Embed Size (px)
Citation preview

214 lllOCHIMICA I4"I" BIOPHYSICA A('TA
B~,~, 35914
P U R I F I C A T I O N AND P R O P E R T I E S OF CYTOCHROME c 3 01:
D E S U L F O V I B R I O VULGARIS , M I Y A Z A K I
T A T S U H I K O Y A G I AND K 1 Y O F U M I M A R U Y A M A
Department of Chemistry, Shizuoha University, Shizuoka 420 (Japan)
(Received March 22nd, 1971)
SUMMARY
Cytochrome c 3 was isolated in a homogeneous state from Desulfovibrio vulgaris, Miyazaki, and its properties examined and compared with those of the cytochrome c3 from D. vulgaris, Hildenborough.
The absorption spectrum of the ultraviolet region of ferrocytochrome c 3 was recorded for the first time. The spectra of the cytochrome c a from the Miyazaki strain has five peaks (at 552, 523, 419, 323 and 275 nm) in the ferro-form, three peaks (at 53o with a shoulder at 56o, 41o and 35o nm) in the ferri-form, and eight isosbestic points (at 56o, 542, 532, 5o8, 432, 412, 343 and 254 nm). Upon contact with CO, the spectrum of the ferro-form changed. I ts isoelectric point is near lO.6, and its redox potential, - -o .29 V.
The Miyazaki strain cytochrome c3 contains 4 hemes. I ts N-terminus is histi- dine. Arginine, as well as isoleucine, is absent. These features are all different from those reported for the cytochrome c 3 of D. vulgaris, Hildenborough.
INTRODUCTION
Cytochrome c a is present in sulfate-reducing bacteria (Desulfovibrio). I t has been isolated from D. vulgaris, Hildenborough *,1,2 and from D. vulgaris, Miyazaki *,3-s, and is also present in other species of Desulfovibrio °-8. I ts isoelectric point and molecular weight were reported to be near lO.5 and 13 ooo 2 , i.e. comparable to those of mammal ian and yeast cytochromes c. I ts iron content was, however, reported to be o.90/o 2,5, which is twice as high as tha t of mammalian and yeast cytochromes c. On this basis, POSTGATE 2 concluded tha t the molecule of cytochrome cz from D. vulgaris, Hildenborough, contains two hemes; HORIO AND KAMEN 9 confirmed tile
Abbrev i a t i on : dansyl , I - d ime thy l aminonaph tha l ene -5 - su l fony l . " D. vulgaris, Hildenborough , was descr ibed as D. desulfurieans, Hildenborough , in refs. i,
2, 6 and 9, and D. vulgaris, Miyazaki , as D. desulfuricans w i t h o u t speci fy ing any s t ra in n a m e in refs.3 5 and I I . Accord ing to the recent nomenc l a tu r e proposed by POSTGATE AND CAMPBELL TM, t h e y are to be cal led D. vulgaris. Since the l a t t e r s t ra in was i so la ted from a p a d d y field in Miyazaki , Japan , we propose to call th is organism, D. vulgaris, Miyazaki .
Biochim. Biophys. Acta, 243 (I97 I) 214 224

T. YAGI, K. MARUYAMA 215
double heine structure for their crystalline cytochrome c 3 also obtained from D. vulgaris, Hildenborough. Recently, DRUCKER et al.7, 8 purified the cytochrome c a from D. vulgaris, Hildenborough, D. salexigens, and D. desulfuricans, VC, and reported that these cytochromes have three hemes.
In our laboratory, cytochrome c a was highly purified from D. vulgaris, Miyazaki, and characterized as an electron carrier for hydrogenase (H2 :ferricytochrome c a oxido- reductase) n. The properties of the cytochrome have been extensively studied and the absorption spectra of the ferri- and ferro-forms of cytochrome ca in the ultraviolet as well as visible regions recorded. These properties as well as the heme content and amino acid composition, some of which are different from those reported by earlier workers, will be described in this paper.
MATERIALS AND METHODS
Bacteria Desulfovibrio vulgaris, Miyazaki, was cultured on a large scale as described by
YAGI et al. n.
Chemicals and reagents I-Dimethylaminonaphthalene-5-sulfonyl (dansyl) chloride was purchased from
Seikagaku Kogyo Co., Tokyo. Yeast cytochrome c was a generous gift from Sankyo Pharmaceutical Co., Tokyo. Highly purified hydrogenase preparations were obtained by the procedure described by YAGI TM. Spotfilm for thin-layer chromatography (25o-#m-thick silicagel plate) was a product of Tokyo Kasei Kogyo Co., Tokyo.
Amino acid composition Cytochrome c 3 was hydrolyzed with 6 M HC1 in evacuated glass tubes for 24
and 48 h at i i o °, and the amount of amino acids except cystine were determined with a Hitachi automatic amino acid analyzer (KLA-3B) by the procedure described by SPACKMAN et al. ~a. Corrections were made for the destruction of serine and threonine by extrapolation to zero time of hydrolysis. For assay of cysteine, heine was removed from the cytochrome and the protein moiety was treated with performic acid as described by AMBLER 14. The mixture was then dried in a stream of N 2 at room temperature, and hydrolyzed with 6 M HC1 in evacuated glass tubes for 24 and 48 h at i i o °. The amount of cysteate was also determined with an amino acid analyzer and corrected for destruction by extrapolation to zero time of hydrolysis.
