Embed Size (px)
Citation preview

1. Euk. Microbiol.. 44(4). 1997 pp. 345-351 0 1997 by the Society of Protozoologists
Rapid Methylation of Cell Proteins and Lipids in Trypanosoma brucei BURT GOLDBERG,*,****,',* NIGEL YARLETT,*,** DONNA RATTENDI,*
DAVID LLOYD**** and CYRUS J. BACCHI*,*** *Haskins Laboratories, and
**Department of Chemistry and Physical Sciences, and ***Department of Biology, Pace University, 41 Park Row, New York, New York 10038 USA, and
****University of Wales, School of Pure & Applied Biology, Cardiff CFI 3TL, Wales, UK
ABSTRACT. The fate of the [ m e t h ~ l - ~ ~ C ] group of S-adenosylmethionine (AdoMet) in bloodstream forms of Trypanosoma brucei brucei, was studied. Trypanosomes were incubated with either [methyl-'4C]methionine, [U-14C]methionine, S-[methyl-I4C]AdoMet or ["Slmethionine and incorporation into the total TCA precipitable fractions was followed. Incorporation of label into protein through methylation was estimated by comparing molar incorporation of [methyl-I4C] and [U-I4C]methionine to [35S]methionine. After 4-h incu- bation with [U-14C]methionine. [methyl-'4C]methionine or ["S]methionine, cells incorporated label at mean rates of 2,880 pmol, 1,305 pmol and 296 pmol per mg total cellular protein, respectively. Cells incubated with [U-I4C] or [methyl-'4C]methionine in the presence of cycloheximide (50 pg/ml) for four hours incorporated label eight- and twofold more rapidly, respectively, than cells incubated with [35S]methionine and cycloheximide. [Methyl-I4C] and [U-14C]methionine incorporation were > 85% decreased by co-incubation with unlabeled AdoMet (1 mM). The level of protein methylation remaining after 4-h treatment with cycloheximide was also inhibited with unlabeled AdoMet. The acid precipitable label from [U-14C]methionine incorporation was not appreciably hydrolyzed by DNAse or RNAse treatment but was 95% solubilized by proteinase K. [U-14C]methionine incorporated into the TCA precipitable fraction was susceptible to alkaline borate treatment, indicating that much of this label (55%) was incorporated as carboxymethyl groups. The rate of total lipid methylation was found to be 1.5 times that of protein methylation by incubating cells with [U-14C]methionine for six hours and differential extraction of the TCA lysate. These studies show T. b. brucei maintains rapid lipid and protein methylation, confirming previous studies demonstrating rapid conversion of methionine to AdoMet and subsequent production of post-methylation products of AdoMet in African trypanosomes.
Supplementary key words. Lipid methylation, protein methylation, S-adenosylmethionine.
ETHIONINE is a critical metabolite for eukaryotic cells, hav- M ing multifunctional roles in the cell for protein synthesis, as a precursor of AdoMet the methyl group donor in transme- thylation reactions, and as decarboxylated AdoMet, the ami- nopropyl group donor in polyamine biosynthesis [ 121. AdoMet synthetase is a tightly regulated enzyme in mammalian cells [ 121, while the product of transmethylation reactions, S-aden- osylhomocysteine (AdoHcy), is rapidly hydrolyzed to adeno- sine and homocysteine. The latter can be converted to cysteine by transsulfuration, or recycled to methionine via methionine synthetase [9]. A delicate equilibrium therefore exists for uti- lization of methionine in protein synthesis, in transmethylation and in polyamine synthesis, and for recycling of byproducts of transmethylation reactions as methionine or transsulfuration metabolites (e.g. cysteine, cystathionine).
African trypanosomes are endemic in Equatorial Africa, in- fecting humans, domestic and wild animals [14]. Recent studies have demonstrated these organisms have an unregulated AdoMet synthetase which, although normally producing a con- stant pool of AdoMet, oversynthesizes AdoMet when the or- ganism is exposed to antagonists of polyamine synthesis and agents blocking transmethylation [e.g. DL-a-difluoromethylor- nithine, MDL 73811, sinefungin: 3, 6, 25-27].
In mammalian cells, nucleic acids, proteins and lipids are major recipients of methyl groups from AdoMet. Although methylation of nucleic acids has been implicated in cellular regulatory events, the roles of protein and lipid methylation are now becoming evident. For example, the addition of methyl groups to protein terminal cysteine moieties of signal transduc- tion proteins has been found to guide their placement in cell membranes [7, 131. Protein methylation has been shown to have
I In partial fulfillment of requirements for the Doctoral degree, Uni- versity of Wales, Cardiff, Wales, UK.
* To whom correspondence should be addressed. Telephone: 212-346- 1246; Fax: 212-346-1586.
Abbreviations: AdoHcy, S-adenosylhomocysteine; AdoMet, S-aden- osylmethionine; DFMO, DL-a-difluromethylornithine; HMI- 18, Isco- ve's modified Dulbecco's medium, with 20% fetal bovine semm (FBS): MDL 7381 1, 5 ' - ( [(Z)-4-amino-2-butenyl]-methylamino)-5'-deoxy- adenosine TCA, trichloroacetic acid.
a role in the maintenance and repair of cellular proteins, in the function of chemotaxis receptors and as a modulator of cal- modulin sensitivity [13, 18, 241.
Our previous work has shown that the predominant form of methylation of trypanosomal proteins is carboxylmethylation catalyzed by protein methylase I1 {26], however the overall de- gree of protein methylation and the fate of methyl groups in trypanosomes is unknown. The present study examines the ca- pability of the parasite to carry out methylation of proteins dur- ing normal growth and after blockade of protein synthesis by cycloheximide utilizing [35S] and ['4C]methionine, and S-[methylJ4C]AdoMet, and examines the partitioning of me- thionine for methylation of proteins and lipids versus that de- voted to protein synthesis.
MATERIALS AND METHODS
Organism. Bloodstream trypomastigotes of Trypanosoma brucei brucei Lab 110 EATRO were grown in rats, (Wistar Female, 200 g , Ace Animals, Boyertown, PA), harvested by cardiac puncture after CO, asphyxiation and separated from blood elements at 0-4" C by DEAE-cellulose chromatography in TSG buffer [0.1 M Tris-HC1 buffer, pH 7.4 containing 40 mM glucose, plus 50 U penicillin and 50 pg/ml streptomycin: 4, 151. This process took 4 h, during which trypanosomes were in an essentially methionine-free environment. Viability was checked microscopically.
