Embed Size (px)
Citation preview

Biochem. J. (1981) 200. 573-581 573Printed in Great Britain
Quantitative measurements of the proton-motive force and its relation tosteady state lactose accumulation in Escherichia coli
Sohail AHMED and Ian R. BOOTHDepartment ofMicrobiology, University ofA berdeen, Marischal College, A berdeen AB9 IAS, U.K.
(Received 7 July 1981/Accepted 28 August 1981)
The magnitude of AVg (membrane potential), ApH (pH gradient), lactose accumulationand cytoplasmic volume have been determined over a range of experimental conditions.A study of two probes of ApH, benzoate and dimethyloxazolidene-2,4-dione (DMO),and four probes of AV/, Rb+, K+, tetraphenylphosphonium (TPP+) and 3,3'-dipropylthiodicarbocyanine iodide, has been carried out. Benzoate and DMO are shownto be equivalent at pH values above the pK of DMO, but the latter may be less accuratebelow this pH. The cations TPP+ and Rb+ were found, by a number of criteria, to beequivalent, and TPP+ may be used in cells not pretreated with EDTA. These studies arean essential prerequisite to the use of TPP+ as a quantitative probe in untreated cells.
The chemiosmotic model of energy couplingplaces the proton in a central position in bacterialenergetics. In a respiring bacterium, such asEscherichia coli, electron flow to oxygen is asso-ciated with proton expulsion generating an electro-chemical gradient of protons across the membrane,the proton-motive force. Since the advent of themodel (Mitchell, 1963) the general principles ofchemiosmotic coupling have been verified for a widevariety of membrane-bound energy transducingsystems (Hamilton, 1975; Harold, 1976). Sub-sequently, the attention of bioenergeticists has beenfocused on the adequacy of the bulk phase proton-motive force, Ap, to account for the work doneeither in ATP synthesis or in solute transport (Kell,1979; Booth & Hamilton, 1980a; Mitchell, 1981).At the present time there is considerable contro-versy surrounding the finer detail of the chemi-osmotic model (Ramos & Kaback, 1977; Kell,1979; Kell et al., 198 1).Any systematic study of the chemiosmotic model
of transport requires an accurate assessment of themagnitude of the components of Ap and of trans-port itself. Ap consists of a charge gradient, themembrane potential (AV), and a pH gradient, ApH:
Ap=AV-ZApH (1)Abbreviations used: pH1, internal pH; pHo, external
pH; Ay/, membrane potential; ApH, pH gradient; Ap,proton-motive force; Apiac, the lactose accumulation ratioexpressed in mV; TPP+, tetraphenylphosphonium cation:DMO, dimethyloxazolidene-2,4-dione: Mes, 4-morpholine-ethanesulphonic acid; cyanine diS-C3-(5),3,3'-dipropylthiodicarbocyanine iodide.
Vol. 200
where Z is equal to 59 at 250C and convertslogarithmic concentration gradients to mV. Thisdescription of the bulk phase Ap as the driving forcefor solute uptake has been criticized (Kell, 1979) butnone the less remains the only working model(Mitchell, 198 1; Kell et al., 198 1).The magnitude of AV is usually determined by the
transmembrane distribution of a lipophilic cation.The accuracy of the method relies on the passivepermeation of the cation in response to the b/tilkphase Ay with no contribution to its accumulationfrom either binding or transport systems. In reality ithas proved difficult to find ideal probes and as aconsequence each has its own limitation. In the past,K+ or Rb+ in the presence of saturating con-centrations of the ionophore valinomycin have beenthe probes of choice because of the simplicity of theirassay. There is no evidence for their binding to cellcomponents, but the presence of transport systemsin bacteria for K+ and the major physiological roleof the ion has provoked criticism of the method(Maloney et al., 1975). The choice of Rb+ over K+in E. coli removes some of these criticisms, since thecation is discriminated against by the K+ transportsystems (Rhoads et al., 1977). In the last few years,lipophilic cations have achieved prominence, sincethey combine many features of the valinomycin/Rb+couple with the added advantage that they arenon-physiological cations and thus are less likely tobe actively transported. However, their lipophilicnature leads to great non-specific binding of thecations which may complicate quantification(Zaritsky et al., 1981). The assay of these cations
0306-3283/81/120573-09$01.50/1 ©) 1981 The Biochemical Society

S. Ahmed and I. R. Booth
requires the use of relatively dense cell suspensions,which may in itself cause problems with aeration.Cationic fluorescent dyes on the other hand havethe advantage that through their optical propertiesa continuous monitor of AVt' can be obtained atrelatively low densities of cells. Thus although noone probe is ideal, a combination of several withdifferent limitations should provide corroborativedata with which to test the chemiosmotic model.
