Embed Size (px)
Citation preview

Purification of Surfactant from Lung Washings and Washings Contaminated with Blood Constituents SUE A. SHELLEY, JUNE E. PACIGA, and JOHN U. BALLS, Department of Pathology, Loyola University Stritch School of Medicine, Maywood, I L 60153, and V.A.Hospital, Hines, IL 60141
ABSTRACT
A rapid and simple method capable of purifying surfactant from rabbit alveolar washings and from washings contami- nated with serum has been developed. The sample, containing 16% NaBr, is placed beneath a two-layer discontinuous gradient of NaBr. After centrifugation, the surfactant is found near the top of the gradient tube at a density of 1.085 at 4 C while the contaminating material remains near the bottom. The lipid com- position of surfactant from lung washings of n o r m a l animals isolated by this method compares quite favorably with s u r f a c t a n t i so la ted by much more elaborate and time consuming methods. Surfactant purified from mixtures of 3H-palmitate labeled rabbit serum and lung washings (1.6 mg serum phospho- lipid:l mg washing phospholipid) con- tained less than 3% of the phospholipid radioactivity. The phospholipid composi- tion of this band was quite similar to that of surfactant from normal lung washings, but the protein content was much higher. A second density gradient centrifugation removed 90% of this protein resulting in a surfactant fraction with a phospholipid to protein ratio similar to that of surfactant f rom n o r m a l l u n g washings. These findings demonstrate that this purifica- tion method is capable of removing a large proportion of both Serum phospho- lipids and proteins from lung washings contaminated with serum, making this
method uniquely suitable for evaluation of surfactant in pathologic conditions of the lung.
INTRODUCTION
A major problem in the study of lung sur- factant, particularly in pathologic conditions, is the lack of appropriate methods to separate the surfactant from blood components transudated into the air spaces. Current methods for the isolation and purification of lung surfactant (1-7) are not applicable to contaminated material because early in the procedure dif- ferential centrifugation is used. Recently we have reported that sedimentation results in the entrapment of protein within aggregates of sur- factant liposomes, with the amount of protein in the sediment being directly dependent on the amount of protein in the original sample (8). These aggregates are not readily dispersed; therefore, density gradient centrifugation of the entire noncellular washing appears to be a better procedure than purifying the sedimented surfactant fraction on density gradients.
This paper describes a relatively simple method of density gradient centrifugation for purifying lung surfactant from both clean lung washings and contaminated samples. It differs from other methods in that no sedimentation step is used and also that the sample is placed beneath the gradient instead of being spread throughout the tube or on top of the gradient.
EXPERIMENTAL PROCEDURES
Lung washings were obtained from New
TABLE I
Density Gradients for Isolation of Lung Surfactant
Rotor and tube (Spinco) SW25.2 SW27.1 SW50L Dimensions of tube 1�88 x 3~A in. 5/8 x 4 in. �89 x 2 in. Total volume of gradient 60 ml 17 ml 5 ml Gradients layers
