Embed Size (px)
Citation preview

ANALYTICAL BIOCHEMISTRY 147, 525-528 (1985)
Proteolytic Release of Glycopeptides from Glycoproteins Transferred to Nitrocellulose following Gel Electrophoresis’
ARNOLDS.DION~ANDANTHONYA.POMENTI
Department of Molecular Biology, Institute for Medical Research, Copewood Street. Camden. New Jersey 08103
Received January 7. 1985
To determine whether glycopeptides could be released from glycoproteins bound to nitrocel- lulose, the glycoproteins of murine mammary tumor virus (MuMTV) were radiolabeled by the periodate oxidation/tritiated sodium borohydride reduction technique and separated by gel electrophoresis followed by diffusion transfer. Pronase digestion of nitrocellulose filter strips containing labeled glycoproteins (gp55 or gp34) revealed a rapid release of glycopeptides. i.e.. approximately total release within 4 h. The released glycopeptides were similar in size. as determined by molecular sieving chromatography, to glycopeptides obtained by proteolytic digestion of MuMTV glycoproteins from dried gel strips (A. Zilberstein et al.. 1980. Cell 21, 417-427) or in solution (M. J. Yagi et al., 1978. Virology 91, 291-304). o 1985 Academic
Press, Inc.
KEY WORDS: glycopeptides; filter transfers: murine mammary tumor virus (MuMTV).
Polyacrylamide gel electrophoresis, under sodium dodecyl sulfate denaturation condi- tions (SDS-PAGE),3 offers a relatively simple technique for high-resolution separations of protein mixtures (1,2) and is especially ap- plicable for studies of individual glycoproteins which are available in limited amounts. In this regard, Zilberstein et al. (3) demonstrated the efficacy of releasing glycopeptides by pro- nase digestion of gel segments after radioau- tographic localization of radiolabeled glyco- proteins of vesicular stomatitis virus following SDS-PAGE.
In the studies presented here, we found that glycoproteins transferred to nitrocellulose could be proteolytically digested to rapidly effect the release of glycopeptides. In addition. the molecular weights of glycopeptides re-
’ This investigation was supported in part by PHS Grant CA30636 awarded by the National Cancer Institute, DHHS.
* To whom correspondence should be addressed. 3 Abbreviations used: SDS-PAGE, sodium dodecyl
sulfate-polyacrylamide gel electrophoresis; MuMTV, murine mammary tumor virus: PBS. phosphate-buffered saline.
leased from extrinsically labeled, MuMTV- coded glycoproteins (gp55 and gp34) by the present technique were similar to those pre- pared by the technique of Zilberstein et al. (3). The advantages of glycopeptide release from nitrocellulose membrane filters include rapid radiolabeled glycoprotein localization by radioautography and glycopeptide release because glycoproteins are immobilized on a thin filter as compared to thicker gel seg- ments.
MATERIALS AND METHODS
Biological and chemical reagents. Milk- born murine mammary tumor virus (MuMTV) of the isogeneic RI11 mouse strain was purified by velocity sedimentation and isopycnic-gra- dient centrifugation (4). Fetuin (Type III), stachyose, dithiothreitol, pronase, sodium periodate, and sodium borohydride were ob- tained from Sigma Chemical Corporation (St. Louis, MO.). [3H]NaBH4 (sp act 282 mCi/mmol) was purchased from NEN (Boston, Mass.). Nitrocellulose membrane fil- ters (0.45 ym, Type HA) were obtained
525 0003-2697185 $3.00 Copyright Q 1985 by Academic Press. Inc. All rights of reproduction in any form reserved.

526 DION AND POMENTI
from Millipore Corporation (Bedford, Mass.). Ovalbumin (2X crystallized) and ultrapure urea were provided by Schwartz/Mann Inc. (Spring Valley, N. Y.).
Radiolabeling of MuMTV glycoproteins. MuMTV [ 1.5 mg in 0.5 ml phosphate-buff- ered saline (PBS)] was reacted with freshly prepared sodium periodate (2 mM final con- centration) for 30 min at 0°C and the virus was recovered by centrifugation (40,000 rpm, Beckman 50.1 rotor for 45 min). Tritium labeling of oxidized carbohydrate groups was then performed by resuspending the pellet in 0.5 ml PBS containing [3H]NaBH4 (10 mCi/ ml) and incubation at room temperature for 30 min, followed by the addition of unlabeled NaBH4 (1 mg/20 ~1) and further incubation for 15 min at ambient temperature. The virus was again pelleted by centrifugation, as described above, and resuspended in 0.1 ml PBS.
