Embed Size (px)
Citation preview

238
Biochimica et Biophysica Acta, 476 (1977) 238--252 © Elsevier/North-Holland Biomedical Press
BBA 98929
PROTEINS OF THE POLYSOMAL MESSENGER RIBONUCLEOPROTEIN ARE RESPONSIBLE FOR ITS ASSOCIATION WITH THE 4 0 ~ RIBOSOMAL SUBUNIT IN HeLa CELLS
J.-P. LIAUTARD
Biologisches Inst i tut der Universitiit Stuttgart, Ulmer Strasse 227, D-7000 Stuttgart 60 (G.F.R.)
(Received September 8th, 1976)
Summary
The role of proteins bound to mRNA is one of the key problems in the s tudy of polysomal messenger ribonucleoprotein complexes. In this paper it is shown that the use of ethylenediamine tetraacetic (EDTA) should be avoided during the purification of the mRNA • protein complexes. The property of cul- tured cells to release mRNA from polysomes at high temperature (43°C) was used to purify polysomal messenger ribonucleoprotein complexes. The poly- somal messenger ribonucleoprotein complexes so obtained bind to salt washed ribosomes even at 0°C. Under the same conditions naked mRNA does not bind to ribosomes. These experiments indicate that certain proteins attached to mRNA are essential for its activity. The polysomal messenger ribonucleopro- tein complexes are bound to the small subunits and their mRNA is partially protected by the ribosomes. This association is not an artificial aggregation.
The study of the complexes reveals that neither poly(A) sequences nor double-stranded regions of the mRNA, but a protein of the polysomal mes- senger ribonucleoprotein complexes are involved in the binding.
The described polysomal messenger r ibonucleoprotein, ribosome complex does not represent a complete initiation complex because some initiation factors are absent and must be added to assure protein synthesis.
Introduct ion
The existence of natural messenger RNA protein complexes (mRNP) in eukaryotic cells is firmly established [1--7], although the function of their pro- teins is still debated [8].
Abbreviat ions: m R N P , c o m p l e x e s of messenger R N A wi th proteins~ E D T A , e thy lened iamine tetraacetic acid.

2 3 9
Lebleu et al. [5], have shown that mRNP can bind to deoxycholate-washed ribosomes under conditions where mRNA does not, which led then to the con- clusion that the proteins are necessary for binding mRNA to ribosomes. But the in vitro translation of mRNP and mRNA does not exhibit any difference in activity [9,10--12]. If the proteins bound to the mRNA are specific, as demon- strated [5--7, 13,14] they have some function in the living cell. It is of course possible that this function does not interfere with protein synthesis, but it is also possible that the absence of difference in activity in vitro between messen- ger RNA • protein and naked RNA is due to inadequate experimental condi- tions. It is quite clear that the first difficulty in the study of the function of the proteins bound to mRNA arises when unfractionated preparations are em- ployed (i.e. $100 supernatant, pH 5 enzyme or unwashed ribosomes}. The original work of Lebleu et al. [5] supports this assumption. These authors found that mRNP and messenger RNA bind to unwashed ribosomes equally well, but only messenger RNA • protein complexes associate with deoxycholate- washed ribosomes, These results suggest the use of washed particles when studying the relationship between:ribosomes and mRNA.
A second problem in the study of the function of proteins bound to mRNA can arise during the purification procedure of mRNA. The release of messenger r ibonucleoproteins f rom polysomes generally involves the use of EDTA. Although ~it is accepted that EDTA irreversibly denatures ribosomal subunits [15,16], it has been claimed that mRNPs are resistant to this agent [2,17]. This assumption was made from buoyant density analysis and not on activity measurements.
In this paper it is shown that, at least under the experimental conditions used, the polysomal mRNA are seriously affected by EDTA treatment. To overcome this difficulty the polysomal messenger r ibonucleoproteins were pre- pared by in vivo temperature shock followed by a high salt treatment. Further, salt-washed ribosomes and salt-washed polysomal messenger r ibonucleoproteins can rebind under conditions where purified mRNA does not. The s tudy of this binding shows that this is not artificial aggregation. A mRNA-associated protein (Mr I 48 000) seems to be involved in the binding step.
Materials and Methods
1. Buffers used. buffer A: 10 mM Tris . HC1 (pH 7.4)/10 mM KC1/3 mM MgCI2/7 mM mercaptoethanol; buffer B: 10 mM Tris. HC1 (pH 7.4)/500 mM KC1/3 mM MgC12/7 mM mercaptoethanol; buffer C: 50 mM Tris. HC1 (pH 8.1}/10 mM NH4C1/3 mM MgC12/7 mM mercaptoethanol; buffer D: 10 mM Tris . HC1 (pH 7.4)/140 mM KC1/3 mM MgC12/; buffer E: 50 mM Tr i s -HCI (pH 7.4)/1 mM EDTA/100 mM NaC1/0.1% Sarkosyl; buffer F: 20 mM phos- phate (pH 7.5)/0.2% SDS/15 mM mercaptoethanol; buffer G: 10 mM Tris • HC1 (pH 7.4)/60 mM KCI/3 mM magnesium acetate/3.5 mM mercaptoethanol.
2. Cell culture and labeling procedure. HeLa cells were grown as suspension culture to a concentration of 5 • 10 s cells/ml in Eagle's medium supplemented with 5% calf serum. When cells were labeled, they were first concentrated ten times by low speed centrifugation. To the concentrated suspension were added: act inomycin D (0.05 #g/ml) and 20 min thereafter radioactive uridine (1.5 pCi/

240
ml). The suspension was stirred for an additional 3 h. The cells were either har- vested or the temperature was raised to 43°C by incubating the flask in a water bath for 20 min.
3. Cell harvest and fractionation procedure. Cells were poured on crushed frozen buffer D (--80°C), washed once, and resuspended in cold buffer A. After 10 min swelling the cells were homogenized by ten strokes in a Dounce homogenizer. Nuclei were sedimented by low speed centrifugation (700 × g, 4 min, 4°C). The supernatant was centrifuged to remove membranes and mito- chondria (Sorvall 30 000 X g, 20 min, 4°C), yielding a postmitochondrial supernatant.
4. RNA purification and analysis. RNA was extracted by the chloroform/ phenol method according to Perry et al. [18]. Purified RNA was further frac- t ionated by electrophoresis in agarose-acrylamide gels, as described by Thiollais et al. [19]. Special care was taken to avoid traces of RNAase: hence the buffers contained 0.2% sodium dodecylsulfate. The gels were made of 2.2% acrylamide and 0.5% agarose in 12 cm tubes. Electrophoresis was carried out at 9V per tube for 2 h using bromophenolblue as tracking dye. At the end of the run the gels were removed, frozen and cut into 1 mm slices. In order to elute t tNA, the slices were agitated in 0.5 ml of water overnight. The eluate was added to 6 ml of Bray's scintillation fluid [53], and the radioactivity was determined.
5. Poly(A) analysis. They presence of poly(A) was determined by its ability to bind poly(U)-sepharose according to the procedure of Adesnik et al. [20] modified by Lindberg and Person [21]. Poly(U)-Sepharose purchased from Pharmacia was used.
6. Purification of polysomal messenger ribonucleoprotein by centrifugation in Cs2S04. The postmitochondrial supernatant from cells treated at 43°C was layered onto a 1.5 ml 30% sucrose cushion in buffer B and centrifuged 16 h at 25 000 rev./min in a SW 50.1 rotor. The pellet was suspended in buffer B and diluted with an equal amount of 15% (w/v) Cs2SO4 solution (Cs2SO4 solution actually containing 10 mM Tris • HC1 pH 7.4 and 2 mM MgC12). 20 A260 of the r ibosome solution was layered onto 15--44% (w/w) Cs2SO4 preformed gra- dients and centrifuged 24 h at 32 000 rev./min in a SW 40 rotor (4°C). The gra- dients were fractionated and the absorbance at 254 nm monitored. The selec- tivity labeled mRNA bands at a density of p = 1.36 g/cm 3. This fraction does not contain detectable rRNA. Three major proteins (MA 75 000, 50 000, 3 6 0 0 0 ) are bound to the m R N A (to be published). This fraction was used for reassociation experiments, mRNA was extracted from this fraction with the chloroform/phenol method as described by Perry et al. [18].
