases may also contribute to the ligand-mediated degradation of the AhR (Dale and Eltom, 2006).Treatment of cells with SPI-serum in the presence of MG132,but not by calpain inhibitor, inhibited the AhR degradation.Moreover, the 26S proteasome-mediated AhR degradation waspreceded by its ubiquitination. This led us to hypothesize that anSPI-associated factor triggers AhR degradation in a mannersimilar to exogenous ligands, in particular TCDD, since most, ifnot all, studies have used TCDD to study ligand-mediated AhRdegradation. However, the mechanism must differ because (1)SPI consumption in rats and SPI-serum treatment in cells reducebasal and DMBA-mediated CYP1A1 induction (Singhal et al.,2007); whereas, treatment with TCDD and other AhR ligandslike B[a]P and DMBA results in enormous CYP1A1 induction;and (2) TCDD-mediated AhR degradation is inhibited followingcycloheximide treatment (Pollenz et al., 2005), while we ob-served that SPI-serum treatment in the presence of cyclohex-imide further stimulates AhR degradation (Singhal et al., 2007).
Several studies have evaluated the effect of geldanamycin(GA) on AhR-mediated signaling (Pollenz et al., 2005; Songand Pollenz, 2002; Chen et al., 1997). GA is a benzoquinoneansamycin that binds directly to ATP binding site of HSP90 anddisrupts the recruitment of cochaperone p23, which has beenshown to stabilize the interaction between HSP90 and HSP90interacting proteins (Kazlauskas et al., 1999;Whitesell andCook,1996), to HSP90 heterocomplex formation, for example, HSP90complex formation with pp60v-src (Whitesell et al., 1994). Chenet al. (1997) demonstrated that GA directly binds with AhR-HSP90 complex and treatment of cells with GA results in rapiddepletion of AhR levels (Chen et al., 1997; Pollenz et al., 2005;Song and Pollenz, 2002) without having any affect on the AhRchaperone proteins—XAP2 and HSP90 (Chen et al., 1997;Meyer et al., 2000). The GA treatment also results in AhR-nuclear accumulation, transformation into a form that does not
bind with CYP1A1 DNA and degradation by 26S proteasome(Song and Pollenz, 2002). Interestingly, we observed that similarto the effects of GA on AhR-mediated signaling, treatment ofcells with SPI-serum results in no changes in the total cellularlevels of chaperone proteins or ARNT (Fig. 1); AhR degradationby 26S proteasome (Fig. 2); nuclear accumulation of GFP-tagged AhR protein (Fig. 4A); lack of dimerization with ARNT(Fig. 4B); no binding of nuclear translocated AhR at CYP1A1enhancer region (Fig. 5); and reduced ligand-mediated AhRtransformation and CYP1A1 induction (Singhal et al., 2007).Moreover, both SPI-associated factors and GA-mediated AhRdegradation were not inhibited by cycloheximide treatment(Singhal et al., 2007; Pollenz et al., 2005; Song and Pollenz,2002; Kazlauskas et al., 2001). A study performed with thetruncation of COOH-terminal transactivation domain of AhRsuggests that this domain is necessary for CHX-sensitivedegradation of AhR (Pollenz et al., 2005). One might assumethat SPI serum and GA act in a similar fashion since theirtreatment protects AhR from a cycloheximide-sensitive factorresponsible for protection of AhR from being degraded. Thissuggests that (1) there could be multiple pathways involved in26S proteasome-mediated degradation of AhR; and (2) theremight be a similarity between the mechanisms of AhR degra-dation assisted by SPI-associated factor and GA. Contrary to thehypothetical model drawn by Song et al. for GA-mediated AhRdegradation (Song and Pollenz, 2002) and as discussed above forits similarities with SPI-associated factor-mediated AhR degra-dation, Kazlauskas et al. concluded that GA blocks the nucleartranslocation of the AhR by interfering in the p23 binding withHSP90. It was concluded that GA affects both the maturation ofAhR–chaperone complex formation as well as promotes disso-ciation of XAP2 protein from the mature complex (Kazlauskaset al., 2001). As shown in Fig. 3, our data also indicate reducedassociation of XAP2 with the immunoprecipitated AhR.Numerous studies performed to evaluate the role of XAP2 inAhR complex suggest that XAP2 is critical in maintaining AhRlocalization in cytoplasm. Reduced XAP2 binding has beenshown to result in nuclear localization, ubiquitination anddegradation of AhR (Kazlauskas et al., 2000; LaPres et al., 2000;Meyer and Perdew, 1999). However, if SPI-associated factor isacting by the samemechanism asGA then like GA, it should alsolead to the degradation of other proteins such as c-Raf1, glu-cocorticoid receptor and CDK4 which interact with HSP90 andmature in a fashion similar to AhR (Chen et al., 1997). As shownin Fig. 1, no effect of SPI-serum treatment was observed on GRprotein expression and levels. Moreover, XAP2 has been knownto associate with AhR-HSP90, but not with GR–HSP90complex (Carver et al., 1998) which indicates that mechanismsof actions of the SPI-associated factor and GA, though verymuch similar, are not exactly the same.
