Embed Size (px)
Citation preview

Pergamon 0022-1910(94)00093-X
J. Insect Physiol. Vol. 41, No. 1, pp. 17-22, 1995 Copyright 0 1995 Elsevier Science Ltd
Printed in Great Britain. All rights reserved 0022-1910/95 $9.50+0.00
Quantification of the Effect of Bacillus thuringiensis Toxins on Short-circuit Current in the Midgut of Bombyx mori BAERBEL LIEBIG,*? DAVID L. STETSONJ DONALD H. DEAN?5
Received 10 June 1994; revised 26 July 1994
We examined the abilities of two Bacillus thuringiensis insecticidal toxins, CryIAa and CryIAc, prepared from Escherichia c&cloned gene products, to inhibit short-circuit current in midgut epithelia of Bombyx mod. Voltage-clamp studies were conducted on isolated midguts, measuring the inhibition of short-circuit current (I& by trypsin-activated toxins. Three bathing solutions were compared and the medium of Chamberlin [(1990) J. exp. Biol. 150, 467-4711 was found to maintain the highest Zsc for the longest time. For CryIAa, a concentration range between 0.33 and 8.0 ng/ml resulted in inhibition of Zsc at the rates of - 0.91 gA/min (lag time, 9 min) and - 7.13 ZrA/min (Iag time, 4 min) respectively, showing a correlation between toxin concentration and inhibitory response. Concentrations greater than 1.6 ng/ml showed diminishing additional effects on the Zs, response, indicating an approach to saturation. The lag times decreased with increasing concentration of toxin applied. For CryIAc, the lowest concentration that gave a response was 3.2 ng/ml (slope, -0.31 pAlmin; lag time, 2 min). There also was a linear correlation between toxin concentration and response for CryIAc, but effective concentrations of CryIAc were approximately 2 orders of magnitude greater than those of CryIAa.
&Endotoxin CryIA Insecticidal crystal proteins
INTRODUCTION
Previous studies on the effect of Bacillus thuringiensis toxin on the electrophysiological properties of the insect midgut have generally reported qualitative effects of the toxin-spore mixture (Harvey and Wolfersberger, 1979; Wolfersberger and Spaeth, 1987). In the one study where purified proteins were used (Crawford and Harvey, 1988), the preparation consisted of the mixture of Cry proteins present in the HD- 1 strain of B. thuringiensis var. kurstaki. In the present study, we take advantage of purified, cloned gene products (Ge et al., 1990) and electrophysiological techniques to compare the effect of individual toxins on short-circuit current in the midgut of Bombyx mori.
Transporting epithelia can be modeled as equivalent electrical circuits, with electron-motive forces represent- ing the driving forces of ion pumps, conductances representing the permeability of the epithelia to movement of ions, and currents representing the flow of
*Present address: Justus von Liebig St. 27, 64401 Gross-Bieberau,
Germany.
TDepartment of Biochemistry, The Ohio State University, Columbus, OH 43210, U.S.A.
fDepartment of Zoology, The Ohio State University, Columbus, OH 43210, U.S.A.
§To whom all correspondence should be addressed.
ions from one side of the epithelium to the other. The voltage-clamp technique permits the determination of each of these circuit parameters (using Ohm’s law) as net values for the entire epithelium and to monitor the flow
of the ions across the epithelium during physiological maneuvers that might change the rates of transepithelial ion flow.
The lepidopteran midgut goblet cell secretes ions into the gut lumen (Wolfersberger, 1992). Flux measurements by Harvey and Zerahn (1972) showed that potassium is the principal ion transported by the midgut. The actions of proton pumps and potassium-H+ antiporters create a potassium flux across the apical membrane (Wieczorek et al., 1991). This in turn creates a potential difference (PD) that has been measured as 180 mV (lumen positive) in Manduca sexta, the tobacco hornworm (Harvey and Wolfersberger, 1979); and 6&70 mV for three other insects, Philosamia Cynthia, Macrothylatia rubi, and B. mori (Giordana and Sacchi, 1977).
Another important process in the goblet cells is the activity of carbonic anhydrase. This enzyme hydrates CO*, creating bicarbonate anions and hydrogen ions. Cellular potassium ion is exchanged for luminal hydrogen ion forming potassium carbonate. The sum of these processes leads to an alkaline pH in the midgut lumen. Both conditions are believed to enable the lepidopteran
17

18 BAERBEL LIEBIG et al.
intestine to buffer the tannic acid in the dietary foliage (Dow, 1986) or otherwise enhance digestion. Potassium ions and the transepithelial potential difference also participate in transport of amino acids into the columnar epithelial cells by driving amino acid symports (Giordana et al., 1982).
