Embed Size (px)
Citation preview

Rational design of new leishmanicidal agents: In silico and In vitro evaluation
S. Andrade Ochoa*,1,2, K.F. Chacón Vargas*,1, J. Correa Basurto2, L.M. Rodríguez Váldez3, G.V. Nevárez Moorillón3 and L. E. Sánchez Torres**,1
1 ENCB-IPN. Prolongación de Carpio y Plan de Ayala s/n, Col. Casco de Santo Tomas, Delegación Miguel Hidalgo, C.P. 11340, México, D.F. Mexico
2 ESM-IPN. Plan de San Luis y Díaz Mirón s/n, Col. Casco de Santo Tomas, Delegación Miguel Hidalgo, C.P. 11340, México, D.F. Mexico
3 FCQ-UACH. Campus Universitario #2 s/n, Circuito Universitario. C.P. 31125, Chihuahua, Chihuahua. Mexico * Both authors contributed equally to this work ** Corresponding author: email: [email protected]
Leishmaniasis is a public health problem caused by protozoan of the genus Leishmania. This parasitic vector-borne disease, presents different clinical manifestations including cutaneous, mucocutaneous and visceral leishmaniasis. The lack of an effective and nontoxic antiparasitic treatment highlights the need to develop new leishmanicidal agents. To address this challenge, the use of bioinformatics and molecular modelling will help on the identification of specific biological targets and to determine the structural and molecular properties that confer biological activity to specific compounds. On the other hand, biological evaluation is a critical aspect in the discovery of new chemical structures with chemotherapeutic properties, and flow cytometry has emerged as a potent tool for the realization of biological assays. The conjunction of in silico and in vitro studies provide valuable information to direct the selection of molecules for in vivo experiments, reducing costs, time and the excessive use of experimental animals. This chapter aims to discuss theoretical techniques and in vitro evaluation tools available for a rational design of new, effective and low cost leishmanicidal agents.
Keywords: Leishmanicidal Activity, Flow Cytometry, QSAR, Docking Molecular, Molecular Modeling
1. Leishmaniasis
According to the World Health Organization (WHO), Leishmaniasis is one of the main Neglected Tropical Diseases that primarily affect the poorest and marginalized populations of the world, with direct impact on the socioeconomic development of underdeveloped regions. Leishmaniasis includes a spectrum of diseases related to species of Leishmania and the host immune system. There are three main forms of manifestation of the disease: cutaneous leishmaniasis (CL) characterized by ulcers at the site of infection; mucocutaneous leishmaniasis (MCL), a progressive infection with invasion and destruction of mucosal; and visceral leishmaniasis (VL), which is the most severe form where the infection spreads to other organism, including the spleen and liver and is also known as Kala-azar [1-3]. Drugs currently used to treat Leishmaniasis are inadequate, since they are not effective and have several side effects that caused treatment abandonment. These drawbacks include a low efficacy, an excessive toxicity of the drug and an increasing resistance by parasites [4]. Therefore, many experts has urged on the need for developing new drugs to control and cure Leishmaniasis [5]. An alternative has been the use of natural-derived and synthetic compounds. Attractive targets for new Leishmaniasis drugs are molecules that participate in parasite-specific metabolic pathways. Affection of this kind of targets implies the death of the parasite and the control of the infection. Table 1 shows some interesting targets that are actually been using for the identification of new drugs against Leishmania. Pharmaceutical industry is facing the challenge of increasing productivity and innovation. The procedure for developing a new drug is too long and requires an important monetary investment; it must be remembered that for a drug to reach the market, it must go through a long process that involves synthesis or isolation, purification and several biological evaluations, both in vitro and in vivo, first in animals and eventually in humans [6]. In fact only one in 10,000 molecules tested, passes to the development phase, one of every 100,000 molecules in the development stage, succeed clinical trials and only 3 of 10 newly registered drugs, recovers its initial investment [7]. Therefore, the current problem is the selection of new molecules of the vast universe that has the potential to be biologically active but reducing time and costs. In silico methodologies and in general, rational drug design, can help on the innovation and on the optimization of time and costs required for new drugs development. In this chapter the utility of the rational design and the in vitro evaluation are exposed, in order to improve the research of new potential chemotherapeutic agents in particular as leishmanicidals. Figure 1 shows the process through which, the rational drug design can accomplish this objective.
The Battle Against Microbial Pathogens: Basic Science, Technological Advances and Educational Programs (A. Méndez-Vilas, Ed.)
© FORMATEX 2015
__________________________________________________________________________________________________________
564

Table 1 Most studied biological targets for the design of new leishmanicidals agents.
Target Species PDB Code Reference
Trypanothione Reductase L. infantum 2JK6 [8] [9]
Nicotinamidase L. infantum 3R2J [10] Phosphoglucose Isomerase L. mexicana 1Q50 [11]
Mitogen-Activated Protein Kinase L. mexicana Homology modelling [12] [13]
Pyruvate kinase L. mexicana 1PKL [10] Triosephosphate isomerase L. mexicana 2VXN [11] Pteridine reductase L. donovani PTR1 [14] DNA topoisomerase L. donovani 2B9S [15] Spermidine synthase L. donovani Homology modelling [16] N-Myristoyltransferase L. donovani 2WUU [11] Dihydrofolate reductase L. donovani Homology modelling [17]
Didydroorotate dehydrogenase L. donovani L. major
3C61 3GYE [10]
Leishmanolysin L. major 1LML [18] Farnesyl diphosphate synthase L. major 4JZB [19]
Fig. 1 Process of rational drug design for leishmanicidals.
2. Compound and Target Identification
Drugs fail in clinical trials for two reasons; for their ineffectiveness or because they are not safe. One of the most important steps in developing a new medicine is the identification of possible molecular targets -including proteins, genes and RNA-. A molecule is considered as a potential therapeutic agent if it is effective, safe, and if it meets clinical and commercial needs, especially if it is "druggable". A molecule is defined as "druggable" when by attaching to their biological target, it provokes a response that can be measured both in vitro and in vivo [20]. In the initial process, a hypothesis is generated on which an extract or compound is evaluated. As result, a compound or group of compounds considered candidates for development are selected. Advances in bioinformatics have led to a significant improvement in the identification of new potential compounds and possible biological targets. The data currently available include proteomic data, data profiling compounds, gene expression data, etc. There are extensive literature reviews that includes a wide range of natural and synthetic compounds with relevant leishmanicidal activities. Reviews conducted by Chan and Peña [21], Takahashi et al., [22] Rocha et al., [23] and Patil et al. [24] are powerful guidelines for the search and identification of new candidates, which can be subjected to computer-aided design, structural optimization, preclinical development, and if everything is successful, a clinical trial and a new drug in the market.
The Battle Against Microbial Pathogens: Basic Science, Technological Advances and Educational Programs (A. Méndez-Vilas, Ed.)