With the current understanding of AhRdegradation paradigm,we speculate that treatment with SPI-associated factor concealsthe transactivation domain of AhR, and makes conformationalchanges in AhR complex followed by nuclear translocation,dissociation of XAP2 from the mature complex and ubiquitina-tion of AhR followed by proteasome-mediated degradation.Serum fractionation studies are in progress to identify the SPI-

282 R. Singhal et al. / Toxicology and Applied Pharmacology 227 (2008) 275–283
associated factor responsible for this phenomenon. The currentstudy provides not only an important model to assess the alter-native degradation pathway and more insight into AhR-mediatedsignaling, but also a novel mechanism of action of disease pre-vention by dietary components like soy.
Acknowledgment
The financial assistance of the USDA (CRIS# 6251-51000-005-03S) is gratefully acknowledged.
References
Andersson, P., McGuire, J., Rubio, C., Gradin, K., Whitelaw, M.L., Pettersson,S., Hanberg, A., Poellinger, L., 2002. A constitutively active dioxin/arylhydrocarbon receptor induces stomach tumors. Proc. Natl. Acad. Sci. U. S. A.99, 9990–9995.
Angrand, P.O., Kallenbach, S., Weiss, M.C., Rousset, J.P., 1990. An exogenousalbumin promoter can become silent in dedifferentiated hepatoma variants aswell as intertypic hybrids. Cell Growth Differ. 1, 519–526.
Badger, T.M., Ronis, M.J., Simmen, R.C., Simmen, F.A., 2005. Soy proteinisolate and protection against cancer. J. Am. Coll. Nutr. 24, 146S–149S.
Bock, K.W., 1994. Aryl hydrocarbon or dioxin receptor: biologic and toxicresponses. Rev. Physiol Biochem. Pharmacol. 125, 1–42.
Bock, K.W., Kohle, C., 2006. Ah receptor: dioxin-mediated toxic responses ashints to deregulated physiologic functions. Biochem. Pharmacol. 72, 393–404.
Boutillier, A.L., Kienlen-Campard, P., Loeffler, J.P., 1999. Depolarization re-gulates cyclin D1 degradation and neuronal apoptosis: a hypothesis aboutthe role of the ubiquitin/proteasome signalling pathway. Eur. J. Neurosci. 11,441–448.
Brown, N.M., Wang, J., Cotroneo, M.S., Zhao, Y.X., Lamartiniere, C.A., 1998.Prepubertal genistein treatment modulates TGF-alpha, EGF and EGF-re-ceptor mRNAs and proteins in the rat mammary gland. Mol. Cell. Endo-crinol. 144, 149–165.
Carver, L.A., LaPres, J.J., Jain, S., Dunham, E.E., Bradfield, C.A., 1998. Cha-racterization of the Ah receptor-associated protein, ARA9. J. Biol. Chem.273, 33580–33587.
Casper, R.F., Quesne, M., Rogers, I.M., Shirota, T., Jolivet, A., Milgrom, E.,Savouret, J.F., 1999. Resveratrol has antagonist activity on the aryl hydro-carbon receptor: implications for prevention of dioxin toxicity. Mol. Phar-macol. 56, 784–790.