No NaBr 8 ml 4 ml 1 ml 13% NaBr 32 ml 8 ml 3 ml Sample in 16% NaBr 20 ml 5 ml t ml
Centrifugaiotn g max 90,000 116,000 98,000 t ime 3 hr 2 hr 1 hr
505
506 SUE A. SHELLEY, JUNE E. PACIGA, AND JOHN U. BALIS
1800
1600
1400
~1000
!: "~6~
4011
2 ~
-- : Phospholipid o - - o Protein ~- -~ Density
1.14
1.12
,
tl.O8 .06
1.04
Bottom 0 3 0 40 Top5o~ Tube
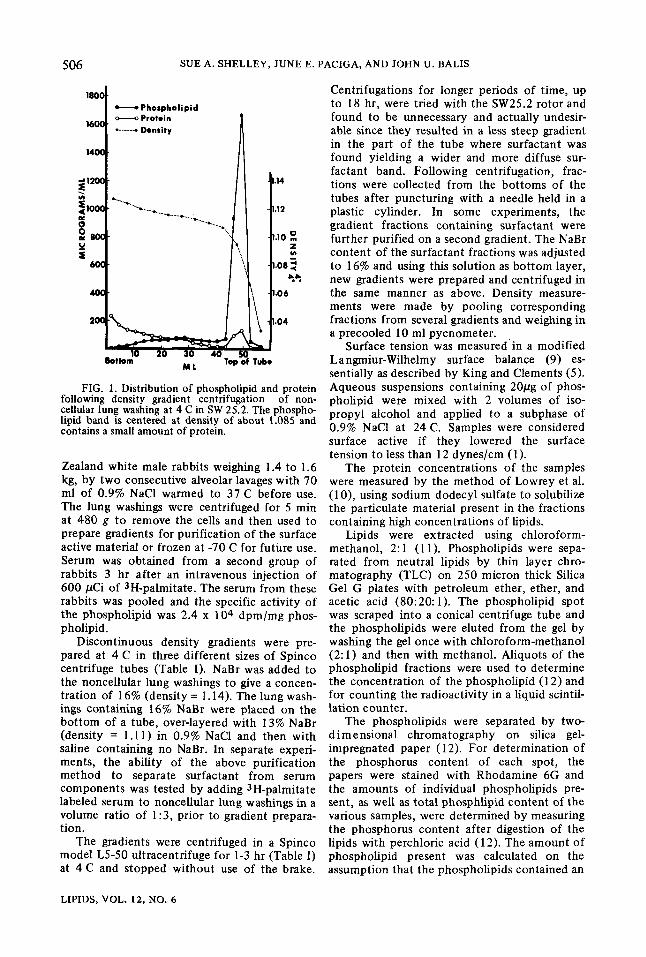
FIG. 1. Distribution of phospholipid and protein following density gradient centrifugation of non- cellular lung washing at 4 C in SW 25.2. The phospho- lipid band is centered at density of about 1.085 and contains a small amount of protein.
Zealand white male rabbits weighing 1.4 to 1.6 kg, by two consecutive alveolar lavages with 70 ml of 0.9% NaCI warmed to 37 C before use. The lung washings were centrifuged for 5 min at 480 g to remove the cells and then used to prepare gradients for purification of the surface active material or frozen at -70 C for future use. Serum was obtained from a second group of rabbits 3 hr after an intravenous injection of 600/uCi of 3H-palmitate. The serum from these rabbits was pooled and the specific activity of the phospholipid was 2.4 x 104 dpm/mg phos- pholipid.
Discontinuous density gradients were pre- pared at 4 C in three different sizes of Spinco centrifuge tubes (Table I). NaBr was added to the noncellular lung washings to give a concen- tration of 16% (density = 1.14). The lung wash- ings containing 16% NaBr were placed on the bottom of a tube, over-layered with 13% NaBr (density = 1.11) in 0.9% NaC1 and then with saline containing no NaBr. In separate experi- ments, the ability of the above purification method to separate surfactant from serum components was tested by adding 3H-palmitate labeled serum to nonceUular lung washings in a volume ratio of 1:3, prior to gradient prepara- tion.
The gradients were centrifuged in a Spinco model L5-50 ultracentrifuge for 1-3 hr (Table I) at 4 C and stopped without use of the brake.
Centrifugations for longer periods of time, up to 18 hr, were tried with the SW25.2 rotor and found to be unnecessary and actually undesir- able since they resulted in a less steep gradient in the part of the tube where surfactant was found yielding a wider and more diffuse sur- factant band. Following centrifugation, frac- tions were collected from the bottoms of the tubes after puncturing with a needle held in a plastic cylinder. In some experiments, the gradient fractions containing surfactant were further purified on a second gradient. The NaBr content of the surfactant fractions was adjusted to 16% and using this solution as bottom layer, new gradients were prepared and centrifuged in the same manner as above. Density measure- ments were made by pooling corresponding fractions from several gradients and weighing in a precooled 10 ml pycnometer.
Surface tension was measured in a modified Langrniur-Wilhelmy surface balance (9) es- sentially as described by King and Clements (5). Aqueous suspensions containing 20/~g of phos- pholipid were mixed with 2 volumes of iso- propyl alcohol and applied to a subphase of 0.9% NaC1 at 24 C. Samples were considered surface active if they lowered the surface tension to less than 12 dynes/cm (1).