© FORMATEX 2015
__________________________________________________________________________________________________________
565

3. In Silico Target Validation: Molecular Docking Studies
Once the biological target is identified, it is necessary to proceed to validation techniques, which include the use of in vitro tools including whole animal models or molecular markers to identify and locate the biological target; in silico tools can also be used to calculate the interaction of target site with the candidate compounds. Although the use of a single technique could be valid, a multi-validation approach increases confidence in the results. In our laboratory, working with both -in vitro and in silico- systems gives us a better view of the process. A computer simulation is an adaptation of a process that is impossible or difficult to see under normal conditions [24]. Molecular modeling is a general term that covers a variety of theoretical methods and computational techniques to model or mimic the behavior of molecules, often taking advantage of the tools offered by computational chemistry [25]. Using specialized software, computational chemistry can describe molecular properties, simulate physical changes or chemical reactions of a molecule and can also determine the electronic parameters of chemical structures [26]. The advantage of in silico studies is that it can make comparisons between models, to predict the behavior of a molecule even before in vitro evaluation. Molecular docking is an in silico technique used to identify correct conformations of small molecule ligands and also to estimate the strength of the protein-ligand interaction, typically a receptor and a ligand [27,28]. The most common connection programs and softwares include Autodock [29], Autodock Vina [30], GOLD [31], and FlexX [32]. Docking of ligands that bind to a receptor through noncovalent interactions, is relatively conventional today, most of the research for new drug development, focuses on this type of prediction [33]. There are several reports in which the molecular docking to design new antiparasitic agents has been used. Ferreira et al (2010) reported the leishmanicidal activity of 3-(3,4,5-trimethoxyphenyl) propanoic acid (TMPP) against promastigotes of Leishmania amazonensis, this compound is a phenylpropane present in different plant species [34]. The mechanism of action of phenylpropanes is not entirely described, but it has been reported that some phenylpropanes, as cinnamaldehyde, has its antimicrobial action on cell walls and membranes of microorganisms [35]. It has also been reported that the main mode of action of cinnamaldehyde is through the inhibition of ergosterol biosynthesis or interaction with the molecule [36]. Ueda-Nakamura et al., (2006) showed changes in plasma membranes of Leishmania amazonensis when exposed to eugenol [37], another phenylpropane structurally similar to TMPP. This evidence shows that the search for biological target should be linked to an objective related to the integrity of cell membranes. Another example describes the interaction of TMPP and 14�-demethylase of Leishmania infantum by molecular docking. The 14�-demethylase is an enzyme involved in the biosynthetic pathway of sterols. Sterols define the membrane fluidity and permeability an also modulate the activity of membrane-bound proteins and ion channels [38]. TMPP has a binding energy of - 5.69 kcal/mol, so TMPP has the ability to interact with this biological target. In Figure 2 the 14�-demethylase reported by Hargrove et al., [39] is illustrated (PDB code = 3L4D) as well as TMPP interaction with this enzyme.
Fig. 2 Docking plot of sterol 14�-demethylase interacting with TMPP. a) Chemical structure of the enzyme sterol 14�-demethylase. b) Prediction of binding affinities of TMPP. A molecular docking study to measure the interactions between 14�-demethylase and a large number of phenylpropane as possible leishmanicidal candidates, has given important results. The main advantage of using sterol 14�-demethylase as a target, is that it has been efficiently used as antifungal agents in clinical medicine and agriculture [40]. Many of the research done for the development of new therapies for leishmaniasis includes the blind detection of compounds that have already expressed antiparasitic effects in in vitro experiments. If further studies of molecular screening alongside experimental studies were included, the time and costs for the discovery of new leishmanicidals would be reduced considerably.
The Battle Against Microbial Pathogens: Basic Science, Technological Advances and Educational Programs (A. Méndez-Vilas, Ed.)
© FORMATEX 2015
__________________________________________________________________________________________________________
566

4. In Vitro Assays
In the biological evaluation of extracts or compounds, antiparasitic activity is related to damage. This injury can be exerts inhibiting proliferation (leishmanistatic) or inducing the parasite to death (leishmanicidal). Both are useful, but the second is desirable. There are several mechanism of cell death [41], but necrosis and apoptosis are the most relevant when leishmanicidal activity is evaluated. Necrosis is considered a process that involves drastic cell changes, since it involves a severe mitochondrial dysfunction, nuclear damage and finally an increased cellular volume that affects the integrity of the cellular membrane; cytolysis occurs and an inflammatory response in the site of necrosis can be present. On the other hand apoptotic process is more organized, with a coordinated sequence of morphological and biochemical changes that dismantle the cellular structure. Characteristics of a cell dying by apoptosis include reduction in cell volume, phosphatidylserine translocation, mitochondrial alterations, chromatin condensation and fragmentation of the cell and of the genetic material. An important characteristic is the retention of the cell membrane integrity until a phagocytic cells internalize the apoptotic cell or the vesicles derived from it -called “apoptotic bodies”- so, an inflammatory process is not evoke. As apoptosis occur normally in eukaryotes to maintain homeostasis, it is often called “Programmed Cell Death (PCD)” [42,43,44]. For many years, PCD has been defined as a fundamental process in pluricellular plants and animals. Recently, the existence of PCD in unicellular organisms has been described [45,46]. In 1996, the first evidence of an apoptotic mechanism in Leishmania was described by Moreira et al. [47]. A decade later, in 2005, Wanderley et al., mentioned that during the infection, death by apoptosis in some promastigotes, promotes the survival of the rest of the population, since apoptotic cells do not induce an inflammatory response [48]. Furthermore, there are reports on the description of PCD as the mechanism of cell death induced by some leishmanicidal drugs. Table 2 summarizes some of the studies when the biological activity correlates with induction of cellular death. According to Galluzzi et at. (2009), for higher eukaryotes, “the most convenient technique to study cell death on a per-cell basis is cytofluorometry” [41]. In our experience, this technology can also be extended to prokaryotes. Cytofluorometry or flow cytometry, is an analytical method that has emerged as a potent tool for biological assays. This technique allows a rapid measurement of several parameters, in order to establish direct effects of the compounds under study on cell populations. Additionally, the use of optical and/or fluorescence microscopy allows a better characterization of the effect evaluated. Figure 3 shows a general scheme of the steps followed by our laboratory when leishmanicidal activity is evaluated.
4.1 Evaluation of metabolic inhibition
There are different methodologies to detect the in vitro leishmanistatic and leishmanicidal effect, two of them are traditional assays: direct count and colorimetric and fluorometric methods; the first is simple and cheap, but requires more time and the results is determined visually. Methods such as MTT and Alamar Blue, are also simple and inexpensive, can handle many samples and the results can be automatically recorded [62]. With these methodologies, it is possible to determine the effect of an extract or the molecule on the parasite, and if this effect is related to leishmanistatic or leishmanicidal activity; the results is related to the whole parasite population [63]. Additionally, these techniques are useful to determine CL50, CL90 values on parasites and CC50 on mammalian macrophages in order to calculate the selectivity index of the extract or molecule under investigation. These parameters are good enough to do an initial screening for selection of candidates that can follow a more detailed biological evaluation The next step is to stablish the mechanism of induced cell death. It is necessary to remember that apoptotic and necrotic cells have different characteristics that can be measured by microscopy and by flow cytometry as morphological changes, plasma membrane alteration, disruption of mitochondrial membrane potential (MMP) and DNA damage. The determination of one or a combination of these parameters, allows the identification and quantification of live, apoptotic and necrotic cells in a mixed population. It can also give valuable information on the molecular pathways activated in dying cells [64]. Specifically, in 1997, Azas et al proposed for the first time the use of flow cytometry as a sensitive and specific tool for assessing the effect of drugs on cellular functions of L. infantum promastigotes [65]. On Figure 4, the dose-response effect of the amphotericin B (Amp B) on L. mexicana is shown, as an example of all the parameters that can be evaluated for leishmanicidal potential of an extract or a molecule.