Measurement of ApH has presented fewer prob-lems and there is general agreement on its magni-tude (Padan et al., 1976; Booth et al., 1979).However, for accurate quantification there is nosingle probe whch can span a wide range ofexperimental pH values. Benzoic acid and DMO areroutinely used probes, but the former is a well knownfood preservative and can interfere with cellularmetabolism (Eklund, 1980), while the latter has ahigh pK (pK6.32, Addanki et al., 1968) which maylead to inaccuracy at acid pH. Recent measure-ments of ApH by 31P n.m.r. have generally corro-borated the data obtained with weak acids. How-ever, the conditions necessary for n.m.r. are farremoved from the ideal for transport experiments(Navon et al., 1977; Slonczewski et al., 1981).A further critical point in terms of quantification
is that of the magnitude of the intracellular volume.Since it was discovered that the periplasm consti-tuted a significant component of the total cell volumein E. coli and other Gram-negative bacteria(Stock et al., 1977) more care has been exercised inthe choice of probe to measure the true cytoplasmicvolume (Booth et al., 1979; Kashket, 1981).However, few studies adequately determine theeffects of variations in the experimental conditionson cytoplasmic volume, with consequent errors inthe measured parameters, Ay and ApH. Accuratequantification requires that all the potential vari-ables should be measured under identical condi-tions.The present communication concerns a detailed
study of the magnitude of AV, ApH, and cyto-plasmic volume under a variety of experimentalconditions. The data presented indicate the limi-tations of some of the probes of Ay/ and ApH, andare discussed in the context of lactose accumulationin E. coli (Booth et al., 1979).
Materials and methods
MaterialsAll materials were as previously specified (Booth
et al., 1979; Kroll & Booth, 1981) with the followingadditions: 86RbCl and [14C]TPP+ were purchasedfrom The Radiochemical Centre, Amersham;[3HITPP+ was generously donated by Dr. R. Kaback(Roche, Nutley, NJ, U.S.A.) and Dr. P. Overath
(MPI fur Biologie, Tibingen, Germany). Cyaninedye diS-C3-(5) was a generous gift from Dr. A. S.Waggoner, Amherst College, Amherst, MA 01002,U.S.A.
Bacterial strainsExperiments were carried out using E. coli
ML308-225 (lac-i-z-y+a+) supplied by Dr. I. C.West, University of Newcastle, Newcastle-upon-Tyne, U.K. and E. coli TK2240 (F-, trk DJ,trkA405,nag A,lac Z, rha, thi) supplied by Dr. W.Epstein, University of Chicago, Chicago, IL, U.S.A.Growth conditions andEDTA treatment
E. coli ML308-225 was grown and maintained aspreviously described (Booth et al., 1979). E. coliTK2240 was grown on K20 glucose minimalmedium, to repress the Kdp K+ transport system, aspreviously described (Kroll & Booth, 1981). EDTAtreatment was as previously described (Booth et al.,1979).Centrifugation based assays
In order to quantify accurately cytoplasmicvolumes, Ay/, ApH, and Ap ac' the concentration ofradioactively labelled probe inside the cell must beestimated. This can be done either by monitoringchanges in the external concentration using flowdialysis, or by separating the cells from the incu-bation medium using Millipore filtration or centri-fugation. Each of these methods has its advantagesand disadvantages, the ultimate choice being deter-mined by the type of experiments to be done.Centrifugation was the main method chosen in thisstudy for the following reasons. It avoids thepossible binding of probe either to the filter or tosilicone oil. Unlike with flow dialysis, a potential of0-30mV (1-3.2-fold accumulation of probe) can beestimated accurately. Finally all parameters can bemeasured under identical conditions and in parallel.Lactose transport
Cells (2mg dry wt./ml) were equilibrated for 5 minat 250C in a stirred glass vessel with a largesurface-to-volume ratio to ensure continuous aera-tion. [glucono-14C]Lactose (5,uM final concn.;5.77Ci/mol) was then added to the cell suspensiontogether with catalase (250Qg/ml final concn.) and3H20 (1-2,uCi/ml) as a water marker. After afurther 20 min incubation (steady state accumulationunder all experimental conditions was achieved by20min) 500,p samples were transferred to Eppen-dorf centrifuge tubes (1.6ml capacity) containinglO,p of 2vol. H202. These were then spun in anEppendorf centrifuge (model S412) at 12 OOOg for20s. A sample of supernatant (lOO,l) was removedand transferred to a centrifuge tube containing asimilar pellet of non-radioactive cells. The remain-ing supernatant was aspirated off with a vacuum line
1981
574

Quantitative measurements of proton-motive force
(the total time taken from sampling to removal of thesupernatant was approx. 1 min). The cell pellets wereresuspended to a final volume of 300,ul with theappropriate buffer and then transferred to a vialcontaining 2ml of Triton X-1()0/toluene/PPO/POPOP scintillant (333ml of Triton X-100, 667mlof toluene, 4g of PPO and 100mg of POPOP). Thesamples of supernatant and pellet were counted onthe preset 3H/'4C program of a Packard 3255liquid-scintillation counter. Calculation of the lac-tose accumulation ratio requires a knowledge of theexternal water space. Since 3H20 distributes equallythroughout all phases, it overestimates the externalwater space, leading to an underestimation ofinternal probe concentration. This inaccuracy can becorrected for by a knowledge of c.p.m. of 3H20/pll ofsupernatant and ul/mg dry wt. (cytoplasmicvolume).
After correction for contaminating 14C in the cellpellet, the lactose accumulation ratio can be cal-culated by dividing the internal radioactivity by theexternal radioactivity and then adjusting for therelevant volume differences.
