Embed Size (px)
Citation preview

J O U R N A L O F P R O T E O M I C S 9 4 ( 2 0 1 3 ) 2 8 9 – 3 0 1
Ava i l ab l e on l i ne a t www.sc i enced i r ec t . com
ScienceDirect
www.e l sev i e r . com/ loca te / j p ro t
Proteomics study reveals the molecular
mechanisms underlying water stress toleranceinduced by Piriformospora indica in barleyMehdi Ghaboolia,b, Behnam Khatabic, Farajolah Shahriary Ahmadid, Mozhgan Sepehrie,Mehdi Mirzaeif, Ardeshir Amirkhanig, Jesús V. Jorrín-Novoh, Ghasem Hosseini Salekdeha,i,⁎aDepartment of Systems Biology, Agricultural Biotechnology Research Institute of Iran, Karaj, IranbDepartment of Agronomy, Faculty of Agriculture, Malayer University, Malayer, IrancDepartment of Entomology and Plant Pathology, University of Tennessee, Knoxville, TN, USAdDepartment of Crop Biotechnology and Breeding, Faculty of Agriculture, Ferdowsi University of Mashhad, Mashhad, IraneDepartment of Soil Science, Faculty of Agriculture, Isfahan University of Technology, Isfahan, IranfAustralian School of Advanced Medicine, Faculty of Human Sciences, Macquarie University, Sydney, NSW, AustraliagAustralian Proteome Analysis Facility (APAF), Department of Chemistry and Biomolecular Sciences, Macquarie University, Sydney, AustraliahAgricultural and Plant Biochemistry and Proteomics Research Group, Dept. of Biochemistry and Molecular Biology, University of Córdoba,Córdoba, SpainiDepartment of Molecular Systems Biology at Cell Science Research Center, Royan Institute for Stem Cell Biology and Technology, ACECR,Tehran, Iran
A R T I C L E I N F O
⁎ Corresponding author at: Agricultural Biotefax: +98 26 32704539.
E-mail address: [email protected] (G
1874-3919/$ – see front matter © 2013 Elseviehttp://dx.doi.org/10.1016/j.jprot.2013.09.017
A B S T R A C T
Article history:Received 6 August 2013Accepted 30 September 2013Available online 11 October 2013
Piriformospora indica is a mutualistic root endophytic fungus, which transfers several benefits tohosts including enhance plant growth and increase yield under both normal and stressconditions. It has been shown that P. indica root-colonization enhances water stress tolerancebased on general and non-specific plant-species mechanism. To better understand themolecular mechanism of P. indica-mediated drought stress tolerance, we designed a set ofcomparative experiments to study the impact of P. indica on barely plants cultivar “GoldenPromise” grown under different drought levels [Filed capacity (F.C.) and 25% F.C.]. P. indicaenhanced root and shoot biomass of colonized plants under both well-watered andwater-deficit conditions. Proteome analysis of P. indica-colonized barley leaves underwell-treated and water-deficit conditions resulted in detection of 726 reproducibly proteinspots.Mass spectrometry analysis resulted in the identification of 45 differentially accumulatedproteins involved in photosynthesis, reactive oxygen scavenging, metabolisms, signaltransduction, and plant defense responses. Interestingly, P. indica increased the level of proteinsinvolved in photosynthesis, antioxidative defense system and energy transport. We proposethat P. indica-mediated drought stress tolerance in barely is through photosynthesisstimulation, energy releasing and enhanced antioxidative capacity in colonized plants.
Biological significancePlant mutualistic symbionts offer long-term abiotic stress tolerance through the hostadaptation to environmental stress. There have been a few published proteomic studies ofplant symbionts to drought, and this is thought to be the first proteomic analysis,
Keywords:DroughtSymbiosis associationPiriformospora indicaPhotosynthesisAntioxidative capacityBarleyProteomics
chnology Research Institute of Iran, P.O. Box 31535-1897, Karaj, Iran. Tel.: +98 26 32700845;
.H. Salekdeh).
r B.V. All rights reserved.

290 J O U R N A L O F P R O T E O M I C S 9 4 ( 2 0 1 3 ) 2 8 9 – 3 0 1
demonstrating the impact of endophyte on barley plant under drought stress. For some ofidentified proteins like TCTP and PCNA, a connection to physiological function in plants isnovel, and can be the best candidates for sources of drought tolerance in future studies.
© 2013 Elsevier B.V. All rights reserved.
1. Introduction
Plants may establish association with beneficial microorganismsto increase tolerance/resistance to abiotic and biotic stresses. Themost frequently foundmutualistic fungal symbiont is ArbuscularMycorrhizal Fungi (AMF), having symbiosis between more than80% of land plants and fungi [1,2]. Endophytes are another groupof beneficial microorganisms that complete part of their lifewithinplant tissueswithout causing apparent injury tohost plant[3]. Endophytic fungi are able to fix nitrogen or enhance phos-phorus uptake aswell as improve plant growth and development[4]. Endophytesprovideprotectionandhost survival by improvinghost tolerance to stress, insect, and disease-resistance [3].Recently, fungal endophytes have been subjected to molecularstudies to understand molecular basis of symbiotic interactions.
Piriformospora indica is a root-interacting mutualistic endo-phytic fungus, which is phylogenetically close to mycorrhizalendosymbionts of orchid and ericoid roots [5]. P. indica isolatedfrom Thar Desert of India in 1997. It is easily grown onvarious complex and minimal substrates and asexuallyforms chlamydospores containing 8 to 25 nuclei [6]. P. indicapromotes plant growth and yield in many plant species,particularly under environmentally harsh conditions that arenot optimal for plants [7,8]. P. indica-root colonization isinitiated with a biotrophic growth phase and continued witha cell death-dependent phase in Arabidopsis and barely. Inspite of the cell death phase induced by P. indica during rootcolonization, the colonized plant showed increase in plantgrowth and yield [9]. This data described P. indica as a novelmutualistic symbiont compared to known mycorrhizae.
Drought is one of the major constraints to crop productivityworldwide, which adversely affects plant growth and yield. Overthe last decade, many efforts have been made to unravel themechanisms of drought tolerance in many plant species. Incontrast, only recently, few molecular studies have beenundertaken to study the impact of plant mutualistic symbiontsin conferring water stress tolerance to hosts [10].
Proteomicshasproved tobeapowerful approach indiscoveringthe molecular mechanisms of plant response to abiotic stresses[11–13]. P. indica-root colonization led to drought stress tolerance inseveral plant species including Arabidopsis, barley, and Chinesecabbage [7,14,15]. In this respect, the contribution of P. indica toimprove plant drought tolerance is point to the natural habitat ofits desert origin.P. indica-mediateddrought stress tolerance is likelybased on general and non-specific plant-species. In Arabidopsis,P. indica-mediated drought stress tolerance is associated withincrease in the host antioxidative capacity through the activationof the glutathione-ascorbate redox pathway; induction ofdrought-related genes, and targeting the plastid-localized Ca2+
sensing regulator protein [14,15]. Currently, only a few proteomicstudies are performed to study the host adaptive mechanismsdeliberated by plant mutualistic symbionts to the environmentalextremes especially drought and salinity, which are particularlyimportant for agriculture worldwide [10,16,17].
In this study, we integrated physiological and proteomicsdata to unravel the molecular mechanisms underlying P. indica-mediated drought stress tolerance in barley. Our results showedthat P. indica promoted growth of barley (Hordeum vulgare L.), andaltered the proteome pattern of P. indica-colonized barley leavesunder drought stress condition. Finally, the potential ofidentified proteins in themechanisms-conferring drought stresstolerance are discussed.
2. Materials and methods
2.1. Fungus, plant inoculation and growth conditions
P. indica was grown on CM (complex medium) and the sporesuspension collected after 28 days by gently scratching the fungussurface on the Petri dishes with a spatula until the spores werereleased. The spore suspension was filtered through cheeseclothto remove the excess medium and washed three times withdistilled water (dH2O) containing Tween-20. After each washingstep, the spores were collected by centrifugation at 4000 g for7 min. The spore pellet was finally suspended again in dH2O andadjusted to 5 × 105 spores per mL. For plant inoculation, barleyseeds of the cultivar “Golden Promise” were surface-sterilizedwith 70% ethanol (v/v) for 30 s followed by 6% sodiumhypochlorite (NaOCl) for 15 min. They were then rinsed inwater, and were germinated for 2 days. Barley seedlings wereinoculated by immersing in the spore suspension solution withgentle shaking for 1–2 h. The mock-treated seedlings weredipped in sterile water only. Inoculated and non-inoculatedbarley seedlings were later transferred into pots, filled withnormal soil and then placed in the growth chamber at 22/18 °Cday/night cycle, 60% relativehumidity, anda photoperiod of 16 h(220 μmol·m−2·s−1 photon flux density). Plants were irrigated bydistilled water. The experiment was conducted in a completelyrandomized design under two treatments (P. indica-inoculatedand non-inoculated plants), and two levels of drought treat-ments [Filed capacity (F.C.) and 25% F.C.] in three biologicalreplications. Drought was initiated 14 days after plant inocula-tion by withholding water and drought-stressed pots werere-watered, when the soil moisture reached 25% F.C. Thewell-watered treatment was maintained near F.C. The shootand root samples from P. indica-colonized and non-inoculatedplants under well-watered and water-deficit conditions wereharvested 28 days after fungal inoculation. Three plants fromeach pot were merged together, and considered as a singlereplication and used for physiological and proteomic analyses.
