Embed Size (px)
Citation preview

journa l homepage : www.e l sev i er . com/ loca te /mycres
m y c o l o g i c a l r e s e a r c h 1 1 1 ( 2 0 0 7 ) 9 3 9 – 9 4 6
Proteome analysis of an ectomycorrhizal fungusBoletus edulis under salt shock
Yu LIANG*, Hui CHEN, Mingjuan TANG, Shihua SHEN*
Key laboratory of Photosynthesis and Environmental Molecular Physiology, Institute of Botany, Chinese Academy of Sciences,
Beijing 100093, P. R. China
a r t i c l e i n f o
Article history:
Received 19 January 2007
Received in revised form
25 May 2007
Accepted 10 June 2007
Published online 29 June 2007
Corresponding Editor:
Daniel C. Eastwood
Keywords:
Edible mushrooms
ESI Q-TOF MS/MS
LC MS/MS
Mycorrhizas
Proteins
Saline soils
Two-dimensional electrophoresis
a b s t r a c t
Soil salinization has become a severe global problem and salinity is one of the most severe
abiotic stresses inhibiting growth and survival of mycorrhizal fungi and their host plants.
Salinity tolerance of ectomycorrhizal fungi and survival of ectomycorrhizal inocula is es-
sential to reforestation and ecosystem restoration in saline areas. Proteomic changes of
an ectomycorrhizal fungus, Boletus edulis, when exposed to salt stress conditions (4 %
NaCl, w/v) were determined using two-dimensional electrophoresis (2DE) and mass spec-
trometry (MS) techniques. Twenty-two protein spots, 14 upregulated and 8 downregulated,
were found changed under salt stress conditions. Sixteen changed protein spots were iden-
tified by nanospray ESI Q-TOF MS/MS and liquid chromatography MS/MS. These proteins
were involved in biosynthesis of methionine and S-adenosylmethionine, glycolysis, DNA
repair, cell cycle control, and general stress tolerance, and their possible functions in
salinity adaptation of Boletus edulis were discussed.
ª 2007 The British Mycological Society. Published by Elsevier Ltd. All rights reserved.
Introduction
Soil salinization has become a severe global problem, with
950 M ha of the earth’s soil being saline soil (Li et al. 2005). In
China, for example, there are more than 30 M ha of saline
soil and 9 M ha of secondary salinized soil (Wang 1993; Li
et al. 2005). The methods to improve the stress resistance
and survival of plant seedlings have become more and more
important for reforestation and ecosystem reestablishment
in saline areas. There are ca 6 K ectomycorrhizal fungal
(EMF) species (Molina et al. 1992), and over 3 % of plant species
could form ectomycorrhizas with EMF. The important role of
EMF in forest establishment and recovery has been well docu-
mented, and the formation of ectomycorrhizae is essential for
the survival of EM trees, as well as successful reforestation in
saline areas (Lamhamedi et al. 1992; Martins et al. 1996; Amar-
anthus 1998; Dunabeitia et al. 2004; Turjaman et al. 2005). Two
intrinsic characteristics may influence the functioning of EMF
species in reforestation in saline areas: (1) host specificity of
the fungus in the soil; (2) the tolerance of inoculated or native
EMF species to salt stress.
King bolete (Boletus edulis) is an EMF widely distributed in
temperate and subtropical forests (Mao 1997; Dunstan et al.
1998; Hall et al. 1998; Vance et al. 2001). It belongs to EMF spe-
cies with a broad host range and could form mycorrhizal rela-
tionships with conifers, oak, birch, fir, etc (Molina et al. 1992;
Mao 1997). Abiotic stress tolerance varies among different
EMF species, and B. edulis was reported as a tolerant species
* Corresponding authors.E-mail addresses: [email protected], [email protected]
0953-7562/$ – see front matter ª 2007 The British Mycological Society. Published by Elsevier Ltd. All rights reserved.doi:10.1016/j.mycres.2007.06.005

940 Y. Liang et al.
and could grow even at a water potential of –3 MPa (Coleman
et al. 1989). The broad host range and high stress tolerance
have made B. edulis a suitable EMF species that could be intro-
duced with various host tree species in reforestation and eco-
system reestablishment in saline areas.
High salinity usually causes water deficit, ion toxicity, and
nutrient deficiency leading to molecular damage, growth ar-
rest, and even death of organisms. Some fungal species, e.g.
yeasts, can tolerate high saline environments and have devel-
oped defence systems, including osmotic adjustment by
osmolyte synthesis and cation transport systems for sodium
exclusion under salt stress (Garcia et al. 1997). Transcriptional
data from some recent studies on yeast have indicated that re-
sponses to salt stress require the activity of several pathways
and many genes are involved (e.g. Gasch & Werner-
Washburne 2000; Posas et al. 2000). However, the lack of knowl-
edge regarding genetic mutations and genomic information of
ECM fungi, make it difficult to understand the molecular
mechanisms of these fungi in response to salinity stress.
Proteomics is a useful tool to study organism responses
under abiotic stresses, offering a powerful tool for the identi-
fication of proteins associated with a particular environmen-
tal signal (e.g. Novotna et al. 2003; Kav et al. 2004; Qi et al.