Incubation studies. Purified trypanosomes from rat blood were used for short term methionine uptake studies by resus- pending cells in HMI- 18 medium [modified Iscove's medium: 111 supplemented with 20% FBS. Cultures were maintained in HMI-18 medium in 24-well culture plates (Falcon 3047). Prior to incorporation studies, all cells were cultured for 48 h in 250-ml tissue culture flasks (20 ml per flask). Cells were ex- amined microscopically for viability, and harvested by centrif- ugation to be utilized as described below. As a control for try- panosomes harvested from rats, initial incorporation studies also utilized trypanosomes continuously cultivated in HMI- 18 [111.
Time course incorporation experiments utilized trypano- somes suspended in HMI-18 - methionine, containing 20%
345

346 J. EUK. MICROBIOL., VOL. 44, NO. 4, JULY-AUGUST 1997
(v/v) dialyzed FBS. Cells were transferred to 24-well culture plates at 5 X 106cells/0.5 ml of medium and incubated with 4 nmol of either L-[35S]methionine, L-[U-I4C]methionine or L-[methyl-I4C]methionine per well. For inhibitor studies, try- panosomes were exposed to cycloheximide at 50 p g / d or 2 nM sinefungin for 4 h in complete HMI-18, centrifuged and incubated for an additional 1-4 h in HMI-18 - methionine and dialyzed FBS, but containing inhibitor and radiolabeled methi- onine.
Analysis of methionine incorporation. At specific time points (0.25-4 h) cells were centrifuged (12,000 g) for 1 min, resuspended in fresh medium minus label and methionine, cen- trifuged and precipitated with 10% TCA. After a 30-min in- cubation (0-4" C), suspensions were centrifuged for 5 min. The supernatant was frozen at -75" C while the precipitate was resuspended in ice cold 10% TCA, and washed onto 25 mm Whatman GFB filters prewet with 10% TCA. Filters were washed twice with 10% TCA, once with 10% TCA + 40 mM nonradiolabeled methionine and twice with 95% ethanol (to re- move lipids). All wash solutions were ice cold. Filters were air dried, added to vials containing Omniflor and counted.
Calculations. [35S]Methionine (1,170 Ci/mmol), [U- I4C]methionine (290 mCi/mmol), [methyl-'4C]methionine (57.4 mCdmmol) and S-[methyl-'4C]adenosylmethionine (57.6 mCd mmol). Each well of cells was given either 4.0 nmol of [3sS]methionine (1.0 mCi, 0.89 nmol, + 3.1 nmol nonradiola- beled methionine), 4.0 nmol of [U-I4C]methionine (1.16 pCi), 4.4 nmol [methyl-I4C]methionine (0.25 pCi) or 3.47 nmol (0.2 pCi) of S-[methyl-I4C]AdoMet. Larger quantities of label, es- pecially [3sS]methionine, could not be added because of exces- sively high counts. Since T. b. brucei is very sensitive to eth- anol, [methyl-'4C]methionine as an ethanol preparation was evaporated over N, and brought to volume with HMI-18 - methionine + FBS before use.
Incorporation was calculated as pmoVmg total cellular pro- tein. Protein determination was carried out by lysing cells with sodium dodecyl sulfate and estimated by the method of Lowry et al. [17].
Data were analyzed by Sigma Plot (Jandel Scientific, San Rafael, CA), using best fit for data points and data from mul- tiple separate experiments. All experiments were carried out in triplicate. Data points are mean values. The specific activity for the incorporation of methyl groups using [methyl- l4C1methionine was calculated as one-fifth of the activity of [U- ''Clmethionine. The normal incorporation of [35S]methionine into protein structure was determined to be only 510% of that incorporated as protein methylation using [methyl-'T]methionine. Economically it was more feasible to use the universal label than the methyl-labeled methionine.
Enzymatic fractionation of TCA precipitates. T. b. brucei blood forms (20 ml of 5 X 10Vml) were incubated for 4 h with 10 pCi [U-'4C]methionine (35 nmol) in HMI-18 - methionine, supplemented with 20% dialyzed FBS, at 37" C. After incuba- tion cells were harvested by centrifugation and the pellets were treated with lml 10% TCA and incubated for at least 30 min at 4" C. The precipitates were centrifuged for 10 min and the pellets were washed with two volumes of ethanol: ether (1:l v/v), air dried, and resuspended in 0.01 M Tris-HC1, pH 7.5. Aliquots (250 p1) of resuspended pellets were then digested with DNAse, RNAse, or proteinase K in the following reaction mixtures: DNAse (40 U/ml), 10 mM Tris-HC1, 50% (v/v) glyc- erol pH 7.5, 10 mM MgCl, and 10 mM CaCl,; RNAse (10 U/ml), 20 mM Tris:HCl, 15 mM NaCl; proteinase K (400 Ulml), 10 mh4 Tris HCl pH 7.5. Reactions were incubated at 37" C for 16 h, and stopped by precipitating with 10% TCA.
The precipitated fractions were then filtered, washed and count- ed.
Determination of the initial rate of protein methylation. To determine the rate of protein methylation, trypanosomes (1 X lO9/ml) were incubated for up to 10 min with 0.2 pCi (3.47 nmol) S-[methyl-I4 CIAdoMet in HMI-18 - methionine without serum. Cells were centrifuged at room temperature for 1 rnin and resuspended in HMI- 18 - methionine without supplemen- tal AdoMet or serum. Cells were harvested at 1, 5 and 10 min, washed once with media, repelleted and lysed with 100 pl of 10% TCA. The lysates were incubated at least 30 min on ice and centrifuged for 1 min. Incorporation was measured for the pellets as described previously. Supernatants were spotted on filters (GFB, 25 mm), dried and counted, to determine label in cytosol. This allowed AdoMet uptake to be distinguished from methyl donation to protein by AdoMet.
Analysis of carboxylrnethylation of proteins. The amount of [U-I4C]methionine incorporated into protein as carboxylme- thy1 groups was determined by alkaline hydrolysis, extraction and quantitation of labile [I4CH,] in the resulting methanol as described [26]. T. b. brucei bloodstream forms from rats were grown in culture for 48 h and labeled as detailed above. Labeled cells were treated with 10% TCA and the precipitate was re- suspended in 1 ml 0.5 M borate buffer, pH 11.0, incubated overnight at room temperature, and extracted with 6 ml of (4: 1 v/v) isoamyl alcoho1:toluene. The radioactivity in the organic phase was determined before and after overnight evaporation at room temperature with the nonrecoverable volatile fraction assumed to be methanol, as previously shown [23, 241.
