Embed Size (px)
Citation preview

ORIGINAL PAPER
Purification and characterization of heterologously expressednitrilases from filamentous fungi
Alena Petříčková & Alicja Barbara Veselá & Ondřej Kaplan & David Kubáč &
Bronislava Uhnáková & Anna Malandra & Jürgen Felsberg & Anna Rinágelová &
Philip Weyrauch & Vladimír Křen & Karel Bezouška & Ludmila Martínková
Received: 19 March 2011 /Revised: 21 July 2011 /Accepted: 3 August 2011 /Published online: 3 September 2011# Springer-Verlag 2011
Abstract Nitrilases from Aspergillus niger CBS 513.88, A.niger K10, Gibberella moniliformis, Neurospora crassaOR74A, and Penicillium marneffei ATCC 18224 wereexpressed in Escherichia coli BL21-Gold (DE3) after IPTG
induction. N. crassa nitrilase exhibited the highest yieldof 69,000 UL−1 culture. Co-expression of chaperones(GroEL/ES in G. moniliformis and P. marneffei; GroEL/ES and trigger factor in N. crassa and A. niger CBS513.88) enhanced the enzyme solubility. Specific activitiesof strains expressing the former two enzymes increasedapproximately fourfold upon co-expression of GroEL/ES.The enzyme from G. moniliformis (co-purified withGroEL) preferred benzonitrile as substrate (Km of0.41 mM, Vmax of 9.7 μmol min−1 mg−1 protein). The P.marneffei enzyme (unstable in its purified state) exhibitedthe highest Vmax of 7.3 μmol min−1 mg−1 protein in cell-free extract, but also a high Km of 15.4 mM, for 4-cyanopyridine. The purified nitrilases from A. niger CBS513.88 and N. crassa acted preferentially on phenylacetoni-trile (Km of 3.4 and 2.0 mM, respectively; Vmax of 10.6 and17.5 μmol min−1 mg−1 protein, respectively), and hydrolyzedalso (R,S)-mandelonitrile with higher Km values. Significantamounts of amides were only formed by the G. moniliformisnitrilase from phenylacetonitrile and 4-cyanopyridine.
Keywords Nitrilase . Chaperones . Aspergillus niger .
Gibberella moniliformis .Neurospora crassa . Penicilliummarneffei
Introduction
Nitrilases are versatile enzymes in biocatalysis, mediatingthe hydrolysis of diverse nitriles under mild conditions.Some of their products, notably hydroxy acids, such asmandelic (Rustler et al. 2008; Sosedov et al. 2009), 3-hydroxyvaleric (Wu et al. 2007), or glycolic acid (Wu et al.2008), are of significant industrial importance. Theseenzymes can also be used to produce amides, which are
Alena Petříčková and Alicja B. Veselá contributed equally to this work.
Electronic supplementary material The online version of this article(doi:10.1007/s00253-011-3525-7) contains supplementary material,which is available to authorized users.
A. Petříčková :A. B. Veselá :O. Kaplan :D. Kubáč :B. Uhnáková :A. Malandra :A. Rinágelová : P. Weyrauch :V. Křen :K. Bezouška : L. Martínková (*)Institute of Microbiology, Centre of Biocatalysis andBiotransformation,Vídeňská 1083,CZ-142 20 Prague, Czech Republice-mail: [email protected]
J. FelsbergInstitute of Microbiology, Centre for DNA Sequencing,Academy of Sciences of the Czech Republic,Vídeňská 1083,CZ-142 20 Prague, Czech Republic
A. Petříčková :A. B. Veselá :K. BezouškaDepartment of Biochemistry, Faculty of Science,Charles University in Prague,Hlavova 8,CZ-128 40 Prague, Czech Republic
A. MalandraDepartment of Chemistry, Chemical Engineering and Materials,University of L’Aquila,Via Campo di Pile - Zona industriale di Pile,67100 L’Aquila, Italy
P. WeyrauchInstitute of Molecular Microbiology and Biotechnology,Westfalian Wilhelms-University Münster,Correnstrasse 3,48149 Münster, Germany
Appl Microbiol Biotechnol (2012) 93:1553–1561DOI 10.1007/s00253-011-3525-7

their usual by-products and even major products in somecases (e.g., Osswald et al. 2002; Fernandes et al. 2006;Kaplan et al. 2006a; Sosedov et al. 2009).
Most of the nitrilases studied and explored as biocatalystshave been bacterial in origin except for a few fungal and plantenzymes (for reviews see, O'Reilly and Turner 2003; Thuku etal. 2009). Besides this, nitrilases of unknown origin wereobtained from metagenomic DNA (Robertson et al. 2004).According to both activity screens (Kato et al. 2000) andgene database searches (Martínková et al. 2009), filamentousfungi seem to be a rich source of nitrilases. The first twofungal nitrilases were purified in the Fusarium genus andcharacterized (Harper 1977; Goldlust and Bohak 1989)without determining their amino acid sequences. The othertwo nitrilases from the same genus were purified andcharacterized by ourselves and their partial amino acidsequences determined (Vejvoda et al. 2008, 2010). Thenitrilase studied by us in Aspergillus niger K10 was the firstfungal nitrilase to be purified from both its native (Kaplan etal. 2006a) and heterologous producer (Escherichia coli;Kaplan et al. 2011a). Gene databases proved to be anabundant source of hypothetical nitrilase sequences and threeof these enzymes were recently expressed in E. coli by us,namely a nitrilase from A. niger CBS 513.88, Neurosporacrassa OR74A, and Gibberella moniliformis (Kaplan et al.2011b). Synthetic genes were prepared according to pub-lished sequences, and functional enzymes were obtained in allcases. The aim of this work was to characterize theseenzymes in order to define their potential biocatalytic use.A further two enzymes were also included in this study,namely a nitrilase from Penicillium marneffei ATCC 18224and the aforementioned enzyme from A. niger K10. Bothenzymes were expressed from the corresponding syntheticgenes but the latter also from the gene amplified from thecDNA of the native organism to compare the efficiency ofboth methods. Aiming to improve the heterologous expres-sion of nitrilases, we also examined the effect of molecularchaperones (GroEL/ES, dnaK-dnaJ-grpE, trigger factor (TF)),which are known for their ability to facilitate protein foldingand hence to improve the production of active proteins(Nishihara et al. 2000).
