Embed Size (px)
Citation preview

DATASET BRIEF
Proteomic analysis of chicory root identifies proteins
typically involved in cold acclimation
Herve Degand1, Anne-Marie Faber1, Nicolas Dauchot2, Dominique Mingeot3,Bernard Watillon3, Pierre Van Cutsem2, Pierre Morsomme1 and Marc Boutry1
1 Universite catholique de Louvain, Institut des Sciences de la Vie, Louvain-la-Neuve, Belgium2 Unite de Recherche en Biologie Cellulaire et Moleculaire Vegetale, Facultes Universitaires Notre-Dame de la Paix
Namur, Namur, Belgium3 Centre wallon de Recherches agronomiques, Biotechnology Department, Gembloux, Belgium
Received: September 18, 2008
Revised: January 21, 2009
Accepted: January 24, 2009
Chicory (Cichorium intybus) roots contain high amounts of inulin, a fructose polymer used as
a storage carbohydrate by the plant and as a human dietary and prebiotic compound. We
performed 2-D electrophoretic analysis of proteins from root material before the first freezing
period. The proteins were digested with trypsin and the peptides analyzed by MS (MALDI-
TOF/TOF). From the 881 protein spots analyzed, 714 proteins corresponded to a database
accession, 619 of which were classified into functional categories. Besides expected proteins
(e.g. related to metabolism, energy, protein synthesis, or cell structure), other well-represented
categories were proteins related to folding and stability (49 spots), proteolysis (49 spots), and
the stress response (67 spots). The importance of abiotic stress response was confirmed by the
observation that 7 of the 21 most intense protein spots are known to be involved in cold
acclimation. These results suggest a major effect of the low temperature period that preceded
root harvesting.
Keywords:
Abiotic stress / Anaerobic stress / Oxidative stress / Vegetative storage protein
Chicory (Cichorium intybus) is a biannual crop plant. During
the first year, the roots accumulate inulin-type fructans that
are soluble fructose polymers (mainly b (2-1)). They are
not digested and thus represent interesting dietary
compounds that cause significant changes in the composi-
tion of the gut microflora and can thus be considered as a
prebiotic [1].
At the end of the growing season, inulin is the most
abundant storage carbohydrate molecule in chicory root (up
to 20% of the root fresh weight). In chicory, fructans have
been proposed to serve as an energy reserve (chicory
contains little starch) and osmolyte source and also to play a
role during abiotic stress [1].
Although some major proteins involved in inulin meta-
bolism in chicory roots have been characterized [1] little is
known about other proteins typical of chicory roots. In
particular, as a biannual crop plant, chicory goes through a
period of cold and freezing temperatures, which induces
expression of proteins that protect the plant against cold
stress. Indeed, proteomic analyses of different species have
shown an increased abundance of several classes of proteins
[2]. Data concerning the proteins involved in the cold stress
response of chicory are limited. A few enzymes, e.g. fructan
1-exohydrolase [3] and pectin acetylesterase [4], have been
shown to be synthesized in chicory in higher amounts
during cold stress. However, no comprehensive analysis of
the proteins expressed in chicory roots has been performed.
In this report, we analyzed the proteome complement of a
chicory root (Sample P, Supporting Information Fig. 1)
harvested after a period with several days with a minimum
temperature below 51C and before a period with freezing
Abbreviations: 1-FFT, fructan:fructan 1-fructosyl transferase;
G3PDH, glyceraldehyde 3-phosphate dehydrogenase; VSP,
vegetative storage protein
Correspondence: Dr. Marc Boutry, Universite catholique de
Louvain, Institut des Sciences de la Vie, Croix du Sud 5–15, 1348
Louvain-la-Neuve, Belgium
E-mail: [email protected]
Fax: 132-10-473872
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com
Proteomics 2009, 9, 2903–2907 2903DOI 10.1002/pmic.200800744

temperatures. Because chicory is not a clonal variety, we
performed protein identification on a single root, thus
avoiding mixing roots with possibly different genotypes.
Root proteins were analyzed by 2-DE (Fig. 1). From this
gel, 881 protein spots were excised and digested with trypsin
and the peptides analyzed by MALDI-TOF/TOF. Identifica-
tion of the proteins (Supporting Information Fig. 2 and
Supporting Information Table 1) was performed in two ways.