Extraction and analysis of total lipids. The TCA lysate from cells labeled with [U-'4C]methionine for 4 h was extracted with two volumes of ch1oroform:methanol (2:l v/v) at room temperature for 8 h [20]. The organic phase was removed and the remaining aqueous phase was washed two times with an equal volume of chloroform:methanol:H,O (48:47:5 v/v). Or- ganic as well as aqueous phases were spotted on filters, dried and counted. Organic phases were then combined and evapo- rated. The residues were resuspended in ch1oroform:methanol (2:l v/v) and counted.
Sources of chemicals. S-[methyl-'4C]adenosyl-L-methionine (58 mCi/mmol), L-[methyl-'4C]methionine (57 mCi/mmol), L-[U-14C]methionine (290 mCi/mmol), L-[3sS]methionine (1,170 Cdmmol) were from Dupont, NEN Research Products (Boston, MA). Iscove's modified Dulbecco's medium, and all sera were from GibcolBRL (Grand Island, NY). All other bio- chemicals were from Sigma (St. Louis, MO).
RESULTS Incorporation of rnethionine into protein. Throughout the
study, trypanosomes harvested from rat blood and subcultured for 48 h were used for incorporation studies, since the number obtainable far exceeded that from continuous culture (5 X 10Vml culture vs. 109/ml blood). The incorporation of [35S]methionine into TCA precipitates from these cells was first compared to incorporation by continuously cultured cells and found to be similar over the 4-h incubation period in several experiments.
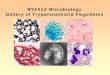
The rates of incorporation of [35S]methionine and [U- I4C]methionine into protein over a 4-h period were compared (Fig. 1). Incorporation was linear over the 4-h period and in- creased as the concentration of methionine increased (not shown). The amount of incorporation with 4.0 nmol [U- I4C]methionine was approximately tenfold greater than with an equimolar amount of ['5S]methionine: 296 (t 120; n = 9) pmollmg total cellular protein for [35S]methionine and 2,880 (i. 120; n = 3) pmollmg total cellular protein for [U-
347 GOLDBERG ET AL.-METHYLATION OF TRYPANOSOME PROTEIN
3500 5
9) 3000
0
R 2000
m
E 1500
v)
.- c
2500
- 9) 1000 - 0
500 E
0 1 2 3 4 a time (h)
Fig. 1. Incorporation of [U-I4C]methionine and 13%] into the acid precipitable fraction of T. b. brucei. Bloodstream trypomastigotes (5 X lo6 cells/0.5 ml) were incubated in HMI-18 medium containing 4 nmol of radiolabel as described in the Materials and Methods. Data is ex- pressed as mean values (? SD) of methionine incorporated as pmoVmg protein from 0-4 h: 0, [U-14C]methionine; H, [35S]methionine.
I4C]methionine (Fig. 1). Previous work demonstrated that try- panosomes incubated with [35S]methionine over this time in- corporate 75% of the label in the cytosol 131, with -40% ap- pearing as [35S]AdoMet, and 37% as post-methylation products of AdoMet, an indication that most of cellular methionine is incorporated into AdoMet and that the majority of the AdoMet is being used for methylation reactions.
Incorporation of [U-14C]methioNne and [methyl-14C]methionine into protein was also compared (Fig. 2) to directly quantitate transfer of the labeled methyl group of [methyl-'4C]methionine and [U-14C]methionine. At the end of 4 h of exposure the in- corporation rate was 2,880 (? 120; n = 3) vs. 1,305 (? 5; n = 3) pmol/mg total cellular protein for [UI4C] and [methyl-I4C] label, respectively. The twofold difference between the two ['*C] labels is not significant when compared to the four- and tenfold lower rates of [35S]methionine incorporation into protein primary structure (296 pmol/mg total cellular protein). For this reason and because of the significantly lower cost, [U- 14C]methionine was used in subsequent studies, and calculations were based on one-fifth of its specific activity to reflect pri- marily incorporation of the S-methyl group.
Figure 2 also demonstrates the effects of cycloheximide on incorporation of [35S], [U-I4C] and [methyl-14C]methionine. Cells were preincubated for 4 h with 50pg/ml of cyclohexi- mide, then harvested and resuspended with either 4.0 nmol of [35S]methionine, [U-14C]methionine or 4.4 nmol [methyl- I4C]methionine for an additional 4 h in the presence of cyclo- heximide. Incorporation of each label into protein was inhibited to about the same extent after 4 h: 90% for [35S]methionine, 89% for [U-I4C]methionine, and 96% for [methyl-14C]methionine (Fig. 2). However, the rate of [U-I4C]methionine incorporation, 311 (t 283; n = 6) pmoVmg total cellular protein, remained approximately eightfold higher than that of [35S]methionine in- corporation, 30 (+ 29; n = 6) pmol/mg total cellular protein, in the presence of cycloheximide after a 4 h incubation (Fig. 2) and that of [methyl-'4C]methionine incorporation, 67 ( 5 2; n = 2) pmoVmg total cellular protein. One reason for the in- hibition of [U-I4C] and [methyl-14C]methionine incorporation in
c T- 10000
.-
: Iooo i j a
loo 1 0)
E . -
a 1 ° 4
0 50 100 150 200 250 300
Time (rnin.)
Fig. 2. Effect of cycloheximide on the incorporation of 4.0 nmol [15S], [U-I4C] and 4.4 nmol [methyl-'4C]methionine into acid precipi- table fractions. Incorporation of ['5S], [U-'4C] and [methyl- I4C]methionine was compared in control cells and cells pretreated for 4 h with 50pg/ml cycloheximide, as described in the Materials and Methods: 0, [U-I4C]methionine control; ., [U-"C] + cycloheximide; A, [methyl-'4C]methionine control; V, [methy1-l4C] + cycloheximide; + , [35S]methionine control; filled hexagon, ['%I + cycloheximide.
the presence of cycloheximide may be concerned with the find- ing that selective methylation of newly formed protein occurs at the level of the polysome, linking protein synthesis to meth- ylation [20].