2004). Unlike methods to study individual proteins, proteo-
mics is used to study a global expression profile of proteins,
which better enable us to understand the systematic and
integrative responses of organism cells to environmental
stresses, as well as interactions and inter-regulations among
proteins under these stresses. Proteomics has been widely
applied to study cellular responses of plants and microbes
under abiotic stress (e.g. Cui et al. 2005; Topanurak et al.
2005; Yan et al. 2005), and great achievements have been
made to understand the integrative molecular regulation
under these stresses.
In the present study, proteomic changes of the EMF, Boletus
edulis, under saline stress (4 % NaCl, w/v) were studied, and
our objective was to find which proteins were involved in re-
sponses to salinity and to better understand the adaptation
mechanisms of B. edulis to salt stress.
Materials and methods
Fungal materials and salt treatment
Boletus edulis was provided by professor Liang-dong Guo and
the fungal materials were kept in the Center for Microbial
Resources of the institute of Microbiology, Chinese Academy
of Sciences. Mycelia of B. edulis were first grown on potato-
dextrose agar (PDA) for a week (25 �C). Rounded pieces of my-
celia (5.5 mm diam) were taken from the colony margin and
grown at 25 �C on potato-dextrose agar media containing 0,
1, 2, 4, 8 % NaCl (w/v), respectively. Radial growth of B. edulis
colonies at different NaCl concentrations was measured.
Mycelia of B. edulis were grown in sterilized potato-
dextrose liquid culture (25 �C, 160 rev min�1). After one-week
of growth, solid NaCl was added to obtain a final concentra-
tion of 4 % (w/v). B. edulis treated with NaCl for 6 h and
untreated controls (three replications) were centrifuged at
6000 g for 5 min at 4 �C to retrieve the mycelia.
Protein extraction
Proteins were extracted using a modified protocol according
to Shen et al. (2003). Five hundred milligram samples were
ground into fine powder in liquid nitrogen with a pre-
cooled mortar and pestle, and homogenized in 2 ml homog-
enization buffer containing 20 mM Tris–HCl (pH 7.5), 250 mM
sucrose, 10 mM ethylene glycol-bis (b-aminoethylether)-
N,N,N0,N0-tetraacetic acid (EGTA), 1 mM phenylmethylsul-
phonyl fluoride (PMSF), 1 mM dithiothreitol (DTT) and 1 %
Triton X-100. The homogenate was transferred into an
eppendorf tube and centrifuged at 15,000 g for 15 min at
4 �C. The supernatant was transferred to a new tube and
the protein was precipitated using 1/4 volume 50 % cold tri-
chloroacetic acid (TCA) in an ice water-bath for 30 min. The
mixture was centrifuged at 15,000 g for 15 min at 4 �C, and
the supernatant was discarded. The pellet was washed with
acetone three times, centrifuged and vacuum-dried. The-
dried powder was dissolved in sample buffer containing
9 M urea, 4 % Nonidet P-40 (NP-40), 2 % ampholine pH
3.5-10 (GE Healthcare Bio-Science, Little Chalfont) and 5 %
2-mercaptoethanol. The protein concentrations in the buffer
were determined according to Bradford (1976), with bovine
serum albumin as the standard.
Two-dimensional gel electrophoresis (2DE)
2DE was carried out according to Shen et al.(2003). First-
dimensional isoelectric focusing (IEF) was performed in
a 13 cm-long glass tube, 3 mm diam. The gel mixture con-
sisted of 4 % acrylamide, 8 M urea, 5 % ampholine (1 part
pH 3.5–10, 1 part pH 5–8) and 2 % NP-40. IEF was performed
at 200, 400, and 800 V for 30 min, 15 h and 1 h, respectively.
About 500 mg protein was loaded. After the first-dimen-
sional run, gels were incubated in equilibration buffer
[0.05 M Tris–HCl pH 6.8, 2.5 % SDS, 10 % (v/v) glycerol and
5 % 2-mercaptoethanol] for 15 min twice. The second-
dimensional electrophoresis was performed on vertical
slab gels (135� 170� 1 mm), and a Laemmli buffer system
(1970) was used to cast 5 % stacking gel and 15 % resolving
gel. After electrophoresis, the gels were stained with 0.1 %
Coomassie brilliant blue (CBB) R-250.
Image analysis
The stained gels were scanned at 600 dots per inch (dpi) reso-
lution using a UMAX Power Look 2100XL scanner (Maxium
Tech, Taipei). The transparency mode was used to obtain
a greyscale image. The image analysis was performed with
ImageMaster� 2D Platinum software (GE Healthcare Bio-Sci-
ence). The optimized parameters were as follows: saliency 2,
partial threshold 4, and minimum area 50. The percentage of
spot volumes (%) between control and salt treatment were
compared using Student’s t-test, and only significantly
changed protein spots ( p< 0.05) were subject to protein
identification.