Measurement ofApHThe magnitude of the pH gradient was estimated
from the distribution of a weak acid as previouslydescribed (Booth et al., 1979). Either [2-'4CIDMO(5,M final concn.; 28.6Ci/mol) or [7-'4C]benzoicacid (2,UM final concn.; 24.4 Ci/mol) were used.Assay conditions were identical to those describedfor lactose transport. ApH was calculated asdescribed by Booth et al. (1979).
Measurement of AVtThe magnitude of the membrane potential was
estimated in the following ways.(i) Uptake of radioactive cations. 86Rb+ or
[14C]TPP+ (5,M and 4,UM final concn.; -0.2,uCi/mland 8.7 Ci/mol respectively) were used. Assayconditions were as described above except forexperiments with E. coli TK 2240, where cells wereincubated with the probes for 10min beforesampling.
(ii) K+ distribution byflame photometry. This wasas previously described (Booth et al., 1979) exceptcells were incubated for 15min before sampling. Acorrection based on the uncoupler-insensitive cationbinding to cells was applied to (i) and (ii). Themembrane potential was calculated by substitutionof concentration ratios into the Nernst equation. Anactivity coefficient for 86Rb, ['4ClTPP and K+ of 1was assumed.
(iii) Cvanine diS-C3-(5) fluorescence. Measure-ments were made on a Perkin-Elmer 3000 fluori-meter, with excitation at 638nm and emission at670nm. Assays were carried out with 20,ug dry wt.of cells/ml at 25°C in a 3ml quartz cuvette. The
cyanine dye was present at a final concentration of.0.166 uM. All additions were made with Hamiltonmicrosyringes and followed by vigorous stirring. Toensure that the dye was responding to AV alone, K+was added to 150mM, final concn., at the end ofeach assay, causing reversal of fluorescence quench-ing. This concentration of K+ collapsed AV asmeasured by 86Rb+ distribution under similar condi-tions.
Cytoplasmic volumesCytoplasmic volumes were estimated under all
experimental conditions with E. coli ML 308-225 aspreviously described (Booth et al., 1979). In experi-ments with E. coli TK2240 cytoplasmic volumeswere assumed to be 1 ,ul/mg dry wt., as only acomparison of AYIRb and AVTpp was being made,rather than absolute values determined. The proto-col for measurement of cytoplasmic volumes was aspreviously described (Booth et al., 1979). Lactose,Rb+, TPP+, DMO, and benzoic acid at the con-centrations used in this study had no effect oncytoplasmic volumes.
Triple labellingThe simultaneous measurement of AV along with
ApH or lactose accumulation, or AV by both['4C1TPP+ and Rb+ was facilitated by triple labell-ing. 3H20 was present as the water marker, 86Rb+was used for AV and an appropriate 14C-labelledprobe for the third parameter. The Packard 3255liquid-scintillation counter was set up such thatchannels counted 86Rb+ alone, 86Rb+ plus 14C and14C plus 3H. A small, 1-2%, spillover of 86Rb+ intothe 3H channel was unavoidable, but could beeliminated as a source of error by increasing theratio of 3H c.p.m. to 86Rb c.p.m. to approx. 10 to 1.Standard samples of 86Rb+ and 14C were used tocalculate the spillover into the lower energy channel.Thus calculation of the accumulation ratio of 14Cand 86Rb probes involved a correction of the 14Cchannel for Rb2+ overlap and a subsequent cor-rection of the 3H channel for 14C overlap using thecorrected 14C data. An independent validation of themethod was carried out using 3H/14C couples and3H/86Rb+ couples for estimation of AV.Oxygen generating systemTo ensure aerobic conditions were maintained
throughout centrifugation-based assays a catalase/H202 mixture was used to generate oxygen. Theconcentrations of catalase and H202 required tomaintain aerobic conditions for 1 min was calcu-lated from anaerobic oxygen pulse experiments. Thisprecaution of maintaining aeration was found toreduce the variability of ApH measurements andprevented a 40mV depolarization of A,v which wasobserved in centrifuge tubes lacking H202.
Vol. 200
575

S. Ahmed and I. R. Booth
Results
Accurate measurements of the parameters of Aprequire a detailed knowledge of the cytoplasmicvolume and any changes in this volume broughtabout by manipulation of the cells. Recently wereported that the volume of E. coli cells is varied byconditions of growth and experimental protocol(Kroll & Booth, 1981). In the present study thecytoplasmic volume was dependent upon the con-centration of the respiratory substrate, succinate,and also upon the external potassium concentration,but independent of the external pH (pHO). In theabsence of succinate the cytoplasmic volume was1.1 + 0.13 p1 of water/mg dry wt. of cells which wasclose to the previously reported value (Booth et al.,1979). In the presence of 2mM- and 16mM-suc-cinate, however, the volume increased to 1.39 + 0.21and 1.6+0.19,u1 of water/mg dry wt., respectively.When cells were treated with EDTA or EDTA/valinomycin the cell volume increased from1.1+0.13 to 1.8+0.27 and 2.2+0.25pl of water/mg dry wt., respectively, as the K+ concentrationwas raised to 150mM at constant osmolarity.Account must be taken of these changes duringmeasurements of AV and ApH. Failure to correct forthe increase in volume of EDTA/valinomycin-treated cells, for example, would cause A,V to beoverestimated by 20-25 mV at high external K+concentrations.
