Embed Size (px)
Citation preview

Rab GAP cascade regulates dynamics of Ypt6 in theGolgi trafficYasuyuki Sudaa,1, Kazuo Kurokawaa, Ryogo Hirataa,2, and Akihiko Nakanoa,b
aLive Cell Molecular Imaging Research Team, RIKEN Center for Advanced Photonics, Wako, Saitama 351-0198, Japan; and bDepartment of Biological Sciences,Graduate School of Science, The University of Tokyo, Bunkyo-ku, Tokyo 113-0033, Japan
Edited by Suzanne R. Pfeffer, Stanford University School of Medicine, Stanford, CA, and accepted by the Editorial Board October 10, 2013 (received for reviewMay 8, 2013)
The Golgi apparatus functions as the central station of membranetraffic in cells, where newly synthesized proteins moving alongthe secretory pathway merge with proteins recycled from sub-sequent membrane organelles such as endosomes. A series of RabGTPases act consecutively and in concert with the maturation ofcis- to-trans cisternae of the Golgi apparatus. Rab GTPases controlvarious steps in intracellular membrane traffic by recruiting down-stream effector proteins. Here, we report the dynamics of Ypt6,a yeast member of the Rab GTPase family, which mediates thefusion of vesicles from endosomes at the Golgi apparatus. Ypt6resides temporarily at the Golgi and dissociates into the cytosolupon arrival of Ypt32, another Rab GTPase functioning in the lateGolgi. We found that Gyp6, a putative GTPase-activating protein(GAP) for Ypt6, specifically interacts with Ypt32, most likely as aneffector. Disruption of GYP6 or introduction of a Rab–GAP activity-deficient mutation in GYP6 resulted in continual residence of Ypt6at the Golgi. We propose that Ypt32 acts to terminate endosome-to-Golgi traffic through a Rab–GAP cascade as it does for cis-to-trans intra-Golgi traffic. Simultaneous disruption of GAP for early-acting Rab proteins in the Golgi showed appreciable defects inpost-Golgi trafficking, but did not significantly affect cell growth.
Rab GTPase cycles between the active, membrane-boundform and the inactive, cytosolic form via the action of in-
trinsic GTP exchange factor (GEF) and GTPase-activatingprotein (GAP). Once Rab GTPase is activated and targeted tothe membrane, various downstream effectors are recruited ontothe membrane to fulfill various intracellular membrane traf-ficking processes (1). Each Rab GTPase exhibits a unique spa-tiotemporal localization pattern at a particular membranouscompartment to precisely regulate vesicular traffic. Thus, RabGTPase and its effectors define a compartment’s identity (2, 3).The Rab–GEF cascade consists of sequential regulation of
Rab GTPases in which an upstream Rab recruits a GEF fordownstream Rab activation. This continuous activation of Rabproteins occurs at several organelles such as endosomes, trans-port vesicles, and the Golgi apparatus. For example, Rab5recruits the Mon1–Ccz1 complex, which is a GEF for Rab7 andfacilitates Rab5-Rab7 conversion during early to late endosomematuration in yeast and animal cells (4, 5). In the exocyticpathway of the budding yeast Saccharomyces cerevisiae, transitionfrom Ypt1 to Ypt31/32 occurs at the Golgi apparatus by re-cruitment of Ypt31/32 GEF as a Ypt1 effector (6). Once Ypt31/32 is activated, Sec2, a GEF for Sec4, is recruited onto themembrane to activate Sec4 at post-Golgi transport vesicles (7).Although the Rab–GEF cascade regulates the continuity ofdifferent Rab GTPases in such organelles (1), it also potentiallycreates instability because continuous activation of early RabGTPase leads to the concomitant existence of two Rab proteinson the same membrane. Recent work has provided evidence fora Rab–GAP cascade, in which a GAP of a former Rab is theeffector of subsequent Rab proteins at the Golgi apparatus inyeast (8). Termination of early acting Ypt1 is facilitated by therecruitment of Gyp1, a GAP for Ypt1, through binding to Ypt32as an effector at a single cisterna of the Golgi.This counter current activity of Rab GTPases could be a main
mechanism of compartment maturation (9, 10). Because Rab
defines the identity of a membrane compartment through the re-cruitment of effector proteins, the transition from one Rab to an-other changes the properties of the membrane where they exist. Inthe Golgi apparatus, the disruption of the boundary between twoRab proteins would affect cisternal maturation. This appears to beindeed the case in yeast in the transition of an early acting Rab-containing compartment to another containing a late-acting Rab(8).Nevertheless,Gyp1,aGAPresponsible for theRab–GAPcascadein the Golgi, is dispensable for yeast cell growth. Other mechanismsmay exist, in which transient contact between two compartmentscontributes to the maturation of the Golgi apparatus (11).Ypt6, the yeast counterpart of Rab6, has been reported to as-
sociate with the Golgi apparatus and regulate the fusion of vesi-cles from endosomes (12). In mammalian cells, accumulatingevidence suggests that Rab6 localizes at the trans-Golgi regionand functions in retrograde protein transport and in Golgi struc-tural organization (13, 14). Ypt6 may very well function in the yeastGolgi maintenance. Ypt6 is activated in to the GTP-bound state bythe Ric1–Rgp1 GEF complex and associates with the Golgi mem-brane, where it recruits a tethering factor called the Golgi-associ-ated retrograde protein (GARP) complex (15, 16). S. cerevisiae haseight Rab GAP proteins containing the conserved Tre2-Bub2-Cdc16 (TBC) domain, which provides the catalytic activity towardRab GTPases. Putative GAP proteins for Ypt6, including Gyp6,have been identified and characterized in vitro (17, 18). However,TBC domain-containing proteins have broad GAP activities towardmultiple Rab proteins in contrast to the high specificity betweenRab–GEF pairs (19). In the case of Ypt6, not only Gyp6 but alsoGyp2 and Gyp8 show in vitro GAP activity toward Ypt6 (20, 21).Thus, the precise mechanisms controlling Ypt6 dynamics and the
Significance
The Golgi apparatus functions as the central station of mem-brane traffic in cells. A series of Rab GTPases, which controlvarious steps in membrane traffic, act consecutively during thecourse of Golgi maturation. Here, we report that Ypt6, a Rab6homologue in yeast, resides temporarily at the Golgi and dis-sociates into the cytosol upon arrival of Ypt32, another RabGTPase functioning in the late Golgi. We have found that Gyp6,a putative GTPase-activating protein for Ypt6, specifically inter-acts with Ypt32 as an effector. Taken together with the pre-viously proposed Rab cascade within the Golgi, we propose thatmultiple Rab cascades interact at the intersection of secretoryand endosomal pathways and play significant roles in trafficwithin and around the Golgi apparatus.
