Embed Size (px)
Citation preview

Int. J. Mol. Sci. 2013, 14, 12607-12619; doi:10.3390/ijms140612607
International Journal of
Molecular Sciences ISSN 1422-0067
www.mdpi.com/journal/ijms
Review
Quorum Sensing and Phytochemicals
Filomena Nazzaro *, Florinda Fratianni and Raffaele Coppola
Institute of Food Science, ISA-CNR, Via Roma 64, Avellino 83100, Italy;
E-Mails: [email protected] (F.F.); [email protected] (R.C.)
* Author to whom correspondence should be addressed; E-Mail: [email protected];
Tel.: +39-825-299102; Fax: +39-825-781585.
Received: 17 May 2013; in revised form: 31 May 2013 / Accepted: 4 June 2013 /
Published: 17 June 2013
Abstract: Most infectious diseases are caused by bacteria, which proliferate within
quorum sensing (QS)-mediated biofilms. Efforts to block QS in bacteria and disrupt
biofilms have enabled the identification of bioactive molecules that are also produced by
plants. This mini review primarily focuses on natural QS inhibitors, which display potential
for treating bacterial infections and also enhance the safety of food supply.
Keywords: quorum sensing; phytochemicals
1. Introduction
During their evolution, bacterial communities have developed several sophisticated ways of
interacting and associating with the environment they inhabit. Microorganisms may respond to a
perceived change by altering their phenotype so that their metabolism and other activities are
successful in the new environment. In the last few decades, it has also become evident that bacteria
coordinate both bacterium-bacterium interactions and associations with higher organisms through
intercellular communication systems that are often based on the expression of new genes, known as
quorum sensing (QS) systems. QS-controlled behaviors take place only when bacteria reach a specific
cell density. Such behaviors are unproductive if undertaken by a singular bacterium but become
effective when the action is simultaneously performed by a group of bacteria. QS can regulate a
number of activities, such as bioluminescence, virulence factor expression, sporulation, biofilm
formation and mating. QS is realized through the bacterial production of chemical signaling molecules,
also known as autoinducers or bacterial pheromones. These signals are produced while the bacterial
OPEN ACCESS

Int. J. Mol. Sci. 2013, 14 12608
population grows until a threshold concentration perceived by the bacteria is reached, resulting in the
activation or repression of specific genes. The accumulation of a stimulatory amount of such molecules
can occur only when a specific, sufficient number of cells, referred to as a quorum, are present [1].
The QS system can be seen as being based on the following crucial elements:
1. The autoinducers;
2. The signal synthase;
3. The signal receptor;
4. The signal response regulator;
5. The regulated genes (which form the so-called QS regulon).
The autoinducers can differ depending on the bacteria. They can be classified into three
subclasses [2]: Autoinducers belonging to the first class are used by Gram-negative bacteria; the
molecules of the second class are used by Gram-positive bacteria; and the third class of biomolecules
can be used both by Gram-positive and Gram-negative bacteria.
Gram-negative bacteria generally produce acylated homoserine lactone (AHL) as autoinducers,
which are synthesized by a LuxI-type enzyme (signal synthase), encoded by the first gene of the lux
operon, as shown in Figure 1. At a low bacterial cell density, the low level of transcription of the lux
operon is insufficient for the activation of LuxR. When the cell density increases and signal levels
reach a specified threshold level, LuxR activation can take place. The LuxR/3-oxo-C6-HSL complex
thereby activates transcription via the lux operon promoter, giving rise to the expression of other genes,
including (in Vibrio fischeri) lux AB genes encoding luciferase and lux CDE, which encodes the
enzymes that produce the substrate for luciferase and, hence, bioluminescence. These molecules
passively diffuse through the bacterial membrane and accumulate both intra- and extracellularly in
proportion to the cell density. Quorum-sensing circuits have been identified in over 25 species of
Gram-negative bacteria. Using the quorum-sensing mechanism, Gram-negative bacteria can efficiently
couple gene expression to fluctuations in cell population density. Among these species, the V. fischeri,
Pseudomonas aeruginosa, Pseudomonas fluorescens, Pseudomonas chlororaphis, Erwinia carotovora
and Agrobacterium tumefaciens systems can be considered the best understood. More recently, the
quorum system mechanism of Chromobacterium violaceum has been studied and used to evaluate the
quorum and antiquorum activity of its biocomponents [3]. In the opportunistic human pathogen
P. aeruginosa, a hierarchical LuxI/LuxR circuit regulates QS. Two pairs of LuxI/LuxR homologues,
LasI/LasR and RhlI/RhlR, exist in P. aeruginosa. Both LasI and RhlI are autoinducer synthases, which
catalyze the formation of the HSL autoinducers N-(3-oxododecanoyl)-homoserine lactone and
N-(butryl)-homoserine lactone, respectively. These two regulatory circuits act in tandem to control the
expression of a number of P. aeruginosa virulence factors. The P. aeruginosa quorum-sensing circuit
functions as follows: At a high cell density, LasR binds its HSL autoinducer, and together, they
combine with promoter elements immediately preceding the genes encoding a number of secreted
virulence factors that are responsible for host tissue destruction during the initiation of the infection
process. These pathogenicity determinants include elastase, encoded by lasB, a protease encoded by
lasA; ExotoxinA, encoded by toxA, and alkaline phosphatase, which is encoded by the aprA gene. The
LasR-autoinducer complex also activates the expression of the second quorum-sensing system of
P. aeruginosa through the activation of a second class of specific target genes, encoding the stationary

Int. J. Mol. Sci. 2013, 14 12609
phase sigma factor rhamnosyl transferase, which is involved in the synthesis of the
biosurfactant/hemolysin rhamnolipid, as well as genes involved in pyocyanin antibiotic synthesis and
the lecA gene, which encodes a cytotoxic lectin [4]. In Cromobacterium violaceum, a Gram-negative
microorganism found in water and soil, the phenotypic response to AHLs involves the production of a
variety of factors, including antibiotics, hydrogen cyanide, proteases, chitinase and particularly
violacein, a water-insoluble purple pigment with antibacterial activity [3,5].