The microvillated apical membrane of the columnar epithelial cell is the target of the B. thuringiensis
8-endotoxin. The &endotoxin protoxin is proteolytically activated by trypsin-like midgut proteases and binds to receptors on the epithelial microvilli. The toxin forms pores in the epithelial cell, reducing the electrochemical gradient for potassium. In a voltage-clamp experiment, if b-endotoxin is introduced to the luminal side of the membrane, a decline in the net short-circuit current (Is,) may be observed. This decline occurs after a lag time and is caused either by a decrease in the basal-to-apical current or by an increase in the current in the opposite direction. As Harvey and Wolfersberger (1979) have shown, the B. thuringiensis toxin decreases the Is, by increasing the apical-to-basal K+ flux.
In the present study, we show that there is a quantifiable relationship between the amount of applied toxin and the resulting decline in Zsc. As a parameter for our observation, we have chosen the change in short-circuit current over time, measured as a negative slope of the Zsc decay. We compared two different types of CryIA lepidopteran-specific toxin which differ in their larvacidal activity to B. mori.
MATERIALS AND METHODS
Insect rearing
The eggs and larvae of B. mori (Lepidoptera, Bombycidae) were a gift of Ross Milne, Forestry Pest Management Institute (FPMI), Sault Ste. Marie, Ontario, Canada. The rearing conditions followed the protocol described in Nayar and Fraenkel (1963). Mulberry (Torus afba) leaves were either provided by FPMI, picked from local mulberry trees in Columbus, OH, or from trees in our greenhouse (obtained from FPMI). For voltage-clamp studies, fifth instar larvae, weighing 0.6-0.9 g, were used.
Preparation of insecticidal crystal protein
Escherichia coli strains, which over-express cryZA genes from B. thuringiensis var. kurstaki on plasmid JMl03, were described previously in Ge et al. (1990). OSU4101 produces CryIAa protoxin, and OSU4201 produces CryIAc protoxin. The proteins were examined by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS- PAGE) (Laemmli and Favre, 1973). Protein concen- trations were determined using Coomassie Blue Protein Assay Reagent or BCA Protein Assay Reagent (Pierce Chemical Company). The protoxin was solubilized in 0.05 M sodium bicarbonate buffer (pH 9.5) with l&30 mM dithiothreitol at 36°C for up to 3 h. Active toxin was generated by digestion with l-2% trypsin
(Sigma) for 2 h at 37°C. Subsequently, a SDS-PAGE gel (9% or 12%) was prepared in order to monitor the degree of digestion and the amount of toxin generated. The percentage of active toxin was determined by scanning the gel with a densitometer (Transmittance/Reflectance Scanning Densitometer, Hoefer Scientific Instruments). A plotter recorded the areas of density as curves, which were integrated by an Electronics Graphics Calculator (Numonics Corporation). Solubilized protein was stored at 4°C. The quality of the proteins was checked monthly by a SDS-PAGE and bioassay.
Voltage clamping
The overall setup followed that described by Harvey et al. (1990). The amplifier equipment for voltage clamp consisted of a d.c. 1000 Voltage/Current Clamp, an A-3 10 Accupulser (both from World Precision Instruments), and a strip chart recorder (Kipp and Zonen). These were linked to a Macintosh computer with a MacLab data acquisition system. Transepithelial resistance was deter- mined from Ohm’s law based on the current response of the epithelium to a unidirectional, transepithelial voltage pulse of 5 mV, 200 ms in duration, applied by the stimulator.
Calomel electrodes served as voltage measuring devices. KC1 agar bridges connected the electrodes to the chamber and were made from polyethylene tubing PE-190 with i.d. 1.19 mm (Clay Adams Becton, Dickinson & Co.) filled with 4% agar in 3 M KCl. Silver plate electrodes passed the short-circuit current.
The chambers, holders, and disks (Dow et al., 1985) were constructed by the Medical Store, Ohio State University, Columbus, OH, and were modeled from the chambers provided by M. Wolfersberger, Temple University, Philadelphia, PA, with two exceptions. The variable electrode has been removed and the chambers have been made of equal volume; each side holds 3.5 ml liquid. The apertures of the disk used in this study were 3.9 mm in dia.