Proteomic responses of Boletus edulis to salt shock 941
Protein identification
Protein spots were excised from the gels manually and cut
into small pieces. Protein digestion was performed accord-
ing to Shen et al. (2003) with slight modification. Each small
gel piece with protein was washed with 25 % (v/v) ethanol
and 7 % acetic acid for about 12 h or overnight at room
temperature, destained with 50 mM NH4HCO3 in 50 % (v/v)
methanol for 1 h at 40 �C. The protein in the gel piece
was reduced with 10 mM DTT in 100 mM NH4HCO3 for 1 h
at 60 �C and incubated with 40 mM iodoacetamide in
100 mM NH4HCO3 for 30 min at room temperature. The gel
pieces were minced and lyophilized, then rehydrated in
25 mM NH4HCO3 with 10 ng sequencing grade modified tryp-
sin (Promega, Madison, WI) at 37 �C overnight. After diges-
tion, the protein peptides were collected, and the gels
were washed with 0.1 % trifluoroacetic acid (TFA) in 50 %
acetonitrile three times to collect the remaining peptides.
The peptides were desalted by ZipTipC 18� pipette tips
(Millipore, Bedford, MA) and co-crystallized with one vol-
ume of saturated a-cyano-4-hydroxycinnamic acid in 50 %
(v/v) acetonitrile containing 1 % TFA.
The desalted protein samples were subject to nanospray
electrospray ionization quadrupole time-of-flight mass spec-
trometry (ESI Q-TOF MS/MS) or liquid chromatography mass
spectrometry (LC-MS/MS).
Q-TOF MS/MS
The applied spray voltage was 800–1000 V, with a sample cone
working on 25–40 V. The MCP detector working voltage was
2250 V, and energy adjustable collision cell was filled with
pure argon gas. MS/MS data were processed using MassLynx
3.5 and searched against NCBInr protein sequence databases
with the MS/MS ion searching program MASCOT (http://
www.matrixscience.com).
LC MS/MS
Liquid chromatography (LC) was performed on a surveyor
LC system (Thermo Finnigan, San Jose, CA). The C18 col-
umn was obtained form Column Technology (Fremont,
CA). Mobile phase A was 0.1 % formic acid in water, and
mobile phase B was 0.1 % formic acid in acetonitrile. The
tryptic peptide mixtures were eluted using a gradient of
2–98 % B over 180 min. The MS/MS was performed on
a LTQ linear ion trap mass spectrometer (Thermo Finnigan,
San Jose, CA) equipped with an electrospray interface and
operated in positive ion mode. The capillary temperature
was set to 170 �C and the spray voltage was 3.4 kV. The ac-
quired MS/MS spectra were compared against the NCBI
Basidiomycota protein database using the TurboSEQUEST
program in the BioWorks 3.1 software suite (Thermo Finni-
gan). An accepted SEQUEST result had to have a ¢Cn score
of at least 0.1 (regardless of charge state). Peptides with
a þ1 charge state were accepted if they were fully trypti-
case digested and had a cross-correlation (Xcorr) of at least
1.9. Peptides with a þ2 charge state were accepted if they
had an Xcorr� 2.2. Peptides with a þ3 charge state were
accepted if they had an Xcorr� 3.75.
Results and discussions
Mycelial growth of Boletus edulis under differentNaCl concentrations
The radial growth of the Boletus edulis mycelia on PDA with dif-
ferent NaCl concentrations is shown in Fig 1. Mycelial growth
of B. edulis was inhibited by NaCl except on the PDA medium
with 1 % NaCl. After 2 d, the radial growth B. edulis on PDA
with NaCl concentration equal or lower than 4 % was found
to fit a linear growth model. The growth of B. edulis was greatly
inhibited on PDA medium with 8 % NaCl. After one-month of
growth, the diameter of mycelial colony reached to 5.8�0.4 cm (mean� S.E., data not shown).
Tresner & Hayes (1971) determined the NaCl tolerance of
975 terrestrial fungal species and found that Basidiomycetes
were the least tolerant with about 75 % of species being unable
to withstand 4 % NaCl. Our results indicated that B. edulis had
a relatively higher NaCl tolerance and could grow well on PDA
medium containing 4 % NaCl. The high salinity tolerance en-
ables B. edulis to survive and grow in almost all saline areas.
Considered the wide host range of B. edulis, it would be a suit-
able EMF species to apply to reforestation and ecosystem res-
toration of saline areas.
2-DE and image analysis
Over 700 reproducible protein spots were detected on the each
gel of control and salt stress treatment with the pH range of
3.5–10 and the mass range of 20–120 kDa (Fig 2). Eighteen pro-
tein spots, comprising ten upregulated (U1-U10) and eight
downregulated proteins (D1-D8), were found significantly
changed under salt stress. The folds, which are defined as
the ratio of mean percentages of spots volume (vol%) on the
treated gels to those on the control gels, ranged from 1.79 to
17.55 for the upregulated proteins, and 0.05 to 0.52 for the
downregulated proteins.
0
1
2
3
4
5
6
7
8
0 1 2 3 4 5 6 7
01248
Time after transferring to the media (day)
Dia
met
er o
f fu
ngal
col
ony
(cm
)
Fig 1 – Radial growth of Boletus edulis mycelia on PDA with
different NaCl concentrations (bars indicate standard
deviations).

