Embed Size (px)
Citation preview

JouRiNA OF BACTERIOLOGYVol. 88, No. 4, p. 974-980 October, 1964Copyright a 1964 American Society for Microbiology
Printed in U.S.A.
PROTEOLYTIC ATTACK OF THE CHROMOPROTEINS OFPORPHYRA BY MARINE BACTERIAL ENZYMES1
JOSEPH R. MERKEL2, GORDON D. BRAITHWAITE3, AND HENRY KRITZLER4Fort Johnson Marine Biological Laboratory, College of Charleston, Charleston, South Carolina
Received for publication 28 May 1964
ABSTRACT
MERKEL, JOSEPH R. (College of Charleston,Charleston, S.C.), GORDON D. BRAITHWAITE, ANDHENRY KRITZLER. Proteolytic attack of the chro-moproteins of Porphyra by marine bacterial en-zymes. J. Bacteriol. 88:974-980. 1964.-Threechromoproteins were isolated from the marine redalga, Porphyra leucosticta (Thuret), and werepurified by adsorption chromatography on calciumphosphate gel and by ammonium sulfate fractiona-tion. The algal chromoproteins were subjected tothe proteolytic action of cultures of actively grow-ing marine bacteria, marine bacterial culturefiltrates, trypsin, and various marine bacterialenzyme preparations. Digestion of the chromo-proteins was followed through various incubationintervals by measuring the loss of chromophorecolor and the increase in either ninhydrin-positivematerial (570 mp) or 280-m,u light-absorbing mate-rial. There was good correlation between the lossof color and enzymatic hydrolysis of the proteins.This was particularly evident when a marinepseudomonad proteinase preparation was used asthe proteolytic agent. In every digestion mixture,phycoerythrin was more resistant to degradationthan was phycocyanin or allophycocyanin. Phyco-erythrin was also more resistant to heat than wasphycocyanin or allophycocyanin.
Species of the marine red alga Porphyra containat least three chromoproteins (bilichromoproteinsor biliproteins): phycoerythrin, phycocyanin, andallophycocyanin. [There is still some question
' A preliminary account of a portion of thisstudy was reported at the Annual Meeting of theAmerican Society for Microbiology, Chicago, Ill.(Bacteriol. Proc., p. 59, 1961).
2 Present address: Department of Chemistryand the Marine Science Center, Lehigh University,Bethlehem, Pa.
Present address: National Institute for Re-search in Dairying, Shinfield, Reading, Berkshire,England.
4 Present address: Department of Biology, BardCollege, Annandale-on-Hudson, N.Y.
regarding the number of naturally occurring algalchromoproteins. Phycoerythrin and phycocyaninin the present text are the R types referred to byothers. See references by Fujiwara (1955) ando hEocha (1963) for examples of various algalchromoproteins.] Phycoerythrin is red, and diluteaqueous salt solutions of the native materialfluoresce orange in the visible light range. Solu-tions of phycocyanin are purple and fluorescered, and those of allophycocyanin are blue withred fluorescence. The color-producing prostheticgroup of each of the bilichromoproteins is atetrapyrrole derivative. To exhibit normalfluorescence, the tetrapyrrole group apparentlymust be attached to the native protein (Kylin,1910) or complexed with zinc ions (6 hEocha,1958, 1963; Lemberg, 1930). According to Kylin(1910), one of the first signs of protein denatura-tion is a loss of fluorescence.Most procedures for obtaining the chromopro-
teins from Porphyra include as a preliminary stepthe incubation of the comminuted thalli infreshwater, salt solutions, or seawater (Kylin,1910; 6 hEocha, 1958; Fujiwara, 1955; Haxo,(5 hEocha, and Norris, 1955). Probably throughthe combined effects of autolysis and bacterialdigestion the chromoproteins are freed from theirintracellular complexes (Fujiwara, 1961), andare released into the medium where further deg-radation occurs. We observed differences in theresistance of the free chromoproteins to bacterialand enzymatic digestion, and these observationsare the basis of this communication.
MATERIALS AND METHODSAlga. Most of the Porphyra used in these
studies was obtained during the winter andearly spring periods from the intertidal zonein the vicinity of Charleston, S.C. (A smallamount of Porphyra was collected in the vicinityof Woods Hole, Mass.) The South Carolina algawas identified by Louis G. Williams as P. leu-costicta (Thuret). [Identification (per.sonal com-
974
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
22
Nov
embe
r 20
21 b
y 20
0.23
6.25
1.12
7.