Bathing solutions
For most experiments, we used the medium described by Chamberlin (1990): 2.0 mM Hepes, 5.0 mM MgCl*, 1.0 mM CaCl,, 5.8 mM KOH, 9.0 mM NaOH, 3.0 mM Na-methylsulfate, 7.7 mM K-citrate, 2.8 mM Na-succi- nate, 5.6 mM malic acid, 2.0 mM glucose, 27.0 mM trehalose dihydrate, 9.4 mM glutamine, 8.9 mM serine, 7.4 mM proline, 12.8 mM glycine, 4.6 mM threonine, 3.6 mM alanine, 23.1 mM N-methyl-D-glucamine, 140 M PEG-400. After bubbling the medium vigorously with O2 for 2 h, the pH was adjusted to 6.7. Afterwards, the medium was sterile-filtered through a 0.2 pm membrane (Millipore). During the experiment, the bathing solution was bubbled continuously with 02. The bathing solution was identical on both sides of the midgut. All experiments, except for the two indicated, were performed with the Chamberlin medium as the bathing solution. In optimizing the method, we compared the Chamberlin
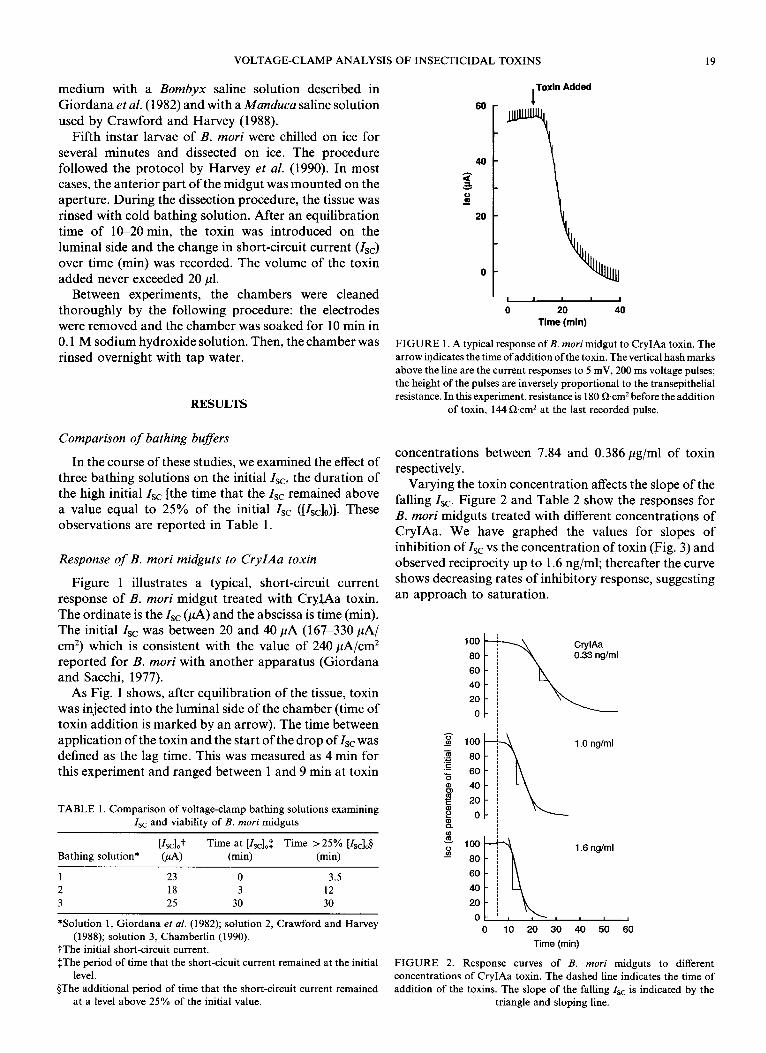
VOLTAGE-CLAMP ANALYSIS OF INSECTICIDAL TOXINS 19
medium with a Bombyx saline solution described in Giordana et al. (1982) and with a Manduca saline solution used by Crawford and Harvey (1988).
Fifth instar larvae of B. mori were chilled on ice for several minutes and dissected on ice. The procedure followed the protocol by Harvey et al. (1990). In most cases, the anterior part of the midgut was mounted on the aperture. During the dissection procedure, the tissue was rinsed with cold bathing solution. After an equilibration time of l&20 min, the toxin was introduced on the luminal side and the change in short-circuit current (I& over time (min) was recorded. The volume of the toxin added never exceeded 20 ~1.