Figure 1. A typical quorum-sensing system in Gram-negative bacteria.
2. Quorum System in Gram Positive Bacteria
Gram-positive bacteria also regulate a variety of processes in response to an increasing cell
population density, but in contrast to Gram-negative bacteria, which use HSL autoinducers,
Gram-positive bacteria employ secreted peptides processed from precursors, that can be used as
autoinducers for QS (Figure 2). Signals are actively exported outside the cell, where they interact with
the external domains of membrane-bound sensor proteins. The transduction of a signal generated by a
phosphorylation cascade culminates in the activation of a DNA-binding protein, that influences the
transcription of specific genes, so that each sensor protein is highly selective for a given peptide signal.
In Gram-positive bacteria, a peptide signal precursor locus is translated into a precursor protein that is
cleaved to produce the processed peptide autoinducer signal. This signal is then usually transported out
of the cell via an ABC transporter. When the extracellular concentration of the peptide signal
accumulates to the minimal stimulatory level, a histidine sensor kinase protein belonging to a
two-component signaling system detects it. The sensor kinase autophosphorylates a conserved
histidine residue (H), and the phosphoryl group is subsequently transferred to a cognate
response-regulator protein. The response regulator is phosphorylated on a conserved aspartate residue
(D), which activates the transcription of a target gene or genes [4]. Similar to Gram-negative bacteria,
Gram-positive bacteria can employ multiple autoinducers and sensors. However, some peptides can
exclusively act from outside the bacterial cell, eliciting a specific set of gene expression changes by
being transported into the cell, where they can activate a different set of behavioral changes. A

Int. J. Mol. Sci. 2013, 14 12610
furanosyl boronated diester molecule termed AI-2 and a non-boronated diester molecule, vA1-2, are
used by Gram-positive and Gram-negative bacteria for intra- and interspecies communication [1].
Many Gram-negative bacteria use AHL autoinducers and also produce AI-2. Likewise, many
Gram-positive bacteria display oligopeptide signaling systems as well as AI-2. Making and responding
to combinations of these and potentially other types of chemical signals could allow bacteria to take a
census of their own population nitrogen levels as well as the population density of other species in the
vicinity. A distinct response to each signal, or a response based on a combinatorial sampling of a
variety of signals, could enable bacteria to continuously modulate their behavior depending on the
species present in a consortium.
Figure 2. A typical quorum-sensing system in Gram-positive bacteria.
D
H
Peptide autoinducerprecursor locus
Target genes
Transport
H and D are the phosphorylated residues histidineand aspartate on the signaling proteins.
Processing
Peptide autoinducers
3. Antiquorum Sensing Activity of Phytochemicals
The role of QS in food ecology has been investigated mainly in the last years [6,7]. In some studies,
various signaling compounds, including AI-1 and AI-2, have been found to be present and even
particularly concentrated in different foods, such as milk, meat and vegetables [8]. Little is known
about the influence of food processing and storage on the production and release of these molecules by
bacteria. As food matrices are generally solid environments (with the exception of liquid foods, of
course), they give rise to microbial cells that become entrapped and localized at high densities at
certain points, where they can show increased rates of growth and form biofilms. The presence and/or
growth of a certain species, as well as its pathogenic activity and biofilm formation, will be affected by
the presence of other microbial species and by all cell-cell interactions occurring in the solid food
matrix, thus influencing the capability of each species to produce QS signals. The results from several
reports suggest that targeting QS could be a new strategy for fighting biofilm infections [9].
Phytochemicals from fruits and vegetables are also capable of inhibiting QS-related processes in human
pathogens [10,11], which is a particularly appealing property. For example, plant food-associated with
QS activity may be of therapeutic interest, as the regular presence in the diet may positively affect the
intestinal microbiota, preventing the concurrent invasion of pathogens. The continuing search for novel

Int. J. Mol. Sci. 2013, 14 12611
antimicrobial and antipathogenic agents has focused on exploiting the fact that plants surviving in an
environment with a high bacterial density may possess protective mechanisms for combating
infections [12]. Based on this argument, researchers are increasingly investigating herbal products in
the quest for new therapeutic and antipathogenic agents that might act as nontoxic inhibitors of QS,
thus controlling infections without encouraging the appearance of resistant bacterial strains [13]. In the
current literature, it is estimated that while 10% of all terrestrial flowering plants have been used by
different communities for treating diseases, only approximately 1% have gained recognition and been
validated [14]. Thus, phytochemicals may represent the richest available reservoir of novel
therapeutics [15]. Although the antimicrobial activities of plant extracts are beyond doubt, in many
instances, their exact mechanism of antimicrobial functionality is not well understood [16,17].
Several types of QS-inhibiting phytochemicals, such as polyphenols, are capable of affecting
biofilm formation in some bacteria [18–21]. However, the antiquorum sensing activity of herbal plants
is still poorly understood, and it is very likely that it will be found that the antimicrobial efficacy is
mediated by QS inhibition. The plant kingdom is a well-known source of medicines and contributes
extensively to the development of pharmaceuticals [22]. Several quorum-quenching molecules have
been identified from plants (Table 1). Many plants have co-evolved and established tightly regulated
symbiotic or syntrophic associations with bacteria. Thus, it is not particularly surprising that higher
organisms are capable of perceiving and responding to these molecules [10,23,24]. Both extracts and
individual molecules from various types of fruits, herbs and spices have shown an ability to inhibit
QS [11,19,25,26]. The best-investigated example of a eukaryotic organism that is capable of producing
metabolites that specifically interfere with bacterial communication is the Australian red alga
Delisea pulchra, which produces metabolites known as halogenated furanones. These compounds show
a wide range of biological activities, including displaying antifouling and antimicrobial properties [27,28].