We recently reported an EST database of 12 226 EST clones
from chicory, most of them from root tissues [5]; hence,
the MS data were compared with these EST data and the
proteins identified according to the EST annotation. A
parallel search was performed using the NCBI database
restricted to green plants. The majority of the spots were
identified by both methods. In most of these cases, the same
protein was identified although the accession numbers, and
sometimes the protein names, were different. This is
most likely due to the differences between these approaches:
comparison with the NCBI databases was based on
tryptic fragments scattered all along the protein while
comparison with the EST database was restricted to the
C-terminal region of the protein for which the sequence is
available. In some cases, identification did not match between
both methods. However, in most of these cases, inspection of
the MS data indicated that both proteins were identified by
each approach, but in a different order of score (only the
highest score is displayed in Supporting Information Table
1). This is a strong indication of the presence of two distinct
proteins in the same spot. Finally, 26% of the spots were not
identified, either because MS data were available for only a
very few peptides or because protein identification gave too
low a score.
Functional classification of the proteins was performed
according to [6] (Supporting Information Table 1). The
distribution of the identified proteins according to this clas-
sification is shown in Supporting Information Table 2. Not
surprisingly, the most highly represented categories were
‘‘metabolism’’ and ‘‘energy’’. Of these, the majority were
involved in sugar/polysaccharide metabolism or glycolysis,
respectively. The vast majority (98/105) of the proteins in the
category ‘‘Protein destination and storage’’ were subcategor-
ized into ‘‘Folding and stability’’ (49 proteins) or ‘‘Proteolysis’’
(49 proteins). Another well-represented subcategory was
‘‘Stress responses’’ (67 proteins).
The identity of 21 intense spots (labeled in Fig. 1) is
reported in Table 1. The most intense spots (452, 473, and
485 in Fig. 1) were very close together and were identified as
fructan:fructan 1-fructosyl transferase (1-FFT). This enzyme
is involved in chain elongation of inulin-type fructans, but
can also use sucrose as an acceptor for fructosyl units from
larger fructans, resulting in shorter polymers and an
increased fructan concentration. This might contribute to
cold resistance (discussed in [1]). Whether these spots
represent different products of different genes or different
post-translational modifications of the same protein cannot
yet be unambiguously determined, since the chicory
genome has not yet been sequenced. However, currently,
extensive investigation of 1-FFT polymorphism or chicory
EST library data mining does not substantiate the existence
of a 1-FFT multigenic family (N. Dauchot and P. Van
Cutsem, personal communication).
Another protein represented as several spots (circled in
Fig. 2A), including three intense ones (962, 966, and 968), is
the vegetative storage protein (VSP), which is thought to
Figure 1. 2-DE image of chicory root proteins.
Sample P (Supporting Information Fig. 1) was
analyzed as indicated in the Supporting
Information Materials and methods. The
most abundant spots are circled and their
identification number indicated. Their func-
tional category is defined in Table 1. The
dotted rectangles and the letters indicate the
regions shown in detail in the corresponding
panels in Fig. 2.
2904 H. Degand et al. Proteomics 2009, 9, 2903–2907
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

play a role in the storage of nitrogen. Several VSP spots
were previously identified on 2-D gel analysis and
considered as resulting from post-translational modifica-
tions of a unique protein [7, 8]. However, examination of the
chicory EST databases indicated that seven different
sequences presenting significant identity with VSP are
expressed in chicory root tissues (Supporting Information
Fig. S3). These seven sequences could all be detected in a
subset of the EST library obtained from a single root. Gene
polymorphism can therefore explain, in part, VSP protein
polymorphism.
Not surprisingly, intense spots were seen corresponding
to the cytoskeleton proteins actin (spot 567) and tubulin
(942) and to glycolytic enzymes: enolase (388), triosepho-
sphate isomerase (881), and glyceraldehyde 3-phosphate
dehydrogenase (G3PDH) (683 and 690). G3PDH is also
present in more than 20 spots of lower intensity (Fig. 2B).
Three major spots, superoxide dismutase (953), catalase
(425), and ascorbate peroxidase (874), are antioxidant
enzymes, particularly involved in the abiotic stress response.