Materials and methods
Nitrilase expression
The expression of all nitrilases was performed in E. coliBL21-Gold (DE3) (Agilent Technologies—StratageneProducts, USA). The enzyme from A. niger K10 (GenBankABX75546) was expressed using either the gene amplifiedfrom the wild-type producer (Kaplan et al. 2011a) or thesynthetic gene (GeneArt, Regensburg, Germany), the codon
frequency of which was optimized using GeneArt's ownsoftware. A strain transformed with plasmid pOK101 wasused to express the former gene (Kaplan et al. 2011a). Thelatter gene was cloned into the Nde I and Hind III sites ofplasmid pET 30a(+) to construct plasmid pOK106 andexpressed in E. coli without the His-tag. The expression ofnitrilases from N. crassa OR74A (GenBank CAD70472),G. moniliformis (GenBank ABF83489), and A. niger CBS513.88 (GenBank XP_001397369) was performed usingstrains transformed with plasmids pOK103, pOK104, andpOK105, respectively (Kaplan et al. 2011b). The geneencoding the nitrilase in P. marneffei ATCC18224 (GenBankXP_002144951 (protein)) was synthesized with GeneArtaccording to the published sequence, which was obtainedusing the programs BLASTX and BLASTP (Altschul et al.1997; http://blast.ncbi.nlm.nih.gov/Blast.cgi). The gene wascloned into the Nde I and Hind III sites of plasmid pET 28a(+) to construct plasmid pOK107 and expressed in E. coliwith an N-terminal His-tag. The optimized gene sequencesare shown in supplementary Fig. S1 (P. marneffei, GeneBankJN012233; A. niger K10, GeneBank JN243351) and in theprevious work (A. niger CBS 513.88, GeneBank JN012230;G. moniliformis, GeneBank JN012231; N. crassa, GeneBankJN012232; Kaplan et al. 2011b, Electronic supplementarymaterial).
Nitrilase and chaperone co-expression
E. coli BL21-Gold (DE3) (Agilent Technologies—StratageneProducts, USA) was transformed with one of the plasmidscontaining nit genes and one of the chaperone plasmids(pG-KJE8, pGro7, pKJE7, pG-Tf2 or pTf16; Takara BioInc., Japan), resulting in 30 strain harboring different nit andchaperone gene combinations. Strains lacking chaperonegenes served as controls. The strains were grown in LBwith kanamycin (50 μg mL−1) and chloramphenicol(20 μg mL−1). Chaperone screening was performed at25°C in 10-mL culture volumes in shaken 50-mL Falcontubes closed with screw caps. After OD610 reached approx-imately 0.5, the expression of chaperones was induced with
L-arabinose (2 gL−1) or tetracycline (5 μg L−1) according tothe manufacturer's instructions (Takara). At the same time,expression of the nitrilase was induced with 0.5 mM IPTG,and the cultivation continued under the same conditions for afurther 17 h.
Selected cultures were scaled up to a 200-mL volumeusing shaken 500-mL Erlenmeyer flasks closed with cottonplugs. The cultivation was carried out at 37°C for 3.5–4 h(until the culture reached an OD610 of approximately 0.5).Then, the cultivation temperature was decreased to 25°Cand the expression of chaperones and nitrilase was inducedas described above. The cells were harvested after a further17 h.
1554 Appl Microbiol Biotechnol (2012) 93:1553–1561

Nitrilase purification
The cells were disrupted by sonication and cell debrisremoved by centrifugation (13,000 g, 4°C, 15 min). Thenitrilases from A. niger CBS 513.88 and N. crassa werepurified from cell-free extracts on a Hi-Prep 16/10 Q FFcolumn (GE Healthcare), with a linear gradient of NaCl(0.15–1 M) in Tris/HCl buffer (50 mM, pH 8.0) at2 mL min−1 in the first step. Active fractions were pooled,concentrated using an Amicon Ultra-4 unit (cut-off 30 kDa;Millipore) and applied on a Superdex 200 HR 10/300 GLcolumn (GE Healthcare) using a Tris/HCl buffer (50 mM,pH 8.0, 150 mM NaCl) at 0.4 mL min−1. Alternatively,HiPrep 16/60 Sephacryl S-200 HR (GE Healthcare) wasused in this step for nitrilase from G. moniliformis. Enzymewas eluted with the same buffer at 2 mL min−1. Activefractions were pooled, concentrated as described above andstored at −80°C.
The nitrilase from P. marneffei was partially purified byaffinity chromatography on Talon Metal Affinity Resin(Clontech). The cell-free extract was incubated with theresin at 4°C for 30 min, centrifuged (750 rpm, 5 min) andthe pellet resuspended in Tris/HCl buffer (50 mM, pH 8.0).The mixture was transferred into a 25-ml column andproteins eluted with a stepwise gradient of imidazole (0–1–150 mM) in the same buffer. Active fractions (eluted at150 mM imidazole) were pooled and concentrated asdescribed above.