The protein concentrations of the samples were measured by the method of Lowrey et al. (10), using sodium dodecyl sulfate to solubilize the particulate material present in the fractions containing high concentrations of lipids.
Lipids were extracted using chloroform- methanol, 2:1 (11). Phospholipids were sepa- rated from neutral lipids by thin layer chro- matography (TLC) on 250 micron thick Silica Gel G plates with petroleum ether, ether, and acetic acid (80:20:1). The phospholipid spot was scraped into a conical centrifuge tube and the phospholipids were eluted from the gel by washing the gel once with chloroform-methanol (2:1) and then with methanol. Aliquots of the phospholipid fractions were used to determine the concentration of the phospholipid (12)and for counting the radioactivity in a liquid scintil- lation counter.
The phospholipids were separated by two- d imensional chromatography on silica gel- impregnated paper (12). For determination of the phosphorus content of each spot, the papers were stained with Rhodamine 6G and the amounts of individual phospholipids pre- sent, as well as total phosphlipid content of the various samples, were determined by measuring the phosphorus content after digestion of the lipids with perchloric acid (12). The amount of phospholipid present was calculated on the assumption that the phospholipids contained an
LIPIDS, VOL. 12, NO. 6

PURIFICATION OF LUNG SURFACTANT
TABLE II
Distribution of Phospholipids
507
Percent of total phospholipids a
LPC Sph PC PI PE PG
Noncellular lung washings
Surfactant from lung washings
Serum-lung washing mixture (1:3)
Surfactant from serum-lung washing mixture - 1 gradient
Surfactant from serum-lung washing mixture - 2 gradients
ND b ND 87.4 2.4 2.5 7.7 (1.7)c (0.6) (0.6) (1.5)
ND ND 85.9 2.5 3.1 8.5 (1.1) (0.2) (0.6) (0.7)
11.1 2.9 72.8 5.1 S.0 3.1 (2.2) (1.1) (3.0) (1.7) (0.6) (0.6) 0.8 1.1 81.3 3.7 5.0 8.1
(0.3) (0.6) (1.3) (0.5) (1.2) (0.5) 0.6 0.7 84.7 2.6 2.8 8.6
(0.3) (0.4) (1.2) (0.4) (0.6) (1.4)
aLPC = lysophosphatidyl choline, Sph = sphingomyelin, PC = phosphatidyl choline, PI = phosphatidyl inositol, PE = phosphatidyi ethanolamine, PG = phosphatidyl glycerol.
bND = none detected. CNumbers in parentheses indicate standard error.
average of 4% phosphorus. For determination of the fatty acid composi-
tion of the phosphatidyl choline (PC) lipids were separated by 2-dimensional TLC on Silica Gel G p l a t e s w i t h chloroform-methanol- ammonium hydroxide (140:50:7) in the first d i r ec t ion and chloroform-methanol-acetone- acetic acid-water (100:20:40:20:10) in the second direction. After development, the plates were dried and the lipid spots detected under ultraviolet light after spraying with 0.01% Rhodamine 6G in water. The PC spots were scraped into tubes and the methyl ester deriva- tives of the acyl groups were prepared by alkaline methanolysis (13). The fatty acid methyl esters were separated by gas liquid chromatography using a Varian model 2100 with 2 mm x 6 ft column packed with 10% EGSS-X on 100/120 mesh Gas Chrom P. The column was standardized using the National Institutes of Health fatty acid methyl ester standards (Applied Science Laboratories, State College, PA). The fatty acids were identified by comparison of the relative retention times with standards and by interpolation from a semilog plot of the carbon number of the free acid versus the logarithm of its respective retention time.
RESULTS
Isolation of Surfactant from Lung Washings
Following centrifugation of density gradi- ents prepared with lung washings, a band of white particulate material was readily observed near the top of the gradient tube at a density of 1.085 at 4 C. Fractions collected from these gradient tubes were analyzed for phospholipid
and protein content and the distribution of phosphol ip id and protein from one such gradient is shown in Figure 1. The fractions corresponding to the visible band of material contained 80% (+ 3.5 S.E.) of the phospholipid but only 12% (-+ 1.5) of the protein present in the gradient. Recentrifugation of this surfactant on a second gradient resulted in recovery of more than 95% of both the phospholipid and protein in the surfactant band. Both the non- cellular lung washings used to prepare the gradients and the gradient fraction from the phospholipid-rich band were surface active with the purified surfactant lowering the surface tension to 5 dynes/cm. Other fractions from the density gradient tubes showed no surface activity indicating that the lung surfactant was concentrated in the narrow visible band.
