Embed Size (px)
Citation preview

ARTICLE IN PRESS
1438-4221/$ - se
doi:10.1016/j.ijm
�Correspondfax: +499131 8
E-mail addr
(M. Hensel).
International Journal of Medical Microbiology 297 (2007) 401–415
www.elsevier.de/ijmm
Protein secretion systems and adhesins: The molecular armory of
Gram-negative pathogens
Roman G. Gerlach, Michael Hensel�
Institut fur Klinische Mikrobiologie, Immunologie und Hygiene, Universitatsklinikum Erlangen, Wasserturmstraße 3-5,
D-91054 Erlangen, Germany
Received 30 January 2007; received in revised form 22 March 2007; accepted 23 March 2007
Abstract
Protein secretion is a basic cellular function found in organisms of all kingdoms of life. Gram-negative bacteria haveevolved a remarkable number of pathways for the transport of proteins across the cell envelope. The secretion systemsfulfill general cellular functions but are also essential for pathogenic bacteria during the interaction with eukaryotichost cells. Secretion systems range from relatively simple structures such as type I secretion systems composed of threesubunits that only secrete one substrate protein to complex machines such as type III and IV secretion systemscomposed of more than 20 subunits that can translocate large sets of effector proteins into eukaryotic target cells. Inthis review, the main structural and functional features of secretion systems are described. One subgroup of substrateproteins of secretion systems are protein adhesins. Despite the conserved function in binding to host cell ligands or toabiotic surfaces, the assembly of the various bacterial adhesins is highly divergent. Here we give an overview on therecent understanding of the assembly of fimbrial and non-fimbrial adhesins and the role of type I, III and V secretionsystems and specialized branches of the general secretion pathway in their biogenesis.r 2007 Elsevier GmbH. All rights reserved.
Keywords: Bacterial protein secretion; Fimbrial adhesins; Non-fimbrial adhesins; Gram-negative pathogen; Cell envelope
Introduction
The transport of proteins across the bacterial cellenvelope is a basic function found in all groups ofbacteria. The analyses of a large number of bacterialgenomes indicate that up to 17% of Proteobacteriagenomes encode for proteins with signal sequences forthe general secretory pathway (GSP) (Bendtsen et al.,2005), with many additional secretion systems and
e front matter r 2007 Elsevier GmbH. All rights reserved.
m.2007.03.017
ing author. Tel.: +499131 852 3640;
52 2531.
ess: [email protected]
substrate proteins being present in most species.Secreted proteins have various functions in processessuch as the biogenesis of the cell envelope, theacquisition of nutrients, motility, intercellular commu-nication and many more. Aggressive bacterial virulencefactors that enable a progressive colonization of hostorganisms are commonly secreted proteins, with toxinsand translocated effector proteins as well-studiedexamples. Protein adhesins are another group ofsecreted proteins found in pathogenic as well asenvironmental bacterial species.
The GSP and its components are found in all threekingdoms of life: in bacteria, archaea and eukaryoticorganelles (chloroplasts but also in the endoplasmic

ARTICLE IN PRESSR.G. Gerlach, M. Hensel / International Journal of Medical Microbiology 297 (2007) 401–415402
reticulum) and provides a generic mechanism for thetransport of proteins across the cytoplasm or organellemembrane. While this transport process is sufficient forthe secretion of proteins in Gram-positive species,Gram-negative species are posed with a specific pro-blem, the transport across a second membrane system,the outer membrane (OM).
The OM is a remarkable structure that enables Gram-negative bacteria to colonize host environments such asthe intestinal lumen, and provides a protective barrieragainst various antimicrobial host defenses as well asagainst antibiotics. However, the OM is also a barrierfor the secretion of proteins, and it is a particularproblem to energize the transport across the OM. Gram-negative bacteria have evolved a remarkable array ofmechanisms for the secretion of proteins across the cellenvelope. In this review, we will briefly describe the keystructural and functional features of the various secre-tion systems, with some systems being studied in greatdetail over several decades and others been discoveredvery recently.
All protein adhesins have to fulfill the same basicrequirements, i.e., to bring a receptor-like domain orsubunit into close contact with a ligand on the host cellsurface and link the domain or subunit to the bacterialsurface. There are two major classes of protein adhesins,(i) the fimbrial adhesins with pili composed of hetero-polymers of several subunits and (ii) non-fimbrialadhesins consisting of a single protein or homotrimers.The assembly of fimbrial as well as non-fimbrialadhesins involves the function of different secretionsystems, and for several adhesins, specific branches ofcommon secretion pathways have evolved. The secondpart of this review will describe our current under-standing of the various adhesins with focus on thesecretion mechanisms underlying the adhesin assembly.
Bacterial protein secretion systems
For Gram-negative bacteria a classification of thesecretion pathways in type I–VI was made primarily dueto the characteristics of the OM secretion mechanisms.A schematic overview of the most common secretionand translocation systems described in more detailbelow is given in Fig. 1.