Nitrilase assay
Strains were screened for optimum nitrilase–chaperonecombinations using reaction mixtures (total volume0.5 mL each) containing an appropriate amount of wholecells resuspended in Tris/HCl buffer (50 mM, pH 8.0;150 mM NaCl) and 25 mM benzonitrile (from 500 mMstock solution in methanol). The suspensions were shakenat 30°C for 5 min, the reaction initiated by the addition ofbenzonitrile and terminated after a further 10 min incuba-tion under the same conditions. Cells from cultivations on200-mL scale were assayed in an analogous way with slightmodifications; activities of G. moniliformis and P. marneffeiwere assayed with 25 mM benzonitrile after a 5 minreaction and those of N. crassa and A. niger with 25 mMphenylacetonitrile after a 1 min reaction.
To determine the nitrilase activity in the soluble andinsoluble fractions, the cells were disrupted by sonicationand cell debris removed by centrifugation (13,000×g, 4°C,30 min). The pellet was resuspended in Tris/HCl buffer(50 mM, pH 8.0; 150 mM NaCl) and it and the supernatantwere used as the insoluble and soluble fraction, respectively.An appropriate amount of each fraction was taken for theabove nitrilase assay. The nitrilases from G. moniliformis and
P. marneffei were assayed with 25 mM benzonitrile and thenitrilases from N. crassa and A. niger CBS 513.88 with25 mM phenylacetonitrile under the above conditions.
Km and Vmax values were determined with 1–25 mM ofsubstrates.
Temperature and pH optima were assayed at 15–60°Cand pH 4.0–12.0 (using 100 mM acetic acid/boric acid/phosphoric acid/NaOH buffers). The residual activities afterpre-incubation of enzymes at 25–60°C or pH 4.5–12.0 wereassayed at optimum temperature of each enzyme (43°C and30°C for nitrilases from G. moniliformis and P. marneffei,respectively; 38°C for other enzymes) and pH 8.0.
Analytical HPLC
Benzonitrile, its analogues, phenylacetonitrile, (R,S)-2-phenylpropionitrile, (R,S)-mandelonitrile, and their reactionproducts (acids, amides) were analyzed using a ChromolithFlash RP-18 (Merck, 25×4.6 mm) with a mobile phaseconsisting of CH3CN (10–25%) and H3PO4 (0.1%) in waterat 2 mL min−1 and 35°C. 4-Cyanopyridine and its reactionproducts were analyzed as described previously (Malandraet al. 2009); 2- and 3-cyanopyridine and their reactionproducts were analyzed in the same way.
Protein assay
Protein concentration was determined according to Bradford(1976) using Bradford Protein Assay (Bio-Rad Laboratories,USA) with bovine serum albumin as the standard.
SDS-PAGE
Proteins were analyzed by 12% sodium dodecyl sulfatepolyacrylamide gel electrophoresis (SDS-PAGE) (Laemmli1970) followed by Coomassie Brilliant Blue R-250staining.
Results
Variability of fungal nitrilases
Putative fungal nitrilases/cyanide hydratases available inthe GenBank can be classified into a number of sequenceclades (see supplementary Fig. S2). With the aim to exploitthe diversity of these enzymes, we selected four of them,which shared a relatively low homology degree. All ofthem were only distantly related (with less than 40% aminoacid identities) to the nitrilase from A. niger K10, for whichboth its amino acid sequence and its biochemical propertieshave been already known (Kaplan et al. 2011a). Thenitrilases from A. niger CBS 513.88, N. crassa OR74A,
Appl Microbiol Biotechnol (2012) 93:1553–1561 1555

and G. moniliformis exhibited less than 50% amino acididentities to each other (see supplementary Fig. S3 formultiple sequence aligment). The P. marneffei enzymeshared a higher identity to G. moniliformis than the otherenzymes (56%). The identity degree of all these fungalnitrilases to well-known bacterial nitrilases was approxi-mately 40%. The nitrilase from N. crassa is “relatedto aliphatic nitrilase” according to GenBank, and theenzymes from G. moniliformis and P. marneffei werepostulated to act on aromatic nitriles due to their similarityto characterized nitrilases from genus Fusarium (Vejvoda etal. 2010).
Preliminary whole-cell experiments confirmed theexpected substrate specificities, indicating that the substratepreference of the enzymes differed between the nitrilasefrom G. moniliformis on one side and nitrilases from N.crassa and A. niger on the other (Kaplan et al. 2011b). Thelatter enzymes acted preferentially on phenylacetonitrileunlike that from G. moniliformis and previously character-ized fungal nitrilases in general (Martínková et al. 2009),which exhibited their maximum activities for benzonitrileor 4-cyanopyridine. High relative activities of E. coli cellsexpressing the nitrilase from P. marneffei for benzonitrileand 4-cyanopyridine suggested that this enzyme was alsoan aromatic nitrilase (data not shown).
The enzyme studied previously by us in A. niger K10(Kaplan et al. 2011a) was more closely related to cyanidehydratases (with approximately 60–85% amino acid iden-tities) than to other fungal nitrilases studied here (maximumamino acid identity of approximately 38% with P. marneffeinitrilase). In compliance with this finding, it produced ahigh ratio of by-product amide, which is the sole product ofcyanide hydratase (i.e., formamide from HCN; O'Reilly andTurner 2003).
These enzymes served as a representative set of diversenitrilases from filamentous fungi for comparing theirheterologous expression in the presence and absence ofmolecular chaperones and for investigating their catalyticproperties.
Nitrilase in Aspergillus niger K10
It was hypothesized that the enzyme from A. niger K10 wasnot correctly folded in E. coli, as its specific activity forbenzonitrile decreased significantly and its substrate spec-ificity changed compared to the enzyme isolated from thenative organism (Kaplan et al. 2011a). MALDI-TOFanalysis showed that the recombinant enzyme retained aC-terminal 46-amino acid peptide which was cleaved in thenative organism (ibid.). It is possible that enzyme misfold-ing led to this missing posttranslational modification. Asimilar posttranslational modification in Rhodococcus rho-dochrous J1 was probably due to autocatalytic cleavage
(Thuku et al. 2007) and the misfolded enzyme from A.niger may have lost this function. However, molecularchaperones did not improve this enzyme's activity signifi-cantly (Kaplan et al. 2011a; this work—supplementaryTable S4).