Identification of dansyl-amino acid This was carried out on thin-layer chromatograms on spotfilms developed in
Solvent I I (toluene-chloroethano1-25% ammonia (6:1o:4, by vol.)) with synthetic dansyl-amino acids as standards, as described by GROS AND LABOUESSE 15.
RESULTS
Purification of cytochrome c 3 Bacterial sonicate prepared from 33 g wet cells 1~ was centrifuged at 80 ooo × g
for 60 rain. The supernatant fluid (240 ml) containing about 8 mg cytochrome c 3 was
Biochim. Biophys. Acta, 243 (i97 I) 214 224
216 CYTOCHROME C a OF Desulfovibrio vulg, aris
15
°lo
15 20 25 30 Fraction Number (3.56 ml/tube)
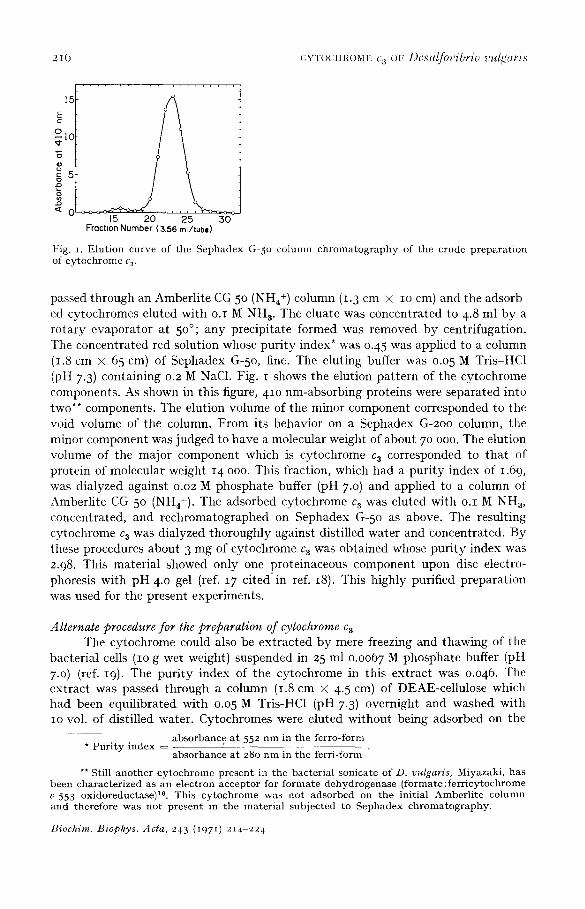
Fig. I, E lu t ion curve of the Sephadex G-5o co lumn c h r o m a t o g r a p h y of the crude p r epa ra t i on of cy toch rome c 3.
passed through an Amberlite CG 50 (NH4+) column (I.3 cm × Io cm) and the adsorb- ed cytochromes eluted with o.I M NH 3. The eluate was concentrated to 4.8 ml by a rotary evaporator at 5o°; any precipitate formed was removed by centrifugation. The concentrated red solution whose purity index" was o.45 was applied to a column (1.8 cm × 65 cm) of Sephadex G-5o, fine. The eluting buffer was o.o5 M Tris-HCl (pH 7.3) containing o.2 M NaC1. Fig. I shows the elution pat tern of the cytochrome components. As shown in this figure, 41o nm-absorbing proteins were separated into t w o " components. The elution volume of the minor component corresponded to the void volume of the column. From its behavior on a Sephadex G-2oo column, the minor component was judged to have a molecular weight of about 7 ° ooo. The elution volume of the major component which is cytochrome c 3 corresponded to that of protein of molecular weight 14 ooo. This fraction, which had a puri ty index of 1.69, was dialyzed against o.o2 M phosphate buffer (pH 7.o) and applied to a column of Amberlite CG 5 ° (NH4+). The adsorbed cytochrome c 3 was eluted with o.I M NH 3, concentrated, and rechromatographed on Sephadex G-5 o as above. The resulting cytochrome ca was dialyzed thoroughly against distilled water and concentrated. By these procedures about 3 mg of cytochrome c a was obtained whose purity index was 2.98. This material showed only one proteinaceous component upon disc electro- phoresis with pH 4.o gel (ref. 17 cited in ref. 18). This highly purified preparation was used for the present experiments.
Alternate procedure for the preparation of cytoehrome c a The cytochrome could also be extracted by mere freezing and thawing of the
bacterial cells (IO g wet weight) suspended in 25 ml 0.0067 M phosphate buffer (pH 7.0) (ref. 19). The purity index of the cytochrome in this extract was 0.046. The extract was passed through a column (1.8 cm × 4-5 cm) of DEAE-cellulose which had been equilibrated with 0.05 M Tris-HC1 (pH 7.3) overnight and washed with io vol. of distilled water. Cytochromes were eluted without being adsorbed on the
absorbance a t 552 n m in the ferro-form " P u r i t y index =
absorbance a t 28o nm in the ferr i-form
*" Stil l ano the r cy toch rome presen t in the bac te r i a l sonicate of D. vulgaris, Miyazaki , has been charac te r ized as an e lect ron acceptor for fo rmate dehydrogenase ( fo rma te : f e r r i cy toch rome c-553 oxidoreductase) 16. This cy toch rome was not adsorbed on the in i t i a l Amber l i t e column and therefore was not p resen t in the ma te r i a l subjec ted to Sephadex c h r o m a t o g r a p h y .