Chang, C.Y., Puga, A., 1998. Constitutive activation of the aromatic hydro-carbon receptor. Mol. Cell Biol. 18, 525–535.
Chen, H.S., Singh, S.S., Perdew, G.H., 1997. The Ah receptor is a sensitivetarget of geldanamycin-induced protein turnover. Arch. Biochem. Biophys.348, 190–198.
Ciechanover, A., 1998. The ubiquitin–proteasome pathway: on protein deathand cell life. EMBO J. 17, 7151–7160.
Ciolino, H.P., Daschner, P.J., Wang, T.T., Yeh, G.C., 1998. Effect of curcuminon the aryl hydrocarbon receptor and cytochrome P450 1A1 in MCF-7human breast carcinoma cells. Biochem. Pharmacol. 56, 197–206.
Constantinou, A.I., Mehta, R.G., Vaughan, A., 1996. Inhibition of N-methyl-N-nitrosourea-induced mammary tumors in rats by the soybean isoflavones.Anticancer Res. 16, 3293–3298.
Dale, Y.R., Eltom, S.E., 2006. Calpain mediates the dioxin-induced activationand down-regulation of the aryl hydrocarbon receptor. Mol. Pharmacol. 70,1481–1487.
Davarinos, N.A., Pollenz, R.S., 1999. Aryl hydrocarbon receptor imported intothe nucleus following ligand binding is rapidly degraded via the cytosplasmicproteasome following nuclear export. J. Biol. Chem. 274, 28708–28715.
DeMartino, G.N., Slaughter, C.A., 1999. The proteasome, a novel proteaseregulated by multiple mechanisms. J. Biol. Chem. 274, 22123–22126.
Denison, M.S., Heath-Pagliuso, S., 1998. The Ah receptor: a regulator of thebiochemical and toxicological actions of structurally diverse chemicals.Bull. Environ. Contam. Toxicol. 61, 557–568.
Denison, M.S., Pandini, A., Nagy, S.R., Baldwin, E.P., Bonati, L., 2002.Ligand binding and activation of the Ah receptor. Chem. Biol. Interact.141, 3–24.
Eason, R.R., Velarde, M.C., Chatman Jr., L., Till, S.R., Geng, Y., Ferguson, M.,Badger, T.M., Simmen, R.C., 2004. Dietary exposure to whey proteins altersrat mammary gland proliferation, apoptosis, and gene expression duringpostnatal development. J. Nutr. 134, 3370–3377.
Fang, N., Yu, S., Badger, T.M., 2004. Comprehensive phytochemical profile ofsoy protein isolate. J. Agric. Food Chem. 52, 4012–4020.
Harper, P.A., Riddick, D.S., Okey, A.B., 2006. Regulating the regulator: factorsthat control levels and activity of the aryl hydrocarbon receptor. Biochem.Pharmacol. 72, 267–279.
Hochstrasser, M., 1996. Ubiquitin-dependent protein degradation. Annu. Rev.Genet. 30, 405–439.
Huang, G., Elferink, C.J., 2005. Multiple mechanisms are involved in Ahreceptor-mediated cell cycle arrest. Mol. Pharmacol. 67, 88–96.
Kazlauskas, A., Poellinger, L., Pongratz, I., 1999. Evidence that the co-chaperonep23 regulates ligand responsiveness of the dioxin (Aryl hydrocarbon) receptor.J. Biol. Chem. 274, 13519–13524.
Kazlauskas, A., Poellinger, L., Pongratz, I., 2000. The immunophilin-like proteinXAP2 regulates ubiquitination and subcellular localization of the dioxinreceptor. J. Biol. Chem. 275, 41317–41324.
Kazlauskas, A., Sundstrom, S., Poellinger, L., Pongratz, I., 2001. The hsp90chaperone complex regulates intracellular localization of the dioxin receptor.Mol. Cell Biol. 21, 2594–2607.
Kewley, R.J., Whitelaw,M.L., Chapman-Smith, A., 2004. The mammalian basichelix–loop–helix/PAS family of transcriptional regulators. Int. J. Biochem.Cell Biol. 36, 189–204.