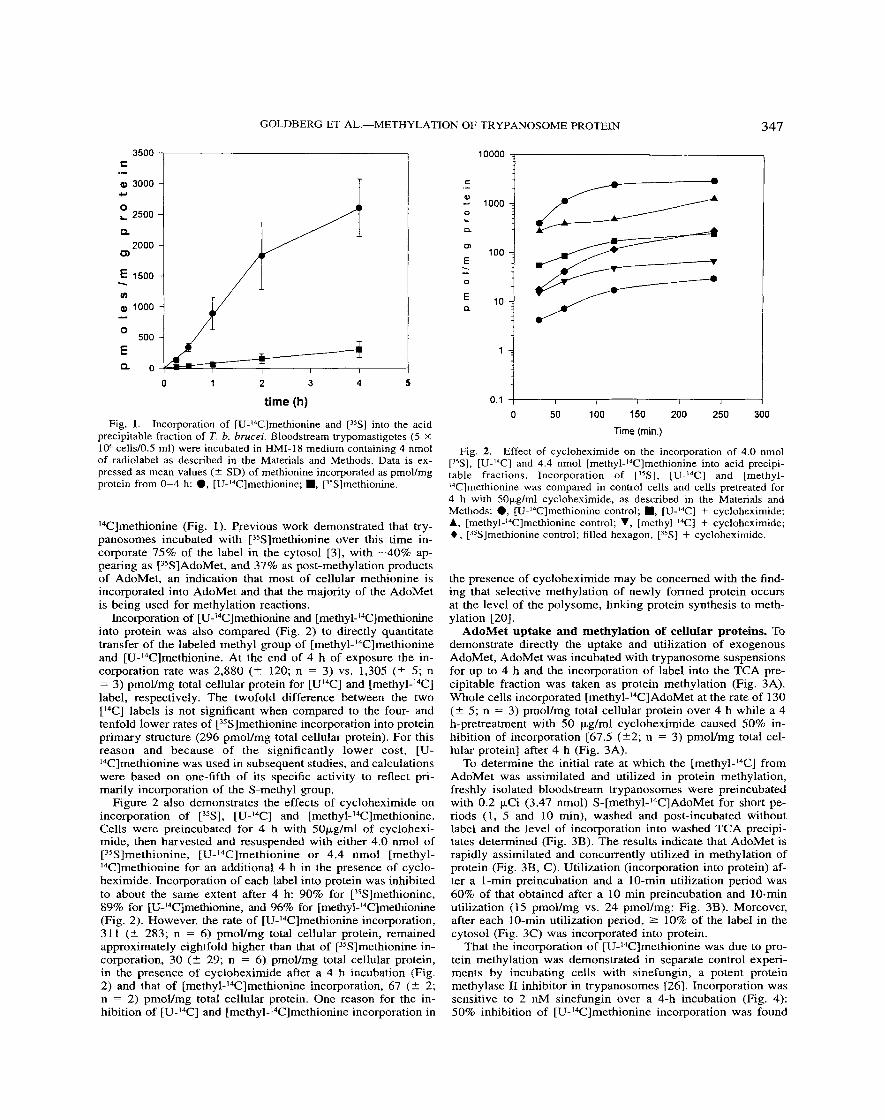
AdoMet uptake and methylation of cellular proteins. To demonstrate directly the uptake and utilization of exogenous AdoMet, AdoMet was incubated with trypanosome suspensions for up to 4 h and the incorporation of label into the TCA pre- cipitable fraction was taken as protein methylation (Fig. 3A). Whole cells incorporated [methyl-I4C]AdoMet at the rate of 130 (? 5 ; n = 3) pmoVmg total cellular protein over 4 h while a 4 h-pretreatment with 50 pg/ml cycloheximide caused 50% in- hibition of incorporation [67.5 (?2; n = 3) pmol/mg total cel- lular protein] after 4 h (Fig. 3A).
To determine the initial rate at which the [methyl-14C] from AdoMet was assimilated and utilized in protein methylation, freshly isolated bloodstream trypanosomes were preincubated with 0.2 pCi (3.47 nmol) S-[methyl-14C]AdoMet for short pe- riods (1, 5 and 10 min), washed and post-incubated without label and the level of incorporation into washed TCA precipi- tates determined (Fig. 3B). The results indicate that AdoMet is rapidly assimilated and concurrently utilized in methylation of protein (Fig. 3B, C). Utilization (incorporation into protein) af- ter a 1-min preincubation and a 10-min utilization period was 60% of that obtained after a 10 min preincubation and 10-min Utilization (15 pmoYmg vs. 24 pmol/mg: Fig. 3B). Moreover, after each 10-min utilization period, 2 10% of the label in the cytosol (Fig. 3C) was incorporated into protein.
That the incorporation of [U-I4C]methionine was due to pro- tein methylation was demonstrated in separate control experi- ments by incubating cells with sinefungin, a potent protein methylase I1 inhibitor in trypanosomes [26]. Incorporation was sensitive to 2 nM sinefungin over a 4-h incubation (Fig. 4): 50% inhibition of [U-14C]methionine incorporation was found
348 J. EUK. MICROBIOL., VOL. 44, NO. 4, JULY-AUGUST 1997
150
c 140 - A - . 130 - 120 - 110 - 100 - 90- E
2 80 - * 70 -
*
rn
m
I
0 50 100 150 200 250 300
lime (mln.)
30
25
20 C a .- I
P a E l5
E :
10
5
0
240
200
E a - -
180
E E 5
120
80
40
B
I I I I I
0 2 4 0 6 10 12
Time (min.)
0 2 c 8 8 10 12
Time (mln.)