The Battle Against Microbial Pathogens: Basic Science, Technological Advances and Educational Programs (A. Méndez-Vilas, Ed.)
© FORMATEX 2015
__________________________________________________________________________________________________________
567

Table 2 Leishmanicidal evaluation in different systems and by different methodologies. Parasites Instar Compounds Source Cell death marker Fluorochrome Reference
L. donovani
Promastigotes (transgenic)
Marine sponge Haliclona exigua Extract Viability
proliferation GFP [49]
Promastigotes Cryptolepine and 11 analogues Synthetic
Traslocation PS Viability
AnnV-FITC PI [50] Cell cycle BE
MMP JC-1 Promastigotes LV82 Naphthoquinones Viability RFP [51]
Promastigotes Withanolides
Isolated from leaves Whitania somnifera
DNA fragmentation TUNEL
[52]
Cell cycle PI Traslocation PS Viability
AnnV-FITC PI
ROS H2DCFDA MMP JC-1
L. major
Promastigotes Amastigotes tansfected
Herbal drugs Extract MMP DiBAC4(3)
[53] Viability Syto 9 PI Viability intracellular GFP
Promastigotes Nitroheteroaryl-1,3,4-thiadiazole derivatives
Synthetic Traslocation PS Viability
Ann V-FITC 7-ADD [54]
DNA fragmentation TUNEL
Promastiogtes Essential oil Thymus capitellatus
Extract
Cell cycle PI
[55] Traslocation PS Viability
AnnV-FITC PI
MMP JC-1
L. infantum
Promastigotes Allopurinol Synthetic Cell proliferation CFSE
[56] Viability SYBR-14 and PI Protein synthesis FITC
Promastigotes 9-chloro and 9-amino-2-methoxyacridines
Synthetic Cell cycle PI
[57] Membrane potential DIOC5(3) MMP TMRM
Promastigotes Harmane harmine harmaline
Reagents
Cell cycle PI
[58] MMP TMRM Membrane potential DIOC5(3) Viability PI
Promastigotes Antimony sulfide nanoparticles Synthetic Translocation PS
Viability AnnV-FITC PI [59]
Promastiogtes Essential oil Thymus capitellatus
Extract
Cell cycle PI
[55] Traslocation PS Viability
AnnV-FITC PI
MMP JC-1
L. amazonensis
Promastigotes Amastigotes tansfected
Herbal drugs Extract MMP DiBAC4(3)
[53] Viability Syto 9 PI Viability intracellular GFP
Amastigotes axenic Parthenolides Extract Viability PI [60] MMP Rho 123
Promastigotes Algae Dictyota mertensii
Extract Viability PI
[61] MMP Rho 123
L. mexicana
Promastigotes Amastigotes tansfected
Herbal drugs Extract MMP DiBAC4(3)
[53] Viability Syto 9 PI Viability intracellular GFP
L. tropica Promastigotes
Essential oil Thymus capitellatus
Extract
Cell cycle PI
[55] Traslocation PS Viability
AnnV-FITC PI
MMP JC-1
The Battle Against Microbial Pathogens: Basic Science, Technological Advances and Educational Programs (A. Méndez-Vilas, Ed.)
© FORMATEX 2015
__________________________________________________________________________________________________________
568

Fig. 3 Process of in vitro evaluation for potential leishmanicidals.
4.2 Morphological changes and cell volume
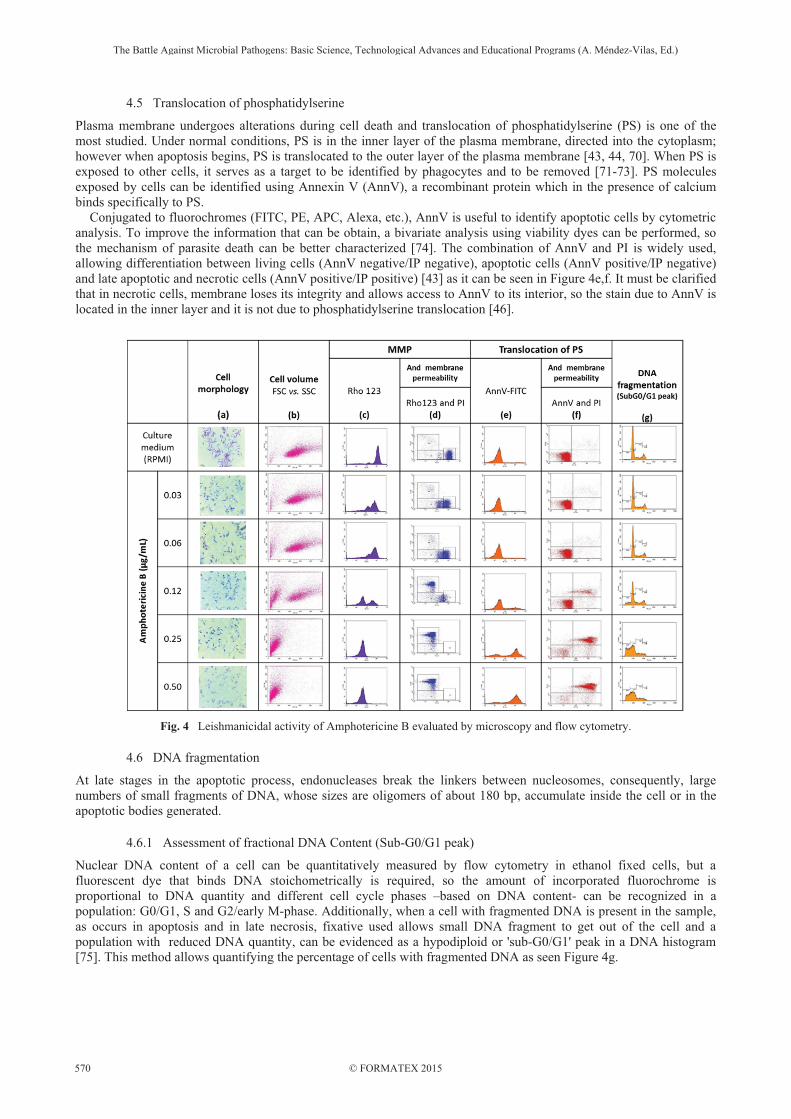
As previously described, apoptotic death in metazoa and protozoa, is characterized by shrinkage of the cell or loss of cellular volume; in the case of death by necrosis, cells suffer cytolysis and small cell fragments are evident. In the morphologic analysis done by microscopy, non-treated Giemsa stained Leishmania promastigotes are enlarged, flagellated and with a well-defined nucleus and kinetoplast; as the drug concentration increases, alteration in cell morphology can be observed. Figure 4a shows that parasites turned round, small and with enrolled flagella in response to Amp A. Although these characteristics can be evaluated by microscopy, the quantification is difficult. In flow cytometry, cells traversing through the focus of a laser beam scatter the laser light. Analysis of the scattered light provides information about the cell size and internal complexity. The cytometer can analyze volume changes by variations in the Forward Scattered (FSC) light and in internal composition by variation in Side Scatter (SSC), such that FSC correlate with cell volume and can be used to detect cellular changes. A useful tool is using a bivariate plot analysis showing the properties of FSC and SSC of the population of parasites evaluated [44]. In correlation with the microscopic evaluation, cytometric analysis shows volume and granularity diminution due to the presence of AmpA as can be noticed in Figure 4b.