Author contributions: Y.S., K.K., R.H., and A.N. designed research; Y.S. performed re-search; Y.S. analyzed data; and Y.S. and A.N. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission. S.R.P. is a guest editor invited by the EditorialBoard.1To whom correspondence should be addressed. E-mail: [email protected] address: Chemical Biology Laboratory, RIKEN, Wako, Saitama 351-0198, Japan.
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1308627110/-/DCSupplemental.
18976–18981 | PNAS | November 19, 2013 | vol. 110 | no. 47 www.pnas.org/cgi/doi/10.1073/pnas.1308627110
relationship between multiple Rab GTPases in the Golgi apparatusremain obscure.Here, we report roles of Rab–GAP in regulation of Ypt6 dy-
namics in S. cerevisiae. We have found that Ypt6 resides at the me-dial-Golgi and dissociates from themembrane at the onset ofYpt31/32 arrival during maturation. Gyp6, a putative GAP for Ypt6, bindsto Ypt32 and is recruited to the membrane, facilitating the dissoci-ation of Ypt6. This counteracting regulation of Ypt6 requires theGAP activity of Gyp6, suggesting that a Rab–GAP cascade definesthe membrane traffic from the endosomes to the Golgi apparatus.
ResultsYpt6 Resides in the Boundary Between the Cis- and Trans-Golgi.Rab–GEF and –GAP cascades have been proposed to regulate thesequential activation and deactivation of the two Rab GTPases,Ypt1 and Ypt31/32, within the Golgi apparatus (6, 8). However,the regulation of another Rab GTPase, Ypt6, which is a Rab6homolog and mediates the fusion of vesicles from endosomesto the Golgi apparatus, is obscure. Because Golgi cisternae inS. cerevisiae are unstacked and dispersed in the cytoplasm (22),the timing and duration of Ypt6 recruitment to the Golgi can beefficiently estimated by measuring the colocalization with knownGolgi cisternae markers. We analyzed the extent of colocalizationof Rab GTPases with cis- and trans-Golgi resident proteins, cis-
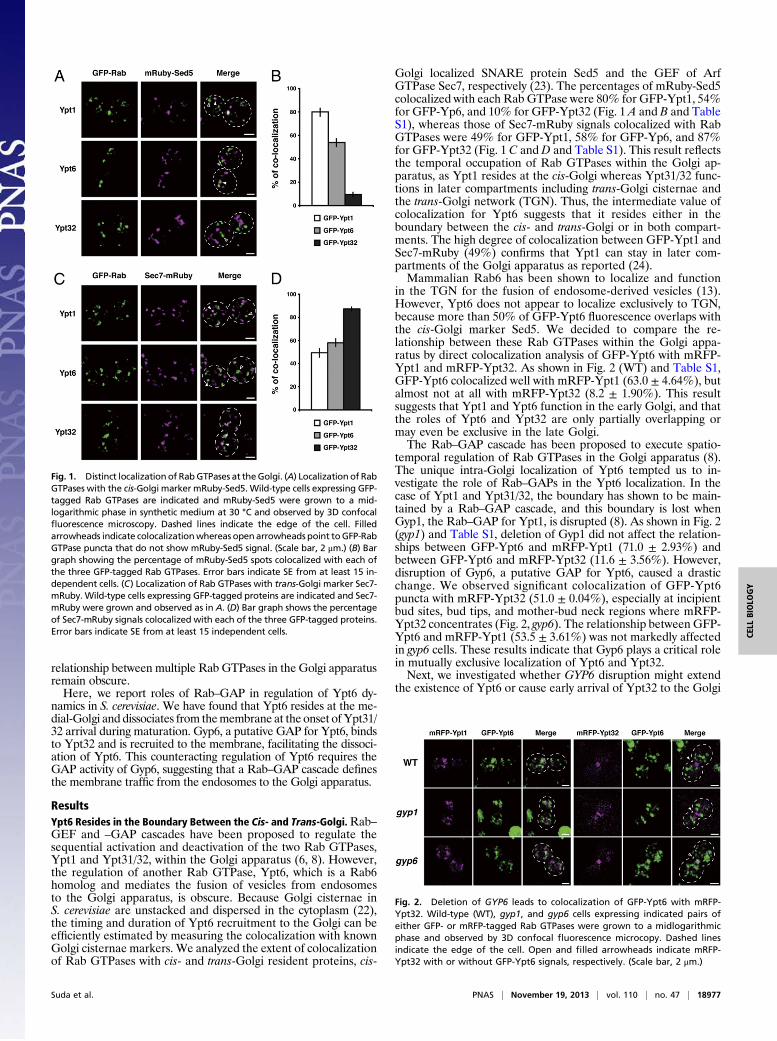
Golgi localized SNARE protein Sed5 and the GEF of ArfGTPase Sec7, respectively (23). The percentages of mRuby-Sed5colocalized with each Rab GTPase were 80% for GFP-Ypt1, 54%for GFP-Yp6, and 10% for GFP-Ypt32 (Fig. 1 A and B and TableS1), whereas those of Sec7-mRuby signals colocalized with RabGTPases were 49% for GFP-Ypt1, 58% for GFP-Yp6, and 87%for GFP-Ypt32 (Fig. 1 C andD and Table S1). This result reflectsthe temporal occupation of Rab GTPases within the Golgi ap-paratus, as Ypt1 resides at the cis-Golgi whereas Ypt31/32 func-tions in later compartments including trans-Golgi cisternae andthe trans-Golgi network (TGN). Thus, the intermediate value ofcolocalization for Ypt6 suggests that it resides either in theboundary between the cis- and trans-Golgi or in both compart-ments. The high degree of colocalization between GFP-Ypt1 andSec7-mRuby (49%) confirms that Ypt1 can stay in later com-partments of the Golgi apparatus as reported (24).Mammalian Rab6 has been shown to localize and function
in the TGN for the fusion of endosome-derived vesicles (13).However, Ypt6 does not appear to localize exclusively to TGN,because more than 50% of GFP-Ypt6 fluorescence overlaps withthe cis-Golgi marker Sed5. We decided to compare the re-lationship between these Rab GTPases within the Golgi appa-ratus by direct colocalization analysis of GFP-Ypt6 with mRFP-Ypt1 and mRFP-Ypt32. As shown in Fig. 2 (WT) and Table S1,GFP-Ypt6 colocalized well with mRFP-Ypt1 (63.0 ± 4.64%), butalmost not at all with mRFP-Ypt32 (8.2 ± 1.90%). This resultsuggests that Ypt1 and Ypt6 function in the early Golgi, and thatthe roles of Ypt6 and Ypt32 are only partially overlapping ormay even be exclusive in the late Golgi.The Rab–GAP cascade has been proposed to execute spatio-
