Embed Size (px)
Citation preview

Psychoactive drugs against effects of stress in infectious and non-infectious viral diseases
S. Novío1, M.J. Núñez-Iglesias1, M. Freire-Garabal1 1 Lennart Levi Stress and Neuroimmunology Laboratory, Department of Pharmacology, School of Medicine, C/ San
Francisco s/n, 15782 Santiago de Compostela, Spain
Stress qualitatively and quantitatively impairs immune function via the activation of main two neuroendocrine pathways, the hypothalamic-pituitary-adrenal (HPA) axis and the sympathetic branch of the autonomic nervous system (ANS). Stress-induced immune impairments appear to be large enough to alter the course of viral diseases, including enhancing the risk for more severe viral infections, delaying the healing time, reducing the immune response to antiviral vaccines, and reactivating latent herpesviruses (e.g. herpes simplex virus) and latent tumor promoting viruses (e.g. Epstein-Barr virus). Nowadays, physicians emphasize the importance of accurately identifying and effectively managing with psychopharmacological interventions stressful life experiences, since psychoactive drugs such as serotonergic antidepressant reduce the suppressive effects of stress on immune function and improve the progression of medical diseases.
Keywords anxiolytic; antidepressant; psychological stress; serotonin; viral disease
1. Psychoneuroimmunoendocrinology
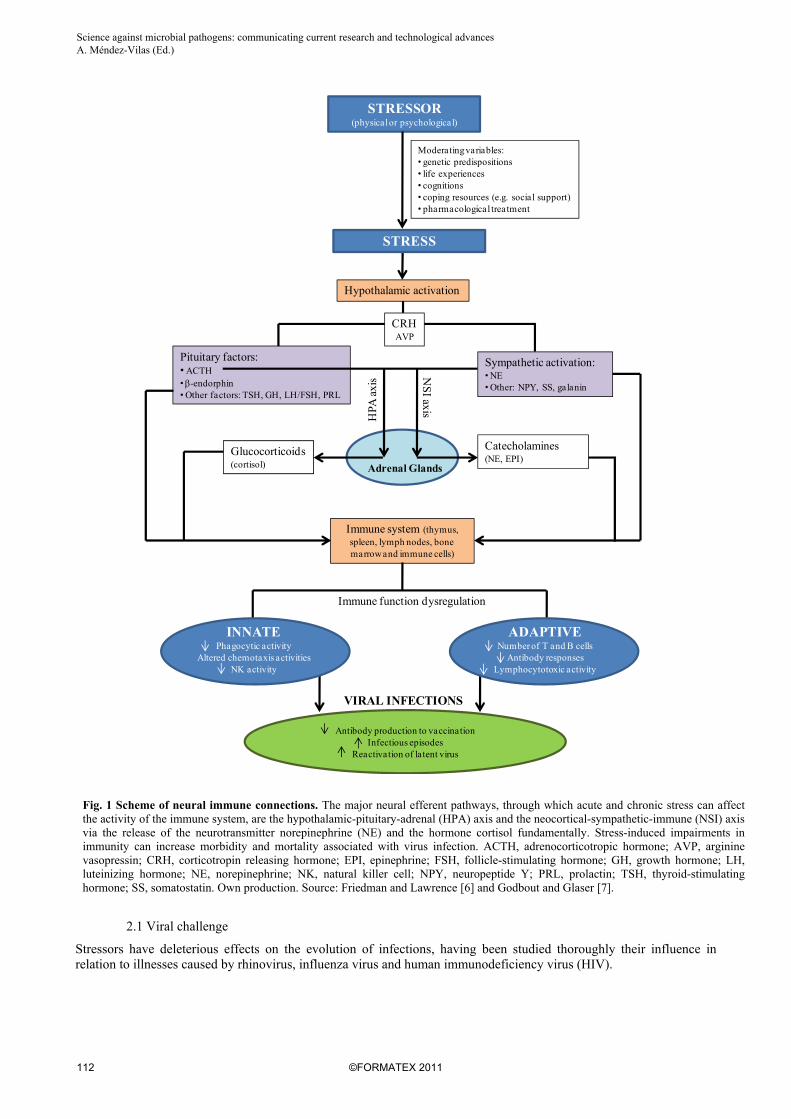
Stress is a state of altered physiological homeostasis, which influences an individual´s ability to cope with life events. Psychological stress, like chemical and physical stressors, affect host defenses comprising neuronal, endocrine, and immune reactions. This complex network of bi-directional signals plays a vital role in determining the outcome of the stress response, since when the balance among the three systems is altered, the risk of disease is increased [1]. Popular conception has it that stress impairs the immune system, however the duration and the course of the stress are the pivotal factors that determine the nature of the stress-induced immune change and any health-related outcome [2, 3]. In this way, acute or transient stress is suggested facilitating some aspects of immune function. For example, individuals exposed to a brief experimental stressor (videotaped speech) [4] exhibit transient increases in natural killer-cell (NK) numbers and function. More intense and/or long-lived stressors, on the other hand, typically have a deleterious effect on immunity. Pike et al. [5] have demonstrated that human subjects with histories of prolonged life stress exhibit reduced NK activity during and after a laboratory speech stress paradigm. Fig. 1 shows as stress impairs both natural and specific immune responses predisposing the host to more severe virus infection [6, 7]. Changes in the absolute number of lymphocytes, T-lymphocytes, T-helper and T-suppressor cells have been reported [8, 9]. Stress also interferes with several immune responses such as splenic cytotoxic activities, mediated by NK cells and cytotoxic T lymphocytes [10], the activity of phagocytosis [11, 12], the delayed type hypersensitivity (DTH) response [9, 13, 14], the blastogenic response of spleen lymphoid cells [8, 9] and T-dependent antibody responses [15]. Research into the mechanisms by which stressors are translated into impaired immune function and vulnerability to disease has focused primarily on two pathways: the hypothalamic-pituitary-adrenal (HPA) axis and the sympathetic branch of the autonomic nervous system (ANS). Whereas increased sympathetic adrenal activity appears to play a major role in immune changes observed after acute stress, HPA axis-activity together with sympathetic mechanisms are mainly responsible for the inhibition of cellular and humoral immune responses following chronic stress exposure [2]. The importance of these systems is so high that numerous studies demonstrate that maladaptive neuroendocrine hyper- or hypoactive responses of the HPA axis or the sympathetic nervous system (SNS) to stress can function as risk factors of specific diseases, such as viral infections. This is illustrated in studies showing that stress (i) increases severity, duration and susceptibility to infectious viral diseases; (ii) contributes to impaired responses to virus vaccines; (iii) promotes reactivation of latent viruses, and; (iv) may even be a cofactor for tumor development and progression.
2. Stress and infectious viral diseases
A number of paradigms have been utilized to examine the effects of stress on infectious viral disease outcomes. The most rigorous of these have involved viral challenge, response to vaccinations, and a focus on reactivation of latent viruses.