The general secretory pathway (GSP)
The transport of unfolded proteins into the periplasmis accomplished by a multi-subunit translocon. Thebacterial GSP consists of two heterotrimeric complexes,SecYEG (Yahr and Wickner, 2000) and SecDFYajCwhich are inserted into the inner membrane (IM), andan accessory component, SecA (Rusch and Kendall,
2007). Proteins targeted for the Sec pathway aretranslated as pre-proteins and possess an N-terminalsignal peptide. Co-translational translocation is possiblewhere the newly synthesized signal peptide of thenascent peptide chain is recognized by SecA or SecAtogether with the ‘general chaperone’ SecB and subse-quently targeted to the SecYEG pore complex. SecAitself might also exhibit a cytosolic chaperone activity(Eser and Ehrmann, 2003) and forms presumablydimers (Woodbury et al., 2002). ATP hydrolysiscatalyzed by SecA along with the proton-motive forcedrives the translocation of the pre-protein into theperiplasm. The SecDFYajC complex is thought toenhance translocation through SecYEG by promotingmembrane cycling of SecA (Duong and Wickner, 1997).Upon translocation, the signal peptide is cleaved off thepre-protein by specific periplasmic signal peptidases andthe mature protein is released in the periplasmic space. Ifadditional signals are present, these might be recognizedby components of the terminal branch pathwaysfor subsequent translocation across or insertion intothe OM.
There are three different terminal branch pathwaysallowing subsequent secretion of periplasmic intermedi-ates through the OM: (i) a complex secreton (type II),(ii) an intrinsic activity of the substrate protein (auto-transporter (AT) or type V) or (iii) by the chaperoneusher (CU) pathway (see below).
Type I secretion systems
Type one secretion systems (T1SS) or ATP-bindingcassette (ABC) transporters are heterotrimeric com-plexes consisting of an IM ABC exporter, a membranefusion protein (MFP) and a pore-forming, outer-membrane protein (OMP). T1SS allow the secretion ofa wide range of substrates (proteinaceous and non-proteinaceous) from the cytoplasm to the extracellularspace in a single step, without a stable periplasmicintermediate. Most protein substrates described so farpossess a C-terminal signal sequence which is character-ized by loosely conserved secondary structures (Stanleyet al., 1991) and is not cleaved off during secretion. Thisimplies that co-translational secretion is not possible(reviewed in Delepelaire (2004)). The mechanism of typeI secretion was studied in great detail on the basis of thea-hemolysin (HlyA) secretion found in some uropatho-genic Escherichia coli (UPEC) (Thanabalu et al., 1998).The HlyA secretion system consists of the ABC exporterHlyB (Schmitt et al., 2003), the MFP HlyD (Schulein etal., 1992) and the common OMP TolC (Koronakis etal., 2000). ABC proteins are believed to form homo-dimers and are responsible for substrate recognition andspecificity as well as for energizing the translocation byATP hydrolysis (Binet and Wandersman, 1995). HlyD

ARTICLE IN PRESS
To
lC
HlyB
Hly
D
SecB
C
N
HlyA
N FimC
FimD
FimA
FimF
FimG
FimH
C
N IgAp
C
SecY
Se
cE
Se
cG S
ecD
Se
cF
Ya
jCSec
ASec
A
C
SP
SecB
C
PulD
Pul
G-J,F,
K-N
Pu
lC
PulSPulS
PulE
PulA
DsbA
DegP
ATP
ADP
ATP
ADP
ATP
ADP
VirB2
VirB5
VirB7
VirB
10
VirB
10
VirB6/8B4
B11
ATP
ADP
ATP
ADP
VirB9
B7B7
C
N
T-DNA
T-DNA
VirD4
VirE2
Prg
I
SipD
SipBC
Prg
JInvGInvH
PrgHKInvAKLN
SpaQS
InvCATP
ADP
InvB
C
N
SipA
SopE
etc.
T1SS T4SS T3SS T2SSChaperone/
usher
T5SS
A. tumefaciens
VirB/D4
S. enterica
SPI1
K. oxytoca
pullulanase
E. coli
Type 1 pili
N. gonorrhoeae
IgA1 protease
Sec-dependent pathways
eukaryotic cell
membrane
OM
IM
periplasm
cytoplasm
extracellular
space
VirB1
Fig. 1. Overview of secretion systems in Gram-negative bacteria. Protein transport across the outer membrane of Gram-negative
bacteria can be subdivided into Sec-independent and Sec-dependent pathways. The three main terminal branches allowing further
transport of Sec-transported periplasmic intermediates are: the CU pathway for the synthesis of fimbrial adhesins, the
autotransporter (T5SS) pathway and the complex type II secretion system (T2SS). No stable periplasmic intermediates are found in
type I, type III and type IV secretion systems (T1SS, T3SS, T4SS). Each of the three export mechanisms features a protein-
conducting channel able to span the two bacterial membranes and, in the case of T3SS and T4SS, one additional membrane of the
host cell. Examples for pathogens utilizing the various secretion systems are indicated at the bottom of the figure. See text for further
details (modified from Kostakioti et al. (2005) and Capitani et al. (2006)).
