Embed Size (px)
Citation preview

HIkan Persson,$ Michel Perricaudet,g Aslihan Tolun,$ Lennart Philipson,+ and Ulf Pettersson$ From the Department of Microbiology, The Biomedical Center, S- 751 2<3 Uppsala, Sweden, and §Recombinaison et Emression GPnPtiaue Institut National de la SantC et de la Recherche Medicale (lJl6.3), Unit& de GEnie GknnPtique, Institut Pasteur, 75015 Pa& France
A simple method for selection of RNA-DNA hybrids has been developed and applied to the purification of adenovirus-specific messenger RNA. Cytoplasmic RNA prepared from adenovirus type 2 (ad2)-infected HeLa cells or from an ad2-transformed rat cell line was hy- bridized in solution to the complementary strands of ad2 DNA. The hybridization mixture was subsequently fractionated by chromatography on a Sepharose 2B column. The intact probe DNA as well as the RNA-DNA hybrids are excluded from the gel matrix and elute with the void volume. Nonhybridized RNA, in contrast, is included into the gel matrix and elutes as a broad peak well separated from the excluded fractions. Fractions corresponding to the void volume, were collected and the RNA-DNA hybrids were denatured in 90% form- amide. The selected RNA was separated from the DNA by affinity chromatography on poly(U)-Sepharose. Re- striction endonuclease fragments of DNA with a large enough size to make them excluded from the agarose column were also used for hybridization. In these ex- periments hybridizations were carried out under con- ditions which would allow R-loop formation (Thomas, M., White, R. L., and Davis, R. W. (1976) Proc. N&Z. Acad. Sci. U. S. A. 73, 2294-2298) and the hybridized RNA was separated from unhybridized RNA by Seph- arose chromatography. The validity of the method was demonstrated by programming an in vitro protein-syn- thesizing system with selected RNA.
recovery from the phase system. Simian virus 40 (SV40) DNA, both double- and single-stranded, has been covalently linked to a solid support and used for the selection of SV40 mRNA. The coupling of DNA to a solid support is time-consuming and usually requires large quantities of DNA (5). All the described systems are capable of selecting messenger RNA but an unspecific background hybridization makes them less useful for mRNA species which are present in small quantities.
Exclusion chromatography on agarose permits the fraction- ation of high molecular weight nucleic acids and thus provides a simple alternative method to achieve separation between RNA-DNA hybrids and nonhybridized RNA. In the present communication we describe the purification of RNA-DNA hybrids by exclusion chromatography on agarose (Sepharose 2B).
EXPERIMENTAL PROCEDURES
Cell and Virus-Ad2 was propagated in suspension cultures of HeLa S3 cells in Eagle’s spinner medium supplemented with 7% calf serum as described before (6). Suspension cultures of one ad2-trans- formed rat cell line (A,T,C,, see Ref. 7) were cultivated in Eagle’s spinner medium supplemented with 10% calf serum.
Methods to separate RNA-DNA hybrids from unhybridized RNA are of great importance for mapping of genes for differ- ent polypeptides on viral or bacterial genomes. Programming an in vitro protein-synthesizing system with mRNA that has been selected by hybridization makes it possible to identify the gene product which is specified by a given DNA sequence. Several methods designed for this purpose have previously been described; RNA hybridized to denaturated double- stranded DNA can be isolated from unhybridized RNA by hydroxylapatite chromatography in the presence of urea as described by Smith et al. (1). This method was used by Lewis et al. (2,3) to isolate adenovirus-specific mRNA hybridized to defined restriction endonuclease fragments of ad2 DNA. Mak et al. (4) described the purification of ad2 mRNA by an aqueous polymer two-phase system, a method which, how- ever, requires large amounts of viral RNA due to a poor
Preparation of Restriction h’ndonuclease Fragments and Com- plementary Strands of ad2 DNA-Restriction endonuclease frag- ments of ad2 DNA were prepared as previously described (6). Before use, the purity of the fragments was documented by agarose gel electrophoresis. Intact viral l- or r-strands were prepared by equilib- rium centrifugation in CsCl of denaturated ad2 DNA in the presence or ribopoly(lJ,G) as described by Tibbetts et al. (8). The viral r-strand refers to the strand of ad2 DNA which is transcribed in the rightward direction. The leftward transcribed strand of ad2 DNA is referred to as the l-strand (see Ref. 9 concerning the nomenclature of the com- plementary strands from ad2 DNA).
Extraction of RNA-HeLa cells were infected with purified ad2 at an input multiplicity of 20,000 virus particles/cell. Cycloheximide (25 pg/ml) was added at 2 h postinfection, the cells harvested at 7 h postinfection, and “early” cytoplasmic RNA was prepared. Late-ad- enovirus cytoplasmic RNA was prepared from cells harvested 18 h postinfection with an input multiplicity of 2000 virus particles/cell. RNA was in both cases extracted by the method of Brawerman et al. (19) and precipitated twice with ethanol.