111©FORMATEX 2011
Science against microbial pathogens: communicating current research and technological advances A. Méndez-Vilas (Ed.)_______________________________________________________________________________
Antibody production to vaccinationInfectious episodes
Reactivation of latent virus
Moderating variables:• genetic predispositions• life experiences• cognitions• coping resources (e.g. social support)• pharmacological treatment
STRESSOR (physicalor psychological)
Pituitary factors:• ACTH• -endorphin• Other factors: TSH, GH, LH/FSH, PRL
STRESS
CRHAVP
Hypothalamic activation
Sympathetic activation:• NE• Other: NPY, SS, galanin
Adrenal Glands
Catecholamines (NE, EPI)
HPA
axi
s NS
I axis
Immune system (thymus, spleen, lymph nodes, bonemarrowand immune cells)
Immune function dysregulation
Glucocorticoids(cortisol)
INNATEPhagocytic activity
Altered chemotaxis activitiesNK activity
ADAPTIVENumber of T and B cells
Antibody responsesLymphocytotoxic activity
VIRAL INFECTIONS
Fig. 1 Scheme of neural immune connections. The major neural efferent pathways, through which acute and chronic stress can affect the activity of the immune system, are the hypothalamic-pituitary-adrenal (HPA) axis and the neocortical-sympathetic-immune (NSI) axis via the release of the neurotransmitter norepinephrine (NE) and the hormone cortisol fundamentally. Stress-induced impairments in immunity can increase morbidity and mortality associated with virus infection. ACTH, adrenocorticotropic hormone; AVP, arginine vasopressin; CRH, corticotropin releasing hormone; EPI, epinephrine; FSH, follicle-stimulating hormone; GH, growth hormone; LH, luteinizing hormone; NE, norepinephrine; NK, natural killer cell; NPY, neuropeptide Y; PRL, prolactin; TSH, thyroid-stimulating hormone; SS, somatostatin. Own production. Source: Friedman and Lawrence [6] and Godbout and Glaser [7].
2.1 Viral challenge
Stressors have deleterious effects on the evolution of infections, having been studied thoroughly their influence in relation to illnesses caused by rhinovirus, influenza virus and human immunodeficiency virus (HIV).
112 ©FORMATEX 2011
Science against microbial pathogens: communicating current research and technological advances A. Méndez-Vilas (Ed.)______________________________________________________________________________

2.1.1 Rhinovirus
About one third of the people with confirmed viral infection do not show evidence of cold symptoms, being their development related to prior life events. Cohen et al. [16] have observed that individuals with higher perceived stress scores have an increased risk of developing acute infectious respiratory illness caused by rhinovirus (rhinovirus type 2, 9, or 14, respiratory syncytial virus, and coronavirus type 229E). On the other hand, stressors also influence the duration and severity of viral diseases. Higher psychological stress is associated with a greater expression of illness (measured by more severe symptoms and greater mucus production) [17] and a delay in the recovery from these infections [18]. Little is known about the physiological mediators of these effects since attempts to link hormonal and immunologic processes to viral outcomes in this paradigm have been largely unsuccessful. However, animal studies suggest three major pathways linking stress exposure to natural resistance to influenza viral infection: the response of pro-inflammatory cytokines, -chemokines, and NK cells. In addition, the adaptive immune response, involving antigen-specific T cell activation, also plays a role in this process [19]. These may be important targets for future influenza viral challenge studies in humans.
2.1.2 Influenza virus
Influenza virus provides a continuous threat of epidemic disease in man. Cellular and humoral immunity plays an important role in the clearing of influenza virus and in the recovery phase of infection [20, 21]. In fact, the neutralizing anti- hemagglutinin antibody has a protective role, preventing the entry of the viruses into susceptible cells [22]. However, there is evidence that influenza virus infection affects several immune functions such as NK activity, DTH response and IL-2 production [22]. This injury to elements of the immunological apparatus increases the vulnerability of host to infection. The adverse effects of stress on infection by influenza virus have been corroborated by numerous studies. Animal researches show that stress exacerbates influenza virus PR-8/34 infection. Freire-Garabal et al. [23, 24] have observed significantly earlier times of death as well as an increase in lung virus titers and permeability index amongst stressed mice in comparison with unstressed controls. Some researches in humans suggest similar findings. An analysis of the incidence of colds in volunteers exposed experimentally to influenza virus showed more evidence of increased nasal secretion after infection between people with higher scores of psychoneurotic symptoms [25]. Also, Clover et al. [26] found that infection with influenza B virus was associated with family cohesion and adaptability, suggesting that family dysfunction may be related to altered immune response. Studies reported in the last decade have demonstrated considerable individual variation in susceptibility to disease. These individual differences may be at least in part due to predisposition to illness [27]. Although the origin and nature of these predispositions have not been fully explained, it is known that stress in the first years of life alters the severity and kinetics of the response to influenza viral infection [28], suggesting that long lasting effects of negative life events on health and disease may be the basis for the individual differences in host susceptibility to infection. Thus, although influenza infection is usually self-limiting and does not require any special treatment, in some individuals other symptoms and complications such as viral pneumonia may develop [29].
2.1.3 HIV
HIV-infected people display physiological changes consistent with elevated resting levels of cortisol [30] and accelerated immune decline [31]. Stressful life events, depression and psychological and emotional reactions associated with negative views of the self, stigmatization or rejection are associated with clinical disease course in HIV-infected people. It is supported by studies which demonstrate that psychological stress in HIV-positive patients: a) predicates significant declines in CD4+ and CD8+ lymphocyte numbers [32, 33]; b) is associated with a more rapid progression into AIDS and an acceleration of HIV replication mediated by sympathetic activation [33-35] and; c) predicts onset of HIV-related symptoms and mortality, as well as a weaker response to antiviral therapy in terms of HIV viral load [34]. Importantly, behavioral interventions (such as social support, spirituality, and stress management techniques) designed to alleviate the distress associated with HIV infection are effective in improving the immunological and virologic indices of disease course [36]. For example, cognitive behavioral stress management intervention (sometimes combined with coping skills training) is associated with decreased cortisol [37], better immunologic control of viral pathogens [38], increases in markers of immune system reconstitution [39], and decreases in HIV viral RNA in peripheral blood up to 15 months after intervention [40]. HPA axis and SNS have been invoked as potential mediators of effects of psychological factors and HIV progression. With regard to HPA axis activity, in vitro studies have shown that corticosteroids can enhance HIV-1 replication [41] and prolong viral gene expression [42]. Moreover, Leserman et al. [31] have demonstrated that cortisol levels predict indicators of HIV progression in vivo. On the other hand, it is known that SNS neurons innervate lymphoid tissues that can become infected with HIV [43] and that norepinephrine (NE) can enhance HIV replication in vitro, demonstrating a dose–response relationship between amount of NE and replication rate [33, 35].
113©FORMATEX 2011
Science against microbial pathogens: communicating current research and technological advances A. Méndez-Vilas (Ed.)_______________________________________________________________________________

2.2 Response to viral vaccinations
Psychological stress can modulate the immune response to several antiviral vaccines, such as hepatitis B virus [44], rubella virus [45], meningitis virus [46], and influenza virus [47-49], down-regulating both virus-specific antibody responses and T-cell responses to antigens. In support of this relationship, Glaser et al. [44] have demonstrated that stress influences the ability to generate an antibody response to the HBsAg recombinant vaccine. Thus, those people who are more anxious and more stressed show a delay in production of antibodies, lower antibody titers to the Hep B vaccine after seroconversion, and decreased virus-specific T-cell response to a Hep B surface antigen. In this way, it is also noteworthy to mention that when these anxious or stressed people receive sessions of cognitive behavioral therapy their immune response to the vaccine is stronger. Host responses to vaccinations are good indicators of how a person would respond to a pathogen. Individuals who have attenuated immune responses, for example against influenza virus vaccination, also experience higher rates of clinical illness and longer clinical episodes [50]. These impaired immune responses to influenza virus vaccination are paralleled in stress studies. For instance, caregivers of a demented spouse, who have elevated concentrations of salivary cortisol that are correlated with higher perceived stress scores [49], show dramatic deficits in both cellular and humoral immune responses to an influenza virus vaccine compared with noncaregivers [47, 48]. On the other hand, several studies note that elevated IL-6 serum levels, which are associated with mental health complications such as depression and cognitive impairment [51], have been found after influenza virus vaccination in older adults [52]. This is of great medical concern since normal aging has been associated with changes in all cellular components of the immune system which may make these individuals more susceptible to stress-associated decreases in cellular immunity [53]. Taken together, these findings suggest that since individuals who are more stressed and more anxious seroconvert later, they might also be slower to develop an antibody response to other pathogens; thus they could be at greater risk for developing more severe infectious diseases.