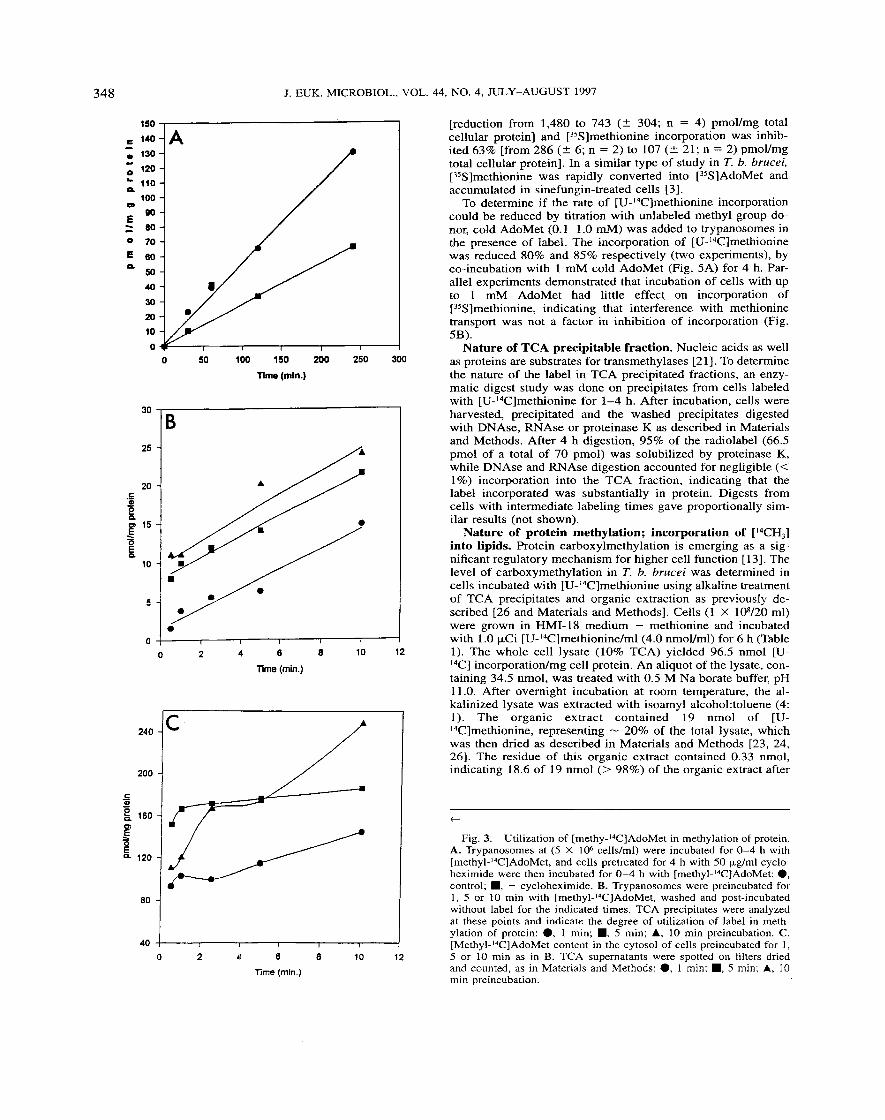
[reduction from 1,480 to 743 ( 2 304; n = 4) pmovmg total cellular protein] and [35S]methionine incorporation was inhib- ited 63% [from 286 (5 6; n = 2) to 107 ( 2 21; n = 2) pmoVmg total cellular protein]. In a similar type of study in T. b. brucei, [35S]methionine was rapidly converted into [3SSlAdoMet and accumulated in sinefungin-treated cells 131.
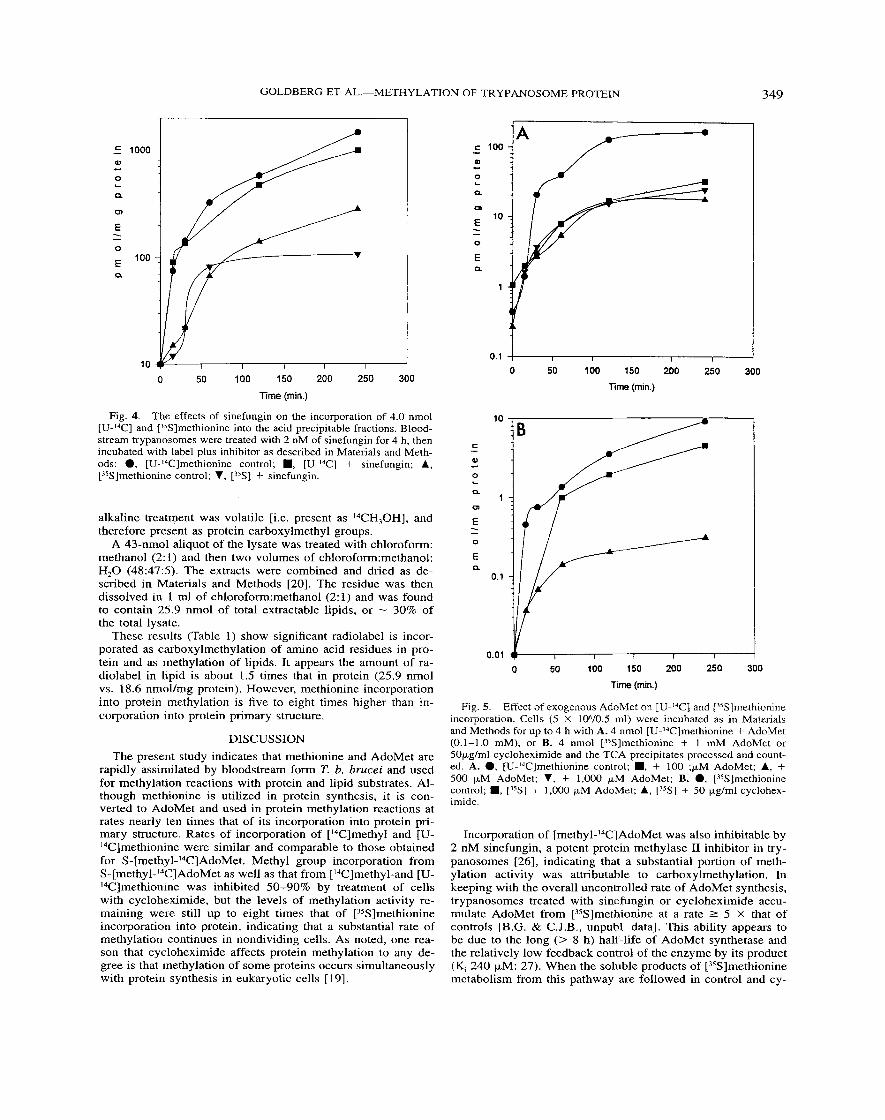
To determine if the rate of [U-I4C]methionine incorporation could be reduced by titration with unlabeled methyl group do- nor, cold AdoMet (0.1-1.0 mM) was added to trypanosomes in the presence of label. The incorporation of [U-I4C]methionine was reduced 80% and 85% respectively (two experiments), by co-incubation with 1 mM cold AdoMet (Fig. 5A) for 4 h. Par- allel experiments demonstrated that incubation of cells with up to 1 mM AdoMet had little effect on incorporation of [35S]methionine, indicating that interference with methionine transport was not a factor in inhibition of incorporation (Fig. 5B).