* This work was supported by grants from the Swedish Cancer Society and the Swedish Medical Research Council. Part of this work was aided by a European Molecular Riology Organization short term fellowship to M. I’. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “ad~~ertisement” in accordance with 18 1J.S.C. Section 1734 solely to indicate this fact.
Liquid Hybridization-The RNA was melted in 80% formamide at 65°C for 15 min immediately before hybridization. The formamide concentration was adjusted to 50% and the RNA hybridized to sepa- rated strands of ad2 DNA in a buffer which contained: 0.4 M NaCI, O.l’%, sodium dodecyl sulfate, 1 mM ethylendiaminetetraacetic acid (EDTA) and 0.02 M piperazine-N,N’-bis(2-ethanesulfonic acid, pH 6.4. Usually 1 to 3 1.18 of DNA was hybridized with 200 to 500 pg of cytoplasmic RNA in a total volume of 1 ml at 5O’C for 16 h. Restriction endonuclease fragments of ad2 DNA (1 to 2 pg) were denatured in 0.3 M sodium hydroxide for 10 min at room temperature, neutralized, and hybridized to a cytoplasmic RNA as described above except that the formamide concentration was increased to 80%. Form- amide for the hybridization experiments was purchased from Merck, Darmstadt, West Germany, and purified as described previously (11).
Sepharose 2B Chromatography-A column (0.9 X 20 cm) of Seph- arose 2B (Pharmacia Fine Chemicals) was prepared in a buffer which
Purification of RNA-DNA Hybrids by Exclusion Chromatography*
(Received for publication, March 2, 1979)
7999
by guest on June 17, 2020http://w
ww
.jbc.org/D
ownloaded from
8000 Selection of Adenovirus mRNA
contained 0.5 M LiCI, 0.5%) SDS,’ 1 mM EDTA, and 0.05 M Tris-HCI, pH 7.9. The hybridization mixture was applied to the column and fractions of 0.5 ml were collected at 5-min intervals. The void volume of the column was determined by including a small amount of ,‘lP- labeled viral DNA (12) in the sample before fractionation. Alterna- tively the optical density at 260 nm of each fraction was measured. Fractions corresponding to the void volume (usually Fractions 7 to 9) were pooled, purified Escherichia coli 16 S ribosomal RNA (25 pg) was added, and the samples precipitated with 2 volumes of cold ethanol overnight at -2O’C. The precipitates were collected by cen- trifugation and dissolved in 0.5 ml of 90% FA buffer (90% deionized formamide, 2 mM EDTA, 0.5 M NaCl, 0.1% SDS and 0.05 M Tris-HCl, pH 7.9). The RNA-DNA hybrids (in 90% FA buffer) were denaturated by incubation for 15 min at 65°C. The formamide concentration was decreased to 25% by addition of NaCl to 0.5 M, 2 mru EDTA, 0.1% SDS and 0.05 M Tris-HCl, pH 7.9, and the sample immediately applied to a poly(U)-Sepharose column as described below.
Poly(U)-Sepharose Chromatography-One-milliliter columns of poly(U)-Sepharose (Pharmacia Fine Chemicals) were prepared and washed with a buffer which contained 25% deionized formamide, 2 mM EDTA, 0.5 M NaCl, 0.1% SDS and 0.05 M Tris-HCl, pH 7.9 (25% FA buffer). Samples in 25% FA buffer were applied to the columns which subsequently were washed with 4 column volumes of 25% FA buffer. Poly(A)-containing RNA was eluted with 4 ml of the 90% FA buffer. Purified E. coli 16 S rRNA (25 pg) and potassium acetate (pH 6.0) (KAc) (final concentration 0.15 M) were added to the eluted samples which were precipitated with 2 volumes of cold ethanol over night at -20°C. The precipitates were collected by centrifugation, dissolved in 0.5 ml of 0.15 M KAc, and reprecipitated with 2 volumes of cold ethanol. The precipitates were again collected by centrifuga- tion and dissolved in the appropriate volume of sterile distilled water (usually 15 ~1).
The poly(U)-Sepharose chromatography step was in some cases omitted when restriction endonuclease fragments were used as probes for the hybridization. In such cases the precipitated fractions from the void volume were divided into two equal aliquots, one was melted as described above, followed by ethanol precipitation, while the other was directly precipitated with ethanol. Both samples were reprecipi- tated with ethanol, dissolved in 15 ~1 of distilled water and 2 to 3 ~1 of each sample was translated in vitro as described below.