2.3 Reactivation of latent viruses
There is a large literature suggesting that psychological stress is a key factor to the appearance, development, duration, severity and recurrence of herpesvirus infections (herpes simplex virus type 1 and 2 [HSV-1 and HSV-2] [54, 55], Epstein–Barr virus [EBV] [56], herpes zoster virus [VZV] [57] and, cytomegalovirus [CMV]) [58]. However, no all stressors produce the same health outcomes. For example, whereas persistent stressors are associated with the frequency of genital herpes recurrences, single or acute stressors do not [55]. The loss of immunological control over the expression/replication of latent herpesviruses is associated with modulation of the cellular immune response. When cellular immunity is compromised (e.g. in patients with AIDS), one or more herpesviruses can be reactivated, which frequently results in a severe infection and even death. For instance, psychologically stressful life events are associated with increased incidence of VZV infections in older individuals [59]. In the same way that, Glaser et al. [60] have observed that stressed-people have significantly higher HSV-1 IgG antibody titers. Increases in antibody titers to a latent herpesvirus reflect a down-regulation of the virus-specific memory T-cell response to the virus and the ability to control the steady-state expression of the latent virus. Although the mechanism of how latent viruses are reactivated in vivo is not well understood, several authors have suggested that psychological stress can produce a shift in the Th1/Th2 cytokine balance towards a Th2 response [61]. This imbalance favors virus induced pathogenesis and increases the susceptibility to infections, because it down-regulates the production of cytokines important for the T cell response to the virus [62]. According to the two previous mentioned sections, psychosocial interventions also contribute to decrease the frequency of recurrences of herpetic infections [63]. Collectively, these findings support the notion that stress-induced changes in the cellular immune response are associated with the reactivation of latent herpesviruses, being these deleterious effects of stressors precipitated by psychiatric illnesses, life events, and upsets.
3. Stress and non-infectious viral diseases
3.1 Tumor development and progression
The literature on stress and cancer is not extensive, and although there are studies suggesting that stress may play a role in tumor initiation, growth, progression and survival [64-66], well designed studies that do not demonstrate this relation have also been carried out [67]. One possible connection between stress and cancer development is the reactivation of latent tumor promoting viruses. EBV is a human tumor virus implicated in several malignant diseases including nasopharyngeal carcinoma, Burkitt´s lymphoma (BL), non-Hodgkin´s lymphoma, and posttransplant lymphomas [68]. Taking into account this, it is plausible that there is a relationship between stress and EBV-associated tumors. Another example should be the human papilomavirus (HPV). Antoni et al. [36] have observed that stress and pessimism are predictive factors of greater severity of HPV-associated cervical intraepithelial neoplasia, a precursor to cervical cancer.
114 ©FORMATEX 2011
Science against microbial pathogens: communicating current research and technological advances A. Méndez-Vilas (Ed.)______________________________________________________________________________

The effects of stress models on the induction and growth of virus-associated tumors in animals have also been widely studied. It was found that mild stress (only that associated with daily injection of placebo) enhances the incidence and shortens the latent period for mammary tumors in C3H/He mice infected with the MTV mammary virus [69-71]. Psychological stressors induce cell damage which activates cellular processes of repair and apoptosis. These processes are critical since faulty DNA repair is associated with increased incidence of cancer [72] and, the survival of abnormal cells can lead to growth transformation and malignantly transformed tumor cells [73]. Mechanisms linking the physiological changes induced by stress and the observed changes in DNA repair and apoptosis are not known, however since the HPA axis and ANS are activated by stress, it is possible that one or more stress hormones could mediate these responses through signaling pathways, perhaps involving NF-kB [74]. Likewise, it is noteworthy to note the played role by stress-related immunosuppression. So, several authors have managed to suggest that natural or experimental stressors modulate the evolution of malignancies in humans as well as other animals [75], suppressing lymphocyte proliferation [76] and NK cell activity [77], one of the first lines of immune defense against foreign (including cancerous) cells. Furthermore, as it was previously discussed, a shift in the Th1/Th2 balance towards Th2 dominance [61] may be permissive to virus replication, increasing the frequency of tumor promotion [64]. In sum, these studies support the possibility that stress-induced immune dysregulation could be a cofactor for increasing the risk of tumor development, particularly of immunogenic tumors induced by oncogenic viruses. Alleviation of psychological distress has been shown to significantly extend life expectancy of individuals with metastatic cancers [78, 79]. Patients with metastatic melanoma were found to benefit from limited sessions of psychosocial interventions, thereby significantly increasing their survival rate [79]. These findings have set the stage to that researchers wonder themselves if the efficacy of cancer vaccines might be compromised under certain circumstances. Since depression and anxiety are commonly encountered disorders in cancer patients, it is going to be important that the conditions for utilizing such vaccines are to be optimized to assure the best possible immune response to them. Utilizing stress-reducing modalities in conjunction with cancer vaccine studies might result in a more vigorous immune response [73].
4. Serotonergic mechanisms against diseases related to virus infections
Serotonin (5HT), first isolated in 1933, is recognized as a very important neurotransmitter of the CNS. Serotonergic pathways are involved in the pathogenesis of depression, being drugs that block the reuptake of 5HT the mainstay of pharmacological antidepressant treatment [80]. The numerous investigations which explore the relationship between serotonergic transmission and different diseases, including that caused by or associated to virus, is justified by i) high prevalence of depression (lifetime prevalence aprox. 16%), isolated or associated to other disorders, and ii) frequent prescription of serotonergic antidepressants. The serotonergic effects observed in relation to viral processes, can be classified according to if they are dependent or not of immunological mechanisms.
4.1 Effects dependent of immunological mechanisms
Viruses are initially detected and destroyed non-specifically by innate immune mechanisms, but if the viruses escape from the early defense mechanisms, they are detected and eliminated specifically by adaptive immune mechanisms. There are more and more studies dealing with 5HT as signaling molecule in immune cells [81]. The last findings about 5HT synthesis and transport in defence cells, as well as the main 5HT receptor subtypes and their functions in innate and adaptive immune cells have been recently summarized [81]. Serotonergic modulation of the immune system has been demonstrated principally in relation to: tumor processes, infectious disorders and complications associated with viral diseases.
4.1.1 Tumor processes
5HT1A agonists and 5HT2A antagonists could be beneficial for tumor patients, since buspirone and nefazodone, respectively, have been shown to reverse the enhanced incidence and the shortened latent period by stress for mammary tumors in stressed-mice infected with the MTV mammary virus [70, 82]. Furthermore, 5HT1A agonists have interesting modulator effects on blood flow, since they constrict more dramatically the microvasculature feeding the tumors than that feeding normal tissue [83]. Immunoprotective effects of these drugs should justify the decreased lethality of cancer animals exposed to psychological stress as well as the higher surveillance of individuals with malignant tumors [66, 70]. Thus, 5HT1A agonist anxiolytics and/or 5HT2A antidepressants have been found to induce a reduction of the suppressive effects of stress on T-cell populations [8, 84], on the blastogenic response of spleen cells to concanavalin A [8, 84], on the thymus and spleen cellularity [8, 84], on the NK cell activity [85], on the in vivo and in vitro activity of phagocytosis [84, 85] and on the DTH response [86]. The mechanism of action of these serotonergic drugs on the immune system is unknown. However, knowing that buspirone and/or nefazodone regulate the level or secretion of numerous hormones (e.g. ACTH [8, 87, 88], corticosterone-cortisol [87], GH [89], TSH [90] and PRL [87, 91]), the
115©FORMATEX 2011
Science against microbial pathogens: communicating current research and technological advances A. Méndez-Vilas (Ed.)_______________________________________________________________________________

most accepted theory, and which at least should partially help to explain the immunoprotective effects of these drugs, points that probably they result from mediators involved in expressing the drug´s effect [8, 66, 85, 86, 92].