Slab gel electrophoresis. MuMTV proteins were separated by slab gel electrophoresis using the discontinuous buffer conditions of Laemmli (2) in the presence of 4 M urea (5). The stacking and separating gels consisted of 3 and 9% acrylamide, respectively. For ra- dioautographic localization of radiolabeled glycoproteins, the gel was dried on a Bio- Rad slab gel dryer and exposed to X-ray film.
D#iision transfers to nitrocellulose. Fol- lowing SDS-PAGE, proteins were transferred to nitrocellulose by the technique of Bowen et al. (5). Briefly, SDS was removed by immersing the slab gel for 3 h at room tem- perature in 10 mM Tris . HCl buffer contain- ing 50 mM NaCI, 2 mM EDTA, 0.1 mM
dithiothreitol, and 4 M urea. Two-dimen- sional transfers were then obtained by placing the slab gel between pairs of nitrocellulose filters, sponges, and supports and immersing the “sandwiched” gel in the same buffer without urea (5). Diffusion transfers were allowed to proceed for 40 to 48 h with at least two buffer changes. Radiolabeled pro- teins transferred to filters were located by radioautography or by liquid scintillation counting of 5-mm filter segments.
Glycopeptide release. Proteolytic release of glycopeptides from glycoproteins was per- formed under the conditions previously de- scribed by Zilberstein et al. (3). Dried gel or filter strips were cut into segments (1 to 2 mm by 5 mm), placed into a conical test tube, and covered with 0.5 ml predigested promise [0.4% (w/v) in 0.1 M Tris - HCl buffer (pH 8.0) containing 10 mM CaC& and 0.5 mM sodium azide]. Digestions were per- formed at 50°C. Additional details for the digestion of filter segments are presented in the text and figure legends. For gels, addi- tional Pronase [50 ~1 of a 2% (w/v) solution] was added at 12 and 24 h, the liquid was removed at 36 h, the rehydrated gel segments were soaked overnight in 2 ml of the same buffer without Pronase, and the liquid was pooled. For filter strips additional Pronase, as described for the gel segments, was added at 24 h only.
Analytical techniques. Protein was deter- mined by the procedure of Lowry et al. (6) with bovine serum albumin as the standard protein. Molecular weight estimations of pro- nase-released glycopeptides were performed on a Bio-Gel PlO/P6 column (7). The void volume (V,) was determined from the elution volume of dextran blue, and molecular weight markers included fetuin and ovalbumin gly- copeptides, obtained by exhaustive Pronase digestion (8) and stachyose. Elution volumes for the molecular weight standards were monitored by the phenol/sulfuric acid assay for neutral sugars (9) and radiolabeled gly- copeptides were located by liquid scintillation counting using Hydromix fluor (Yorktown Research, New Hyde Park, N. Y.).
RESULTS AND DISCUSSION
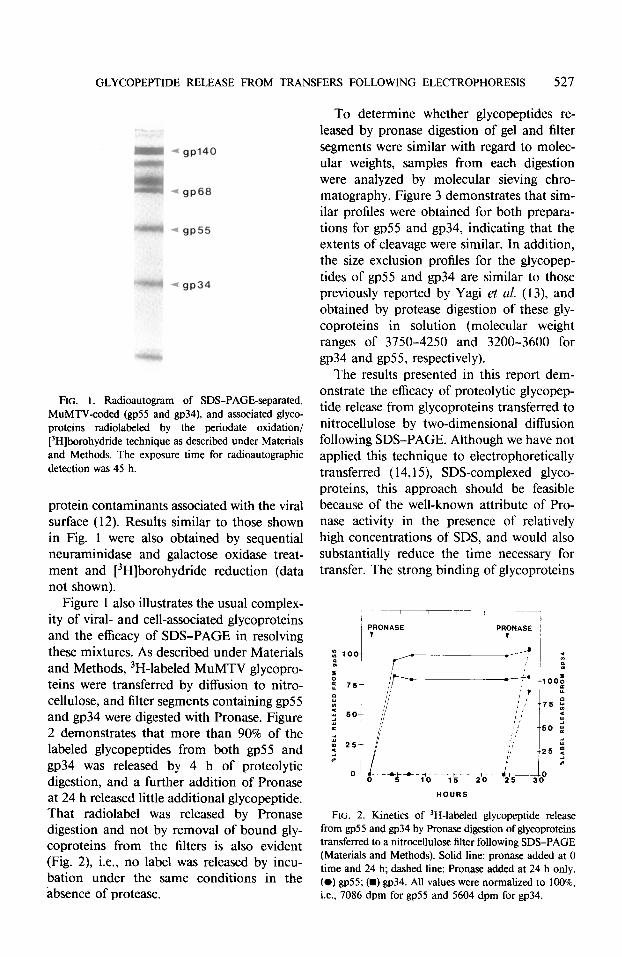
The radioautographic localization of peri- odate-oxidized/tritiated borohydride-reduced glycoproteins of MuMTV is illustrated in Fig. 1. Of these labeled glycoproteins, only two are MuMTV-coded, i.e., gp55 and gp34, derived from the env polyprotein precursor ( IO,1 1). Higher molecular weight glycopro- teins have been routinely observed to be host
GLYCOPEPTIDE RELEASE FROM TRANSFERS FOLLOWING ELECTROPHORESIS 527
- gp68
* 9P55
- clP34
FIG. 1. Radioautogram of SDS-PAGE-separated, MuMTV-coded (gp55 and gp34), and associated glyco- proteins radiolabeled by the periodate oxidation/ [‘Hlborohydtide technique as described under Materials and Methods. The exposure time for radioautographic detection was 45 h.