In Vitro Translation-Micrococcal nuclease-treated rabbit retic- ulocyte lysates were prepared as described by Pelham and Jackson (13). Unfractionated early- or late-ad2 cytoplasmic RNA was added at a final concentration of 160 to 320 pg/ml and the RNA translated in the nuclease-treated lysate as described previously (14). Cytoplas- mic RNA selected by hybridization to 1 to 3 pg of complementary strand-specific ad2 DNA was usually dissolved in 15 ~1 of sterile water. Two or three microliters of the RNA were added per reaction volume (25 ~1). After incubation for 2 h at 35°C the translation mixtures were treated with pancreatic RNase and sodium hydroxide as previously described (14). An equal volume of 2~ gel sample buffer (15) was added and the samples were analyzed by SDS-polyacrylamide gel electrophoresis as described below.
Immunoprecipitation-Samples from the in vitro translation mix- tures were immunoprecipitated with an antiserum raised in syngeneic rats against an ad2-transformed rat cell line (AzT&,) (16) or with a control serum from nonimmunized rats as described previously (14). The immunoprecipitates were analyzed by SDS-polyacrylamide gel electrophoresis.
SDS-Polyacrylamide Gel Electrophoresis-Samples were ana- lyzed in 1.5.mm-thick and 140.mm-long 13% polyacrylamide slab gels as described by Maize1 (15). Electrophoresis was for 10 h at 100 V and the gels were subsequently analyzed by fluorography (17).
RESULTS AND DISCUSSION
Selection of RNA-DNA Hybrids by Exclusion Chromatog- raphy--Several exclusion chromatography matrices are pres- ently available for fractionation of large macromolecules, such as nucleic acids. Among those we decided to investigate whether 2% agarose (Sepharose 2B) would be useful for sep- aration of RNA-DNA hybrids from unhybridized RNA. Com- plementary strands of the ad2 genome (11.5 X 10” daltons) labeled with [“Hlthymidine were fist fractionated on a Seph- arose 2B column in a buffer which contained 0.5 M LiCl,
’ The abbreviation used is: SDS, sodium dodecyl sulfate.
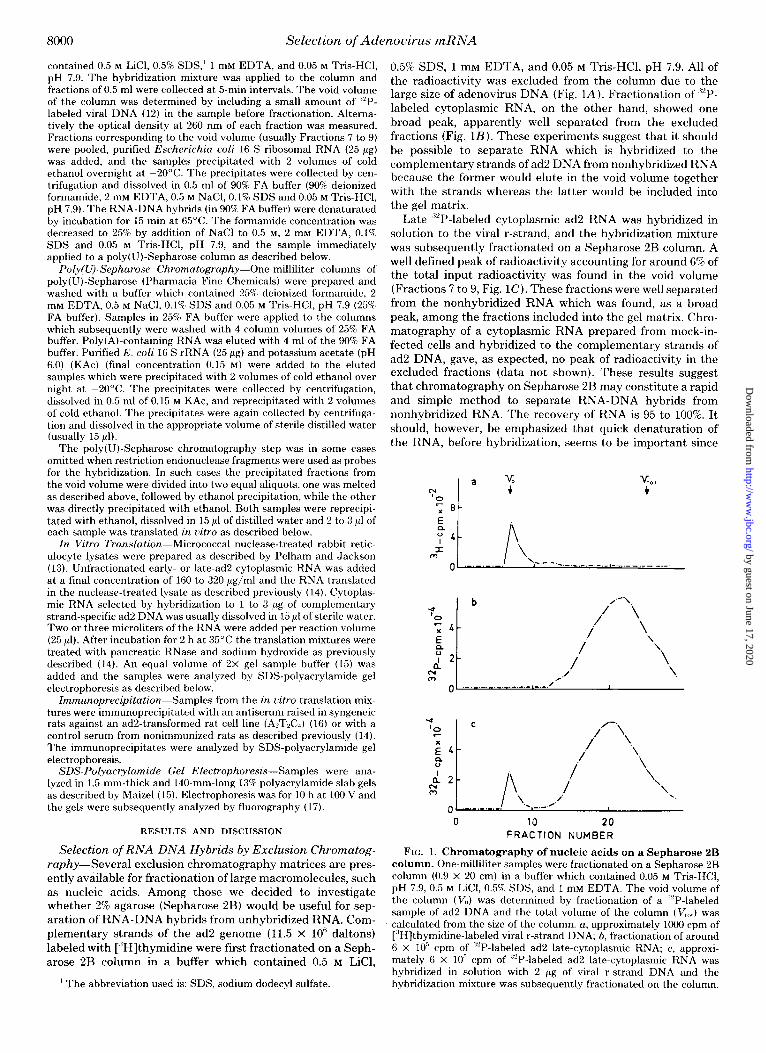
0.5% SDS, 1 mM EDTA, and 0.05 M Tris-HCl, pH 7.9. All of the radioactivity was excluded from the column due to the large size of adenovirus DNA (Fig. L4). Fractionation of “‘P- labeled cytoplasmic RNA, on the other hand, showed one broad peak, apparently well separated from the excluded fractions (Fig. 1B). These experiments suggest that it should be possible to separate RNA which is hybridized to the complementary strands of ad2 DNA from nonhybridized RNA because the former would elute in the void volume together with the strands whereas the latter would be included into the gel matrix.