Biochim. Biophys. Acta, 243 (1971) 214-224
CYTOCHROME C 3 OF Desulfovibrio vulgaris 217
column. The p u r i t y index of the cy tochrome in the eluate was o . I I . This was lyo- phil ized, dissolved in 3.0 ml water , and subjec ted to Sephadex G-5o column chro- m a t o g r a p h y as descr ibed in the preceding section. The elut ion pa t t e rn of the cyto- chromes is shown in Fig. 2. The fract ions e luted in tubes between No. 21 and 25 were collected, and d ia lyzed against 0.02 M phospha te buffer. The pu r i t y index of cy tochrome ca in this p repa ra t ion was 0.28. This p repa ra t ion is b y no means a pure cy tochrome c a p repara t ion , bu t is the p repa ra t ion obta inable b y one of the mi ldes t procedures available. This could be purified fur ther by adsorp t ion on and elut ion from Amber l i t e CG 50 (NH4+) column as descr ibed in the preceding section to give a p repa ra t ion with a pu r i ty index comparab le to t ha t of the p repara t ion descr ibed in the preceding section.
c£ 0.5
o o.4
"8 02
0,2 g
< 0 15 20 25 30 Froction Number (3.56 ml/tube)
30O
200
£ ioo
®
° o
/ . i 2 3
Concentration of Cytochrome-Hem (UM)
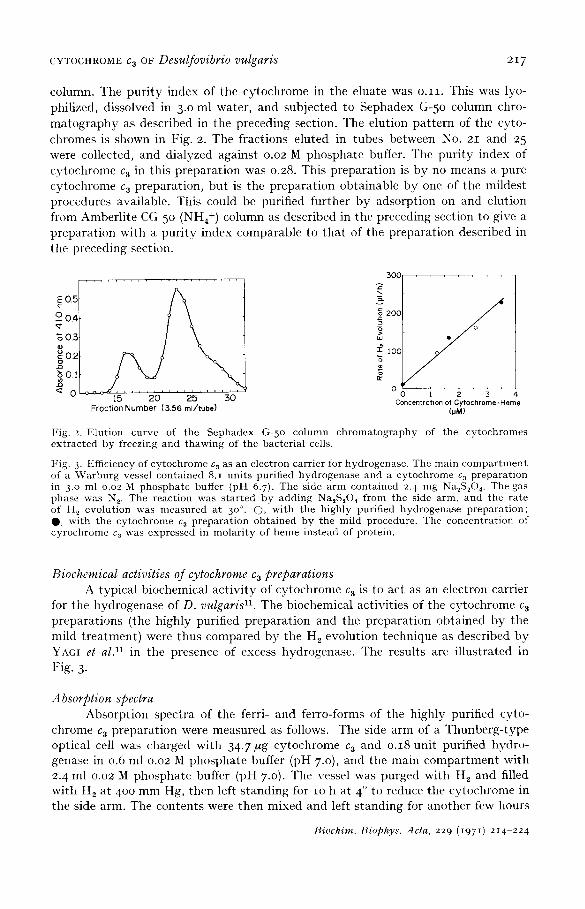
Fig. 2. FAution curve of the Sephadex G-5 o column chromatography of the cytochromes extracted by freezing and thawing of the bacterial cells.
Fig. 3. Efficiency of cytochrome c~ as an electron carrier for hydrogenase. The main compar tmen t of a ~Varburg vessel contained 8.1 units purified hydrogenase and a cytochrome c 3 preparat ion in 3.0 ml o.o2 M phospha te buffer (pH 6.7). The side a rm contained 2. 4 trig Na~S204. The gas phase was N 2. The reaction was s tar ted by adding Na~S204 from the side arm, and the rate of H 2 evolution was nleasured at 3 o°. (2), with the highly purified hydrogenase preparat ion; O, wi th the cytoehrome c 3 prepara t ion obtained by the mild procedure. The concentrat ion of cyrochrome c3 was expressed in molar i ty of heme instead of protein.
Biochemical activities of cytochrome c 3 preparations A typ ica l b iochemical a c t i v i t y of cy tochrome c 3 is to act as an electron carr ier
for the hydrogenase of D. vulgaris 11. The biochemical act ivi t ies of the cy tochrome c 3 p repa ra t ions (the h ighly purif ied p repa ra t ion and the p repa ra t ion ob ta ined b y the mild t r ea tmen t ) were thus compared b y the H 2 evolut ion technique as descr ibed b y YAGI et a l ) 1 in the presence of excess hydrogenase . The results are i l lus t ra ted in Fig. 3.