2.2. Sampling, microscopic analysis and physiologicalanalysis
Root samples from P. indica-inoculated and non-inoculatedplants were collected 14 days after plant inoculation. Rootwere softened by 10% KOH solution for 15 min, acidified with

291J O U R N A L O F P R O T E O M I C S 9 4 ( 2 0 1 3 ) 2 8 9 – 3 0 1
1 M HCl for 10 min, and stained with 0.02% Trypan blueovernight, and then were distained with 50% lacto-phenol for1 h prior to microscopic observation under a light microscope.Physiological traits including fresh and dry weights of rootand shoot of P. indica-colonized and non-inoculated plantsunder well-watered and water-deficit conditions were mea-sured 28 days post inoculation. Plant dry weight wasmeasured after drying leaf and root tissues in oven for 48 hat 70 °C.
2.3. Protein extraction
Protein was extracted according with Dameral et al. (1986) withsome modifications [18]. Leaf sample (1 g per plant) wasgrounded with liquid nitrogen, dissolved in extraction buffer[10% w/v trichloroacetic acid (TCA) in ice-cold acetone with0.07% w/v dithiothreitol (DTT)] for 1 h at −20 °C, and then wascentrifuged at 35,000 g for 15 min at 4 °C. Pellets were washedby ice-cold acetone containing 0.07 w/v DTT, incubated at−20 °C for 1 h, and then were centrifuged at 15,000 g for 15 minat 4 °C. The washing and sedimentation steps were repeatedthree times and finally pellets were lyophilized. The lyophilizedpowder was homogenized in lysis buffer (9 M urea, 2% w/v3-[(3-cholamidopropyl)dimethylamonio]-1-propanesulfonate(CHAPS), 0.8% w/v pharmalyte pH 3–10, 1% w/v DTT) in 37 °Cfor 1.5 h. After removing pellet, protein concentration wasdetermined using Bradford assay (Bio-Rad), and bovine serumalbumin (BSA) was used to generate a standard curve.
2.4. Two dimensional gel electrophoresis and imageanalysis
In first dimension, isoelectric focusing (IEF) was carried out usingthe Immobiline™DryStrip gels (IPG strips) (18 cm, pH 4–7, linear)onMultiphor II (GEhealthcare) for 16 h at roomtemperature (RT).For two-dimensional (2-DE) gel electrophoresis (both analyticaland preparative gels), 350 μg and 3.5 mg of total protein wereloaded on each gel, respectively. The running conditions forelectrophoresis (IEF) were as follows: 500 V for 1 h, followed by
Fig. 1 – Effect of Piriformospora indica on barley growth rates cultifigure represents barley under well-watered [Filed capacity (F.C.)condition (25% F.C.).
1000 V for 1 h, and finally run at 3500 V for 16 h. The focusedstrips were equilibrated twice for 15 min in 10 mL equilibrationsolution. The first equilibration was carried out in the presenceof solution containing 6 Murea, 30% v/v glycerol, 2%w/v SDS, 1%w/v DTT, and 50 mM Tris–HCl, pH 8.8. The second equilibrationwas performed in the same solution modified by thereplacement of DTT by 2.5% w/v iodoacetamide. Strips wereloaded on 12.5% SDS-PAGE gels using a Dodeca Cell (Bio-Rad).Analytical 2-DE gels were stained with silver nitrate [19], andpreparative gels were stained with colloidal Coomassie brilliantblue (CBB) G-250 [20]. Silver-stained gels were immediatelyscanned using GS-800 densiometer (Bio-Rad), and gel imageswere analyzed by Melanie 6 software (Genebio, Geneva,Switzerland). Protein spot pairs were performed visually and byscatter plots between gels of each data point that were displayedto estimate gel similarity and experimental errors. Molecularweights of proteins on gels were calculated by standard proteinmarkers (GE healthcare). One two-dimensional gel per plant wasrun and percent volume of each spot was estimated andanalyzed. Two-way analysis of variance (ANOVA), and comparedtreatment means were calculated. Protein spots were foundsignificantly up or down, when p ≤ 0.05 and had consistentlypresent in all three biological replications. The induction factorwas calculated by dividing the percent volume of spots of 25%F.C. condition by the percent volume of spots corresponding toF.C. condition.
2.5. Mass spectrometry analysis
Protein spots with significant changes, were excised frompreparative CBB-stained gels [20]. Gel slices were destainedwith freshly prepared wash solution [100% acetonitrile/50 mMammonium bicarbonate (NH4CHO3) (50:50 v/v)] for 1 h at RT.Wash solution was removed and protein spots were air-driedfor 30 min at 37 °C. Proteins were digested using a trypsinsolution (12 ng/ml trypsin in 50 mM NH4CHO3). The reactionproceeded by incubation for 45 min at 47 °C. Excess of trypsinsolution was removed and replaced by 50 mM NH4CHO3, andgel pieces were incubated at 37 °C overnight. All samples were
var “Golden Promise” 28 days after fungal inoculation. Left], and right figure shows barley plants exposed to drought
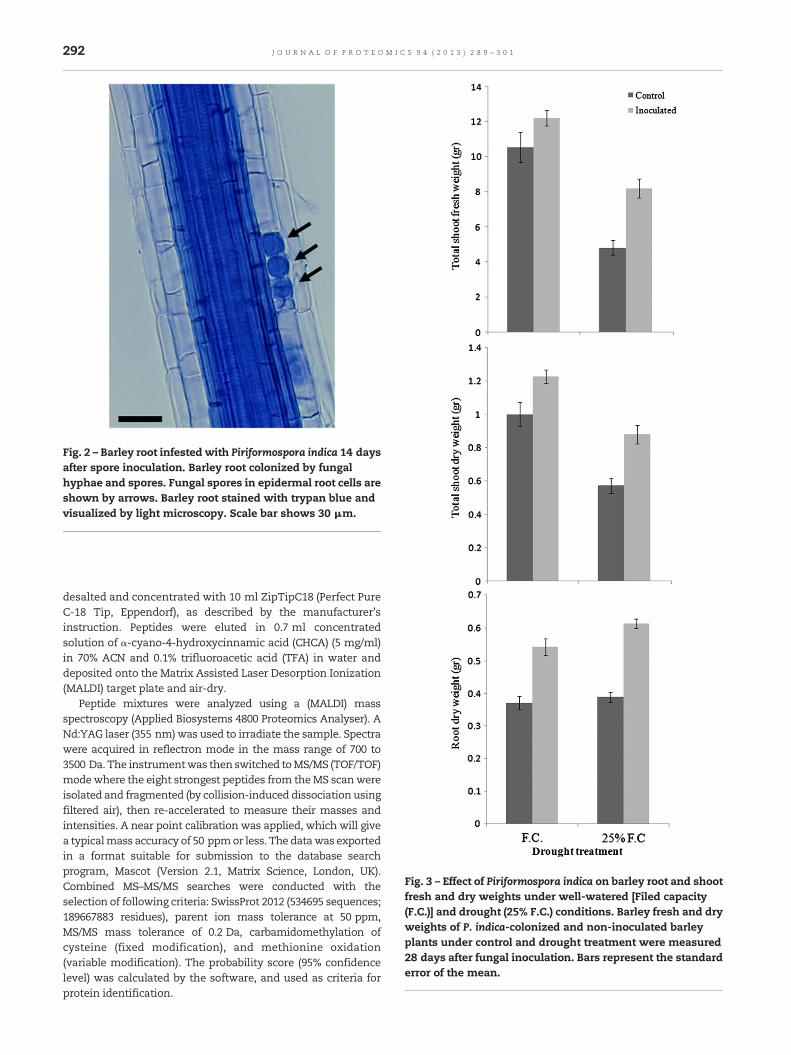
Fig. 2 – Barley root infested with Piriformospora indica 14 daysafter spore inoculation. Barley root colonized by fungalhyphae and spores. Fungal spores in epidermal root cells areshown by arrows. Barley root stained with trypan blue andvisualized by light microscopy. Scale bar shows 30 μm.