protein contaminants associated with the viral surface (12). Results similar to those shown in Fig. 1 were also obtained by sequential neuraminidase and galactose oxidase treat- ment and [3H]borohydride reduction (data not shown).
Figure 1 also illustrates the usual complex- ity of viral- and cell-associated glycoproteins and the efficacy of SDS-PAGE in resolving these mixtures. As described under Materials and Methods, 3H-labeled MuMTV glycopro- teins were transferred by diffusion to nitro- cellulose, and filter segments containing gp55 and gp34 were digested with Pronase. Figure 2 demonstrates that more than 90% of the labeled glycopeptides from both gp55 and gp34 was released by 4 h of proteolytic digestion, and a further addition of Pronase at 24 h released little additional glycopeptide. That radiolabel was released by Pronase digestion and not by removal of bound gly- coproteins from the filters is also evident (Fig. 2) i.e., no label was released by incu- bation under the same conditions in the absence of protease.
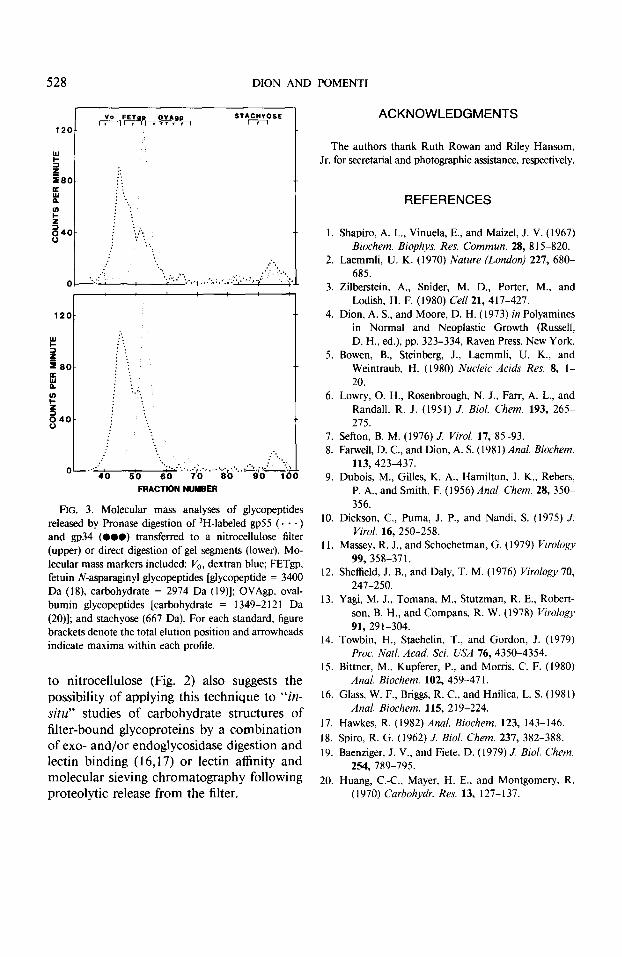
To determine whether glycopeptides re- leased by pronase digestion of gel and filter segments were similar with regard to molec- ular weights, samples from each digestion were analyzed by molecular sieving chro- matography. Figure 3 demonstrates that sim- ilar profiles were obtained for both prepara- tions for gp55 and gp34, indicating that the extents of cleavage were similar. In addition, the size exclusion profiles for the glycopep- tides of gp55 and gp34 are similar to those previously reported by Yagi et al. (13) and obtained by protease digestion of these gly- coproteins in solution (molecular weight ranges of 3750-4250 and 3200-3600 for gp34 and gp55, respectively).