Between experiments, the chambers were cleaned thoroughly by the following procedure: the electrodes were removed and the chamber was soaked for 10 min in 0.1 M sodium hydroxide solution. Then, the chamber was rinsed overnight with tap water.
RESULTS
Comparison of bathing buglers
In the course of these studies, we examined the effect of three bathing solutions on the initial Zsc, the duration of the high initial Zsc [the time that the Zsc remained above a value equal to 25% of the initial Zsc ([Z&J]. These observations are reported in Table 1.
Response of B. mori midguts to CryZAa toxin
Figure 1 illustrates a typical, short-circuit current response of B. mori midgut treated with CryIAa toxin. The ordinate is the I,, (PA) and the abscissa is time (min). The initial Is, was between 20 and 40 ,uA (167-330 PA/ cm’) which is consistent with the value of 240 ,uA/cm* reported for B. mori with another apparatus (Giordana and Sacchi, 1977).
As Fig. 1 shows, after equilibration of the tissue, toxin was injected into the luminal side of the chamber (time of toxin addition is marked by an arrow). The time between application of the toxin and the start of the drop of I,, was defined as the lag time. This was measured as 4 min for this experiment and ranged between 1 and 9 min at toxin
TABLE 1. Comparison of voltage-clamp bathing solutions examining Zsc and viability of B. mori midguts
lZscl0,t Time at [Zsc],$ Time ~25% [Is&
Bathing solution* ‘&A) (min) (min)
1 23 0 3.5
2 18 3 12
3 25 30 30
*Solution 1, Giordana er al. (1982); solution 2, Crawford and Harvey
(1988); solution 3, Chamberlin (1990).
tThe initial short-circuit current. $The period of time that the short-cicuit current remained at the initial
level.
§The additional period of time that the short-circuit current remained at a level above 25% of the initial value.
, Toxin Added
60
s 8 t \
20
0
t L I
I I 1
0 20 40
Time (min)
FIGURE 1. A typical response of B. mori midgut to CryIAa toxin. The
arrow indicates the time ofaddition of the toxin. The vertical hash marks
above the line are the current responses to 5 mV, 200 ms voltage pulses;
the height of the pulses are inversely proportional to the transepithelial
resistance. In this experiment, resistance is 180 &cm2 before the addition of toxin, 144 0cm2 at the last recorded pulse.
concentrations between 7.84 and 0.386 pg/ml of toxin respectively.
Varying the toxin concentration affects the slope of the falling Is,. Figure 2 and Table 2 show the responses for B. mori midguts treated with different concentrations of CryIAa. We have graphed the values for slopes of inhibition of Zsc vs the concentration of toxin (Fig. 3) and observed reciprocity up to 1.6 ng/ml; thereafter the curve shows decreasing rates of inhibitory response, suggesting an approach to saturation.
1 .O ng/ml
3 P 1.6 ng/ml
0 10 20 30 40 50 60
Time (min)
FIGURE 2. Response curves of B. mori midguts to different concentrations of CryIAa toxin. The dashed line indicates the time of
addition of the toxins. The slope of the falling Zsc is indicated by the
triangle and sloping line.