Fig. 3 – Effect of Piriformospora indica on barley root and shootfresh and dry weights under well-watered [Filed capacity(F.C.)] and drought (25% F.C.) conditions. Barley fresh and dryweights of P. indica-colonized and non-inoculated barleyplants under control and drought treatment were measured28 days after fungal inoculation. Bars represent the standarderror of the mean.
292 J O U R N A L O F P R O T E O M I C S 9 4 ( 2 0 1 3 ) 2 8 9 – 3 0 1
desalted and concentrated with 10 ml ZipTipC18 (Perfect PureC-18 Tip, Eppendorf), as described by the manufacturer'sinstruction. Peptides were eluted in 0.7 ml concentratedsolution of α-cyano-4-hydroxycinnamic acid (CHCA) (5 mg/ml)in 70% ACN and 0.1% trifluoroacetic acid (TFA) in water anddeposited onto the Matrix Assisted Laser Desorption Ionization(MALDI) target plate and air-dry.
Peptide mixtures were analyzed using a (MALDI) massspectroscopy (Applied Biosystems 4800 Proteomics Analyser). ANd:YAG laser (355 nm)was used to irradiate the sample. Spectrawere acquired in reflectron mode in the mass range of 700 to3500 Da. The instrumentwas then switched toMS/MS (TOF/TOF)modewhere the eight strongest peptides from theMS scanwereisolated and fragmented (by collision-induced dissociation usingfiltered air), then re-accelerated to measure their masses andintensities. A near point calibration was applied, which will givea typicalmass accuracy of 50 ppmor less. The datawas exportedin a format suitable for submission to the database searchprogram, Mascot (Version 2.1, Matrix Science, London, UK).Combined MS–MS/MS searches were conducted with theselection of following criteria: SwissProt 2012 (534695 sequences;189667883 residues), parent ion mass tolerance at 50 ppm,MS/MS mass tolerance of 0.2 Da, carbamidomethylation ofcysteine (fixed modification), and methionine oxidation(variable modification). The probability score (95% confidencelevel) was calculated by the software, and used as criteria forprotein identification.

Fig. 4 – Two dimensional (2-D) gel electrophoresis analyses of proteins extracted from barley leaves “Golden Promise” underwell-watered (F.C.) condition. Three hundred fifty micrograms of total protein was loaded on SDS-PAGE gel, with a well formolecular weight standards. Proteins were visualized by silver staining and arrows represent responsive proteins described inTable 1. The changes in protein abundance between Piriformospora indica-inoculated and non-inoculated plants under controland stress condition have been presented in Table 1.
Fig. 5 – Functional annotation of the identified proteins.
293J O U R N A L O F P R O T E O M I C S 9 4 ( 2 0 1 3 ) 2 8 9 – 3 0 1
2.6. Gene expression analysis using real-time reversetranscription-PCR
Total RNA, from the same samples used for proteomeanalysis, was extracted using Trizol reagent (Invitrogen/Lifetechnologies). Two micrograms of total RNA was used for cDNAsynthesis using RevertAid First Strand cDNA Synthesis Kit(Thermo Scientific). The list of selected genes and designedprimers used in this study is presented in Table S1. qRT-PCRreaction was performed using iQ SYBR Green Supermix Kit(Bio-Rad). PCR profile was as follows: 4 min at 95 °C, 45 cycles of15 s at 95 °C, 20 s at 60 °C, 30 s at 72 °C, and 1 min at 72 °C,followed melting curve analysis (55 °C to 95 °C in increasingsteps of 0.5 °C). All replication reactions were performed in six(three biological replications and two technical replicates). Thebarley ubiquitin gene was selected as a housekeeping gene andused in parallel for each sample. The 2−ΔΔCT method was usedto analysis qRT-PCR data [21]. The mean value, varianceand standard error were calculated from three independentcombined values for each targeted gene.
3. Results
3.1. Microscopic analysis and plant growth
We first studied the interaction of P. indica with barley rootsunder drought stress condition by microscopy. Barley roots
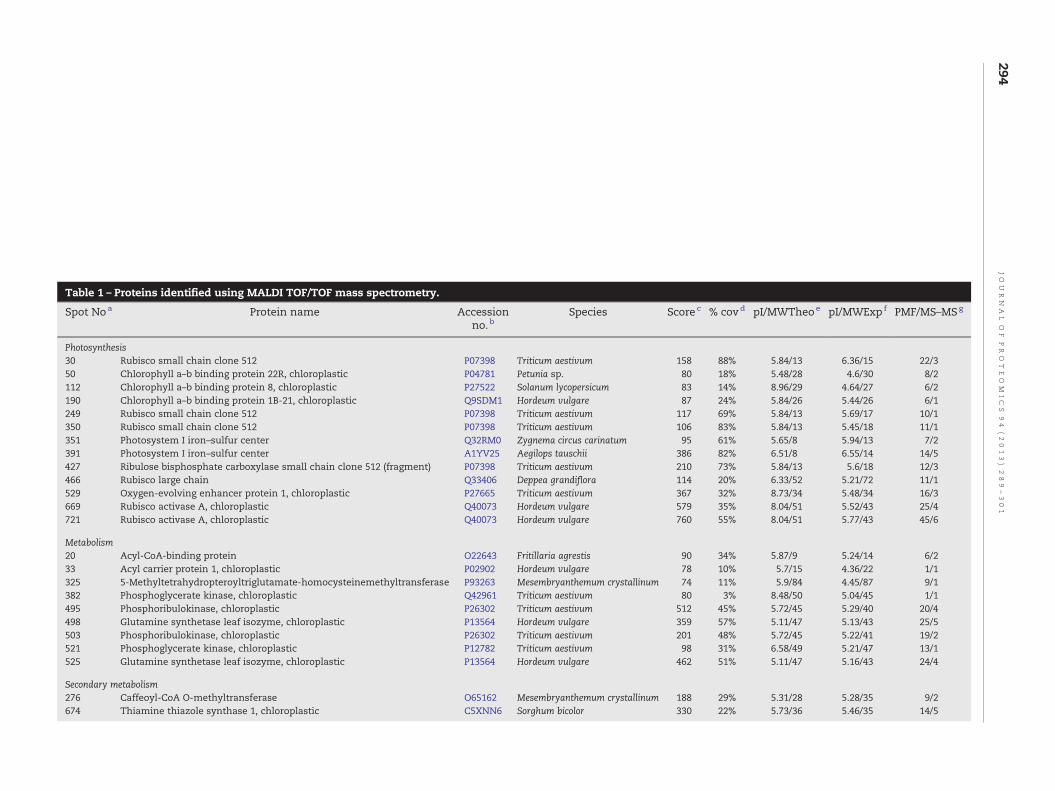
Table 1 – Proteins identified using MALDI TOF/TOF mass spectrometry.
