Embed Size (px)
Citation preview

Insect Biochem., 1976, Vol. 6, pp. 631 to 635. Pergamon Press. Printed in Great Britain
RATE OF rRNA SYNTHESIS DURING METAMORPHOSIS OF THE BLOWFLY, CALLIPHORA VICINA
ALBERTO PROTZEL and LEo LEVErOOOK
Laboratory of Physical Biology, National Institute of Arthritis, Metabolism and Digestive Diseases, National Institutes of Health, Bethesda, MD. 20014, U.S.A.
(Received 7 June 1976)
A~traet--The rate of rRNA biosynthesis during metamorphosis of the blowfly Calliphora vicina has been determined by measuring the short-term incorporation of injected 3H-uridine into rRNA, and the specific activity of the soluble (precursor) UTP pool. The highest rate of rRNA synthesis (106 #g rRNA/insect/day) occurs in the mature larva. This rate declines during early pharate adult development to become approximately constant at 46 #g/insect/day up to adult emergence. The good agreement between the rate of rRNA synthesis and the previously determined rate of degradation is consistent with the observed constancy of the rRNA pool size during pharate adult development.
I N T R O D U C T I O N
IN A RELATED paper (PROTZEL et al., 1976) we measured a decrease in ribosomal RNA (rRNA) sped- tic activity during metamorphosis of C. vicina follow- ing long-term labeling. In these experiments the rRNA was maximally labeled at the start of the exper- iments by raising larvae on a semi-defined diet con- taining labeled uridine.
As previously noted (SRrDrt~,A and LEVE~mOOK, 1974) and confirmed below, the rRNA pool remains relatively constant during most of pharate adult de- velopment of this insect. Thus, the observed decrease in rRNA specific activity is due to de novo rRNA synthesis from either unlabeled precursors, or precur- sors of lower specific activity than that of the rRNA. The former case (which is in fact hypothetical), leads to a rRNA half-life of 8.5 days, the latter, (based on the measured U T P pool specific activity), to a half-life of 2.2 days. This lower value was used to calculate that about 31% of the rRNA pool is replaced per insect per day during pharate adult development.
The present experiments were designed to obtain a more direct estimate of the rate(s) of rRNA syn- thesis during metamorphosis of C. vicina. Unlike the long term labeling technique previously employed, we have now measured the appearance of label in rRNA following injection (i.e. a 'pulse') of labeled precursor. The rates determined by the two types of experiments are shown to be in good agreement.
MATERIALS AND M E T H O D S
Rearing methods for blowflies, the chemicals used and the experimental procedures except for those described below are given by PROTZEL et al. (1976).
For short term labeling of the insect rRNA, CO2 anaes- thetized 3rd instar larvae or chilled pharate adult stages were individually injected with 1.0/d containing about
0.5 #Ci of either 14C-uridine (496 mCi/m-mole) or 3H-uri- dine (26.4Ci/m-mole). At selected times after injection, RNA was extracted from individual insects by a procedure (PRorZEL et al., 1976) modified from that of Greenberg (1969). Labeled rRNA and heterogeneous nuclear RNA (HnRNA) were then isolated together by passage of the RNA extract through a Sephadex G-200 column as pre- viously described (PRorZEL et al., 1976). Of the total radioactivity thus obtained in the void vol, the counts per- taining to rRNA were independently determined, as fol- lows.
Pharate adults aged 3 and 5 days post pupariation were individually injected with about 1.0 gCi of 14C-uridine. At intervals of 1, 2, and 3 hr after injection, rRNA and HnRNA was extracted from groups of three insects as de- scribed above. Carrier E. coil tRNA (kindly donated by Dr. J. Seidman) was added to the void volume at a concen- tration of 3.0 O.D.26o units per ml, and the RNA precipi- tated with 2.5 vol. (v/v) of ethanol at -21°C overnight. The precipitated RNA was then analysed by polyacryla- mide gel electrophoresis (PAGE) according to LOENING (1969), and the distribution of radioactivity along the gel determined. A typical 2hr pattern is depicted in Fig. 1. The fraction of void vol counts in 26S and 18S RNA was then measured according to GIRARD et al. (1965). Three correction factors for the time sequence were thus obtained, and used to correct total void vol counts to rRNA counts in the individual experimental insects.