Certain furanones specifically interfere with AHL-regulated processes [29] by accelerating the
degradation of the AHL receptor protein [30]. Teplitski et al. [23] demonstrated that several plants
secrete substances that mimic bacterial AHLs and subsequently affect quorum-sensing-regulated
behaviors in the bacteria associated with these plants. Thus, the detection of anti-pathogenic
phytochemicals that inhibit the QS regulation of bacterial colonization and virulence factor production
may provide very promising alternative anti-infective agents [31,32]. Plant extracts can act as QSIs
due to the similarity of their chemical structure to those of QS signals (AHL) and/or their ability to
degrade signal receptors (LuxR/LasR) [11,24]. GABA (γ-aminobutyric acid), which is produced by
some plants, acts to promote the degradation of the OHC8HSL AHL signal by lactonase (AttM)
in A. tumefaciens, thus limiting the QS-dependent infection process [33,34]. Emblica officinalis,
through the presence of pyrogallol and its analogues, exhibits antagonism against AI-2 [35].
Medicago truncatula modulates AhyR, CviR and LuxR reporter activities in different organisms [22] as
well as QS in general in P. aeruginosa and S. meliloti [36]. Curcuma longa, through the production of
curcumin, inhibits the expression of virulence genes in P. aeruginosa PA01 [37]. Extracts from some
varieties of apple (e.g., Annurca) and apple derivatives (e.g., cider) show demonstrated QSI activity,
most likely due to the presence of different polyphenols, such as hydroxycinnamic acids, rutin and
epicatechin, which act as AQS agents in synergistic manner against C. violaceum [38,39]. Antiquorum
sensing activities have also been observed for extracts of L. nobilis, S. oleraceus, R. officinalis,
T. capensis, J. sambac, P. alba and P. nigra, which are capable of decreasing violacein production [25].

Int. J. Mol. Sci. 2013, 14 12612
Cinnamaldehyde and its derivatives affect a wide range of QS-regulated activities, such as biofilm
formation in P. aeruginosa and AI-2-mediated QS in different Vibrio spp. [40,41]. Grapefruit, due to
the presence of furo-coumarins, has been shown to inhibit the AI-1 and AI-2 activities of V. harveyi as
well as biofilm formation by pathogens such as E. coli, S. typhimurium and P. aeruginosa [42].
Extracts of sour orange seeds containing limonoids, such as isolimonic acid, ichangin and deacetyl
nomilinic acid 17 β-D-glucopyranoside, can cause >90% inhibition of AI-2 activity in V. harveyi at a
concentration of 100 μg/mL and show activity against HAI- and AI-2-mediated bioluminescence [43].
Flavanones, flavonoids abundant in Citrus, have been shown to interfere with quorum sensing (QS)
and affect related physiological processes [44]. Flavonoids, such as naringenin, kaempferol, quercetin
and apigenin, inhibit HAI-1- or AI-2-mediated bioluminescence in V. harveyi BB886 and MM32.
Flavanones (i.e., naringenin eriodictyol and taxifolin) can be capable to significantly reduce the
production of pyocyanin and elastase in P. aeruginosa without affecting bacterial growth. Naringenin
and taxifolin also reduce the expression of several QS-controlled genes (i.e., lasI, lasR, rhlI, rhlR,
lasA, lasB, phzA1 and rhlA) in P. aeruginosa PAO1. Naringenin also dramatically reduce the
production of the acylhomoserine lactones N-(3-oxododecanoyl)-l-homoserine lactone
(3-oxo-C12-HSL) and N-butanoyl-l-homoserine lactone (C4-HSL), driven by the lasI and rhlI gene
products, respectively [45]. Quercetin, sinensetin, apigenin and naringenin display anti-biofilm
formation activity against V. harveyi BB120 and E. coli O157:H7 [44,46]. Flavan-3-ol catechin can
reduce the production of QS-mediated virulence factors, such as pyocyanin and elastase, and biofilm
formation by P. aeruginosa PAO1 [47,48]. AHL-degrading abilities have been reported for a large
number of legumes, including alfalfa, clover, lotus, peas and yam beans [49–51]. Biofilm formation by
E. coli can even be disrupted by grapefruit juice and by rosmarinic acid produced by the roots of
Ocimum basilicum (sweet basil) [11]. Phenolic plant secondary metabolites such as salicylic acid
stimulate AHL-lactonase enzyme expression [52]. Ursolic acid at 10 μg/mL is capable of decreasing
biofilm formation by 79% in E. coli and by 57%–95% in V. harveyi and P. aeruginosa PAO1 [53].
Aqueous extracts of edible plants and fruits such as Ananas comosus, Musa paradiciaca, Manilkara
zapota and Ocimum sanctum have been found to show QSI activity against violacein production by
C. violaceum and against pyocyanin pigment, staphylolytic protease and elastase production in
P. aeruginosa PAO1 as well as its biofilm formation ability [54]. Broccoli extracts and its constituents
can inhibit expression of QS-associated genes, thereby down-regulating the virulence attributes of
E. coli O157:H7 both in vitro and in vivo, suggesting that this vegetal, like other Brassicaeae, has the
potential to be developed as an anti-infective agent [55]. Other extracts, such as ethanol and ethyl
acetate extracts from Hypericum connatum, exhibit QSI activity against C. violaceum, limiting its
production of violacein [56]. Polyphenol compounds with a gallic acid moiety, such as
epigallocatechin gallate, ellagic acid and tannic acid, which are commonly produced by many plants,
are capable of specifically interfering with AHL-mediated signaling by blocking AHL-mediated
communication between bacteria [19,20]. For example, pomegranates and berries are rich in
ellagitanins such as punicalagin and ellagic acid, showing concentrations higher than
300 mg/100 gr [57]. In the gut, ellagitanins are hydrolyzed to ellagic acid by the microbiota and
subsequently metabolized to form urolithin-A and urolithin-B; these metabolites can then accumulate
in the human intestine, where they have important functions. Urolithin-A, for example is capable
of modulating the growth of bacteria in rat intestines before and after chemically induced
Int. J. Mol. Sci. 2013, 14 12613
inflammation [58]. Urolithin A and urolithin B are also able to inhibit QS-associated processes
(by up to 40%) and decrease the levels of AHLs produced by the entheropathogen Y. entherocolitica
(by up to 45%) [59]. 4',5'-O-dicaffeoylquinic acid (4',5'-ODCQA) can act as a pump inhibitor with a
potential of targeting efflux systems in a wide panel of Gram-positive human pathogenic bacteria [60].