VOL. 88, 1964 PROTEOLYTIC ATTACK OF PORPHYRA CHROMOPROTEINS
munication) was made while Dr. Williams was onthe staff of Furman University. His present ad-dress is the National Water Quality Network,U.S. Public Health Service, Cincinnati, Ohio.]
Chromoproteins were extracted from thefreshly collected algal fronds, or the algae wereimmediately frozen or dried after collection, andwere stored under these conditions until needed.The most successful method of collecting andpreserving the algal fronds for long storageperiods was found to be the following: (i) collect-ing the algal fronds at low tide when they hadbeen partially dried, (ii) further air-drying in theshade for 6 to 8 hr, followed by (iii) oven-dryingof thinly spread fronds at 75 to 78 C (for 12 to 24hr, (iv) grinding of the dry fronds in a WaringBlendor, and (v) storing in a vacuum desiccatorover calcium chloride at 0 to -10 C.
Chromoprotein isolations and separations. Algalfronds, dried, freshly picked, or frozen, weresuspended in enough unsterile seawater or 1%sodium chloride solution to immerse the plantmaterial. The fronds were then chopped in aWaring Blendor for 2 to 3 min, transferred to500-ml Erlenmeyer flasks, and incubated at 25 Cin the dark. At 24-hr intervals, the supernatantfluids were decanted, and the remaining sedimentswere resuspended in seawater or saline solutionand reincubated. The process of incubation wascontinued usually for 72 hr at 25 C and an addi-tional 96 hr at 4 C. Bacterial activity was quiteevident at the end of the incubation period. Thesupernatant fluid from each 24-hr digestion periodwas centrifuged at about 17,000 X g to removethe particulate matter, and the soluble chro-moproteins were precipitated in the cold withammonium sulfate (30%, w/v; Baker, purified).The precipitated proteins were stored in theammonium sulfate solutions at 4 C until all ofthe supernatant fluids were precipitated. Theprotein precipitates were redissolved in a 1%aqueous NaCl solution and were dialyzed for 48hr against large amounts of distilled water or 1%NaCl.The dialyzed chromoproteins were then sepa-
rated on calcium phosphate gel columns, preparedaccording to the directions of Swingle and Tiselius(1951), and by ammonium sulfate fractionation.Elution of the proteins from the gel columns wasaccomplished with increasing amounts of 0.067 Mphosphate buffer (pH 7.4) added to 1% NaCl.Fractions containing the major amounts of each
of the chromoproteins were precipitated withammonium sulfate (30%, w/v), redissolved in 1%NaCl, and dialyzed against distilled water or 1%NaCl. The chromatographic process was repeateduntil a single component was obtained from eachof the major fractions. The purity of the isolatedchromoproteins was monitored by spectral meas-urements (Beckman DU spectrophotometer).
Proteolytic attack of chromoproteins. Millipore-filtered solutions of the individual chromoproteinsand of mixtures of the three chromoproteins weresubjected to proteolytic digestion by growingsuspensions of mixed marine bacteria, by Milli-pore filtrates of the media in which the marinebacteria had grown, and by bacteria-free enzymepreparations which were obtained from filtratesof Aeromonas proteolytica (nov.sp.) (Merkel et al.,1964; Prescott and Willms, 1960) by ammoniumsulfate (40 to 50% saturated) precipitation. [Itwas recently shown (Prescott and Willms, 1960)that the principal proteinase is actually a mixtureof at least two enzymes.] This was accomplishedby adding the chromoproteins to sterile seawaterwhich, in some experiments, was enriched withpeptones; inoculating the protein solutions withthe sterile, crude source of enzyme or heavy 18- to24-hr bacterial suspension; and incubating at25 C in the dark. The loss of chromophore colorduring incubation was followed visually andrecorded quantitatively with a Beckman DUspectrophotometer by reading the absorbance ata maximum for the chromophore of each protein.Boiled preparations were always added to controltubes to distinguish between proteolytic activityand physical and chemical losses.
Pure chromoproteins were used to determinethe relationship between the loss of chromophorecolor and protein hydrolyzed. In a typical experi-ment, 6 ml of each of the sterile chromoproteinsolutions dissolved in 0.067 M phosphate bufferat pH 7.4 were inoculated with 0.2 ml of steriletrypsin (twice-crystallized preparation; Nutri-tional Biochemicals Corp., Cleveland, Ohio; 10mg/ml in pH 7.4 buffer). Controls were inoculatedwith the same trypsin preparation after boilingit for 30 min. The optical densities of the mixtureswere read at the specific absorption maximaimmediately after adding the enzyme solutionand after 8 hr of incubation. The reaction wasstopped by adding 1 ml of 10% trichloroaceticacid to 2.5 ml of the enzymae-protein mixtures.The mixtures were boiled for 30 min, the precipi-
975
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
22
Nov
embe
r 20
21 b
y 20
0.23
6.25
1.12
7.