To assess the recovery of 1"C in certain experiments, and to measure the 14C-labeled rRNA pool size by isotopic dilution (cf. HIRscn, 1967), tritiated insect rRNA was added to the initial extraction medium. The 3H-rRNA (660 dis/min/#g) was prepared from ribosomes isolated from 200 larvae raised on a semi-defined diet (Crmr~ and LEV~mOOK, 1966), containing 150/~Ci 5-3H-uridine in 75ml of medium. This procedure for estimating the amount of rRNA was judged to be more accurate that the analytical procedure previously employed (SRIDrt~tA and LEV~NaOOK, 1974).
Injection of labeled uridine leads to the appearance of both labeled UMP and CMP in the insect rRNA (PROTZEL et al., 1976). For a proper interpretation of the kinetic data
631
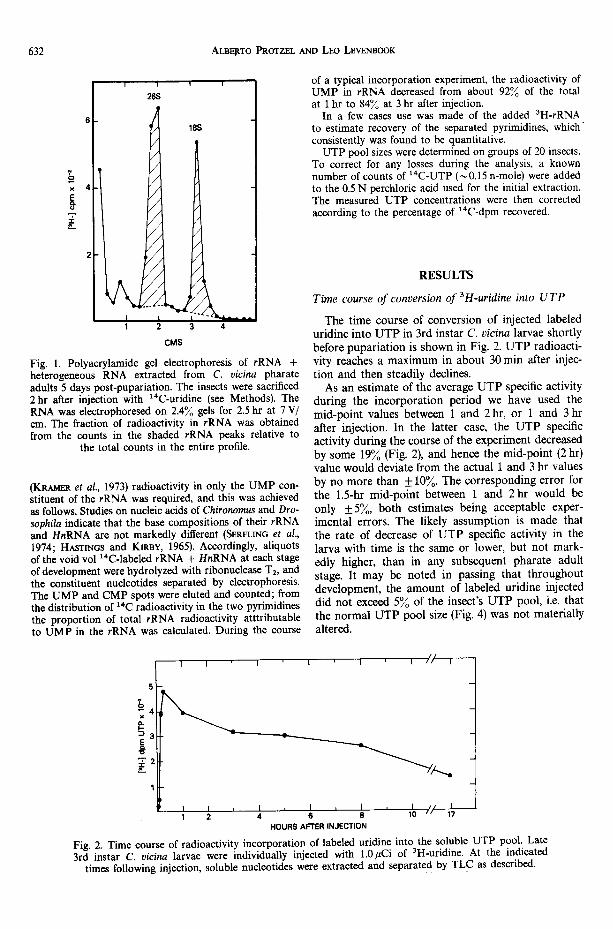
6 3 2 ALBEp.TO P R O T Z E L A N D L E O L E V E N B O O K
x 4 E= o .-r
I ] i i
2 6 S
18s
I 1
|
CMS
4
Fig. 1. Polyacrylamide gel electrophoresis of rRNA + heterogeneous RNA extracted from C. vicina pharate adults 5 days post-pupariation. The insects were sacrificed 2 hr after injection with ~4C-uridine (see Methods). The RNA was electrophoresed on 2.4% gels for 2.5 hr at 7 V/ cm. The fraction of radioactivity in rRNA was obtained from the counts in the shaded rRNA peaks relative to
the total counts in the entire profile.