Phospholipid analysis revealed no significant differences between the phospholipid composi- tion of the washings and of the surfactant band (Table II). Both the washings and the surfactant band contained more than 85% PC. A signifi- cant amount of phosphatidyl glycerol (PG) and smaller quantities of phosphatidyl ethanol- amine (PE) and phosphatidyl inositol (PI) were also present. The fatty acid composition of the PC from the lung washings and the surfactant pur i f i ed from the washings differed only slightly (Table III). The surfactant band con- tained slightly more palmitic acid (16:0) and less 18-carbon fatty acids than did the original washings.
Isolation of Surfactant from Serum-Lung Washing Mixtures
In order to determine if this simple method of surfactant purification was capable of sepa-
LIPIDS, VOL. 12, NO. 6
508 SUE A. SHELLEY, JUNE E. PACIGA, AND JOHN U. BALIS
TABLE II1
Major Fatty Acids of Phosphatidyl Choline
Percent of total fatty acids
14:0 16:0 16:1 18:0 18:1 18:2 18:3 20:4
Noncellular lung washings
Surfactant from lung washings
Serum-lung washing mixture (1:3)
Surfactant from serum-lung washing mixture-1 gradient
Surfactant from serum-lung washing mixture-2 gradients
2.2 62.3 5.8 2.1 18.0 8.6 1.0 trace (0.2) a (1.8) (0.3) (0.2) (1.3) (0.7) (0.1) 2.7 67.8 5.6 1.7 14.4 6.9 0.9 trace
(0.5) (1.7) (0.3) (0.2) (1.1) (0.9) (0.1)
1.4 37.4 2.7 11.5 19.3 23.6 2.1 2.0 (0.6) (1.1) (0.2) (0.9) (0.7) (0.2) (0.2) (0.4) 1.5 60.2 5.2 3.2 1'7.9 10.6 I.I 0.3
(0.3) (0.2) (0.3) (0.1) (0.3) (0.5) (0.1) (0.1) 1.4 60.9 4.4 2.7 19.5 9.9 1.2 trace
(0.3) (0.3) (0.2) (0.2) (0.3) (0.2) (0.1)
aNumbers in parentheses indicate standard error
1 2 0 0 0
!0000 .,, i
8000
6oo9
400
300 U
- -- Phos pholipid I ~-, c- ~ Protein
I \ ..... c ' / I \ V~
\
1 \ \ A t,,00 t
1 $ lO 15
Bottom Top of Tube ML
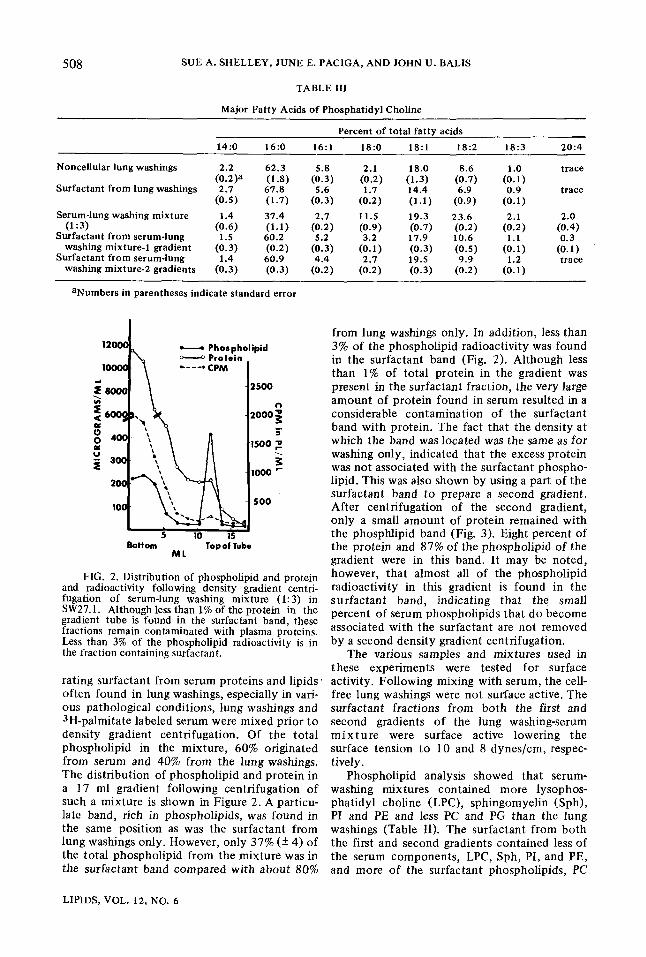
FIG. 2. Distribution of phospholipid and protein and radioactivity following density gradient centri- fugation of serum-lung washing mixture (1:3) in SW27.1. Although less than 1% of the protein in the gradient tube is found in the surfactant band, these fractions remain contaminated with plasma proteins. Less than 3% of the phospholipid radioactivity is in the fraction containing surfactant.