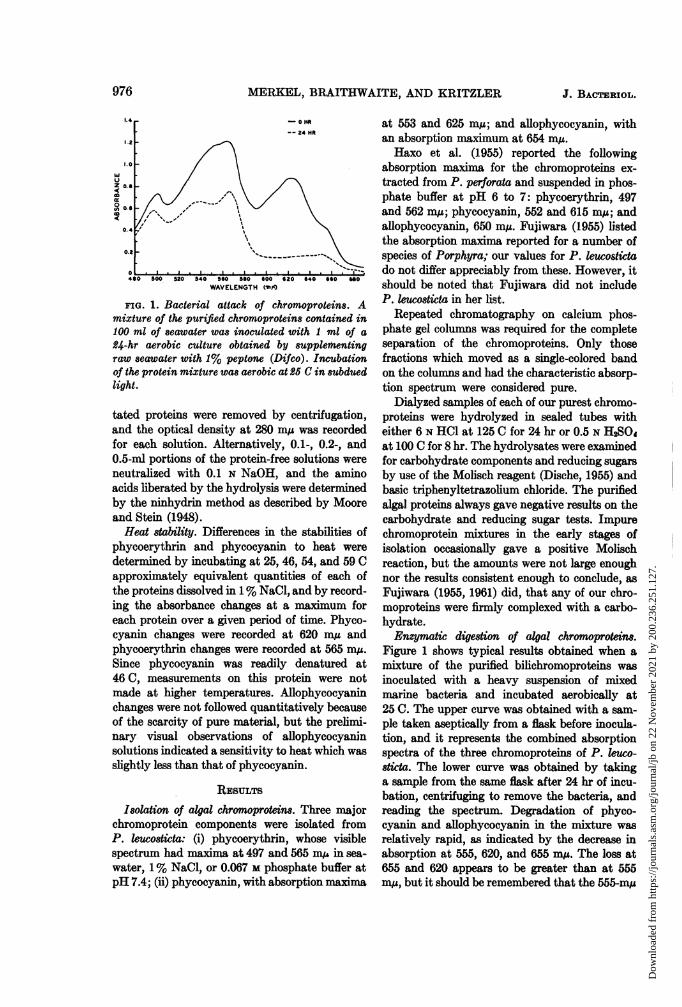
MERKEL, BRAITHWAITE, AND KRITZLER
Z
0.4
0.2
WAVELENGTH (O"$
FIG. 1. Bacterial attack of chromoproteins. Amixture of the purified chromoproteins contained in100 ml of seawater was inoculated with 1 ml of a
24-hr aerobic culture obtained by -supplementingraw seawater with 1% peptone (Difco). Incubationof the protein mixture was aerobic at 2£ C in subduedlight.
tated proteins were removed by centrifugation,and the optical density at 280 mu was recordedfor each solution. Alternatively, 0.1-, 0.2-, and0.5-nd portions of the protein-free solutions wereneutralized with 0.1 N NaOH, and the aminoacids liberated by the hydrolysis were determinedby the ninhydrin method as described by Mooreand Stein (1948).Heat stability. Differences in the stabilities of
phycoerythrin and phycocyanin to heat were
determined by incubating at 25, 46, 54, and 59 Capproximately equivalent quantities of each ofthe proteins dissolved in 1% NaCl, and by record-ing the absorbance changes at a maximum foreach protein over a given period of time. Phyco-cyanin changes were recorded at 620 m,u andphycoerythrin changes were recorded at 565 m,u.Since phycocyanin was readily denatured at46C, measurements on this protein were notmade at higher temperatures. Allophycocyaninchanges were not followed quantitatively becauseof the scarcity of pure material, but the prelimi-nary visual observations of allophycocyaninsolutions indicated a sensitivity to heat which wasslightly less than that of phycocyanin.
REsuLTs
Isolation of algal chromoprotins. Three majorchromoprotein components were isolated fromP. leucosticta: (i) phycoerythrin, whose visiblespectrum had maxima at 497 and 565 m, in sea-water, 1% NaCl, or 0.067 M phosphate buffer atpH 7.4; (ii) phycocyanin, with absorption maxima
at 553 and 625 m,u; and allophycocyanin, withan absorption maximum at 654 m,u.Haxo et al. (1955) reported the following
absorption maxima for the chromoproteins ex-tracted from P. perforata and suspended in phos-phate buffer at pH 6 to 7: phycoerythrin, 497and 562 m,u; phycocyanin, 552 and 615 m,u; andallophycocyanin, 650 m,u. Fujiwara (1955) listedthe absorption maxima reported for a number ofspecies of Porphyra; our values for P. leucostictado not differ appreciably from these. However, itshould be noted that Fujiwara did not includeP. leostticta in her list.