Spot Noa Protein name Accessionno. b
Species Score c % covd pI/MWTheoe pI/MWExp f PMF/MS–MSg
Photosynthesis30 Rubisco small chain clone 512 P07398 Triticum aestivum 158 88% 5.84/13 6.36/15 22/350 Chlorophyll a–b binding protein 22R, chloroplastic P04781 Petunia sp. 80 18% 5.48/28 4.6/30 8/2112 Chlorophyll a–b binding protein 8, chloroplastic P27522 Solanum lycopersicum 83 14% 8.96/29 4.64/27 6/2190 Chlorophyll a–b binding protein 1B-21, chloroplastic Q9SDM1 Hordeum vulgare 87 24% 5.84/26 5.44/26 6/1249 Rubisco small chain clone 512 P07398 Triticum aestivum 117 69% 5.84/13 5.69/17 10/1350 Rubisco small chain clone 512 P07398 Triticum aestivum 106 83% 5.84/13 5.45/18 11/1351 Photosystem I iron–sulfur center Q32RM0 Zygnema circus carinatum 95 61% 5.65/8 5.94/13 7/2391 Photosystem I iron–sulfur center A1YV25 Aegilops tauschii 386 82% 6.51/8 6.55/14 14/5427 Ribulose bisphosphate carboxylase small chain clone 512 (fragment) P07398 Triticum aestivum 210 73% 5.84/13 5.6/18 12/3466 Rubisco large chain Q33406 Deppea grandiflora 114 20% 6.33/52 5.21/72 11/1529 Oxygen-evolving enhancer protein 1, chloroplastic P27665 Triticum aestivum 367 32% 8.73/34 5.48/34 16/3669 Rubisco activase A, chloroplastic Q40073 Hordeum vulgare 579 35% 8.04/51 5.52/43 25/4721 Rubisco activase A, chloroplastic Q40073 Hordeum vulgare 760 55% 8.04/51 5.77/43 45/6
Metabolism20 Acyl-CoA-binding protein O22643 Fritillaria agrestis 90 34% 5.87/9 5.24/14 6/233 Acyl carrier protein 1, chloroplastic P02902 Hordeum vulgare 78 10% 5.7/15 4.36/22 1/1325 5-Methyltetrahydropteroyltriglutamate-homocysteinemethyltransferase P93263 Mesembryanthemum crystallinum 74 11% 5.9/84 4.45/87 9/1382 Phosphoglycerate kinase, chloroplastic Q42961 Triticum aestivum 80 3% 8.48/50 5.04/45 1/1495 Phosphoribulokinase, chloroplastic P26302 Triticum aestivum 512 45% 5.72/45 5.29/40 20/4498 Glutamine synthetase leaf isozyme, chloroplastic P13564 Hordeum vulgare 359 57% 5.11/47 5.13/43 25/5503 Phosphoribulokinase, chloroplastic P26302 Triticum aestivum 201 48% 5.72/45 5.22/41 19/2521 Phosphoglycerate kinase, chloroplastic P12782 Triticum aestivum 98 31% 6.58/49 5.21/47 13/1525 Glutamine synthetase leaf isozyme, chloroplastic P13564 Hordeum vulgare 462 51% 5.11/47 5.16/43 24/4
Secondary metabolism276 Caffeoyl-CoA O-methyltransferase O65162 Mesembryanthemum crystallinum 188 29% 5.31/28 5.28/35 9/2674 Thiamine thiazole synthase 1, chloroplastic C5XNN6 Sorghum bicolor 330 22% 5.73/36 5.46/35 14/5
294JO
UR
NA
LO
FPR
OT
EO
MIC
S94
(2013)
289–301
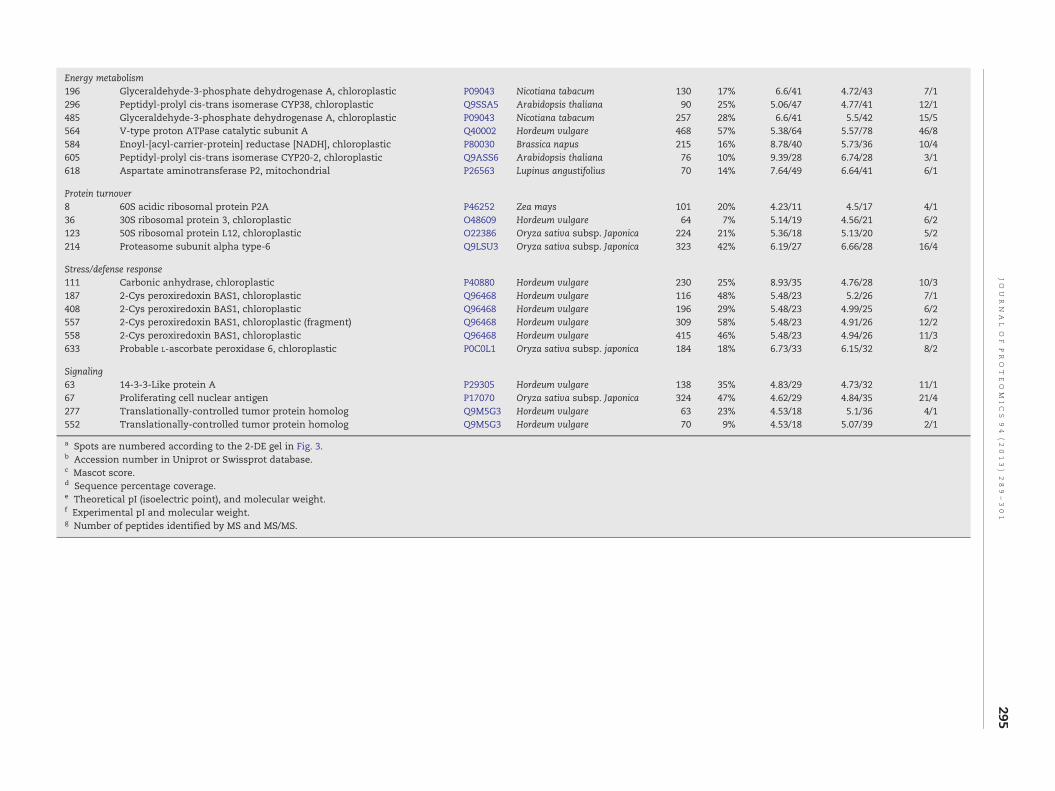
Energy metabolism196 Glyceraldehyde-3-phosphate dehydrogenase A, chloroplastic P09043 Nicotiana tabacum 130 17% 6.6/41 4.72/43 7/1296 Peptidyl-prolyl cis-trans isomerase CYP38, chloroplastic Q9SSA5 Arabidopsis thaliana 90 25% 5.06/47 4.77/41 12/1485 Glyceraldehyde-3-phosphate dehydrogenase A, chloroplastic P09043 Nicotiana tabacum 257 28% 6.6/41 5.5/42 15/5564 V-type proton ATPase catalytic subunit A Q40002 Hordeum vulgare 468 57% 5.38/64 5.57/78 46/8584 Enoyl-[acyl-carrier-protein] reductase [NADH], chloroplastic P80030 Brassica napus 215 16% 8.78/40 5.73/36 10/4605 Peptidyl-prolyl cis-trans isomerase CYP20-2, chloroplastic Q9ASS6 Arabidopsis thaliana 76 10% 9.39/28 6.74/28 3/1618 Aspartate aminotransferase P2, mitochondrial P26563 Lupinus angustifolius 70 14% 7.64/49 6.64/41 6/1
Protein turnover8 60S acidic ribosomal protein P2A P46252 Zea mays 101 20% 4.23/11 4.5/17 4/136 30S ribosomal protein 3, chloroplastic O48609 Hordeum vulgare 64 7% 5.14/19 4.56/21 6/2123 50S ribosomal protein L12, chloroplastic O22386 Oryza sativa subsp. Japonica 224 21% 5.36/18 5.13/20 5/2214 Proteasome subunit alpha type-6 Q9LSU3 Oryza sativa subsp. Japonica 323 42% 6.19/27 6.66/28 16/4
Stress/defense response111 Carbonic anhydrase, chloroplastic P40880 Hordeum vulgare 230 25% 8.93/35 4.76/28 10/3187 2-Cys peroxiredoxin BAS1, chloroplastic Q96468 Hordeum vulgare 116 48% 5.48/23 5.2/26 7/1408 2-Cys peroxiredoxin BAS1, chloroplastic Q96468 Hordeum vulgare 196 29% 5.48/23 4.99/25 6/2557 2-Cys peroxiredoxin BAS1, chloroplastic (fragment) Q96468 Hordeum vulgare 309 58% 5.48/23 4.91/26 12/2558 2-Cys peroxiredoxin BAS1, chloroplastic Q96468 Hordeum vulgare 415 46% 5.48/23 4.94/26 11/3633 Probable L-ascorbate peroxidase 6, chloroplastic P0C0L1 Oryza sativa subsp. japonica 184 18% 6.73/33 6.15/32 8/2
Signaling63 14-3-3-Like protein A P29305 Hordeum vulgare 138 35% 4.83/29 4.73/32 11/167 Proliferating cell nuclear antigen P17070 Oryza sativa subsp. Japonica 324 47% 4.62/29 4.84/35 21/4277 Translationally-controlled tumor protein homolog Q9M5G3 Hordeum vulgare 63 23% 4.53/18 5.1/36 4/1552 Translationally-controlled tumor protein homolog Q9M5G3 Hordeum vulgare 70 9% 4.53/18 5.07/39 2/1
a Spots are numbered according to the 2-DE gel in Fig. 3.b Accession number in Uniprot or Swissprot database.c Mascot score.d Sequence percentage coverage.e Theoretical pI (isoelectric point), and molecular weight.f Experimental pI and molecular weight.g Number of peptides identified by MS and MS/MS.