Nature of TCA precipitable fraction. Nucleic acids as well as proteins are substrates for transmethylases [21]. To determine the nature of the label in TCA precipitated fractions, an enzy- matic digest study was done on precipitates from cells labeled with [U-I4C]methionine for 1-4 h. After incubation, cells were harvested, precipitated and the washed precipitates digested with DNAse, RNAse or proteinase K as described in Materials and Methods. After 4 h digestion, 95% of the radiolabel (66.5 pmol of a total of 70 pmol) was solubilized by proteinase K, while DNAse and RNAse digestion accounted for negligible (< 1 %) incorporation into the TCA fraction, indicating that the label incorporated was substantially in protein. Digests from cells with intermediate labeling times gave proportionally sim- ilar results (not shown).
Nature of protein methylation; incorporation of [I4CH3] into lipids. Protein carboxylmethylation is emerging as a sig- nificant regulatory mechanism for higher cell function [13]. The level of carboxymethylation in T. b. hrucei was determined in cells incubated with [U-I4C]methionine using alkaline treatment of TCA precipitates and organic extraction as previously de- scribed [26 and Materials and Methods]. Cells ( I X 108/20 ml) were grown in HMI-18 medium - methionine and incubated with 1.0 FCi [U-'4C]methionine/ml (4.0 nmovml) for 6 h (Table 1). The whole cell lysate (10% TCA) yielded 96.5 nmol [U- "C] incorporatiodmg cell protein. An aliquot of the lysate, con- taining 34.5 nmol, was treated with 0.5 M Na borate buffer, pH 11.0. After overnight incubation at room temperature, the al- kalinized lysate was extracted with isoamyl alcoho1:toluene (4: 1). The organic extract contained 19 nmol of [U- l4C1methionine, representing - 20% of the total iysate, which was then dried as described in Materials and Methods [23, 24, 261. The residue of this organic extract contained 0.33 nmol, indicating 18.6 of 19 nmol (> 98%) of the organic extract after
t
Fig. 3. Utilization of [methy-'4C]AdoMet in methylation of protein. A. Trypanosomes at (5 X lo6 cells/ml) were incubated for 0-4 h with [methyl-14C]AdoMet, and cells pretreated for 4 h with 50 pglml cyclo- heximide were then incubated for 0-4 h with [methyl-14C]AdoMet: 0, control; W, + cycloheximide. B. Trypanosomes were preincubated for 1, 5 or 10 min with [methyl-'4C]AdoMet, washed and post-incubated without label for the indicated times. TCA precipitates were analyzed at these points and indicate the degree of utilization of label in meth- ylation of protein: 0, 1 min; W, 5 min; A, 10 min preincubation. C. [Methyl-14C]AdoMet content in the cytosol of cells preincubated for 1, 5 or 10 min as in B. TCA supernatants were spotted on filters dried and counted, as in Materials and Methods: 0, 1 min; H, 5 min; A, 10 min preincubation.
.E 1000 Q
0
- L
n
0)
E . - 0
100
P
10
GOLDBERG ET AL.-METHYLATION OF TRYPANOSOME PROTEIN 349
0 50 100 150 200 250 300 Time (min.)
Fig. 4. The effects of sinefungin on the incorporation of 4.0 nmol [U-I4C] and [35S]methionine into the acid precipitable fractions. Blood- stream trypanosomes were treated with 2 nM of sinefungin for 4 h, then incubated with label plus inhibitor as described in Materials and Meth- ods: 0, [U-I4C]methionine control; ., [U-I4C] + sinefungin; A, [3SS]methionine control; V, [%I + sinefungin.
alkaline treatment was volatile [i.e. present as I4CH,OH], and therefore present as protein carboxylmethyl groups.
A 43-nmol aliquot of the lysate was treated with chloroform: methanol (2: 1 ) and then two volumes of ch1oroform:methanol: H,O (48:47:5). The extracts were combined and dried as de- scribed in Materials and Methods [20]. The residue was then dissolved in 1 ml of chlorofomxmethanol (2:l) and was found to contain 25.9 nmol of total extractable lipids, or - 30% of the total lysate.
These results (Table 1) show significant radiolabel is incor- porated as carboxylmethylation of amino acid residues in pro- tein and as methylation of lipids. It appears the amount of ra- diolabel in lipid is about 1.5 times that in protein (25.9 nmol vs. 18.6 nmollmg protein). However, methionine incorporation into protein methylation is five to eight times higher than in- corporation into protein primary structure.
DISCUSSION The present study indicates that methionine and AdoMet are
rapidly assimilated by bloodstream form T. b. brucei and used for methylation reactions with protein and lipid substrates. Al- though methionine is utilized in protein synthesis, it is con- verted to AdoMet and used in protein methylation reactions at rates nearly ten times that of its incorporation into protein pri- mary structure. Rates of incorporation of [I4C]methyl and [U- I4C]methionine were similar and comparable to those obtained for S-[methyl-I4C]AdoMet. Methyl group incorporation from S-[methyl-14C]AdoMet as well as that from [I4C]methyl-and [U- I4C]methionine was inhibited 50-90% by treatment of cells with cycloheximide, but the levels of methylation activity re- maining were still up to eight times that of [35S]methionine incorporation into protein, indicating that a substantial rate of methylation continues in nondividing cells. As noted, one rea- son that cycloheximide affects protein methylation to any de- gree is that methylation of some proteins occurs simultaneously with protein synthesis in eukaryotic cells [19].
c 100 - 0
0
a
a
- L
10 E - - 0
E n
1
0.1 I I I I I I
0 50 100 150 200 250 300 Time (min.)
10
c
Q
0
.-
I
L
a 1
W
E . - 0
E a
0.1
0.01 4 I I I I I
50 100 150 200 250 300 0 lime (min.)
Fig. 5. Effect of exogenous AdoMet on [U-l'Cl and [35S]methionine incorporation. Cells (5 X 106/0.5 ml) were incubated as in Materials and Methods for up to 4 h with A. 4 nmol [U-14C]methionine + AdoMet (0.1-1.0 mM), or B. 4 nmol [35S]methionine + 1 mM AdoMet or 50yg/ml cycloheximide and the TCA precipitates processed and count- ed. A. 0, [U-I4C]methionine control; ., + 100 ;yM AdoMet; A, + 500 yM AdoMet; V, + 1.000 )LM AdoMet; B. 0, [ZsS]methionine control; ., ['5S] + 1,000 yM AdoMet; A, ["S] + 50 yg/ml cyclohex- imide.