4.3 Membrane permeability
Cytoplasmic membrane integrity is essential for life, so cells with irreversible membrane damage are dead, a fact that occurs early in necrosis and late in apoptosis. During the apoptotic cell death, plasma membrane integrity is preserved and most membrane functions remain unchanged [66]. Loss of the integrity of the plasma membrane can be easily demonstrated by using "viability dyes" such as propidium iodide (PI) and 7-Aminoactinomycin D (7-AAD) which are excluded by intact membranes [67,68]. Parasites with membrane integrity are PI negatives and those with membrane damage are PI positives; the evaluation of the membrane permeability can be used in combination with other probes, as discussed above, for discrimination between apoptotic and necrotic cell death.
4.4 Mitochondrial membrane potential (MMP)
In mitochondrial membranes of living cells, there is a relatively large electrical potential due to the respiratory process that generates a negative charge inside the mitochondria, this feature is also used to discriminate between competent and respiratory-deficient cells as a parameter of viability. The ability to maintain the MMP can be measured by flow cytometry using permeable cationic lipophilic fluorochrome such as Rhodamine 123 (Rho 123) and 3,3’-dipentyloxacarbocyanine iodide (DiOC5(3)). When cells are alive, they retain the fluorochrome inside the mitochondria and show a high mean fluoresce intensity (MFI). In apoptotic parasites, the MFI decreases, due to alterations in MMP and when necrotic parasites are present, the activity of their mitochondria is zero and the retention of the fluorochrome is minimum [46, 69] (Figure 4c). Simultaneous staining with viability dyes enables the distinction between live, apoptotic and necrotic parasites as show in Figure 4d.
The Battle Against Microbial Pathogens: Basic Science, Technological Advances and Educational Programs (A. Méndez-Vilas, Ed.)
© FORMATEX 2015
__________________________________________________________________________________________________________
569
4.5 Translocation of phosphatidylserine
Plasma membrane undergoes alterations during cell death and translocation of phosphatidylserine (PS) is one of the most studied. Under normal conditions, PS is in the inner layer of the plasma membrane, directed into the cytoplasm; however when apoptosis begins, PS is translocated to the outer layer of the plasma membrane [43, 44, 70]. When PS is exposed to other cells, it serves as a target to be identified by phagocytes and to be removed [71-73]. PS molecules exposed by cells can be identified using Annexin V (AnnV), a recombinant protein which in the presence of calcium binds specifically to PS. Conjugated to fluorochromes (FITC, PE, APC, Alexa, etc.), AnnV is useful to identify apoptotic cells by cytometric analysis. To improve the information that can be obtain, a bivariate analysis using viability dyes can be performed, so the mechanism of parasite death can be better characterized [74]. The combination of AnnV and PI is widely used, allowing differentiation between living cells (AnnV negative/IP negative), apoptotic cells (AnnV positive/IP negative) and late apoptotic and necrotic cells (AnnV positive/IP positive) [43] as it can be seen in Figure 4e,f. It must be clarified that in necrotic cells, membrane loses its integrity and allows access to AnnV to its interior, so the stain due to AnnV is located in the inner layer and it is not due to phosphatidylserine translocation [46].
Fig. 4 Leishmanicidal activity of Amphotericine B evaluated by microscopy and flow cytometry.
4.6 DNA fragmentation
At late stages in the apoptotic process, endonucleases break the linkers between nucleosomes, consequently, large numbers of small fragments of DNA, whose sizes are oligomers of about 180 bp, accumulate inside the cell or in the apoptotic bodies generated.
4.6.1 Assessment of fractional DNA Content (Sub-G0/G1 peak)
Nuclear DNA content of a cell can be quantitatively measured by flow cytometry in ethanol fixed cells, but a fluorescent dye that binds DNA stoichometrically is required, so the amount of incorporated fluorochrome is proportional to DNA quantity and different cell cycle phases –based on DNA content- can be recognized in a population: G0/G1, S and G2/early M-phase. Additionally, when a cell with fragmented DNA is present in the sample, as occurs in apoptosis and in late necrosis, fixative used allows small DNA fragment to get out of the cell and a population with reduced DNA quantity, can be evidenced as a hypodiploid or 'sub-G0/G1' peak in a DNA histogram [75]. This method allows quantifying the percentage of cells with fragmented DNA as seen Figure 4g.
The Battle Against Microbial Pathogens: Basic Science, Technological Advances and Educational Programs (A. Méndez-Vilas, Ed.)
© FORMATEX 2015
__________________________________________________________________________________________________________
570

4.6.2 Detection of DNA strand breaks (TUNEL Assay)
Another method by which, fragmented DNA is detected is TUNEL assay [76]. Enzyme, terminal deoxynucleotidyl transferase (Tdt), adds labelled nucleotides (for example, FITC-dUTP ) to the 5’ ends of the DNA fragments, so positive cells are easily detected by flow cytometry. In addition, DNA content can be simultaneously measure in the parasite. Once the in vitro studies have been done and results have been analyzed, the selected compounds can be further subjected to optimization or in vivo assays.
5. Optimization: QSAR Studies and Quantum Mechanic Studies
The optimization of the most promising molecules is one of the most important steps in drug development. The chemical structure of the most biologically active compounds are used as starting point for chemical modifications in order to improve their potency, selectivity and pharmacokinetic parameters. One of the tools used for system optimization are the quantitative structure-activity relationship (QSAR) studies. QSAR studies assume that there is a relationship between the biological activity of a molecule and its structure, by a simple mathematical relationship that can reproduce (and subsequently predict) a biological activity for a set of compounds, under the assumption that the activity observed is the result of the contributions of several structural and molecular factors, which act independently. To perform a QSAR study is necessary to determine the biological activity of a series of molecules, and is also necessary to know their structural (number and types of atoms, number of functional groups, molecular weight, etc.), topological (Balaban index, eccentricity, etc.), molecular (lipophilicity, molecular refractivity, total loads, etc.) and/or quantum mechanical (dipole moment, electronegativity, total energy of the system, etc.) properties that may be related to the biological activity; these properties are known as molecular descriptors or molecular indices. The basic strategy in QSAR studies is to find a mathematical equation that considers the biological activity as a function of the structure and molecular properties of candidate compounds [77]. The strategy is directed through a mathematical-statistical treatment by least squares multivariate methods and by principal component analysis applied to the descriptors, so that the biological activity of a set of compounds is adjusted to a multilinear model through a multiple regression analysis. QSAR models help on the understanding of the relationship between structure and biological activity of the compound, as well as to evaluate the biological activity of known molecules and to predict the activity of compounds even before its synthesis and / or purification. They also allow us to identify the molecular properties that have more influence on the biological activity, which is critical in rationalizing the mechanisms of action [78]. Genetic algorithms are an evolutionary method widely used for complex optimization problems in diverse scientific disciplines such as robotics, chemistry and pharmaceuticals [79]. A specific application of genetic algorithms is the variable subset selection (GA-VSS). MobyDigs is a software which extended the genetic strategy based on the evolution of a single population of models to a more complex genetic strategy based on the development of more than one population strategy, allowing to obtain statistically valid models [80]. In Figure 5 terpenes structures are observed with their inhibitory concentrations determined by our research group. Terpenes are volatile compounds present in plants to whichvarious biological properties are attributed.