R.G. Gerlach, M. Hensel / International Journal of Medical Microbiology 297 (2007) 401–415 403
seems to be involved in HlyA recognition (Thanabalu etal., 1998) but its main functions are possibly to makecontact to the portions of TolC extending into theperiplasm and maybe to trigger the opening of the TolCchannel in an iris-like fashion. The TolC OMP acts as atrimeric OM channel for several exporter systems suchas T1SS as well as RND (resistance, nodulation,division) type systems. Each monomer of TolC providesfour b-strands in the OM and four very long (100 (A) a-helices in the periplasmic space which combine in thetrimeric form to a large hydrophilic channel. A MFP-triggered unwinding of the coiled–coiled a-helices mightprovide a channel opening with a diameter of 16–20 (Awhich would allow the transport of partially foldedproteins (reviewed in Koronakis et al. (2004)).
Type II secretion systems
This pathway depends on the Sec machinery and isused by a wide variety of Gram-negative bacteria to
secrete enzymes and toxins across the OM. It is the mainterminal branch of the GSP, and best characterized sofar is the secretion of pullulanase (PulA) by Klebsiella
oxytoca (Pugsley et al., 1997). Cryo-electron microscopyrevealed that PulD forms a ring-shaped complex of12–14 subunits with a central cavity in the OM. Forproper targeting and insertion of PulD, the smalllipoprotein PulS is required (Nouwen et al., 1999). Thetype II secreton consists of a multi-subunit complexinserted into the IM, some of the components exhibitingextensive cytoplasmic domains. Interestingly, the pro-teins PulG, H, I and J show a limited homology to thetype IV pilus structural subunit pilin (‘pseudopilins’, seebelow). Little is known about the role of the othercomponents of the secreton (PulF, K, L, M, N) and howthe transport is energized. In the homologous esp systemof Vibrio cholerae, it could be shown that thecytoplasmic EspE (a homolog of PulE) possesses anATP-binding motif and has autokinase activity. EspE isrecruited to the IM by interaction with EspL (Sandkvist

ARTICLE IN PRESSR.G. Gerlach, M. Hensel / International Journal of Medical Microbiology 297 (2007) 401–415404
et al., 1995). This model assumes that EspE hydrolyzesATP which induces conformational changes within theIM components which are transmitted to the periplas-mic domains and finally to PulD/S.
Type III secretion system
Type III secretion system (T3SS) are complex,supramolecular structures which span the IM, theperiplasmic space, the OM, the extracellular space anda host cellular membrane. These complex assemblies arestructurally and evolutionarily related to the flagellasystems. T3SS have been isolated in species of severalGram-negative bacteria (Salmonella, Yersinia, Shigella,Escherichia, Pseudomonas) and were shown to consist ofat least 20 different subunits which enable these bacteriato translocate substrates (effectors) directly into thecytoplasm of the host cell to exert a broad range ofvirulence functions (reviewed in Ghosh (2004)). Becauseof their shape and their ability to translocate proteins ina cell contact-dependent manner, they are also referredto as ‘injectisomes’ or ‘molecular needles’ (Cornelis,2006).
Two oligomeric rings form the major part of the so-called ‘basal body’. The ring complex inserted into theIM consists of multiple copies of at least three proteins.In the Salmonella pathogenicity island 1 (SPI1)-encodedT3SS, the complex is composed of PrgK and PrgH(Kimbrough and Miller, 2000). A central periplasmiccylinder, the ‘inner rod’, is built of PrgJ subunits andprotrudes from an IM ring socket structure (Marlovitset al., 2004). To the cytoplasmic face of the basal bodythe ATPase is bound, presumably as a double-hexamericring, that energizes the translocation (Muller et al.,2006). A characteristic of T3SS is the presence ofcognate chaperones, small acidic proteins. These cha-perones are considered to stabilize and prevent terminalfolding of effector proteins. The energy of ATPhydrolysis by the ATPase is conducted by release ofthe chaperone from an effector chaperone complex andsubsequent loading of the unfolded substrate into theT3SS apparatus (Akeda and Galan, 2005). Anotherfunction of the effector-specific chaperones could be themasking of domains needed for membrane targetingwithin the host cell (Letzelter et al., 2006).
The OM ring complex consists of a member of thesecretin family. Secretins are also involved in theformation of OM pores in type II and type IV secretionsystems (T4SS). The pore monomers are transported bythe Sec-dependent pathway and inserted into the OM ina pilotin-dependent manner (Lario et al., 2005). In T3SSsecretins are believed to exert a stabilizing and anchor-ing function for the needle complex.
The rigid extracellular needle forms a protein-conducting channel extending the inner rod. The hollow
cylinder is made of several hundred copies of proteins ofthe YscF family (Cordes et al., 2003) and shows an innerdiameter of 20–30 (A, which only supports translocationof at least partially unfolded substrates.
Interestingly, the needle length of all T3SS character-ized so far varies in a narrow band around 60 (A. There issome controversy about the mechanism of needle lengthcontrol: The Cornelis group found that the needle lengthin Yersinia depends in a linear fashion on the length of arepetitive domain within YscP (Journet et al., 2003). Inthis model, YscP would act as a molecular ruler andextends through the entire length of the protein-conduct-ing channel of the translocon and is anchored to theneedle tip and the basal body. Once YscP is fullyelongated by the growing needle, it would signal to thebasal body of the T3SS to switch substrate specificityfrom needle components to effectors (Cornelis, 2006). InSalmonella Typhimurium a kinetic regulation wasproposed: The component of the inner rod (PrgJ) andthe needle protein PrgI are secreted as long as thepolymerization of the inner rod is completed. This InvJ-dependent assembly of the inner rod leads to stableanchoring of the needle to the basal body and subsequentsubstrate switch to effectors (Marlovits et al., 2006).Needle length control is believed to be essential for theoptimal function of T3SS. In Yersinia, host cell contact isaccomplished with the major adhesins YadA and Inv.For this reason, the T3SS needle must span at least thedistance determined by these surface molecules to interactwith the target cell (Mota et al., 2005). In the Shigella
system, it was shown that lipopolysaccharide (LPS)length was evolutionary adapted to facilitate T3SSfunction. The LPS is condensed to half of its length byspecific glycosylation to compensate for an optimalneedle length (West et al., 2005).