942 Y. Liang et al.
Protein identification
Twelve changed protein spots, comprising six upregulated
and six downregulated proteins, were identified using ESI
Q-TOF MS/MS and LC MS/MS (Table 1). These identified pro-
teins were involved in cellular processes, such as biosynthesis
of methionine and S-adenosylmethionine (U1, D1, D2), glycol-
ysis (U2), DNA repair (D4), cell cycle control and development
(U3, U4, D3, D5), and general stress tolerance (U5, U6). One pro-
tein (D6) was identified as hypothetical protein UM02452.1
with unknown function.
Function of changed proteins under salt stress
Proteins related to biosynthesis of L-methionineand S-adenosylmethionine
Cystathionine gamma-synthase (CGS)(D1), arginine N-methyltrans-
ferase 3(RMT3) (D2) and 5-methyltetrahydropteroyltriglutamate-
U1D5
D6
D2
D3
D1
D4
D7
D8
U4
U5U6
U3
U2
U7
U8U9
U10
U1D5
U4
D6
U5U6
D2
D3
D1
D4U3
U2
U7
U8U9
D7
D8U10
A
B
Fig 2 – Representative 2DE gel patterns of Boletus edulis.
(A) Control, (B) salt shock (4 % NaCl, 6 h).
homocysteine S-methyltransferase (MHMT) (U1) are involved
in biosynthesis of methionine (Met) and S-adenosylmethio-
nine (AdoMet; Fig 3). CGS catalyses the synthesis of cystathio-
nine from cysteine and O-phospho-L-homoserine (OPH) in a
g-replacement reaction (Ravanel et al. 1998). MHMT is involved
in L-methionine biosynthesis from 5-methyltetrahydropter-
oyltri-L-glutamate (5-CH3-H4-pteroyl-(glu)3) and homocysteine
(Whitfield et al. 1970; Urbanowski et al. 1987). RMT3 catalyses
the transfer of a methyl group from S-adenosylmethionine
(AdoMet) to a guanidino nitrogen of arginine, resulting in
S-adenosyl-homocysteine (AdoHcy) and methylarginine
(Bedford & Richard 2005).
Biosynthesis of Met and AdoMet might play an important
role in fungal responses to salt stress. It has been reported
that a yeast methionine biosynthetic gene HAL2(MET22), is ca-
pable of improving growth under salt stress, and methionine
supplementation may also improve the tolerance of yeast to
NaCl (Glaser et al. 1993). AdoMet is an important metabolite
participating in many cellular processes, e.g. methylation of
proteins, nucleic acids, polysaccharides and fatty acids, bio-
synthesis of ethylene and polyamines, cell wall synthesis
and modifications (Espartero et al. 1994).
The upregulation of MHMT is expected to enhance the bio-
synthesis of methionine and the downregulation of RMT3
may reduce the AdoMet consumption in arginine methylation
of proteins. The changes of these two enzymes may cause the
accumulation of methionine and AdoMet in the fungus, which
could meet the increased demand in stress-tolerance-related
processes such as hormone synthesis and cell wall modifica-
tions. It is interesting to find that CGS was downregulated in
our study. One possible reason, as suggested by Kim et al.
(2002), may be the negative regulation of methionine accumu-
lation. The role of CGS in the biosynthesis of methionine and
S-adenosylmethionine is still unclear, and results of trans-
genic studies showed that CGS repression did not caused a de-
crease of cystathionine concentrations in plants (Kreft et al.
2003).
Proteins involved in glycolysis
Enhanced glycolysis and increased expression of related en-
zymes under abiotic stresses have been well documented
(e.g. Umeda & Uchimiya 1994; Suzuki et al. 2005). In the present
study, glyceraldehyde-3-phosphate dehydrogenase (U2) in-
volved in glycolysis was upregulated, which may be related
to glycolysis enhancement and be responsible for the energy
supply under salt stress. Another possible function of in-
creased glycolysis under salt stress is osmoregulation, for
the biosynthesis of glycerol, a common osmolyte of fungi, is
connected with glycolysis (Blomberg 2000). In a recent study
on proteomic changes of yeast to salt stress, enzymes in gly-
colysis also changed significantly (Gori et al. 2007), which indi-
cates the important role of glycolysis regulation during
salinity adaptation of fungi.