Measurement ofApHWe compared two acids, DMO and benzoic acid,
as indicators of the internal pH as a function of pH.and in the presence and absence of succinate.Endogenously-respiring cells were found to have acytoplasmic pH (pH,) between 6.5 and 8.25 whenthe external pH was varied from 5.0 to 8.5 (resultsnot shown). DMO indicated a higher value of theintracellular pH than benzoate when pHo was below6.5. This we interpret to be due to the sensitivity ofthe estimation of ApH by DMO to small errors inthe accumulation ratio at pHo values below its pK.The equation derived by Waddell & Butler (1959)for ApH is as follows:
ApH = log [l(o IloPK-PHo+ 1)-lOPK-PH] (2)
where Cl/Co is the transmembrane concentrationratio of the weak acid. At values of pHo below thepK of the acid the expression (IOPK-PHo) becomeslarge and any small error in C1/C0 is magnified.When ApH was reduced at pHO 5.5 by eitheraddition of acetylsalicylic acid or the uncouplertetrachlorosalicylanilide a similar discrepancy wasobserved between the magnitude of ApH as indi-cated by benzoate and DMO (Fig. 1). At low valuesof ApH the difference increased, as does the effect of
1.75
1.50
1.25
.5m 1.00.2
r_c
v 0.75
/00
/ 0, A
, A*
* AA
, In, a
, In
/ 0
AA
, AZ0L0
0 0.25 0.50 0.75 1.00 1.25 1.50 1.75ApHDMO
Fig. 1. Comparison ofApH as measured by benzoate andDMO at pH,S.S in the presence of agents that reduce
ApHEDTA/valinomycin-treated cells were incubated ina Tris (5 mM)/Mes (5 mM)/choline chloride (150mM)buffer system containing 0.2mM-KCI (pH,5.5) at2mg dry wt./ml (Booth et al., 1979). The pHgradient was reduced either by tetrachlorosalicyl-anilide (0-0.8 um, final concn.) or by acetylsalicylicacid (0-9 mm, final concn. at pH, 5.5). The cellswere respiring endogenous reserves and were incu-bated with the probe under the appropriate condi-tions for 5min. (0) Tetrachlorosalicylanilide pre-sent; (A) acetylsalicylic acid present.
.,mall errors in the measurement of the accumula-tion ratio (results not shown). Thus, it may be thatbenzoate more accurately reflects the magnitude ofApH at acid values of pHo.When 2 mM-succinate was added to cell sus-
pensions, the respiratory rate increased 4-fold;however, the effect on ApH was small but sig-nificant (results not shown). Above pHO 5.5 bothbenzoate and DMO indicated the same value ofApH. This reflected a doubling of the internalconcentration of benzoate while that of DMOremained essentially constant. A possible explana-tion for this observation might be that due to thehigh concentration of the undissociated form ofDMO, this weak acid is relatively insensitive to smallchanges in intracellular pH. In the presence ofsuccinate, pH, was relatively constant at pH, 7.1over the range pHO5.0-7.0, but increased rapidlyabove pHO7.0 (results not shown). Above pHO7.5the pH gradient was inverted and was estimated byweak acid exclusion to be more acid by 0.1 pH units.When the succinate concentration was increased to
1981
576
Quantitative measurements of proton-motive force
16mM both pHi and the pH0 at which inversion ofthe pH gradient occurred were found to move to themore acidic values of 6.5 and 7.0, respectively(results not shown). This may reflect a depression ofthe internal pH due to significant accumulation ofthe acidic respiratory substrate. An importantfeature of this result is that failure to correct for thechange in the cytoplasmic volume entirely com-pensates for the decreased DMO accumulation andpHi then appears not to be affected by 16mM-suc-cinate.
Measurement ofAvThe cations, K+, Rb+ and TPP+ performed
almost identically in EDTA/valinomycin-treatedcells of E. coli ML308-225. Using Rb+ and TPP+,the measurement of A V was found to be time-dependent at all values of pH.. At pHo6.0, AVincreased rapidly to 120mV and then rose slowly toa steady value of around 140mV after 15min (Fig.2a). On the other hand, at pH, 8.0 AV was initiallymeasured as around 180mV but then declined sothat after 20min AV was only 150-160mV (Fig.2b). These characteristics were not a function ofEDTA/valinomycin treatment of the cells sincesimilar results were observed in untreated cells withTPP+ as AV probe (Fig. 2a,b). In such cells TPP+responded more slowly. The time dependency couldnot be attributed to properties of either probeindividually. At pH06.0 or 8.0 addition of the probeto cells after a period of incubation led immediatelyto an indicated value of A V equal to that whichwould have been observed had the probe beenpresent from zero time (results not shown). How-ever, the time-dependency could be abolished byincreasing the external K+ concentration to 1 mM
(results not shown) and thus it may relate to somefeature of potassium transport in these cells (Kroll &Booth, 1981).