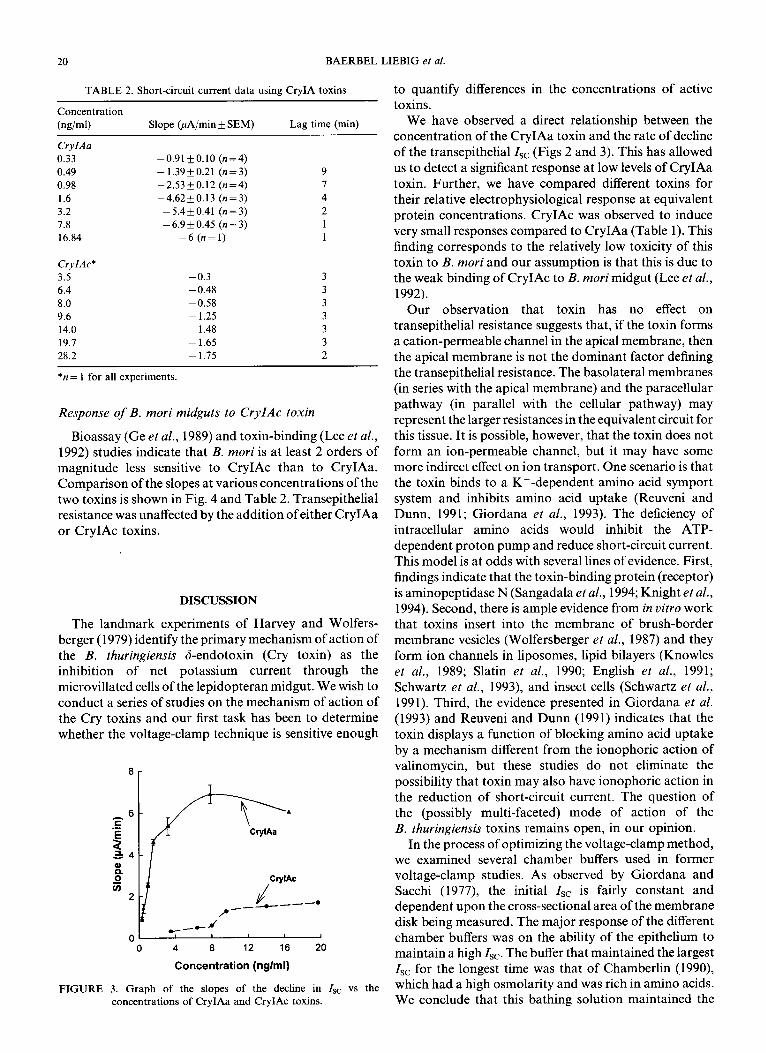
20 BAERBEL LIEBIG et al.
TABLE 2. Short-circuit current data using CryIA toxins
Concentration
(ngiml) Slope (hA/min + SEM) Lag time (min)
CryIAa
0.33
0.49
0.98
1.6
3.2
7.8
16.84
CryIAc*
3.5
6.4
8.0
9.6
14.0
19.7
28.2
-0.91 kO.10 (n=4)
-1.39k0.21 (n=3)
-2.53+0.12 (n=4)
-4.62k0.13 (n=3)
-5.4*0.41 (n=3)
- 6.9 f 0.45 (n = 3)
-6 (n= 1)
-0.3
- 0.48
-0.58
- 1.25
- 1.48
-1.65
-1.75
*n = 1 for all experiments.
Response of B. mori midguts to CryIAc toxin
Bioassay (Ge et al., 1989) and toxin-binding (Lee et al.,
1992) studies indicate that B. mori is at least 2 orders of magnitude less sensitive to CryIAc than to CryIAa. Comparison of the slopes at various concentrations of the two toxins is shown in Fig. 4 and Table 2. Transepithelial resistance was unaffected by the addition of either CryIAa or CryIAc toxins.
DISCUSSION
The landmark experiments of Harvey and Wolfers- berger (1979) identify the primary mechanism of action of the B. thuringiensis d-endotoxin (Cry toxin) as the inhibition of net potassium current through the microvillated cells of the lepidopteran midgut. We wish to conduct a series of studies on the mechanism of action of the Cry toxins and our first task has been to determine whether the voltage-clamp technique is sensitive enough
8r
.
0 4 8 12 16 20
Concentration (nglml)
FIGURE 3. Graph of the slopes of the decline in Is, vs the concentrations of CryIAa and CryIAc toxins.
to quantify differences in the concentrations of active toxins.
We have observed a direct relationship between the concentration of the CryIAa toxin and the rate of decline of the transepithelial1sc (Figs 2 and 3). This has allowed us to detect a significant response at low levels of CryIAa toxin. Further, we have compared different toxins for their relative electrophysiological response at equivalent protein concentrations. CryIAc was observed to induce very small responses compared to CryIAa (Table 1). This finding corresponds to the relatively low toxicity of this toxin to B. mori and our assumption is that this is due to the weak binding of CryIAc to B. mori midgut (Lee et al., 1992).