Repeated chromatography on calcium phos-phate gel columns was required for the completeseparation of the chromoproteins. Only thosefractions which moved as a single-colored bandon the colums and had the characteristic absorp-tion spectrum were considered pure.
Dialyzed samples of each of our purest chromo-proteins were hydrolyzed in sealed tubes witheither 6 N HC1 at 125 C for 24 hr or 0.5 N H2SO4at 100 C for 8 hr. The hydrolysates were examinedfor carbohydrate components and reducing sugarsby use of the Molisch reagent (Dische, 1955) andbasic triphenyltetrazolium chloride. The purifiedalgal proteins always gave negative results on thecarbohydrate and reducing sugar tests. Impurechromoprotein mixtures in the early stages ofisolation occasionally gave a positive Molischreaction, but the amounts were not large enoughnor the results consistent enough to conclude, asFujiwara (1955, 1961) did, that any of our chro-moproteins were firmly complexed with a carbo-hydrate.Enzymatic digestion of algal chromoprotins.
Figure 1 shows typical results obtained when amixture of the purified bilichromoproteins wasinoculated with a heavy suspension of mixedmarine bacteria and incubated aerobically at25 C. The upper curve was obtained with a sam-ple taken aseptically from a flask before inocula-tion, and it represents the combined absorptionspectra of the three chromoproteins of P. leuco-8ticta. The lower curve was obtained by takinga sample from the same flask after 24 hr of incu-bation, centrifuging to remove the bacteria, andreading the spectrum. Degradation of phyco-cyanin and allophycocyanin in the mixture wasrelatively rapid, as indicated by the decrease inabsorption at 555, 620, and 655 mu. The loss at655 and 620 appears to be greater than at 555m,u, but it should be remembered that the 555-mg
976 J. BACTERIOL..
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
22
Nov
embe
r 20
21 b
y 20
0.23
6.25
1.12
7.
VOL. 88, 1964 PROTEOLYTIC ATTACK OF PORPHYRA CHROMOPROTEINS
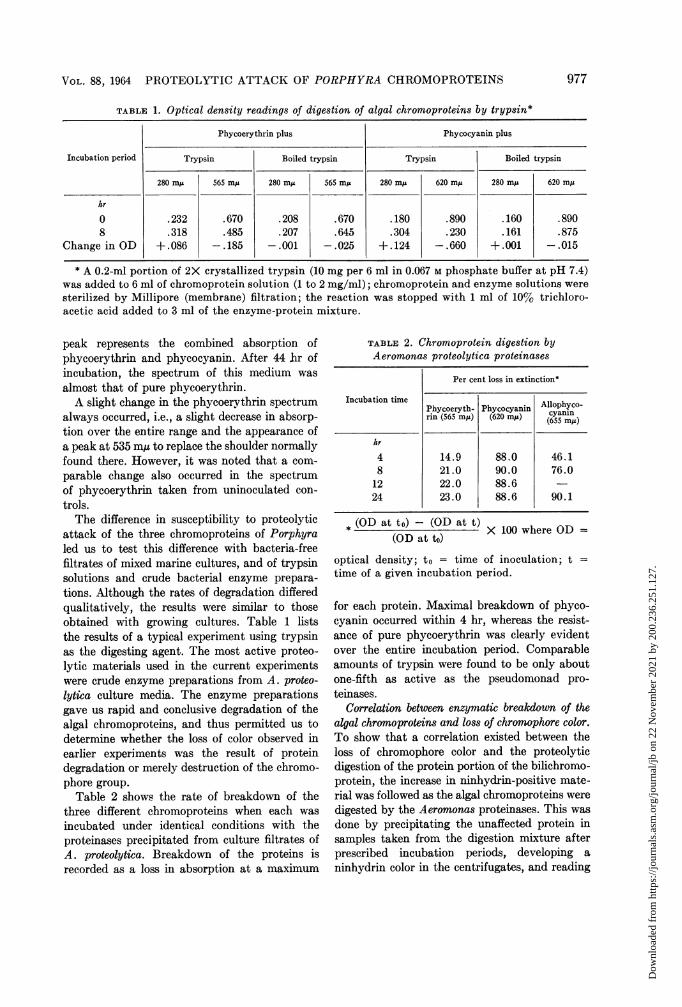
TABLE; 1. Optical density readings of digestion of algal chromoproteins by trypsin*
Phycoerythrin plus Phycocyanin plus
Incubation period Trypsin Boiled trypsin Trypsin Boiled trypsin
280 myA 565 m,u 280 m,u 565 m,u 280 m,u 620 mys 280 mxj 620 m,u
hr
0 .232 .670 .208 .670 .180 .890 .160 .8908 .318 .485 .207 .645 .304 .230 .161 .875
Change in OD +.086 -.185 -.001 -.025 +.124 -.660 +.001 -.015
* A 0.2-ml portion of 2X crystallized trypsin (10 mg per 6 ml in 0.067 M phosphate buffer at pH 7.4)was added to 6 ml of chromoprotein solution (1 to 2 mg/ml); chromoprotein and enzyme solutions weresterilized by Millipore (membrane) filtration; the reaction was stopped with 1 ml of 10% trichloro-acetic acid added to 3 ml of the enzyme-protein mixture.