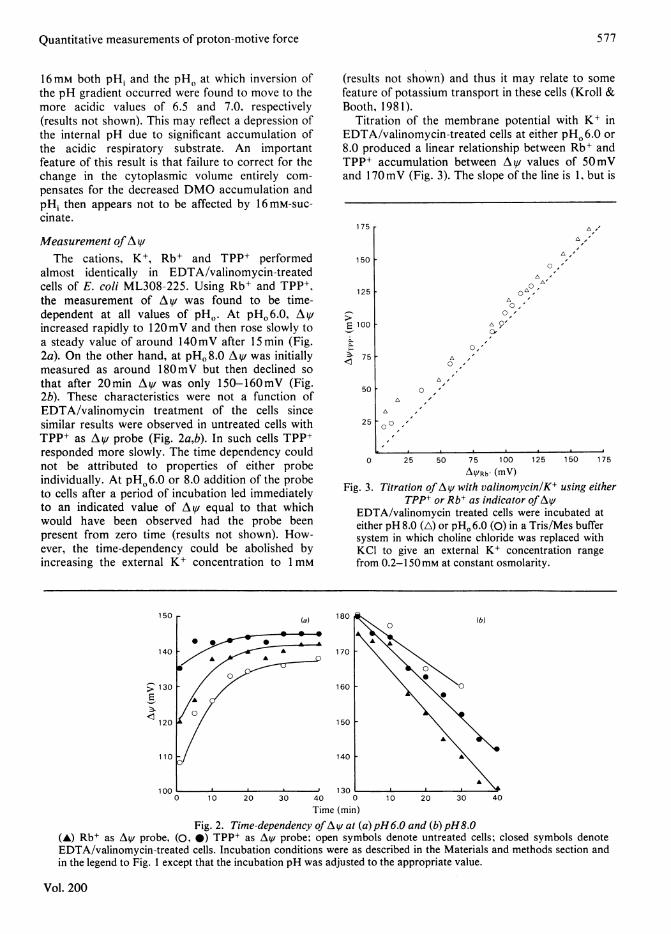
Titration of the membrane potential with K+ inEDTA/valinomycin-treated cells at either pHO6.0 or8.0 produced a linear relationship between Rb+ andTPP+ accumulation between AV values of 50mVand 170mV (Fig. 3). The slope of the line is 1, but is
175 r
150 F
125 [
50
25
0,A-0-"
L'O,'
cvI,
O, '
0 ,
A
0 I0
0 25 50 75 100 125 150 175
AYRb (mV)
Fig. 3. Titration ofAV with valinomycin/K+ using eitherTPP+ or Rb+ as indicator of At1/
EDTA/valinomycin treated cells were incubated ateither pH 8.0 (A) or pH. 6.0 (0) in a Tris/Mes buffersystem in which choline chloride was replaced withKCI to give an external K+ concentration rangefrom 0.2-150mM at constant osmolarity.
150
140
(a)
0 1 0 20 30 40
Time (min)Fig. 2. Time-dependency ofAy at (a) pH6.O and (b)pH8.0
(A) Rb+ as AV probe, (0, *) TPP+ as AV probe; open symbols denote untreated cells; closed symbols denoteEDTA/valinomycin-treated cells. Incubation conditions were as described in the Materials and methods section andin the legend to Fig. 1 except that the incubation pH was adjusted to the appropriate value.
Vol. 200
577
1-
E 1001-10.M.
;.). 75< I
S. Ahmed and I. R. Booth
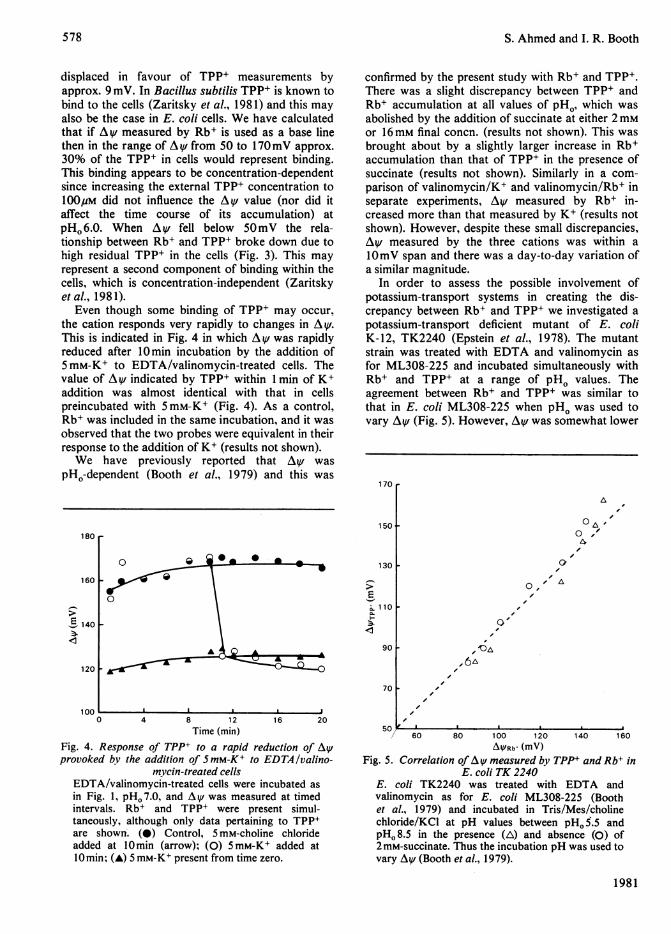
displaced in favour of TPP+ measurements byapprox. 9 mV. In Bacillus subtilis TPP+ is known tobind to the cells (Zaritsky et al., 1981) and this mayalso be the case in E. coli cells. We have calculatedthat if A V measured by Rb+ is used as a base linethen in the range of A^V from 50 to 170mV approx.30% of the TPP+ in cells would represent binding.This binding appears to be concentration-dependentsince increasing the external TPP+ concentration to100pM did not influence the Av value (nor did itaffect the time course of its accumulation) atpHO 6.0. When Av fell below 5OmV the rela-tionship between Rb+ and TPP+ broke down due tohigh residual TPP+ in the cells (Fig. 3). This mayrepresent a second component of binding within thecells, which is concentration-independent (Zaritskyetal., 1981).