In this work, the enzyme from A. niger K10 wasproduced from both the gene amplified from the nativeorganism and the synthetic gene optimized according to E.coli codon bias. However, the latter approach was lessefficient with an active nitrilase production an order ofmagnitude lower (about 20 vs. 200 UL−1 of culture). It ispossible that using the optimized gene led to a translationspeed that was too high, which hampered the folding of therecombinant product and could finally result in itsdegradation or aggregation. Regions that are comprised ofrare codons can delay peptide elongation and thus providetime for correct folding of the nascent protein. Indeed, theoptimization of codon frequency in heterologouslyexpressed genes was reported to lead to an enhancedproportion of misfolded and aggregated protein (for areview, see Sabate et al. 2010).
Nitrilases in Aspergillus niger CBS 513.88 and Neurosporacrassa OR74A
A screening of molecular chaperone effects on total nitrilaseproduction (supplementary Table S4) did not indicate anysignificant effect of chaperone co-expression in strainsexpressing the A. niger CBS 513.88 or N. crassa enzyme.Only in strains simultaneously expressing GroEL/ESchaperones and trigger factor was a slight increase innitrilase activity per liter of culture observed (by a factor of1.25 times). Activity measurement in the soluble andinsoluble cell fractions showed that the percentage ofnitrilase activity in the soluble fraction was also increasedin strains co-expressing these chaperones compared to thecontrols (from 53% to 69% and 65% to 77% in strainsexpressing nitrilases from N. crassa and A. niger, respec-tively). Therefore, these strains were used to purify theenzymes. A more significant increase in nitrilase productionwas, however, achieved by modifying the cultivationmethod in terms of the culture volume (200 vs. 10 mL),type of cultivation vessels (flasks vs. Falcon tubes), andinitial cultivation temperature (37°C vs. 25°C). Thus, thecells expressing nitrilases from N. crassa and A. nigeryielded up to 69,000 and 23,000 UL−1 of culture (6,700and 3,650 UL−1 of cell suspension of OD610=1), respec-tively, for phenylacetonitrile.
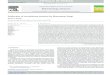
Purification of enzymes from N. crassa and A. niger tonear homogeneity (Fig. 1) was accomplished in two stepswith a 2.4 and threefold increase in specific activity,respectively, and approximately 50% yields. SDS-PAGEsuggested that traces of GroEL could co-purify with
1556 Appl Microbiol Biotechnol (2012) 93:1553–1561

nitrilase from A. niger. According to purification results(data not shown), the nitrilases in N. crassa and A. nigerformed approximately 42% and 34% of the total solubleprotein, respectively.
An examination of the substrate specificity of thepurified nitrilases confirmed the findings made with wholecells (Kaplan et al. 2011b); the enzymes from N. crassa andA. niger CBS 513.88 acted preferentially on phenylacetoni-trile. The purified enzyme from N. crassa exhibited anapproximately 1.6 times higher Vmax value for phenyl-acetonitrile than that from A. niger (Table 1). (R,S)-Mandelonitrile was also hydrolyzed at significant rates,but, compared to phenylacetonitrile, the Km values for thissubstrate were higher, especially in the enzyme from A.niger (more than three times; Table 1). The Km valuesdetermined with 2-phenylpropionitrile were the lowest, butthe Vmax values were 30–175 times lower than withphenylacetonitrile (ibid.). Benzonitrile and cyanopyridineswere transformed at approximately 100 times lower ratesthan phenylacetonitrile by both enzymes (data now shown).The enzymes formed almost no amide from the examinedsubstrates. The temperature optima of both enzymes wereapproximately 38°C and the pH optima at slightly alkalinevalues. The nitrilase from A. niger exhibited a broader pH
optimum (7.0–9.5) than the enzyme from N. crassa (8.0–8.5)(data not shown). The former enzyme was also more stableat alkaline pH values (Fig. 2a). Both enzymes were relativelystable at temperatures below 35°C and still retained 40–60%activity after 1-h preincubation at 40°C (Fig. 2b).
Nitrilases in Gibberella moniliformis and Penicilliummarneffei
The enzymes from G. moniliformis (Kaplan et al. 2011b)and P. marneffei were both expressed at lower levelscompared to the above arylaliphatic nitrilases (Fig. 1). Ascreening of strains harboring various chaperone plasmidsindicated that the activities of strains with the pGro7plasmid exhibited a nitrilase activity per liter of culturethat was six to seven times higher than that of the controls(supplementary Table S4). Other plasmids had no positiveeffect on the nitrilase activities. A more detailed examina-tion of this effect in larger culture volumes (200 vs. 10 mL;Table 2) gave a similar result in G. moniliformis, totalactivity increasing by a factor of approximately 6.8 inpresence of GroEL/ES. The strain expressing the enzymefrom P. marneffei exhibited an approximately 2.9-foldincrease in nitrilase activity on co-expression of GroEL/ES under the same conditions. This effect was mainly dueto increased specific activities of the cultures. In G.moniliformis, the specific activities of the culture brothand cell-free extract were a factor of 4.4 and 2.3 timeshigher than the controls, respectively. There was also asignificant increase in the proportion of activity in thesoluble fractions. Similar effects were observed in strainsproducing the nitrilase from P. marneffei; the coexpression ofGroEL/ES increased the specific activity of the culture brothand cell-free extract approximately 4.3 and 2.9 times,respectively. The increase in nitrilase solubility was evenmore significant than in the previous enzyme. In both cases,the nitrilases were purified from the strains with chaperones.