The root sample used for proteomic analysis was subjected
to temperatures below 51C and it is known that cold accli-
mation induces the expression of anti-oxidative stress
proteins [9, 10]. Another abundant spot (983), nucleotide
diphosphate kinase B, can be added to this list of abiotic
stress response proteins. Although this enzyme is primarily
involved in the synthesis of nucleoside triphosphates other
than ATP, it also regulates the expression of genes involved
in the oxidative stress response [11].
An abundant cysteine proteinase inhibitor was also observed
(spot 989). The physiological role of this protein is not clearly
defined, but might involve the regulation of endogenous
proteases, programmed cell death, and the response to biotic as
well as abiotic stress. In particular, cold stress has been shown
to upregulate the expression of this protein [12, 13].
Other major spots include proteins known to be involved in
water stress, which is related to cold stress. A dehydrin protein
was identified as a major chicory root spot (829). Among water
stress proteins, dehydrins are believed to form complexes with
other macromolecules and protect them from freeze-induced
desiccation [14]. Another major spot (936) is similar to Medicagotruncatula cold acclimation-specific protein 15 (CAS15). This
protein contains the characteristic dehydrin K- and S-segments
and is highly cold-responsive [15]. Spot 938 is homologous to a
water stress-induced protein, which shows high identity with
CAS15. Although not major spots, other proteins related to
water stress were identified, such as the 25 kDa dehydrin-like
protein. Another dehydrin, ERD14, was identified in nine spots
(Supporting Information Table 1). In conclusion, of the 21
major spots analyzed, 7 were putatively involved in oxidative,
water or cold stress, all possibly related to cold acclimation. This
abundance is in agreement with the large number (67 spots) of
proteins classified as ‘‘Stress response’’ (Supporting Informa-
tion Table 1).
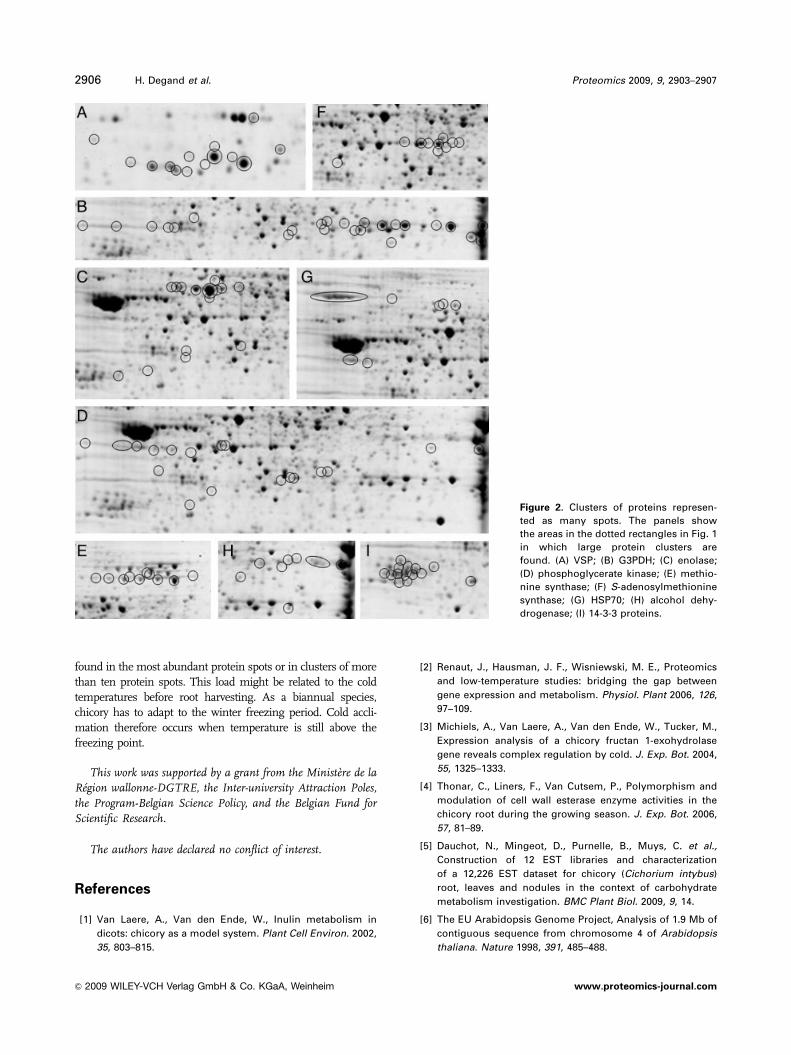
Focusing only on major spots can be misleading, as some
proteins might be scattered over a large number of spots,
either because they are encoded by a multigene family or
because they undergo various post-translational modifica-
tions. We have already mentioned the VSP and G3PDH
families. Other protein families represented by a large
number of spots are shown in Fig. 2. These include the
metabolic enzymes enolase (C), phosphoglycerate kinase
(D), methionine synthase (E), S-adenosylmethionine
synthase (F), and alcohol dehydrogenase (H). Alcohol
dehydrogenase together with pyruvate dehydrogenase and
lactate dehydrogenase both seen as several spots (Support-
ing Information Table 1), is probably involved in the
fermentation pathway, which occurs in root tissues subjec-
ted to low oxygen stress. Many HSPs were also identified,