Fig. 5 Chemical structure of terpenes with leishmanicidal activity. a) p-Cymene; b) �-Citronellol; c) Geranial; d) Geraniol; e) Lavandulol; f) Linallol; g) Myrcene; h) Nerol; i) Ocimene; j) 3,7-dimetiloctane The QSAR study begins with the construction and optimization of each of the molecular compounds evaluated. There are different programs for system optimization, the most popular are Gaussian [81] and Spartan [82]. Once validated the minimum potential energy of the system, molecular descriptors or indices are calculated. Structural and constitutional descriptors are those that provide us with information on chemical structure. There are several servers on the network that calculate various descriptors, the DRAGON System program [83] is a package developed by Milano Chemometrics to obtain a large number of descriptors that are based on the structure of the systems. After the descriptors are obtained, the mathematical relationship between the biological activity and descriptors calculated is obtained. Equation (1) shows the QSAR model that predicts leishmanicidal activity from structural descriptors systems; the model was obtained by genetic algorithms.
The Battle Against Microbial Pathogens: Basic Science, Technological Advances and Educational Programs (A. Méndez-Vilas, Ed.)
© FORMATEX 2015
__________________________________________________________________________________________________________
571
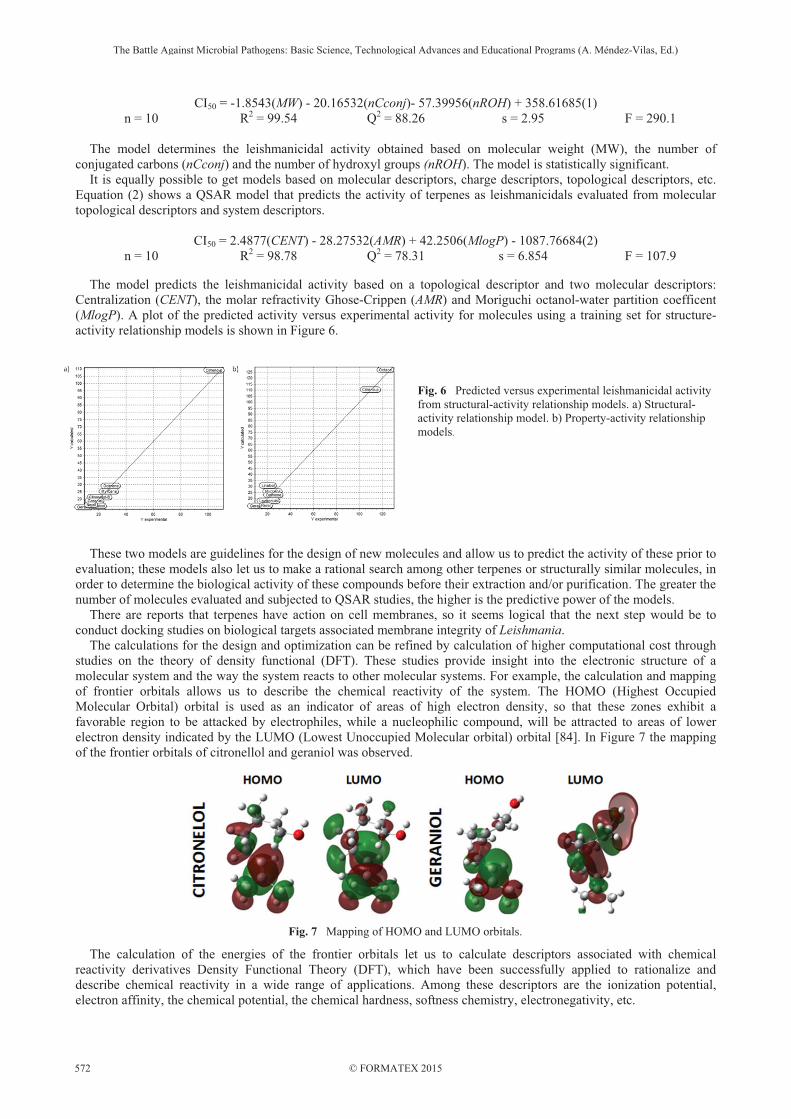
CI50 = -1.8543(MW) - 20.16532(nCconj)- 57.39956(nROH) + 358.61685(1) n = 10 R2 = 99.54 Q2 = 88.26 s = 2.95 F = 290.1
The model determines the leishmanicidal activity obtained based on molecular weight (MW), the number of conjugated carbons (nCconj) and the number of hydroxyl groups (nROH). The model is statistically significant. It is equally possible to get models based on molecular descriptors, charge descriptors, topological descriptors, etc. Equation (2) shows a QSAR model that predicts the activity of terpenes as leishmanicidals evaluated from molecular topological descriptors and system descriptors.
CI50 = 2.4877(CENT) - 28.27532(AMR) + 42.2506(MlogP) - 1087.76684 (2) n = 10 R2 = 98.78 Q2 = 78.31 s = 6.854 F = 107.9
The model predicts the leishmanicidal activity based on a topological descriptor and two molecular descriptors: Centralization (CENT), the molar refractivity Ghose-Crippen (AMR) and Moriguchi octanol-water partition coefficent (MlogP). A plot of the predicted activity versus experimental activity for molecules using a training set for structure-activity relationship models is shown in Figure 6.
Fig. 6 Predicted versus experimental leishmanicidal activity from structural-activity relationship models. a) Structural-activity relationship model. b) Property-activity relationship models.
These two models are guidelines for the design of new molecules and allow us to predict the activity of these prior to evaluation; these models also let us to make a rational search among other terpenes or structurally similar molecules, in order to determine the biological activity of these compounds before their extraction and/or purification. The greater the number of molecules evaluated and subjected to QSAR studies, the higher is the predictive power of the models. There are reports that terpenes have action on cell membranes, so it seems logical that the next step would be to conduct docking studies on biological targets associated membrane integrity of Leishmania. The calculations for the design and optimization can be refined by calculation of higher computational cost through studies on the theory of density functional (DFT). These studies provide insight into the electronic structure of a molecular system and the way the system reacts to other molecular systems. For example, the calculation and mapping of frontier orbitals allows us to describe the chemical reactivity of the system. The HOMO (Highest Occupied Molecular Orbital) orbital is used as an indicator of areas of high electron density, so that these zones exhibit a favorable region to be attacked by electrophiles, while a nucleophilic compound, will be attracted to areas of lower electron density indicated by the LUMO (Lowest Unoccupied Molecular orbital) orbital [84]. In Figure 7 the mapping of the frontier orbitals of citronellol and geraniol was observed.
Fig. 7 Mapping of HOMO and LUMO orbitals.
The calculation of the energies of the frontier orbitals let us to calculate descriptors associated with chemical reactivity derivatives Density Functional Theory (DFT), which have been successfully applied to rationalize and describe chemical reactivity in a wide range of applications. Among these descriptors are the ionization potential, electron affinity, the chemical potential, the chemical hardness, softness chemistry, electronegativity, etc.