Both nitrilases co-purified with GroEL as indicated bySDS-PAGE (Fig. 1). The two-step purification of theenzyme from G. moniliformis resulted in an approximatelyninefold increase in the specific activity. Replacement ofSephacryl S-200 by Superdex 200 in the second step didnot improve the purity of the enzyme significantly asconcluded from SDS-PAGE (data not shown), while yield
Fig. 1 Expression in E. coli and purification of nitrilases fromNeurospora crassa OR74A, Aspergillus niger CBS 513.88, Gibberellamoniliformis, and Penicillium marneffei ATCC 18224. Lane 1 marker.Lanes 2, 4, 6, 8 cell-free extracts from strains expressing nitrilasesfrom N. crassa, A. niger, G. moniliformis, and P. marneffei,respectively. Lanes 3, 5, 7, 9 (partially) purified nitrilases from N.crassa, A. niger, G. moniliformis, and P. marneffei, respectively.Nitrilase from P. marneffei was purified by affinity chromatography.Other enzymes were purified in two steps (Q-Sepharose, Superdex200). Upper bands in lanes 5–9 indicate co-purification with GroEL
Table 1 Substrate specificity ofarylaliphatic nitrilases Substrate Aspergillus niger Neurospora crassa
Km (mM) Vmax (U mgprotein−1) Km (mM) Vmax (U mg−1 protein)
Phenylacetonitrile 3.4 10.6 2.0 17.5
(R,S)-Mandelonitrile 11.4 12.4 3.4 9.9
2-Phenylpropionitrile 0.80 0.35 1.3 0.10
Appl Microbiol Biotechnol (2012) 93:1553–1561 1557

was higher with Sephacryl S-200 (58%) than Superdex 200(36%). The nitrilase from P. marneffei was purifiedapproximately fourfold with 27.5% yield in one step. Thereason for the relatively low yield of the enzyme may havebeen its instability, which also caused a significant activityloss during storage at −80°C or 4°C. Therefore, effects oftemperature and pH on enzyme stability as well as thesubstrate specificity of this enzyme were examined withcell-free extracts (Fig. 2; Table 3).
The nitrilase from G. moniliformis exhibited the highestVmax and lowest Km values for benzonitrile, but phenyl-acetonitrile was transformed with much lower Vmax andhigher Km values (Table 3). 3- and 4-Cyanopyridine weregood substrates of the enzyme but 2-cyanopyridine was nottransformed at all probably due to steric hindrance. Thenitrilase from G. moniliformis was highly active over abroader pH range (7–10; data not shown) than thearylaliphatic nitrilases, and a similar observation was maderegarding pH stability (Fig. 2a). It also exhibited the highesttemperature optimum (approximately 43°C; data notshown) and thermostability (Fig. 2b) of the enzymesexamined.
In contrast, the temperature optimum of the nitrilasefrom P. marneffei was the lowest (30–35°C; data notshown) and the enzyme stability decreased more sharplyat temperatures over 35°C compared to the other enzymes(Fig. 2b). The enzyme was relatively stable over a pHregion of 6.5–8.5 (Fig. 2a). Its substrate specificity wasbroad (Table 3), comparable Vmax and Km values beingdetermined for benzonitrile and phenylacetonitrile. Theenzyme was also active with cyanopyridines including thesterically hindered 2-cyanopyridine but the high Km valuesof these compounds indicated their inefficient binding bythe enzyme. 4-Cyanopyridine was transformed by thisenzyme with the highest Vmax of all substrates tested.
The enzyme from P. marneffei did not produce anysignificant amount of amides. The production of amidefrom benzonitrile and chlorobenzonitriles by nitrilase fromG. moniliformis was low, not exceeding 6% in total product,but this enzyme produced 42% and 54% of amide fromphenylacetonitrile and 4-cyanopyridine, respectively.
Discussion
The role of molecular chaperones consists of assistingnewly synthetized polypeptide folding (Sabate et al. 2010).The bacterial chaperone network involves the Hsp70(DnaK, DnaJ, GrpE) and GroEL/ES systems and TF (Gupta
Fig. 2 pH (a) and temperature (b) stabilities of purified nitrilasesfrom Neurospora crassa OR74A (filled squares), Aspergillus nigerCBS 513.88 (filled diamonds), and Gibberella moniliformis (filledtriangles) and cell-free extract of cells producing nitrilase fromPenicillium marneffei ATCC 18224 (filled circles) (0.2, 0.2, 0.3 and1.6 mg protein per milliliter, respectively). a Enzymes were pre-incubated at pH 4.5–12.0 and room temperature for 2 h. b Enzymeswere pre-incubated at 25–60°C and pH 8.0 for 1 h. Specific activites,which were determined under standard conditions prior to preincuba-tion of the enzymes, were taken as 100%
Table 2 Effect of GroEL/ES chaperone co-expression on nitrilase production and solubility in Escherichia coli strains harboring nit genes fromGibberella moniliformis and Penicillium marneffei ATCC 18224
Gene source Chaperone Total activity[U L−1 culture]
Specific activity[U L−1 cultureof OD610=1]
Specific activity[U mg−1 cell-freeextract]
Percentage (%)of activity in
soluble fraction insoluble fraction
Gibberella moniliformis – 233±13 76±7 0.28±0.12 57±3 43±3
GroEL/ES 1584±145 335±87 0.64±0.03 70±4 30±4
Penicillium marneffei – 48±3 7±0.2 0.024±0.004 31±3 69±3
GroEL/ES 141±22 30±2 0.070±0.014 52±8 48±8
Data are means of three independent experiments
1558 Appl Microbiol Biotechnol (2012) 93:1553–1561

et al. 2010). Though co-expression of these chaperones wasoften used as a means to facilitate overexpression ofeukaryotic proteins in E. coli, the outcome of this methodis hard to predict (ibid.). Plasmids containing genes codingfor components of the above chaperone network (individ-ually or in combination) were designed (Nishihara et al.2000) and became commercially available.