Late ““P-labeled cytoplasmic ad2 RNA was hybridized in solution to the viral r-strand, and the hybridization mixture was subsequently fractionated on a Sepharose 2B column. A well defined peak of radioactivity accounting for around 6% of the total input radioactivity was found in the void volume (Fractions 7 to 9, Fig. 1C). These fractions were well separated from the nonhybridized RNA which was found, as a broad peak, among the fractions included into the gel matrix. Chro- matography of a cytoplasmic RNA prepared from mock-in- fected cells and hybridized to the complementary strands of ad2 DNA, gave, as expected, no peak of radioactivity in the excluded fractions (data not shown). These results suggest that chromatography on Sepharose 2B may constitute a rapid and simple method to separate RNA-DNA hybrids from nonhybridized RNA. The recovery of RNA is 95 to 100%. It should, however, be emphasized that quick denaturation of the RNA, before hybridization, seems to be important since
0 10 20 FRACTION NUMBER
FIG. 1. Chromatography of nucleic acids on a Sepharose 2B column. One-milliliter samples were fractionated on a Sepharose 2B column (0.9 x 20 cm) in a buffer which contained 0.05 M Tris-HCl, pH 7.9, 0.5 M LiCl, 0.5% SDS, and 1 mM EDTA. The void volume of the column (V,,) was determined by fractionation of a ,‘“P-labeled sample of ad2 DNA and the total volume of the column (V,,,,) was calculated from the size of the column. a, approximately 1000 cpm of [“Hjthymidine-labeled viral r-strand DNA; b, fractionation of around 6 x IO” cpm of ““P-labeled ad2 late-cytoplasmic RNA; c, approxi- mately 6 x lo” cpm of “2P-labeled ad2 late-cytoplasmic RNA was hybridized in solution with 2 pg of viral r-strand DNA and the hybridization mixture was subsequently fractionated on the column.
by guest on June 17, 2020http://w
ww
.jbc.org/D
ownloaded from
Selection of Adenovirus mRNA 8001
Ad2 A B c D E Ad2 i
-E75K - E60K
- ‘-
1 EL2-
_ 50K
- E26K
- -El&5K -E17.5K -E16K -E15K
.-E12.5K
-EllK
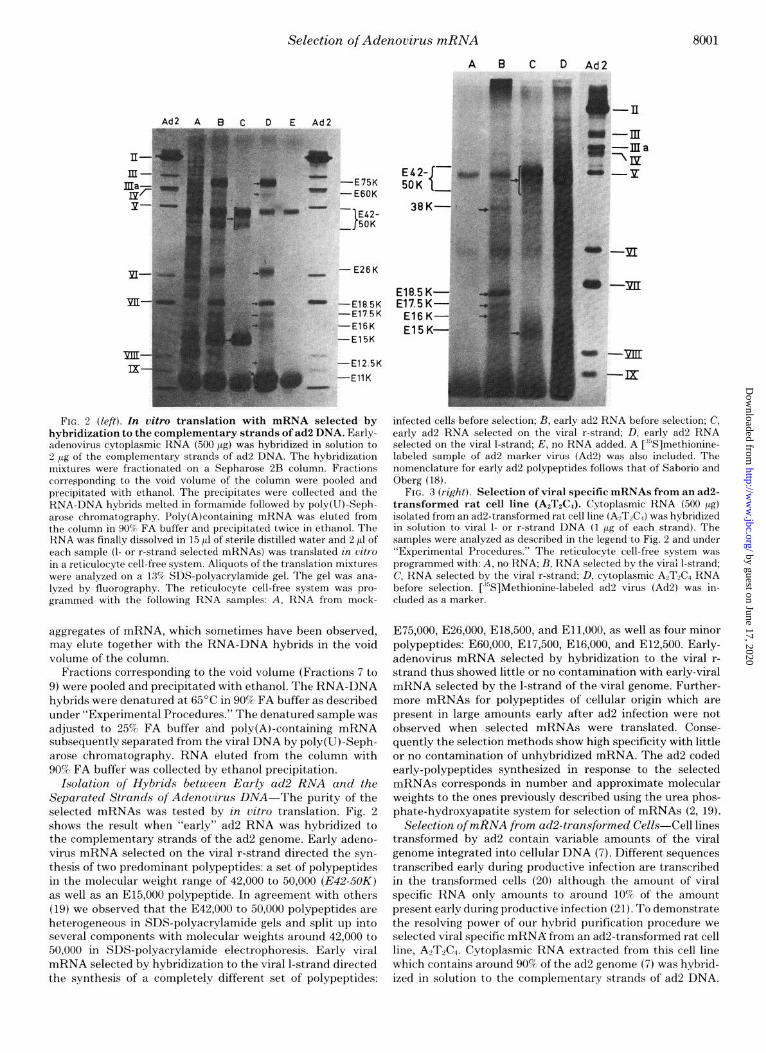
FIG. 2 (left). In vitro translation with mRNA selected by hybridization to the complementary strands of ad2 DNA. Early- adenovirus cytoplasmic RNA (500 pg) was hybridized in solution to 2 pg of the complementary strands of ad2 DNA. The hybridization mixtures were fractionated on a Sepharose 2B column. Fractions corresponding to the void volume of the column were pooled and precipitated with ethanol. The precipitates were collected and the RNA-DNA hybrids melted in formamide followed by poly(U)-Seph- arose chromatography. Poly(A)containing mRNA was eluted from the column in 90% FA buffer and precipitated twice in ethanol. The RNA was finally dissolved in 15 11 of sterile distilled water and 2 ~1 of each sample (I- or r-strand selected mRNAs) was translated in vitro in a reticulocyte cell-free system. Aliquots of the translation mixtures were analyzed on a 13% SDS-polyacrylamide gel. The gel was ana- lyzed by fluorography. The reticulocyte cell-free system was pro- grammed with the following RNA samples: A, RNA from mock-
aggregates of mRNA, which sometimes have been observed, may elute together with the RNA-DNA hybrids in the void volume of the column.