295JO
UR
NA
LO
FPR
OT
EO
MIC
S94
(2013)
289–301
296 J O U R N A L O F P R O T E O M I C S 9 4 ( 2 0 1 3 ) 2 8 9 – 3 0 1
inoculated with P. indica showed a high degree of rootcolonization under both well-watered and water-deficitconditions. After spore germination, the fungus penetratesepidermal cells and spreads throughout the cortex tissue andspore formed both inter- and intracellularly. P. indica had agrowth-promoting effect on colonized barley plants as theygrow faster, more vigorously and establishedmore tillers thannon-inoculated plants under both well-watered and droughtcondition (Fig. 1). Microscopic analyses showed that P. indicacolonized barley roots efficiently under both control anddrought conditions as measured by production of the numberof chlamydospores at 2 weeks after spore inoculation. Nocolonization was observed in non-inoculated plant roots.P. indica-root colonization was begun with the germinationof chlamydospores that penetrated the root cortex, ram-ifying intercellularly from the point of penetration andsubsequently, formed chlamydospores within cortical cells(Fig. 2). P. indica increased fresh and dry weight ofP. indica-colonized barley plants by 1.23 and 1.46 times higherthan non-inoculated plants under well-watered condition,respectively at 28 days after fungal inoculation (Fig. 3). As weexpected, drought had a negative effect on growth rates ofboth P. indica-colonized barley and non-inoculated plants.Under drought condition (25% F.C.), the shoot fresh and dryweight of P. indica-colonized barley plants were up to 1.70 and1.54 times higher than non-inoculated plants. The root dryweight of P. indica-colonized and non-inoculated plantswere increased by 13% and 5%, respectively under drought
Fig. 6 – Classification of 45 individual barley protein spots showeindica-inoculated (I) and non-inoculated (NI) barley plants only atexpressed as log2 (abundance in drought-treated plants/abundanexpression pattern: (A) up-regulated in both NI and I plants, (B) dand NI, but in opposite directions, (D) up-regulated only in NI planin I plants, (G) down-regulated only in I plants. Solid markers remarkers represent a non-significant difference (p-value > 0.05) innon-inoculated samples.
condition, compared to well-watered condition. P. indicapromoted root growth by 1.58 times higher compared to theroot dry weight of non-inoculated plants under drought stresscondition (Fig. 3). However, the increase in dry root weight ofnon-inoculated plants under drought condition was notsignificant compared to the well-watered condition.
3.2. Proteome responses of barley leaves to drought andP. indica
We analyzed the overall changes in protein patterns ofbarley leaves in response to drought stress and P. indica-rootcolonization. In total, 726 protein spots were detectable insilver-stained gel (Fig. 4). Protein spots with an averageexpression level of 1.5-fold changes were considered asresponsive proteins to drought treatments and in response toP. indica compared to proper control (well-watered and non-inoculated plants). Of these detected spots, 62 protein spotsshowed significant differences in their abundance betweenP. indica-colonized plants compared with non-inoculated plantsin response to drought stress. Protein identification using massspectrometry resulted in identification of 45 proteins (Tables 1and S2). These proteins were grouped in several fundamentalprocesses, including cellular energy metabolism, stress/defenseresponse, photosynthesis, and protein synthesis/assemblyand signaling (Fig. 5). Overall, 37 proteins were differentiallyexpressed in P. indica-colonized barley plants under droughtstress compared to mock-treated barley plants under drought
d statistically significant changes in Piriformosporadrought stress condition compared with control. The ratio isce in control samples). Proteins are grouped according to the
own-regulated in both NI and I plants, (C) modulated in both Its, (E) down-regulated only in NI plants, (F) up-regulated onlypresent a significant difference (p-value r 0.05) and openresponse to salt stress in P. indica-inoculated and

297J O U R N A L O F P R O T E O M I C S 9 4 ( 2 0 1 3 ) 2 8 9 – 3 0 1
treatments. This suggests that the differentially expressedproteins in P. indica-colonized barley plants might play roles inP. indica-mediated drought stress tolerance.
3.3. Identification and grouping of drought and P. indicaresponsive proteins
Forty-five identified proteins were divided into seven majorcategories (Fig. 6): (A) the abundance of three proteins wasincreased in both non-inoculated and P. indica-colonized barleyplants under drought stress. These proteins included Rubiscosmall chain (spot 249), caffeoyl-CoA O-methyltransferase(CCOMT) (spot 276) and acyl-CoA-binding protein (spot 20).In this group, the abundance of the acyl-CoA-binding proteinwas more induced in the P. indica-colonized plants; (B) theabundance of three proteins reduced in both the P. indica-colonized and the non-inoculated plants. These includetwo phosphoribulokinases (PRKs) (spots 503 and 495) and acylcarrier protein (spot 33); (C) the abundance of glyceraldehyde-3-phosphate dehydrogenase (spot 196) and proliferating cellnuclear antigen (PCNA) (protein spot 67) changed in oppositedirections in P. indica-colonized and non-inoculated plants;(D) the abundance of phosphoglycerate kinase (spot 521),enoyl-[acyl-carrier-protein] reductase (spot 584), aspartateaminotransferase (spot 618), and Rubisco small chain (proteinspot 350) increased only in non-inoculated barley plants; (E) theabundance of four proteins included photosystem I iron–sulfurcenter (spot 391), chlorophyll a–b binding protein (spot 50),60S acidic ribosomal protein (spot 8), and 50S ribosomal protein(spot 123) were decreased in response of drought treatment.Nonetheless, the abundance of two proteins of this groupreturned to control levels after P. indica-inoculation; (F) theabundance of 21 proteins including oxygen-evolving enhancerprotein (spot 529), V-type proton ATPase (spot 564), probableL-ascorbate peroxidase (APX) (spot 633), 2-Cys peroxiredoxin(2-Cys Prx) (spots 187, 408, 557 and 558), 5-methyltetra-hydropteroyltriglutamate (spot 325), Rubisco small chain(spots 30 and 427), 30S ribosomal protein 3 (spot 36), carbonicanhydrase (spot 111), photosystem I iron–sulfur center (spot351), translationally-controlled tumor protein (TCTP) (spots 277and 552), proteasome subunit alpha type-6 (spot 214), chlo-rophyll a–b binding protein (spots 190 and 112), peptidyl-prolylcis-trans isomerase (spot 605), phosphoglycerate kinase (spot466) and Rubisco large chain (spot 408) were increased only inP. indica-colonized barley plants; (G) The abundance of eightproteins including peptidyl-prolyl cis-trans isomerase (spot296), Rubisco activase (spots 669 and 721), thiamine thiazolesynthase (protein spot 674), glutamine synthetase leaf isozyme(spots 498 and 525), 14-3-3-like protein A (spot 63), andglyceraldehyde-3-phosphate dehydrogenase (spot 485) werereduced only in P. indica-colonized barley plants (Fig. 6).
3.4. Correlation between RNA transcript and protein levels
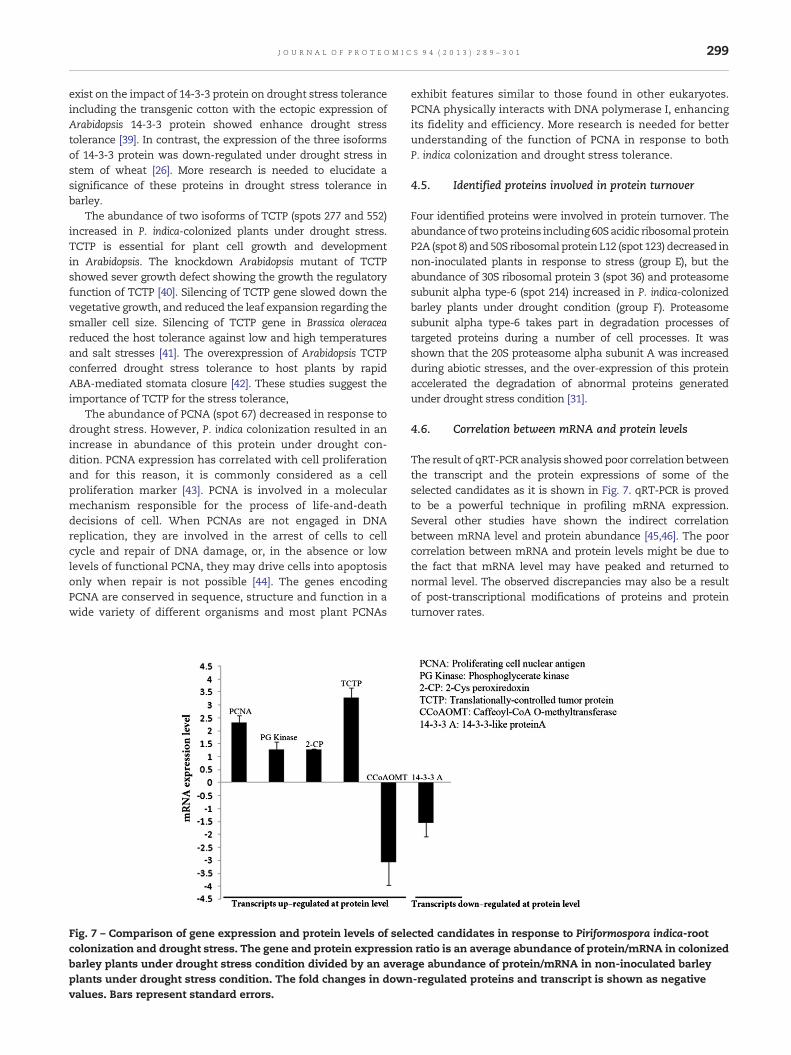
We analyzed the gene expression profile of six selectedproteins responsive to P. indica-root colonization. Based onthe proteomics analysis, all of these selected proteins weresignificantly altered at protein level (Fig. 6). As shown in Fig. 6,the direct correlation between gene expression and protein levelwas found only for the TCTP (spot 552) and PCNA (spot 67).