(KRA~mR et al., 1973) radioactivity in only the UMP con- stituent of the rRNA was required, and this was achieved as follows. Studies on nucleic acids of Chironomus and Dro- sophila indicate that the base compositions of their rRNA and HnRNA are not markedly different (SERFLIr~G et al., 1974; HASTINGS and KIRBY, 1965). Accordingly, aliquots of the void vol ~4C-labeled rRNA + HnRNA at each stage of development were hydrolyzed with ribonuclease T2, and the constituent nucleotides separated by electrophoresis. The UMP and CMP spots were eluted and counted; from the distribution of t4C radioactivity in the two pyrimidines the proportion of total rRNA radioactivity atttributable to UMP in the rRNA was calculated. During the course
of a typical incorporation experiment, the radioactivity of UMP in rRNA decreased from about 92% of the total at 1 hr to 84% at 3 hr after injection.
In a few cases use was made of the added 3H-rRNA to estimate recovery of the separated pyrimidines, which" consistently was found to be quantitative.
UTP pool sizes were determined on groups of 20 insects. To correct for any losses during the analysis, a known number of counts of t4C-UTP (~0.15 n-mole) were added to the 0.5 N perchloric acid used for the initial extraction. The measured UTP concentrations were then corrected according to the percentage of 14C-dpm recovered.
RESULTS
Time course o f conversion o f 3H-uridine into U T P
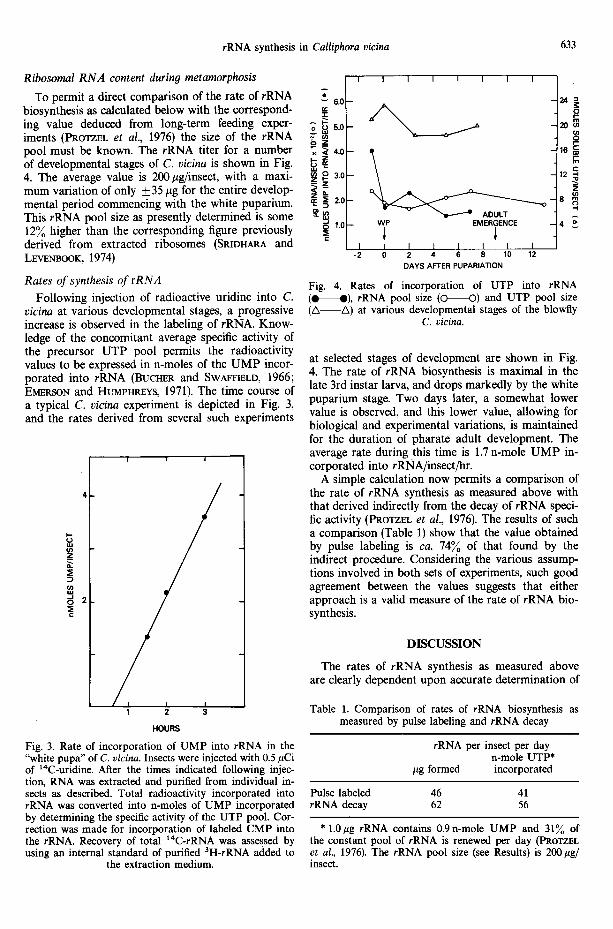
The time course of conversion of injected labeled uridine into U T P in 3rd instar C. vicina larvae shortly before pupariation is shown in Fig. 2. U T P radioacti- vity reaches a maximum in about 30 min after injec- tion and then steadily declines.
As an estimate of the average U T P specific activity during the incorporation period we have used the mid-point values between 1 and 2 hr, or 1 and 3 hr after injection. In the latter case, the U T P specific activity during the course of the experiment decreased by some 19% (Fig. 2), and hence the mid-point (2 hr) value would deviate from the actual 1 and 3 hr values by no more than ___ 10%. The corresponding error for the 1.5-hr mid-point between 1 and 2 hr would be only +__5%, both estimates being acceptable exper- imental errors. The likely assumption is made that the rate of decrease of U T P specific activity in the larva with time is the same or lower, but not mark- edly higher, than in any subsequent pharate adult stage. It may be noted in passing that throughout development, the amount of labeled uridine injected did not exceed 5% of the insect's U T P pool, i.e. that the normal U T P pool size (Fig. 4) was not materially altered.