Our observation that toxin has no effect on transepithelial resistance suggests that, if the toxin forms a cation-permeable channel in the apical membrane, then the apical membrane is not the dominant factor defining the transepithelial resistance. The basolateral membranes (in series with the apical membrane) and the paracellular pathway (in parallel with the cellular pathway) may represent the larger resistances in the equivalent circuit for this tissue. It is possible, however, that the toxin does not form an ion-permeable channel, but it may have some more indirect effect on ion transport. One scenario is that the toxin binds to a K+-dependent amino acid symport system and inhibits amino acid uptake (Reuveni and Dunn, 1991; Giordana et al., 1993). The deficiency of intracellular amino acids would inhibit the ATP- dependent proton pump and reduce short-circuit current. This model is at odds with several lines of evidence. First, findings indicate that the toxin-binding protein (receptor) is aminopeptidase N (Sangadala et al., 1994; Knight et al., 1994). Second, there is ample evidence from in vitro work that toxins insert into the membrane of brush-border membrane vesicles (Wolfersberger et al., 1987) and they form ion channels in liposomes, lipid bilayers (Knowles et al., 1989; Slatin et al., 1990; English et al., 1991; Schwartz et al., 1993), and insect cells (Schwartz et al., 1991). Third, the evidence presented in Giordana et al. (1993) and Reuveni and Dunn (1991) indicates that the toxin displays a function of blocking amino acid uptake by a mechanism different from the ionophoric action of valinomycin, but these studies do not eliminate the possibility that toxin may also have ionophoric action in the reduction of short-circuit current. The question of the (possibly multi-faceted) mode of action of the B. thuringiensis toxins remains open, in our opinion.
In the process of optimizing the voltage-clamp method, we examined several chamber buffers used in former voltage-clamp studies. As observed by Giordana and Sacchi (1977) the initial Z,, is fairly constant and dependent upon the cross-sectional area of the membrane disk being measured. The major response of the different chamber buffers was on the ability of the epithelium to maintain a high Isc. The buffer that maintained the largest Zsc for the longest time was that of Chamberlin (1990) which had a high osmolarity and was rich in amino acids. We conclude that this bathing solution maintained the

VOLTAGE-CLAMP ANALYSIS OF INSECTICIDAL TOXINS 21
viability of the midgut cells and hence the activity of the
ion pumps. Chamberlin medium has an osmotic pressure of 416 mOsm (courtesy of Dr F. Hink, The Ohio State
University), and F. Hink (personal communication)
recommends that the osmotic pressure for insect tissue cultures should range between 360 and 370 mOsm. This is consistent with the measurement of 370 mOsm as the osmolarity of the hemolymph from fifth instar larvae of B. mori (Rouschal, 1940). The chamber buffers which were used by Harvey and Wolfersberger (1979) and Giordana et al. (1982) are lower than the recommended value ( < 300 mOsm).
Perhaps of equal importance to the preservation and viability of the midgut tissue during the voltage-clamp experiment, is the high amino acid content of the Chamberlin buffer relative to the other chamber buffers.
Parenti et al. (1985) showed that the midgut uses amino acids, particularly those involved in metabolism directly related to the tricarboxylic acid cycle, as the primary source of energy for the potassium pump activity in the B. mori midgut.
Maddrell et al. (1988) reported the effects of the hemolytic toxin, CytA, from B. thuringiensis var.
israelensis on fluid secretion of Malpighian tubules of the insect Rhodnius prolixus. As with our results, they
observed an initial lag time followed by a rapid reduction of the assayed phenomenon (in their case, rate of fluid secretion by the tubules). They explored the cumulative
effects of serially-added subtoxic amounts of CytA toxin and compared, using Poisson statistics, the rate of formation of active complexes to the theoretical number of “hits” (toxin molecules) required to give the same rate (degree of intoxication per time). They calculated that 12 molecules must attach to a small given area of membrane to form an active complex. There are several reasons why we have not attempted to apply such statistical approaches to our system. The most obvious is the differences between the two toxins. CytA is 27 kDa,
has no amino acid homology, and is structurally quite different from the 65 kDa CryIAa toxin used in our study
(Hofte and Whiteley, 1989). The nature of the responses of the two toxins to the two tissues is also different. In their system, the lag time (time before onset of secretion failure) varied with toxin concentration, whereas in our work, we observe that the slope of the inhibition of I,, varied with toxin concentration. Furthermore, we did not observe the cumulative dose effect described by Maddrell et al. (1988). When we apply several subtoxic doses of CryIAa toxin to
B. mori membranes, we observe very shallow slopes approximating the smallest amounts of applied toxin
(E. Alcantara, personal observation). This does not necessarily indicate one-hit kinetics. However, since we have previously observed that CryIA toxins exist in solution as oligomers of molecular weight greater than 500 kDa (Becktel et al., 1992) CryIA toxins may initially bind as oligomers rather than, or in addition to, accumulating after binding as proposed for CytA by Maddrell et al. (1988).