7. Cell-free protein synthetizing system. The cell-free protein synthetizing system (100 t~l samples) contained: 1 A~0 ribosomes, 0.1 pmol ATP, 0.25 pmol phosphoenolpyruvate, 0.05 t~mol GTP. 1 pg phosphoenolpyruvate kinase, 20 #l S100 (approx. 15 pg tRNA and 0.1 mg proteins) if indicated, 20 pl ribosom- al wash (approx. 40 t~g proteins) 0.125 #mol of each amino acid, 1 pmol Tris • HC1 pH 7.4, 6 ttmol KCI, 0.3 ttmol magnesium acetate, 0.35 ttmol mercapto- ethanol, 0.03 pCi [3H]leucine and 0.01 pmol aurin tricarboxylic acid when mentioned. The incubation time was 1 h at 37°C. The reaction was stopped and analysed according to Mans and Novelli [22]. The amino acid incorpora- tion was linear for approximately 30 min. This system was total ly inhibited

241
by cycloheximide but not by aurin tricarboxylic acid. 8. Other methods employed. The sucrose gradient electrophoresis procedure
has been described previously [23]. Ultracentrifugation was performed at 4°C with Beckman L2 centrifuges, and the sedimentation coefficients were deter- mined according to McEven [24].
For the radioactivity measurement, trichloroacetic acid-precipitated frac- tions were filtered on glass-fibre filters. The samples were counted in a Packard scintillation counter. Counting efficiency was about 50%.
Protein gel electrophoresis was performed in 7.5% acrylamid with 0.1% SDS [25] using a Shandon apparatus; the gels were stained with Coomassie brilliant blue [25]. The preparation of proteins associated with mRNA has been described in detail previously [14].
Results
I. Damaging effect o f EDTA on polysomal messenger ribonucleoproteins o f HeLa cells
To reconstruct an initiation complex in vitro it is essential to obtain all com- ponents in a non-denatured form.
Messenger RNA can be dislodged from polysomes as ribonucleoprotein com- plexes, and many laboratories have prepared polysomal messenger ribonucleo- proteins from polysomes by suspending them in buffered EDTA [1,3--5]. However, working with HeLa cells, polysomal messenger ribonucleoproteins that can be rebound to salt washed ribosomes were not obtainable by this pro- cedure.
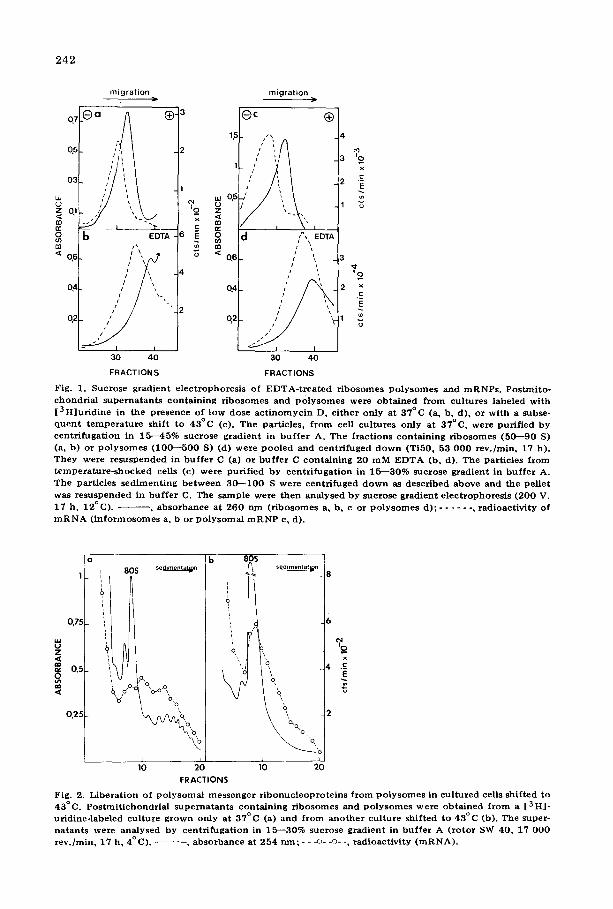
EDTA-treated ribonucleoprotein particles were closely investigated by sucrose gradient electrophoresis. This is a rather sensitive procedure for detect- ing minor surface changes [23], showing that EDTA-treated HeLa cell ribo- somes and inforrnosomes have drastically altered mobilities in comparison to untreated particles (Figs. la and b). Dissociation of a polysomal fraction in buf- fered EDTA/saline yields ribosomal subunits and polysomal messenger ribo- nucleoproteins which migrate very fast (Fig, ld). They have mobilities analo- gous to EDTA-treated free cytoplasmic messenger ribonucleoproteins (i.e. informosomes) and native subunits.
Apparently the increased mobility reflects a change in the particles structure. Ribosomal particles seem to be affected to the same extent (Fig, 1), and it has been shown that EDTA-treated ribosomes and irreversibly damaged and cannot support protein synthesis [16,26]. Therefore, loss of activity of the proteins associated with mRNA may be due to the damaging effect of EDTA. Thus, it was attempted to obtain polysomal messenger ribonucleoproteins without using chelating agents.
II. Polysomal messenger ribonucleoproteins are released from polysomes o f temperature-shocked cells
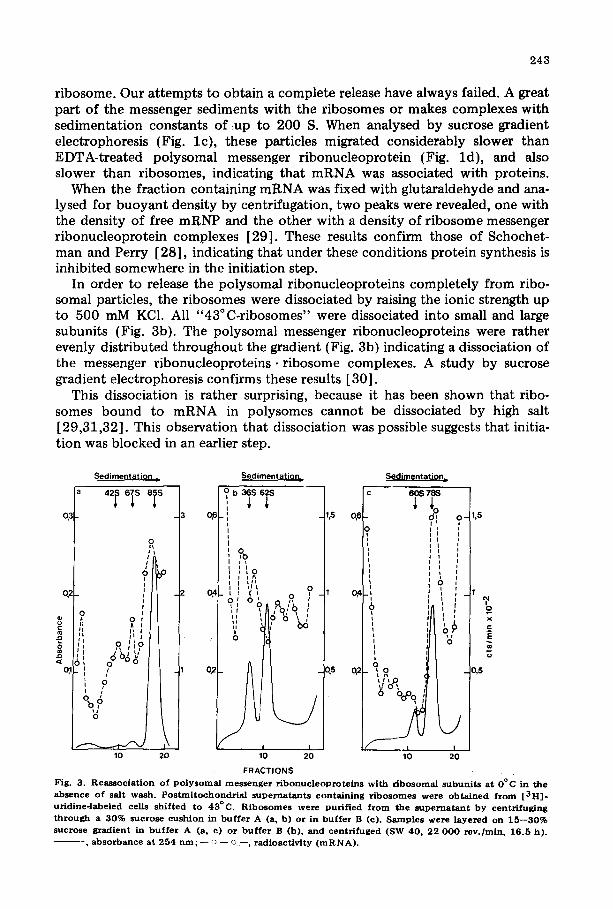
As shown by McCormick and Penman [27], cells shifted to a relatively high temperature (42°C) sto protein synthesis. Results shown in Fig. 2 confirm these observations. Schochetman and Perry [28] have shown that part of the messenger RNA is released as mRNP, but that another part is still bound to the
242
migration migration
1,5 , \ 4 J ,
0,5 f ,'* 2 /" ' '~
? i L , 3 "o 1 x
o3 ,, ! 2
z 01 z
- - ~ ^ I
0 b EOTA - 6 ~ O d / , EDTl
<m o,e ,, ,
09 / ', 09 2
/ '_ 2 0,2 02 1 ,~
30 40 30 40 FRACTIONS FRACTIONS
Fig. 1. Sucrose g rad ien t e l ec t rophores i s of E D T A - t r e a t e d r i b o so mes P o l y s o m e s and m R N P s . Pos tmi to - chondr ia l s u p e r n a t a n t s con t a in ing r i bos ome s and p o l y s o m e s were o b t a i n e d f rom cul tures labeled wi th [ 3 H ] u r i d i n e in the p resence of low dose a c t i n o m y c i n D, e i the r only at 37°C (a, b, d) , or wi th a subse- q u e n t t e m p e r a t u r e shif t to 43°C (c). The par t ic les , f r o m cell cu l tu res only a t 37°C, were pur i f ied by c e n t r i f u g a t i o n in 15 - -45% sucrose g rad ien t in b u f f e r A. Th e f rac t ions con ta in ing r i bosomes (5 0 - -9 0 S) (a, b) or p o l y s o m e s ( 1 0 0 - - 5 0 0 S) (d) we re p o o l e d and cen t r i fuged d o w n (Ti50 , 53 0 0 0 r ev . /min , 17 h). T h e y were r e suspended in b u f f e r C (a) or b u f f e r C con ta in ing 20 m M E D T A (b, d). Th e par t ic les f rom t e m p e r a t u r e - s h o c k e d cells (c) were pur i f i ed by een t r i fuga t i on in 15 - -30% sucrose grad ien t in b u f f e r A. The par t ic les s e d i m e n t i n g b e t w e e n 3 0 - - 1 0 0 S were cen t r i fuged d o w n as desc r ibed above and the pe l le t was r e s uspended in b u f f e r C. The sample were t h e n ana lysed by sucrose g rad ien t e l ec t rophores i s (200 V, 17 h, 12°C) . - - , ab so rbanee at 260 n m ( r ibosomes a, b, c or p o l y s o m e s d); . . . . . . , r ad ioac t iv i ty of m R N A ( i n f o r m o s o m e s a0 b or p o l y s o m a l m R N P c, d).