sl ~ 4 x
53 E= "o
1
I I I I I I / / /
I I , I , I , I , I 0 / / I 1 2 4 6 s 1 17
HOURS AFTER INJECTION
Fig. 2. Time course of radioactivity incorporation of labeled uridine into the soluble UTP pool. Late 3rd instar C. vicina larvae were individually injected with 1.0#Ci of 3H-uridine. At the indicated
times following injection, soluble nucleotides were extracted and separated by TLC as described.
rRNA synthesis in Calliphora vicina 633
Ribosomal R N A content during metamorphosis
To permit a direct comparison of the rate of rRNA biosynthesis as calculated below with the correspond- ing value deduced from long-term feeding exper- iments (PROTZEL e t al., 1976) the size of the rRNA pool must be known. The rRNA titer for a number of developmental stages of C. vicina is shown in Fig. 4. The average value is 200 #g/insect, with a maxi- mum variation of only + 35 #g for the entire develop- mental period commencing with the white puparium. This rRNA pool size as presently determined is some 12~o higher than the corresponding figure previously derived from extracted ribosomes (SRIDHARA and LEVE~mOOK, 1974)
Rates of synthesis of rRNA
Following injection of radioactive uridine into C. vicina at various developmental stages, a progressive increase is observed in the labeling of rRNA. Know- ledge of the concomitant average specific activity of the precursor UTP pool permits the radioactivity values to be expressed in n-moles of the UMP incor- porated into rRNA (BucI-mR and SWAFFIELD, 1966; EMERSON and HUMPHREYS, 1971). The time course of a typical C. vicina experiment is depicted in Fig. 3, and the rates derived from several such experiments
4
ffl
!2
1 2 3
HOURS
Fig. 3. Rate of incorporation of UMP into rRNA in the "white pupa" of C. vicina. Insects were injected with 0.5 #Ci of 14C-uridine. After the times indicated following injec- tion, RNA was extracted and purified from individual in- sects as described. Total radioactivity incorporated into rRNA was converted into n-moles of UMP incorporated by determining the specific activity of the UTP pool. Cor- rection was made for incorporation of labeled CMP into the rRNA. Recovery of total t4C-rRNA was assessed by using an internal standard of purified 3H-rRNA added to
the extraction medium.
6.0
-g ! 6.o x ~ 4.0--
~ 3.0
i 1 .0 - - c
I -2
O .
o~ - O
=le E 130 N
z 8 ~
4 ~
I I I I I I I
WP EMERGENCE
I I t I I I I 0 2 4 6 8 10 12
DAYS AFTER PUPARIATION
Rates of incorporation of UTP into Fig. 4. rRNA ( H ) , rRNA pool size (O O) and UTP pool size (A /x) at various developmental stages of the blowfly
C. vicina.
at selected stages of development are shown in Fig. 4. The rate of rRNA biosynthesis is maximal in the late 3rd instar larva, and drops markedly by the white puparium stage. Two days later, a somewhat lower value is observed, and this lower value, allowing for biological and experimental variations, is maintained for the duration of pharate adult development. The average rate during this time is 1.7 n-mole UMP in- corporated into rRNA/insect/hr.
A simple calculation now permits a comparison of the rate of rRNA synthesis as measured above with that derived indirectly from the decay of rRNA speci- fic activity (PgoTZEL et al., 1976). The results of such a comparison (Table 1) show that the value obtained by pulse labeling is ca. 74~o of that found by the indirect procedure. Considering the various assump- tions involved in both sets of experiments, such good agreement between the values suggests that either approach is a valid measure of the rate of rRNA bio- synthesis.
DISCUSSION
The rates of rRNA synthesis as measured above are clearly dependent upon accurate determination of
Table 1. Comparison of rates of rRNA biosynthesis as measured by pulse labeling and rRNA decay
rRNA per insect per day n-mole UTP*
#g formed incorporated
Pulse labeled 46 41 rRNA decay 62 56
* 1.0#g rRNA contains 0.9n-mole UMP and 31% of the constant pool of rRNA is renewed per day (PROTZEL et al., 1976). The rRNA pool size (see Results) is 200 #g/ insect.