The results presented in this report dem- onstrate the efficacy of proteolytic glycopep- tide release from glycoproteins transferred to nitrocellulose by two-dimensional diffusion following SDS-PAGE. Although we have not applied this technique to electrophoretically transferred ( 14,15), SDS-complexed glyco- proteins, this approach should be feasible because of the well-known attribute of Pro- nase activity in the presence of relatively high concentrations of SDS, and would also substantially reduce the time necessary for transfer. The strong binding of glycoproteins
PRONASE PRONASE r v
HOURS
FIG. 2. Kinetics of ‘H-labeled gtycopeptide release from gp55 and gp34 by Pronase digestion of glycoproteins transferred to a nitrocellulose filter following SDS-PAGE (Materials and Methods). Solid line: pronase added at 0 time and 24 h; dashed line: Pronase added at 24 h only. (0) gp55; (w) gp34. All values were normalized to lOO%, i.e., 7086 dpm for gp55 and 5604 dpm for gp34.
o L _,....' \ ; . . . '.. .
':.. 4’0 50 60
: .~..,.',,..~..... ~ ..,.,. . . . . '.2 I. iJ
70 60 90 1 0
FRACTION NUMBER
FIG. 3. Molecular mass analyses of glycopeptides released by Pronase digestion of 3H-labeled gp55 (a . . ) and gp34 (000) transferred to a nitrocellulose filter (upper) or direct digestion of gel segments (lower). Mo- lecular mass markers included: V,, dextran blue; FETgp, fetuin N-asparaginyl glycopeptides [glycopeptide = 3400 Da (18) carbohydrate = 2974 Da ( 19)]; OVAgp. oval- bumin glycopeptides [carbohydrate = 1349-212 I Da (20)]; and stachyose (667 Da). For each standard, figure brackets denote the total elution position and arrowheads indicate maxima within each profile.
to nitrocellulose (Fig. 2) also suggests the possibility of applying this technique to “in- situ” studies of carbohydrate structures of filter-bound glycoproteins by a combination of exo- and/or endoglycosidase digestion and lectin binding (16,17) or lectin affinity and molecular sieving chromatography following proteolytic release from the filter.
528 DION AND POMENTI
ACKNOWLEDGMENTS
The authors thank Ruth Rowan and Riley Hansom, Jr. for secretarial and photographic assistance, respectively.
REFERENCES
1. Shapiro, A. L., Vinuela, E., and Maizel, J. V. (1967) Biochem. Biophys. Rex Commun. 28, 815-820.
2. Laemmli, U. K. (1970) Nature (London) 227, 680-
685. 3. Zilberstein, A., Snider, M. D., Porter, M.. and
Lodish, H. F. (1980) Cell 21, 417-427. 4. Dion, A. S., and Moore, D. H. (I 973) in Polyamines
in Normal and Neoplastic Growth (Russell, D. H., ed.), pp. 323-334, Raven Press. New York.
5. Bowen, B., Steinberg, J., Laemmli, U. K., and Weintraub, H. (1980) Nucleic Acids Rex 8, l- 20.
6. Lowry, 0. H., Rosenbrough, N. J., Farr, A. L., and Randall, R. J. (1951) J. Biol. Chem. 193, 265- 275.
7. Sefton, B. M. (1976) J. Viral. 17, 85-93.
8. Far-well, D. C., and Dion, A. S. (I 98 1) Anal. Biochem.
113, 423-437. 9. Dubois. M., Gilles, K. A., Hamilton, J. K., Rebers,
P. A., and Smith, F. (1956) Anal. Chem. 28, 350-
356. 10. Dickson, C., Puma, J. P., and Nandi. S. (1975) J.
Virol. 16, 250-258. 1 I. Massey, R. J., and Schochetman, G. (1979) Virology
99, 358-37 I. 12. Sheffield, J. B., and Daly, T. M. (1976) Virol0g.v 70,
247-250. 13. Yagi, M. J., Tomana, M., Stutzman, R. E., Robert-
son, B. H., and Compans. R. W. (1978) Virology
91, 29 I-304. (1979)
(1980)
14. Towbin, H., Staehelin, T., and Gordon, J. Proc. Natl. Acad. Sci. USA 76, 4350-4354
15. Bittner, M., Kupferer, P., and Morris, C. F. Anal. Biochem. 102, 459-411.
16. Glass, W. F., Brings, R. C., and Hnilica, L. S. Anal. Biochem. 115, 219-224.
(1970) Curbohydr. Res. 13, 127-137.
(1981)
17. Hawkes, R. (1982) Anal. Biochem. 123, 143-146. 18. Spiro, R. G. (1962) J. Biol. Chem. 237, 382-388.
19. Baenziger, J. V., and Fiete, D. (1979) J. Biol. Chem. 254, 789-795.
20. Huang, C.-C., Mayer. H. E.. and Montgomery, R.





