temporal regulation of Rab GTPases in the Golgi apparatus (8).The unique intra-Golgi localization of Ypt6 tempted us to in-vestigate the role of Rab–GAPs in the Ypt6 localization. In thecase of Ypt1 and Ypt31/32, the boundary has shown to be main-tained by a Rab–GAP cascade, and this boundary is lost whenGyp1, the Rab–GAP for Ypt1, is disrupted (8). As shown in Fig. 2(gyp1) and Table S1, deletion of Gyp1 did not affect the relation-ships between GFP-Ypt6 and mRFP-Ypt1 (71.0 ± 2.93%) andbetween GFP-Ypt6 and mRFP-Ypt32 (11.6 ± 3.56%). However,disruption of Gyp6, a putative GAP for Ypt6, caused a drasticchange. We observed significant colocalization of GFP-Ypt6puncta with mRFP-Ypt32 (51.0 ± 0.04%), especially at incipientbud sites, bud tips, and mother-bud neck regions where mRFP-Ypt32 concentrates (Fig. 2, gyp6). The relationship between GFP-Ypt6 and mRFP-Ypt1 (53.5 ± 3.61%) was not markedly affectedin gyp6 cells. These results indicate that Gyp6 plays a critical rolein mutually exclusive localization of Ypt6 and Ypt32.Next, we investigated whether GYP6 disruption might extend
the existence of Ypt6 or cause early arrival of Ypt32 to the Golgi
Fig. 1. Distinct localization of RabGTPases at theGolgi. (A) Localization of RabGTPases with the cis-Golgi marker mRuby-Sed5. Wild-type cells expressing GFP-tagged Rab GTPases are indicated and mRuby-Sed5 were grown to a mid-logarithmic phase in synthetic medium at 30 °C and observed by 3D confocalfluorescence microscopy. Dashed lines indicate the edge of the cell. Filledarrowheads indicate colocalizationwhereas openarrowheads point toGFP-RabGTPase puncta that do not show mRuby-Sed5 signal. (Scale bar, 2 μm.) (B) Bargraph showing the percentage of mRuby-Sed5 spots colocalized with each ofthe three GFP-tagged Rab GTPases. Error bars indicate SE from at least 15 in-dependent cells. (C) Localization of Rab GTPases with trans-Golgi marker Sec7-mRuby. Wild-type cells expressing GFP-tagged proteins are indicated and Sec7-mRuby were grown and observed as in A. (D) Bar graph shows the percentageof Sec7-mRuby signals colocalized with each of the three GFP-tagged proteins.Error bars indicate SE from at least 15 independent cells.
Fig. 2. Deletion of GYP6 leads to colocalization of GFP-Ypt6 with mRFP-Ypt32. Wild-type (WT), gyp1, and gyp6 cells expressing indicated pairs ofeither GFP- or mRFP-tagged Rab GTPases were grown to a midlogarithmicphase and observed by 3D confocal fluorescence microcopy. Dashed linesindicate the edge of the cell. Open and filled arrowheads indicate mRFP-Ypt32 with or without GFP-Ypt6 signals, respectively. (Scale bar, 2 μm.)
Suda et al. PNAS | November 19, 2013 | vol. 110 | no. 47 | 18977
CELL
BIOLO
GY
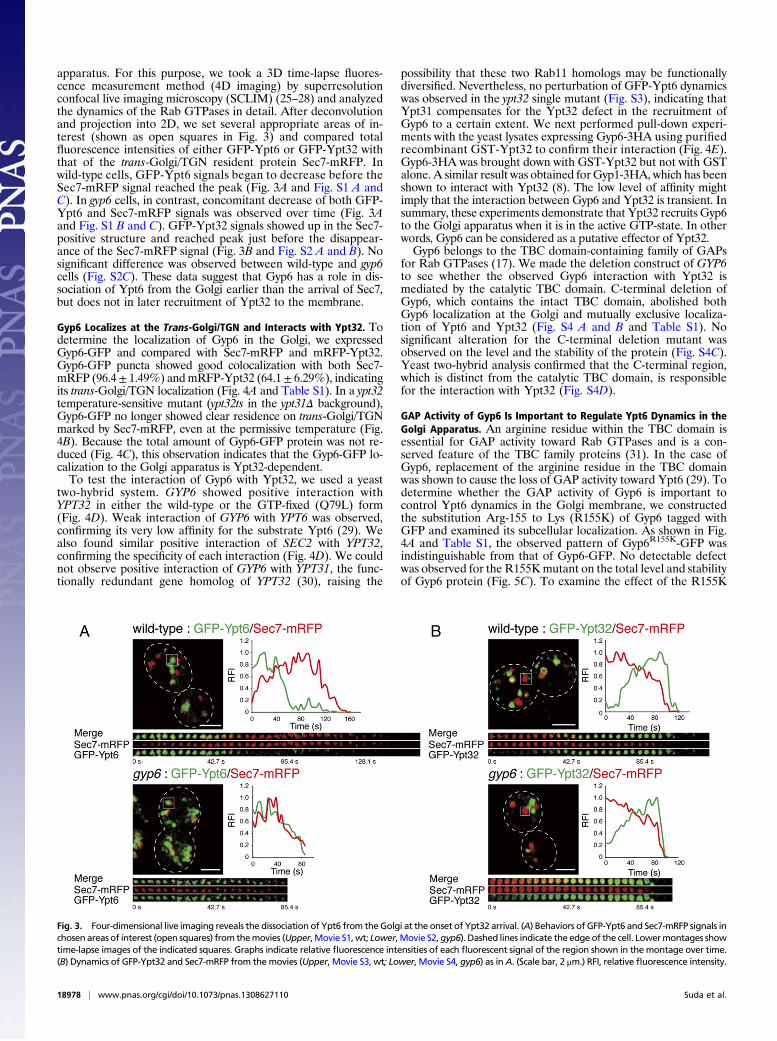
apparatus. For this purpose, we took a 3D time-lapse fluores-cence measurement method (4D imaging) by superresolutionconfocal live imaging microscopy (SCLIM) (25–28) and analyzedthe dynamics of the Rab GTPases in detail. After deconvolutionand projection into 2D, we set several appropriate areas of in-terest (shown as open squares in Fig. 3) and compared totalfluorescence intensities of either GFP-Ypt6 or GFP-Ypt32 withthat of the trans-Golgi/TGN resident protein Sec7-mRFP. Inwild-type cells, GFP-Ypt6 signals began to decrease before theSec7-mRFP signal reached the peak (Fig. 3A and Fig. S1 A andC). In gyp6 cells, in contrast, concomitant decrease of both GFP-Ypt6 and Sec7-mRFP signals was observed over time (Fig. 3Aand Fig. S1 B and C). GFP-Ypt32 signals showed up in the Sec7-positive structure and reached peak just before the disappear-ance of the Sec7-mRFP signal (Fig. 3B and Fig. S2 A and B). Nosignificant difference was observed between wild-type and gyp6cells (Fig. S2C). These data suggest that Gyp6 has a role in dis-sociation of Ypt6 from the Golgi earlier than the arrival of Sec7,but does not in later recruitment of Ypt32 to the membrane.