The importance of chaperones in nitrilase folding wasindicated by experiments with nitrilases, where they co-purified with some of these molecules. The enzymes fromwild-type producing strains belonging to Pseudomonasfluorescens and Bacillus pallidus co-purified with proteinsclosely related to GroEL (Layh et al. 1998; Almatawah etal. 1999). The recently described nitrilase from A. niger co-purified with hsp60, the eukaryotic homologue of GroEL(Kaplan et al. 2006a).
When co-expressing GroEL/ES in E. coli with analiphatic nitrilase from Comamonas testosteroni, the spe-cific activity of the cells increased fivefold and the solubleenzyme fraction from 10% to 58% (Lévy-Schil et al. 1995).The same chaperonin system also exhibited a positive effecton the functional expression in E. coli of another enzyme ofthe nitrilase superfamily, D-carbamoylase (Chao et al. 2000;Sareen et al. 2001). As suggested in a recent study of tenprotein kinases (Haacke et al. 2009), the effect ofchaperones may be case-specific. Similar conclusions weremade in a study of endostatin, ORP150 protein and humanlysozyme (Nishihara et al. 2000). Therefore, we screenedfor chaperone effects using a large array of chaperone–nitrilase combinations. The chaperone effects were actuallydifferent in strains expressing different nitrilases. Signifi-cant benefits of GroEL/ES, which binds to unfoldedproteins and creates a protected environment for theirfolding (Lund 2009), were observed in aromatic nitrilasesfrom Gibberella and Penicillium but not in arylacetonitri-lases. This effect could be also dependent on the nitrilaseexpression levels, which were higher in the latter enzymes.
Chaperone-assisted overexpression facilitated purifica-tion of the enzymes by increasing the specific activity of
the cells and the enzyme ratio in the soluble fractions. Thelatter effect was also significant in strains expressingarylaliphatic nitrilases from N. crassa and A. niger with acombination of GroEL/ES and TF chaperones. Therefore,the corresponding strains were used for enzyme purifica-tion. As a result, four new nitrilases (two aromatic and twoarylaliphatic) were purified or partially purified in thisstudy. The nitrilases from G. moniliformis and P. marneffeiwere not obtained as homogeneous proteins, as they co-purified with GroEL as did a number of the wild-typenitrilases. This co-purification could indicate incompleteenzyme folding; a similar phenomenon was observed withprotein kinases and the GroEL and DnaJ chaperones(Haacke et al. 2009).
Aromatic nitrilases occur frequently in bacteria, primarilyrhodococci (for a review, see O'Reilly and Turner 2003). Allof the fungal nitrilases purified previously also belonged tothis group of enzymes (Harper 1977; Goldlust and Bohak1989; Vejvoda et al. 2008, 2010). Comparison of the kineticdata of the nitrilase from G. moniliformis for (hetero)aromatic nitriles and phenylacetonitrile suggested this en-zyme could be also classified as an aromatic nitrilase.Nevertheless, the Vmax value of this enzyme for benzonitrilewas more than 10 times lower than the specific activities ofmost of the nitrilases from the related taxon Fusarium(Goldlust and Bohak 1989; Vejvoda et al. 2008, 2010). Thiscould be at least partly caused by the co-purification ofGroEL, but could be also due to partial misfolding. On theother hand, the recombinant enzyme and the wild-typeenzymes from related species exhibited similar substratespecificities and optimum reaction conditions.
A nitrilase activity was recently reported in Penicilliummulticolor by ourselves (Kaplan et al. 2006b), but theenzyme was not purified. No other nitrilases were describedin the Penicillium genus as far as we know. The advantageof the new enzyme from P. marneffei is its relatively widesubstrate specificity. Similarly to the nitrilase in P. multi-color it exhibited, however, a lower temperature stabilitythan most other nitrilases.
Table 3 Substrate specificity ofaromatic nitrilases
aPartially purified enzymebCell-free extractcNo activity
Substrate Gibberella moniliformisa Penicillium marneffeib
Km (mM) Vmax (U mgprotein−1) Km (mM) Vmax (U mgprotein
−1)
Benzonitrile 0.41 9.7 7.9 0.16
3-Chlorobenzonitrile 0.93 6.6 11.1 1.9
4-Chlorobenzonitrile 1.3 3.9 2.6 0.13
2-Cyanopyridine –c –c 11.7 0.46
3-Cyanopyridine 3.2 4.2 34.4 0.21
4-Cyanopyridine 2.2 7.3 15.4 7.3
Phenylacetonitrile 0.75 1.2 9.1 0.10
Appl Microbiol Biotechnol (2012) 93:1553–1561 1559

The substrate specificity of the enzymes from N. crassaand A. niger indicated that these enzymes were arylaceto-nitrilases. This category of nitrilases has significant indus-trial potential in the production of enantiopure carboxylicacids and amides (e.g., Fernandes et al. 2006; Rustler et al.2008; Banerjee et al. 2009; Sosedov et al. 2009). They haveso far been reported in the bacteria of the Alcaligenes (forreviews, see O'Reilly and Turner 2003; Thuku et al. 2009),Pseudomonas (Kiziak et al. 2005), or Halomonas (Chmuraet al. 2008) genera. The nitrilase from P. fluorescens is oneof the best described arylacetonitrilases (Kiziak et al. 2005,2007; Kiziak and Stolz 2009). The enzymes from Alcali-genes faecalis or P. fluorescens (GenBank AAW79573) onlyshared an approximately 40% amino acid sequence simi-larity with the new arylacetonitrilases from fungi. However,both of these enzymes showed similar substrate specificitiesas the bacterial arylacetonitrilases.