rating surfactant from serum proteins and lipids' often found in lung washings, especially in vari- ous pathological conditions, lung washings and 3H-palmitate labeled serum were mixed prior to density gradient centrifugation. Of the total phospholipid in the mixture, 60% originated from serum and 40% from the lung washings. The distribution of phospholipid and protein in a 17 ml gradient following centrifugation of such a mixture is shown in Figure 2. A particu- late band, rich in phospholipids, was found in the same position as was the surfactant from lung washings only. However, only 37% (+ 4) of the total phospholipid from the mixture was in the surfactant band compared with about 80%
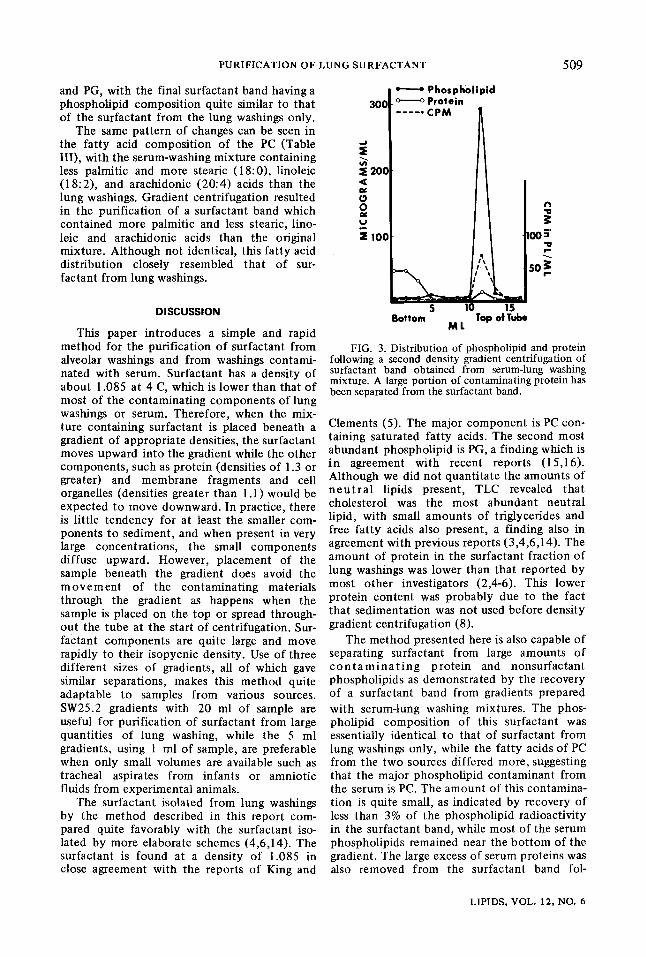
from lung washings only. In addition, less than 3% of the phospholipid radioactivity was found in the surfactant band (Fig. 2). Although less than 1% of total protein in the gradient was present in the surfactant fraction, the very large amount of protein found in serum resulted in a considerable contamination of the surfactant band with protein. The fact that the density at which the band was located was the same as for washing only, indicated that the excess protein was not associated with the surfactant phospho- lipid. This was also shown by using a part of the surfactant band to prepare a second gradient. After centrifugation of the second gradient, only a small amount of protein remained with the phosphlipid band (Fig. 3). Eight percent of the protein and 87% of the phospholipid of the gradient were in this band. It may be noted, however, that almost all of the phospholipid radioactivity in this gradient is found in the surfactant band, indicating that the small percent of serum phospholipids that do become associated with the surfactant are not removed by a second density gradient centrifugation.