Proteins related to DNA repair
Two parallel pathways, one error free and another error prone
(mutagenic), were found in the ubiquitin-related DNA repair
system. In Saccharomyces cerevisiae the homologue of
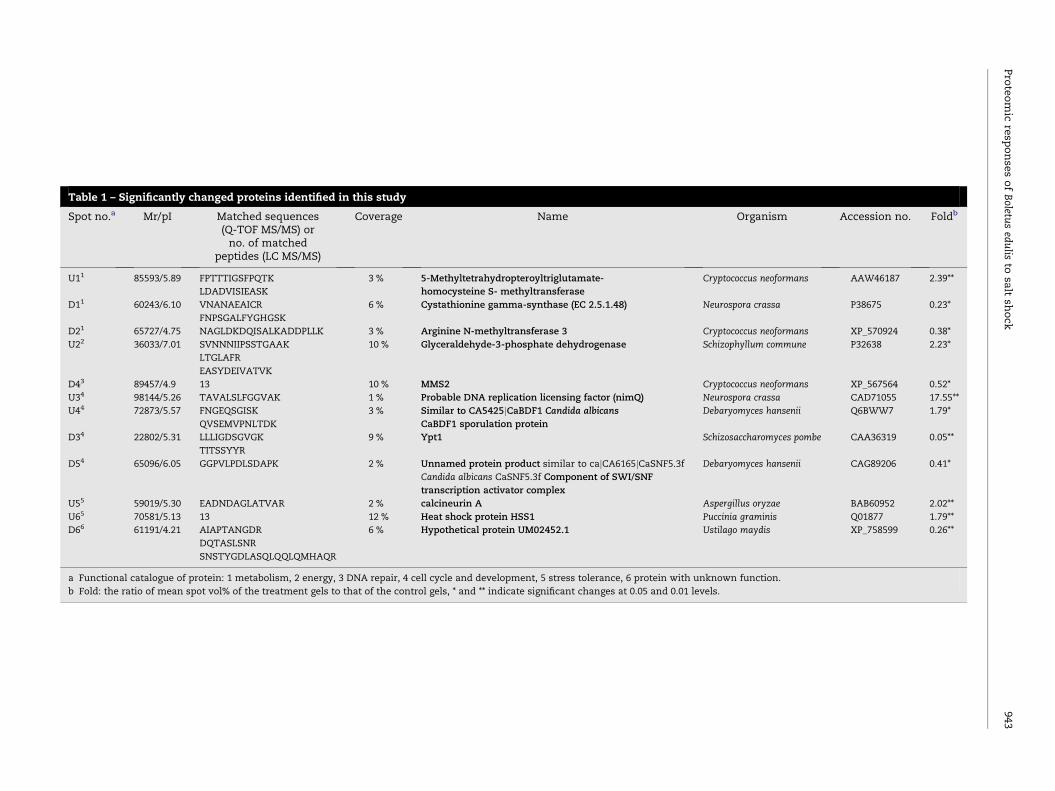
Table 1 – Signifi
Spot no.a M Organism Accession no. Foldb
U11 8559 Cryptococcus neoformans AAW46187 2.39**
D11 6024 Neurospora crassa P38675 0.23*
D21 6572 Cryptococcus neoformans XP_570924 0.38*
U22 3603 Schizophyllum commune P32638 2.23*
D43 8945 Cryptococcus neoformans XP_567564 0.52*
U34 9814 Neurospora crassa CAD71055 17.55**
U44 7287 Debaryomyces hansenii Q6BWW7 1.79*
D34 2280 Schizosaccharomyces pombe CAA36319 0.05**
D54 6509 .3f Debaryomyces hansenii CAG89206 0.41*
U55 5901 Aspergillus oryzae BAB60952 2.02**
U65 7058 Puccinia graminis Q01877 1.79**
D66 6119 Ustilago maydis XP_758599 0.26**
a Functional catal ith unknown function.
b Fold: the ratio o .01 levels.
Pro
teo
mic
resp
on
ses
of
Boletu
sed
ulis
tosa
ltsh
ock
943
cantly changed proteins identified in this study
r/pI Matched sequences(Q-TOF MS/MS) or
no. of matchedpeptides (LC MS/MS)
Coverage Name
3/5.89 FPTTTIGSFPQTK 3 % 5-Methyltetrahydropteroyltriglutamate-
homocysteine S- methyltransferaseLDADVISIEASK
3/6.10 VNANAEAICR 6 % Cystathionine gamma-synthase (EC 2.5.1.48)
FNPSGALFYGHGSK
7/4.75 NAGLDKDQISALKADDPLLK 3 % Arginine N-methyltransferase 3
3/7.01 SVNNNIIPSSTGAAK 10 % Glyceraldehyde-3-phosphate dehydrogenase
LTGLAFR
EASYDEIVATVK
7/4.9 13 10 % MMS2
4/5.26 TAVALSLFGGVAK 1 % Probable DNA replication licensing factor (nimQ)
3/5.57 FNGEQSGISK 3 % Similar to CA5425jCaBDF1 Candida albicans
CaBDF1 sporulation proteinQVSEMVPNLTDK
2/5.31 LLLIGDSGVGK 9 % Ypt1
TITSSYYR
6/6.05 GGPVLPDLSDAPK 2 % Unnamed protein product similar to cajCA6165jCaSNF5
Candida albicans CaSNF5.3f Component of SWI/SNF
transcription activator complex
9/5.30 EADNDAGLATVAR 2 % calcineurin A
1/5.13 13 12 % Heat shock protein HSS1
1/4.21 AIAPTANGDR 6 % Hypothetical protein UM02452.1
DQTASLSNR
SNSTYGDLASQLQQLQMHAQR
ogue of protein: 1 metabolism, 2 energy, 3 DNA repair, 4 cell cycle and development, 5 stress tolerance, 6 protein w
f mean spot vol% of the treatment gels to that of the control gels, * and ** indicate significant changes at 0.05 and 0

944 Y. Liang et al.
O-phospho-L-homoserine(OPH)
cystathionine
homocysteine
S-adenosyl-L-Met
S-adenosyl-L-Hcy 5-CH3-H4-pteroyl-(glu)3
H4-pteroyl-(glu)3L-methionine
CGS
MHMT
Protein-L-arginine
Protein-N-methyl-L-arginine
RMT
Fig 3 – Biosynthesis of L-methionine and S-adenosylmethionine. Abbreviations: CGS, cystathionine gamma-synthase; RMT,
arginine N-methyltransferase; MHMT, 5-methyltetrahydropteroyltriglutamate-homocysteine S-methyltransferase.