Chlorogenic and vanillic acids and rutin, which can be found in berries, resveratrol (grapes), kinurenic
acid (honey), daidzein (soy), dimethyl-esculetin (artemisia) and pomegranate extract can all be used as
positive controls for QS inhibition [61]. Flavanols and proanthocyanidins form a complex with the
spores and hyphae of pathogenic fungi of fruit crops [62], and phenolic polymer deposition is related
to a decrease in bacterial multiplication rates. QS-inhibitory activity has been demonstrated for several
unifloral honeys, which were found to be capable of inhibiting the production of AHLs: Chestnut
honey showed the highest inhibitory activity, whereas orange and rosemary honeys were less effective.
Truchado et al. [63] hypothesized that one of the factors affecting this inhibitory activity may be its
floral origin, independent of the geographic location.
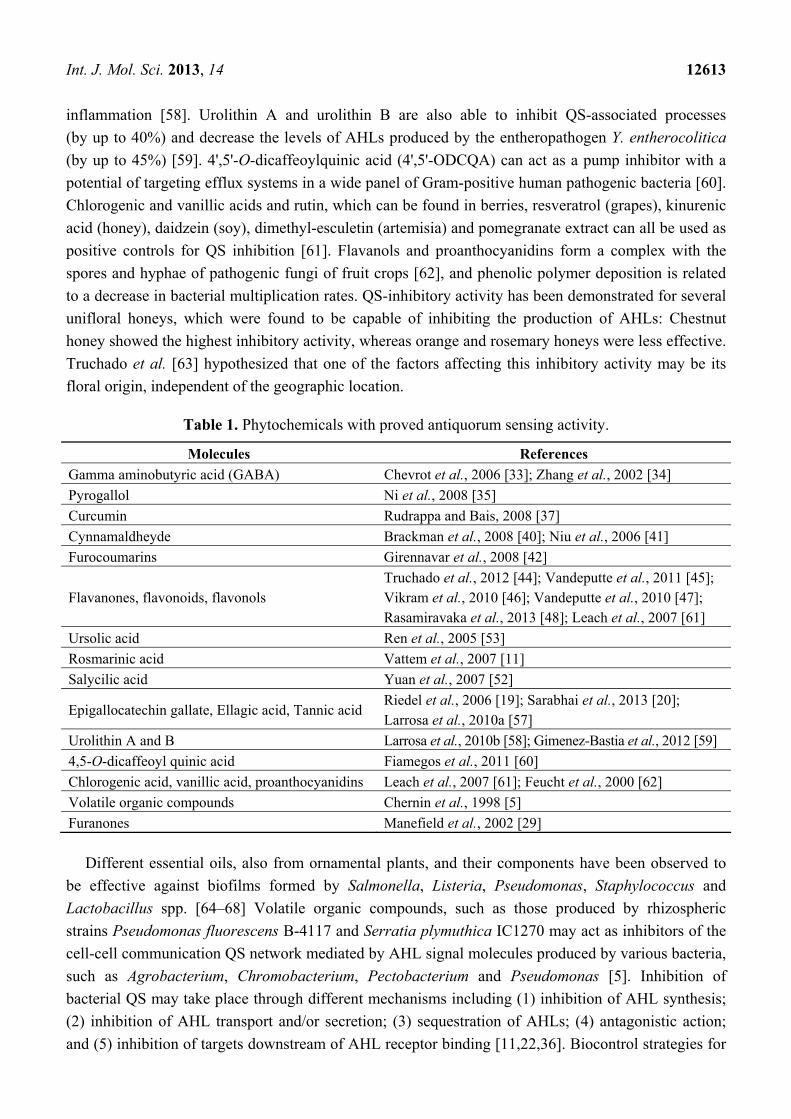
Table 1. Phytochemicals with proved antiquorum sensing activity.
Molecules References
Gamma aminobutyric acid (GABA) Chevrot et al., 2006 [33]; Zhang et al., 2002 [34]
Pyrogallol Ni et al., 2008 [35]
Curcumin Rudrappa and Bais, 2008 [37]
Cynnamaldheyde Brackman et al., 2008 [40]; Niu et al., 2006 [41]
Furocoumarins Girennavar et al., 2008 [42]
Flavanones, flavonoids, flavonols Truchado et al., 2012 [44]; Vandeputte et al., 2011 [45]; Vikram et al., 2010 [46]; Vandeputte et al., 2010 [47]; Rasamiravaka et al., 2013 [48]; Leach et al., 2007 [61]
Ursolic acid Ren et al., 2005 [53]
Rosmarinic acid Vattem et al., 2007 [11]
Salycilic acid Yuan et al., 2007 [52]
Epigallocatechin gallate, Ellagic acid, Tannic acid Riedel et al., 2006 [19]; Sarabhai et al., 2013 [20]; Larrosa et al., 2010a [57]
Urolithin A and B Larrosa et al., 2010b [58]; Gimenez-Bastia et al., 2012 [59]
4,5-O-dicaffeoyl quinic acid Fiamegos et al., 2011 [60]
Chlorogenic acid, vanillic acid, proanthocyanidins Leach et al., 2007 [61]; Feucht et al., 2000 [62]
Volatile organic compounds Chernin et al., 1998 [5]
Furanones Manefield et al., 2002 [29]
Different essential oils, also from ornamental plants, and their components have been observed to
be effective against biofilms formed by Salmonella, Listeria, Pseudomonas, Staphylococcus and
Lactobacillus spp. [64–68] Volatile organic compounds, such as those produced by rhizospheric
strains Pseudomonas fluorescens B-4117 and Serratia plymuthica IC1270 may act as inhibitors of the
cell-cell communication QS network mediated by AHL signal molecules produced by various bacteria,
such as Agrobacterium, Chromobacterium, Pectobacterium and Pseudomonas [5]. Inhibition of
bacterial QS may take place through different mechanisms including (1) inhibition of AHL synthesis;
(2) inhibition of AHL transport and/or secretion; (3) sequestration of AHLs; (4) antagonistic action;
and (5) inhibition of targets downstream of AHL receptor binding [11,22,36]. Biocontrol strategies for

Int. J. Mol. Sci. 2013, 14 12614
combating bacterial QS and biofilm formation through the use of natural phytochemicals can enhance
the safety and security of foods and improve human health by reducing the capability of pathogens to
invade the intestine, providing an opportunity for (1) downregulation of microbial spoilage activity and
(2) altering microbial activity and survival. The first strategy for combating microbial spoilage activity
presents the interesting advantage of preventing other undesirable microorganisms from colonizing a
particular niche, while down-regulating the expression of enzymes such as proteases and lipases,
resulting in a reduction in food damage. The second approach may be more desirable in the case of
foodborne pathogens. The use of a bacterium’s own QS system against itself may minimize the
possibility that the bacterium will adapt and become resistant to the applied QS inhibitors. Again, use
of vegetal extracts and/or pure compounds can contribute to the limitation of food spoilage and the
formation of bacterial biofilms. Extracts of garlic have been shown to block QS by P. aeruginosa,
limiting the production of biofilms and thereby supporting the clearance of the bacteria [69]. Similarly,
vanilla extracts are capable of interfering with QS in C. violaceum, suggesting that the consumption of
vanilla-containing foods may be beneficial [70]. The presence of ascorbic acid, an AI-2 analog present
in several fruits, may result in reduction of toxin production by Clostridium perfringens as well as
decreased spore production when added to ground meat extract [71].