The Battle Against Microbial Pathogens: Basic Science, Technological Advances and Educational Programs (A. Méndez-Vilas, Ed.)
© FORMATEX 2015
__________________________________________________________________________________________________________
572

Finally, the association of all these theoretical and experimental procedures improves the search and evaluation of new potential drug with leishmanicidal activity, reducing time and cost, besides facilitating the understanding of the possible mechanisms of action.
References [1] PAHO. Guía para el desarrollo de planes integrados de acción para la prevención, control y eliminación de las enfermedades
infecciosas desantendidas. Pan American Health Organization. Washington DC. 2011. p. 12-20. [2] Kayser O, Kiderlen AF, Croft SL, Koch-Institut R. Natural Products as potential antiparasitic drugs. Stud Nat Prod Chem.
2002; 26: 779–848. [3] WHO (World Health Organization). (2014). Health topics. Retrieved from http://www.who.int/topics/en/ [4] Bezerra de Menezes JP, Sampaio Guedes CE, Oliverida AL, Bittencourt MB, Sampaio TP. Advances in Development of New
Treatment for Leishmaniasis. BioMed Res Int. 2015;11. [5] CORDIS. Mejora de los fármacos contra las infecciones por tripanosomáticos. [Internet] Community Research and
Development Information Service. European Commission 2013. Avalaible from: http://cordis.europa.eu/news/rcn/36486_es.html
[6] San Román, L. Desarrollo de nuevos fármacos: desde la invención a la Farmacia. España: Universidad de Salamanca. 2013. p 13-20
[7] Kolb HC, Finn MG, Sharpless KB. Click chemistry: diverse chemical function from a few good reactions. Angew Chem Int Ed. 2001; 40(11):2004-2021.
[8] Shukla AK, Patra S, Dubey VK. Evaluation of selected antitumor agents as subversive substrate and potential inhibitor of trypanothione reductase: an alternative approach for chemotherapy of Leishmaniasis. Mol Cell Biochem, 2011; 352(1-2):261-70.
[9] Venkatesan SK, Shukla AK, Dubey VK. Molecular docking studies of selected tricyclic and quinone derivatives on trypanothione reductase of Leishmania infantum. J Comput Chem. 2010; 31(13):2463-75.
[10] Ogungbe IV, Erwin WR, Setzer WN. Antileishmanial phytochemical phenolics: molecular docking to potential protein targets. J Mol Graphics Modell. 2014; 48:105-117.
[11] Ogungbe IV, Setzer WN. In-silico Leishmania target selectivity of antiparasitic terpenoids. Molecules. 2013; 18(7):7761-7847. [12] Awale M, Kumar V, Saravanan P, Mohan CG. Homology modeling and atomic level binding study of Leishmania MAPK with
inhibitors. J Mol Model. 2010; 16(3): 475-88. [13] Saravanan P, Venkatesan SK, Mohan CG, Patra S, Dubey VK. Mitogen-activated protein kinase 4 of Leishmania parasite as a
therapeutic target. Eur J Med Chem. 2010; 45(12):5662-70. [14] Kumar P, Kumar A, Verma SS, Dwivedi N, Singh N, Siddiqi MI, Singh N. Leishmania donovani pteridine reductase 1:
biochemical properties and structure-modeling studies. Exp Parasitol. 2008; 120(1):73-79. [15] Misra P, Khaliq T, Dixit A, SenGupta S, Samant M, Kumari S, Dube A. Antileishmanial activity mediated by apoptosis and
structure-based target study of peganine hydrochloride dihydrate: an approach for rational drug design. J Antimicrob Chemother. 2008; 62(5): 998-1002.
[16] Grover A, Katiyar SP, Singh SK, Dubey VK, Sundar D. A leishmaniasis study: Structure-based screening and molecular dynamics mechanistic analysis for discovering potent inhibitors of spermidine synthase. BBA-Proteins Proteom. 2012; 1824(12): 1476-83.
[17] Maganti L, Manoharan P, Ghoshal N. Probing the structure of Leishmania donovani chagasi DHFR-TS: comparative protein modeling and protein–ligand interaction studies. J Mol Model. 2010; 16(9):1539-47.
[18] Shaukat A, Mirza HM, Ansari AH, Yasinzai M, Zaidi SZ, Dilshad S, Ansari FL. Benzimidazole derivatives: synthesis, leishmanicidal effectiveness, and molecular docking studies. Med Chem Res. 2013; 22(8): 3606-20.
[19] Mukherjee P, Desai PV, Srivastava A, Tekwani BL, Avery MA. Probing the structures of leishmanial farnesyl pyrophosphate synthases: Homology modeling and docking studies. J Chem Inf Model. 2008; 48(5):1026-40.
[20] Hughes JP, Rees S, Kalindjian SB, Philpott KL. Principles of early drug discovery. Br J Pharmacol. 2011; 162(6):1239-49. [21] Chan-Bacab MJ, Peña-Rodríguez LM. Plant natural products with leishmanicidal activity. Nat Prod Rep. 2001; 18(6):674-88. [22] Takahashi M, Fuchino H, Sekita S, Satake M. In vitro leishmanicidal activity of some scarce natural products. Phytor Res.
2004; 18(7): 573-78. [23] Rocha LG, Almeida JRGS, Macedo RO, Barbosa-Filho JM. A review of natural products with antileishmanial
activity.Phytomedicine. 2005; 12(6):514-35. [24] Patil RS, Patil MS, Kshirsagar SS, Chaudhari PS, Bayas JP, Oswal RJ. Synthetic and natural products against leishmaniasis: A
review. World J Public Health Sc. 2012; 1:7-22. [25] Ren X, Rinke P, Joas C, Scheffler M. Random-phase approximation and its applications in computational chemistry and
materials science. J Mater Sci. 2012; 47(21):7447-71. [26] Lipinski CA, Lombardo F, Dominy BW, Feeney PJ. Experimental and computational approaches to estimate solubility and
permeability in drug discovery and development settings. Adv Drug Deliv Rev. 2012; 64:4-17. [27] Feller D. The role of databases in support of computational chemistry calculations. J Comput Chem. 1996;17(13):1571-86. [28] Yuriev E, Agostino M, Ramsland PA. Challenges and advances in computational docking: 2009 in review. J Mol Recognit.
2011; 24(2):149-164. [29] Kumalo HM, Bhakat S, Soliman ME. Theory and applications of covalent docking in drug discovery: merits and
pitfalls. Molecules. 2015; 20(2): 1984-2000. [30] Morris GM, Huey R, Lindstrom W, Sanner MF, Belew RK, Goodsell DS, Olson AJ. AutoDock4 and AutoDockTools4:
Automated Docking with Selective Receptor Flexibility. J Comput Chem. 2009; 30:2785–91.
The Battle Against Microbial Pathogens: Basic Science, Technological Advances and Educational Programs (A. Méndez-Vilas, Ed.)
© FORMATEX 2015
__________________________________________________________________________________________________________
573

[31] Trott O, Olson AJ. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J Comput Chem. 2010; 31:455–61.
[32] Jones G, Willett P, Glen RC, Leach AR, Taylor R. Development and validation of a genetic algorithm for flexible docking. J Mol Biol. 1997; 267:727–48.