Absorption spectra Absorp t ion spec t ra of the ferri- and ferro-forms of the h ighly purified cyto-
chrome c 3 p repa ra t ion were measured as follows. The side arm of a Thunbe rg - type opt ical cell was charged with 34.7 ktg cy toehrome c 3 and o.18 uni t purified hydro- genase in 0.6 ml 0.02 M phospha te buffer (pH 7.o), and the main c o m p a r t m e n t wi th 2. 4 ml 0.02 M phospha te buffer (pH 7.0). The vessel was purged with H 2 and filled wi th H 2 at 400 m m Hg, then left s t and ing for IO h at 4 ° to reduce the cy tochrome in the side arm. The contents were then mixed and left s t and ing for ano ther few hours
Biochim. Biophys. Acta, 229 (1971 ) 214 224
2 I ~ T . Y A ( ; I , K . M A I , ~ U Y A M : \
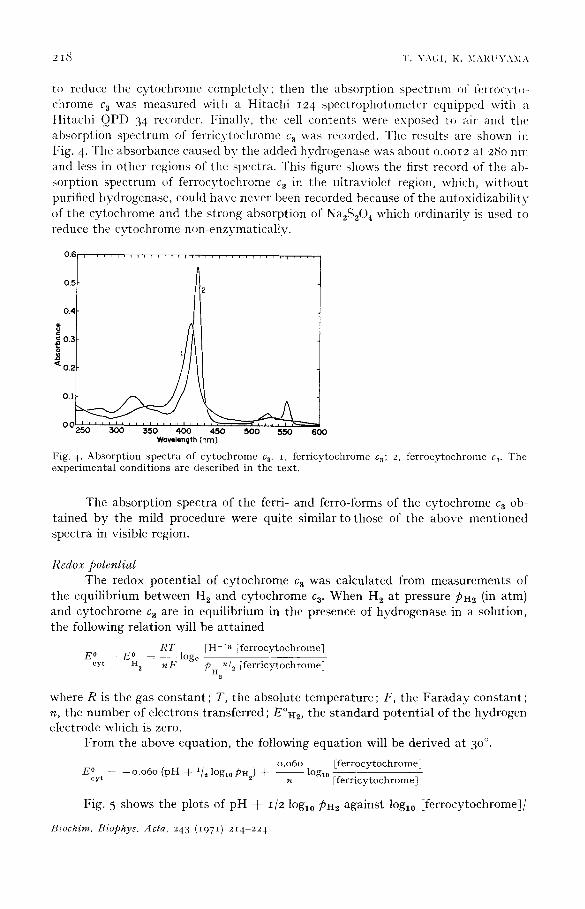
to reduce the cytochrome completely; then the absorption spectrum of l\,rrocvt~- chrome c 3 was measured with a Hitachi i24 spectrophotometer equipped with a Hitachi QPD 34 recorder. I~inally, the cell contents were exposed to air and the' absorption spectrum of ferricytochrome c a was recorded. The results are shown in l"ig. 4. The absorbance caused bv the added hydrogenase was about o.ooI2 at 28o nm and less in other regions of the spectra• This figure shows the first record of the ab- sorption spectrum of ferrocytochrome c 3 in the ultraviolet region, which, without purified hydrogenase, could have never been recorded because of the autoxidizabili ty of the cytochrome and the strong absorption of Na2S204 which ordinarily is used to reduce the cvtochrome non-enzymatically.
0 .6 , . . . . , . , , , , . . . . , , , . . , . . . . , . . . . , . . . .
0 .5
0.2
* ~ 0 . i
0.
0 0 I , , , , = , I i , i , i J , i , , ~
• 250 3 0 0 350 4 0 0 4 5 0 5 0 0 550 6 0 0 Wavelength (nm)
F i g . 4 . A b s o r p t i o n s p e c t r a o f c y t o c h r o m e c 3. i , f e r r i c y t o c h r o m e c3; 2 , f e r r o c y t o c h r o m e c.~. T h e
e x p e r i m e n t a l c o n d i t i o n s a r e d e s c r i b e d i n t h e t e x t .
The absorption spectra of the ferri- and ferro-forms of the cytochrome ca ob- tained by the mild procedure were quite similar to those of the above mentioned spectra in visible region.
Redox potential The redox potential of cytochrome c a was calculated from measurements of
the equilibrium between H 2 and cytochrome c 3. When H 2 at pressure PH2 (in atm) and cytochrome c 3 are in equilibrium in the presence of hydrogenase in a solution, the following relation will be at tained
R T [ H + I n i f e r r o c y t o c h r o m e ] E ° - - E ° - - l o g e
cy t H 2 nF PH2n/e i f e r r i c y t o c h r o m e ]
where R is the gas constant ; T, the absolute temperature; F, the Faraday constant ; n, the number of electrons transferred ; E°H2, the s tandard potential of the hydrogen electrode which is zero.
From the above equation, the following equation will be derived at 3 °° .