[a
o,z
o.-'
o,2,'
i 80S sediment atJ~n
i 1~) 20
FRACTIONS
80S £
,p 'L '
J
sedimentat~n
b
1C) 20
x ¢ .E
E
5
Fig. 2. L ibe ra t i on of p o l y s o m a l m e s s e n g e r r i b o n u c l e o p r o t e i n s f r o m p o l y s o m e s in cu l tu red cells sh i f ted to 43°C. Pos tmi t i chondr i a l s u p e r n a t a n t s con ta in ing r i bosomes and p o l y s o m e s were ob t a ined f r o m a [ 3 H ] - uridineolabeled cu l tu re g rown only a t 37°C (a) and f rom a n o t h e r cu l tu re sh i f ted to 43°C (b). The super- na t an t s were ana lysed by cen t r i fuga t i on in 15 - -30% sucrose grad ien t in b u f f e r A ( ro to r SW 40, 17 0 0 0 r ev . /min , 17 h, 4°C) . - , ab so rbance at 254 run ; - - - o - - o - - , r ad ioac t iv i ty ( m R N A ) .
243
ribosome. Our at tempts to obtain a complete release have always failed. A great part of the messenger sediments with the ribosomes or makes complexes with sedimentation constants of ~up to 200 S. When analysed by sucrose gradient electrophoresis (Fig. lc) , these particles migrated considerably slower than EDTA-treated polysomal messenger r ibonucleoprotein (Fig. ld ) , and also slower than ribosomes, indicating that mRNA was associated with proteins.
When the fraction containing mRNA was fixed with glutaraldehyde and ana- lysed for buoyant density by centrifugation, two peaks were revealed, one with the density of free mRNP and the other with a density of r ibosome messenger r ibonucleoprotein complexes [29]. These results confirm those of Schochet- man and Perry [28], indicating that under these conditions protein synthesis is inhibited somewhere in the initiation step.
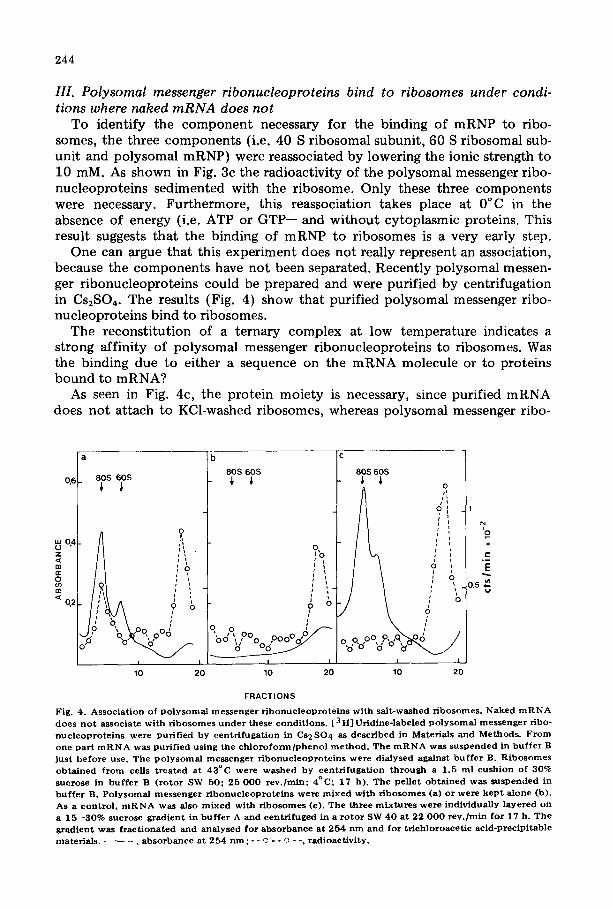
In order to release the polysomal ribonucleoproteins completely from ribo- somal particles, the ribosomes were dissociated by raising the ionic strength up to 500 mM KCI. All "43°C-ribosomes '' were dissociated into small and large subunits (Fig. 3b). The polysomal messenger ribonucleoproteins were rather evenly distributed throughout the gradient (Fig. 3b) indicating a dissociation of the messenger r ibonucleoproteins , r ibosome complexes. A study by sucrose gradient electrophoresis confirms these results [30].
This dissociation is rather surprising, because it has been shown that ribo- somes bound to mRNA in polysomes cannot be dissociated by high salt [29,31,32]. This observation that dissociation was possible suggests that initia- tion was blocked in an earlier step.
q~
o
o,1
Sedimentation L Sedimentation_
?
I I
11 I
0 l l O ," I , / oo ~o ~
/ o
U
10 20
? I
b 36S 62S
I ~ o I I I I 1 111
I I I I | t'~ T
,I ?
I I 10 20
FRACTIONS
SedimentationL
c 6OS 78S
,s 0,~ ? 1,s I
I ,,
~ s
6oOo iI
I I 10 20
Fig . 3 . R e a s s o c i a t i o n o f P o l y s o m a l m e s s e n g e r r i b o n u c l e o p r o t e i n s w i t h r i b o s o m a l s u b u n i t s a t 0 ° C in the absence o f s a l t w a s h . P o s t m i t o c h o n d r i a l s u p e r n a t a n t s c o n t a i n i n g r i b o s o m e s w e r e obta ined f r o m [ 3 H ] - u r i d i n e - l a b e l e d cel ls sh i f t ed t o 4 3 ° C . R i b o s o m e s were puri f ied f rom the supernatant by c e n t r i f u g i n g t h r o u g h a 3 0 % sucrose cush ion i n b u f f e r A (a , b ) o r i n b u f f e r B (c ) . Sa mples w e r e layered on 15- - -30% sucrose gradient in b u f f e r A (a , c ) o r b u f f e r B ( b ) , and centr i fuged ( S W 4 0 , 2 2 0 0 0 rev . /min , 1 6 . 5 h ) .