Gyp6 Localizes at the Trans-Golgi/TGN and Interacts with Ypt32. Todetermine the localization of Gyp6 in the Golgi, we expressedGyp6-GFP and compared with Sec7-mRFP and mRFP-Ypt32.Gyp6-GFP puncta showed good colocalization with both Sec7-mRFP (96.4± 1.49%) andmRFP-Ypt32 (64.1± 6.29%), indicatingits trans-Golgi/TGN localization (Fig. 4A and Table S1). In a ypt32temperature-sensitive mutant (ypt32ts in the ypt31Δ background),Gyp6-GFP no longer showed clear residence on trans-Golgi/TGNmarked by Sec7-mRFP, even at the permissive temperature (Fig.4B). Because the total amount of Gyp6-GFP protein was not re-duced (Fig. 4C), this observation indicates that the Gyp6-GFP lo-calization to the Golgi apparatus is Ypt32-dependent.To test the interaction of Gyp6 with Ypt32, we used a yeast
two-hybrid system. GYP6 showed positive interaction withYPT32 in either the wild-type or the GTP-fixed (Q79L) form(Fig. 4D). Weak interaction of GYP6 with YPT6 was observed,confirming its very low affinity for the substrate Ypt6 (29). Wealso found similar positive interaction of SEC2 with YPT32,confirming the specificity of each interaction (Fig. 4D). We couldnot observe positive interaction of GYP6 with YPT31, the func-tionally redundant gene homolog of YPT32 (30), raising the
possibility that these two Rab11 homologs may be functionallydiversified. Nevertheless, no perturbation of GFP-Ypt6 dynamicswas observed in the ypt32 single mutant (Fig. S3), indicating thatYpt31 compensates for the Ypt32 defect in the recruitment ofGyp6 to a certain extent. We next performed pull-down experi-ments with the yeast lysates expressing Gyp6-3HA using purifiedrecombinant GST-Ypt32 to confirm their interaction (Fig. 4E).Gyp6-3HA was brought down with GST-Ypt32 but not with GSTalone. A similar result was obtained forGyp1-3HA, which has beenshown to interact with Ypt32 (8). The low level of affinity mightimply that the interaction between Gyp6 and Ypt32 is transient. Insummary, these experiments demonstrate that Ypt32 recruits Gyp6to the Golgi apparatus when it is in the active GTP-state. In otherwords, Gyp6 can be considered as a putative effector of Ypt32.Gyp6 belongs to the TBC domain-containing family of GAPs
for Rab GTPases (17). We made the deletion construct of GYP6to see whether the observed Gyp6 interaction with Ypt32 ismediated by the catalytic TBC domain. C-terminal deletion ofGyp6, which contains the intact TBC domain, abolished bothGyp6 localization at the Golgi and mutually exclusive localiza-tion of Ypt6 and Ypt32 (Fig. S4 A and B and Table S1). Nosignificant alteration for the C-terminal deletion mutant wasobserved on the level and the stability of the protein (Fig. S4C).Yeast two-hybrid analysis confirmed that the C-terminal region,which is distinct from the catalytic TBC domain, is responsiblefor the interaction with Ypt32 (Fig. S4D).
GAP Activity of Gyp6 Is Important to Regulate Ypt6 Dynamics in theGolgi Apparatus. An arginine residue within the TBC domain isessential for GAP activity toward Rab GTPases and is a con-served feature of the TBC family proteins (31). In the case ofGyp6, replacement of the arginine residue in the TBC domainwas shown to cause the loss of GAP activity toward Ypt6 (29). Todetermine whether the GAP activity of Gyp6 is important tocontrol Ypt6 dynamics in the Golgi membrane, we constructedthe substitution Arg-155 to Lys (R155K) of Gyp6 tagged withGFP and examined its subcellular localization. As shown in Fig.4A and Table S1, the observed pattern of Gyp6R155K-GFP wasindistinguishable from that of Gyp6-GFP. No detectable defectwas observed for the R155Kmutant on the total level and stabilityof Gyp6 protein (Fig. 5C). To examine the effect of the R155K
Fig. 3. Four-dimensional live imaging reveals the dissociation of Ypt6 from theGolgi at the onset of Ypt32 arrival. (A) Behaviors of GFP-Ypt6 and Sec7-mRFP signals inchosen areas of interest (open squares) from themovies (Upper,Movie S1,wt; Lower,Movie S2,gyp6). Dashed lines indicate the edge of the cell. Lowermontages showtime-lapse images of the indicated squares. Graphs indicate relative fluorescence intensities of each fluorescent signal of the region shown in themontage over time.(B) Dynamics of GFP-Ypt32 and Sec7-mRFP from themovies (Upper, Movie S3, wt; Lower, Movie S4, gyp6) as inA. (Scale bar, 2 μm.) RFI, relative fluorescence intensity.
18978 | www.pnas.org/cgi/doi/10.1073/pnas.1308627110 Suda et al.

mutation of Gyp6 on the dissociation of Ypt6 from the Golgimembrane, we examined GFP-Ypt6 dynamics relative to Sec7-mRFP by 4D imaging using SCLIM, in the gyp6 cells expressingeither Gyp6-3HA or Gyp6R155K-3HA. As shown in Fig. 5A andFig. S5 A and C, GFP-Ypt6 dissociation occurred in a similarfashion to the wild type in Gyp6-3HA cells, confirming that theGyp6-3HA construct was functional. However, in the Gyp6R155K-3HA–expressing cells, GFP-Ypt6 failed to reduce the signalsbefore the signal reduction of Sec7-mRFP (Fig. 5B and Fig. S5 BandC). These observations demonstrate that the GAP activity ofGyp6 is critical for the dissociation of Ypt6 during the courseof Golgi maturation.