REFERENCES
Becktel W. J., Qi F., Wu S.-J., Lee M. and Dean D. H. (1992)
Conformational analysis of &endotoxins of Bacillus thuringiensis.
Biophys. J. 61, a474. (Abstract.)
Chamberlin M. E. (1990) Luminal alkalinization by the isolated midgut
of the tobacco homworm (Manduca sexta). J. exp. Biol. 150,
467471.
Crawford D. N. and Harvey W. R. (1988) Barium and calcium block
Bacillus thuringiensis subspecies kurstaki d-endotoxin inhibition of
potassium current across isolated midgut of larval Manduca sexta.
J. exp. Biol. 137, 277-286. Dow J. A. T. (1986) Insect midgut function. Adv. Insect Physiol. 19,
187-328. Dow J. A. T., Harvey W. R., Wolfersberger M. G. and Boyes B. (1985)
An improved chamber for the short-circuiting of epithelia. J. exp.
Biol. 114, 685-689.
English L. H., Readdy T. L. and Bastian A. E. (1991).
Delta-endotoxin-induced leakage of 86Rb+-K+ and HrO from
phospholipid vesicles is catalyzed by reconstituted midgut
membrane. Insect Biochem. 21, 177-184. Ge A. Z., Pfister R. M. and Dean D. H. (1990) Hyperexpression of a
Bacillus thuringiensis delta-endotoxin-encoding gene in Escherichia coli: properties of the product. Gene 93, 49-54.
Ge A. Z., Shivarova N. I. and Dean D. H. (1989) Location of the
Bombyx mori specificity domain on a Bacillus thuringiensis 6-endotoxin protein. Proc. natn. Acad. Sci. U.S.A. 86, 40374041.
Giordana B. and Sacchi F. (1977) Some ionic and electrical parameters
of the intestinal epithelium in three mature larvae of lepidoptera.
Comp. Biochem. Physiol. 56A, 95-99. Giordana B., Sacchi V. F. and Hanozet G. M. (1982) Intestinal amino
acid absorption in lepidopteran larvae. Biochim. biophys. Acta 692, 8 l-88.
Giordana B., Tasca M., Villa M., Chiantore C., Hanozet G. M. and
Parenti P. (1993) Bacillus thuringiensis subsp. aizawai 6-endotoxin
inhibits the K+/amino acid cotransporters of lepidopteran larval
midgut. Camp. Biochem. Physiol. 106C, 403407. Harvey W. R. and Wolfersberger M. G. (1979) Mechanism ofinhibition
of active potassium transport in isolated midgut of Manduca sexta
by Bacillus thuringiensis endotoxin. J. exp. Biol. 83, 293-304. Harvey W. R. and Zerahn K. (1972) Active transport of potassium and
other alkali metals by the isolated midgut of the silkworm. Curr.
Topics membr. Tramp. 3, 3688392.
Harvey W. R., Crawford D. N. and Spaeth D. D. (1990) Isolation,
voltage clamping and flux measurements in lepidopteran midgut.
Meth. Enzym. 192, 599608. Hiifte H. and Whiteley H. R. (1989) Insecticidal crystal proteins of
Bacillus thuringiensis. Microbial. Rev. 53, 242-255. Knight P. J. K., Crickmore N. and Ellar D. J. (1994). The receptor for
Bacillus thuringiensis CryIA(c) delta-endotoxin in the brush border
membrane of the lepidoperan Manduca sexta is aminopeptidase N. Molec. Microbial. 11, 4299436.
Knowles B. H., Blatt M. R., Tester M., Horsnell J. M., Carroll J.,
Menestrina G. and Ellar D. J. (1989) A cytolytic &endotoxin from
Bacillus thuringiensis var. israelensis forms cation-selective channels
in planar lipid bilayers. PEBS Lett. 244, 259-262. Laemmh U. K. and Favre R. (1973) Maturation of the head of
bacteriophage T,-I.DNA packaging events. J. molec. Biol. 80, 575-599.
Lee M. K., Milne R. E., Ge A. Z. and Dean D. H. (1992) Location of
a Bombyx mori receptor binding region on a Bacillus thuringiensis &endotoxin. J. biol. Chem. 267, 31153121.