Even though some binding of TPP+ may occur,the cation responds very rapidly to changes in AV.This is indicated in Fig. 4 in which A v was rapidlyreduced after 10min incubation by the addition of5mM-K+ to EDTA/valinomycin-treated cells. Thevalue of Av indicated by TPP+ within 1 min of K+addition was almost identical with that in cellspreincubated with 5mM-K+ (Fig. 4). As a control,Rb+ was included in the same incubation, and it wasobserved that the two probes were equivalent in theirresponse to the addition of K+ (results not shown).We have previously reported that Ay was
pHo-dependent (Booth et al., 1979) and this was
confirmed by the present study with Rb+ and TPP+.There was a slight discrepancy between TPP+ andRb+ accumulation at all values of pHo, which wasabolished by the addition of succinate at either 2mMor 16mm final concn. (results not shown). This wasbrought about by a slightly larger increase in Rb+accumulation than that of TPP+ in the presence ofsuccinate (results not shown). Similarly in a com-parison of valinomycin/K+ and valinomycin/Rb+ inseparate experiments, Ai,v measured by Rb+ in-creased more than that measured by K+ (results notshown). However, despite these small discrepancies,A,t measured by the three cations was within alOmV span and there was a day-to-day variation ofa similar magnitude.
In order to assess the possible involvement ofpotassium-transport systems in creating the dis-crepancy between Rb+ and TPP+ we investigated apotassium-transport deficient mutant of E. coliK-12, TK2240 (Epstein et al., 1978). The mutantstrain was treated with EDTA and valinomycin asfor ML308-225 and incubated simultaneously withRb+ and TPP+ at a range of pHo values. Theagreement between Rb+ and TPP+ was similar tothat in E. coli ML308-225 when pHo was used tovary Ay/ (Fig. 5). However, A ,v was somewhat lower
170
150 .180 r
160
E 140
120
100
0 I0 a 0 a
0
*A. , F,~~~A--
130
1-E
90 I
O -
0
0 , A
0'
/
70 F
0 4 8 12 16 20Time (min)
Fig. 4. Response of TPP+ to a rapid reduction of Ay/provoked by the addition of SmM-K+ to EDTA/valino-
mycin-treated cellsEDTA/valinomycin-treated cells were incubated asin Fig. 1, pH. 7.0, and Ay/ was measured at timedintervals. Rb+ and TPP+ were present simul-taneously, although only data pertaining to TPP+are shown. (0) Control, 5 mM-choline chlorideadded at 10min (arrow); (0) 5mM-K+ added at10min; (A) 5 mM-K+ present from time zero.
Fi
60 80 100 120 140 160A5Rb' (mV)
ig. 5. Correlation of Ay/ measured by TPP+ and Rb+ inE. coli TK 2240
E. coli TK2240 was treated with EDTA andvalinomycin as for E. coli ML308-225 (Boothet al., 1979) and incubated in Tris/Mes/cholinechloride/KCI at pH values between pHo 5.5 andpHo 8.5 in the presence (A) and absence (0) of2 mM-succinate. Thus the incubation pH was used tovary Ay/ (Booth et al., 1979).
1981
578

Quantitative measurements of proton-motive force
in this strain possibly due to errors in the volume,but also due to strain differences (Kroll & Booth,1981). Thus, the presence of potassium transportsystems does not unduly influence the relationshipbetween Rb+ and TPP+ as membrane potentialprobes.
In conclusion, these three cations, which aresubject to different limitations of applicability,suggest similar values of AV in E. coli ML308-225.This study also reveals that other factors maycontribute to the magnitude of the indicated AV.Cyanine-dye fluorescenceThe fluorescent dye cyanine-diS-C3-(5) can be
used as a membrane potential indicator in E. colicells (Letellier & Schechter, 1979; Ghazi et al.,1981). We studied this probe to ascertain itsusefulness as a continuous AV/ indicator. However,while this work was in progress a similar study hasbeen published which is in essence identical with ourown (Ghazi et al., 1981) and thus we will onlysummarize our observations.We calibrated the quenching of dye fluorescence
at 670nm by valinomycin/K+ titration at pHO8.0and found that the relationship was non-linear overthe whole range of Ay/ from 1OOmV to 170mV.Below 1OOmV changes in fluorescence quenchingwere not evident. In a time course there was no timedependency of AV, but the dye did respond in thepredicted manner to addition of 5mM-K+ (resultsnot shown). On the other hand, addition of 4mM-lactose did not depolarize the cells, which wasobserved to occur with either TPP+ or Rb+ as A ,vprobe. Thus, the dye responded rapidly to largechanges in Ay but did not appear to be sensitive tosmaller slow changes. The quenching of dyefluorescence showed a similar dependence on pHo asAy measured by other probes, but below pHo 7.0 thevalues indicated on the basis of calibration at pHo 8.0were some 2OmV lower. Thus, although the cyaninedye indicates broadly similar characteristics of AVits quantification appears to be of very limitedvalidity.