Failure of Rab Transition Perturbs Post-Golgi Membrane Trafficking,but Does Not Significantly Affect the Cisternal Maturation of theGolgi. The Rab conversion has been proposed as a key charac-teristic of compartment maturation (4, 32, 33). We examinedwhether the cisternal maturation of the Golgi is affected by thedisruption of the boundary between Golgi-resident Rab GTPa-ses, by looking at the gyp1 gyp6 double mutant. In gyp1 gyp6 cells,Ypt1, Ypt6, and Ypt32 appear to more or less colocalize in theGolgi (Fig. 6A and Table S1). This is a clear piece of evidence
confirming the disruption of the boundary between the RabGTPases. To estimate the rates of maturation, we marked Golgicisternae with GFP-Gos1 (medial-Golgi) and Sec7-mRFP (trans-Golgi) (23), and then measured the ratio of the red/green fluo-rescence transition and calculated its doubling time. The colortransition of cisternae in wild-type cells had a ratio doubling timeof 22.5 ± 8.3 s (±SD; n = 13), whereas in gyp1 gyp6 cells it was∼34% increased compared with wild-type cells, 30.1 ± 5.9 s (n =13) (Fig. 6B). Statistically, the difference of these values was notsignificant (Student t test, P > 0.05). These observations suggestthat efficient transition of these Rab GTPases at the Golgi is notcritical but may be important for the speed of maturation.We also analyzed protein recycling and export in gyp1 gyp6
cells. Snc1 cycles between post-Golgi organelles and the plasmamembrane and primarily localizes to the plasma membrane (34).The localization of GFP-Snc1 was normal in gyp6 cells, whereasaccumulation of internal structures was seen in gyp1 and gyp1 gyp6cells (Fig. S6A), suggesting recycling defects in these mutants. Wealso observed mislocalization of Vps10, a sorting receptor forvacuolar hydrolases, which cycles between the late-Golgi andprevacuolar endosome-like compartments (PVCs) (35). Vps10-3xGFP was found to be in punctate structures in wild-type andgyp6 cells as reported previously. In gyp1 and gyp1 gyp6 cells,Vps10-3xGFP was mislocalized to the vacuolar membrane andoccasionally to perivacuolar dot-like structures (Fig. S6B). Thedot-like structures are similar to the exaggerated PVC-like struc-tures characteristic of the mutants that are defective in the proteinexport from the PVC, also suggesting recycling defects. Colony blotanalysis (36) indicated gyp1, gyp6, and gyp1 gyp6 cells showedweak but detectable missecretion of caboxypeptidase Y (Fig. S6C).Additionally, activity-staining gel electrophoresis indicated thatinvertase is underglycosylated in gyp1 and gyp1 gyp6 mutants (Fig.S6D). These results indicate that the perturbation of the Rab reg-ulation at the Golgi appreciably affected the Golgi functions butwas not deleterious to the cells.
DiscussionIntracellular logistics based on membrane traffic is tightly regu-lated to ensure cellular homeostasis. Rab GTPases play pivotalroles in regulation of multiple traffic events (1). They show strict
Fig. 4. Gyp6 resides in the Golgi and interacts with Ypt32 as a putativeeffector. (A) Colocalization of Gyp6-GFP with Sec7-mRFP. Wild-type cellsexpressing Gyp6-GFP or Gyp6R155K-GFP and either Sec7-mRFP or mRFP-Ypt32were grown to a midlogarithmic phase, and imaged by 3D confocal fluores-cence microscopy. Dashed lines indicate the edge of the cell. (Scale bar, 2 μm.)(B) ypt32 temperature-sensitivemutant cells (ypt32ts) expressingGyp6-GFP andSec7-mRFP were grown at 25 °C to a midlogarithmic phase and imaged byfluorescence microscopy as in A. (C) Total lysates of wild-type and ypt32ts cellsexpressing Gyp6-GFP after incubation either at 25 °C or 37 °C for 30 min wereseparated by SDS/PAGE followed by immunoblotting analysis using anti-GFPantibodies. Dolichol phosphate mannose synthase, Dpm1, was also probed asa control. (D) Yeast two-hybrid assay between GYP6 and Rab GTPase alleles ofwild-type GTP- and GDP-fixed states. Growth on the plate lacking tryptophan/leucine (WL) indicates the cells harboring plasmids. Growth on the plate lackingtryptophan/leucine/histidine (WLH) with or without 5 mM 3-AT indicatesa positive interaction. (E) Pull-down of Gyp6-3HA and Gyp1-3HA by GST-tag-ged Ypt32. Lysates from yeast cells overexpressing Gyp6-3HA or Gyp1-3HAwere incubated with glutathione Sepharose preincubated with either GST- orGST-Ypt32 in the presence of GTPγS. Samples were separated by SDS/PAGE andsignals were detected by immunoblotting using an antibody against HA. One-percent input of yeast total lysate was run as a control.
Fig. 5. Four-dimensional live imaging demonstrates the requirement ofGAP-activity of Gyp6 for the dissociation of Ypt6 from the Golgi. (A and B)Behaviors of GFP-Ypt6 and Sec7-mRFP from the movies (Upper, Movie S5,GYP6-3HA; Lower, Movie S6, gyp6R155K-3HA). Dashed lines indicate the edgeof the cell. Lower montages show time-lapse images of the areas marked bysquares. Graphs indicate relative fluorescence intensities of each fluorescentsignal of the region shown in the montage over time. (Scale bar, 2 μm.) (C)The steady-state levels of wild-type and R155K mutant Gyp6-3HA. Theamount of Gyp6-3HA was examined by Western blot analysis of the total celllysates using anti-HA monoclonal antibody. Dpm1 was used as a control.
Suda et al. PNAS | November 19, 2013 | vol. 110 | no. 47 | 18979
CELL
BIOLO
GY

localization when they are in the active GTP state and thus arebelieved to define the identity of compartments (2, 3). The dif-ferentiated roles of Rab GTPases in the Golgi apparatus are ofparticular interest because the mechanism of compartmentali-zation of the Golgi is still a question of dispute. High-resolutionlive imaging is a very powerful approach to address this question,and the budding yeast S. cerevisiae has been used as an advan-tageous organism for this purpose because its Golgi cisternae areunstacked and easy to resolve by fluorescence microscopy (22,37). Cisternae of the Golgi change their identity from cis to transnature by maturation (23, 38). Regarding Rab GTPases, Ypt1functions in cis-Golgi cisternae and Ypt31/32 reside in trans-Golgi cisternae/TGN, and the efficient transition from early-acting Ypt1 to subsequent Ypt31/32 is fulfilled by the action ofRab–GEF and –GAP cascades (8). In the present paper, wedemonstrate that another Rab–GAP cascade is present in theyeast Golgi apparatus in the transition from Ypt6 at the medial-Golgi to Ypt31/32 at the trans-Golgi/TGN.