The various samples and mixtures used in these experiments were tested for surface activity. Following mixing with serum, the cell- free lung washings were not surface active. The surfactant fractions from both the first and second gradients of the lung washing-serum m i x t u r e were surface active lowering the surface tension to 10 and 8 dynes/cm, respec- tively.
Phospholipid analysis showed that serum- washing mixtures contained more lysophos- phatidyl choline (LPC), sphingomyelin (Sph), PI and PE and less PC and PG than the lung washings (Table 1I). The surfactant from both the first and second gradients contained less of the serum components, LPC, Sph, PI, and PE, and more of the surfactant phospholipids, PC
LIPIDS, VOL. 12, NO. 6
PURIFICATION OF LUNG SURFACTANT 509
and PG, with the final surfactant band having a phospholipid composition quite similar to that of the surfactant from the lung washings only.
The same pattern of changes can be seen in the fatty acid composition of the PC (Table III), with the serum-washing mixture containing less palmitic and more stearic (18:0), linoleic (18:2), and arachidonic (20:4) acids than the lung washings. Gradient centrifugation resulted in the purification of a surfactant band which contained more palmitic and less stearic, lino- leic and arachidonic acids than the original mixture. Although not identical, this fatty acid distribution closely resembled that of sur- factant from lung washings.
DISCUSSION
This paper introduces a simple and rapid method for the purification of surfactant from alveolar washings and from washings contami- nated with serum. Surfactant has a density of about 1.085 at 4 C, which is lower than that of most of the contaminating components of lung washings or serum. Therefore, when the mix- ture containing surfactant is placed beneath a gradient of appropriate densities, the surfactant moves upward into the gradient while the other components, such as protein (densities of 1.3 or greater) and membrane fragments and cell organelles (densities greater than 1.1) would be expected to move downward. In practice, there is little tendency for at least the smaller com- ponents to sediment, and when present in very large concentrations, the small components diffuse upward. However, placement of the sample beneath the gradient does avoid the m o v e m e n t of the contaminating materials through the gradient as happens when the sample is placed on the top or spread through- out the tube at the start of centrifugation. Sur- factant Gomponents are quite large and move rapidly to their isopycnic density. Use of three different sizes of gradients, all of which gave similar separations, makes this method quite adaptable to samples from various sources. SW25.2 gradients with 20 ml of sample are useful for purification of surfactant from large quantities of lung washing, while the 5 ml gradients, using 1 ml of sample, are preferable when only small volumes are available such as tracheal aspirates from infants or amniotic fluids from experimental animals.
The surfactant isolated from lung washings by the method described in this report com- pared quite favorably with the surfactant iso- lated by more elaborate schemes (4,6,14). The surfactant is found at a density of 1.085 in close agreement with the reports of King and
-" : Phospholipid
i,ool
Ill i !oo oo~
5 10 15 Bottom Top of Tube
ML
FIG. 3. Distribution of phospholipid and protein following a second density gradient centrifugation of surfactant band obtained from serum-lung washing mixture. A large portion of contaminating protein has been separated from the surfactant band.
Clements (5). The major component is PC con- taining saturated fatty acids. The second most abundant phospholipid is PG, a finding which is in agreement with recent reports (15,16). Although we did not quantitate the amounts of n e u t r a l lipids present, TLC revealed that cholesterol was the most abundant neutral lipid, with small amounts of triglycerides and free fatty acids also present, a finding also in agreement with previous reports (3,4,6,14). The amount of protein in the surfactant fraction of lung washings was lower than that reported by most other investigators (2,4-6). This lower protein content was probably due to the fact that sedimentation was not used before density gradient centrifugation (8).