Lactose transportThe uptake of lactose was measured under the
same conditions as AV, ApH and cytoplasmicvolume and was found to reach a steady stateat around 20 min. The accumulation, Apiac, wassimilar to that observed previously (Booth et al.,1979) and was only significantly stimulated by suc-cinate between pHO5.0 and 6.5 (results not shown).The relationship of the lactose accumulation to Ap(APiac/AP) has been defined as the 'effective orapparent stoichiometry' (Booth, 1981). As can beseen, the value of the apparent stoichiometry dependsto a significant extent on the experimental condi-tions (Fig. 6) and upon the probes used. In the
Vol. 200
1.0 r
0.9
a 0.8
0.7
0.6 L
0 O
0
5.0 6.0 7.0 8.0pH.
Fig. 6. Variation ofthe apparent stoichiometrv (APlac/AP)as a/unction ofpH0
The apparent stoichiometry was calculated from thelactose accumulation (results not shown) and themagnitude of AV (Rb+) and ApH (benzoate). Theincubation system was as described in Fig. 1 withthe pH of the buffer system adjusted to theappropriate pH value. (0) endogenously-respiringcells, (A) cells respiring succinate (2mM). Allmeasurements were made at the steady state, 20minafter the addition of lactose.
presence of 2 mM-succinate, the least equivocalresults were obtained since benzoate and DMOwere in agreement on ApH, and Rb+ and TPP+similarly agreed on AiV. Thus in this instance theapparent stoichiometry is markedly pHo-dependent,rising from 0.68 at pHO5.0 to 0.94 at pHO8.0. Inendogenously-respiring cells the apparent stoichio-metry is dependent upon which probes are used forApH and Ay/. Using the lowest values (benzoate andRb+, respectively) the apparent stoichiometry waslargely pHo-independent between pHO 5.0 andpHo 7.0 at around 0.82; above pHo 7.0 the apparentstoichiometry increased to 0.96 at pHO8.0 (Fig. 6).Notwithstanding these differences the conclusion isthat the bulk phase Ap is adequate to account for thelactose accumulation observed in cells.
Discussion
The data presented in this paper confirm andextend our earlier observations on the relationshipbetween Ap and lactose accumulation (Booth et al.,1979; Booth & Hamilton, 1980b; Booth, 1980).However, the importance of this study is that itestablishes that independent measurements of AsVand ApH using different probes corroborate eachother. With respect to Ay/ probes this has beenlacking from previous studies (Ghazi et al., 1981)which also fall down particularly on assumptionsrelating to the cytoplasmic volume. In the presentstudy Ay, ApH, cytoplasmic volume and lactose
5-79

580 S. Ahmed and 1. R. Booth
transport have been examined under the sameconditions, using centrifugation-based assays.The cytoplasmic volume of cells was found to be
a significant experimental variable. This was shownmost markedly when AV was titrated to below50mV, where underestimation of the cell volumeleads to overestimation of AV, by up to 3OmV.Similarly, if the volume is overestimated this maylead to an underestimate of the measured para-meter. With regard to ApH, the effect of volumechanges is most dramatic when the accumulation ofthe weak acid is small, as the case of DMO atpHO6.5 in the presence of 16mM-succinate. Failureto correct for the volume changes raise ApH from0.31 units to 0.53 units. Thus, it is essential inquantitative studies of bioenergetics that the cyto-plasmic volume be treated as an experimentalvariable.
In an earlier study of ApH in endogenously-respiring cells we used DMO and a membrane-filtration technique to separate the cells from theincubation medium. However, a subsequent studyrevealed that DMO bound in a pH-dependentfashion to membrane filters (C. V. Salmond, W. A.Hamilton & I. R. Booth, unpublished results). Thepresent study confirms the general trend of ourearlier results and indeed emphasizes the lack of aconstant internal pH in endogenously-respiring cells.Two weak acids of dissimilar structure and differentpK values gave close agreement at pHo values abovetheir pK's. DMO at an incubation pH below its pKgave slightly higher values than did benzoic acid andthis may be related to the sensitivity of the assay tosmall errors under these conditions. Benzoic acid is afood preservative; however, at the concentrationused it did not lower ApH as measured by DMO(C. V. Salmond, W. A. Hamilton & I. R. Booth,unpublished results). The failure ofDMO to show anincrease in ApH when succinate was added to thecells may reflect its insensitivity to small changes inApH when used at pHo values below its pK. WhenApH was titrated with uncoupler or acetylsalicylicacid at pHO5.5, the discrepancy between ApHmeasured by DMO and that indicated by benzoicacid increased as ApH decreased (Fig. 1). Thisappears to confirm the suggestion that DMO is notwholly reliable at such low values of pHo.
With cells respiring succinate (2mM), pHi wasmaintained relatively constant and above pH0 7.5ApH was inverted to become acid inside. Thisexplains one of the significant differences betweenour previous work (Booth et al., 1979) and that ofother groups (Padan et al., 1976; Zilberstein et al.,1979). Our earlier failure to find ApH inversion-waswith endogenously-respiring cells, whereas the othergroups provided succinate as a respiratory sub-strate. However, there is still a slight discrepancy inthe actual value of pHi.