• a b s o r b a n c e a t 2 5 4 n m ; - - o - - o - - , r a d i o a c t i v i t y ( m R N A ) .
244
III. Polysomal messenger ribonucleoproteins bind to ribosomes under condi- tions where naked m R N A does not
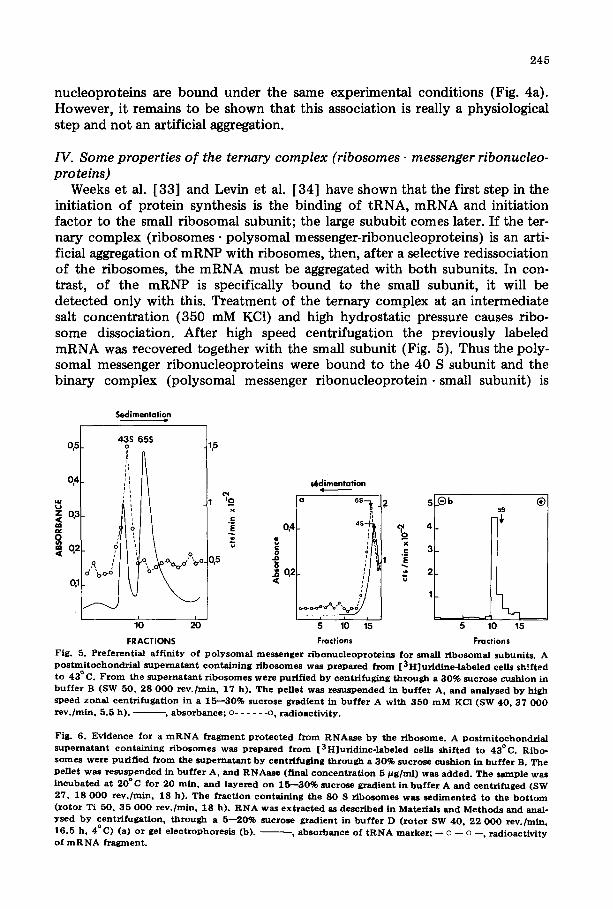
To identify the componen t necessary for the binding of mRNP to ribo- somes, the three components (i.e. 40 S ribosomal subunit, 60 S ribosomal sub- unit and polysomal mRNP) were reassociated by lowering the ionic strength to 10 mM. As shown in Fig. 3c the radioactivity of the polysomal messenger ribo- nucleoproteins sedimented with the ribosome. Only these three components were necessary. Furthermore, this reassociation takes place at 0°C in the absence of energy (i.e. ATP or GTP-- and wi thout cytoplasmic proteins. This result suggests that the binding of mRNP to ribosomes is a very early step.
One can argue that this experiment does not really represent an association, because the components have not been separated. Recent ly polysomal messen- ger r ibonucleoproteins could be prepared and were purified by centrifugation in Cs2SO4. The results (Fig. 4) show that purified polysomal messenger ribo- nucleoproteins bind to ribosomes.
The reconsti tut ion of a ternary complex at low temperature indicates a strong affinity of polysomal messenger r ibonucleoproteins to ribosomes. Was the binding due to either a sequence on the mRNA molecule or to proteins bound to mRNA?
As seen in Fig. 4c, the protein moiety is necessary, since purified mRNA does not attach to KCl-washed ribosomes, whereas polysomal messenger ribo-
o,6ta 80s~, 60S~,
A 04 ",
z ,,
<~ 0,2 ' " 0
oOo )P "o"
i
10 20
_ r _
80S 60S 80S 60S i i i I
) i
°,o : ,
i i ~ 0
o Io ooo o' ~b o'°',,,,°°o ,Ooo o, /
I I I I
10 20 10 20
1
% ¢-
0,5
FRACTIONS F i g . 4 . A s s o c i a t i o n o f P o l y s o m a l m e s s e n g e r r i b o n u e l e o p r o t e i n s w i t h s a l t - w a s h e d r i b o s o m e s . N a k e d m R N A d o e s n o t a s s o c i a t e w i t h r i b o s o m e s u n d e r t h e s e c o n d i t i o n s . [ 3 H ] U r i d i n e - l a b e l e d p o l y s o m a l m e s s e n g e r r i b o -
n u c l e o p r o t e i n s w e r e p u r i f i e d b y c e n t r i f u g a t i o n in C s 2 S O 4 as d e s c r i b e d in M a t e r i a l s a n d M e t h o d s . F r o m o n e p a r t m R N A w a s p u r i f i e d u s i n g t h e c h l o r o f o r m / p h e n o l m e t h o d . T h e m R N A w a s s u s p e n d e d in b u f f e r B j u s t b e f o r e u s e . T h e p o l y s o m a l m e s s e n g e r r i b o n u c l e o p r o t e i n s w e r e d i a l y s e d a g a i n s t b u f f e r B. R i b o s o m e s o b t a i n e d f r o m ce l l s t r e a t e d a t 4 3 ° C w e r e w a s h e d b y c e n t r i f u g a t i o n t h r o u g h a 1 . 5 m l c u s h i o n o f 3 0 % s u c r o s e i n b u f f e r B ( r o t o r S W 5 0 ; 2 5 0 0 0 r e v . / m i n ; 4 ° C ; 17 h ) . T h e p e l l e t o b t a i n e d w a s s u s p e n d e d i n
b u f f e r B. P o l y s o m a l m e s s e n g e r r i b o n u c l e o p r o t e i n s w e r e m i x e d w i t h r i b o s o m e s (a ) o r w e r e k e p t a l o n e (b ) . A s a c o n t r o l , m R N A w a s a l so m i x e d w i t h r i b o s o m e s (c ) . T h e t h r e e m i x t u r e s w e r e i n d i v i d u a l l y l a y e r e d o n a 1 5 - - 3 0 % s u c r o s e g r a d i e n t i n b u f f e r A a n d c e n t r i f u g e d i n a r o t o r SW 4 0 a t 2 2 0 0 0 r e v . / m i n f o r 17 h. T h e g r a d i e n t w a s f r a c t i o n a t e d a n d a n a l y s e d f o r a b s o r b a n c e a t 2 5 4 n m a n d f o r t r i c h l o r o a c e t i c a c i d - p r e c i p i t a b l e
m a t e r i a l s . , a b s o r b a n c e a t 2 5 4 n m ; - - o - - o - - , r a d i o a c t i v i t y .
245
nucleoproteins are bound under the same experimental conditions (Fig. 4a). However, it remains to be shown that this association is really a physiological step and not an artificial aggregation.