peak represents the combined absorption ofphycoerythrin and phycocyanin. After 44 hr ofincubation, the spectrum of this medium was
almost that of pure phycoerythrin.A slight change in the phycoerythrin spectrum
always occurred, i.e., a slight decrease in absorp-tion over the entire range and the appearance ofa peak at 535 m, to replace the shoulder normallyfound there. However, it was noted that a com-
parable change also occurred in the spectrumof phycoerythrin taken from uninoculated con-
trols.The difference in susceptibility to proteolytic
attack of the three chromoproteins of Porphyraled us to test this difference with bacteria-freefiltrates of mixed marine cultures, and of trypsinsolutions and crude bacterial enzyme prepara-
tions. Although the rates of degradation differedqualitatively, the results were similar to thoseobtained with growing cultures. Table 1 liststhe results of a typical experiment using trypsinas the digesting agent. The most active proteo-lytic materials used in the current experimentswere crude enzyme preparations from A. proteo-lytica culture media. The enzyme preparationsgave us rapid and conclusive degradation of thealgal chromoproteins, and thus permitted us todetermine whether the loss of color observed inearlier experiments was the result of proteindegradation or merely destruction of the chromo-phore group.
Table 2 shows the rate of breakdown of thethree different chromoproteins when each was
incubated under identical conditions with theproteinases precipitated from culture filtrates ofA. proteolytica. Breakdown of the proteins isrecorded as a loss in absorption at a maximum
TABLE 2. Chromoprotein digestion byAeromonas proteolytica proteinases
Per cent loss in extinction*
Incubation timePhycoeryth- Phycocyanin cyanlynrin (565 m/A) (620 MAs) cy5 mAni
hr
4 14.9 88.0 46.18 21.0 90.0 76.012 22.0 88.624 23.0 88.6 90.1
* (OD at to)- (OD at t) X 100 where OD =(GD at to)
optical density; to = time of inoculation; t =
time of a given incubation period.
for each protein. Maximal breakdown of phyco-cyanin occurred within 4 hr, whereas the resist-ance of pure phycoerythrin was clearly evidentover the entire incubation period. Comparableamounts of trypsin were found to be only aboutone-fifth as active as the pseudomonad pro-teinases.
Correlation between enzymatic breakdown of thealgal chromoproteins and loss of chromophore color.To show that a correlation existed between theloss of chromophore color and the proteolyticdigestion of the protein portion of the bilichromo-protein, the increase in ninhydrin-positive mate-rial was followed as the algal chromoproteins weredigested by the Aeromonas proteinases. This wasdone by precipitating the unaffected protein insamples taken from the digestion mixture afterprescribed incubation periods, developing aninhydrin color in the centrifugates, and reading
977
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
22
Nov
embe
r 20
21 b
y 20
0.23
6.25
1.12
7.