Attached to the tip of the needle, a structure known asthe ‘needle extension’ was identified. This structure isthought to mediate the formation of the translocationpore and was shown to consist of LcrV in Yersinia
(Mueller et al., 2005) and in Salmonella SPI2 presum-ably of SseB (Chakravortty et al., 2005). The transloca-tion pore inserted into the host cell membrane consistsof the ‘translocator’ proteins, best studied for Yersinia
YopB and YopD (reviewed in Viboud and Bliska(2005)). Using purified translocators of enteropatho-genic E. coli or of Pseudomonas aeruginosa reconstitu-tion of pore complexes was done (Ide et al., 2001;Schoehn et al., 2003). These studies revealed asymme-trical pores with an inner diameter ranging between 30and 50 (A.
Type IV secretion systems (T4SS)
T4SS are characterized by the ability to translocateproteins or complexes of protein and single-stranded

ARTICLE IN PRESSR.G. Gerlach, M. Hensel / International Journal of Medical Microbiology 297 (2007) 401–415 405
DNA. Based on sequence similarities, T4SS are believedto have evolved from bacterial conjugation machineries(reviewed in Cascales and Christie (2003)). The T-DNAtransfer system of Agrobacterium tumefaciens is theprototypical type A T4SS (reviewed in Burns (1999)).This well-studied T4SS translocates protein–DNA com-plexes but is somehow distinct from the T4SS ofpathogens of humans and animals that appear totranslocate proteins only.
Again by sequence comparison, T4SS are categorizedinto two subclasses: type IVA (T4ASS) and type IVB(T4BSS) (Christie and Vogel, 2000). For substraterecruitment and targeting to the IM parts of thetransenvelope protein complex, a homohexamer ofVirD4 acts as the so-called ‘coupling protein’ (CP) inT4ASS. A stable interaction of the CP with homologs ofVirB10, a part of the multi-subunit transenvelopeprotein complex, was demonstrated (Llosa et al.,2003). The components of the transenvelope complexare members of the mating-pair formation (Mpf) proteinfamily. Different functions could be assigned to sets ofVirB proteins: VirB3 and VirB6–10 might form thechannel traversing the periplasmic space. VirB9 is asecretin-like protein which could form the OM pore.The periplasmic VirB1 is a peptidoglycan hydrolase thatis thought to contribute to channel formation (Ward etal., 2002). VirB4 and VirB11 are both IM proteins withputative ATPase activity which could energize thetranslocation and perhaps the pilus polymerization.The T-pilus is an elongated extracellular structure whichis mainly composed of cyclized VirB2 subunits, butVirB5 and VirB7 might also contribute to pilus assemblyor being part of the pilus.
Recent work demonstrated the roles of T4ASS inseveral important human bacterial pathogens. Pertussistoxin of Bordetella pertussis is secreted in a contact-independent manner, while CagA of Helicobacter pylori
is a translocated T4SS effector protein that inducesinflammatory responses and cytoskeletal alterations inthe host cell. T4ASS have also been identified in Brucella
spp. and Bartonella henselae, and the translocatedeffectors have central functions in the intracellularlifestyle of these pathogens. Further details can befound in a recent review by Backert and Meyer (2006).
In contrast to the T4ASS, T4BSS are less wellunderstood. One example of a T4BSS is the virulence-associated dot/icm machinery of Legionella pneumophila.The system was discovered by screening for mutantsunable to survive within macrophages (Vogel andIsberg, 1999). Individual mutants were assigned to dot
(defect in organelle trafficking) or icm (intracellularmultiplication) according to their respective phenotypes.To date only in L. pneumophila substrates of a T4BSShave been described (Segal et al., 2005). Amongst theover hundred experimentally identified or predictedsubstrates, RalF was shown to be secreted into the
cytoplasm of the host cell and acts as a guanidineexchange factor on the host protein ADP ribosylationfactor (ARF-1) (Nagai et al., 2002).
Type V secretion systems
Type V secretion system (T5SS) includes severalmechanisms: AT secretion, the two-partner system(TPS) and the oligomeric coiled-coil adhesin (Oca)system. AT proteins are of modular composition: AnN-terminal signal sequence targets the protein to the Secmachinery at the IM, the passenger domain harbors thespecific effector function and the C-terminal transloca-tion unit forms, once inserted into the OM, a b-barrelsecondary structure which allows secretion of thepassenger domain. ATs are synthesized as pre-pro-proteins, after cleavage of the signal peptide the pro-protein is released into the periplasm. The passengerdomain is, depending on the particular AT, cleaved offfrom its translocation unit after passing the OM and theprotein is released into the extracellular milieu.