4. Conclusions
Interventions targeting bacterial QS in food are largely unexplored at present. Consumer
expectations of obtaining pathogen-free food with an acceptable shelf life without the use of chemical
additives represent a great challenge for the food industry based on currently available technologies.
Biocontrol approaches using vegetal extracts or pure compounds obtained from plants offer some
specific advantages compared to more conventional treatments. Indeed, thus-treated food is perceived
as more “natural” and “green.” Although no one bio-control approach can currently address this
problem in its entirety, such approaches, together, generate a toolbox of options that may act in
synergistic and complementary ways, through various mechanisms of attack, that can be applied
individually or in combination. Indeed, it is becoming increasingly evident that the use of
phytochemical-based biocontrol strategies in food shows great potential for more realistic outcomes,
although much work is still necessary to move these treatments from the laboratory bench to market.
Additionally, better understanding the potential of phytochemicals to inhibit QS activity is of great
relevance to the field of research aimed at the identification and development of novel anti-QS
compounds capable of preventing bacterial infections in humans. Such ‘‘antipathogenic” compounds,
in contrast to antibacterial compounds, would not kill bacteria or completely stop their growth and
would therefore most likely prevent the development of resistant strains [72]. Finally, in recent years,
plant bioengineering could be helpful, which in particular, could influence their associated bacteria, as
demonstrated for example by QS strategies suppressing the influence of pathogens of the genus
Pectobacterium [73].
Conflict of Interest
The authors declare no conflict of interest.

Int. J. Mol. Sci. 2013, 14 12615
References
1. Bassler, B.L. Small talk: Cell-to-cell communication in bacteria. Cell 2002, 109, 421–424.
2. Reading, N.C.; Sperandio, V. Quorum sensing: The many languages of bacteria. FEMS
Microbiol. Lett. 2006, 254, 1–11.
3. McClean, K.H.; Winson, M.K.; Fish, L.; Taylor, A.; Chhabra, S.R.; Camara, M.; Daykin, M.;
Lamb, J.H.; Swift, S.; Bycroft, B.W.; et al. Quorum sensing and Chromobacterium violaceum:
Exploitation of violacein production and inhibition for the detection of N-acylhomoserine
lactones. Microbiology 1997, 143, 3703–3711.
4. Miller, M.B.; Bassler, B.L. Quorum sensing in bacteria. Annu. Rev. Microbiol. 2001, 55, 165–199.
5. Chernin, L.S.; Winson, M.K.; Thompson, J.M.; Haran, S.; Bycroft, B.W.; Chet, I.; Williams, P.;
Gordon, S.; Stewart, A.B. Chitinolytic activity in Chromobacterium violaceum: Substrate analysis
and regulation by quorum sensing. J. Bacteriol. 1998, 180, 4435–4441.
6. Smid, E.J.; Lacroix, C. Microbe-microbe interactions in mixed culture food fermentations.
Curr. Opin. Biotechnol. 2013, 24, 148–154.
7. Skandamis, P.N.; Nychas, G.-J. Quorum sensing in the context of food microbiology.
Appl. Environ. Microbiol. 2012, 78, 5473–5482.
8. Ammor, M.S.; Michaelidis, C.; Nychas, G.J.E. Insights into the role of quorum sensing in food
spoilage. J. Food Protect. 2008, 171, 1510–1525.
9. Musk, D.J., Jr.; Hergenrother, P.J. Chemical countermeasures for the control of bacterial biofilms:
Effective compounds and promising targets. Curr. Med. Chem. 2006, 13, 2163–2177.
10. Adonizio, A.L.; Downum, K.; Bennett, B.C.; Mathee, K. Antiquorum sensing activity of
medicinal plants in southern Florida. J. Ethnopharmacol. 2006, 105, 427–435.
11. Vattem, D.A.; Mihalik, K.; Crixell, S.H.; McClean, R.J.C. Dietary phytochemicals as quorum
sensing inhibitors. Fitoterapia 2007, 78, 302–310.
12. Cos, P.; Vlietink, A.J.; Berghe, D.V.; Maes, L. Anti-infective potential of natural products:
How to develop a stronger in vitro “proof of concept”. J. Ethnopharmacol. 2006, 106, 290–302.
13. Hentzer, M.; Givskov, M. Pharmacological inhibition of quorum sensing for the treatment of
chronic bacterial infections. J. Clin. Invest. 2003, 112, 1300–1307.
14. Lewis, K.; Ausubel, F.M. Prospects of plant derived antibacterials. Nat. Biotechnol. 2006, 24,
1504–1507.
15. Kumar, V.P; Chauhan, N.S.; Rajani, H.P.M. Search for antibacterial and antifungal agents from
selected Indian medicinal plants. J. Ethnopharmacol. 2006, 107, 182–188.