4.1.2 Infectious disorders
Killer lymphocytes (NK cells and CD8+ T cells) are key cells in the immunopathogenesis of HIV/AIDS. NK cells and CD8+ cells exert anti-HIV effects by both direct killing of HIV-infected cells as well as through the production of HIV suppressive factors (chemokines, cytokines, receptors and co-receptors) blocking viral entry and replication [93]. Since depression has been associated with suppression of NK and CD8+ T cell immunity in HIV-infected individuals [32, 94], psychological state is a risk factor for morbidity and mortality in HIV/AIDS [94, 95]. However, when depression-associated alterations of immunity are reversed by treatment with antidepressants, viral disease improves. For instance, Cruess et al. [96] observed significant increases in NK cytolytic activity in HIV-seropositive women associated with the resolution of depression; in the same way that Benton et al. [97] showed that HIV replication in latently infected cell lines and the HIV viral infectivity of macrophages were significantly decreased by immunological effects of selective serotonin reuptake inhibitors (SSRIs).
4.1.3 Complications associated with viral diseases
Drug abuse has been closely linked with viral infectious diseases, being it a risk factor for serious complications, such as neurotoxicity, in these patients. The harmful effect of drug abuse is double-strong: i) by weakness of immune system and, ii) by favoring of invasion. On the one hand, it is known that cocaine has an important suppressive effect on immunity. General depression of phagocyte activity, of the number of thymocytes and white blood cells, of antibody responses and of DTH reactions has been observed following the administration of cocaine. Besides, these drugs also induce a dose-related suppression of NK cell activity and, T cells from cocaine-treated mice fail to generate cytotoxic T-lymphocytes in vitro and in vivo [98]. On the other hand, recently it has been observed that cocaine directly, without participation of the immune system, increases HIV-1 invasion through the brain–blood barrier [99] and neurotoxicity of viral proteins (e.g. Tat, gp120) [100]. Several findings suggest that serotonergic transmission could be important for the cocaine-mediated enhancement of HIV-1 neurotoxicity: a) cocaine binds to transporter sites for 5HT with high affinity and inhibits the uptake of this monoamine into presynaptic neurons [101], b) cocaine exhibits indirect 5HT agonist properties [102]; c) cocaine-induced locomotion is attenuated by 5HT receptor antagonists [103]. However, a study carried out by Arsenov et al. [104] revealed that inhibition of 5HT transporter function by SSIRs did not contribute to the ability of cocaine to enhance the toxicity of HIV proteins, being this effect mediated by dopaminergic systems.
4.2 Effects non-dependent of immunological mechanisms
4.2.1 Tumorigenic effects
A controversial issue about serotonergic antidepressants, principally SSRI antidepressants, is their relationship with the development of tumors. SSRIs have been reported to be both tumor promoters and antineoplastic agents [105-109], and even some studies have failed to confirm their relationship [110]. Crowson and Magro [107] postulated that cutaneous pseudolymphomas were associated with antidepressant therapy (fluoxetine hydrochloride and amitriptyline hydrochloride), possibly reflecting perturbation of lymphoid function by inhibiting the binding of endogenous histamine to an intracellular histamine receptor (HIC). Likewise, Brandes et al. [105] found that fluoxetine: a) decreased the latency and increased the growth of tumors induced by the subcutaneous injection of C-3 fibrosarcoma cells; b) increased aggregate tumor weight (s.c. injected melanoma) without decreasing time to death for pulmonary metastasis (i.v. injected melanoma); c) decreased the latency and increased the frequency of DBMA-induced mammary tumors. By contrast, Abdul et al. [108] observed that the growth of subcutaneous, PC-3 xenografts (prostate carcinoma) in athymic nude mice was significantly inhibited by fluoxetine in a dose-dependent manner. Likewise, Bendele et al. [109] tested fluoxetine hydrochloride for carcinogenicity in rodents. They found no evidence of an increased incidence of any type of unusual or commonly occurring spontaneous neoplasm in either rats or mice, constating even statistically significant decreases in a few commonly occurring neoplasms. The same was observed by other authors. Fluoxetine was shown to sensitize the cytotoxic potential of conventional anticancer agents in drug resistance cells and human xenograft mouse tumor models [111], to decrease the viability of human colon cells [112] and to induce cytotoxicity in some tumor derived cells [108]. A recent study showed that fluoxetine has a selective antiproliferative effect to undifferentiated neural cells and modulates the levels of differentiation markers in differentiated cells [113]. In spite of that the protective effect of this SSRI could be explained by its immunostimulant action [9], serious doubts have been raised about it [114-116]. Taking together, these publications do not support a clear association between the use of antidepressants and cancer, so SSRIs are neither complete carcinogens nor antineoplastic agents.
116 ©FORMATEX 2011
Science against microbial pathogens: communicating current research and technological advances A. Méndez-Vilas (Ed.)______________________________________________________________________________

There are few researches investigating if protective and/or deleterious effects of SSRIs are applicable to cancer usually associated to virus-induced immunosupression. Outside of endemic regions (equatorial Africa, northeastern Brazil, and Papua New Guinea), the incidence of Burkitt´s lymphoma (BL) has increased dramatically because of its association with HIV infection. BL is treated by aggressive combination chemotherapy, usually over a 6-month period. Although there is a 3-year relapse-free survival rate of approximately 80% for patients with local disease, patients with disseminated tumor respond worse to chemotherapy and have poor survival rates. Hopes have been raised by the report that lymphoid cells express the 5HT transporter, which may be targeted to elicit apoptosis in BL [117]. In fact, SSRIs were reported to specifically induce apoptotic death in biopsy-like BL cell lines [118]. These findings afford an important challenge: the identification of a SSRI which can be an alternative therapy especially in countries in which access to treatments of first choice are difficult, expensive and complicated.
4.2.2 Improvement in adherence to therapy
Adherence to antiviral therapy has shown to be a crucial factor to chronic patients. Without adequate adherence, antiretroviral agents are not maintained at sufficient concentrations to suppress viral replication in infected cells and to lower the plasma viral load. In addition to being associated with poor short-term virological response, poor adherence to antiviral medication favors the appearance of drug-resistant pathogens. Therefore, overcoming the factors that reduce adherence to antiretroviral agents is of utmost importance for prolonged viral load suppression. The critical factors that influence adherence fall into 4 main groups: (1) patient factors, such as drug use, age, mood states, etc; (2) medication regimen, such as dosing complexity, number of pills, etc; (3) the patient-health-care provider relationship; and (4) the system of care [119, 120]. Patients with chronic infections (e.g. hepatitis C, HIV/AIDS) have high prevalence of psychiatric symptoms such as depression and anxiety [121], which may be even aggravated or induced by the antiviral treatment (e.g. interferon-alpha) [122] requiring dose reduction or premature discontinuation of therapy.
Treatment of depression associated with viral states has shown to be an effective strategy to avoid premature termination of antiviral therapy. For instance, Horberg et al. [120] showed that depression, which was diagnosed in 42% of individuals with HIV, caused a worse adherence to antiretroviral regimes being it reversed by compliant SSRI use. Thus, it can be suggested that the prescription of antidepressants represents an additional strategy to manage nonadherence to antiviral regimes and more attention should be paid to factors that worsen treatment adherence, particularly stress, anxiety and depression in order to increase the effectiveness of antiviral therapy.
References
[1] Masood A, Banerjee B, Vijayan VK, Ray A. Modulation of stress-induced neurobehavioral changes by nitric oxide in rats. European Journal of Pharmacology. 2003;458:135-139.
[2] Glaser R, Kiecolt-Glaser JK. Stress-induced immune dysfunction: implications for health. Nature Reviews. Immunology. 2005;5:243-251.
[3] Segerstrom SC, Miller GE. Psychological stress and the human immune system: a meta-analytic study of 30 years of inquiry. Psychological Bulletin. 2004;130:601-630.
[4] Larson MR, Ader R, Moynihan JA. Heart rate, neuroendocrine, and immunological reactivity in response to an acute laboratory stressor. Psychosomatic Medicine. 2001;63:493-501.