MMS2(D4) encodes a ubiquitin-conjugating enzyme-like pro-
tein, which was involved in the error-free post-replication
reparations (Broomfield et al. 1998). The downregulation of
MMS2 (D4) in our study might inhibit the error-free reparation,
and might, in turn, result in increased mutagenesis. These
results are in agreement with the fact that mutagenesis is
often increased as a consequence of stress-induced genetic
pathways, which is considered an adaptive strategy that could
potentially enhance the chances of surviving the stress
(Tenaillon et al. 2004).
Proteins related to cell cycle control and development
Ypt1 (D3), a Ras-like small GTPase involved in the endoplasmic
reticulum (ER)-to-Golgi step of the secretory pathway, is es-
sential for cell growth (Miyake & Yamamoto 1990). The down-
regulation of Ypt1 indicated the repression of cell growth and
division under salt stress.
Swi/Snf complexes (D5) have been suggested as transcrip-
tion repressors (Martens & Winston 2003) and most Swi/Snf
target genes are able to respond to growth conditions or to
the developmental state of the cells (Muchardt & Yaniv
1999). DNA replication licensing factor (nimQ) (U3) is responsi-
ble for the initiation of DNA replication and involved in control
of cell cycle (Ye et al. 1997). Protein spot (U4) is similar to
Candida albicans CA5425jCaBDF1 sporulation protein. Under
extreme stress conditions many bacteria and fungi form
stress-resistant spores. Sporulation can be regarded as the
ultimate form of growth control and cell cycle regulation dur-
ing stress (Kultz 2005). No spore production was found in the
present study, which might be due to the short duration of salt
stress. The upregulation of DNA replication licensing factor,
and sporulation protein, and downregulation of the compo-
nent of the SWI/SNF complex indicated the activation of
a cell cycle control system. Considering the changes of the
protein related to cell growth (Ypt1), these proteins may be
part of the B. edulis switch from normal growth to salt stress.
Salt stress tolerance proteins
Heat shock protein HSS1 (U6) belongs to the heat shock pro-
tein family. Proteins in this family are induced by a variety
of environmental stresses and have been mentioned as con-
tributing to salt tolerance (Lewis et al. 1995; Lahav et al.
2004). Calcineurin A (U5) is the catalytic subunit of calcineurin,
a Ca2þ and calmodulin-dependent serine-threonine protein
phosphatase (Rusnak & Mertz 2000). Calcineurin is essential
for NaCl tolerance of yeast and could promote yeast survival
under environmental stress (Mendoza et al. 1994; Bultynck
et al. 2006). The role of calcineurin in salt tolerance is thought
to be mediated by transcriptional and post-translational
mechanisms (Rusnak & Mertz 2000), and it was reported that
calcineurin-deficient yeast exhibited decreased tolerance to
Naþ and Liþ (Nakamura et al. 1993; Mendoza et al. 1994). The
upregulation of two stress tolerance proteins, calcineurin A
and heat shock protein HSS1, may enhanced salt tolerance
of Boletus edulis and promotes survival of the fungus under
salt stress.
In conclusion, proteins related to multiple cellular pro-
cesses, e.g. metabolisms, energy related processes, DNA re-
pair, cell cycle control, and stress tolerance, were found
involved in the stress responses of B. edulis to salt stress in
the present study. Paying more attention to these cellular pro-
cesses rather than individual stress-tolerance related proteins
will be helpful for better understanding of the global response
mechanisms of organisms to abiotic stresses.
Acknowledgements
This work was supported by the Knowledge Innovation Pro-
ject of the Institute of Botany, the Chinese Academy of
Sciences (C-810). We are grateful to Liang-dong Guo for provid-
ing fungal isolates. We also thank Kate Partner and two anon-
ymous reviewers for their comments and suggestion on an
early version of this paper.
r e f e r e n c e s
Amaranthus MP, 1998. The Importance and Conservation of Ectomy-corrhizal Fungal Diversity in Forest Ecosystems: lessons fromEurope and the Pacific Northwest [General technical reportPNW 0(431)]. USDA Forest Service, Seattle.
Bedford MT, Richard S, 2005. Arginine methylation: an emergingregulator of protein function. Molecular Cell 18: 263–272.
Blomberg A, 2000. Metabolic surprises in Saccharomyces cerevisiaeduring adaptation to saline conditions: questions, someanswers and a model. FEMS Microbiology Letters 182: 1–8.

Proteomic responses of Boletus edulis to salt shock 945
Bradford MM, 1976. A rapid and sensitive method for thequantitation of microgram quantities of protein utilizing theprinciple of protein-dye binding. Analytical Biochemistry 72:248–254.