16. Mowrey, D.B. The Scientific Validation of Herbal Medicine; McGraw-Hill: New York, NY,
USA, 1990; pp. 789–799.
17. Rios, J.L.; Recio, M.C. Medicinal plants and antimicrobial activity. J. Ethnopharmacol. 2005,
100, 80–84.
18. Huber, B.; Eberl, L.; Feucht, W.; Polster, J. Influence of polyphenols on bacterial biofilm
formation and quorum-sensing. Z. Naturforsch. 2003, 58, 879–884.
19. Riedel, K.; Kothe, M.; Kramer, B.; Saeb, W.; Gotschlich, A.; Ammendola, A.;
Eberl, L. Computer-aided design of agents that inhibit the cep quorum-sensing system of
Burkholderia cenocepacia. Antimicrob. Agents Chemother. 2006, 50, 318–323.

Int. J. Mol. Sci. 2013, 14 12616
20. Sarabhai, S.; Sharma, P.; Capalash, N. Ellagic acid derivatives from Terminalia chebula Retz.
downregulate the expression of quorum sensing genes to attenuate Pseudomonas aeruginosa
PAO1 virulence. PLoS One 2013, 8, e53441.
21. Cragg, G.M.; Newman, D.J.; Snader, K.M. Natural products in drug discovery and development.
J. Nat. Prod. 1997, 60, 52–60.
22. Gao, M.; Teplitski, M.; Robinson, J.B.; Bauer, W.D. Production of substances by
Medicago truncatula that affect bacterial quorum sensing. Mol. Plant Microbe Interact. 2003,
16, 827–834.
23. Teplitski, M.; Robinson, J.B.; Bauer, W.D. Plants secrete substances that mimic bacterial
N-acyl homoserine lactone signal activities and affect population density-dependent behaviors in
associated bacteria. Mol. Plant Microbe Interact. 2000, 13, 637–648.
24. Al-Hussaini, R.; Mahasneh, A.M. Microbial growth and quorum sensing antagonist activities of
herbal plants extracts. Molecules 2009, 14, 3425–3435.
25. Brackman, G.; Hillaert, U.; van Calenbergh, S.; Nelis, H.J.; Coenye, T. Use of quorum sensing
inhibitors to interfere with biofilm formation and development in Burkholderia multivorans and
Burkholderia cenocepacia. Res. Microbiol. 2009, 160, 144–151.
26. Vikram, A.; Jesudhasan, P.R.; Jayaprakasha, G.K.; Pillai, B.S.; Patil, B.S. Grapefruit
bioactive limonoids modulate E. coli O157:H7 TTSS and biofilm. Int. J. Food Microbiol. 2010,
140, 109–116.
27. De Nys, R.; Wright A.D.; König, G.M.; Sticher, O. New halogenated furanones from the marine
alga Delisea pulchra. Tetrahedron 1993, 49, 11213–11220.
28. Givskov, M.; de Nys, R.; Manefield, M.; Gram, L.; Maximilien, R.; Eberl. L.; Molin, S.;
Steinberg, P.D.; Kjelleberg, S. Eukaryotic interference with homoserine lactone-mediated
prokaryotic signaling. J. Bacteriol. 1996, 178, 6618–6622.
29. Manefield, M.; Rasmussen, T.B.; Henzter, M.; Andersen, J.B.; Steinberg, P.; Kjelleberg, S.;
Givskov, M. Halogenated furanones inhibit quorum sensing through accelerated LuxR turnover.
Microbiology 2002, 148, 1119–1127.
30. Coates, A.; Hu, Y.; Bax, R.; Page, C. The future challenges facing the development of new
antimicrobial drugs. Nat. Rev. Drug Discov. 2002, 1, 895–910.
31. Martin, C.A.; Hoven, A.D.; Cook, A.M. Therapeutic frontiers: Preventing and treating
infectious diseases by inhibiting bacterial quorum sensing. Eur. J. Clin. Microbiol. 2008, 27,
635–642.
32. Kalia, C.V. Quorum sensing inhibitors: An overview. Biotechnol. Adv. 2013, 31, 224–245.
33. Chevrot, R.; Rosen, R.; Haudecoeur, E.; Cirou, A.; Shelp, B.J.; Ron, E.; Faure, D. GABA controls
the level of quorum-sensing signal in Agrobacterium tumefaciens. Proc. Natl. Acad. Sci. USA
2006, 103, 7460–7464.
34. Zhang, H.B.; Wang, L.H.; Zhang, L.H. Genetic control of quorum-sensing signal turnover in
Agrobacterium tumefaciens. Proc. Natl. Acad. Sci. USA 2002, 99, 4638–4643.
35. Ni, N.; Choudhary, G.; Li, M.; Wang, B. Pyrogallol and its analogs can antagonize bacterial
quorum sensing in Vibrio harveyi. Bioorg. Med. Chem. Lett. 2008, 18, 1567–1572.

Int. J. Mol. Sci. 2013, 14 12617
36. Mathesius, U.; Mulders, S.; Gao, M.; Teplitski, M.; Caetano-Anolles, G.; Rolfe, B.G.; Bauer, W.D.
Extensive and specific responses of a eukaryote to bacterial quorum-sensing signals. Proc. Natl.
Acad. Sci. USA 2003, 100, 1444–1449.
37. Rudrappa, T.; Bais, H.P. Curcumin, a known phenolic from Curcuma longa, attenuates the
virulence of Pseudomonas aeruginosa PAO1 in whole plant and animal pathogenicity models.
J. Agric. Food Chem. 2008, 56, 1955–1962.
38. Fratianni, F.; Coppola, R.; Nazzaro, F. Phenolic composition and antimicrobial and antiquorum
sensing activity of an ethanolic extract of peels from the apple cultivar Annurca. J. Med. Food
2011, 14, 957–963.
39. Fratianni, F.; de Giulio, A.; Sada, A.; Nazzaro, F. Biochemical characteristics and biological
properties of annurca apple cider. J. Med. Food 2012, 15, 18–23.