LaPres, J.J., Glover, E., Dunham, E.E., Bunger, M.K., Bradfield, C.A., 2000.ARA9 modifies agonist signaling through an increase in cytosolic arylhydrocarbon receptor. J. Biol. Chem. 275, 6153–6159.
Ma, Q., Baldwin, K.T., 2000. 2,3,7,8-tetrachlorodibenzo-p-dioxin-induceddegradation of aryl hydrocarbon receptor (AhR) by the ubiquitin-protea-some pathway. Role of the transcription activaton and DNA binding of AhR.J. Biol. Chem. 275, 8432–8438.
Ma, Q., Whitlock Jr., J.P., 1996. The aromatic hydrocarbon receptor modulatesthe Hepa 1c1c7 cell cycle and differentiated state independently of dioxin.Mol. Cell. Biol. 16, 2144–2150.
Marlowe, J.L., Puga, A., 2005. Aryl hydrocarbon receptor, cell cycle regulation,toxicity, and tumorigenesis. J. Cell. Biochem. 96, 1174–1184.
McGehee Jr., R.E., Ronis, M.J., Cowherd, R.M., Ingelman-Sundberg, M.,Badger, T.M., 1994. Characterization of cytochrome P450 2E1 induction ina rat hepatoma FGC-4 cell model by ethanol. Biochem. Pharmacol. 48,1823–1833.
Meyer, B.K., Perdew, G.H., 1999. Characterization of the AhR–hsp90–XAP2core complex and the role of the immunophilin-related protein XAP2 inAhR stabilization. Biochemistry 38, 8907–8917.
Meyer, B.K., Petrulis, J.R., Perdew, G.H., 2000. Aryl hydrocarbon (Ah) receptorlevels are selectivelymodulated by hsp90-associated immunophilin homologXAP2. Cell Stress Chaperones 5, 243–254.
Moennikes, O., Loeppen, S., Buchmann, A., Andersson, P., Ittrich, C., Poellinger,L., Schwarz, M., 2004. A constitutively active dioxin/aryl hydrocarbonreceptor promotes hepatocarcinogenesis inmice. Cancer Res. 64, 4707–4710.
Oesch-Bartlomowicz, B., Huelster, A., Wiss, O., Antoniou-Lipfert, P., Dietrich,C., Arand, M., Weiss, C., Bockamp, E., Oesch, F., 2005. Aryl hydrocarbonreceptor activation by cAMP vs. dioxin: divergent signaling pathways. Proc.Natl. Acad. Sci. U. S. A. 102, 9218–9223.
Palermo, C.M., Westlake, C.A., Gasiewicz, T.A., 2005. Epigallocatechin gallateinhibits aryl hydrocarbon receptor gene transcription through an indirectmechanism involving binding to a 90 kDa heat shock protein. Biochemistry44, 5041–5052.
Persky, V., Van Horn, L., 1995. Epidemiology of soy and cancer: perspectivesand directions. J. Nutr. 125, 709S–712S.
Petrulis, J.R., Perdew, G.H., 2002. The role of chaperone proteins in the arylhydrocarbon receptor core complex. Chem. Biol. Interact. 141, 25–40.
Poland, A., Knutson, J.C., 1982. 2,3,7,8-tetrachlorodibenzo-p-dioxin and re-lated halogenated aromatic hydrocarbons: examination of the mechanism oftoxicity. Annu. Rev. Pharmacol. Toxicol. 22, 517–554.

283R. Singhal et al. / Toxicology and Applied Pharmacology 227 (2008) 275–283
Pollenz, R.S., 2002. The mechanism of AH receptor protein down-regulation(degradation) and its impact on AH receptor-mediated gene regulation.Chem. Biol. Interact. 141, 41–61.
Pollenz, R.S., 2007. Specific blockage of ligand-induced degradation of the Ahreceptor by proteasome but not calpain inhibitors in cell culture lines fromdifferent species. Biochem. Pharmacol. 74, 131–143.
Pollenz, R.S., Popat, J., Dougherty, E.J., 2005. Role of the carboxy-terminaltransactivation domain and active transcription in the ligand-induced andligand-independent degradation of the mouse Ahb-1 receptor. Biochem.Pharmacol. 70, 1623–1633.