[5] Pike JL, Smith TL, Hauger RL, Nicassio PM, Patterson TL, McClintick J, Costlow C, Irwin MR. Chronic life stress alters sympathetic, neuroendocrine, and immune responsivity to an acute psychological stressor in humans. Psychosomatic Medicine. 1997;59:447-457.
[6] Friedman EM, Lawrence DA. Environmental stress mediates changes in neuroimmunological interactions. Toxicological Sciences. 2002;67:4-10.
[7] Godbout JP, Glaser R. Stress-induced immune dysregulation: implications for wound healing, infectious disease and cancer. Journal of Neuroimmune Pharmacology. 2006;1:421-427.
[8] Freire-Garabal M, Belmonte A, Suárez-Quintanilla J. Effects of buspirone on the immunosuppressive response to stress in mice. Archives Internationales de Pharmacodynamie et de Thérapie. 1991;314:160-168.
[9] Freire-Garabal M, Núñez MJ, Losada C, Pereiro D, Riveiro MP, González-Patiño E, Mayán JM, Rey-Mendez M. Effects of fluoxetine on the immunosuppressive response to stress in mice. Life Sciences. 1997;60:PL403-413.
[10] Núñez MJ, Balboa J, Rodrigo E, Brenlla J, González-Peteiro M, Freire-Garabal M. Effects of fluoxetine on cellular immune response in stressed mice. Neuroscience Letters. 2006;396:247-251.
[11] Freire-Garabal M, Núñez MJ, Fernández-Rial JC, Couceiro J, García-Vallejo L, Rey-Méndez M. Phagocytic activity in stressed mice: effects of alprazolam. Research in Immunology. 1993;144:311-316.
[12] Freire-Garabal M, Núñez MJ, Balboa JL, González-Bahillo J, Belmonte A. Effects of midazolam on the activity of phagocytosis in mice submitted to surgical stress. Pharmacology, Biochemistry and Behavior. 1993;46:605-608.
[13] Varela-Patiño MP, Nuñez MJ, Losada C, Pereiro D, Castro-Bolaño C, Saburido XL, Otero JM, Mayán JM, Blanco A, Rey-Méndez M, Freire-Garabal M. Effects of alprazolam on delayed type hypersensitivity (DTH) response in stressed mice. Research Communications in Psychology, Psychiatry and Behavior. 1994;19:69-77.
[14] Nuñez MJ, Riveiro MP, Varela M, Liñares D, Lopez P, Maña P, Martinez-Bahamonde F, Nuñez L, Mayan JM, Rey-Mendez M, Freire-Garabal M. Effects of nefazodone on delayed type hypersensitivity response in stressed mice). Research Communications in Biological Psychology and Psychiatry. 1998;23:19-27.
117©FORMATEX 2011
Science against microbial pathogens: communicating current research and technological advances A. Méndez-Vilas (Ed.)_______________________________________________________________________________

[15] Fukui Y, Sudo N, Yu XN, Nukina H, Sogawa H, Kubo C. The restraint stress-induced reduction in lymphocyte cell number in lymphoid organs correlates with the suppression of in vivo antibody production. Journal of Neuroimmunology. 1997;79:211-217.
[16] Cohen S, Tyrrell DA, Smith AP. Psychological stress and susceptibility to the common cold. The New England Journal of Medicine. 1991;325:606-612.
[17] Cohen S, Doyle WJ, Skoner DP. Psychological stress, cytokine production, and severity of upper respiratory illness. Psychosomatic Medicine. 1999;61:175-180.
[18] Cohen S, Frank E, Doyle WJ, Skoner DP, Rabin BS, Gwaltney JM Jr. Types of stressors that increase susceptibility to the common cold in healthy adults. Health Psychology. 1998;17:214-223.
[19] Kemeny ME, Schedlowski M. Understanding the interaction between psychosocial stress and immune-related diseases: a stepwise progression. Brain, Behavior, and Immunity. 2007;21:1009-1018.
[20] Ada GL, Jones PD. The immune response to influenza infection. Current Topics in Microbiology and Immunology. 1986;128:1-54.
[21] Ennis A. Some newly recognized aspects of resistance against and recovery from influenza. Archives of Virology. 1982;73:207-217.
[22] Del Gobbo V, Villani N, Marini S, Balestra E, Caliò R. Suppressor cells induced by influenza virus inhibit interleukin-2 production in mice. Immunology. 1990;69:454-459.
[23] Freire-Garabal M, Balboa JL, Fernandez-Rial JC, Núñez MJ, Belmonte A. Effects of alprazolam on influenza virus infection in stressed mice. Pharmacology, Biochemistry and Behavior. 1993;46:167-172.
[24] Freire-Garabal M, Núñez MJ, Losada C, Pereiro D, Castro-Bolaño C, Gándara J, Varela-Patiño MP, Rey-Méndez M. Effects of buspirone on influenza A virus infection in stressed mice. Life Sciences. 1995;56:PL187-193.
[25] Broadbent DE, Broadbent MH, Phillpotts RJ, Wallace J. Some further studies on the prediction of experimental colds in volunteers by psychological factors. Journal of Psychosomatic Research. 1984;28:511-523.
[26] Clover RD, Abell T, Becker LA, Crawford S, Ramsey CN Jr. Family functioning and stress as predictors of influenza B infection. The Journal of Family Practice. 1989;28:535-539.
[27] Caldji C, Diorio J, Meaney MJ. Variations in maternal care in infancy regulate the development of stress reactivity. Biological Psychiatry. 2000;48:1164-1174.
[28] Avitsur R, Hunzeker J, Sheridan JF.Role of early stress in the individual differences in host response to viral infection. Brain, Behavior and Immunity. 2006;20:339-348.
[29] Julkunen I, Sareneva T, Pirhonen J, Ronni T, Melén K, Matikainen S. Molecular pathogenesis of influenza A virus infection and virus-induced regulation of cytokine gene expression. Cytokine & Growth Factor Reviews. 2001;12:171-180.
[30] Lortholary O, Christeff N, Casassus P, Thobie N, Veyssier P, Trogoff B, Torri O, Brauner M, Nunez EA, Guillevin L. Hypothalamo-pituitary-adrenal function in human immunodeficiency virus-infected men. The Journal of Clinical Endocrinology and Metabolism. 1996;81:791-796.
[31] Leserman J, Petitto JM, Golden RN, Gaynes BN, Gu H, Perkins DO, Silva SG, Folds JD, Evans DL. Impact of stressful life events, depression, social support, coping, and cortisol on progression to AIDS. The American Journal of Psychiatry. 2000;157:1221-1228.
[32] Leserman J, Petitto JM, Perkins DO, Folds JD, Golden RN, Evans DL. Severe stress, depressive symptoms, and changes in lymphocyte subsets in human immunodeficiency virus-infected men. A 2-year follow-up study. Archives of General Psychiatry. 1997;54:279-285.
[33] Sloan E, Collado-Hidalgo A, Cole S. Psychobiology of HIV infection. In: Ader R, ed. Psychoneuroimmunology. San Diego: Academic Press; 2007:869-895.
[34] Cole SW, Kemeny ME, Fahey JL, Zack JA, Naliboff BD. Psychological risk factors for HIV pathogenesis: mediation by the autonomic nervous system. Biological Psychiatry. 2003;54:1444-1456.
[35] Cole SW, Naliboff BD, Kemeny ME, Griswold MP, Fahey JL, Zack JA. Impaired response to HAART in HIV-infected individuals with high autonomic nervous system activity. Proceedings of the National Academy of Sciences of the United States of America. 2001;98:12695-12700.
[36] Antoni MH, Schneiderman N, Penedo F. Behavioral interventions: immunologic mediators and disease outcomes. In: Ader R, ed. Psychoneuroimmunology. San Diego: Academic Press; 2007:675-703.