The method presented here is also capable of separating surfactant from large amounts of c o n t a m i n a t i n g protein and nonsurfactant phospholipids as demonstrated by the recovery of a surfactant band from gradients prepared with serum-lung washing mixtures. The phos- pholipid composition of this surfactant was essentially identical to that of surfactant from lung washings only, while the fatty acids of PC from the two sources differed more, suggesting that the major phospholipid contaminant from the serum is PC. The amount of this contamina- tion is quite small, as indicated by recovery of less than 3% of the phospholipid radioactivity in the surfactant band, while most of the serum phospholipids remained near the bottom of the gradient. The large excess of serum proteins was also removed from the surfactant band fol-
LIPIDS, VOL. 12, NO. 6

510 SUE A. SHELLEY, JUNE E. PACIGA, AND JOHN U. BALIS
lowing two centr i fugat ions . Current ly used m e t h o d s for surfactant puri-
fication generally involve several successive pe r i ods of cent r i fugat ion and ra ther large amoun t s o f lung tissue or washings as s tar t ing material and , therefore , are not applicable to large numbers of samples or to samples where the am oun t of available tissue or washings is l imited. In addi t ion , these m e t h o d s have se ldom been applied to any samples o the r than those from hea l thy animals, presumably because they are not adequa te to separate sur fac tant f rom large amoun t s of con tamina t ing material found in v a r i o u s p a t h o l o g i c a l condi t ions . They likewise have not been widely applied to human material , since samples recovered at au topsy are also con t amina t ed wi th blood. The m e t h o d of purif icat ion in t roduced in this s tudy has been successfully applied to normal and edema tous human lung washings, amni to t i c fluid samples, pharyngeal aspirates f rom newborn infants , and t r a chea l aspirates col lected rout ine ly f rom infants on respirators (1 8). Use of this m e t h o d permits clinical evaluation of matura t ion and pathological changes in the funct ional com- ponen t s of the surfactant system.
ACKNOWLEDGMENT
This work was supported in part by U.S. Public Health Service Grant HD-04434 and USAMRDC Con-
tract DADAI 7-70-C-0041.
REFERENCES
I. Abrams, M.E., J. Appl. Physiol. 21:718 (1966). 2. Klein, R.M., and S. Margolis, Ibid. 25:654 (1968). 3. Steim, J.M., Biochem. Biophys. Res. Commun.
34:434 (1969). 4. Frosolono, M.F., B.L. Charms, R. Pawlowski, and
S. Slivka, J. Lipid Res. 11:439 (19"70). 5. King, R.J., and J.A. Clements, Am. J. Physiol.
223:707 (! 972). 6. Gil, J., and O.K. Reiss, J. Cell Biol. 58:152
(19'73). 7. C o l a c i c c o , G.C., A.R. Buckelew, and E.M.
Scarpelli, J. Appl. Physiol. 34:743 0973). 8. Shelley, S.A., M.V. L'Heureux, and J.U. Balls, J.
Lipid Res. 16:224 (1975). 9. Greenfield, L.J., and G.O. Kimmell, J. Surg. Res.
7:276 (1967). I0. Lowry, O.H., N.J. Rosebrough, A.L. Farr, and
R.J. Randall, J. Biol. Chem. 193:265 (1951). I 1. Folch, J., M. Lees, and G.H. SIoan-Stanley, Ibid.
226:497 (I 957). 12. Wuthier, R.E., J. Lipid Res. 7:544 (1966). 13. Sun, G.Y., and L.A. Horrocks, Lipids 5:1006
(1970). 14. King, R.J., and J.A. Clements, Am. J. Physiol.
223:715 (1972). 15. Pfleger, R.C., R.F. Henderson, and J. Waide,
Chem. Phys. Lipids 9:51 (1972). 16. Rooney, S.A., P.M. Canavan, and E.K. Motoyama,
Biochim. Biophys. Acta 360:56 (1974). 17. Tierney, D.F., and R.P. Johnson, J. Appl. Physiol
20:1253 (1965). 18. Balls, J.U., M. Kovacevic, J.E. Paciga, and S.A.
Shelley, Pediat, Res. 10:457 (1976).
[Received February 7, 1977]
LIPIDS, VOL. 12, NO. 6





