0 . 0 6 0 [ f e r r o c y t o c h r o m e ] E ° = - - 0 . 0 6 0 ( p H + 1 / z l o g t 0 P H 2 ) + - - l o g t o
cy t n [ f e r r i c y t o c h r o m e l
Fig. 5 shows the plots of pH + 1/2 lOgl0 PH2 against log10 [ferrocytochrome]/
Biochim. Biophys. Acta, 2 4 3 ( I 9 7 1 ) 2 1 4 - 2 2 4
CYXOCHROME C a OF Desulfovibrio vulgaris 2I 9
6.5
T Q. 6.0
+ el
5.5
o -leJ
5.C
0.0 0.5 1.0 1.5 [cyt C3 2+]
log [cyt c3 3÷]
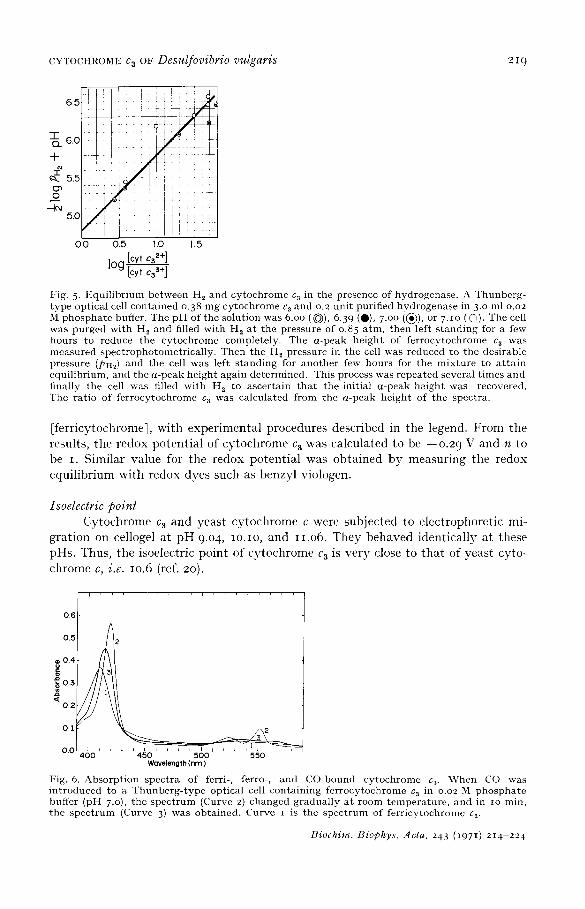
Fig. 5. Equi l ibr ium between H 2 and cytochrome c 3 in the presence of hydrogenase. A Thunberg- type optical cell contained o.38 mg cytochrome ca and o.2 unit purified hydrogenase in 3.o ml o.o2 M phospha te buffer. The p H of the solution was 6.oo ( ~ ) , 6.39 (O), 7.oo (~)), or 7.1o ( 0 ) . The cell was purged with H 2 and filled wi th H , at the pressure of o.85 atm, then left s tanding for a few hours to reduce the cytochrome completely. The a-peak height of ferrocytochronle c 3 was measured spectrophotometrical ly. Then the H 2 pressure in the cell was reduced to the desirable pressure (PHi) and the cell was left s tanding for another few hours for the mixture to a t ta in equilibrium, and the a-peak height again determined. This process was repeated several t imes and finally the cell was filled with H~ to ascertain tha t the initial a-peak height was recovered. The ratio of ferrocytochrome c a was calculated from the a-peak height of the spectra.
[ferricytochromel, with experimental procedures described in the legend. From the results, the redox potential of cytochrome c a was calculated to be --o.2 9 V and n to be I. Similar value for the redox potential was obtained by measuring tile redox equilibrium with redox dyes such as benzyl viologen.
Isoelectric point Cytochrome c3 and yeast cytochrome c were subjected to electrophoretic mi-
gration on cellogel at pH 9.04, IO.IO, and 11.o6. They behaved identically at these pHs. Thus, the isoelectric point of cytochrome c a is very close to that of yeast cyto- chrome c, i.e. lO.6 (ref. 20).
0 . 6
0.5
~o.4 o~ ~ 0 .3
< 0 . 2
I = i I i i i 0.0 ' ' ' ' ' r ' - 400 450 500 550
Wavelength (nm)
Fig. 6. Absorpt ion spectra of ferri-, ferro-, and CO-bound cytochrome c 3. When CO was introduced to a Thunberg- type optical cell containing ferrocytochrome c 3 in o.o2 M phospha te buffer (pH 7.o), the spect rum (Curve 2) changed gradually at room temperature , and in to min, the spect rum (Curve 3) was obtained. Curve I is the spect rum of ferricytochrome ca.
Biochim. Biophys. Acta, 243 (1971) 214-224

220 T. YA(;I, K. MARUYAMA
Binding of CO When CO was introduced to a Thunberg-type optical cell containing ferro-
cytochrome c a prepared by reducing the highly purified ferricytoehrome c a with Na2S204 at pH 7.o, the spectral change was observed (Fig. 6). The spectrum of ferro- cytochrome c 3 (Curve 2) was changed to Curve 3. The a, fl, and y peaks shifted toward the spectrum of the ferri-form (Curve I). However, the presumed CO-compound does not appear to be in the ferri-form, because its spectral curve does not cut the isosbestic points of the ferro- and ferricytochrome curves.
The spectral change was also observed when the ferrocytochrome c a preparation obtained by the mild procedure was treated with CO. The spectrum of the CO-com- pound was identical with the Curve 3 in Fig. 6.
POSTGATE 2 previously had stated that no spectral change occurred upon treat- ment of cytochrome c a with CO.