Fractions corresponding to the void volume (Fractions 7 to 9) were pooled and precipitated with ethanol. The RNA-DNA hybrids were denatured at 65°C in 90% FA buffer as described under “Experimental Procedures.” The denatured sample was adjusted to 25% FA buffer and poly(A)-containing mRNA subsequently separated from the viral DNA by poly(U)-Seph- arose chromatography. RNA eluted from the column with 90% FA buffer was collected by ethanol precipitation.
Isolation of Hybrids between Early ad2 RNA and the Separated Strands of Adenouirus DNA-The purity of the selected mRNAs was tested by in vitro translation. Fig. 2 shows the result when “early” ad2 RNA was hybridized to the complementary strands of the ad2 genome. Early adeno- virus mRNA selected on the viral r-strand directed the syn- thesis of two predominant polypeptides: a set of polypeptides in the molecular weight range of 42,000 to 50,000 (E42-50K) as well as an E15,OOO polypeptide. In agreement with others (19) we observed that the E42,OOO to 50,000 polypeptides are heterogeneous in SDS-polyacrylamide gels and split up into several components with molecular weights around 42,000 to 50,000 in SDS-polyacrylamide electrophoresis. Early viral mRNA selected by hybridization to the viral l-strand directed the synthesis of a completely different set of polypeptides:
A B C D Ad2 *
infected cells before selection; B, early ad2 RNA before selection; C, early ad2 RNA selected on the viral r-strand; D, early ad2 RNA selected on the viral l-strand; E, no RNA added. A [ ‘“Slmethionine- labeled sample of ad2 marker virus (Ad2) was also included. The nomenclature for early ad2 polypeptides follows that of Saborio and Oberg (18).
FIG. 3 (r&t). Selection of viral specific mRNAs from an ad2- transformed rat cell line (AzT&). Cytoplasmic RNA (500 pg) isolated from an ad2-transformed rat cell line (ALTLC,) was hybridized in solution to viral l- or r-strand DNA (1 pg of each strand). The samples were analyzed as described in the legend to Fig. 2 and under “Experimental Procedures.” The reticulocyte cell-free system was programmed with: A, no RNA; B, RNA selected by the virai l-strand; C, RNA selected by the viral r-strand; D, cytoplasmic ALTL RNA before selection. [“S]Methionine-labeled ad2 virus (Ad2) was in- cluded as a marker.
E75,000, E26,000, E18,500, and Ell,OOO, as well as four minor polypeptides: E60,000, E17,500, E16,000, and E12,500. Early- adenovirus mRNA selected by hybridization to the viral r- strand thus showed little or no contamination with early-viral mRNA selected by the l-strand of the viral genome. Further- more mRNAs for polypeptides of cellular origin which are present in large amounts early after ad2 infection were not observed when selected mRNAs were translated. Conse- quently the selection methods show high specificity with little or no contamination of unhybridized mRNA. The ad2 coded early-polypeptides synthesized in response to the selected mRNAs corresponds in number and approximate molecular weights to the ones previously described using the urea phos- phate-hydroxyapatite system for selection of mRNAs (2, 19).