40. Brackman, G.; Defoirdt, T.; Miyamoto, C.; Bossier, P.; van Calenbergh, S.; Nelis, H.; Coenye, T.
Cinnamaldehyde and cinnamaldehyde derivatives reduce virulence in Vibrio spp. by decreasing
the DNA-binding activity of the quorum sensing response regulator LuxR. BMC Microbiol. 2008,
8, 149–162.
41. Niu, C.; Afre, S.; Gilbert, E.S. Sub-inhibitory concentrations of cinnamaldehyde interfere with
quorum sensing. Lett. Appl. Microbiol. 2006, 43, 489–494.
42. Girennavar, B.; Cepeda, M.L.; Soni, K.A.; Vikram, A.; Jesudhasan, P.; Jayaprakasha, G.K.;
Pillai, S.D.; Patil, B.S. Grapefruit juice and its furocoumarins inhibits autoinducer signaling and
biofilm formation in bacteria. Int. J. Food Microbiol. 2008, 125, 204–208.
43. Vikram, A.; Jesudhasan, P.R.; Jayaprakasha, G.K.; Pillai, S.D.; Patil, B.S. Citrus limonoids
interfere with Vibrio harveyi cell-cell signaling and biofilm formation by modulating the response
regulator LuxO. Microbiology 2011, 157, 99–110.
44. Truchado, P.; Giménez-Bastida, J.-A.; Larrosa, M.; Castro-Ibáñez, I.; Espı́n, J.C.;
Tomás-Barberán, F.A.; Garcı́a-Conesa, M.T.; Allende, A. Inhibition of quorum sensing (QS) in
Yersinia enterocolitica by an orange extract rich in glycosylated flavanones. J. Agric. Food Chem.
2012, 60, 8885–8894.
45. Vandeputte, O.M.; Kiendrebeogo, M.; Rasamiravaka, T.; Stévigny, C.; Duez, P.; Rajaonson, S.;
Diallo, B.; Mol, A.; Baucher, M.; El Jazir, M. The flavanone naringenin reduces the production of
quorum sensing-controlled virulence factors in Pseudomonas aeruginosa PAO1. Microbiology
2011, 157, 2120–2132.
46. Vikram, A.; Jayaprakasha, G.K.; Jesudhasan, P.R.; Pillai, S.D.; Patil, B.S. Suppression of
bacterial cell-cell signalling, biofilm formation and type III secretion system by citrus flavonoids.
J. Appl. Microbiol. 2010, 109, 515–527.
47. Vandeputte, O.M.; Kiendrebeogo, M.; Rajaonson, S.; Diallo, B.; Mol, A.; Jaziri, M.E.;
Baucher, M. Identification of catechin as one of the flavonoids from Combretum albiflorum bark
extract that reduces the production of quorum-sensing-controlled virulence factors in
Pseudomonas aeruginosa PAO1. Appl. Environ. Microbiol. 2010, 71, 243–253.
48. Rasamiravaka, T.; Jedrzejowski, A.; Kiendrebeogo, M.; Rajaonson, S.; Randriamampionona, D.;
Rabemanantsoa, S.; Andriantsimahavandy, A.; Rasamindrakotroka, A.; El Jaziri, M.;
Vandeputte, O.M. Endemic Malagasy Dalbergia species inhibit quorum sensing in
Pseudomonas aeruginosa PAO1. Microbiology 2013, doi:10.1099/mic.0.064378-0.

Int. J. Mol. Sci. 2013, 14 12618
49. Delalande, L.; Faure, D.; Raffoux, A.; Uroz, S.; D’Angelo-Picard, C.; Elasri, M.; Carlier, A.;
Berruyer, R.; Petit, A.; Williams, P.; Dessaux, Y. N-hexanoyl-L-homoserine lactone, a mediator of
bacterial quorum-sensing regulation, exhibits plant-dependent stability and may be inactivated by
germinating Lotus corniculatus seedlings. FEMS Microbiol. Ecol. 2005, 52, 13–20.
50. Fatima, Q.; Zahin, M.; Khan, M.S.A.; Ahmad, I. Modulation of quorum sensing controlled
behavior of bacteria by growing seedling, seed and seedling extracts of leguminous plants. Indian
J. Microbiol. 2010, 50, 238–242.
51. Gotz, C.; Fekete, A.; Gebefuegi, I.; Forczek, S.T.; Fuksova, K.; Li, X.; Englmann, M.;
Gryndler, M.; Hartmann, M.; Matucha, M.; et al. Uptake, degradation and chiral discrimination of
N-acyl-D/L-homoserine lactones by barley (Hordeum vulgare) and yam bean (Pachyrhizus erosus)
plants. Anal. Bioanal. Chem. 2007, 389, 1447–1457.
52. Yuan, Z.C.; Edlind, M.P.; Liu, P.; Saenkham, P.; Banta, L.M.; Wise, A.A.; Ronzone, E.;
Binns, A.N.; Kerr, K.; Nester, E.W. The plant signal salicylic acid shuts down expression of the
vir regulon and activates quormone-quenching genes in Agrobacterium. Proc. Natl. Acad. Sci. USA
2007, 104, 11790–11795.
53. Ren, D.; Zuo, R.; González-Barrios, A.F.; Bedzyk, L.A.; Eldridge, G.R.; Pasmore, M.E.;
Wood, T.K. Differential gene expression for investigation of Escherichia coli biofilm inhibition
by plant extract ursolic acid. Appl. Environ. Microbiol. 2005, 71, 4022–4034.
54. Musthafa, K.S.; Ravi, A.V.; Annapoorani, A.; Packiavathy, I.S.V.; Pandian, S.K. Evaluation of
anti-quorum-sensing activity of edible plants and fruits through inhibition of the
N-acyl-homoserine lactone system in Chromobacterium violaceum and Pseudomonas aeruginosa.
Chemotherapy 2010, 56, 333–339.