[37] Antoni MH, Cruess S, Cruess DG, Kumar M, Lutgendorf S, Ironson G, Dettmer E, Williams J, Klimas N, Fletcher MA, Schneiderman N. Cognitive-behavioral stress management reduces distress and 24-hour urinary free cortisol output among symptomatic HIV-infected gay men. Annals of Behavioral Medicine. 2000;22:29-37.
[38] Cruess S, Antoni M, Cruess D, Fletcher MA, Ironson G, Kumar M, Lutgendorf S, Hayes A, Klimas N, Schneiderman N. Reductions in herpes simplex virus type 2 antibody titers after cognitive behavioral stress management and relationships with neuroendocrine function, relaxation skills, and social support in HIV-positive men. Psychosomatic Medicine. 2000;62:828-837.
[39] Antoni MH, Cruess DG, Klimas N, Maher K, Cruess S, Kumar M, Lutgendorf S, Ironson G, Schneiderman N, Fletcher MA. Stress management and immune system reconstitution in symptomatic HIV-infected gay men over time: effects on transitional naive T cells (CD4(+)CD45RA(+)CD29(+)).The American Journal of Psychiatry. 2002;159:143-145.
[40] Antoni MH, Carrico AW, Durán RE, Spitzer S, Penedo F, Ironson G, Fletcher MA, Klimas N, Schneiderman N. Randomized clinical trial of cognitive behavioral stress management on human immunodeficiency virus viral load in gay men treated with highly active antiretroviral therapy. Psychosomatic Medicine. 2006;68:143-151.
[41] Markham PD, Salahuddin SZ, Veren K, Orndorff S, Gallo RC. Hydrocortisone and some other hormones enhance the expression of HTLV-III. International Journal of Cancer. 1986;37:67-72.
[42] Ayyavoo V, Rafaeli Y, Nagashunmugam T, Mahalingham S, Phung MT, Hamam A, Williams WV, Weiner DB. HIV-1 viral protein R (Vpr) as a regulator of the target cell. Psychoneuroendocrinology. 1997;22(Suppl 1):S41-49.
118 ©FORMATEX 2011
Science against microbial pathogens: communicating current research and technological advances A. Méndez-Vilas (Ed.)______________________________________________________________________________

[43] Felten DL, Felten SY, Bellinger DL, Carlson SL, Ackerman KD, Madden KS, Olschowki JA, Livnat S. Noradrenergic sympathetic neural interactions with the immune system: structure and function. Immunological Reviews. 1987;100:225-260.
[44] Glaser R, Kiecolt-Glaser JK, Bonneau RH, Malarkey W, Kennedy S, Hughes J. Stress-induced modulation of the immune response to recombinant hepatitis B vaccine. Psychosomatic Medicine. 1992;54:22-29.
[45] Morag M, Morag A, Reichenberg A, Lerer B, Yirmiya R. Psychological variables as predictors of rubella antibody titers and fatigue--a prospective, double blind study. Journal of Psychiatric Research. 1999;33:389-395.
[46] Burns VE, Drayson M, Ring C, Carroll D. Perceived stress and psychological well-being are associated with antibody status after meningitis C conjugate vaccination. Psychosomatic Medicine. 2002;64:963-970.
[47] Sheridan JF, Feng NG, Bonneau RH, Allen CM, Huneycutt BS, Glaser R. Restraint stress differentially affects anti-viral cellular and humoral immune responses in mice. Journal of Neuroimmunology. 1991;31:245-255.
[48] Kiecolt-Glaser JK, Glaser R, Gravenstein S, Malarkey WB, Sheridan J. Chronic stress alters the immune response to influenza virus vaccine in older adults. Proceedings of the National Academy of Sciences of the United States of America. 1996;93:3043-3047.
[49] Vedhara K, Cox NK, Wilcock GK, Perks P, Hunt M, Anderson S, Lightman SL, Shanks NM. Chronic stress in elderly carers of dementia patients and antibody response to influenza vaccination. Lancet. 1999;353:627-631.
[50] Patriarca PA. A randomized controlled trial of influenza vaccine in the elderly. Scientific scrutiny and ethical responsibility. JAMA. 1994;272:1700-1701.
[51] Penninx BW, Kritchevsky SB, Yaffe K, Newman AB, Simonsick EM, Rubin S, Ferrucci L, Harris T, Pahor M. Inflammatory markers and depressed mood in older persons: results from the Health, Aging and Body Composition study. Biological Psychiatry. 2003;54:566-572.
[52] Glaser R, Robles TF, Sheridan J, Malarkey WB, Kiecolt-Glaser JK. Mild depressive symptoms are associated with amplified and prolonged inflammatory responses after influenza virus vaccination in older adults. Archives of General Psychiatry. 2003;60:1009-1014.
[53] Song C, Vandewoude M, Stevens W, De Clerck L, Van der Planken M, Whelan A, Anisman H, Dossche A, Maes M. Alterations in immune functions during normal aging and Alzheimer's disease. Psychiatry Research. 1999;85:71-80.
[54] Bonneau RH, Sheridan JF, Feng NG, Glaser R. Stress-induced suppression of herpes simplex virus (HSV)-specific cytotoxic T lymphocyte and natural killer cell activity and enhancement of acute pathogenesis following local HSV infection. Brain, Behavior, and Immunity. 1991;5:170-192.
[55] Cohen F, Kemeny ME, Kearney KA, Zegans LS, Neuhaus JM, Conant MA. Persistent stress as a predictor of genital herpes recurrence. Archives of Internal Medicine. 1999;159:2430-2436.
[56] Cacioppo JT, Kiecolt-Glaser JK, Malarkey WB, Laskowski BF, Rozlog LA, Poehlmann KM, Burleson MH, Glaser R. Autonomic and glucocorticoid associations with the steady-state expression of latent Epstein-Barr virus. Hormones and Behavior. 2002;42:32-41.
[57] Mehta SK, Cohrs RJ, Forghani B, Zerbe G, Gilden DH, Pierson DL. Stress-induced subclinical reactivation of varicella zoster virus in astronauts. Journal of Medical Virology. 2004;72:174-179.
[58] Mehta SK, Stowe RP, Feiveson AH, Tyring SK, Pierson DL. Reactivation and shedding of cytomegalovirus in astronauts during spaceflight. The Journal of Infectious Diseases. 2000;182:1761-1764.
[59] Schmader K, Studenski S, MacMillan J, Grufferman S, Cohen HJ. Are stressful life events risk factors for herpes zoster?. Journal of the American Geriatrics Society. 1990;38:1188-1194.
[60] Glaser R, Kiecolt-Glaser JK. Chronic stress modulates the virus-specific immune response to latent herpes simplex virus type 1. Annals of Behavioral Medicine. 1997;19:78-82.
[61] Glaser R, MacCallum RC, Laskowski BF, Malarkey WB, Sheridan JF, Kiecolt-Glaser JK. Evidence for a shift in the Th-1 to Th-2 cytokine response associated with chronic stress and aging. The journals of gerontology. Series A, Biological sciences and medical sciences. 2001;56:M477-482.
[62] Glaser R, Litsky ML, Padgett DA, Baiocchi RA, Yang EV, Chen M, Yeh PE, Green-Church KB, Caligiuri MA, Williams MV. EBV-encoded dUTPase induces immune dysregulation: Implications for the pathophysiology of EBV-associated disease. Virology. 2006;346:205-218.
[63] VanderPlate C, Kerrick G. Stress reduction treatment of severe recurrent genital herpes virus. Biofeedback and Self-Regulation. 1985;10:181-188.
[64] Glaser R, Padgett DA, Litsky ML, Baiocchi RA, Yang EV, Chen M, Yeh PE, Klimas NG, Marshall GD, Whiteside T, Herberman R, Kiecolt-Glaser J, Williams MV. Stress-associated changes in the steady-state expression of latent Epstein-Barr virus: implications for chronic fatigue syndrome and cancer. Brain, Behavior, and Immunity. 2005;19:91-103.