Maddrell S. H. P., Lane N. J., Harrison J. B., Overton J. A. and Moreton R. B. (1988) The initial stages in the action of an insecticidal
6-endotoxin of Bacillus thuringiensis var. israelensis on the epithehal
cells of the malpighian tubules of the insect, Rhodniusprolixus. J. cell Sci. 90, 131-144.
Nayar J. K. and Fraenkel G. (1963) Practical methods of year-round
laboratory rearing of the silkworm, Bombyx mori (L.) (Lepidoptera,
Bombycidae). Ann. Ent. Sot. Am. 56, 122-123. Parenti P., Giordana B., Sacchi V. F., Hanozet G. M. and Guerritore

22 BAERBEL LIEBIG et al.
A. (1985) Metabolic activity related to the potassium pump in the midgut of Bombyx mori larvae. J. exp. Biol. 116, 69978.
Reuveni M. and Dunn P. E. (1991) Differential inhibition by Bacillus
thuringiensis 6-endotoxin of leucine and aspartic acid uptake into
BBMV from midgut of Manduca sexta. Biochem. biophys. Res.
Commun. 181, 1089-1093.
Rouschal W. (1940) Osmotische werte wirbelloser landtiere und ihre
oekologische bedeutung. Z. wiss. Zool. 153, 196.
Sangadala S., Walters F. S., English L. H. and Adang M. J. (1994).
A mixture of Manduca sexta aminopeptidase and phosphatase
enhances Bacillus thuringiensis insecticidal cryIA(c) toxin binding
and s6Rb+-K+ efflux in vitro. J. biol. Chem. 269, 10088-10092.
Schwartz J.-L., Garneau L., Masson L. and Brousseau R. (1991) Early
response of cultured lepidopteran cells to exposure to F-endotoxins
from Bacillus thuringiensis: involvement of calcium and anionic
channels. Biochim. biophys. Acta 1065, 25&260.
Schwartz J.-L., Garneau L., Savaria D., Masson L., Brousseau R. and
Rousseau E. (1993) Lepidopteran-specific crystal toxins from
Bacillus thuringiensis form cation- and anion-selective channels in
planar lipid bilayers. J. membr. Biol. 132, 53-62.
Slatin S. L., Abrams C. and English L. (1990) Delta-endotoxin form
cation-selective channels in planar lipid bilayers. Biochem. biophys.
Res. Commun. 169, 765-172.
Wieczorek H., Putzenlechner M., Zeiske W. and Klein U. (1991)
A vacuolar-type proton pump energizes K+/H+ antiport in an
animal plasma membrane. J. biol. Chem. 266, 1534G-15347.
Wolfersberger M. G. (1992) V-ATPase-energized epithelia and
biological insect control. J. exp. Biol. 172, 377-386.
Wolfersberger M. G. and Spaeth D. D. (1987) Activity of spore-crystal
preparations from twenty serotypes of Bacillus thuringiensis toward
Manduca sexta larvae in vivo. J. appl. Ent. 103, 138-141.
Wolfersberger M. G., Liithy P., Maurer A., Parenti P., Sacchi F. V.,
Giordana B. and Hanozet G. M. (1987) Interaction of Bacillus
thuringiensis delta-entotoxin with membrane vesicles isolated from
lepidopteran larval midgut Comp. Biochem. Physiol. 86A, 301-308.
Acknowledgements-This work was supported by grant AI 29092 from
the National Institutes of Health and a grant from the USDA Forest
Service, Northeastern Forest Experiment Station, to DHD. We thank
Xuejun Chen for reproducing Fig. 1, and Mike Wolfersberger and Mary
Chamberlin for their comments on the manuscript.



![Analysis of gene expression in the midgut of Bombyx mori … · insect communication, reproduction, and pathogen ex-clusion [2, 6]. Thus, commensal bacteria in the midgut are important](https://img.pdfslide.us/doc/110x75/5acc86b07f8b9a63398cf216/analysis-of-gene-expression-in-the-midgut-of-bombyx-mori-communication-reproduction.jpg)

![RNAi Implications in Entomological Research and Pest Control · T. castaneum Chitin synthases CHS1 and CHS2 are crucialexoskeleton and the midgut peritrophic matrix [28] Bombyx mori](https://img.pdfslide.us/doc/110x75/60c6f3bbff9aed657710bff8/rnai-implications-in-entomological-research-and-pest-control-t-castaneum-chitin.jpg)