In contrast to the classical ATs which are synthesizedas a single polypeptide, in TPSs the passenger domainand the transporter domain are translated as twoseparate proteins which are referred as members of theTpsA and TpsB families, respectively (Jacob-Dubuissonet al., 2004).
A group of non-fimbrial adhesins (see below) belongsto the family of surface-attached oligomeric ATs. Thehallmark of these proteins is that they form oligomericcomplexes at the bacterial surface, and their C-terminaltranslocation units interact to form a b-barrel porewhich functions as a membrane anchor (Koretke et al.,2006). The most prominent member of this family ofmembrane proteins is YadA from Yersinia spp. (Rog-genkamp et al., 2003).
Chaperone usher (CU) pathway
The CU pathway is linked to the assembly of adhesivesurface structures of Gram-negative bacteria. One of thebest characterized systems are type 1 pili from uro-pathogenic E. coli. All the subunits needed for pilusassembly are translated as pre-proteins (pre-pilins) andtranslocated into the periplasm in a Sec-dependentmanner. FimC is a periplasmic chaperone which bindspilus subunits called pilins (FimA, F, G, H) with a 1:1stoichiometry (Vetsch et al., 2004). Complex formationwith the chaperone is essential to prevent prematurepilus assembly in the periplasm and for targeting of thesubunits to the integral OMP FimD. FimD is theassembly platform (usher) from which the nascentpilus is built and anchored. The usher mediatestranslocation of folded subunits through a large pore(Saulino et al., 2000).

ARTICLE IN PRESSR.G. Gerlach, M. Hensel / International Journal of Medical Microbiology 297 (2007) 401–415406
Other secretion pathways
Genome-wide screens for homologs of the T4BSScomponent IcmF revealed a group of pathogenicityislands present in Gram-negative bacteria that havebeen termed IcmF-associated homologous protein(IAHP) cluster. Investigation of the virulence-associatedsecretion cluster in V. cholerae led to the identificationof a new class of secretion systems, termed type VI.This type VI secretion system (T6SS) manages theexport of substrates at least in the extracellular spacewithout the requirement of hydrophobic N-terminalsignal sequences. Using the model host Dictyostelium
discoideum, a virulence function of the T6SS-secreted substrates could be shown, even suggesting atranslocation of these proteins in the cytosol of theamoebae (Pukatzki et al., 2006). Recently, anotherT6SS was identified encoded at the HIS-I locus ofP. aeruginosa (Mougous et al., 2006). In this study anATPase, ClpV1, was identified, presumably energizingthe secretion. Furthermore, the substrate Hcp1 could beshown to be secreted in a T6SS-dependent manner andis present in CF patients with chronic P. aeruginosa
infection.The twin arginine transport (Tat) system and the
YidC system have no known role in pathogenesis andare only briefly described. Tat is the only translocationpathway characterized so far able to transport prefoldedand often oligomeric proteins across membranes (re-viewed in Sargent et al., 2006). A common feature of Tatsubstrates is the presence of a consensus amino acidsequence (SRRxFLK) in the N-terminal signal peptide.This twin-arginine-bearing signal peptide targets pre-cursor proteins to the membrane-associated Tat trans-port apparatus. Hallmarks of this translocationpathway are that it is energized solely by the proton-motive force, and the remarkably low number ofdifferent subunits building up the secretion system.YidC is an E. coli membrane factor involved in thebiogenesis of integral IM proteins, both in conjunctionwith and independent of the Sec system (Luirink et al.,2005).
Bacterial protein adhesins
The ability to adhere to a wide variety of biotic andabiotic surfaces is a feature which can promote bacterialsurvival and is one of the key virulence functions ofmany pathogens. Adhesion is mediated by distinctsurface structures which can be subdivided by theirassembly mechanism and structure into two majorclasses (Fig. 2): fimbrial adhesins and non-fimbrialadhesins (Soto and Hultgren, 1999). The terms non-fimbrial and afimbrial adhesin are often used synony-mously for mono- or oligomeric adhesive surface
structures. A possible distinction could be the presence(non-fimbrial) or absence (afimbrial) of a typicalmembrane anchor in the protein.
Many bacteria are able to express a whole set ofdifferent adhesins, often belonging to different sub-classes, on their surface. This might be an adaptation todifferent environmental conditions or, in the case ofpathogenic bacteria, to different hosts or host tissues. Inthe following section, prototypes of the various adhesinsof Gram-negative bacteria are described.
Fimbrial adhesins
Fimbriae (or pili) are a group of rigid, straightfilamentous appendages on a bacterial surface. They aremost prominent on Gram-negative bacteria, where theyare anchored within the OM. These surface structures,composed of hundreds to thousands of subunits,mediate adhesion via specific interaction with surfacestructures (receptors) present on host cells. Interestingly,it was shown that fimbriae were able to mediateunspecific adhesion by increasing the surface hydro-phobicity of the bacteria (Lindahl et al., 1981).