[65] Antoni MH, Lutgendorf SK, Cole SW, Dhabhar FS, Sephton SE, McDonald PG, Stefanek M, Sood AK. The influence of bio-behavioural factors on tumour biology: pathways and mechanisms. Nature Reviews. Cancer. 2006;6:240-248.
[66] Freire-Garabal M, Nuñez MJ, Losada C, Pereiro D, Riveiro P, Fernandez-Rial JC, Garcia-Vallejo LA, Mayán JM, Rey-Méndez M. Inhibitory effects of buspirone on the enhancement of lung metastases induced by operative stress in rats. Research Communications in Biological Psychology and Psychiatry. 1996;21:13-25.
[67] Spiegel D. Effects of psychotherapy on cancer survival. Nature Reviews. Cancer. 2002;2:383-389. [68] Thompson MP, Kurzrock R. Epstein-Barr virus and cancer. Clinical Cancer Research. 2004;10:803-821. [69] Freire-Garabal M, Núñez MJ, Balboa JL, Suárez JA, Belmonte A. Effects of alprazolam on the development of MTV-induced
mammary tumors in female mice under stress. Cancer Letters. 1992;62:185-189. [70] Freire-Garabal M, Rey-Méndez M, García-Vallejo LA, Balboa J, Suárez JM, Rodrigo E, Brenlla J, Núñez MJ. Effects of
nefazodone on the development of experimentally induced tumors in stressed rodents. Psychopharmacology. 2004;176:233-238.
[71] Freire-Garabal M, Núñez MJ, Balboa JL, Suárez JA, Gallego A, Belmonte A. Effects of amphetamine on the development of MTV-induced mammary tumors in female mice. Life Sciences. 1992;51:PL37-40.
119©FORMATEX 2011
Science against microbial pathogens: communicating current research and technological advances A. Méndez-Vilas (Ed.)_______________________________________________________________________________

[72] Kiecolt-Glaser JK, Glaser R. Psychoneuroimmunology and cancer: fact or fiction?. European Journal of Cancer. 1999;35:1603-1607.
[73] Glaser R. Stress-associated immune dysregulation and its importance for human health: a personal history of psychoneuroimmunology. Brain, Behavior, and Immunity. 2005;19:3-11.
[74] Padgett DA, Glaser R. How stress influences the immune response. Trends in Immunology. 2003;24:444-448. [75] Tausk F, Elenkov I, Moynihan J. Psychoneuroimmunology. Dermatologic Therapy. 2008;21:22-31. [76] Laudenslager ML, Ryan SM, Drugan RC, Hyson RL, Maier SF. Coping and immunosuppression: inescapable but not
escapable shock suppresses lymphocyte proliferation. Science. 1983;221:568-570. [77] Ben-Eliyahu S. The promotion of tumor metastasis by surgery and stress: immunological basis and implications for
psychoneuroimmunology. Brain, Behavior, and Immunity. 2003;17(Suppl 1):S27-36. [78] Kuchler T, Henne-Bruns D, Rappat S, Graul J, Holst K, Williams JI, Wood-Dauphinee S. Impact of psychotherapeutic
support on gastrointestinal cancer patients undergoing surgery: survival results of a trial. Hepato-gastroenterology. 1999;46:322-335.
[79] Fawzy FI, Fawzy NW, Hyun CS, Elashoff R, Guthrie D, Fahey JL, Morton DL. Malignant melanoma. Effects of an early structured psychiatric intervention, coping, and affective state on recurrence and survival 6 years later. Archives of General Psychiatry. 1993;50:681-689.
[80] Delgado PL, Price LH, Miller HL, Salomon RM, Aghajanian GK, Heninger GR, Charney DS. Serotonin and the neurobiology of depression. Effects of tryptophan depletion in drug-free depressed patients. Archives of General Psychiatry. 1994;51:865-874.
[81] Ahern GP. 5-HT and the immune system. Current Opinion in Pharmacology. 2011;11:29-33. [82] Freire-Garabal M, Nuñez M, Balboa JL, Belmonte-Otero A, Belmonte A. Effects of buspirone on the development of MTV-
induced mammary tumors in female mice under stress. Research Communications in Psychology, Psychiatry and Behavior. 1991; 16:107-114.
[83] Stücker O, Laemmel E, Teisseire B, Vicaut E. Specific response of mouse tumor-feeding arterioles to stimulation by 5-HT1 agonists. International Journal of Radiation Oncology, Biology, Physics. 1997;37:1125-1131.
[84] Freire-Garabal M, Varela M, Riveiro P, Balboa J, Liñares D, Mañá P, Mayán JM, Rey-Méndez M, Núñez MJ. Effects of nefazodone on the immune system of mice. European Neuropsychopharmacology. 2000;10:255-264.
[85] Freire-Garabal M, Nunez-Iglesias MJ, Balboa JL, Fernandez-Rial JC, Rey-Mendez M. Effects of buspirone on the immune response to stress in mice. Pharmacology, Biochemistry and Behavior. 1995;51:821-825.
[86] Freire-Garabal M, Nuñez MJ, Losada C, Pereiro D, Riveiro MP, Castro-Bolaño C, Blanco A, Millán J, Mayán JM, Rey-Mendez M. Effects of buspirone on delayed type hypersensitivity (DTH) response in stressed mice. Research Communications in Biological Psychology and Psychiatry. 1995;20:49-57.
[87] Levy AD, Van de Kar LD. Endocrine and receptor pharmacology of serotonergic anxiolytics, antipsychotics and antidepressants. Life Sciences. 1992;51:83-94.
[88] Matheson GK, Knowles A, Guthrie D, Gage D, Weinzapfel D, Blackbourne J. Actions of serotonergic agents on hypothalamic-pituitary-adrenal axis activity in the rat. General Pharmacology. 1997;29:823-828.
[89] Spencer GS, Dobbie P, Bass JJ. Neuroendocrine regulation of growth hormone secretion in sheep. III. Serotoninergic systems. Domestic Animal Endocrinology. 1991;8:383-391.
[90] Broqua P, Baudrie V, Bluet-Pajot MT, Chaouloff F. Buspirone, ipsapirone and 1-(2-pyrimidinyl)-piperazine decrease cold-induced thyrotropin secretion in rats. European Journal of Pharmacology. 1991;204:141-147.
[91] Walsh AE, Cowen PJ. Attenuation of the prolactin-stimulating and hyperthermic effects of nefazodone after subacute treatment. Journal of Clinical Psychopharmacology. 1994;14:268-273.
[92] Freire-Garabal M, Nuñez MJ, Fernandez-Rial JC, Rey-Mendez M, Belmonte A. Effects of buspirone on the resistance, development and passive transfer of immunity to Listeria monocytogenes in mice submitted to stress. Archives Internationales de Pharmacodynamie et de Thérapie. 1993;324:114-123.
[93] Fauci AS, Mavilio D, Kottilil S. NK cells in HIV infection: paradigm for protection or targets for ambush. Nature Reviews. Immunology. 2005;5:835-843.
[94] Evans DL, Ten Have TR, Douglas SD, Gettes DR, Morrison M, Chiappini MS, Brinker-Spence P, Job C, Mercer DE, Wang YL, Cruess D, Dube B, Dalen EA, Brown T, Bauer R, Petitto JM. Association of depression with viral load, CD8 T lymphocytes, and natural killer cells in women with HIV infection. The American Journal of Psychiatry. 2002;159:1752-1759.
[95] Chida Y, Vedhara K. Adverse psychosocial factors predict poorer prognosis in HIV disease: a meta-analytic review of prospective investigations. Brain, Behavior, and Immunity. 2009;23:434-445.