55. Lee, K.-M.; Lim, J.; Nam, S.; Young Yoon, M.; Kwon, Y.-K.; Jung, B.Y.; Park, Y.J.; Park, S.;
Yoon, S.S. Inhibitory effects of broccoli extract on Escherichia coli O157:H7 quorum sensing and
in vivo virulence. FEMS Microbiol. Lett. 2011, 321, 67–74.
56. Fratianni, F.; Nazzaro, F.; Marandino, A.; Fusco M.D.R.; Coppola, R.; de Feo, V.; de Martino, L.
Biochemical composition, antimicrobial activities, and anti-quorum-sensing activities of ethanol and
ethyl acetate extracts from Hypericum connatum Lam. (Guttiferae). J. Med. Food 2013, 16, 454–459.
57. Larrosa, M.; García-Conesa, M.T.; Espín, J.C.; Tomás-Barberán, F.A. Ellagitannins, ellagic acid
and vascular health. Mol. Asp. Med. 2010, 31, 513–539.
58. Larrosa, M.; González-Sarrías, A.; Yánez-Gascón, M.J.; Selma, M.V.; Azorín-Ortuño, M.; Totia, S.;
Tomás-Barberán, F.A.; Dolara, P.; Espín, J.C. Anti-inflammatory properties of a pomegranate
extract and its metabolite urolithin-A in a colitis rat model and the effect of colon inflammation on
phenolics metabolism. J. Nutr. Biochem. 2010, 21, 717–725.
59. Giménez-Bastida, J.A.; Truchado, P., Larrosa, M.; Espín, J.C.; Tomás-Barberán, F.A.; Allende, A.;
García-Conesa, M.T. Urolithins, ellagitannin metabolites produced by colon microbiota, inhibit
Quorum Sensing in Yersinia enterocolitica: Phenotypic response and associated molecular
changes. Food Chem. 2012, 132, 1465–1474.
60. Fiamegos, Y.C.; Panagiotis, L.; Kastritis, X.; Vassiliki, E.; Han, H.; Bonvin, A.M.J.J.; Vervoort, J.;
Lewis, K.; Hamblin, M.R.; Tegos, G.P. Antimicrobial and efflux pump inhibitory activity of
caffeoylquinic acids from Artemisia absinthium against Gram-positive pathogenic bacteria.
PLoS One 2011, 6, e18127.

Int. J. Mol. Sci. 2013, 14 12619
61. Leach, J.E.; Lloyd, L.A.; McGee, J.D.; Hilaire, E.; Wang, X.; Guikema, J.A. Trafficking of Plant
Defense Response Compounds. In Delivery and Perception of Pathogen Signals in Plants;
Keen, N.T., Mayama, S., Leach, J.E., Tsuyumu, S., Eds.; The American Phytopathological
Society (APS) Press: St. Paul, MN, USA, 2007; pp. 1–268.
62. Feucht, W.; Schwalb, P.; Zinkernagel, V. Complexation of fungal structures with monomeric and
prooligomeric flavanols. J. Plant Dis. Prot. 2000, 107, 106–110.
63. Truchado, P.; López-Gálvez, F.; Gil, M.I.; Tomás-Barberán, F.A.; Allende, A. Quorum sensing
inhibitory and antimicrobial activities of honeys and the relationship with individual phenolics.
Food Chem. 2009, 115, 1337–1344.
64. Al-Shuneigat, J.; Cox, S.D.; Markham, J.L. Effects of a topical essential oil-containing formulation
on bio-film-forming coagulase-negative staphylococci. Lett. Appl. Microbiol. 2005, 41, 52–55.
65. Chorianopoulos, N.G.; Giaouris, F.D.; Skendamis, P.N.; Haroutounian, S.A.; Nychas, G.-J.
Disinfectant test against monoculture and mixed-culture biofilms composed of technological,
spoilage and pathogenic bacteria: Bactericidal effect of essential oil and hydrosol of
Satureja thymbra and comparison with standard acid-base sanitizers. J. Appl. Microbiol. 2008,
104, 1586–96.
66. Khan, M.S.; Zahin, M.; Hasan, S.; Husain, F.M.; Ahmad, I. Inhibition of quorum sensing
regulated bacterial functions by plant essential oils with special reference to clove oil. Lett. Appl.
Microbiol. 2009, 49, 354–360.
67. Zaki, A.A.; Shaaban, M.I.; Hashish, N.E.; Amer, M.A.; Lahloub, M.-F. Assessment of anti-quorum
sensing activity for some ornamental and medicinal plants native to Egypt. Sci. Pharm. 2013,
81, 251–258.
68. Szabó, M.A.; Varga, G.Z.; Hohmann, J.; Schelz, Z.; Szegedi, E.; Amaral, L.; Molnár, J. Inhibition
of quorum-sensing signals by essential oils. Phytother. Res. 2010, 24, 782–786.
69. Bjarnsholt, T.; Jensen, P.O.; Rasmussen, T.B.; Christophersen, L.; Calum, H.; Hentzer, M.;
Hougen, H.P.; Rygaard, J.; Moser, C.; Eberl, L.; et al. Garlic blocks quorum sensing and
promotes rapid clearing of pulmonary Pseudomonas aeruginosa infections. Microbiology 2005,
151, 3873–3880.
70. Choo, J.H.; Rukayadi, Y.; Hwang, J.K. Inhibition of bacterial quorum sensing by vanilla extract.
Lett. Appl. Microbiol. 2006, 42, 637–641.
71. Novak, J.S.; Fratamico, P.M. Evaluation of ascorbic acid as a quorum sensing analogue to control
growth, sporulation, and enterotoxin production in Clostridium perfringens. J. Food Sci. 2004, 69,
72–78.
72. Otto, M. Quorum-sensing control in Staphylococci—A target for antimicrobial drug therapy?
FEMS Microbiol. Lett. 2004, 241, 135–141.
73. Barriuso, J.; Solano, B.; Fray, R.G.; Cámara, M.; Hartmann, A.; Mañero, J.G. Transgenic tomato
plants alter quorum sensing in plant growth-promoting rhizobacteria. Plant Biotechnol. J. 2008,
6, 442–452.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article
distributed under the terms and conditions of the Creative Commons Attribution license
(http://creativecommons.org/licenses/by/3.0/).