Broomfield S, Chow BL, Xiao W, 1998. MMS2, encoding a ubiqui-tin-conjugating-enzyme-like protein, is a member of the yeasterror-free postreplication repair pathway. Proceedings of theNational Academy of Sciences USA 95: 5678–5683.
Bultynck G, Heath VL, Majeed AP, Galan J-M, Haguenauer-Tsapis R, Cyert MS, 2006. Slm1 and Slm2 are novel substratesof the calcineurin phosphatase required for heat stress-induced endocytosis of the yeast uracil permease. Molecularand Cellular Biology 26: 4729–4745.
Coleman MD, Bledsoe CS, Lopushinsky W, 1989. Pure cultureresponse of ectomycorrhizal fungi to imposed water stress.Canadian Journal of Botany 67: 29–39.
Cui S, Huang F, Wang J, Ma X, Cheng Y, Liu J, 2005. A proteomicanalysis of cold stress responses in rice seedlings. Proteomics 5:3162–3172.
Dunabeitia MK, Hormilla S, Garcia-Plazaola JI, Txarterina K,Arteche U, Becerril JM, 2004. Differential responses ofthree fungal species to environmental factors and their rolein the mycorrhization of Pinus radiata D. Don. Mycorrhiza 14:11–18.
Dunstan WA, Dell B, Malajczuk N, 1998. The diversity of ecto-mycorrhizal fungi associated with introduced Pinus spp. in theSouthern Hemisphere, with particular reference to WesternAustralia. Mycorrhiza 8: 71–79.
Espartero J, Pintor-Toro JA, Pardo JM, 1994. Differential accumu-lation of S-adenosylmethionine synthetase transcripts in re-sponses to salt stress. Plant Molecular Biology 25: 217–227.
Garcia MJ, Rios G, Ali R, Belles JM, Serrano R, 1997. Comparativephysiology of salt tolerance in Candida tropicalis and Saccharo-myces cerevisiae. Microbiology 143: 1125–1131.
Gasch AP, Werner-Washburne M, 2002. The genomics of yeastresponses to environmental stress and starvation. Functionaland Integrative Genomics 2: 181–192.
Glaser HU, Thomas D, Gaxiola R, Montrichard F, Surdin-Kerjan Y,Serrano R, 1993. Salt tolerance and methionine biosynthesis inSaccharomyces cerevisiae involve a putative phosphatase gene.EMBO Journal 12: 3105–3110.
Gori K, Hebraud M, Chambon C, Mortensen HD, Arneborg N,Jespersen L, 2007. Proteomic changes in Debaryomyceshansenii upon exposure to NaCl stress. FEMS Yeast Research 7:293–303.
Hall IR, Lyon AJE, Wang Y, Sinclair L, 1998. Ectomycorrhizal fungiwith edible fruiting bodies 2. Boletus edulis. Economic Botany 52:44–56.
Kav NNV, Srivastava S, Goonewardene L, Blade SF, 2004.Proteome-level changes in the roots of Pisum sativum inresponse to salinity. Annals of Applied Biology 145: 217–230.
Kim J, Lee M, Chalam R, Martin MN, Leustek T, Boerjan W, 2002.Constitutive overexpression of cystathionine g-synthase inArabidopsis leads to accumulation of soluble methionine andS-methylmethionine. Plant Physiology 128: 95–107.
Kreft O, Hoefgen R, Hesse H, 2003. Functional analysis of cysta-thionine g-synthase in genetically engineered potato plants.Plant Physiology 131: 1843–1854.
Kultz D, 2005. Molecular and evolutionary basis of the cellularstress response. Annual Review of Physiology 67: 225–257.
Laemmli UK, 1970. Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature 227: 680–685.
Lahav R, Nejidat A, Abeliovich A, 2004. Alterations in proteinsynthesis and levels of heat shock 70 proteins in response tosalt stress of the halotolerant yeast Rhodotorula mucilaginosa.Antonie van Leeuwenhoek 85: 259–269.
Lamhamedi MS, Bernier PY, Fortin JA, 1992. Growth, nutritionand response to water stress of Pinus pinaster inoculated
with ten dikaryotic strains of Pisolithus sp. Tree Physiology 10:153–167.
Lewis JG, Learmonth RP, Watson K, 1995. Induction of heat,freezing and salt tolerance by heat and salt shock in Saccha-romyces cerevisiae. Microbiology 141: 687–694.
Li W, Liu X, Qiao H, Sun J, Duan D, 2005. Two phase tillage: quickmethod for crop production in saline soils. Agrifood ResearchReports 68: 66–72.
Mao X-L, 1997. The Macrofungi in China. Henan Technology Press,Zhengzhou.
Martens JA, Winston F, 2003. Recent advances in understandingchromatin remodeling by Swi/Snf complexes. Current OpinionIn Genetics & Development 13: 136–142.
Martins A, Barroso J, Pais MS, 1996. Effect of ectomycorrhizalfungi on survival and growth of micropropagated plants andseedlings of Castanea sativa mill. Mycorrhiza 6: 265–270.