[96] Cruess DG, Douglas SD, Petitto JM, Have TT, Gettes D, Dubé B, Cary M, Evans DL. Association of resolution of major depression with increased natural killer cell activity among HIV-seropositive women. The American Journal of Psychiatry. 2005;162:2125-2130.
[97] Benton T, Lynch K, Dubé B, Gettes DR, Tustin NB, Ping Lai J, Metzger DS, Blume J, Douglas SD, Evans DL. Selective serotonin reuptake inhibitor suppression of HIV infectivity and replication. Psychosomatic Medicine. 2010;72:925-932.
[98] Freire-Garabal M, Nuñez MJ, Balboa JL, Fernandez-Rial JC, Suarez-Quintanilla J, Orallo F, Gonzalez-Bahillo J, Belmonte A. Effects of cocaine on the resistance, development and passive transfer of immunity to Listeria Monocytogenes in mice. Research Communications in Substances of Abuse. 1992;13:315-322.
[99] Fiala M, Eshleman AJ, Cashman J, Lin J, Lossinsky AS, Suarez V, Yang W, Zhang J, Popik W, Singer E, Chiappelli F, Carro E, Weinand M, Witte M, Arthos J. Cocaine increases human immunodeficiency virus type 1 neuroinvasion through remodeling brain microvascular endothelial cells. Journal of Neurovirology. 2005;11:281-291.
[100] Aksenov MY, Aksenova MV, Nath A, Ray PD, Mactutus CF, Booze RM. Cocaine-mediated enhancement of Tat toxicity in rat hippocampal cell cultures: the role of oxidative stress and D1 dopamine receptor. Neurotoxicology. 2006;27:217-228.
120 ©FORMATEX 2011
Science against microbial pathogens: communicating current research and technological advances A. Méndez-Vilas (Ed.)______________________________________________________________________________

[101] Ritz MC, Cone EJ, Kuhar MJ. Cocaine inhibition of ligand binding at dopamine, norepinephrine and serotonin transporters: a structure-activity study. Life Sciences. 1990;46:635-645.
[102] Parsons LH, Koob GF, Weiss F. Serotonin dysfunction in the nucleus accumbens of rats during withdrawal after unlimited access to intravenous cocaine. The Journal of Pharmacology and Experimental Therapeutics. 1995;274:1182-1191.
[103] Reith ME. 5-HT3 receptor antagonists attenuate cocaine-induced locomotion in mice. European Journal of Pharmacology. 1990;186:327-330.
[104] Aksenov MY, Aksenova MV, Silvers JM, Mactutus CF, Booze RM. Different effects of selective dopamine uptake inhibitors, GBR 12909 and WIN 35428, on HIV-1 Tat toxicity in rat fetal midbrain neurons. Neurotoxicology. 2008;29:971-977.
[105] Brandes LJ, Arron RJ, Bogdanovic RP, Tong J, Zaborniak CL, Hogg GR, Warrington RC, Fang W, LaBella FS. Stimulation of malignant growth in rodents by antidepressant drugs at clinically relevant doses. Cancer Research. 1992;52:3796-3800.
[106] Steingart AB, Cotterchio M. Do antidepressants cause, promote, or inhibit cancers? Journal of Clinical Epidemiology. 1995;48:1407-1412.
[107] Crowson AN, Magro CM. Antidepressant therapy. A possible cause of atypical cutaneous lymphoid hyperplasia. Archives of Dermatology. 1995;131:925-929.
[108] Abdul M, Logothetis CJ, Hoosein NM. Growth-inhibitory effects of serotonin uptake inhibitors on human prostate carcinoma cell lines. The Journal of Urology. 1995;154:247-250.
[109] Bendele RA, Adams ER, Hoffman WP, Gries CL, Morton DM. Carcinogenicity studies of fluoxetine hydrochloride in rats and mice. Cancer Research. 1992;52:6931-6935.
[110] Stebbing J, Powles T, Mandalia S, Nelson M, Gazzard B, Bower M. Use of antidepressants and risk of cancer in individuals infected with HIV. Journal of Clinical Oncology. 2008;26:2305-2310.
[111] Peer D, Dekel Y, Melikhov D, Margalit R. Fluoxetine inhibits multidrug resistance extrusion pumps and enhances responses to chemotherapy in syngeneic and in human xenograft mouse tumor models. Cancer Research. 2004;64:7562-7569.
[112] Yue CT, Liu YL. Fluoxetine increases extracellular levels of 3-methoxy-4-hydroxyphenylglycol in cultured COLO320 DM cells. Cell Biochemistry and Function. 2005;23:109-114.
[113] Di Lieto A, Leo D, Volpicelli F, di Porzio U, Colucci-D'Amato L. FLUOXETINE modifies the expression of serotonergic markers in a differentiation-dependent fashion in the mesencephalic neural cell line A1 mes c-myc. Brain Research. 2007;1143:1-10.
[114] Rabkin JG, Rabkin R, Wagner G. Effects of fluoxetine on mood and immune status in depressed patients with HIV illness. The Journal of Clinical Psychiatry. 1994;55:92-97.
[115] Maes M, Meltzer HY, Bosmans E, Bergmans R, Vandoolaeghe E, Ranjan R, Desnyder R. Increased plasma concentrations of interleukin-6, soluble interleukin-6, soluble interleukin-2 and transferrin receptor in major depression. Journal of Affective Disorders. 1995;34:301-309.
[116] Berkeley MB, Daussin S, Hernandez MC, Bayer BM. In vitro effects of cocaine, lidocaine and monoamine uptake inhibitors on lymphocyte proliferative responses. Immunopharmacology and Immunotoxicology. 1994;16:165-178.
[117] Serafeim A, Grafton G, Chamba A, Gregory CD, Blakely RD, Bowery NG, Barnes NM, Gordon J. 5-Hydroxytryptamine drives apoptosis in biopsylike Burkitt lymphoma cells: reversal by selective serotonin reuptake inhibitors. Blood. 2002;99:2545-2553.
[118] Serafeim A, Holder MJ, Grafton G, Chamba A, Drayson MT, Luong QT, Bunce CM, Gregory CD, Barnes NM, Gordon J. Selective serotonin reuptake inhibitors directly signal for apoptosis in biopsy-like Burkitt lymphoma cells. Blood. 2003;101:3212-3219.
[119] Chesney MA. Factors affecting adherence to antiretroviral therapy. Clinical Infectious Diseases. 2000;30(Suppl 2):S171-176.
[120] Horberg MA, Silverberg MJ, Hurley LB, Towner WJ, Klein DB, Bersoff-Matcha S, Weinberg WG, Antoniskis D, Mogyoros M, Dodge WT, Dobrinich R, Quesenberry CP, Kovach DA. Effects of depression and selective serotonin reuptake inhibitor use on adherence to highly active antiretroviral therapy and on clinical outcomes in HIV-infected patients. Journal of Acquired Immune Deficiency Syndromes. 2008;47:384-390.
[121] Kraus MR, Schäfer A, Csef H, Scheurlen M, Faller H. Emotional state, coping styles, and somatic variables in patients with chronic hepatitis C. Psychosomatics. 2000;41:377-384.
[122] Kraus MR, Schäfer A, Faller H, Csef H, Scheurlen M. Paroxetine for the treatment of interferon-alpha-induced depression in chronic hepatitis C. Alimentary Pharmacology & Therapeutics. 2002;16:1091-1099.
121©FORMATEX 2011
Science against microbial pathogens: communicating current research and technological advances A. Méndez-Vilas (Ed.)_______________________________________________________________________________



![*Department of Pharmaceutical Chemistry, College of ... · infectious diseases [2]. Sulfonamides were the first antimicrobial drugs used, which lead to the antibiotic revolution in](https://img.pdfslide.us/doc/110x75/5f08bf277e708231d42385b8/department-of-pharmaceutical-chemistry-college-of-infectious-diseases-2.jpg)

