The two probes of AV which were investigated indetail, Rb+ and TPP+, gave reasonable agreement.There may be a significant binding of TPP+ since itregistered higher values of AiV in endogenously-respiring cells and its accumulation was not com-pletely abolished by 150mM-K+/valinomycin (Fig.3). This, of course, presupposes the validity of thevalinomycin/Rb+ measurements. Certainly anyerror in the Rb+ measurement does not appear to bedue to K+ transport systems, since in the mutantlacking such activities a similar discrepancy betweenTPP+ and Rb+ was observed. However, onedisturbing feature of Rb+ in E. coli ML308-225 wasthat its accumulation increased relative to TPP+ orK+ in the presence of succinate (results not shown).We do not have an adequate explanation for this atpresent.On the whole Rb+ and TPP+ gave similar values
of AV, showed similar time dependencies andresponded identically to sudden partial depolariza-tion of the membrane. Thus, it seems probable thatTPP+ may be a quantitatively useful AV indicator inwhole cells that have not been treated with EDTAand valinomycin. Most importantly, the AV, valuesobserved with TPP+ in untreated cells were verysimilar (within 11 mV) to the magnitude of AV intreated cells. In the succeeding paper we willillustrate the usefulness of this in further examiningenergy coupling to lactose accumulation in E. coli.
The authors thank Professor W. A. Hamilton for hisgenerosity and encouragement throughout this work. Wealso acknowledge our co-workers Dr. R. G. Kroll, Mrs.C. V. Salmond and Miss F. Thomson for their helpfuladvice, and Dr. D. Burke, Department of Pharmacology,for the use of the fluorimeter. Thanks are also due to Dr.R. M. Macnab, Dr. R. Kaback, Dr. P. Overath, Dr. D.Nicholls for preprints of their papers, chemicals andadvice. S. A. is in receipt of financial support from theScience Research Council (grant GR/A.99299) to whomwe express our thanks.
References
Addanki, S., Cahill, F. D. & Sotos, J. F. (1968) J. Biol.Chem. 243, 2337-2348
Booth, I. R. (1980) Biochem. Soc. Trans. 8, 276-278Booth, I. R. (1981) Trends Biochem. Sci., in the pressBooth, I. R. & Hamilton, W. A. (1980a) Biochem. J. 188.467-473
Booth, I. R. & Hamilton, W. A. (1980b) in Micro-organisms and Nitrogen Sources (Payne, J. W., ed.).pp. 171-207, John Wiley and Sons, London
Booth, I. R., Mitchell, W. J. & Hamilton W. A. (1979)Biochem. J. 182, 687-696
Eklund, T. (1980) J. Appl. Bacteriol. 48, 423-432Epstein, W., Whitelaw, V. & Hesse, J. (1978) J. Biol.
Chem. 253, 6666-6668Ghazi, A., Schechter, E., Letellier, L. & Labedan, B.
(1981) FEBS Lett. 125, 197-200
1981

Quantitative measurements of proton-motive force 581
Hamilton, W. A. (1975) Adv. Microbial Phvsiol. 12.1-53
Harold, F. M. (1976) Curr. Top. Bioenerg. 6, 83-149Kashket, E. R. (1981)J. Bacteriol. 146. 377-384Kell, D. B. (1979) Biochim. Biophvs. Acta 549, 55-99Kell, D. B., Clarke, D. J. & Morris, J. G. (1981) FEMS
Microbiol. Lett. 1 1, 1-1I1Kroll, R. G. & Booth, I. R. (1981) Biochem. J. 198,
691-698Letellier, L. & Schechter, E. (1979) Eur. J. Biochem. 102,
441-447Maloney, P. C., Kashket, E. R. & Wilson, T. H. (1975)
Methods Membr. Biol. 5, 1-49Mitchell, P. (1963) in Organisation and Control in
Prokarvotic and Eukarvotic Cells (Charles, P. &Knight, B. C. J. G., eds.), pp. 121-166, CambridgeUniversity Press
Mitchell, P. (1981) Chem. Brit. 17, 14-23
Navon, G., Ogawa, S., Shulman, R. G. & Yamane, T.(1977) Proc. Natl. A cad. Sci. U.SA. 74, 87-97
Padan, E., Zilberstein, D. & Rottenberg, H. (1976) Eur.J. Biochem. 63, 533-541
Ramos, S. & Kaback, H. R. (1977) Biochemistr.v 16,854-859
Rhoads, D. B., Woo, A. & Epstein, W. (1977) Biochim.Biopkvs. Acta 469,45-51
Slonczewski, J. L., Rosen, B. P.. Alger. J. R. & Macnab.R. M. (1981) Proc. Natl. A cad. Sci. U.S.A. in the press
Stock, J., Rauch, B. & Roseman, S. (1977)J. Biol. Chem.252, 7580-7862
Waddell, W. J. & Butler, T. C. (1959) J. Clin. Invest. 38.720-729
Zaritsky, A., Kihara, M. & Macnab, R. M. (1981) J.Membr. Biol., in the press
Zilberstein, D., Schuldiner, S. & Padan, E. (1979)Biochemistrv 18, 669-673
Vol. 200





