Selection of mRNA from ad2-transformed CeZls-Cell lines transformed by ad2 contain variable amounts of the viral genome integrated into cellular DNA (7). Different sequences transcribed early during productive infection are transcribed in the transformed cells (20) although the amount of viral specific RNA only amounts to around 10% of the amount present early during productive infection (21). To demonstrate the resolving power of our hybrid purification procedure we selected viral specific mRNA from an ad2-transformed rat cell line, AnT&. Cytoplasmic RNA extracted from this cell line which contains around 90% of the ad2 genome (7) was hybrid- ized in solution to the complementary strands of ad2 DNA.
by guest on June 17, 2020http://w
ww
.jbc.org/D
ownloaded from
Selection of Adenovirus mRNA
Ad2 A B c D E F G
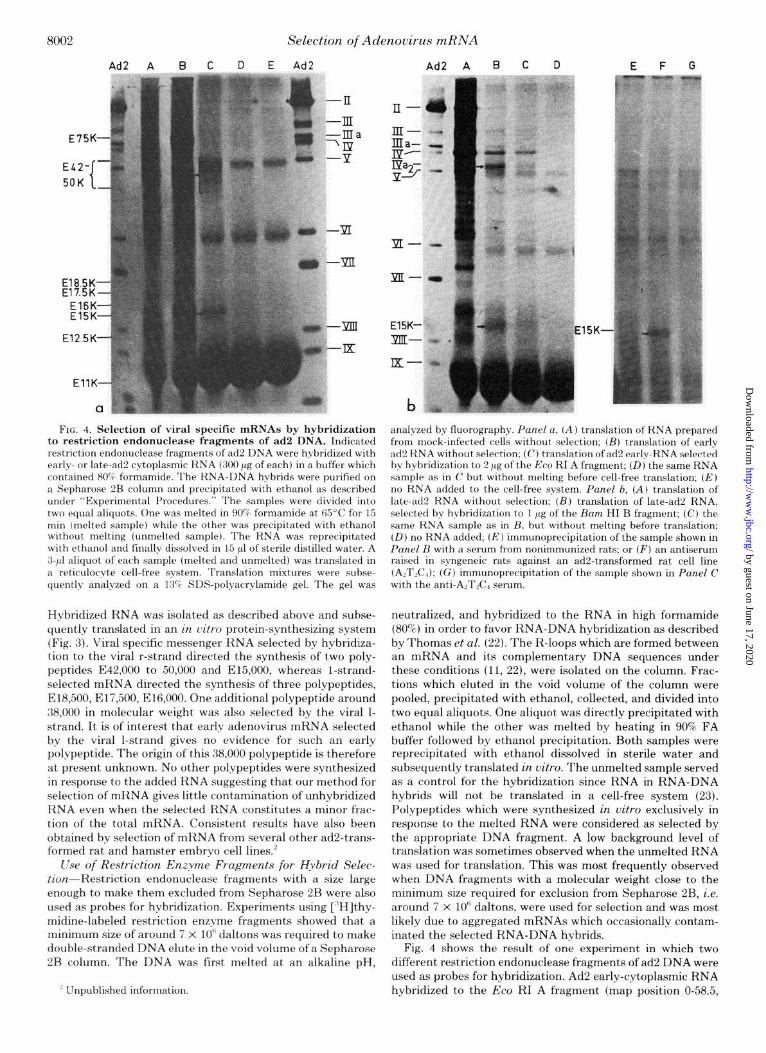
FIG. 4. Selection of viral specific mRNAs by hybridization to restriction endonuclease fragments of ad2 DNA. Indicated restriction endonuclease fragments of ad2 DNA were hybridized with early- or late-ad2 cytoplasmic RNA (300 pg of each) in a buffer which contained 80% formamide. The RNA-DNA hybrids were purified on a Sepharose 2B column and precipitated with ethanol as described under “Experimental Procedures.” The samples were divided into two equal aliquots. One was melted in 90% formamide at 65’C for 15 min (melted sample) whde the other was precipitated with ethanol without melting (unmelted sample). The RNA was reprecipitated with ethanol and finally dissolved in 15 ~1 of sterile distilled water. A 3-pl aliquot of each sample (melted and unmelted) was translated in a reticulocyte cell-free system. Translation mixtures were subse- quently analyzed on a 13% SDS-polyacrylamide gel. The gel was
Hybridized RNA was isolated as described above and subse- quently translated in an in zlitro protein-synthesizing system (Fig. 3). Viral specific messenger RNA selected by hybridiza- t,ion to the viral r-strand directed the synthesis of two poly- peptides E42,OOO to 50,000 and E15,000, whereas l-strand- selected mRNA directed the synthesis of three polypeptides, E18,500, E17,500, E16,OOO. One additional polypeptide around 38,000 in molecular weight was also selected by the viral l- strand. It is of interest that early adenovirus mRNA selected by the viral l-strand gives no evidence for such an early polypeptide. The origin of this 38,000 polypeptide is therefore at present unknown. No other polypeptides were synthesized in response to the added RNA suggesting that our method for selection of mRNA gives little contamination of unhybridized RNA even when the selected RNA constitutes a minor frac- tion of the total mRNA. Consistent results have also been obt.ained by selection of mFtNA from several other ad2-trans- formed rat and hamster embryo cell lines.’