In conclusion, four new nitrilases were purified andcharacterized in heterologous producers expressing fungalnit genes. Two of the enzymes (from A. niger and N.crassa) were expressed at high levels, yielding 23,000 and69,000 units per liter of culture, respectively. The highproduction and straightforward purification of theseenzymes enabled tens of milligrams of them to be obtainedfrom 1 L of E. coli cultures. These new arylacetonitrilasescan be useful in the production of mandelic acid frommandelonitrile, or other carboxylic acids substituted at theα-position. Further studies should demonstrate the substraterange, enantioselectivity, chemoselectivity, and possibleapplication areas of these enzymes.
Acknowledgments The authors wish to thank Hynek Mrazek MSc.for his technical help with nitrilase purification. Financial support viaprojects P504/11/0394, 305/09/H008 (Czech Science Foundation),IAA500200708 (Grant Agency of the Academy of Sciences of theCzech Republic), LC06010, OC09046 (Ministry of Education of theCzech Republic), COST/ESF CM0701 (STSM fellowships COST-STSM-CM0701-4765 and −4766 to A. Malandra) and InstitutionalResearch Concept AV0Z50200510 (Institute of Microbiology) isgratefully acknowledged.
References
Almatawah QA, Cramp R, Cowan DA (1999) Characterization of aninducible nitrilase from a thermophilic bacillus. Extremophiles3:283–291
Altschul SF, Madden TL, Schäffer AA, Zhang JH, Zhang Z, Miller W,Lipman DJ (1997) Gapped BLAST and PSI-BLAST: a newgeneration of protein database search programs. Nucleic AcidsRes 25:3389–3402
Banerjee A, Dubey S, Kaul P, Barse B, Piotrowski M, Banerjee UC(2009) Enantioselective nitrilase from Pseudomonas putida:cloning, heterologous expression, and bioreactor studies. MolBiotechnol 41:35–41
Bradford MM (1976) A rapid and sensitive method for thequantitation of microgram quantities of protein utilizing
the principle of protein-dye binding. Anal Biochem 72:248–254
Chao YP, Chiang CJ, Lo TE, Fu H (2000) Overproduction of D-hydantoinase and carbamoylase in a soluble form in Escherichiacoli. Appl Microbiol Biotechnol 54:348–353
Chmura A, Shapovalova AA, van Pelt S, van Rantwijk F, Tourova TP,Muyzer G, Sorokin DYu (2008) Utilization of arylaliphaticnitriles by haloalkaliphilic Halomonas nitrilicus sp. nov. isolatedfrom soda soils. Appl Microbiol Biotechnol 81:371–378
Fernandes BCM, Mateo C, Kiziak C, Chmura A, Wacker J, vanRantwijk F, Stolz A, Sheldon RA (2006) Nitrile hydrataseactivity of a recombinant nitrilase. Adv Synth Catal 348:2597–2603
Goldlust A, Bohak Z (1989) Induction, purification, and characteriza-tion of the nitrilase of Fusarium oxysporum f. sp. melonis.Biotechnol Appl Biochem 11:581–601
Gupta R, Lakshmipathy SK, Chang HC, Etchells SA, Hartl FU (2010)Trigger factor lacking the PPIase domain can enhance the foldingof eukaryotic multi-domain proteins in Escherichia coli. FEBSLett 584:3620–3624
Haacke A, Fendrich G, Ramage P, Geiser M (2009) Chaperone over-expression in Escherichia coli: apparent increased yields ofsoluble recombinant protein kinases are due mainly to solubleaggregates. Protein Expres Purif 64:185–193
Harper DB (1977) Fungal degradation of aromatic nitriles. Enzymologyof C-N cleavage by Fusarium solani. Biochem J 167:685–692
Kaplan O, Vejvoda V, Plíhal O, Pompach P, Kavan D, Bojarová P,Bezouška K, Macková M, Cantarella M, Jirků V, Křen V,Martínková L (2006a) Purification and characterization of anitrilase from Aspergillus niger K10. Appl Microbiol Biotechnol73:567–575
Kaplan O, Nikolaou K, Pišvejcová A, Martínková L (2006b)Hydrolysis of nitriles and amides by filamentous fungi. EnzymeMicrob Technol 38:260–264
Kaplan O, Bezouška K, Plíhal O, Ettrich R, Kulik N, Vaněk O, KavanD, Benada O, Malandra A, Šveda O, Veselá AB, Rinágelová A,Slámová K, Cantarella M, Felsberg J, Dušková J, Dohnálek J,Kotik M, Křen V, Martínková L (2011a) Heterologous expres-sion, purification and characterization of nitrilase from Aspergil-lus niger K10. BMC Biotechnol 11:2
Kaplan O, Bezouška K, Malandra A, Veselá AB, Petříčková A,Felsberg J, Rinágelová A, Křen V, Martínková L (2011b) Genomemining for the discovery of new nitrilases in filamentous fungi.Biotechnol Lett 33:309–312
Kato Y, Ooi R, Asano Y (2000) Distribution of aldoxime dehydratasein microorganisms. Appl Environ Microbiol 66:2290–2296
Kiziak C, Stolz A (2009) Identification of amino acid residuesresponsible for the enantioselectivity and amide formationcapacity of the arylacetonitrilase from Pseudomonas fluorescensEBC191. Appl Environ Microbiol 75:5592–5599