MERKEL, BRAITHWAITE, AND KRITZLER
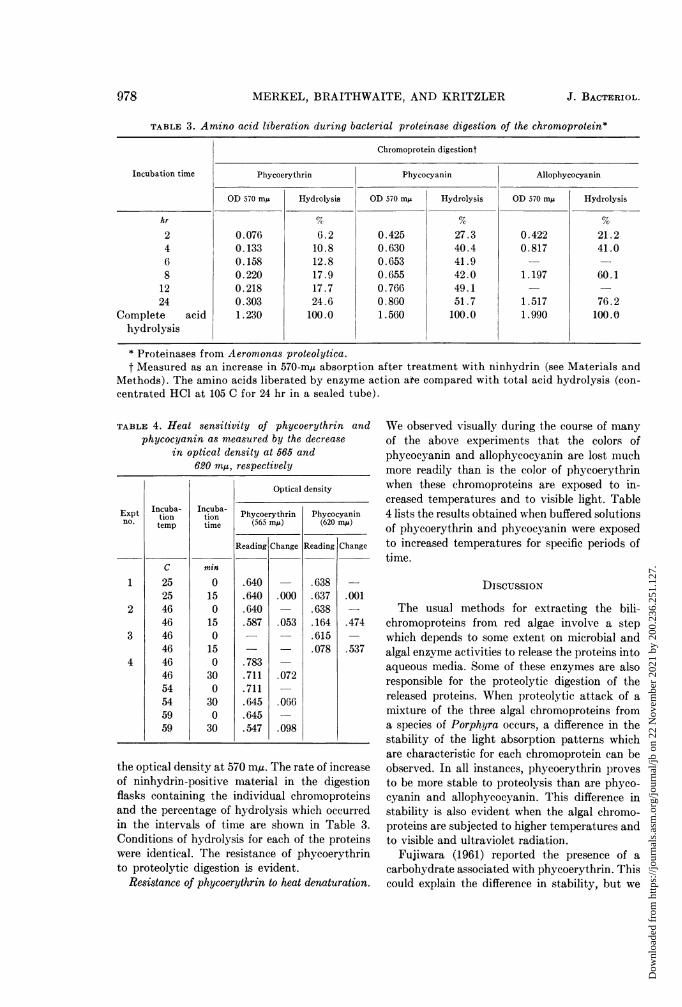
TABLE 3. Amino acid liberation during bacterial proteinase digestion of the chromoprotein*
Chromoprotein digestiont
Incubation time Phycoerythrin Phycocyanin Allophycocyanin
OD 570 my Hydrolysis OD 570 mu Hydrolysis OD 570 mp Hydrolysis
hr 7c G %2 0.076 6.2 0.425 27.3 0.422 21.24 0.133 10.8 0.630 40.4 0.817 41.06 0.158 12.8 0.653 41.9 -
8 0.220 17.9 0.655 42.0 1.197 60.112 0.218 17.7 0.766 49.1 -
24 0.303 24.6 0.860 51.7 1.517 76.2Complete acid 1.230 100.0 1.560 100.0 1.990 100.0
hydrolysis
* Proteinases from Aeromonas proteolytica.t Measured as an increase in 570-m, absorption after treatment with ninhydrin (see Materials and
Methods). The amino acids liberated by enzyme action ate compared with total acid hydrolysis (con-centrated HCl at 105 C for 24 hr in a sealed tube).
TABLE 4. Heat sensitivity of phycoerythrin andphycocyanin as measured by the decrease
in optical density at 565 and620 mAs, respectively
Expt Incuba-n.
tiontemp
1
2
3
4
C
252546464646464654545959
Incuba-tiontime
min
0
150
150
150
300
300
30
Optical density
Phycocyanin(620 mIA)
Reading Change
.638
.637
.638
.164
.615
.078
.001
.474
.537
Phycoerythrin(565 my)
Reading Change
.640
.640
.640
.587
.783
.711
.711
.645
.645
.547
.000
.053
.072
.066
.098
the optical density at 570 mu. The rate of increaseof ninhydrin-positive material in the digestionflasks containing the individual chromoproteinsand the percentage of hydrolysis which occurredin the intervals of time are shown in Table 3.Conditions of hydrolysis for each of the proteinswere identical. The resistance of phycoerythrinto proteolytic digestion is evident.
Resistance of phycoerythrin to heat denaturation.
We observed visually during the course of manyof the above experiments that the colors ofphycocyanin and allophycocyanin are lost muchmore readily than is the color of phycoerythrinwhen these chromoproteins are exposed to in-creased temperatures and to visible light. Table4 lists the results obtained when buffered solutionsof phycoerythrin and phycocyanin were exposedto increased temperatures for specific periods oftime.
DISCUSSION
The usual methods for extracting the bili-chromoproteins from red algae involve a stepwhich depends to some extent on microbial andalgal enzyme activities to release the proteins intoaqueous media. Some of these enzymes are alsoresponsible for the proteolytic digestion of thereleased proteins. When proteolytic attack of amixture of the three algal chromoproteins froma species of Porphyra occurs, a difference in thestability of the light absorption patterns whichare characteristic for each chromoprotein can beobserved. In all instances, phycoerythrin provesto be more stable to proteolysis than are phyco-cyanin and allophycocyanin. This difference instability is also evident when the algal chromo-proteins are subjected to higher temperatures andto visible and ultraviolet radiation.