However, three other genes encoding 14-3-3 like protein (spot63), PG kinase protein (spot 382) and 2-Cys Prx (spot 408) showedno significant changes at transcript level. Surprisingly, therewasdrastic differences in the protein andmRNA expression levels ofCCOMT (spot 276), as the mRNA was down-regulated while theprotein level was up-regulated.
4. Discussion
In present study, we used integrated physiology and proteomicapproach to study the mechanisms underlying P. indica-mediated water stress tolerance in barley. P. indica improveddrought stress tolerance in barley and the comparativeproteomic analyses led to the identification of 45 responsiveproteins involved in photosynthesis, metabolism, oxidativestress defense, and signaling pathway.
4.1. Proteins involved in photosynthesis
In total, we found 13 differentially accumulated proteinsinvolved in photosynthesis. These proteins included Rubiscolarge subunit (spot 466), Rubisco small subunits (spots 30, 249,350 and 427), Rubisco activase (spots 669 and 721), oxygen-evolving enhancer protein (spot 529), chlorophyll a–b bindingprotein (spots 50, 112 and 190), and photosystem I iron–sulfurcenter (spots 351 and 391). We observed that the abundanceof ten photosynthetic proteins was changed in a similardirection in both P. indica-colonized and non-inoculatedplants. Drought decreases the expression level of a large numberof proteins involved in the photosynthesis pathways [22].Specifically, the protein levels of several photosyntheticproteins in groups E and F were decreased under droughtstress condition, but the protein level of most of theseproteins were increased in P. indica-colonized plants (Fig. 6,group F). These results suggest that the growth promotingeffect of P. indica especially under stress condition mightbe associated to photosynthesis stimulation under droughttreatment.
4.2. Proteins involved in cellmetabolismand energy transport
Eighteen of identified proteins were associated to cellularmetabolism (9 proteins), energy metabolism (7 proteins), andsecondary metabolism (2 proteins). Among these proteins,protein spots 8 and 4 were decreased in P. indica-colonized andnon-inoculated plants, respectively. The expressions of proteins7 and 4 were increased in inoculated and non-inoculatedplants, respectively (Fig. 6). One of the identified proteins was5-methyltetrahydropteroyltriglutamate homocysteine methyltransferase (spot 325). This enzyme transfers a methyl group of5-methyltetrahydrofolate to homocysteine resulting in produc-tion of methionine. The activation of methyl cycle is importantin water stress in producing methylated polyols [23]. Theexpression of methionine synthase was increased in barleyunder various abiotic stresses [24]. P. indica-colonized barleyplants showed increased in the expression level of theseproteins under drought stress condition. Based on this data, wespeculate that P. indica activates the methyl cycle in response todrought stress in colonized plants.

298 J O U R N A L O F P R O T E O M I C S 9 4 ( 2 0 1 3 ) 2 8 9 – 3 0 1
We identified two isoforms of phosphoglycerate kinase inwhich one isoform (spot 521) was up-regulated under droughtstress and another isoform (spot 382) was up-regulated inP. indica-colonized plants. In vitro activities of these enzymesin pigeon pea showed that the phosphoglycerate kinase wasdecreased under mild stress, but during increased in waterstress the activity of phosphoglycerate kinase was enhanced[25]. It was shown that the expression of phosphoglyceratekinase increased under drought stress from 30 days afteranthesis in wheat stem [26].
The abundance of two isoforms of PRKs decreased in bothP. indica-colonized and non-inoculated plants under droughtcondition. PRK catalyzes the ATP-dependent phosphorylationof ribulose-5-phosphate to ribulose-1,5-phosphate, a keystep in the pentose phosphate pathway. PRK is especiallyimportant in photosynthesis and the reduction in amounts ofPRK has been associated with the reduction of plant biomassand photosynthetic capacity [27]. Decreased in abundance ofPRK in our study might also be associated with reduction inthe barley biomass and growth under drought stress.
V-type proton ATPase catalytic subunit A fragment(protein spot 564) is another identified protein. V-ATPasesare responsible for generating energy for transportation ofions and metabolites. V-ATPases play a critical role in stressresponse [28]. V-ATPases play a major role in salt adaptation byexclusion of excess sodium from cytoplasmic compartment andaccumulates in vacuoles. V-ATPase also responds to a varietyof other stresses such as drought [29]. The abundance of thisprotein remained unchanged in non-inoculated plants underdrought stress compared to well-watered plants, but P. indicaincreased the expression of this protein under drought stress.Increased abundance of this protein in P. indica-colonized barleyplants under drought stressmight result in higher adaptability ofP. indica-colonized plants in osmoregulation processes.
We identified peptidyl-prolyl cis-trans isomerase CYP20,which remained unchanged under drought stress condition,but its expression was up-regulated in P. indica-colonizedplants. The expression of CYP20 gene was induced inArabidopsis in response to Pseudomonas syringae infection [28].The higher abundance of CYP20 P. indica-colonized plant maysuggest its involvement in a compatible interaction betweenP. indica and barley.
The abundance of CCOMT increased in P. indica-colonizedbarley plants under drought stress condition. It has beendemonstrated that CCOMT plays an essential role in thesynthesis of guaiacyl lignin units as well as in the supply ofsubstrates for the synthesis of syringyl lignin units [30]. Verylittle is known about the effects of drought on lignin bio-synthesis. CCOMT may control the sensitivity of severalregulatory genes specific to short exposure to osmotic stress [31].
4.3. Proteins involved in oxidative stress defense
Six of the forty-five identified proteins were enzymes involvedin stress defense proteins. Reactive oxygen species (ROS)takes part in the regulation of many different cellularprocesses. ROS is required as substrates and signals in cellmetabolism, growth, and differentiation in low concentration.It participates in triggering changes in gene expression, butalso can cause damage to macromolecules and membranes at
elevated concentrations. Consequently, each cell has amulti-level and cross-talking defense system against oxida-tive damage counteracting with antioxidants. One isoform ofcarbonic anhydrase (spot 111) down-regulated under droughtstress, but P. indica increased the abundance of thisproteinunderdrought stress. Carbonic anhydrase is zinc-containing enzymecatalyzing the reversible hydration of CO2and plays animportant role in facilitating the diffusion of CO2 in photo-synthesis [32]. Plants exposed to stress exhibited a reduction ingrowth, photosynthesis, and the activity of nitrate reductase andcarbonic anhydrase. Transgenic Arabidopsis overexpressing thegene encoding carbonic anhydrase showed increased salt stresstolerance at seedling stage in rice [33].
The expression of APX (spot 633) was up-regulated inP. indica-colonized barley plants under drought treatment. TheAPXs belong to one of the most important enzymatic groupsof antioxidant in plants. APXs are scavengers of H2O2, whichgenerates continuously in the cells. The expression of APXgenes can be activated by specific factors such as mechanicaldamage, water deficiency, and salt stress. APX also plays animportant role in controlling the concentration of oxygenradicals that participate in signal transduction in manynaturally occurring physiological processes in cell. Thefunction of APXs is not only limited to antioxidativeprotection, but also connected with the regulation of hostmetabolism. Several studies have shown that overexpressionof APX genes in transgenic plants enhanced tolerance tooxidative and water deficit stresses [34].
We also identified four isoforms of 2-Cys Prx (spots 187, 408,557 and 558). Interestingly, P. indica increased significantly theexpression of these isoforms under drought stress condition.2-Cys Prx are highly conserved and abundant antioxidantenzymes that catalyze the breakdown of peroxides to protectcells from oxidative stress [35]. Peroxiredoxins are involved inthe detoxification of cell-toxic peroxides and the activity ofperoxiredoxin in general, and chloroplast 2-Cys Prx in particular,affect fatty acid signaling by compounds such as traumatin andjasmonic acid. Several independent studies revealed thatP. indica-root colonization associated with altered antioxidativecapacities in host plants [36]. We concluded that P. indicaincreased the antioxidative capacity and enhanced droughtstress tolerance in colonized plants. It has been demonstratedthat Trichoderma harzianum, an efficient root endophyte, protectsplant host against the detrimental effects of drought byenhanced antioxidant activities [37]. Our results suggest thatenhanced foliar antioxidant capacity under stress condition inP. indicaunder stressmay play a role to improve plant adaptationto stress.