Type I and P pili
Pyelonephritis-associated (P) pili as well as type I piliare build up in the OM via the CU pathway. Both typesof pili are found in UPEC and exhibit different bindingspecificities due to the nature of the adhesive tip of theirfibrillum. P pili, encoded on the pap operon, bindthrough the PapG adhesin to terminal sugars ofglycolipids present on the surface of host cells. Hostand tissue specificity varies due to the presence ofdifferent PapG adhesins recognizing different butrelated Gala-(1–4)-Gal sugars (Feria et al., 2001). Thefim gene cluster encodes type I pili, with FimH being thepilus adhesin. FimH variants present on non-pathogenicE. coli show high affinity to trimannose-containingglycoprotein receptors. In contrast, UPEC expresspredominantly FimH variants with high affinity tomonomannose residues, found enriched on surfaceglycoproteins of urinary tract cells. FimH-mediatedreceptor binding was shown to mediate bacterialinternalization within bladder cells, resulting in bacterialpersistence and chronic urinary tract infections (Marti-nez et al., 2000). Using subunit-exchange experiments, itcould be shown recently that the gross bindingspecificity (host and cell tropism) does not only dependon the adhesive tip fibrillum but also on the pilus shaft(Duncan et al., 2005).
In Salmonella enterica serovar Typhimurium, type Ipili encoded by the fim gene cluster were shown to beinvolved in adhesion to HeLa cells (Baumler et al.,1996). The complete genome sequence of S. Typhimu-rium revealed the presence of 13 operons with homology
ARTICLE IN PRESS
PapD
PapC
PapA
PapE
PapF
PapG
PapK
Type IVa
pili
Type I
(P) pili
Intimin/
Tir
T1SS-secreted
adhesins
Integral OM
proteins
T5SS-secreted
adhesins
eukaryotic cell membrane
OM
IM
PS
cytoplasm
extracellular
space
YadA
TAA
Pil
E,V-X,
C, N
PilBPilT,
U ATP
ADP
ATP
ADP
PilA
P. aeruginosa K UPEC EPEC/EHEC Yersinia spp. E. coli Yersinia spp. P. fluorescensB. pertussis
D4
D3
D2
D1
Tir
Intimin
D4
D3
D2
D1
D5
Invasin
OmpA
NC
FhaC
SphB1
FhaC
FHA
TPS
?
LapA
LapB
LapD
LapC
LapE
periplasm
FhaB
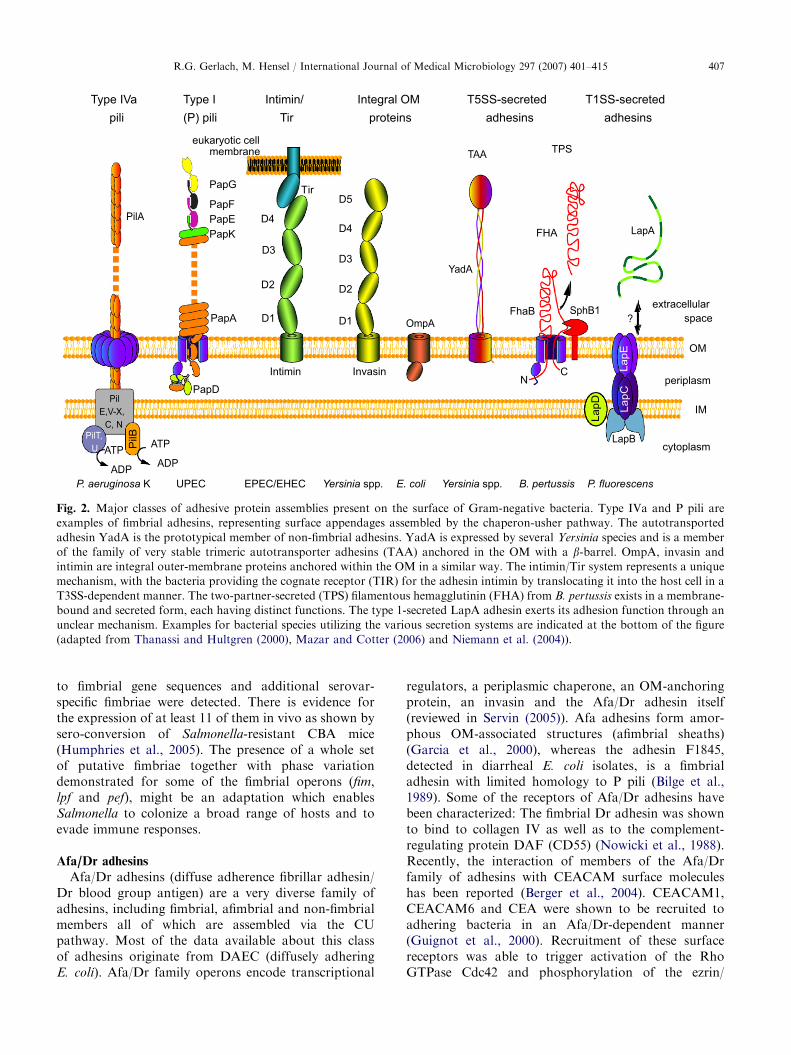
Fig. 2. Major classes of adhesive protein assemblies present on the surface of Gram-negative bacteria. Type IVa and P pili are
examples of fimbrial adhesins, representing surface appendages assembled by the chaperon-usher pathway. The autotransported
adhesin YadA is the prototypical member of non-fimbrial adhesins. YadA is expressed by several Yersinia species and is a member
of the family of very stable trimeric autotransporter adhesins (TAA) anchored in the OM with a b-barrel. OmpA, invasin and
intimin are integral outer-membrane proteins anchored within the OM in a similar way. The intimin/Tir system represents a unique
mechanism, with the bacteria providing the cognate receptor (TIR) for the adhesin intimin by translocating it into the host cell in a
T3SS-dependent manner. The two-partner-secreted (TPS) filamentous hemagglutinin (FHA) from B. pertussis exists in a membrane-
bound and secreted form, each having distinct functions. The type 1-secreted LapA adhesin exerts its adhesion function through an
unclear mechanism. Examples for bacterial species utilizing the various secretion systems are indicated at the bottom of the figure
(adapted from Thanassi and Hultgren (2000), Mazar and Cotter (2006) and Niemann et al. (2004)).