[33] Rarey M, Kramer B, Lengauer T, Klebe G. A fast flexible docking method using an incremental construction algorithm. J Mol Biol. 1996; 261:470–89.
[34] Yuriev E, Ramsland PA. Latest developments in molecular docking: 2010–2011 in review. J Mol Recognit. 2013; 26:215–39. [35] Ferreira MGP, Kayano AM, Silva-Jardim I, Silva TOD, Zuliani JP, Facundo, VA, Stábeli RG. Antileishmanial activity of 3-(3,
4, 5-trimethoxyphenyl) propanoic acid purified from Amazonian Piper tuberculatum Jacq, Piperaceae, fruits. Rev Bras Farmacogn. 2010; 20(6):1003-06.
[36] Bang KH, Lee DW, Park HM, Rhee YH. Inhibition of fungal cell wall synthesizing enzymes by trans-cinnamaldehyde. Biosci, Biotechnol, Biochem. 2000; 64(5):1061-63.
[36] Shreaz S, Sheikh RA, Rimple B, Hashmi AA, Nikhat M, Khan LA. Anticandidal activity of cinnamaldehyde, its ligand and Ni (II) complex: effect of increase in ring and side chain. Microb Pathog. 2010; 49(3):75-82.
[37] Ueda-Nakamura T, Mendonça-Filho RR, Morgado-Díaz JA, Maza PK, Dias Filho BP, Cortez DAG, Nakamura CV. Antileishmanial activity of Eugenol-rich essential oil from Ocimum gratissimum. Parasitol Inter. 2006; 55(2): 99-105.
[38] Lepesheva GI, Waterman MR. Sterol 14alpha-demethylase cytochrome P450 (CYP51), a P450 in all biological kingdoms. Biochim Biophys Acta. 2007; 1770(3): 467–77.
[39] Hargrove TY, Wawrzak Z, Liu J, Nes WD, Waterman MR, Lepesheva GI. Substrate preferences and catalytic parameters determined by structural characteristics of sterol 14�-demethylase (CYP51) from Leishmania infantum. J BiolChem. 2011; 286(30):26838-26848
[40] Zonios DI, Bennet JE. Update on azole antifungals. Semin Respir Crit Care Med. 2008; 29:198–210. [41] Galluzzi L, Aaronson SA, Abrams J, Alnemri ES, Andrews DW, Baehrecke EH, Bazan HG, Blagosklonny MV, Blomgren K,
Borner C, Bredesen DE, Brenner C, Castedo M, Cidlowsk JA, Ciechanover A, Cohen GM, Laurenzi V, De Maria R, Deshmukh M, Dynlacht BD, El-Deiry WS, Flavell RA, Fulda S, Garrido C, Golstein P, Gougeon ML, Green DR, Gronemeyer H, Hajnóczky G, Hardwick JM, Hengartner MO, Ichijo M, Jäättelä M, Kepp O, Kimchi A, Klionsky DJ, Knight RA, Kornbluth S, Kumar S, Levine B, Lipton SA, Lugli E, Madeo F, Malorni W, Marine JCW, Martin SJ, Medema JP, Mehlen P, Melino G, Moll UM, Morselli E, Nagata S, Nicholson DW, Nicotera P, Nuñez G, Oren M, Penninger J, Pervaiz S, Peter ME, Piacentini M, Prehn JM, Puthalakath H, G, Rabinovich A, Rizzuto R, Rodrigues CMP, Rubinsztein DC, Rudel T, Scorrano L, SimonHU, Steller H, Tschopp J, Tsujimoto Y, Vandenabeele P, Vitale I, Vousden KH, Youle RJ, Yuan j, Zhivotovsky B and Kroemer G. Guidelines for the use and interpretation of assays for monitoring cell death in higher eukaryotes. Cell Death Differ, 2009; 16(8):1093-1107.
[42] Fink SL, Cookson BT. Apoptosis, pyroptosis, and necrosis: mechanistic description of dead and dying eukaryotic cells. Infect Immunol. 2005; 73(4):1907-16.
[43] Martínez SM. Técnicas para la detección de apoptosis y scenescencia celular in vitro y su importancia en biotecnología de la salud. Rev Colom de Biotecnol. 2009; 2:152-166.
[44] Jiménez-Ruiz A, Alzate JF, MacLeod ET, Luder CG, Fasel N, Hurd H. Apoptotic markers in protozoan parasites. Parasit Vectors. 2010; 3(104):b48.
[45] Lee N, Bertholet S, Debrabant A, Muller J, Duncan R, Nakhasi HL. Programmed cell death in the unicellular protozoan parasite Leishmania. Cell Death Differ. 2002; 9(1):53-64.
[46] Proto WR, Coombs GH, Mottram JC. Cell death in parasitic protozoa: regulated or incidental?. Nature Rev Microbiol. 2013; 11(1): 58-66.
[47] Moreira MEC, Del Portillo HA, Milder RV, Balanco JMF, Barcinski MA. Heat shock induction of apoptosis in promastigotes of the unicellular organism Leishmania (Leishmania) amazonensis. J Cell Physiol. 1996; 167(2):305-13.
[48] Wanderley JL, M., Benjamin A, Real F, Bonomo A, Moreira MEC, Barcinski MA. Apoptotic mimicry: an altruistic behavior in host/Leishmania interplay. Braz J Med Biol Res. 2005; 38(6):807-12.
[49] Dube A, Singh N, Saxena A, Lakshmi V. Antileishmanial potential of a marine sponge, Haliclona exigua (Kirkpatrick) against experimental visceral leishmaniasis. Parasitol Res. 2007; 101(2):317–24.
[50] Hazra S, Ghosh S, Debnath S, Seville S, Prajapati VK, Wright C. Wright C, Hazra, B. Antileishmanial activity of cryptolepine analogues and apoptotic effects of 2,7-dibromocryptolepine against Leishmania donovani promastigotes. Parasitol Research. 2012; 111(1):195–203.
[51] Lezama-Dávila CM, Isaac-Márquez AP, Kapadia G, Owens K, Oghumu S, Stephen B, Satoskar AR. Leishmanicidal activity of two naphthoquinones against L. donovani. Biol Pharm Bull, 2012; 35(10):1761–64.
[52] Chandrasekaran S, Dayakar A, Veronica J, Sundar S, Maurya R. An in vitro study of apoptotic like death in Leishmania donovani promastigotes by withanolides. Parasitol Inter. 2013; 62(3):253–61.
[53] Plock A. Sokolowska-Köhler W, Presber W. Application of flow cytometry and microscopical methods to characterize the effect of herbal drugs on Leishmania spp. Exp Parasitol. 2001; 97(3):141–53.
[54] Ardestani SK, Poorrajab F, Razmi S, Foroumadi A, Ajdary S, Gharegozlou B, Behrouzi-Fardmoghadam M, Shafiee A. Cell death features induced in Leishmania major by 1,3,4-thiadiazole derivatives. Exp Parasitol. 2012; 132(2):116–22.
[55] Machado M, Dinis AM, Santos-Rosa M, Alves V, Salgueiro L, Cavaleiro C, Sousa MC. Activity of Thymus capitellatus volatile extract, 1,8-cineole and borneol against Leishmania species. Vet Parasitol, 2014; 200(1-2): 39–49.