Use of Restriction Enzyme Fragments for Hybrid Selec- tion--Restriction endonuclease fragments with a size large enough to make them excluded from Sepharose 2B were also used as probes for hybridization. Experiments using [RH]thy- midine-labeled restriction enzyme fragments showed that a minimum size of around 7 x 10’ daltons was required to make double-stranded DNA elute in the void volume of a Sepharose 2B column. The DNA was fist melted at an alkaline pH,
’ Unpublished information.
5K
analyzed by fluorography. Panel a, (A ) translation of RNA prepared from mock-infected cells without selection; (B) translation of early ad2 RNA without selection: (C) translation of ad2 early-RNA selected by hybridization to 2 pg of the Eco RI A fragment; (D) the same RNA sample as in C but without melting before cell-free translation; (E) no RNA added to the cell-free system. Panel b, (A) translation of late-ad2 RNA without selection: (B) translation of late-ad2 RNA, selected by hybridization to 1 pg of the Bum HI B fragment; (C) the same RNA sample as in B, but without melting before translation; (D) no RNA added; (E) immunoprecipitation of the sample shown in Panel B with a serum from nonimmunized rats; or (8’) an antiserum raised in syngeneic rats against an ad2-transformed rat cell line (AATLC$); (G) immunoprecipitation of the sample shown in Panel C with the anti-AzTKI serum.
neutralized, and hybridized to the RNA in high formamide (80%) in order to favor RNA-DNA hybridization as described by Thomas et al. (22). The R-loops which are formed between an mRNA and its complementary DNA sequences under these conditions (11, 22), were isolated on the column. Frac- tions which eluted in the void volume of the column were pooled, precipitated with ethanol, collected, and divided into two equal aliquots. One aliquot was directly precipitated with ethanol while the other was melted by heating in 90% FA buffer followed by ethanol precipitation. Both samples were reprecipitated with ethanol dissolved in sterile water and subsequently translated in uitro. The unmelted sample served as a control for the hybridization since RNA in RNA-DNA hybrids will not be translated in a cell-free system (23). Polypeptides which were synthesized in vitro exclusively in response to the melted RNA were considered as selected by the appropriate DNA fragment. A low background level of translation was sometimes observed when the unmelted RNA was used for translation. This was most frequently observed when DNA fragments with a molecular weight close to the minimum size required for exclusion from Sepharose 2B, i.e. around 7 x 10’ daltons, were used for selection and was most likely due to aggregated mRNAs which occasionally contam- inated the selected RNA-DNA hybrids.
Fig. 4 shows the result of one experiment in which two different restriction endonuclease fragments of ad2 DNA were used as probes for hybridization. Ad2 early-cytoplasmic RNA hybridized to the Eco RI A fragment (map position O-58.5,
by guest on June 17, 2020http://w
ww
.jbc.org/D
ownloaded from

Selection of Adenovirus mRNA 8003
molecular weight 13.6 x 10”) selected two ad2 early mRNAs encoding polypeptides E42,OOO to 50,000 and E15,OOO (Fig. 4A ). Hybridization of a late-cytoplasmic ad2 RNA to fragment Ram HI B of ad2 DNA (map position O-29.1, molecular weight 6.7 x 10”) selected three ad2 mRNAs encoding polypeptides IVa.,, 15,000, and IX (Fig. 4B). Polypeptide IX which is not readily visible in Fig. 4B due to the endogenous globin syn- thesis, was revealed by immunoprecipitation with a monospe- cific antiserum against virion polypeptide IX (14) (data not shown). The 15,000 polypeptide synthesized in response to the selected mRNA was immunoprecipitated with an antise- rum raised in syngeneic rats against an ad2-transformed rat cell line (A,T,C,) (16), suggesting that the mRNA encoding the late 15,000 polypeptide is identical with the mRNA en- coding the early 15,000 polypeptide (E15,OOO). Large scale purification of mRNAs by purification of R-loops on the Sepharose 2B column is, however, hampered by the low efficiency of hybridization between the RNA and its comple- mentary DNA sequence under R-loop conditions.