4.4. Identified proteins involved in cell signaling andproliferation
One of the identified drought-responsive proteins is 14-3-3-likeprotein A (spot 63), which is involved in cell signaling. Theprotein expression level of this protein was decreased inP. indica-colonized plants under drought stress response. 14-3-3-like protein A is a highly conserved protein among plat speciesand plays key roles inmany plant biological processes includingresponding to abiotic and biotic stress either by signaling,transcription activation, or defense [38]. However, a few reports
299J O U R N A L O F P R O T E O M I C S 9 4 ( 2 0 1 3 ) 2 8 9 – 3 0 1
exist on the impact of 14-3-3 protein on drought stress toleranceincluding the transgenic cotton with the ectopic expression ofArabidopsis 14-3-3 protein showed enhance drought stresstolerance [39]. In contrast, the expression of the three isoformsof 14-3-3 protein was down-regulated under drought stress instem of wheat [26]. More research is needed to elucidate asignificance of these proteins in drought stress tolerance inbarley.
The abundance of two isoforms of TCTP (spots 277 and 552)increased in P. indica-colonized plants under drought stress.TCTP is essential for plant cell growth and developmentin Arabidopsis. The knockdown Arabidopsis mutant of TCTPshowed sever growth defect showing the growth the regulatoryfunction of TCTP [40]. Silencing of TCTP gene slowed down thevegetative growth, and reduced the leaf expansion regarding thesmaller cell size. Silencing of TCTP gene in Brassica oleraceareduced the host tolerance against low and high temperaturesand salt stresses [41]. The overexpression of Arabidopsis TCTPconferred drought stress tolerance to host plants by rapidABA-mediated stomata closure [42]. These studies suggest theimportance of TCTP for the stress tolerance,
The abundance of PCNA (spot 67) decreased in response todrought stress. However, P. indica colonization resulted in anincrease in abundance of this protein under drought con-dition. PCNA expression has correlated with cell proliferationand for this reason, it is commonly considered as a cellproliferation marker [43]. PCNA is involved in a molecularmechanism responsible for the process of life-and-deathdecisions of cell. When PCNAs are not engaged in DNAreplication, they are involved in the arrest of cells to cellcycle and repair of DNA damage, or, in the absence or lowlevels of functional PCNA, they may drive cells into apoptosisonly when repair is not possible [44]. The genes encodingPCNA are conserved in sequence, structure and function in awide variety of different organisms and most plant PCNAs
Fig. 7 – Comparison of gene expression and protein levels of selecolonization and drought stress. The gene and protein expressionbarley plants under drought stress condition divided by an averaplants under drought stress condition. The fold changes in downvalues. Bars represent standard errors.
exhibit features similar to those found in other eukaryotes.PCNA physically interacts with DNA polymerase I, enhancingits fidelity and efficiency. More research is needed for betterunderstanding of the function of PCNA in response to bothP. indica colonization and drought stress tolerance.
4.5. Identified proteins involved in protein turnover
Four identified proteins were involved in protein turnover. Theabundanceof twoproteins including60Sacidic ribosomalproteinP2A (spot 8) and 50S ribosomal protein L12 (spot 123) decreased innon-inoculated plants in response to stress (group E), but theabundance of 30S ribosomal protein 3 (spot 36) and proteasomesubunit alpha type-6 (spot 214) increased in P. indica-colonizedbarley plants under drought condition (group F). Proteasomesubunit alpha type-6 takes part in degradation processes oftargeted proteins during a number of cell processes. It wasshown that the 20S proteasome alpha subunit A was increasedduring abiotic stresses, and the over-expression of this proteinaccelerated the degradation of abnormal proteins generatedunder drought stress condition [31].
4.6. Correlation between mRNA and protein levels
The result of qRT-PCR analysis showedpoor correlation betweenthe transcript and the protein expressions of some of theselected candidates as it is shown in Fig. 7. qRT-PCR is provedto be a powerful technique in profiling mRNA expression.Several other studies have shown the indirect correlationbetween mRNA level and protein abundance [45,46]. The poorcorrelation between mRNA and protein levels might be due tothe fact that mRNA level may have peaked and returned tonormal level. The observed discrepancies may also be a resultof post-transcriptional modifications of proteins and proteinturnover rates.
cted candidates in response to Piriformospora indica-rootratio is an average abundance of protein/mRNA in colonizedge abundance of protein/mRNA in non-inoculated barley-regulated proteins and transcript is shown as negative

300 J O U R N A L O F P R O T E O M I C S 9 4 ( 2 0 1 3 ) 2 8 9 – 3 0 1
5. Conclusion
Plant mutualistic symbionts offer long-term abiotic stresstolerance through the host adaptation to environmental stress[4]. There have been a few published proteomic studies of plantsymbionts to drought, and this is thought to be the firstproteomic analysis, demonstrating the impact of endophyte onbarley plant under drought stress. We showed the involvementof some of the previously known proteins in the host adaptationto drought stress. Notably, for someof the identified proteins likeTCTP and PCNA, a connection to physiological function in plantsis novel, and can be the best candidates for sources of droughttolerance in future studies. According to our results, and becausethe fungus does not infest leaves, it is likely that P. indica inducessystemic response to drought stress. P. indica has a clearadvantage over the manipulation of individual componentsunder drought and other abiotic stress tolerance, since it targetsseveral drought related genes simultaneously, and has noobvious side effects for biotechnological application. It is likelythat a more comprehensive analysis will help to unravel themechanisms by which P. indica in targeted host drought-tolerance mechanism.
Supplementary data to this article can be found online athttp://dx.doi.org/10.1016/j.jprot.2013.09.017.
R E F E R E N C E S
[1] Rodriguez R, Redman R. More than 400 million years ofevolution and some plants still can't make it on their own:plant stress tolerance via fungal symbiosis. J Exp Bot2008;59:1109–14.
[2] Harrison MJ. Signaling in the arbuscular mycorrhizalsymbiosis. Annu Rev Microbiol 2005;59:19–42.
[3] Schulz B, Boyle C. The endophytic continuum. Mycol Res2005;109:661–86.
[4] Harman GE. Multifunctional fungal plant symbionts: newtools to enhance plant growth and productivity. New Phytol2011;189:647–9.
[5] Weiss M, Selosse MA, Rexer KH, Urban A, Oberwinkler F.Sebacinales: a hitherto overlooked cosm ofheterobasidiomycetes with a broad mycorrhizal potential.Mycol Res 2004;108:1003–10.
[6] Verma S, Varma A, Rexer KH, Hassel A, Kost G, Sarbhoy A,et al. Piriformospora indica, gen. et sp. nov., a newroot-colonizing fungus. Mycologia 1998;90:896–903.
[7] Waller F, Achatz B, Baltruschat H, Fodor J, Becker K, Fischer M,et al. The endophytic fungus Piriformospora indicareprograms barley to salt-stress tolerance, diseaseresistance, and higher yield. Proc Natl Acad Sci U S A2005;102:13386–91.
[8] Sherameti I, Shahollari B, Venus Y, Altschmied L, Varma A,Oelmüller R. The endophytic fungus Piriformospora indicastimulates the expression of nitrate reductase and thestarch-degrading enzyme glucan-water dikinase in tobaccoand Arabidopsis roots through a homeodomain transcriptionfactor that binds to a conserved motif in their promoters.J Biol Chem 2005;280:26241–7.
[9] Deshmukh S, Hückelhoven R, Schäfer P, Imani J, Sharma M,Weiss M, et al. The root endophytic fungus Piriformosporaindica requires host cell death for proliferation duringmutualistic symbiosis with barley. Proc Natl Acad Sci U S A2006;103:18450–7.
[10] Shoresh M, Harman GE. The molecular basis of shootresponses of maize seedlings to Trichoderma harzianum T22inoculation of the root: a proteomic approach. Plant Physiol2008;147:2147–63.
[11] Kosova K, Vitamvas P, Prasil IT, Renaut J. Plant proteomechanges under abiotic stress—contribution of proteomicsstudies to understanding plant stress response. J Proteome2011;74:1301–22.
[12] Hossain Z, Nouri MZ, Komatsu S. Plant cell organelleproteomics in response to abiotic stress. J Proteome Res2012;11:37–48.