634 ALBERTO PROTZEL AND LEO LEVENBOOK
two parameters, viz. the specific activities of the rRNA precursor pool, and of the UMP in rRNA. Appropriate corrections have been made to ensure accurate measurement of the UMP, but a correct esti- mate of the true precursor is of necessity more doubt- ful. Labeled uridine is readily taken up by most cells (see review by HOeHSTADT, 1974) prior to its sub- sequent phosphorylation to UTP. For mammalian cells in tissue culture there is some controversy as to whether such newly generated UTP is, or is not, part of the true precursor pool for nucleolar RNA synthesis (HocHSTADT, 1974; GOODY and ELLEM, 1975); identity of total UTP from entire insects with the true precursor pool must therefore be at least equally doubtful. However, the good agreement between the rate of rRNA synthesis during pharate adult development as measured directly above, and the value obtained indirectly from long-term labeling after all pools have presumably equilibrated (PRoTZEL et al., 1976) suggests that our estimate of the precur- sor pool is approximately correct. Feed-back of iso- tope from rRNA to precursor pool would lead to an underestimate of the rate of incorporation. The magnitude of this error over the period of incorpor- ation can be calculated from the data of PROTZEL et al. (1976); it does not exceed 570 of the incorpor- ation rate, and has therefore been ignored.
The relative constancy in the rate of rRNA syn- thesis during blowfly pharate adult development is unexpected in view of the extensive morphogenetic changes occurring during this time. The overall rate for the whole organism obviously represents a com- posite of the rates of rRNA synthesis of individual cells and tissues which in themselves may vary widely. A case in point is the developing ovary of the female silkworm, which has a dominant r61e in RNA syn- thesis (CHINZEI and ToJo, 1972). However, blowfly ovaries are incompletely developed at adult emer- gence, and in our study the disproportionate contri- bution of any particular organ affecting the rate of rRNA synthesis has not been detected. Constancy of the rate of UTP incorporation into RNA in whole organisms has also been reported for the entire deve- loping chick (BARBmOLI et al., 1973) and, apparently, in Drosophila embryos (FAUSTO-STERLING et al., 1974). It is worth noting also that the constant rate of rRNA synthesis is markedly different from the U-shaped profiles for the rates of protein synthesis during phar- ate adult development observed in other blowflies (DINAMARCA and LEVENBOOK, 1966; WILLIAMS and BraT, 1972).
The present pulse-labeled estimate of the rate of r R N A synthesis, when integrated over the period from pupariation to adult emergence, leads to a value of 8770 replacement of the rRNA pool; the corre- sponding figure from long-term labeling is 937o (PRO- TZEL et al., 1976). Either value indicates a major de novo synthesis of adult fly ribosomes during metamor- phosis, and supports the view that there is little, if any, carry-over of larval extra-cellular ribosomes (if
these do in fact exist) into adult tissues (LEVENBOOK et al., 1976).
REFERENCES
BARBmOLI B., CORI A., MONTI M. G., and MORUZZ! M. S. (1973) Incorporation of labeled uridine into acid-solu- ble and acid insoluble materials during development of chick embryo. Develop. Biol. 35, 336-369.
BtlCHER N. L. R. and SWAFFIELD M. N. (1966) Rate of incorporation of 6-1*C-orotic acid into uridine 5'-tri- phosphate and cytidine 5'-triphosphate and nuclear ribo- nucleic acid in regenerating rat liver. Biochim. Biophys. Acta 108, 551-567.
CHEN P. S. and LEVENBOOK L. (1966) Studies on the hemo- lymph proteins of the blowfly Phormia regina--II. Syn- thesis and breakdown as revealed by isotopic labelling. J. Insect Physiol. 12, 1611-1627.
CHINZEI Y. and ToJo S. (1972) Nucleic acid changes in the whole body and several organs of the silkworm, Bombyx mori, during metamorphosis. J. Insect Physiol. 18, 1683-1698.