[56] Kamau SW, Hurtado M, Müller-Doblies UU, Grimm F, Nunez R. Flow cytometric assessment of allopurinol susceptibility in Leishmania infantum promastigote. Cytometry. 2000; 40(4):353–60
[57] Di Giorgio C, Delmas F, Filloux N, Robin M, Seferian L, Azas N. Gasquet M, Costa M. Rimon.David P, Galy JP. In Vitro Activities of 7-Substituted 9-Chloro and 9-Amino-2-Methoxyacridines and Their Bis- and Tetra-Acridine Complexes against Leishmania infantum In Vitro Activities of 7-Substituted 9-Chloro and 9-Amino-2-Methoxyacridines and Their Bis- and Tetra-Acri. Antimicrob Agents Ch. 2003; 47(1):174–180.
The Battle Against Microbial Pathogens: Basic Science, Technological Advances and Educational Programs (A. Méndez-Vilas, Ed.)
© FORMATEX 2015
__________________________________________________________________________________________________________
574

[58] Di Giorgio C, Delmas F, Ollivier E, Elias R, Balansard G, Timon-David P. In vitro activity of the beta-carboline alkaloids harmane, harmine, and harmaline toward parasites of the species Leishmania infantum. Exp Parasitol. 2004; 106(3-4):67–74.
[59] Soflaei S, Dalimi A, Ghaffarifar F, Shakibaie M, Shahverdi AR, Shafiepour M. In Vitro Antiparasitic and Apoptotic Effects of Antimony Sulfide Nanoparticles on Leishmania infantum. J Parasitol Res. 2012; 2012
[60] Tiuman TS, Ueda-Nakamura T, Alonso A, Nakamura CV. Cell death in amastigote forms of Leishmania amazonensis induced by parthenolide. BMC Microbiol. 2014; 14(152).
[61] Dos Santos Aliança AS, dos Anjos KFL, de Vasconcelos Reis TN, Higino TMM, Brelaz-de-Castro MCA, Bianco ÉM, de Figueiredo RCBQ. The in vitro biological activity of the Brazilian brown seaweed Dictyota mertensii against Leishmania amazonensis. Molecules. 2014; 19(9):14052–65.
[62] Fumarola L, Spinelli R, Brandonisio O. In vitro assays for evaluation of drug activity against Leishmania spp. Res in Microbiol. 2004; 155:224–30.
[63] Macey MG. Flow Cytometry: Principles and Applications. 3rd ed.New Jersey: Humana Press; 2007. 230-31 [64] Wlodkowic D, Skommer J, Darzynkiewicz Z. Flow Cytometry-Based Apoptosis Detection. In Erhardt P, Toth A, Apoptosis
Methods and Protocols. New York: Humana Press; 2009. p. 19-22 [65] Azas N, Di Giorgio C, Delmas F, Gasquet M. Timon-David P. Leishmania infantum Promastigotes: Flow Cytometry as a
Possible Tool for Assessing the Effects of Drugs on Cellular Functions. Exp Parasitol. 1997; 87:1–7. [66] Shapiro HM. Practical Flow Cytometry 4th ed. New Jersey: Wiley-Liss, Hoboken, 2003. p. 294-98. [67] Darzynkiewicz Z, Bruno S, Del Bino G, Gorczyca W, Holtz MA, Lassota P, Traganos F. Features of apoptotic cells measured
by flow cytometry. Cytometry. 1992; 13:795–808. [68] Ormerod, M. Chapter 9: Cell Death, Including Apoptosis. In Ormerod MG, Flow Cytometry - A Basic Introduction. Oxford:
IRL Press, Oxford University Press. 2008. [69] Fouchet P, Jayat C, Héchard Y, Ratinaud MH, Frelat G. Recent advances of flow cytometry in fundamental and applied
microbiology. Biol. Cell. 1993; 78(1�2):95-109 [70] Fadok VA, Voelker DR, Campbell PA, Cohen JJ, Bratton DL, Henson PM. Exposure of phosphatidylserine on the surface of
apoptotic lymphocytes triggers specific recognition and removal by macrophages. J Immunol. 1992; 148(7):2207-16. [71] Wanderley JL, Pinto da Silva L, Deolindo P, Soong L, Borges VM, Prates DB, Almeida de Souza AP, Aldina B, Cunha do
Nascimiento MT, Saraiva EM, Barcinski MA. Cooperation between apoptotic and viable metacyclics enhances the pathogenesis of Leishmaniasis. PLoS One. 2009:4(5):e5733.
[72] Wanderley JLM, Barcinski MA. Apoptosis and apoptotic mimicry: the Leishmania connection. Cell Mol Life Sci. 2010; 67(10): 1653-1659.
[73] El-Hani, CN, Borges VM, Wanderley JL, Barcinski MA Apoptosis and apoptotic mimicry in Leishmania: an evolutionary perspective. Front Cell Infect Microbiol. 2012; 2(96).
[74] Vermes I, Haanen C, Steffens-Nakken H, Reutellingsperger C. A novel assay for apoptosis flow cytometric detection of phosphatidylserine expression on early apoptotic cells using fluorescein labelled annexin V. J Immunol Methods. 1995; 184(1):39-51.
[75] Gong JP, Traganos F, Darzynkiewicz Z. A selective procedure for DNA extraction from apoptotic cells applicable for gel electrophoresis and flow cytometry. Anal Biochem. 1994; 218(2):314-19.
[76] Gavrieli Y, Sherman Y, Ben-Sasson SA. Identification of programmed cell death in situ via specific labeling of nuclear DNA fragmentation. J Cell Biol. 1992; 119(3):493–501.
[77] Hansch C, Steinmetz WE, Leo AJ, Mekapati SB, Kurup A, Hoekman D. On the role of polarizability in chemical-biological interactions. J Chem Inf Comput Sci. 2003; 43(1):120-25.
[78] Mauri A, Consonni V, Pavan M, Todeschini R. Dragon software: An easy approach to molecular descriptor calculations. Match. 2006; 56(2):237-48.
[79] Wehrens R, Buydens LM. Evolutionary optimisation: a tutorial. TRAC-Trend Anal Chem. 1998; 17(4):193-203. [80] Todeschini R, Consonni V, Mauri A, Pavan M. MobyDigs: software for regression and classification models by genetic
algorithms. Data Handling in Science and Technology. 2003; 23:141-67. [81] Frisch M. J. et al. Gaussian 03, Revision D.01 ed., Gaussian, Inc., 2004. Wallingford, CT. [82] Agarwal M, Frank M. Spartan: A software tool for parallelization bottleneck analysis. In Multicore Software Engineering,
2009: IWMSE'09. ICSE Workshop on (56-63). IEEE. [83] Talete srl. Dragon for Windows (software for Molecular Descriptor Calculations). 2006: Version 5.4, http://www.talete.mi.it/ [84] Kong J, White CA, Krylov AI, Sherrill D, Adamson RD, Furlani TR, Pople JA. Q�Chem 2.0: a high�performance ab initio
electronic structure program package. J Comp Chem. 2000; 21(16):1532-48.
The Battle Against Microbial Pathogens: Basic Science, Technological Advances and Educational Programs (A. Méndez-Vilas, Ed.)
© FORMATEX 2015
__________________________________________________________________________________________________________
575





