Heine content (I) A cytochrome c a solution was dialyzed against distilled water (15 1, 24 h)
and glass distilled water (i 1 2 times, 24 h each). The thoroughly dialyzed preparation thus obtained had an absorbance of 57-52 at 552 nm in the ferro-form. A I-ml portion of this preparation was frozen and lyophilized. Evacuation was continued for 20 h at room temperature after the cytochrome became a dry powder by visual appearance, and finally, for 3 tl at 85 °. Immediately after the vacuum was broken, the contents
T A B L E I
T H E R E S U L T S O F M E T A L A N A L Y S E S O F C Y T O C H R O M E C a
Analyzed by a tomic absorp t ion s pe c t rom e t r y a t I n s t i t u t e for Solid S ta te Physics, Un ive r s i ty of Tokyo. Values are expressed as/*g.
Cytochrome c 3 (z.oo rag) Blank (o. 5 ml diffusate in 0.5 ml aqueous solution of f inal dialysis)
F e I 5 . O -4 O .O 2 5
Zn i ,o o.o C u o . o o . o
Mn o.o o.o
weighed 7.43 mg; the preparation absorbed humidity quickly and, after 2 min, it weighed 7.5o rag. The specific absorbance coefficient of the a-peak of ferrocytochrome c 3 was, thus, 7.74 at pH 7.0. The compound, therefore, contains 3.9 moles heine per 14 ooo g (assumed molecular weight).
(2) The results of metal analyses by atomic absorption spectrometry are sum- marized in Table I. These analyses indicate that cytochrome c 3 contains 3.75 atoms Fe and 0.2 atoms Zn per 14 ooo g.
Amino acid composition The results of the amino acid analyses are shown in Table II.
N-terminal amino acid Dansylation of cytochrome c 3 was carried out as described by GRAY 22. Dansyl
Biochirn. Biophys. Acta, 243 (1971) 214-224

CYTOCHROME C a OF Desulfovibrio vulgaris 221
T A B L E I I
A M I N O A C I D C O M P O S I T I O N OF C Y T O C H R O M E 6 3 OF D. vulgar'is, M I Y A Z A K I
(a). Moles/mole of p ro te in as ca lcu la t ed from amino acid analysis . (b). Moles/mole of pro te in to nea res t whole in teger .
Residue Moles/mole of protein
(~) (b)
Lys ine 2o.9 H i s t i d ine 9.5 Argin ine o.o Cysteate* 7.7 A s p a r t a t e 12.5 Threoninc 6.5 Serine 5.o G l u t a m a t e 5- i Pro l ine 3.9 Glycine 9.7 Alanine i i, 8 H a l f cys t ine 6.8"* Valine 4.5 Meth ionine 2.8 Isoleucine o.o Leucine 3.o Tyros ine 2. 4 P h e n y l a l a n i n e 2. i T r y p t o p h a n
Tota l n u m b e r of res idues Mol.wt. of p ro te in mo ie ty § § Mol.wt. of 4 hemes Mol.wt. of cy toch rome ca
2 1
9-1o o
12-13 6 7
5 5 4
IO
I 2
5 3 o
3 2 3
2
o§
I O 7 - - I I I
I I 4 8 8 - - I 2 0 0 3
2 466 13 949-14 469
Recovered from the performic ac id - t rea ted pro te in der ived by remova l of heroes of cy toch rome c 3.
** High recovery of cys t ine by mere hydro lys i s of the na t ive cy tochrome c 3 was also r epo r t ed by CORAL et al. 21.
*** Obta ined from the cys t ea t e content . Assumed to be zero because of low absorbance a t 28o nm.
.~§ The presence of i 2 - i 6 amido groups is a s sumed from the amino acid analysis .
chloride (5 mg) in 0.25 ml acetone was added to a mixture of cytochrome c 3 (o.14 mg) and 240 m g urea in 0.8 ml 0.025 M NaHCOa buffer (pH 9.6), and the mixture left standing o+ernight at room temperature. The dansylated protein was precipitated by centrifugation, and washed twice with 8 ml water. The precipitate was then hydro- lyzed with 0.5 ml 6 M HC1 in an evacuated glass tube for 5 h at lO5 °. The hydrolyzate was dried in a stream of N2, dissolved in water and dried. The residue was dissolved in 0.02 M acetic acid, and extracted with peroxide-free ether. The ether layer did not contain any dansyl-amino acid. The aqueous layer contained three fluorescent materials as revealed by thin-layer chromatography on spotfilm using Solvent I I of GROS AND LABOUESSE 15. The material with the highest RF was dansyl-OH. That with the lowest RF had a free amino group, because, upon acetylation with acetic anhy- dride-pyridine (2:1, by vol.) (ref. 23), it could be extracted by ether; it was, thus, assumed to be e-dansyl-lysine. The material of intermediate RF was identified as dansyl-histidine upon cochromatography with authentic dansyl-histidine using Sol- vent II.