IV. Some properties of the ternary complex (ribosomes. messenger ribonucleo- proteins)
Weeks et al. [33] and Levin et al. [34] have shown that the first step in the initiation of protein synthesis is the binding of tRNA, mRNA and initiation factor to the small ribosomal subunit; the large sububit comes later. If the ter- nary complex (ribosomes - polysomal messenger-ribonucleoproteins) is an arti- ficial aggregation of mRNP with ribosomes, then, after a selective redissociation of the ribosomes, the mRNA must be aggregated with both subunits. In con- trast, of the mRNP is specifically bound to the small subunit, it will be detected only with this. Treatment of the ternary complex at an intermediate salt concentration (350 mM KC1) and high hydrostatic pressure causes ribo- some dissociation. After high speed centrifugation the previously labeled mRNA was recovered together with the small subunit (Fig. 5). Thus the poly- somal messenger ribonucleoproteins were bound to the 40 S subunit and the binary complex (polysomal messenger r ibonucleoprote in , small subunit) is
I,
, I I
; ! , o
i
10 2C)
Sedimentation
o,s ~s 6ss tl,5 /
o4 i, I x
i ),5
o -e c~
,<
s6dimentation
ss-~
4s j,i
J o.0.~% ,°,
5 1 15 FRACTIONS Fractions
2
._c
.eb ® 5s
5 10 15 Fractions
Fig. 5. Preferen t ia l a f f in i ty of p o l y s o m a l me s s e nge r r i bonuc leop ro t e in s for smal l r i bosoma l subuni ts . A p o s t m i t o c h o n d r i a l s u p e r n a t a n t con t a in ing r i bosomes was p r e p a r e d f r o m [3H]u r id ine - l abe l ed cells sh i f ted to 43 ° C. F r o m the s u p e r n a t a n t r i b o s o m e s were pur i f i ed by cen t r i fug ing t h r o u g h a 30% sucrose cush ion in b u f f e r B (SW 50, 28 000 rev. /min0 17 h). The pe l le t was r e suspended in b u f f e r A, an d ana lysed b y high speed zona l cen t r i fuga t ion in a 15 - -30% sucrose g rad ien t in b u f f e r A wi th 350 m M KCI (SW 40° 37 0 0 0 r ev . /m in , 5.5 h). - - - , abso rbanee ; o . . . . . . o, r ad ioac t iv i ty .
Fig. 6. Evidence for a m R N A f r a g m e n t p r o t e c t e d f rom RNAase by the r ibosome . A p o s t m i t o c h o n d r i a l s u p e r n a t a n t contaJ_ning r i bos ome s was p r e p a r e d f r o m [3H]u r id ine - l abe l ed cells sh i f ted to 43°C. Ribo- somes were pur i f ied f r o m the s u p e r n a t a n t by cen t r i fug ing t h ro u g h a 30% sucrose cush ion in b u f f e r B. The pe l le t was r e suspended in b u f f e r A, and RNAase (final c o n c e n t r a t i o n 5 # g / m l ) was added . The sample was i n c u b a t e d a t 20°C for 20 min , and l aye red on 15---30% sucrose g rad ien t in b u f f e r A an d cen t r i fuged (SW 27, 18 0 0 0 r ev . /min , 18 h). The f rac t ion con ta in ing the 80 S r i b o so mes was s e d i m e n t e d to the b o t t o m ( r o t o r Ti 50, 35 000 r ev . /min , 18 h) . R N A was e x t r a c t e d as descr ibed in Materials and Methods and anal- ysed b y cen t r i fuga t ion , t h rough a 5- -20% sucrose grad ien t in b u f f e r D ( r o t o r SW 40, 22 0 0 0 rev . /min , 16 .5 h, 4°C) (a) or gel e lec t rophores i s (b). - - - , abso rbance of t R N A m a r k e r ; - o - - o - - r ad ioac t iv i ty of m R N A f r agmen t .

246
more stable than the r ibosome against higher concentrated saline at high hydro- static pressure.
If the polysomal messenger r ibonucleoproteins are correctly positioned in the complex, they are located between the two ribosomal subunits. In this case a port ion of the mRNA must be shielded by the r ibosome and escapes a mild RNAase attack. Such protect ion of the mRNA by ribosomes has been found in R17 initiated in vitro [35].
The ternary complex (i.e. polysomal messenger ribonucleoproteins, 40 S and 60 S ribosomal subunits) was digested with a low dose of RNAase (5 pg/ml). This procedure, while removing the bulk of polysomal messenger ribnucleopro- teins, leaves part of the mRNA with the ribosome. The RNA was extracted from the r ibosome (which was separated by sucrose gradient centrifugation), and analysed by sucrose gradient centrifugation and gel electrophoresis. A "mRNA-fragment" of about 6 S (Figure 6) was found. This "l~NA-fragment" found is obviously neither tRNA nor 5 S RNA. If one separates the two sub- units (as described in Fig. 5), the radioactive fragment sediments with the small subunit.
The RNA-fragment is calculated to be 120--150 nucleotides long. This is longer than the length generally found for the mRNA protected by ribosomes in polysomes (40--80 nucleotides) [35]. This comparison suggests that another factor other than the r ibosome alone has participated in the protect ion of the mRNA from the action of RNAase.
V. Analysis of the mRNA fragment Jeffery and Brawerman [36] have suggested that the poly(A) sequence at the
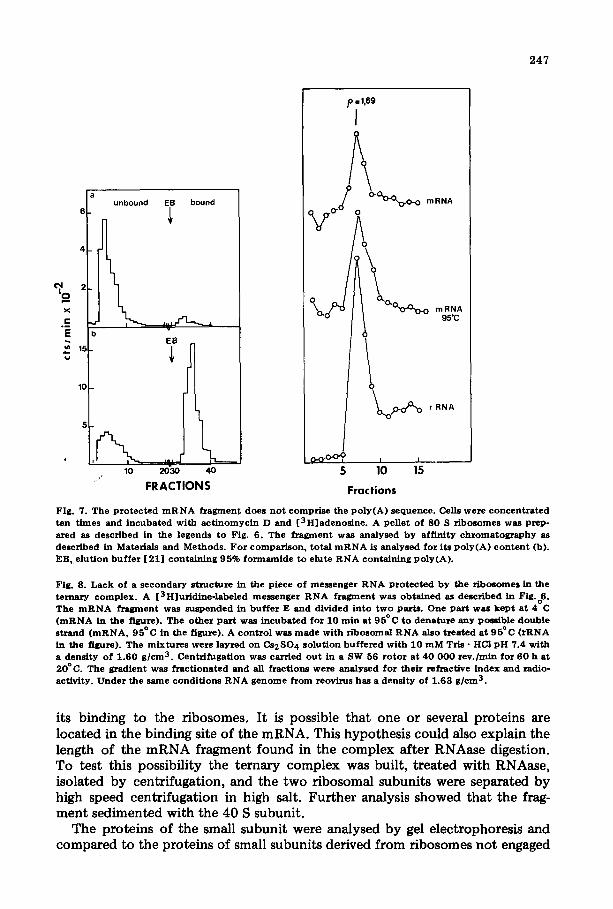
3' end of the messenger is folded back and associated with another part of the messenger. Similar results have been obtained by Bester et al. [37]. These authors suggest that the poly(A) sequence is involved in the initiation of pro- tein synthesis. In our experiments, the presence of a poly(A) sequence {resis- tant to RNAase) could explain the length of the RNAase-protected mRNA frgment reported above. To investigate this hypothesis the m R N A fragment was passed through a poly(U)-Sepharose column as described by Lindberg and Person [21]. Under these conditions 70% of the total mRNA was retained but the mRNA fragment, labeled with [3H]adenosine, passed through (Fig. 7 ) a n d thus does not contain poly(A} sequences. Furthermore, the chloroform/phe- nol-extracted RNA piece was totally degraded by pancreatic RNAase at 150 mM NaC1. In this condit ion poly(A) is not attacked. Consequently poly(A) is not present in this complex.
Another possibility explaining the length of the mRNA piece not at tacked be the RNAase is the presence of double-stranded RNA. Such double-stranded regions have been found in mRNA [38--40]. The fragment was analysed by Cs2804 centrifugation before and after thermal denaturation. The results pres- ented in Fig. 8 show that no double-stranded structure can be detected in the region involved in the binding of mRNA to the ribosome.