[13] Jorrín-Novo JV, Maldonado AM, Echevarría-Zomeño S,Valledor L, Castillejo MA, Curto M, et al. Second-generationproteomic techniques, an appropriate experimental design,and data analysis to fulfill MIAPE standards, increase plantproteome coverage and expand biological knowledge.J Proteome 2009;72:285–314.
[14] Sun C, Johnson JM, Cai D, Sherameti I, Oelmüller R, Lou B.Piriformospora indica confers drought tolerance in Chinesecabbage leaves by stimulating antioxidant enzymes, theexpression of drought-related genes and the plastid-localizedCAS protein. J Plant Physiol 2010;167:1009–17.
[15] Sherameti I, Tripathi S, Varma A, Oelmüller R. Theroot-colonizing endophyte Piriformospora indica confersdrought tolerance in Arabidopsis by stimulating theexpression of drought stress-related genes in leaves. MolPlant-Microbe Interact 2008;21:799–807.
[16] Palmieri MC, Perazzolli M, Matafora V, Moretto M, Bachi A,Pertot I. Proteomic analysis of grapevine resistance inducedby Trichoderma harzianum T39 reveals specific defencepathways activated against downy mildew. J Exp Bot2012;63:6237–51.
[17] Alikhani M, Khatabi B, Sepehri M, Nekouei MK, Mardi M,Salekdeh GH. A proteomics approach to study the molecularbasis of enhanced salt tolerance in barley (Hordeum vulgare L.)conferred by the root mutualistic fungus Piriformospora indica.Mol Biosyst 2013;9:1498–510.
[18] Sarhadi E, Mahfoozi S, Hosseini SA, Salekdeh GH. Coldacclimation proteome analysis reveals close link betweenthe up-regulation of low-temperature associatedproteins and vernalization fulfillment. J Proteome Res2010;9:5658–67.
[19] Blum H, Beier H, Gross HJ. Improved silver staining of plantproteins, RNA and DNA in polyacrylamide gels.Electrophoresis 1987;8:93–9.
[20] Neuhoff V, Arold N, Taube D, Ehrhardt W. Improved stainingof proteins in polyacrylamide gels including isoelectricfocusing gels with clear background at nanogram sensitivityusing Coomassie Brilliant Blue G‐250 and R‐250.Electrophoresis 1988;9:255–62.
[21] Schmittgen TD, Livak KJ. Analyzing real-time PCR data by thecomparative CT method. Nat Protoc 2008;3:1101–8.
[22] Hajheidari M, Abdollahian‐Noghabi M, Askari H, Heidari M,Sadeghian SY, Ober ES, et al. Proteome analysis of sugar beetleaves under drought stress. Proteomics 2005;5:950–60.
[23] Bohnert HJ, Jensen RG. Strategies for engineering water-stresstolerance in plants. Trends Biotechnol 1996;14:89–97.
[24] Narita Y, Taguchi H, Nakamura T, Ueda A, Shi W, Takabe T.Characterization of the salt-inducible methionine synthasefrom barley leaves. Plant Sci 2004;167:1009–16.
[25] Singal H, Sheoran I, Singh R. Effect of water stress onphotosynthesis and in vitro activities of the PCR cycleenzymes in pigeonpea (Cajanus cajan L.). Photosynth Res1985;7:69–76.
[26] Bazargani MM, Sarhadi E, Bushehri AAS, Matros A, Mock HP,Naghavi MR, et al. A proteomics view on the role ofdrought-induced senescence and oxidative stress defense inenhanced stem reserves remobilization in wheat. J Proteome2011;74:1959–73.

301J O U R N A L O F P R O T E O M I C S 9 4 ( 2 0 1 3 ) 2 8 9 – 3 0 1
[27] Banks FM, Driscoll SP, Parry MAJ, Lawlor DW, Knight JS, GrayJC, et al. Decrease in phosphoribulokinase activity byantisense RNA in transgenic tobacco. Relationship betweenphotosynthesis, growth, and allocation at different nitrogenlevels. Plant Physiol 1999;119:1125–36.
[28] Jones AME, Thomas V, Bennett MH, Mansfield J, Grant M.Modifications to the Arabidopsis defense proteome occur priorto significant transcriptional change in response toinoculation with Pseudomonas syringae. Plant Physiol2006;142:1603–20.
[29] Tsiantis MS, Bartholomew DM, Smith JAC. Salt regulation oftranscript levels for the c subunit of a leaf vacuolar H+-ATPasein the halophyte Mesembryanthemum crystallinum. Plant J1996;9:729–36.
[30] Ye Z-H, Zhong R, Morrison Iii WH, Himmelsbach DS. Caffeoylcoenzyme A O-methyltransferase and lignin biosynthesis.Phytochemistry 2001;57:1177–85.
[31] Toorchi M, Yukawa K, Nouri MZ, Komatsu S. Proteomicsapproach for identifying osmotic-stress-related proteins insoybean roots. Peptides 2009;30:2108–17.
[32] Lu YK, Stemler AJ. Extrinsic photosystem II carbonicanhydrase in maize mesophyll chloroplasts. Plant Physiol2002;128:643–9.
[33] Yu S, Zhang X, Guan Q, Takano T, Liu S. Expression of acarbonic anhydrase gene is induced by environmentalstresses in Rice (Oryza sativa L.). Biotechnol Lett2007;29:89–94.
[34] Wang Y, Wisniewski M, Meilan R, Cui M, Webb R, FuchigamiL. Overexpression of cytosolic ascorbate peroxidase in tomatoconfers tolerance to chilling and salt stress. J Am Soc HorticSci 2005;130:167–73.
[35] Veal EA, Findlay VJ, Day AM, Bozonet SM, Evans JM, Quinn J,et al. A 2-Cys peroxiredoxin regulates peroxide-inducedoxidation and activation of a stress-activated MAP kinase.Mol Cell 2004;15:129–39.
[36] Baltruschat H, Fodor J, Harrach BD, Niemczyk E, Barna B,Gullner G, et al. Salt tolerance of barley induced by the rootendophyte Piriformospora indica is associated with a strongincrease in antioxidants. New Phytol 2008;180:501–10.
[37] Mastouri F, Björkman T, Harman GE. Seed treatment withTrichoderma harzianum alleviates biotic, abiotic, andphysiological stresses in germinating seeds and seedlings.Phytopathology 2010;100:1213–21.
[38] Chen F, Li Q, Sun L, He Z. The rice 14-3-3 gene family and itsinvolvement in responses to biotic and abiotic stress. DNARes 2006;13:53–63.
[39] Yan J, He C, Wang J, Mao Z, Holaday SA, Allen RD, et al.Overexpression of the Arabidopsis 14-3-3 protein GF14λ incotton leads to a “stay-green” phenotype and improves stresstolerance under moderate drought conditions. Plant CellPhysiol 2004;45:1007–14.
[40] Berkowitz O, Jost R, Pollmann S, Masle J. Characterization ofTCTP, the translationally controlled tumor protein, fromArabidopsis thaliana. Plant Cell 2008;20:3430–47.
[41] Cao B, Lu Y, Chen G, Lei J. Functional characterization of thetranslationally controlled tumor protein (TCTP) geneassociated with growth and defense response in cabbage.Plant Cell Tiss Org Cult 2010;103:217–26.
[42] Kim YM, Han YJ, Hwang OJ, Lee SS, Shin AY, Kim SY, et al.Overexpression of Arabidopsis translationally controlledtumor protein gene AtTCTP enhances drought tolerance withrapid ABA-induced stomatal closure. Mol cells 2012:1–10.
[43] Pawlikowski M, Gruszka A, Kurnatowska I, Winczyk K,Kunert-Radek J, Radek A. Proliferating cell nuclear antigen(PCNA) expression in pituitary adenomas: relationship to theendocrine phenotype of adenoma. Folia Histochem Cytobiol2006;44:37–41.
[44] Paunesku T, Mittal S, Protic M, Oryhon J, Korolev S,Joachimiak A, et al. Proliferating cell nuclear antigen (PCNA):ringmaster of the genome. Int J Radiat Biol 2001;77:1007–21.
[45] Torabi S, Wissuwa M, Heidari M, Naghavi MR, Gilany K,Hajirezaei MR, et al. A comparative proteome approach todecipher the mechanism of rice adaptation to phosphorousdeficiency. Proteomics 2009;9:159–70.
[46] Taheri F, Nematzadeh G, Zamharir MG, Nekouei MK, NaghaviM, Mardi M, et al. Proteomic analysis of the Mexican lime treeresponse to “Candidatus Phytoplasma aurantifolia” infection.Mol Biosyst 2011;7:3028–35.





