with the HSP70 family being the most abundant (Fig. 2G),
and representing another example of proteins involved in
the stress response. HSP70 has been shown to be a mole-
cular chaperone during cold stress [16]. A final example of a
large protein family is the 14-3-3 protein cluster (Fig. 2I),
which are regulatory proteins involved in many different
signaling pathways, including the cold stress response [17].
In conclusion, investigation of the proteome complement of
chicory root led to the identification of major spots corre-
sponding to 1-FFT, the enzyme involved in inulin synthesis.
More surprising was the large number of proteins known to be
involved in the abiotic stress response, several of which were
Table 1. Identification of the most abundant spots
Spota) Protein identification Functional category
388 Enolase Glycolysis425 Catalase 3 Stress response452, 473, 485 Fructan:fructan 1-
fructosyltransferasePolysaccharide
metabolism519 Alcohol
dehydrogenaseshomologous
Fermentation
567 Actin Cytoskeleton683, 690 Glyceraldehyde 3-
phosphatedehydrogenase
Glycolysis
829 Dehydrin 2 Stress response874 Ascorbate peroxidase Stress response881 Triosephosphate
isomeraseGlycolysis
936 Cold-acclimationspecific protein 15
Stress response
938 Water stress-inducedprotein, putative
Stress response
942 Beta-7 tubulin Cytoskeleton953 Superoxide
dismutase [Cu–Zn]Stress response
962, 966, 968 Vegetative storageprotein
Storage
983 Nucleosidediphosphatekinase B
Nucleosidemetabolism andstress response
S989 Cysteine proteinaseinhibitor
Proteolysis
Proteomics 2009, 9, 2903–2907 2905
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com
found in the most abundant protein spots or in clusters of more
than ten protein spots. This load might be related to the cold
temperatures before root harvesting. As a biannual species,
chicory has to adapt to the winter freezing period. Cold accli-
mation therefore occurs when temperature is still above the
freezing point.
This work was supported by a grant from the Ministere de laRegion wallonne-DGTRE, the Inter-university Attraction Poles,the Program-Belgian Science Policy, and the Belgian Fund forScientific Research.
The authors have declared no conflict of interest.
References
[1] Van Laere, A., Van den Ende, W., Inulin metabolism in
dicots: chicory as a model system. Plant Cell Environ. 2002,
35, 803–815.
[2] Renaut, J., Hausman, J. F., Wisniewski, M. E., Proteomics
and low-temperature studies: bridging the gap between
gene expression and metabolism. Physiol. Plant 2006, 126,
97–109.
[3] Michiels, A., Van Laere, A., Van den Ende, W., Tucker, M.,
Expression analysis of a chicory fructan 1-exohydrolase
gene reveals complex regulation by cold. J. Exp. Bot. 2004,
55, 1325–1333.
[4] Thonar, C., Liners, F., Van Cutsem, P., Polymorphism and
modulation of cell wall esterase enzyme activities in the
chicory root during the growing season. J. Exp. Bot. 2006,
57, 81–89.