VI. One of the major proteins found in the polysomal messenger ribonucleo- proteins is also found in the ribosome, messenger ribonucleoprotein complex after mild RNAase digestion
As shown above, the proteins associated with the mRNA are necessary for
247
2
x
10
5
unbound EB bound
10 2030 40 j ' FRACTIONS
jo •1,69
I
(3-
5 1~0 15 Fractions
Fig . 7. T h e p r o t e c t e d m R N A f r a g m e n t d o e s n o t c o m p r i s e t he p o l y ( A ) s e q u e n c e . Cells w e r e c o n c e n t r a t e d t e n t imes a n d i n c u b a t e d w i t h a c t i n o m y c i n D a n d [ 3 H ] a d e n o s i n e . A pe l l e t o f 8 0 S r i b o s o m e s w a s p r e p - s.red as d e s c r i b e d in t he l egends t o Fig. 6 . The f r a g m e n t w a s a n a l y s e d b y a f f i n i t y c h r o m a t o g r a p h y as d e s c r i b e d in Mate r ia l s a n d M e t h o d s . F o r c o m p a r i s o n , t o t a l m R N A is a n a l y s e d f o r i t s p o l y ( A ) c o n t e n t (b) . EB0 e l u t i o n b u f f e r [ 2 1 ] c o n t a i n i n g 9 5 % f o r m a m i d e t o e lu te R N A c o n t a i n i n g p o l y ( A ) .
Fig . 8. L a c k o f a s e c o n d a r y s t r u c t u r e in t he p i ece o f m e s s e n g e r R N A P r o t e c t e d b y the r i b o s o m e s in t h e t e r n a r y c o m p l e x . A [ 3 H ] u r i d i n e - l a b e l e d m e s s e n g e r R N A f r a g m e n t w a s o b t a i n e d as d e s c r i b e d in Fig.o6. T h e m R N A f r a g m e n t w a s s u s p e n d e d in b u f f e r E a n d d iv ided i n t o t w o pa r t s . O n e p a r t waS k e p t a t 4 C ( m R N A in t h e f igure) . The o t h e r p a r t w a s i n c u b a t e d f o r 1 0 r a i n a t 9 5 ° C t o d e n a t u r e a n y poss ib l e d o u b l e s t r a n d ( m R N A , 9 5 ° C in the f igure) . A c o n t r o l w a s m a d e w i t h r i b o s o m a l R N A a lso t r e a t e d a t 9 5 ° C ( r R N A in the f igure) . T h e m i x t u r e s we re l a y r e d o n C s 2 S O 4 s o l u t i o n b u f f e r e d w i t h 1 0 m M Tris • HCI p H 7 .4 w i t h a d e n s i t y o f 1 . 6 0 g / c m 3. C e n t r i f u g a t i o n was c a r r i e d o u t in a SW 56 r o t o r a t 4 0 0 0 0 r e v . / m i n f o r 6 0 h a t 2 0 ° C . The g r a d i e n t was f r a e t i o n a t e d a n d all f r a c t i o n s w e r e a n a l y s e d f o r t h e i r r e f r ac t i ve i n d e x a n d r ad io - ac t i v i t y . U n d e r t h e s a m e c o n d i t i o n s R N A g e n o m e f r o m reov i rus h a s a d e n s i t y o f 1 .63 g / e ra 3.
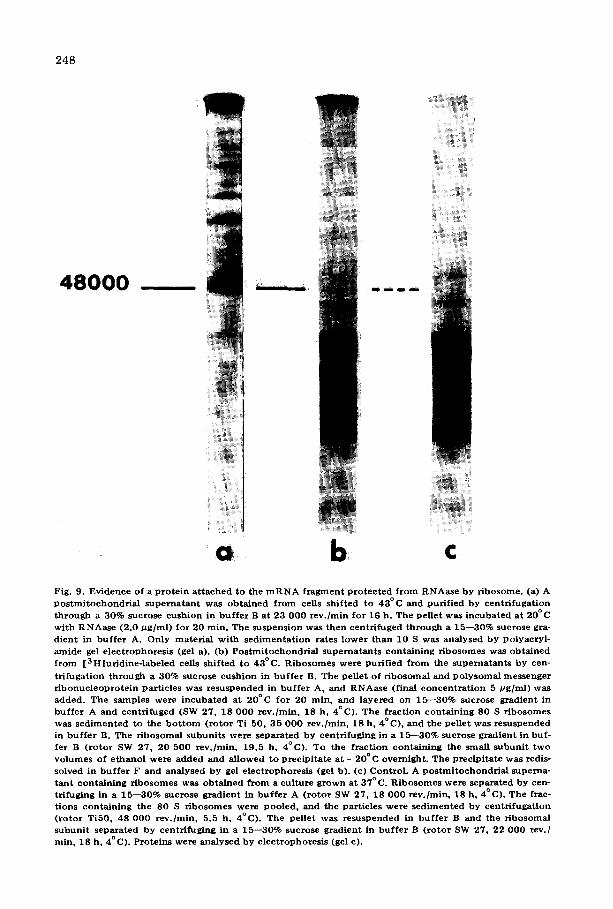
its binding to the ribosomes. It is possible that one or several proteins are located in the binding site of the mRNA. This hypothesis could also explain the length of the mRNA fragment found in the complex after RNAase digestion. To test this possibility the ternary complex was built, treated with RNAase, isolated by centrifugation, and the two ribosomal subunits were separated by high speed centrifugation in high salt. Further analysis showed that the frag- ment sedimented with the 40 S subunit.
The proteins of the small subunit were analysed by gel electrophoresis and compared to the proteins of small subunits derived from ribosomes not engaged
248
4 8 0 0 0
b Fig. 9. Ev idence of a p r o t e i n a t t a c h e d to the m R N A f r a g m e n t p r o t e c t e d f r o m R N A a s e by r ibosome . (a) A p o s t m i t o c h o n d r i a i s u p e r n a t a n t was o b t a i n e d f r o m cells sh i f ted to 4 3 ° C and pur i f i ed by cen t r i fuga t i on t h r o u g h a 30% sucrose cush ion in b u f f e r B a t 23 000 r e v . / m i n fo r 16 h. The pe l le t was i n c u b a t e d a t 20°C wi th R N A a s e (2 .0 # g / m l ) for 20 rain. The suspens ion was t h e n cen t r i fuged t h ro u g h a 15- -30% sucrose gra- d ien t in b u f f e r A. Only m a t e r i a l w i th s e d i m e n t a t i o n ra tes l o w e r t han 10 S was ana lysed by po lyac ry l - amide gel e l ec t rophores i s (gel a). (b) P o s t m i t o c h o n d r i a l s u p e r n a t a n t s con ta in ing r i b o so mes was ob t a ined f r o m [ 3 H ] u r i d i n e - l a b e l e d cells sh i f ted to 43°C. R i b o s o m e s were pur i f i ed f r o m the s u p e r n a t a n t s by cen-
t r i fuga t ion t h r o u g h a 30% sucrose cush ion in b u f f e r B. The pe l le t of r i b o s o m a l and p o l y s o m a i mes sen g e r r i b o n u c l e o p r o t e i n par t ic les was r e s us pe nde d in b u f f e r A, and RNAase (final c o n c e n t r a t i o n 5 /~g/ml) was added . T he samples were i n c u b a t e d a t 20°C for 20 rain, and l ay e r ed on 15 - -30% sucrose g rad ien t in b u f f e r A and cen t r i fuged (SW 27, 18 0 0 0 r ev . /min , 18 h, 4°C) . The f rac t ion con ta in ing 80 S r i bosomes was s e d i m e n t e d to the b o t t o m ( r o t o r Ti 50, 35 0 0 0 r ev . /min , 18 h, 4°C) , and the pe l le t was r e suspended in b u f f e r B. The r i bosoma l subuni t s we re s epa ra t ed b y cen t r i fug ing in a 15---30% sucrose g rad ien t in buf- fer B ( ro to r SW 27, 20 500 r e v . /m in , 19 .5 h, 4°C) . To the f r ac t ion con t a in ing the smal l subun i t t w o v o l u m e s of e t hano l were a dde d and a l lowed to p rec ip i t a t e a t - - 2 0 ° C overn ight . Th e p rec ip i t a t e was redis- solved in b u f f e r F and ana lysed b y gel e l ec t rophores i s (gel b). (c) Contro l . A p o s t m i t o c h o n d r i a l superna- t a n t con t a in ing r i bosom e s was o b t a i n e d f r o m a cu l tu re g rown a t 37°C. R i b o s o m e s were sepa ra ted by cen- t r i fuging in a 15 - -30% sucrose g rad ien t in b u f f e r A ( r o t o r SW 27, 18 0 0 0 r ev . /min , 18 h, 4°C) . The frac- t ions con ta in ing the 80 S r i bos ome s were poo led , and the par t ic les were s e d i m e n t e d by cen t r i fuga t i on ( r o t o r TiS0, 48 000 r ev . /min , 5.5 h, 4°C) . The pe l le t was resusPended in b u f f e r B and the r i bosoma l subun i t s epa ra t ed b y cen t r i fug ing in a 15 - -30% sucrose g rad ien t in b u f f e r B ( r o t o r SW 27, 22 0 0 0 rev. / rain, 18 h, 4°C) . Prote ins were ana lysed by e lec t rophores i s (gel c).