R.G. Gerlach, M. Hensel / International Journal of Medical Microbiology 297 (2007) 401–415 407
to fimbrial gene sequences and additional serovar-specific fimbriae were detected. There is evidence forthe expression of at least 11 of them in vivo as shown bysero-conversion of Salmonella-resistant CBA mice(Humphries et al., 2005). The presence of a whole setof putative fimbriae together with phase variationdemonstrated for some of the fimbrial operons (fim,lpf and pef), might be an adaptation which enablesSalmonella to colonize a broad range of hosts and toevade immune responses.
Afa/Dr adhesins
Afa/Dr adhesins (diffuse adherence fibrillar adhesin/Dr blood group antigen) are a very diverse family ofadhesins, including fimbrial, afimbrial and non-fimbrialmembers all of which are assembled via the CUpathway. Most of the data available about this classof adhesins originate from DAEC (diffusely adheringE. coli). Afa/Dr family operons encode transcriptional
regulators, a periplasmic chaperone, an OM-anchoringprotein, an invasin and the Afa/Dr adhesin itself(reviewed in Servin (2005)). Afa adhesins form amor-phous OM-associated structures (afimbrial sheaths)(Garcia et al., 2000), whereas the adhesin F1845,detected in diarrheal E. coli isolates, is a fimbrialadhesin with limited homology to P pili (Bilge et al.,1989). Some of the receptors of Afa/Dr adhesins havebeen characterized: The fimbrial Dr adhesin was shownto bind to collagen IV as well as to the complement-regulating protein DAF (CD55) (Nowicki et al., 1988).Recently, the interaction of members of the Afa/Drfamily of adhesins with CEACAM surface moleculeshas been reported (Berger et al., 2004). CEACAM1,CEACAM6 and CEA were shown to be recruited toadhering bacteria in an Afa/Dr-dependent manner(Guignot et al., 2000). Recruitment of these surfacereceptors was able to trigger activation of the RhoGTPase Cdc42 and phosphorylation of the ezrin/

ARTICLE IN PRESSR.G. Gerlach, M. Hensel / International Journal of Medical Microbiology 297 (2007) 401–415408
radixin/moesin complex which was accompanied withmembrane rearrangements leading to tight bacterialattachment. Triggering of host cell signaling cascades byinteraction of Afa/Dr adhesins with surface receptorsmight also result in a zipper-like invasion of epithelialcells (Kansau et al., 2004).
CS pili
The assembly of coli surface antigen 1 (CS1) pili ofenterotoxigenic E. coli shows high functional similaritiesto the chaperone/usher pathway but the proteinsinvolved share no detectable sequence similarities. TheCS pili assembly system is therefore termed the‘alternate CU pathway’ (Soto and Hultgren, 1999).
Four linked genes, cooABCD, are essential for thebiogenesis of CS pili. All four proteins are GSP-dependently transported into the periplasm where theperiplasmic chaperone CooB stabilizes CooA, C and D.The minor pilin CooD is thought to initiate pilusassembly and is located on the pilus tip. The OMPCooC presumably is the usher through which CooD andthe major pilin CooA are secreted (Starks et al., 2006).CS1 pili confer binding of E. coli to intestinal epithelialcells and mediate agglutination of bovine erythrocytes;however, the host receptor has not been characterized sofar. Mutational analyses revealed that adhesion as wellas hemagglutination critically depend on amino acid(aa) residues within the minor pilin CooD (Sakellaris etal., 1999).
Type IV pili
Type IV pili are formed in the cytoplasmic membraneby the polymerization of pilin subunits. The assembledpilus structure is extruded across the OM and formslong and flexible surface appendages (reviewed in Craiget al. (2004)). Components of the type IV pilus assemblymachinery are structurally related to T2SS, wherehomologous proteins are called ‘pseudopilins’. TypeIV pilins share a conserved stretch of 25 hydrophobicaa, an unusual N-methylation at their N-terminus and adisulfide-bond at their C-terminus. Based on their aasequence and length type IV pilins are grouped into twosubclasses, type IVa and type IVb. Pili polymerized fromtype IVa and type IVb subunits differ significantly indiameter as well as helical structure (Hansen and Forest,2006). One of the best characterized examples of typeIVa pili are PAK pili, the dominant adhesin ofP. aeruginosa strain K. PAK pili were shown to mediateadherence to mucosal epithelial cells by binding to theglycolipids asalio-GM1 and asalio-GM2 (Krivan et al.,1988). So far, type IVb pili were identified exclusively inbacteria able to colonize the human intestine. InV. cholerae the toxin-coregulated pilus was shown tomediate, besides adhesion, autoaggregation throughhomophilic interactions. This effect is believed to bebeneficial for the bacteria at the site of infection because
microcolonies can be formed and secreted toxins canreach high local concentrations (Kirn et al., 2005). S.