Puga, A., Tomlinson, C.R., Xia, Y., 2005. Ah receptor signals cross-talk withmultiple developmental pathways. Biochem. Pharmacol. 69, 199–207.
Puppala, D., Gairola, C.G., Swanson, H.I., 2007. Identification of kaempferol asan inhibitor of cigarette smoke-induced activation of the aryl hydrocarbonreceptor and cell transformation. Carcinogenesis 28, 639–647.
Reeves, P.G., Nielsen, F.H., Fahey Jr., G.C., 1993. AIN-93 purified diets forlaboratory rodents: final report of the American Institute of Nutrition ad hocwriting committee on the reformulation of the AIN-76A rodent diet. J. Nutr.123, 1939–1951.
Ronis, M.J., Rowlands, J.C., Hakkak, R., Badger, T.M., 2001. Inducibility ofhepatic CYP1A enzymes by 3-methylcholanthrene and isosafrole differs inmale rats fed diets containing casein, soy protein isolate or whey fromconception to adulthood. J. Nutr. 131, 1180–1188.
Rowlands, J.C., He, L., Hakkak, R., Ronis, M.J., Badger, T.M., 2001. Soy andwhey proteins downregulate DMBA-induced liver and mammary glandCYP1 expression in female rats. J. Nutr. 131, 3281–3287.
Santiago-Josefat, B., Fernandez-Salguero, P.M., 2003. Proteasome inhibitioninduces nuclear translocation of the dioxin receptor through an Sp1 andprotein kinase C-dependent pathway. J. Mol. Biol. 333, 249–260.
Sharp, G.B., Lagarde, F., Mizuno, T., Sauvaget, C., Fukuhara, T., Allen, N.,Suzuki, G., Tokuoka, S., 2005. Relationship of hepatocellular carcinoma tosoya food consumption: a cohort-based, case–control study in Japan. Int. J.Cancer 115, 290–295.
Simmen, R.C., Eason, R.R., Till, S.R., Chatman Jr., L., Velarde, M.C., Geng, Y.,Korourian, S., Badger, T.M., 2005. Inhibition of NMU-induced mammarytumorigenesis by dietary soy. Cancer Lett. 224, 45–52.
Singhal, R., Badger, T.M., Ronis, M.J., 2007. Reduction in 7,12-dimethylbenz[a]anthracene-induced hepatic cytochrome-P450 1A1 expression followingsoy consumption in female rats is mediated by degradation of the arylhydrocarbon receptor. J. Nutr. 137, 19–24.
Song, Z., Pollenz, R.S., 2002. Ligand-dependent and independent modulation ofaryl hydrocarbon receptor localization, degradation, and gene regulation.Mol. Pharmacol. 62, 806–816.
Tokunaga, T., 2004. Novel physiological function of fructooligosaccharides.BioFactors 21, 89–94.
Varshavsky, A., 1997. The ubiquitin system. Trends Biochem. Sci. 22,383–387.
Whitesell, L., Cook, P., 1996. Stable and specific binding of heat shock protein90 by geldanamycin disrupts glucocorticoid receptor function in intact cells.Mol. Endocrinol. 10, 705–712.
Whitesell, L., Mimnaugh, E.G., De Costa, B., Myers, C.E., Neckers, L.M.,1994. Inhibition of heat shock protein HSP90-pp60v-src heteroproteincomplex formation by benzoquinone ansamycins: essential role for stressproteins in oncogenic transformation. Proc. Natl. Acad. Sci. U. S. A. 91,8324–8328.
Xiao, R., Badger, T.M., Simmen, F.A., 2005. Dietary exposure to soy or wheyproteins alters colonic global gene expression profiles during rat colontumorigenesis. Mol. Cancer 4, 1.



![Polycyclic aromatic hydrocarbons (PAH) guidance · Polycyclic aromatic hydrocarbons (Benzo[a]pyrene) General Information Key Points Identity The term polycyclic aromatic hydrocarbons](https://img.pdfslide.us/doc/110x75/5af498db7f8b9a8d1c8c6b2b/polycyclic-aromatic-hydrocarbons-pah-guidance-aromatic-hydrocarbons-benzoapyrene.jpg)

