In conclusion, our present results demonstrate that chro- matography on Sepharose 2B is a simple and efficient method for isolation and purification of RNA-DNA hybrids. The selection of early adenovirus mRNA by l- and r-strand DNA clearly shows that the specificity which is obtained with this method is equal to or better than what can be achieved with other methods. Furthermore selection of adenovirus mRNA from an ad2-transformed rat cell line, in which the viral mRNA constitutes less than 1% of the total RNA content of the cell, shows that the method is valid also for systems in which the RNA species to be selected is present in very low amounts.
The size of the DNA probe used for hybrid selection and the possibility to separate its complementary strands appear at present to be the major limitation of the method. This problem can in many cases be circumvented by cloning frag- ments in a suitable bacterial vector. The complementary strands of bacteriophage X, a commonly used cloning vehicle, are easy to separate and of sufficient size to make our present method applicable. Recombinant DNA research is progressing at a very rapid pace and libraries of cloned fragments from different organisms are becoming available. We thus expect that this simple method for isolation and purification of spe- cific mRNA species should have a wide application.
In a recent communication we used this method in combi- nation with the hybrid arrested cell-free translation method
(23) in order to map a late-ad2 minor mRNA encoding a viral maturation protein on the ad2 genome (24).
Achnou$e&menls-We thank Marianne Gustafson and Christina SjSholm for excellent secretarial work.
1.
2.
3.
4.
5.
6.
7.
8.
9. 10.
11.
12.
13.
14.
15.
16.
17.
18. 19. 20. 21. 22.
23.
24.
REFERENCES
Smith, M. J., Hough, B. R., Chamberlin, M. E., and Davidson, E. H. (1974) J. Moi. Biol. 85, 103-126
Lewis. J. B.. Atkins. ,J. F.. Solem. R.. Gesteland. R. F.. and Anderson, C. W. (1976) Cell 7, 14i-15i
Lewis, J. B., Anderson, C. W., and Atkins, J. F. (1978) Cell 12, 37-44
Mak, S., iiberg, B., Johansson, K., and Philipson, L. (1976) Biochemistry 15, 5754-5761
Gilboa. E . . Prives. C. L.. and Aviv. H. (1975) Biochemistrv 14. 4215-4220
_
Pettersson, U., Tibbetts, C., and Philipson, L. (1976) J. Mol. Biol. 101,479-501
Gallimore, P. H., Sharp, P. A., and Sambrook, J. (1974) J. Mol. Biol. 89 49-72
Tibbetts, b., Pettersson. U., Johansson, K., and Philipson. L. (1974) J. Viral. 13,370-377
(1977) cJ. Viral. 22, 830-831 Brawerman. G.. Mendecki. J.. and Lee. S. Y. (1972) Biochemistrv
11,637~641 Casey, J., and Davidson, N. (1977) Nucleic Acids Res. 4, 1539-
1552 ,Johansson, K., Pettersson, U., Philipson, L., and Tibbetts, C.
(1977) J. Viral. 23, 29-35 Pelham, H. R. B., and Jackson, R. J. (1976) Eur. J. Biochem. 67,
247-256 Persson, H., Pettersson, IJ., and Mathews, M. B. (1978) Virology
90,67-79 Maizel, J. V. (1971) Methods in Virologv (Maramorosch, K.. and
Koprowski, H., eds) Academic Press;New York Kvist. S.. Ostbex L., Persson. H.. Phil&on. L., and Peterson. P.
A. (1978) Procy Natl. Acad. Sci. U. S: A. 75, 5674-5678 Bonner, W. M., and Laskey, R. A. (1974) Eur. J. Biochem. 46,
83-88 Saborio, J. L., and Oberg, B. (1976) CJ. Virol. 17, 865-875 Harter, M. L., and Lewis, J. B. (1978) J. Viral. 26, 736-749 Flint, S. d. (1977) Cell 10, 153-166 Flint, S. J., and Sharp, P. A. (1976) J. Mol. Biol. 106, 749-771 Thomas, M., White, -R. L., and Davis, tl. W. (1976) Proc. Natl.
Acad. Sci. U. S. A. 73.2294-2298 Paterson, B. M., Roberts; B. E., and Kuff, E. L. (1977) Proc. Natl.
Acad. Sci. U. S. A. 74,4370-4374 Persson, H., Mathisen, B., Philipson, L., and Pettersson, U. (1979)
Virology 93, 198-208
by guest on June 17, 2020http://w
ww
.jbc.org/D
ownloaded from

H Persson, M Perricaudet, A Tolun, L Philipson and U PetterssonPurification of RNA-DNA hybrids by exclusion chromatography.
1979, 254:7999-8003.J. Biol. Chem.
http://www.jbc.org/content/254/16/7999Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/254/16/7999.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on June 17, 2020http://w
ww
.jbc.org/D
ownloaded from





