Kiziak C, Conradt D, Stolz A, Mattes R, Klein J (2005) Nitrilase fromPseudomonas fluorescens EBC191: cloning and heterologousexpression of the gene and biochemical characterization of therecombinant enzyme. Microbiology 151:3639–3648
Kiziak C, Klein J, Stolz A (2007) Influence of different carboxy-terminal mutations on the substrate-, reaction- and enantiospeci-ficity of the arylacetonitrilase from Pseudomonas fluorescensEBC191. Protein Eng Des Sel 20:385–396
Laemmli UK (1970) Cleavage of the structural proteins duringassembly of the head of bacteriophage T4. Nature 227:680–685
Layh N, Parratt J, Willetts A (1998) Characterization and partialpurification of an enantioselective arylacetonitrilase fromPseudomonas fluorescens DSM 7155. J Mol Catal B-Enz5:467–474
Lévy-Schil S, Soubriere F, Crutz-Le Coq AM, Faucher D, Crouzet J,Pétré D (1995) Aliphatic nitrilase from a soil-isolated Comamo-
1560 Appl Microbiol Biotechnol (2012) 93:1553–1561

nas testosteroni sp.: gene cloning and overexpression, purifica-tion and primary structure. Gene 161:15–20
Lund PA (2009) Multiple chaperonins in bacteria—why so many?FEMS Microbiol Rev 33:785–800
Malandra A, CantarellaM,Kaplan O, Vejvoda V, Uhnáková B, ŠtěpánkováB, Kubáč D, Martínková L (2009) Continuous hydrolysis of 4-cyanopyridine by nitrilases from Fusarium solani and Aspergillusniger K10. Appl Microbiol Biotechnol 85:277–284
Martínková L, Vejvoda V, Kaplan O, Kubáč D, Malandra A, CantarellaM, Bezouška K, Křen V (2009) Fungal nitrilases as biocatalysts:recent developments. Biotechnol Adv 27:661–670
O'Reilly C, Turner PD (2003) The nitrilase family of CN hydrolysingenzymes—a comparative study. J Appl Microbiol 95:1161–1174
Osswald S, Wajant H, Effenberger F (2002) Characterization andsynthetic applications of recombinant AtNIT1 from Arabidopsisthaliana. Eur J Biochem 269:680–687
Robertson DE, Chaplin JA, DeSantis G, Podar M, Madden M, Chi E,Richardson T, Milan A, Miller M, Weiner DP, Wong K, McQuaidJ, Farwell B, Preston LA, Tan X, Snead MA, Keller M, MathurE, Kretz PL, Burk MJ, Short JM (2004) Exploring nitrilasesequence space for enantioselective catalysis. Appl EnvironMicrobiol 70:2429–2436
Rustler S, Motejadded H, Altenbuchner J, Stolz A (2008) Simulta-neous expression of an arylacetonitrilase from Pseudomonasfluorescens and a (S)-oxynitrilase from Manihot esculenta inPichia pastoris for the synthesis of (S)-mandelic acid. ApplMicrobiol Biotechnol 80:87–97
Sabate R, de Groot NS, Ventura S (2010) Protein folding andaggregation in bacteria. Cell Mol Life Sci 67:2695–2715
Sareen D, Sharma R, Vohra RM (2001) Chaperone-assisted over-expression of an active D-carbamoylase from Agrobacteriumtumefaciens AM 10. Protein Expres Purif 23:374–379
Sosedov O, Matzer K, Bürger S, Kiziak C, Baum S, Altenbuchner J,Chmura A, van Rantwijk F, Stolz A (2009) Construction ofrecombinant Escherichia coli catalysts which simultaneouslyexpress an (S)-oxynitrilase and different nitrilase variants for thesynthesis of (S)-mandelic acid and (S)-mandelic amide frombenzaldehyde and cyanide. Adv Synth Catal 351:1531–1538
Thompson JD, Higgins DG, Gibson TJ (1994) CLUSTAL W:Improving the sensitivity of progressive multiple sequencealignment through sequence weighting, position-specific gappenalties and weight matrix choice. Nucleic Acids Res22:4673–4680
Thuku RN, Weber BW, Varsani A, Sewell BT (2007) Post-translationalcleavage of recombinantly expressed nitrilase from Rhodococcusrhodochrous J1 yields a stable, active helical form. FEBS J274:2099–2108
Thuku RN, Brady D, Benedik MJ, Sewell BT (2009) Microbialnitrilases: versatile, spiral forming, industrial enzymes. J ApplMicrobiol 106:703–727
Vejvoda V, Kaplan O, Bezouška K, Pompach P, Šulc M, Cantarella M,Benada O, Uhnáková B, Rinágelová A, Lutz-Wahl S, Fischer L,Křen V, Martínková L (2008) Purification and characterization ofa nitrilase from Fusarium solani O1. J Mol Catal B-Enz 50:99–106
Vejvoda V, Kubáč D, Davidová A, Kaplan O, Šulc M, Šveda O,Chaloupková R, Martínková L (2010) Purification and charac-terization of nitrilase from Fusarium solani IMI196840. ProcBiochem 45:1115–1120
Wu SJ, Fogiel AJ, Petrillo KL, Hann EC, Mersinger LJ, DiCosimo R,O'Keefe DP, Ben-Bassat A, Payne MS (2007) Protein engineeringof Acidovorax facilis 72W nitrilase for bioprocess development.Biotechnol Bioeng 97:689–693
Wu SJ, Fogiel AJ, Petrillo KL, Jackson RE, Parker KN, DiCosimo R,Ben-Bassat A, O'Keefe DP, Payne MS (2008) Protein engineeringof nitrilase for chemoenzymatic production of glycolic acid.Biotechnol Bioeng 99:717–720
Appl Microbiol Biotechnol (2012) 93:1553–1561 1561
Nishihara K, Kanemori M, Yanagi H, Yura T (2000) Overexpression oftrigger factor prevents aggregation of recombinant proteins inEscherichia coli. Appl Environ Microbiol 66:884–889