Ypt6 Resides at the Medial-Golgi During Maturation. Ypt1 functionsin the fusion of ER-derived vesicles with the Golgi at cis-Golgicisternae (39, 40), whereas Ypt31/32 locate at the trans-Golgi/TGN and regulate various traffic events directly and indirectly,e.g., fusion of vesicles from endosomes (41), movement of theGolgi apparatus (42), and subsequent Sec4 activation throughrecruitment of Sec2, a GEF for Sec4 (7). Ypt6 is the yeast ho-molog of the mammalian Rab6, which functions in the fusion ofendosome-derived vesicles to the Golgi by recruiting the GARPcomplex as its effector (12, 15, 16). In mammals and plants, it isgenerally agreed that Rab6 functions in trans-Golgi/TGN (13,14). However, our results showed significant colocalization ofGFP-Ypt6 with mRuby-Sed5, suggesting that Ypt6 also localizesto cis-Golgi cisternae in yeast. Colocalization of GFP-Ypt6 withSec7-mRFP ensures that Ypt6 does localize to trans-Golgi/TGN,however, GFP-Ypt6 was scarcely colocalized with the trans-Golgi/TGN-localized Rab GTPase Ypt32. Live imaging of thedynamics of GFP-Ypt6 and GFP-Ypt32 indicated that Ypt6dissociates from the Golgi at the onset of Ypt32 arrival. To-gether, our data suggest that Ypt6 resides and most probablyfunctions in the medial-Golgi. Ypt6 and Ypt31/32 are bothreported to function in the fusion of vesicles that contain diversesets of cargo proteins such as Snc1 and Vps10 from endosomes(12, 34, 41, 43). Budding yeast might have used a temporal
hierarchy between these Rab GTPases for strict sorting of a di-verse set of cargo proteins upon their arrival at the Golgi.
Ypt32 Recruits Gyp6 to the Golgi as a Putative Effector. We haveshown that Gyp6-GFP colocalizes with trans-Golgi resident pro-teins, Sec7-mRFP and mRFP-Ypt32. This is inconsistent witha previous study claiming the presence of Gyp6 in the endosomal/prevacuolar compartment (44). Considering the role of Gyp6 asa Ypt6 GAP, its Golgi localization looks reasonable. The Golgilocalization of Gyp6 is lost when the Ypt31/32 function is com-promised by a temperature-sensitive mutation. This indicates thatYpt31/32 is important for the localization of Gyp6. The interactionbetween Ypt32 and Gyp6 has been confirmed by yeast two-hybridassay and pull-down experiments, although the nucleotide-dependentinteraction between Ypt32 and Gyp6 has not been detected in vitro.The C-terminal region of Gyp6, which is distinct from the catalyticTBC domain, is responsible for the interaction. The GTP form ofYpt32 appears to be preferred in this interaction in vivo, while nodetectable GAP activity has been reported for Gyp6 toward Ypt32 invitro (19).We have also detected weak interaction betweenGyp6 andYpt6 by yeast two-hybrid, confirming a low affinity of Gyp6 towardYpt6 in vitro (29). These observations strongly suggest that Ypt32binds Gyp6 as a putative effector.
The Rab–GAP Cascade Between Ypt6 and Ypt31/32.We show that thedeletion of GYP6 leads to the coexistence of Ypt6 and Ypt32 inthe Golgi, particularly at the bud neck. This unusual situation isprobably caused by the continual activation of Ypt6 in the post-Golgi transport vesicles that accumulate at the site of cytokinesis.Among eight TBC-domain containing putative GAP proteins inyeast (17, 18), only the GYP6 deletion resulted in such a defect inour analysis. Gyp2 was also reported as a possible GAP for Ypt6(20, 45), but the gyp2 mutant did not show any defect in Ypt6localization (Fig. S7). Deletion of GYP1 did not perturb themutually exclusive localization between Ypt6 and Ypt32 either.This is in agreement with the endocytic events in which dele-tion of Msb3/Gyp3 effects only the localization of Vps21 (19).Taken together, the Rab–GAP substrate specificity might bemuch more rigorous than what has been predicted from in vitrobinding experiments.Our live-cell imaging analysis by SCLIM has demonstrated that
Gyp6 is essential for temporal occupation of Ypt6 at the Golgi,probably at the medial cisternae. The conserved arginine residuein the TBC domain of Gyp6, which is reported important for itsGAP activity toward Ypt6 (29), is critical for this role of Gyp6.Taking all these data together, we conclude that a Rab–GAPcascade exists between Ypt6 and Ypt32.Localization of Ypt1 and Ypt32 overlaps in the Golgi (8), but
that of Ypt6 and Ypt32 is exclusive. Presence of a Rab–GEFcascade for Ypt1/Ypt32 may explain this difference. In mamma-lian cells, a putative Rab–GEF cascade has been proposed be-tween medial-Golgi-localized Rab33B and the trans-Golgi proteinRab6A via recruitment of Ric1, a component of the GEF complexfor Rab6 (14, 46). Whether a similar mechanism might also existin yeast or not awaits further investigation.
Continual Activation of Multiple Rab Proteins at the Golgi Has onlya Slight Effect on Golgi Maturation.We found that the perturbationof Rab–GAP cascade affects normal Golgi function. The simul-taneous deletion of Gyp1 and Gyp6, which strongly affected thenormal localization of the three Golgi Rab GTPases in yeast,resulted in some appreciable transport defects of both secretoryand recycling cargoes. Our results suggest that coordinatedexpression of the Rab GTPases is indeed important for the Golgifunction.Recent studies suggest that mammalian Rab6 is important
not only for membrane traffic but also for organization andmaintenance of the Golgi (13, 14). To examine whether con-current existence of the Golgi Rab GTPases affects the main-tenance of the Golgi structure and/or maturation of cisternae, weconstructed a gyp1 gyp6 double mutant, in which early-acting
Fig. 6. Simultaneous deletion of GYP1 and GYP6 does not affect the Golgicisternal maturation. (A) gyp1 gyp6 cells expressing indicated pairs of eitherGFP- or mRFP-tagged Rab GTPases were grown to a midlogarithmic phaseand observed by 3D confocal fluorescent microcopy. Dashed lines indicatethe edge of the cell. (Scale bar, 2 μm.) (B) A box plot representing the dis-tribution of the ratio-doubling time for the color conversion of cisternaefrom GFP-Gos1 to Sec7-mRFP in wild-type and gyp1 gyp6 cells. The hori-zontal line in each box represents the median value of the distribution. Theboundaries of a box represent the lower and upper quartile values. Thewhiskers extending vertically from the upper and lower portions of each boxrepresent the extent of the rest of the data. Student t test P value was 0.12 incomparison between the wild type and the gyp1 gyp6 double mutant.