[5] Dauchot, N., Mingeot, D., Purnelle, B., Muys, C. et al.,
Construction of 12 EST libraries and characterization
of a 12,226 EST dataset for chicory (Cichorium intybus)
root, leaves and nodules in the context of carbohydrate
metabolism investigation. BMC Plant Biol. 2009, 9, 14.
[6] The EU Arabidopsis Genome Project, Analysis of 1.9 Mb of
contiguous sequence from chromosome 4 of Arabidopsis
thaliana. Nature 1998, 391, 485–488.
Figure 2. Clusters of proteins represen-
ted as many spots. The panels show
the areas in the dotted rectangles in Fig. 1
in which large protein clusters are
found. (A) VSP; (B) G3PDH; (C) enolase;
(D) phosphoglycerate kinase; (E) methio-
nine synthase; (F) S-adenosylmethionine
synthase; (G) HSP70; (H) alcohol dehy-
drogenase; (I) 14-3-3 proteins.
2906 H. Degand et al. Proteomics 2009, 9, 2903–2907
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com

[7] Ameziane, R., Richard-Molard, C., Deleens, E., Morot-
Gaudry, J. F., Limani, A. M., Nitrate (15NO3) limitation
affects nitrogen partitioning between metabolic and storage
sinks and nitrogen reserve accumulation in chicory
(Cichorium intybus L.). Planta 1997, 202, 303–312.
[8] Richard-Molard, C., Brugiere, N., Moille, M., Carrayoln, E.,
Limani, A. M., Molecular characterization of a gene encod-
ing a vegetative storage protein (CiVSP) from Cichorium
intybus and its expression in the root and shoot in relation
to nitrogen status and pathogen resistance. Physiol. Plant
2004, 121, 568–577.
[9] Oktem, H. A., Eyidooan, F., Demirba, D., Bayrac, A. T. et al.,
Antioxidant responses of lentil to cold and drought stress.
J. Plant Biochim. Biotechnol. 2008, 17, 15–21.
[10] Kuk, Y. I., Shin, J. S., Burgos, N. R., Hwang, T. E. et al.,
Antioxidative enzymes offer protection from chilling
damage in rice plants. Crop Sci. 2003, 43, 2109–2117.
[11] Yang, K. A., Moon, H., Kim, G., Lim, C. J. et al., NDP kinase 2
regulates expression of antioxidant genes in Arabidopsis.
Proc. Jpn. Acad. Ser. B 2003, 79, 86–91.
[12] Valdes-Rodriguez, S., Guerrero-Rangel, A., Melgoza-
Villagomez, C., Chagolla-Lopez, A. et al., Cloning of
a cDNA encoding a cystatin from grain amaranth
(Amaranthus hypochondriacus) showing a tissue-
specific expression that is modified by germination
and abiotic stress. Plant Physiol. Biochem. 2007, 45,
790–798.
[13] Pernas, M., Sanchez-Monge, R., Salcedo, G., Biotic and
abiotic stress can induce systatin expression in chestnut.
FEBS Lett. 2000, 467, 206–210.
[14] Kosova, K., Vitamvas, P., Prasil, I. T., The role of
dehydrins in plant response to cold. Biol. Plant. 2007, 51,
601–617.
[15] Pennycooke, J. C., Cheng, H., Stockinger, E., Comparative
genomic sequence and expression analyses of
Medicago truncatula and Alfalfa subspecies falcata COLD-
ACCLIMATION-SPECIFIC genes. Plant Physiol. 2008, 146,
1242–1254.
[16] Zhang, C., Guy, C. L., In vitro evidence of Hsc 70 functioning
as a molecular chaperone during cold stress. Plant Physiol.
Biochem. 2006, 44, 844–850.
[17] Jarillo, J. A., Capel, J., Leyva, A., Martinez Zapater, J. M.,
Salinas, J., Two related low-temperature-inducible genes
of Arabidopsis encode proteins showing high homology to
14-3-3 proteins, a family of putative kinase regulators. Plant
Mol. Biol. 1994, 25, 693–704.
Proteomics 2009, 9, 2903–2907 2907
& 2009 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.proteomics-journal.com





