DINAMARCA M. L. and LEVENBOOK L. (1966) Oxidation, utilization and incorporation into protein of alanine and lysine during metamorphosis of the blowfly, Phormia regina. Arch. Biochem. Biophys. 117, 110-119.
EMERSON C. P. Jr. and HUMPHREYS T. (1971) A simple and sensitive method for quantitative measurement of cellular RNA synthesis. Anal. Biochem. 40, 254-266.
EAUSTO-STERLING A., ZHEUTLIN L. M., and BROWN P. R. (1974) Rates of RNA synthesis during early embryo- genesis in Drosophila melanogaster. Develop. Biol. 40, 78-83.
GIRARD M., LATHAM H., PENMAN S., and DARNELL J. E. (1965) Entrance of newly formed messenger RNA and ribosomes into HeLa cell cytoplasm. J. molec. Biol. 11, 187-201.
GOODY H. E. and ELLEM K. A. O. (1975) Nutritional effects on precursor uptake and compartmentalization of intra- cellular precursor pools in relation to RNA synthesis. Biochim. Biophys. Acta 383, 30-39.
GREENBERG J. R. (1969) Synthesis and properties of riboso- mal RNA in Drosophila. J. molec. Biol. 46, 85-98.
HASTINGS J. R. B. and KIRBY K. S. (1966) The nucleic acids of Drosophila melanogaster. Biochem. J. 100, 532-539.
HIRSCH C. A. (1967) Quantitative determination of the ribosomal ribonucleic acid content of liver and Novikoff hepatoma from fed and fasted rats. d. biol. Chem. 242, 2822-2827.
HOCHSTADT J. (1974) The role of the membrane in the utilization of nucleic acid precursors. CRC Crit. Rev. Biochem. 2, 259-310.
KRAMER G., W1EGERS U., and HILZ H. (1973) mRNA turn- over studies applying labeled uridine require an evalu- ation of specific radioactivities of UTP and RNA-U. Bio- chem. Biophys. Res. Comm. 55, 273-281.
LEVENBOOK L., SRIDHARA S., and LAMBERTSSON A. (1976) Extracellular ribosomes during metamorphosis of the blowfly Calliphora vicina R. D.--A reappraisal of their authenticity. Biochem. Biophys. Res. Comm. 70, 15-21.
LOENING U. E. (1969) The determination of the molecular weight of ribonucleic acid by polyacrylamide-gel electro- phoresis. The effects of changes in conformation. Bio- chem. J. 113, 131-138.

rRNA synthesis in Calliphora vicina 635
PROTZEL A., SRIDHARA S., and LEVENBOOK L. (1976) Turn- over of ribosomes during metamorphosis of the blowfly, Calliphora vicina. Insect. Biochem. (In press).
SERFLING E., MAXIMOVSKY L. F., and WOBUS U. (1974) Synthesis and processing of ribosomal ribonucleic acid in salivary glands of Chironomus thummi. Eur. J. Bio- chem. 45, 277-289.
SRIDHARA S. and LEVENBOOK L. (1974) The contribution of the fat body to RNA and ribosomal changes during
development of the blowfly Calliphora erythrocephala (Meig.). Develop. Biol. 38, 64-72.
WmUAMS K. L. and Brat L. M. (!972) A study of the quantitative significance of protein synthesis during the metamorphosis of the sheep blowfly Lucilia. Insect Bio- chem. 2, 305-320.
Key Word Index: Calliphora vicina, metamorphosis, ribosomal RNA, ribosomes, rates of synthesis.










![Optical(Coherence(Tomography:(Age(estimation(of(Calliphora ......Introduction(Pupae,)a)sedentary)and)long]lasting)stage)of)the)blowfly)lifecycle,)are)often)reported)at)crime)scenes)](https://img.pdfslide.us/doc/110x75/60b54e2575eeff0dad7cccb6/opticalcoherencetomographyageestimationofcalliphora-introductionpupaeasedentaryandlonglastingstageoftheblowflylifecycleareoftenreportedatcrimescenes.jpg)


