Fujiwara (1961) reported the presence of acarbohydrate associated with phycoerythrin. Thiscould explain the difference in stability, but we
978 J . BACTERI OL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
22
Nov
embe
r 20
21 b
y 20
0.23
6.25
1.12
7.

VOL. 88, 1964 PROTEOLYTIC ATTACK OF PORPHYRA CHROMOPROTEINS
found no evidence for carbohydrates in our purechromoprotein preparations. Because of the re-ported similarities of the molecular weights,amino acid composition, and terminal aminoacids (Fujiwara, 1956; Kimmel and Smith,1958), the differences in the stability of the pro-teins must reside in some differences in the struc-tures (primary, secondary, or tertiary) of theproteins. 6 hEocha (1958) speculated on thepossible differences in the globulin structures toaccount for the different phycoerythrins havingsimilar chromophore groups.With the apparent exception of phycocyanin,
there is good correlation between the loss of colorand hydrolysis of the protein during proteolyticdigestion. If this were not the case, one mightassume that the rapid loss of color which weobserved was simply degradation or alteration ofthe chromophore groups. There was degradationof the tetrapyrrole structures, as evidenced bythe complete loss of color in phycocyanin andallophycocyanin solutions which were incubatedwith either growing bacteria or sterilized bacterialproteinases. Even the color of phycoerythrin islost after lengthy incubations with A. proteolyticaproteinases. The difference between the rate andamount of color loss and amino acid productionduring digestion of phycocyanin can be attributedto the instability of phycocyanin at the incuba-tion temperatures and relatively dilute proteinconcentrations used in the above experiment. Wenoted that, after a brief incubation period, phyco-cyanin was precipitating out of solution. Theprecipitated protein no longer contributed to thecolor of the solution, but the proteolytic digestionand, hence, amino acid liberation continued.A slight alteration also occurs in the spectrum
of phycoerythrin when sterile solutions are incu-bated at room temperature. This change mani-fests itself as a slight decrease in the overallabsorption and the appearance of a peak at 535m,u to replace the shoulder located at this pointin the spectrum of the freshly prepared phyco-erythrin solutions. Two-peaked phycoerythrinfrom Phormidium species was reported by6 hEocha and Haxo (1960) as an artifact arisingduring prolonged extraction procedures.
In most of our experiments with bacterialenzymes, the only color remaining at the end ofthe incubation period was that of phycoerythrin.Thinking that the color could be either freephycoerythrobilin (Kitasato, 1925; Lemberg,1930) or the tetrapyrrole attached to a short
peptide (Fujiwara, 1957), we washed portionsof the phycoerythrin digestion mixtures withamyl alcohol, but no pigment was extracted.Fingerprinting of the amino acids and peptidesreleased by the enzyme action gave results similarto prolonged acid digestion; i.e., no chromopep-tide was found. It is still possible that very shortincubation periods with the bacterial enzymeswill show that the chromophore group is attackedor detached before the remainder of the complexis hydrolyzed. Fujiwara (1957) obtained chromo-peptides after 2-hr digestion of phycoerythrinwith pepsin. We have also been able to isolatechromopeptides from our trypsin digestion mix-tures which have been incubated less than 8 hr,and, since our bacterial enzyme preparations areconsiderably more active than the animal en-zymes, we may find chromopeptides at very earlystages of digestion.The chromoproteins of the red and blue-green
algae are generally considered to function asaccessory pigments in photosynthesis. Haxo andBlinks (1950) demonstrated an excellent correla-tion between the absorption spectrum of phyco-erythrin and the photosynthetic action spectrumof a number of red algae. The possibility alsoexists that phycoerythrin has functions otherthan photosynthetic (Boney and Corner, 1960).Our findings concerning the relative stability ofphycoerythrin suggest that this pigment eitheraids in survival or is the one chromoproteincapable of surviving adverse conditions. P.perforata is reported by Haxo and Blinks tocontain excessive amounts of phycoerythrin inthe carposporic areas a few days before the releaseof reproductive cells.