249
in protein synthesis. The small subunit containing the mRNA fragment has one additional protein with a molecular weight of about 48 000 (Fig. 9). This pro- tein corresponds to one of the major proteins found in the polysomal messen- ger ribonucleoproteins {Fig. 9). Therefore, it seems justified to conclude that a least one protein of the polysomal messenger ribonucleoproteins was present in the mRNA • ribosome complex formed in vitro. But if one of the minor pro- teins of the polysomal messenger ribonucleoprotein was also present it was not detectable due to its low concentration. To obtain a better resolution it was attempted to separate the fragment from the ribosomal subunits, but without success. The mRNA fragment always sedimented with the 40 S RNA even after high salt treatment (750 mM KC1), after deoxycholate treatment (known to remove proteins from mRNPs [41]) and after aurin tricarboxylic acid treat- ment (which inhibits mRNP binding [42]). EDTA irreversibly separates the mRNP fragment, but the ribosomes (previously treated by RNAase) were dis- located and ribosomal fragments sedimented in all the gradient. Under this con- dition it was impossible to analyse the protein associated with the mRNA frag- ment. Therefore, the study of minor proteins is still without definitive results.
VII. The ternary complex (ribosomes'messenger ribonucleoproteins) needs initiation factors to assume protein synthesis
The ability of the complex to support in vitro protein synthesis was tested. As shown in Table Ic, the complex was active only if one provided proteins washed from ribosomes by high salt (e.g. initiation factors), and if the trans- lation factors and tRNAs were provided by the addition of cytosol. This result shows that the complex was already an early step of initiation. Furthermore, aurin tricarboxylic acid did not inhibit the protein synthesis in this experiment, implying that mRNA was previously engaged in the right position; since aurin tricarboxylic acid inhibits the binding of mRNP to the ribosome [58].
T A B L E I
I N F L U E N C E O F T H E R I B O S O M A L W A S H O N C E L L - F R E E P O L Y P E P T I D E S Y N T H E S I S B Y T H E
R E A S S O C I A T E D P O L Y S O M A L M E S S E N G E R R I B O N U C L E O P R O T E I N S . R I B O S O M E C O M P L E X
P o s t m i t o c h o n d r i a l s u p e m a t a n t s c o n t a i n i n g 8 0 S r i b o s o m e s a n d p o l y s o m e s w e r e o b t a i n e d f r o m a c u l t u r e grown a t 3 7 ° C (a ) a n d f r o m another temperature-shocked c u l t u r e (b , c) . Fract ions containing p a r t i c l e s a b o v e 3 0 S w e r e pel leted b y centri fugatton through a 3 0 % s u c r o s e c u s h i o n i n b u f f e r A. O n e p a r t (a , b )
w a s r e s u s p e n d e d i n b u f f e r G and the o ther P a s t ( c ) in buf fer B. The l a t t e r s a m p l e w a s t h e n c e n t r i f u g e d t h r o u g h a 3 0 % sucrose cush ion in b u f f e r B; t h e p e l l e t w a s r e s u s p e n d e d i n b u f f e r G , t h e s u p e r n a t a n t over the sucrose cush ion w a s d i a l y s e d a g a i n s t b u f f e r G a n d u s e d as r i b o s o m a l w a s h . The prote in synthes is w a s c a r r i e d o u t as d e s c r i b e d i n M a t e r i a l s a n d M e t h o d s .
R i b o s o m e s S a l t w a s h A u r i n t r i c s x b o x y l i c c p m / A 2 6 0 r i b o s o m e s a d d e d a c i d a d d e d
a) N o r m a l ( 3 7 ° C ) - - - - 1 0 1 0
- - + 9 8 0
b ) S h o c k e d ce l l s ( 4 3 ° C ) - - - - 1 0 8 0
- - + 8 7 0 + - - 1 1 2 9
c) S h o c k e d ce l l s ( 4 3 ° C ) w a s h e d - - - - 8 0
w i t h 0 . 5 M K C I + - - 4 3 3
+ + 4 2 8

250
Discussion
Whether messenger RNA joins the small ribosomal subunits as free RNA molecules or as mRNPs has been an intactable problem. When analysing for the capacity of eukaryotic cells to stimulate protein synthesis, investigators have not been able to detect a difference between the particles (polysomal messen- ger ribonucleoprotein) and mRNAs [9--12] with regard to their activity in a cell-free translation system. The following points try to explain these results:
(a) As shown above, EDTA treatment appears to modify polysomal messen- ger ribonucleoproteins, at least in HeLa cells; one must not expect from such particles a better stimulation than from free mRNAs in the cell-free system.
(b) The proteins involved in the binding of mRNA to the ribosomes are attached to the mRNA, but it is possible that a reserve of these proteins is pres- ent in the cytosol. Recently we have shown that some free proteins with high affinity to RNAs share properties with proteins bound to the mRNA [43]. Consequently, the argument whether the proteins associated with the mRNA are essential for polypeptide synthesis, may simply overridden whenever an $30, S100 of pH 5 enzyme is added to the cell-free system. Cashion and Stan- ley [44] leave no doubt that genuine proteins in the polysomal messenger ribo- nucleoproteins are responsible for the association to the 40 S subunit. Since in their s tudy the puromycin-induced release of the methionine as methionylpuro- mycin did not require cytoplasmic supernatant, no other putative cytoplasmic mRNA binding factor was present. Furthermore, when the reconsti tution of an initiation complex is studied, polysomal messenger ribonucleoproteins are essential. Lebleu et al. [5] have reported that free mRNA does not bind "a t all", but that the protein moiety is necessary for the binding to ribosomes washed by deoxycholate.
Together with the results reported in this paper one can suggest that at least one of the proteins bound to the active mRNA is involved in the initiation process. The protein with a molecular weight of 48 000 is probably involved in this early step of the initiation.
The question now remains whether any of the proteins in the polysomal messenger ribonucleoproteins (especially the protein) 48 000-dalton are identi- cal with one of the hitherto-described initiation factors (see Hellerman and Shafritz [45]). In agreement with Noll and Noll [46] we consider initiation fac- tors as proteins not permanently bound to the ribosome. They are engaged in the initiation process and are detached during elongation. These factors have first been isolated from the KC1 washes; they exist also in the cytosol [47--49], and on native subunits [11]. The proteins considered here are tightly bound to the mRNA (they are not washed off by 0.5 M KC1, or by centrifugation in Cs2SO4) and do not appear in the ribosomal washes. They are rather a perma- nent companion of the mRNA during the translation. Therefore, they should not be classified as true initiation factors; but it is possible that these proteins are identical to a modified form of the initiation factor found to be necessary to bind mRNA (IF4 and IF6) [50].
mRNPs also have been found free in the cytoplasm [2,4]. This free mRNP is regarded to be unactive in protein synthesis [2,52] and differs in its pro- tein composition from the polysomal messenger ribonucleoproteins [14,54]. It

251
was recently shown that the protein with molecular weight of 48 000 is not found in the free mRNP [14]. Thus, it is possible that this protein enhances the activity of mRNA. Wigle [49] has found in the cytoplasm and "initiation factor" with a molecular weight close to this protein (i.e. 50 000). This pro- tein is mRNA specific. Perhaps a modification as phosphorylation of the pro- tein is necessary for its function. It has been demonstrated that the major protein found in polysomal messemger ribonucleoproteins (Mr 50 000) is phos- phorylated [54,55]. This can explain that the cell free systems using naked mRNA need ATP [56,57].