Incorporation of [methyl-14C]AdoMet was also inhibitable by 2 nM sinefungin, a potent protein methylase I1 inhibitor in try- panosomes [26], indicating that a substantial portion of meth- ylation activity was attributable to carboxylmethylation. In keeping with the overall uncontrolled rate of AdoMet synthesis, trypanosomes treated with sinefungin or cycloheximide accu- mulate AdoMet from [35S]methionine at a rate 2 5 X that of controls [B.G. & C.J.B., unpubl. data]. This ability appears to be due to the long (> 8 h) half-life of AdoMet synthetase and the relatively low feedback control of the enzyme by its product (K, 240 pM: 27). When the soluble products of [35S]methionine metabolism from this pathway are followed in control and cy-

350 J. EUK. MICROBIOL., VOL. 44, NO. 4, JULY-AUGUST 1997
Table 1. Fate of [U-14C]methionine in T. b. brucei: incorporation into protein and total lipid."
NmoVmg Total lysate Fraction protein/6 h (90)
Total lysate [U-I4C]methionineb 96.5 100.0 Alkali labile [I4CH,] protein extraction 34.5'
Na borate treated (total) 29.3 -
Organic fraction 19.0 Dried organic residue 0.33 Labile [I4CH,]' 18.6 19.3 Aqueous fraction 0.09
Organic fractiond 25.9 26.7 Aqueous 14.3 14.8
a Blood forms (5 X 106/ml) were grown in HMI-18 with FBS. Cells were incubated for 6 h in methionine-deficient HMI-18 + 20% FBS containing 4 nmol (1.16 FCi/ml) of [U-14C]methionine. Cells were har- vested, washed in TSG buffer and lysed with 10% TCA. Portions of the [U-I4C]methionine lysate were used to determine total incorporation, alkali-labile protein methylation, and incorporation into total lipid.
Proteins were labeled with [U-'4C]methionine, precipitated with cold 10% TCA, collected on filters and washed with cold TCA, TCA + methionine, and 95% ethanol as described in Materials and Methods.
Labile [I4CH,] is determined from the radioactivity measured before and after overnight evaporation of the organic extract at room temper- ature. The difference in [I4CH,] recovered is the volatile fraction, meth- anol.
This fraction is the combined organic extractions of the lysate with ch1oroform:methanol. and chloroform:methanol:H,O, dried and resus- pended in ch1oroform:methanol as described in Materials and Methods.
These fractions are the starting aliquots utilized in subfractionation from the 96.5 nmol of original lysate.
- -
- Lipid extraction 43.w
cloheximide treated cells, they remain unchanged over a 1-h incubation. These products include cystathionine and cysteine which are trans-sulfuration products formed after methyl do- nation by AdoMet [3; B.G. & C.J.B., unpubl. data].
The distribution of methyl groups in trypanosomes incubated for 6 h with [U-I4C]methionine indicated incorporation of meth- yl label in the lipid fraction was - 1.5 X that incorporated into protein methylation (25.9 vs 18.6 nmol/mg protein: Table 1). These values comespond to utilization of 20% of the cellular methionine for carboxylmethylation of protein and 30% for lip- id methylation and are comparable to those obtained for Leish- mania sp. incubated with S-[methyl-3H]AdoMet in which - 60% of the label was incorporated into total lipid and 30% as protein [2]. Another study, also examining fate of S-[methyl- 3H]AdoMet in Leishmania donovani, found 45% of total label as protein, and 25% as lipid [16]. It appears that Leishmania, as well as the African trypanosomes, utilize significant amounts of methionine as AdoMet in protein and lipid methylation re- actions.
The post-translational modification of proteins by methyl- ation has several important roles in cell metabolism including: (a) activation or deactivation specific proteins such as chemo- tactic receptors; (b) methylation of hormones and carrier pro- teins; (c) repair of aging proteins; (d) regulation of calrnodulin- dependent processes [8, 241. Methylation is a reversible post- translational modification of protein, since both methylases and specific esterases exist [12, 241. Within the realm of protein methylation, carboxylmethylation has been of particular interest with the finding that isoprenylation and carboxylmethylation may direct proteins to functional sites on the cell membrane [7]. Studies in progress in our laboratory indicate a substantial portion of trypanosome carboxylmethylation activity is also lo- calized in the cell membrane (N.Y. & CJB, unpubl. data).
The present study was based in part on the ability of blood- stream trypanosomes to assimilate S-[methyl-I4C]AdoMet and incorporate the label in transmethylation reactions. Studies US- ing pathogenic Leishmania sp. have also shown exogenous AdoMet served as source of methyl groups for protein and lipid methylation [2, 161 and strongly suggest that hemoflagellates as a group transport AdoMet which is present at - 70 nM in mammalian semm [22]. Co-incubation of trypanosomes with unlabeled AdoMet and the resulting significant reduction in in- corporation of [U-14C]methionine into protein are indicative of a high uptake rate (Fig. 5A). Since incorporation of [35S]methionine was minimally inhibited by exogenous AdoMet, the > 80% reduction of [U-14C]methionine incorpo- ration was not due to interference with transport. As a group, the ability of hemoflagellates to assimilate AdoMet may be in- dicative of a high methylation rate as well as the need to sal- vage preformed adenine, which is essential for growth [5]. In- deed, purine-starved Crithidia luciliae assimilated S-[methy- 3H]AdoMet and utilized it in protein methylation at a tenfold greater rate than cells cultivated in purine-replete medium [ 11.
There is a need for understanding the partitioning of rnethi- onine between the pathways sharing this essential amino acid in African trypanosomes. This study indicates that methylation of proteins and lipids are major sinks for the methyl group of methionine and far exceed the incorporation of methionine into primary protein structure. In this study we have described the degree of protein methylation that occurs in these organisms. In a related study we examined the effects of well characterized trypanocides (DFMO, sinefungin and MDL 7381 1) on protein methylation [lo]. These studies along with the present work indicate that transmethylation is a major part of trypanosome metabolism and a critical factor in the understanding of the mechanism of action of agents affecting polyamine or AdoMet synthesis.
ACKNOWLEDGEMENTS We thank Hao Zhang for his invaluable help in compiling
the figures used in the work. This study was supported by Na- tional Institutes of Health grant A117340 to CJB.