Biochim. Biophys. Acta, 243 ( i97 I) : '14-224

2 2 2 "I'. YA( ; I , K. M A I { [ ' Y A M A
D I S C U S S I O N
Cytochrome c a was first i sola ted by POSTGATE 2 from D. vulgaris, Hildenborough, and by ISHIMOTO et al. 5 from D. vulgaris, Miyazaki , in 1956- 7. Both authors repor ted tha t the iron content of the cy tochrome was o.9°4, POSTGATE proposed a double heme s t ruc ture for cy tochrome c a because his molecular weight es t imat ion was 13 ooo. Since then, s t ruc tura l analysis has main ly been conduc ted with the cy tochrome c a of the Hi tdenborough s t ra in of D. vulgaris. Thus, in 1 9 6 1 , HORIO AND KAMEN 9 crysta l - lized this cy tochrome c a and, on the basis of the results of thei r analysis of its heine content , suppor t ed the double heme s t ructure . In 197o, DRI:CKER ct al.7, s proposed a t r iple heme s t ruc ture for three p repara t ions of cy tochrome c a ob ta ined from D. vulgaris, Hildenborough, D. salexigens, and D. desulfuricans, VC. According to the amino acid sequence repor ted b y AMBLER 2~, there are two sequences of the type
C y s - X Y Cys His , and two sequences of the type Cys A B C D - C y s - H i s , and no SH or - S - S - l inkage in the molecule for the cy tochrome c a of the Hi ldenborough strain.
We have now ob ta ined cy tochrome c a of D. vulgaris, Miyazaki , in a homogene- ous s ta te and have examined i ts heme content , amino acid composi t ion, and o ther impor t an t propert ies . The results are summar ized in Tables I I I and IV. I t is obvious t ha t the cy tochrome c a of D. wdgaris, Miyazaki , is different from tha t of D. vulgaris, Hildenborough, since t hey are d is t inguishable in amino acid composi t ion and o ther impor t an t propert ies . One of the most s t r ik ing differences is t ha t the cy tochromc c a of D. vulgaris, Miyazaki , contains 4 hemes. This means t ha t the heine moie ty of this cy tochrome amounts to 17-18% of the molecule. Such hemoprote ins have never been
T A B L E 111
C O M P A R I S O N O F T H E A M I N O A C I D C O M P O S I T I O N O F C Y T O C H R O M E S C a
Strain Desulfovibrio vulgaris D. desul- D. sal- furicans, exigens
Miyazaki Hildenborough [:C
L y s i n e 21 20 17 22 H i s t i d i n e 9 - 1 o 9 9 0 A r g i n i n e o 1 o o
A s p a r t a t e 1 2 - i 3 ~ 2 7 9 T h r e o n i n e 6 - 7 5 5 6 S e r i n e 5 6 (~ ~ 2 G l u t a m a t e 5 5 t I 5 P r o l i n e 4 4 6 5 G l y c i n e i o 9 8 i I A l a n i n e 12 i o 0 13 C y s t e i n e 8 8 8 8 V a l i n e 5 8 9 O M e t h i o n i n e 3 3 i 2 I s o l e u c i n e o o i 2 L e u c i n e 3 2 6 4 T y r o s i n e 2 3 3 2 1 P h e n y l a l a n i n e 2 2 3 3 T r y p t o p h a n o o o o N u m b e r o f r e s i d u e s l O 7 - 1 1 1 lO 7 lO8 115
l i e f s . 24 7 8
Biochim. Biophys. Acta, 243 ( i 9 7 I) 2 1 4 - 2 2 4

CYTOCHROME C 3 OF Desulfovibrio vu!garis 223
T A B L E IV
C O M P A R I S O N OF THE P R O P E R T I E S OF TI lE C Y T O C H R O M E S g 3 OF D. IJlglgar~$
D a t a concern ing the Hi ldenborough s t ra in are fronl the references ind ica ted .
Cytochrome c 3 from Refs.
M iyazaki Hildenborough strain strain
Spect ra l d a t a Afl*/Aa* 0.52 0-54; °.51 7; 9 Ay*/Aa* 6. 3 6. 5 7, 9 AT/Aa* 4 .0 4.9; 4 .1 7; 9 A6*/Aa" 1.o 5 A6/Aa* 0.77 0.74 9 da*/A~so 2.98 2.95 9
Upon con tac t w i th CO, the spec t rum Changed Unchanged 2
H e m e con t en t 4 2;3 2, 9; 7 Approx. mol. wt. 14 ooo 13 5 °0 7, 24 N- t e rminus His t id ine Alanine 24 C- te rminus G l u t a m a t e 24 p l Near lO.6 IO-I I 2 Redox po ten t i a l 0.29 V --0.205 V 2, 7
Abbrev ia t ions : Aa*, Aft*, etc., absorbances a t a, fl, etc., peaks of the ferro-form; AT, A6, a n d A28 o, absorbances a t y and 6 peaks, and a t 28o nm of the ferr i-form; pI, isoelectric point .
reported in the literature. The lower iron content a for the Miyazaki strain cytochrome c a reported earlier would be due to contaminat ion with high molecular weight cyto- chrome which escaped detection until Sephadex column chromatography was prac- tised (see Fig. I).
The highly purified cytochrome c a combines with CO in contrast to the obser- vations previously made by POSTGATE 2. One might suspect tha t the cytochrome had been denatured when it was being eluted with o.I M NH a from the Amberlite column, since HORIO AND NAMER 9 reported the unstable nature of cytochrome c a against alkali. The cytochrome Ca preparat ion obtained by one of the mildest procedures was also found to combine with CO. The highly purified cytochrome c a and the cytochrome Ca obtained by the mild procedure behaved identically on Sephadex G-5o column chro- ma tography (Figs. I and 2), had identical absorption spectra, and both of them had identical biochemical act ivi ty in the hydrogenase system (Fig. 3)- I t is thus evident tha t the highly purified cytochrome c a preparat ion is not a denatured artifact.