18980 | www.pnas.org/cgi/doi/10.1073/pnas.1308627110 Suda et al.

Ypt1 and Ypt6 are not eliminated and thus coexist with the later-acting Ypt31/32 in the Golgi. Detailed analysis by 4D fluorescentmicroscopy using SCLIM shows that the maturation speed of theGolgi in gyp1 gyp6 mutant cells is slightly affected, but not sig-nificantly different from that of the wild type. Our observationmight imply the presence of additional fail-safe mechanism(s)that ensures the Golgi compartmentalization and maturation.
Materials and MethodsExperimental details are mostly described in SI Materials and Methods.
Yeast Strains and Media. Standard methods and media were used (47). Thestrains used in this study are listed in Table S2. All strains in this study, exceptAH109 for yeast two-hybrid assay, were from a YPH499 strain background
(48). Gene disruption and insertion were basically done by PCR-mediatedgene replacement (49, 50) and verified by PCR or phenotype.
Plasmids. The plasmids used in this study are listed in Table S3. Plasmid-basedfluorescent protein-tagged constructs were expressed under the control ofthe ADH1 promoter. Site-directed mutagenesis was performed by the gap-repair method (51, 52).
ACKNOWLEDGMENTS. We thank Kalai Madhi Muniandy for technical sup-port and all members of the A.N. laboratory for helpful comments. Thiswork was supported by a Grant-in-Aid for Specially Promoted Research(Grant 20001009 to A.N.) from the Ministry of Education, Culture, Sports,Science, and Technology of Japan and by Extreme Photonics and CellularSystems Biology Projects of RIKEN (The Institute of Physical and ChemicalResearch) (Japan) (A.N.).
1. Grosshans BL, Ortiz D, Novick PJ (2006) Rabs and their effectors: Achieving specificityin membrane traffic. Proc Natl Acad Sci USA 103(32):11821–11827.
2. Zerial M, McBride H (2001) Rab proteins as membrane organizers. Nat Rev Mol CellBiol 2(2):107–117.
3. Pfeffer SR (2001) Rab GTPases: Specifying and deciphering organelle identity andfunction. Trends Cell Biol 11(12):487–491.
4. Nordmann M, et al. (2010) The Mon1-Ccz1 complex is the GEF of the late endosomalRab7 homolog Ypt7. Curr Biol 20(18):1654–1659.
5. Gerondopoulos A, Langemeyer L, Liang J-R, Linford A, Barr FA (2012) BLOC-3 mutatedin Hermansky-Pudlak syndrome is a Rab32/38 guanine nucleotide exchange factor.Curr Biol 22(22):2135–2139.
6. Wang W, Ferro-Novick S (2002) A Ypt32p exchange factor is a putative effector ofYpt1p. Mol Biol Cell 13(9):3336–3343.
7. Ortiz D, Medkova M, Walch-Solimena C, Novick PJ (2002) Ypt32 recruits the Sec4pguanine nucleotide exchange factor, Sec2p, to secretory vesicles; evidence for a Rabcascade in yeast. J Cell Biol 157(6):1005–1015.
8. Rivera-Molina FE, Novick PJ (2009) A Rab GAP cascade defines the boundary betweentwo Rab GTPases on the secretory pathway. Proc Natl Acad Sci USA 106(34):14408–14413.
9. Nottingham RM, Pfeffer SR (2009) Defining the boundaries: Rab GEFs and GAPs. ProcNatl Acad Sci USA 106(34):14185–14186.
10. Segev N (2011) Coordination of intracellular transport steps by GTPases. Semin CellDev Biol 22(1):33–38.
11. Pfeffer SR (2010) How the Golgi works: A cisternal progenitor model. Proc Natl AcadSci USA 107(46):19614–19618.
12. Luo Z, Gallwitz D (2003) Biochemical and genetic evidence for the involvement ofyeast Ypt6-GTPase in protein retrieval to different Golgi compartments. J Biol Chem278(2):791–799.
13. Storrie B, et al. (2012) Electron tomography reveals Rab6 is essential to the traffickingof trans-Golgi clathrin and COPI-coated vesicles and the maintenance of Golgi cis-ternal number. Traffic 13(5):727–744.
14. Starr T, Sun Y, Wilkins N, Storrie B (2010) Rab33b and Rab6 are functionally over-lapping regulators of Golgi homeostasis and trafficking. Traffic 11(5):626–636.
15. Siniossoglou S, Peak-Chew SY, Pelham HRB (2000) Ric1p and Rgp1p form a complexthat catalyses nucleotide exchange on Ypt6p. EMBO J 19(18):4885–4894.
16. Siniossoglou S, Pelham HRB (2001) An effector of Ypt6p binds the SNARE Tlg1p andmediates selective fusion of vesicles with late Golgi membranes. EMBO J 20(21):5991–5998.
17. Neuwald AF (1997) A shared domain between a spindle assembly checkpoint proteinand Ypt/Rab-specific GTPase-activators. Trends Biochem Sci 22(7):243–244.
18. Strom M, Vollmer P, Tan TJ, Gallwitz D (1993) A yeast GTPase-activating protein thatinteracts specifically with a member of the Ypt/Rab family. Nature 361(6414):736–739.
19. Lachmann J, Barr FA, Ungermann C (2012) The Msb3 GAP controls the activity of theRab GTPases Vps21 and Ypt7 at endosomes and vacuoles. Mol Biol Cell 23(13):2516–2526.
20. Albert S, Gallwitz D (1999) Two new members of a family of Ypt/Rab GTPase acti-vating proteins. Promiscuity of substrate recognition. J Biol Chem 274(47):33186–33189.
21. De Antoni A, Schmitzová J, Trepte H-H, Gallwitz D, Albert S (2002) Significance of GTPhydrolysis in Ypt1p-regulated endoplasmic reticulum to Golgi transport revealed bythe analysis of two novel Ypt1-GAPs. J Biol Chem 277(43):41023–41031.