LITERATURE CITED
1. Alleman, M. M., Mann, V. H., Bacchi, C. J., Yarlett, N., Gottlieb, M. & Dwyer, D. M. 1995. Crithidia luciliae: Effect of purine starvation on S-adenosyl-L-methionine uptake and protein methylation. Exper. Parasitol., 81:519-528.
2. Avila, I. L. & Polegre, M. A. 1993. Uptake and metabolism of S-adenosyl-L-methionine by Leishmania mexicana and Leishmania bra- ziliensis promastigotes. Mol. Biochem. Parasitol., 58: 123-134.
3. Bacchi, C. J., Goldberg, B., Garofalo-Hannan, J., Rattendi, D., Lyte, P. & Yarlett, N. 1995. Fate of soluble methionine in African trypanosomes: effects of metabolic inhibitors. Biochem. J . , 309:737- 743.
4. Bacchi, C. J., Garofalo, J., Mockenhaupt, D., McCann, P. P., Diek- ema, K. A,, Pegg, A. E., Nathan, H. C., Mullaney, E. A., Chunosoff, L., Sjoerdsma, A. & Hutner, S. H. 1983. In vivo effects of DL-a- difluoromethylornithine on the metabolism and morphology of Trypanosoma brucei brucei. Mol. Biochem. Parasitol. 7:209-225.
5. Berens, R. L., Krug, E. C. & Marr, J. J. 1995. Purine and pyrim- idine metabolism. in: Marr, J. J. & Muller, M. (ed.), Biochemistry and Molecular Biology of Parasites. New York, Academic Press. Pp. 89- 117.
6. Bitonti, A. J., Byers, T. L., Bush, T. L., Casara, I? J. , Bacchi, C. J., Clarkson, Jr. A. B., McCann, P. P. & Sjoerdsma, A. 1990. Cure of Trypanosoma brucei brucei and Trypanosoma brucei rhodesiense in- fections in mice with an irreversible inhibitor of S-adenosylmethionine decarboxylase. Antimicrob. Agents Chemother., 34: 1485-1490.
7. Clarke, S. 1992. Protein isoprenylatlon and methylation at car- boxyl-terminal cysteine residues. Annu. Rev. Biochem.. 61:356-386.

GOLDBERG ET AL.-METHYLATION OF TRYPANOSOME PROTEIN 35 1
8. Dunten, I? & Koshland, D. J. 1991. Tuning the responsiveness of a sensory receptor via covalent modification. J. Biol. Chem., 266: 1491- 1496.
9. Finkelstein, J. D. 1990. Methionine metabolism in mammals. J. Nutr. Biochem., 1:228-237.
10. Goldberg, B., Rattendi, D., Yarlett, N., Lloyd, D. & Bacchi, C. J. 1997. Effects of carboxymethylation and polyamine synthesis inhib- itors on methylation of Trypanosoma brucei brucei cellular proteins and lipids. J. Euk. Microbiol., 44:352-358.
11. Hirumi, H. & Hirumi, K. 1989. Continuous cultivation of Trypanosoma brucei bloodstream forms in a medium containing a low concentration of serum protein without feeder layer cells. J. Parasitol.,
12. Hoffman, R. M. 1985. Altered methionine metabolism and transmethylation in cancer. Anticancer Res., 5: 1-30.
13. Hrycyna, C. A. & Clarke, S. 1993. Modification of eukaryotic signaling proteins by C-terminal methylation reactions. Pharmac. Ther., 59281-300.
14. Kuzoe, E A. S. 1993. Current situation of African trypanoso- miasis. Acta Tropica, 54: 153-162.
15. Lanham, S. M. & Godfrey, D. G. 1970. Isolation of salivarian trypanosomes from man and other animals using DEAE cellulose. Ex- per. Parasitol., 28321-523.
16. Lawrence, E & Robert-Gero, M. 1993. Distribution of macro- molecular methylations in promastigotes of Leishmania donovani and impact of sinefungin. J. Euk. Microbiol., 4058 1-589.
17. Lowry, 0. H., Rosebrough, N. J., Farr, A. L. & Randall, R. J. 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193:265-275.
18. Najbauer, J., Johnson, B. A. & Aswad, D. W. 1992. Analysis of
75~985-989.
stable protein methylation in cultured cells. Arch. Biochem. Biophys.,
19. Paik, W. K. & Kim, S. 1990. Reevaluation of the enzymology of protein methylation. In: Paik, W. K. & Kim, S. (ed.), Protein Meth- ylation. Boca Raton, CRC Press. Pp. 23-32.
20. Patnaik, I? K., Field, M. C., Menon, A. K., Cross, G . A. M., Yee, M. C. & Butikofer, F! 1993. Molecular species analysis of phospholip- ids from Trypanosoma brucei bloodstream and procyclic forms. Mol. Biochem., Parasitol., 58:97- 106.
21. Razin, A. & Cedar, H. 1991. DNA methylation and gene ex- pression. Microbiological Rev., 55:45 1-458.
22. Stramentinoli, G. 1987. Pharmacologic aspects of S-adenosyl- methionine pharmacokinetics and pharmacodynamics. Am. J. Med.,
23. van Waarde, A. & van Hoof, P. J. M. 1985. Pitfalls in the mea- surement of protein carboxyl methylation during chemotaxis of Dic- tyostelium discoideum. Biochim. Biophys. Acta. 840:344-354.
24. van Waarde, A. 1987. What is the function of protein carboxyl methylation? Comp. Biochem. Physiol., 86B:423-438.
25. Yarlett, N. & Bacchi, C. J. 1988. Effects of DL-a-difluorome- thylornithine on methionine cycle intermediates in Trypanosoma brucei brucei. Mol. Biochem. Parasitol., 27: 1-10,
26. Yarlett, N., Quamina, A. & Bacchi, C. J. 1991. Protein meth- ylases in Trypanosoma brucei brucei: activities and response to DL-a- difluoromethylornithine. J . Gen. Microbiol., 137:7 17-724.
27. Yarlett, N., Garofalo, J., Goldberg, B., Ciminelli, M. A,, Ruggi- ero, V., Sufrin, I. R. & Bacchi, C. J. 1993. S-Adenosylmethionine synthetase in bloodstream Trypanosoma brucei. Biochim. Biophys. Acta, 1181:68-76.
293: 85-92.
83(S~pp1.5A):35-42.
Received 7-8-96, 1-24-97; accepted 3-24-97