The N-terminus of cytochrome c a of D. vulgaris, Miyazaki, is histidine. Arginine, as well as isoleucine, is absent. These structural features are all different from the established amino acid sequence of cytochrome c a of D. vulgaris, Hildenborough 24. In spite of these differences, the amino acid compositions of both cytochromes are very similar (Table I I I ) . Both contain 8 cysteine residues, about 1 7 acidic amino acids (or their amides), and about 2o lysine residues. The number of histidine, proline, methionine and aromatic amino acid units are almost the same in both cytochromes.
The redox potential was est imated to be - -o .2 9 V, instead of - -o .2o 5 V re- ported for the other preparat ions of cytochromes c a (refs. 2, 6, 7). I t is the lowest redox potential ever reported for c-cytochromes. Although the cytochrome c a contains
Biochim. Biophys. Acta, 243 (I97I) 214-224

224 T. YA(;1, K. MAI{UYAMA
4 heroes, it behaves like a one-electron carrier in tile redox system as shown in Fig. 5. This means that 4 heroes nmst be surrounded by an almost identical redox environ- ment and receive electrons independently as if they were retained in separate protein molecules. Elucidation of tile structure and of the mechanism of action of this unusual hemoprotein should help us to unders tand tile mechanism of electron t ransport on a molecular basis.
ACKNOWLEDGEMENTS
We are indebted to Professor H. Inokuchi of Universi ty of Tokyo and Professor T. Ozaki of Shizuoka Universi ty who afforded us facilities used in the present experi- ments; to Professor E. J. Hehre of Albert Einstein College of Medicine for suggestions and advice in preparing manuscript ; to Dr. S. Tamura and Miss T. Miyazawa for the metal analyses; to Dr. K. Takahashi and Mr. K. Asaoka for the amino acid analyses; and to Miss A. Yagi for measuring H2-cytochrome equilibria.
This work was supported in part by a grant from the Ministry of Educat ion
of Japan.
R E F E R E N C E S
I J. R. POSTGATE, Biochem. J., 56 (1954) xi, 58 (1954) ix. 2 J. R. POSTGATE, J. GeE. Microbiol., 14 (1956) 545. 3 3,I. ISHIMOTO, J. KOYAMA AND Y. NAGAI, Bull. Chem. Soc. Japan, 27 (1954) 565. 4 M. ISHIMOTO, J. KOYAMA AND Y. NAGAI, J. Biochem. Tokyo, 41 (1954) 763 . 5 M. [SHIMOTO, J. KOYAMA, T. YAGI AND M. SHIRAKI, J. Biochem. Tokyo, 44 (1957) 413. 6 J. LE GALL, G. MAZZA AND N. DRAGONI, Bioehim. Biophys. Acta, 99 (1965) 385 • 7 U. DRUCKER, E. B. TROUSIL, L. L. CAMPBELL, G. H. BARLOW AND E. MARGOLIASH, Bio-
chemistry, 9 (197 °) 1515. 8 U. DRUCKER, E. B. TROUSIL AND L. L. CAMPBELL, Biochemistry, 9 (197 o) 3395. 9 T. HORIO AND M. D. KAMEN, Bioehim. Biophys. Acta, 48 (1961) 266.
IO J. R. POSTGATE AND L. L. CAMPBELL, Bacteriol. Rev., 3 ° (1966) 732. I I T. YAGI, M. HONYA AND N. TAMIYA, Biochim. Biophys. Acta, 153 (1968) 699. I2 T. YAGI, J. Biochem. Tokyo, 68 (197 o) 649. I 3 D. H. SPACKMAN, V~ 7. H. STEIN AND S. MOORE, Anal. Chem., 3 ° (1958) 119o. i 4 R. P. AMBLER, Biochem. J., 89 (1963) 349. 15 C. GRos AND g . LABOUESSE, Eur. J. Biochem., 7 (1969) 463 • 16 T. YAGI, J. Biochem. Tokyo, 66 (1969) 473. 17 R. A. REISFELD, [J. J. LEWIS AND D. E. WILLIAMS, Nature, 195 (1962) 281. 18 Y. NAGAI, in The Methods in Biochemical Experiments, P a r t IX, TAMPAKUSHITSU-KAKUSAN-
KOSO, 1967, p. 3 (in Japanese) . 19 M. ISHIMOTO, T. YAGI AND M. SHIRAKI, J. Biochem. Tokyo, 44 (1957) 707 . 20 S. MINAKAMI, J. Biochem. Tokyo, 42 (1955) 749. 21 M. L. COVAL, T. HORIO AND M. D. KAMEN, Biochim. Biophys. Aeta, 51 (1961) 246. 22 "W. R. GRAY, Methods Enzymol., I I (1967) 139. 23 A. A. BENSON, H. DANIEL AND J{. WISER, Proc. Natl. Acad. Sci. U.S., 45 (1959) 1582. 24 R. P. AMBLER, Biochem. J., lO9 (1968) 47 P.
Biochim. Biophys. Acta, 243 (1971) 214-224





