Acknowledgements
I would like to express my appreciation to Professor K. KShler for his con- stant interest in this work, and for many useful suggestion and criticism. I thank Drs. O.-G. Issinger and J. Kreft for critical reading of the manuscript. The help of Mr. and Mrs. B. Smith in translating the manuscript is gratefully acknowledged. This work has been supported by the Deutsche Forschungsge- meinschaft.
References
1 Perry, R.P. and Kelley, D.E. (1968) J. Mol. Biol. 35, 37--59 2 Spirin, A.S. (1969) Eur. J. Biochem. 10, 20--35 3 Henshaw, E.C. (1968) J. Mol. Biol. 36, 401--417 4 Spohr, G., Granboulan, N., Morel, C. and Scherrer, K. (1970) Eur. J. Biochem. 17, 296--318 5 Lebleu, B., Marbaix, G., Huez, G., Temmerman, J., Burny, A. and Chantrenne, H. (1971) Eur. J.
Biochcm. 19 ,264- -269 6 Morel, C., Gander, E., Herzberg, M., Dubochet, J. and Scherrer, K. (1973) Eur. J. Biochem. 36,
455---464 7 Blobel, G. (1972) Biochem. Biophys. Res. Commun. 47, 88--95 8 Williamson, R. (1973) FEBS Lett. 37, 1--6 9 Sampson, J., Mathews, M., Osborn, M. and Borghetti, A. (1972) Biochemistry 11, 3636--3640
10 Ernst, V. and Arnstein, H. (1975) Bioehim. Biophys. Acta 378, 251--259 11 Freienstein, C. and Blobel, G. (1974) Proc. Natl. Acad. Sci. U.S. 71, 3435--3439 12 Chen. S.H., Layers, G. and Spector, A. (1976) Biochim. Biophys. Acta 418, 39--51 13 Bryan, R. and Hayashi, M. (1973) Nature New Biol. 244, 271--274 14 Liautard, J.-P., Setyono, B., Spindler, E. and Kohler, K. (1976) Biochim. Biophys. Aeta 425, 373--
383 15 Gesteland, R. (1968) J. Mol. Biol. 18, 356--371 16 Martin, T.E., Rolleston, F.S., Low, R.B. and Wool, I.G. (1969) J. Mol. Biol. 43 ,135 - -147 17 Lissitzky, S., Poiree, J.-C., Cartouzou, G. and Gregoire, J. (1970) Eur. J. Biochem. 12, 104--112 18 Perry, R.P., LaTorre, J., Kelley, D.E. and Grecnberg, J.E. (1972) Biochim. Biophys. Acta 262,
220--226 19 Tiollals, P., Galibert, F., Lepetit , A. and Auger, M. (1972) Biochimie 54, 339--354 20 Adesnik, M., Salditt, M., Thomas, W. and Darnell, J.E. (1972) J. Mol. Biol. 71, 21--30 21 Lindberg, U. and Pergson, T. (1972) Eur. J. Bioehem. 31, 246--252 22 Mans, R.J. and Novelli, G.D. (1961) Arch. Biochem. Biophys. 94, 48--54 23 Liautard, J.-P. and Kohlcr, K. (1974) Anal. Biochem. 60, 581--588 24 MeEven, C.R. (1967) Anal. Biochem. 20, 114--148 25 Weber, K., Pringle, J.R. and Osborn, M. (1972) in Methods in Enzymology (Colowiek, S.P. and
Kaplan, N.O., eds.), Vol. XXVI, pp. 3--27, Academic Press, New York 26 Kohler, K., Lecou, C. and Arends, S. (1972) Rev. Eur. Clin. Biol. 17, 44--53 27 McCormick, W. and Penman, S. (1969) J. Mol. Biol. 39, 315--323 28 Sehochetman, G. and Perry, R.P. (1972) J. Mol. Biol. 63, 577--586 29 Liautard, J.-P. (1976) Ph.D. Thesis, University of Strasbourg 30 Liautard, J.-P. and Kohler, K. (1976) Biochemie 85, 317--323 31 Falvey, A.K. and Stahelin, T. (1970) J. Mol. Biol. 53, 1--19

252
32 Kumar, A. and Lindberg, U. (1972) Proc. Natl. Acad. Sci. U.S. 69, 681--684 33 Weeks, D.P., Verma, D., Seal, S. and Marcus, A. (1972) Nature 236, 167--168 34 Levin, D., Kyner, D. and Acs, G. (1973) J. Biol. Chem. 248, 6416--6425 35 Agetslnger-Steitz, J. (1969) Nature 224, 957--964 36 Jeffery, W. and Brawerman, G. (1975) Biochemistry 14, 3445--3451 37 Bester, A., Kennedy, D. and Heywood, S. (1975) Proc. Natl. Acad. Sci. U.S. 72, 1513--1527 38 Williamson, R., Morrisson, M., Lanyon, G., Eason, R. and Paul, J. (1971) Biochemistry 10,
3014--3021 39 Holder, J. and Lindgrel, J. (1975) Biochemistry 14, 4209--4215 40 Favre, A., Morel, C. and Scherrer, K. (1975) Eur. J. Biochem. 57, 147--157 41 Olsnes, S. (1971) Eur. J. Biochem. 23, 557--566 42 Leubleu, B., Marbaix, G., Werenne, J., Burny, A. and Huez, G. (1970) Biochem. Biophys. Res.
Commun. 40, 731--739 43 Setyono, B., C~rossmalln, M. and Liautard, J.-P. (1977) Bioehimie, in the press 44 Cashion, L. and Stanley, W. (1974) Proc. Natl. Acad. Sci. U.S. 71 , 436 - -440 45 Hellerman, J.G. and Shafritz, D.A. (1975) Proe. Natl. Aead. ScL U.S. 72, 1021--1025 46 Noll, M. and Noll, H. (1972) Nat. New Biol. 238, 225--228 47 Prichard, P.M., Gilbert, J.M., Shafritz, D.A. and Anderson, W.F. (1970) Nature 226, 511--514 48 Zasloff, M. and Ochoa, S. (1972) Proc. Natl. Acad. Sci. U.S. 69, 1796--1799 49 Wigle, D. (1973) Eur. J. Biochem. 35, 11--17 50 Staehelin, T., Schreier, M., Trachsel, H. and Erni, B. (1975) 10th FEBS Meeting, Abstr. 48 51 Jacob-Lorena, M. and Baglioni, C. (1972) Proc. Natl. Acad. Sei. U.S. 69, 1425---1428 52 Scherrer, K. and Marcaud, L. (1968) J. Cell. Physiol. 72, 181--212 53 Bray, G.A. (1960) Anal. Biochem. 1 , 2 7 9 54 Gander, E., Stewart, A., Morel, C. and Scherrer, K. (1973) Eur. J. Biochem. 38, 443--452 55 Auerbach, S. and Pederson, T. (1975) Biochem. Biophys. Res. Commun. 63, 149--156 56 Marcus, A. (1976) J. Biol. Chem. 245, 955--961 57 Schreier, M.H. and Staehelin, T. (1973) J. Mol. Biol. 73, 329--349 58 Liautard, J.P. and Liautard, J. (1977) Biochim. Biophys. Acta 4 7 4 , 5 8 8 - - 5 9 4





