Mendoza I, Rubio F, Rodriguez-Navarro A, Pardo JM, 1994. Theprotein phosphatase calcineurin is essential for NaCltolerance of Saccharomyces cerevisiae. Journal of BiologicalChemistry 269: 8792–8796.
Miyake S, Yamamoto M, 1990. Identification of ras-related, YPTfamily genes in Schizosaccharomyces pombe. EMBO Journal 9:1417–1422.
Molina R, Massicotte H, Trappe JM, 1992. Specificity phenomenain mycorrhizal symbioses: community-ecological conse-quences and practical implications. In: Allen MF (ed.),Mycorrhzal Functioning: An Integrative Plant–Fungal Process.Chapman & Hall, New York, pp. 357–423.
Muchardt C, Yaniv M, 1999. The mammalian SWI/SNF complexand the control of cell growth. Cell and Developmental Biology10: 189–195.
Nakamura T, Liu Y, Hirata D, Namba H, Harada S-I, Hirokawa T,Miyakawa T, 1993. Protein phosphatase type 2B (calcineurin)-mediated, FK506-sensitive regulation of intracellular ions inyeast is an important determinant for adaptation to high saltstress conditions. EMBO Journal 12: 4063–4071.
Novotna J, Vohradsky J, Berndt P, Gramajo H, Langen H, Li X-M,Minas W, Orsaria L, Roeder D, Thompson CJ, 2003. Proteomicstudies of diauxic lag in the differentiating prokaryote Strep-tomyces coelicolor reveal a regulatory network of stress-inducedproteins and central metabolic enzymes. MolecularMicrobiology 48: 1289–1303.
Posas F, Chambersi JR, Heymani JA, Hoeffleri JP, de Nadal E,Arino J, 2000. The transcriptional response of yeast to salinestress. Journal of Biological Chemistry 275: 17249–17255.
Qi S, Yang P, Jing Y, Shen S, Yang S, 2004. Analysis of salt shock-induced proteome of Sinorhizobium fredii. Chinese ScienceBullettin 49: 1828–1833.
Ravanel S, Gakiere B, Job D, Douce R, 1998. The specific features ofmethionine biosynthesis and metabolism in plants. Proceed-ings of the National Academy of Sciences of the United States ofAmerica 95: 7805–7812.
Rusnak F, Mertz P, 2000. Calcineurin: form and function. PhysiologicalReviews 80: 1483–1521.
Shen S, Jing Y, Kuang T, 2003. Proteomics approach to identifywound-response related proteins from rice leaf sheath.Proteomics 3: 527–535.
Suzuki M, Hashioka A, Mimura T, Ashihara H, 2005. Salt stressand glycolytic regulation in suspension-cultured cells of themangrove tree, Bruguiera sexangula. Physiologia Plantarum 123:246–253.
Tenaillon O, Denamur E, Matic I, 2004. Evolutionary significanceof stress induced mutagenesis in bacteria. Trends in Microbiol-ogy 12: 264–270.
Topanurak S, Sinchaikul S, Sookkheo B, Phutrakul S, Chen S-T,2005. Functional proteomics and correlated signalingpathway of the thermophilic bacterium Bacillus stearothermo-philus TLS33 under cold-shock stress. Proteomics 5: 4456–4471.

946 Y. Liang et al.
Tresner HD, Hayes JA, 1971. Sodium chloride tolerance of terres-trial fungi. Applied Microbiology 22: 210–213.
Turjaman M, Tamai Y, Segah H, Limin SH, Cha JY, Osaki M,Tawaraya K, 2005. Inoculation with the ectomycorrhizal fungiPisolithus arhizus and Scleroderma sp. improves early growth ofShorea pinanga nursery seedlings. New Forests 30: 67–73.
Umeda M, Uchimiya H, 1994. Differential transcript levels ofgenes associated with glycolysis and alcohol fermentation inrice plants (Oryza sativa) under submergence stress. PlantPhysiology 106: 1015–1022.
Urbanowski ML, Stauffer LT, Plamann LS, Stauffer GV, 1987.A new methionine locus, metR, that encodes a transactingprotein required for activation of metE and metH in Escherichiacoli and Salmonella typhimurium. Journal of Bacteriology 169:1391–1397.
Vance NC, Borsting M, Pilz D, Freed J, 2001. Special Forests Prod-ucts: Species Information Guide for the Pacific NorthwestGeneral technical report PNW(513). USDA Forest Service, Seattle.
Wang Z, 1993. China Saline Soil. Science press, Beijing.Whitfield CD, Steers EJ, Weissbach H, 1970. Purification and prop-
erties of 5-methyltetrahydropteroyltri-glutamate-homocys-teine transmethylase. Journal of Biological Chemistry 245: 390–401.
Yan S, Tang Z, Su W, Sun W, 2005. Proteomic analysis of saltstress-responsive proteins in rice root. Proteomics 5: 235–244.
Ye XS, Fincher RR, Tang A, McNeal KK, Gygax SE, Wexler AN,Ryan KB, James SW, Osmani SA, 1997. Proteolysis andtyrosine phosphorylation of p34cdc2/cyclin B, the role ofMCM2 and initiation of DNA replication to allow tyrosinephosphorylation of p34cdc2. Journal of Biological Chemistry 272:33384–33393.