22. Suda Y, Nakano A (2012) The yeast Golgi apparatus. Traffic 13(4):505–510.23. Matsuura-Tokita K, Takeuchi M, Ichihara A, Mikuriya K, Nakano A (2006) Live imaging
of yeast Golgi cisternal maturation. Nature 441(7096):1007–1010.24. Sclafani A, et al. (2010) Establishing a role for the GTPase Ypt1p at the late Golgi.
Traffic 11(4):520–532.25. Okamoto M, et al. (2012) High-curvature domains of the ER are important for
the organization of ER exit sites in Saccharomyces cerevisiae. J Cell Sci 125(Pt 14):3412–3420.
26. Ito Y, et al. (2012) cis-Golgi proteins accumulate near the ER exit sites and act as thescaffold for Golgi regeneration after brefeldin A treatment in tobacco BY-2 cells. MolBiol Cell 23(16):3203–3214.
27. Nakano A, Luini A (2010) Passage through the Golgi. Curr Opin Cell Biol 22(4):471–478.
28. Kurokawa K, Ishii M, Suda Y, Ichihara A, Nakano A (2013) Live cell visualization ofGolgi membrane dynamics by super-resolution confocal live imaging microscopy(SCLIM). Methods Cell Biol, in press.
29. Will E, Gallwitz D (2001) Biochemical characterization of Gyp6p, a Ypt/Rab-specificGTPase-activating protein from yeast. J Biol Chem 276(15):12135–12139.
30. Benli M, Döring F, Robinson DG, Yang X, Gallwitz D (1996) Two GTPase isoforms,Ypt31p and Ypt32p, are essential for Golgi function in yeast. EMBO J 15(23):6460–6475.
31. Albert S, Will E, Gallwitz D (1999) Identification of the catalytic domains and theirfunctionally critical arginine residues of two yeast GTPase-activating proteins specificfor Ypt/Rab transport GTPases. EMBO J 18(19):5216–5225.
32. Rink J, Ghigo E, Kalaidzidis Y, Zerial M (2005) Rab conversion as a mechanism ofprogression from early to late endosomes. Cell 122(5):735–749.
33. Poteryaev D, Datta S, Ackema K, Zerial M, Spang A (2010) Identification of the switchin early-to-late endosome transition. Cell 141(3):497–508.
34. Lewis MJ, Nichols BJ, Prescianotto-Baschong C, Riezman H, Pelham HRB (2000) Specificretrieval of the exocytic SNARE Snc1p from early yeast endosomes.Mol Biol Cell 11(1):23–38.
35. Seaman MN, Marcusson EG, Cereghino JL, Emr SD (1997) Endosome to Golgi retrievalof the vacuolar protein sorting receptor, Vps10p, requires the function of the VPS29,VPS30, and VPS35 gene products. J Cell Biol 137(1):79–92.
36. Conibear E, Stevens TH (2002) Studying yeast vacuoles. Methods Enzymol 351:408–432.
37. Papanikou E, Glick BS (2009) The yeast Golgi apparatus: Insights and mysteries. FEBSLett 583(23):3746–3751.
38. Losev E, et al. (2006) Golgi maturation visualized in living yeast. Nature 441(7096):1002–1006.
39. Cao X, Ballew N, Barlowe C (1998) Initial docking of ER-derived vesicles requiresUso1p and Ypt1p but is independent of SNARE proteins. EMBO J 17(8):2156–2165.
40. Suvorova ES, Duden R, Lupashin VV (2002) The Sec34/Sec35p complex, a Ypt1p ef-fector required for retrograde intra-Golgi trafficking, interacts with Golgi SNAREsand COPI vesicle coat proteins. J Cell Biol 157(4):631–643.
41. Chen SH, et al. (2005) Ypt31/32 GTPases and their novel F-box effector protein Rcy1regulate protein recycling. Mol Biol Cell 16(1):178–192.
42. Lipatova Z, et al. (2008) Direct interaction between a myosin V motor and the RabGTPases Ypt31/32 is required for polarized secretion. Mol Biol Cell 19(10):4177–4187.
43. Seaman MN, McCaffery JM, Emr SD (1998) A membrane coat complex essential forendosome-to-Golgi retrograde transport in yeast. J Cell Biol 142(3):665–681.
44. Ali R, Brett CL, Mukherjee S, Rao R (2004) Inhibition of sodium/proton exchange bya Rab-GTPase-activating protein regulates endosomal traffic in yeast. J Biol Chem279(6):4498–4506.
45. Lafourcade C, Galan J-M, Peter M (2003) Opposite roles of the F-box protein Rcy1pand the GTPase-activating protein Gyp2p during recycling of internalized proteins inyeast. Genetics 164(2):469–477.
46. Pusapati GV, Luchetti G, Pfeffer SR (2012) Ric1-Rgp1 complex is a guanine nucleotideexchange factor for the late Golgi Rab6A GTPase and an effector of the medial GolgiRab33B GTPase. J Biol Chem 287(50):42129–42137.
47. Burke D, Dawson D, Stearns T (2000) Methods in Yeast Genetics, 2000 Edition (ColdSpring Harbor Laboratory Press, Plainview, NY).
48. Sikorski RS, Hieter P (1989) A system of shuttle vectors and yeast host strains designedfor efficient manipulation of DNA in Saccharomyces cerevisiae. Genetics 122(1):19–27.
49. Longtine MS, et al. (1998) Additional modules for versatile and economical PCR-basedgene deletion and modification in Saccharomyces cerevisiae. Yeast 14(10):953–961.
50. Janke C, et al. (2004) A versatile toolbox for PCR-based tagging of yeast genes: Newfluorescent proteins, more markers and promoter substitution cassettes. Yeast 21(11):947–962.
51. Muhlrad D, Hunter R, Parker R (1992) A rapid method for localized mutagenesis ofyeast genes. Yeast 8(2):79–82.
52. Oldenburg KR, Vo KT, Michaelis S, Paddon C (1997) Recombination-mediated PCR-directed plasmid construction in vivo in yeast. Nucleic Acids Res 25(2):451–452.
Suda et al. PNAS | November 19, 2013 | vol. 110 | no. 47 | 18981
CELL
BIOLO
GY











![A Golgi-Released Subpopulation of the Trans-Golgi · A Golgi-Released Subpopulation of the Trans-Golgi Network Mediates Protein Secretion in Arabidopsis1[OPEN] Tomohiro Uemura,a,b,2,3,4](https://img.pdfslide.us/doc/110x75/5eda9f5a09f66a09130ba5a1/a-golgi-released-subpopulation-of-the-trans-golgi-a-golgi-released-subpopulation.jpg)