ACKNOWLEDGMENTS
We are grateful for the assistance of SophiaWaring, Mary Bee, Rachel Merkel, WilliamMurphy, and David Somerstein (NationalScience Foundation undergraduate researchparticipant) in some phases of these studies.We are indebted to George Grice, Jr. for sup-
plying the Porphyra from Woods Hole, Mass.,and to J. M. Prescott for the preparation of mostof the Aeromronas proteolytica proteinases used inthis study.The work was aided by a contract between
the College of Charleston and the Office of NavalResearch (Nonr. 2439-01), and was completedwith the aid of a contract between LehighUniversity and the Office of Naval Research
979
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
22
Nov
embe
r 20
21 b
y 20
0.23
6.25
1.12
7.

MERKEL, BRAITHWAITE, AND KRITZLER
(Nonr. 610-05). We also wish to thank SmithKline & French Laboratories for its grant toLehigh University to purchase biochemicalequipment used in some parts of this study.
LITERATURE CITED
BONEY, A. D., AND E. D. S. CORNER. 1960. A possi-ble function of phycoerythrin in intertidalred algae. Nature 188:1042-1043.
DISCHE, Z. 1955. New color reactions for deter-mination of sugars in polysaccharides. Meth-ods Biochem. Analy. 2:320-321.
FUJIWARA, T. 1955. Studies on chromoproteins inJapanese Nori (Porphyra tenera). 1. A newmethod for the crystallization of phyco-erythrin and phycocyanin. J. Biochem.(Tokyo) 42:411-417.
FUJIWARA, T. 1956. Studies on chromoproteins inJapanese Nori (Porphyra tenera). II. Aminoacid composition of phycoerythrin and phyco-cyanin. J. Biochem. (Tokyo) 43:195-203.
FUJIWARA, T. 1957. Studies on chromoproteins inJapanese Nori (Porphyra tenera). III. Chro-mopeptides derived from phycoerythrin bypeptic digestion. J. Biochem. (Tokyo) 44:723-733.
FUJIWARA, T. 1961. Studies on chromoproteins inJapanese Nori (Porphyra tenera). V. On thesugar components of phycoerythrin. J. Bio-chem. (Tokyo) 49:361-367.
HAXO, F. T., AND L. R. BLINKS. 1950. Photosyn-thetic action spectra of marine algae. J. Gen.Physiol. 33:389-422.
HAXO, F., C. 6 HEOCHA, AND P. NORRIS. 1955.Comparative studies of chromatographicallyseparated phycoerythrins and phycocyanins.Arch. Biochem. Biophys. 54:162-173.
KIMMEL, J. R., AND E. L. SMITH. 1958. The amino
acid composition of crystalline pumpkin seedglobulin, edestin, c-phycocyanin, and r-phycoerythrin. Bull. Soc. Chim. Biol. 40:2049-2063.
KITASATO, Z. 1925. Biochemical studies of phyco-erythrin and phycocyanin. Acta Phytochim.2:75-97.
KYLIN, H. 1910. Uber Phykoerythrin und Phyko-cyan bei Ceramium rubrum (Huds.) Ag., H-s.Z. Physiol. Chem. 69:169-239.
LEMBERG, R. 1930. Chromoprotein of red algae. II.Scission with pepsin and acid. Isolation of apyrrole dyestuff. Ann. Chem. 477:195-245.
MERKEL, J. R., E. D. TRAGANZA, B. B. MUKHER-JEE, T. B. GRIFFIN, AND J. M. PRESCOTT. 1964.Proteolytic activity and general characteris-tics of a marine bacterium, Aeromonas proteo-lytica, sp.n. J. Bacteriol. 87:1227-1233.
MOORE, S., AND W. H. STEIN. 1948. Photometricninhydrin method for use in the chroma-tography of amino acids. J. Biol. Chem.176:367-388.
O HEOCHA, C. 1958. Comparative biochemicalstudies of the phycobilins. Arch. Biochem.Biophys. 73:207-219.
dHEOCHA, C., AND F. T. HAXO. 1960. Some atypi-cal chromoproteins. Biochim. Biophys. Acta41:516-520.
O HEOCHA, C. 1963. Spectral properties of thephycobilins. I. Phycocyanobilin. Biochemis-try 2:375-382.
PRESCOTT, J. M., AND C. R. WILLMS. 1960. Somecharacteristics of proteolytic systems of amarine bacterial species. Proc. Soc. Exptl.Biol. Med. 103:410-413.
SWINGLE, S. M., AND A. TIsELIUS. 1951. Tricalciumphosphate as an adsorbent in the chroma-tography of proteins. Biochem. J. 48:171-174.
980 J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
22
Nov
embe
r 20
21 b
y 20
0.23
6.25
1.12
7.





