enterica serovar Typhi uses type IVb PilS pili to bind tointestinal epithelial cells (Zhang et al., 2000). Theepithelial surface receptor of PilS was identified as cysticfibrosis transmembrane conductance regulator (CFTR)(Pier et al., 1998). Furthermore, there is experimentalevidence that S. Typhi is able to actively increase CFTRconcentrations on apical membranes to promote adhe-sion and subsequent uptake (Lyczak and Pier, 2002).
A hallmark of type IV pilus biology is their ability toretract through the bacterial cell wall while the pilus tipremains firmly adhered to a receptor structure. Pilusretraction is required for a specialized movement acrosssemisolid matrices (e.g. mucosal epithelia) called ‘twitch-ing motility’ (Burrows, 2005).
Curli
Another type of fimbrial adhesins are curli or thinaggregative fibers (agf). These surface appendages seemto be commonly present in the genome of Salmonella
spp. and E. coli strains. Biosynthesis of curli in E. coli aswell S. Typhimurium depends on the expression of twodivergently orientated operons: csgDEFG and csgBA
(Hammar et al., 1995; Romling et al., 1998). CsgD is atranscriptional regulator of the LuxR family andpromotes expression of several biofilm-associated genesand the csgBA operon (Brombacher et al., 2003).Besides a function in biofilm formation, curli were alsoshown to mediate adhesion of bacteria to the humanmatrix proteins fibronectin and laminin but also toplasminogen, contact phase proteins and major histo-compatibility complex class I molecules (Ben Nasr et al.,1996; Olsen et al., 1998; Robinson et al., 2006).
Curli are assembled at the bacterial surface throughextracellular nucleation precipitation: the major fibersubunit CsgA polymerizes on the surface-exposednucleator CsgB (Hammar et al., 1996). CsgE and CsgFare two periplasmic proteins which are required foroptimal curli assembly and interact with the OMPCsgG. Recently, the OM lipoprotein CsgG was shownto be an integral part of a secretion complex to whichCsgE and CsgF are recruited. This secreton presumablyfacilitates the transport of CsgA and CsgB across theOM (Robinson et al., 2006).
Non-fimbrial adhesins
Non-fimbrial adhesins are a large family of mono- oroligomeric surface-associated proteins which are notassembled by the CU pathway and without separatesubunits for the adhesive tip and pilus shaft. The groupof secreted adhesins (see below), which are notpredominantly surface-associated, mediate bacterialbinding by mechanisms still to be elucidated.

ARTICLE IN PRESSR.G. Gerlach, M. Hensel / International Journal of Medical Microbiology 297 (2007) 401–415 409
Autotransported adhesins
To better account for the specific features of thisprotein family, the Oca family of T5SS-secretedsubstrates was suggested to be termed ‘trimeric ATadhesins’ (TAA) (Linke et al., 2006). YadA is the best-characterized TAA and the major adhesin of entero-pathogenic Yersinia spp. YadA forms stable, high-molecular-weight trimers which protrude about 28 nmfrom the bacterial OM (Hoiczyk et al., 2000). It has beenobserved that Yad length can vary significantly betweendifferent serotypes of Yersinia spp. (Heise and Dersch,2006). YadA mediates binding to several extracellularmatrix (ECM) proteins such as collagen, laminin andfibronectin (Nummelin et al., 2004). BadA fromBartonella henselae was also shown to mediate bindingto ECM proteins (e.g. collagens and fibronectin) but thehair-like appendages observed were much longer(100–300 nm) (Riess et al., 2004). There are numerousother TAAs present or predicted in human and plantpathogens, all of them share a similar moleculararchitecture: They resemble a ‘lollipop’ containing ahead domain, presumably harboring the adhesionfunction, the neck, the stalk, a fibrous, highly repetitivestructure which contains coiled coils, and the anchordomain (reviewed in Linke et al. (2006)).
In S. enterica serovar Typhimurium, the SPI3-encoded adhesin MisL seems to be a ‘classical’ auto-transported substrate, being not homologous to theTAA family members (Dorsey et al., 2005). Anothermonomeric AT of Salmonella, ShdA, was shown to havea function in adhesion and virulence. The surface-located protein, encoded on the CS54 pathogenicityisland, is induced in vivo within the murine cecum andits expression might play a role in the long-termcolonization of mice. GST-fusion proteins of thepassenger domain of ShdA were shown to bindfibronectin in vitro (Kingsley et al., 2002).
The third branch of T5SS, TPS, is represented byfilamentous hemagglutinin (FHA) of Bordetella pertus-
sis. FHA is the product of N- and C-terminal processingof the precursor FhaB. FhaB is recognized and secretedby the OM pore-forming protein FhaC, a member of theTpsB family (Mazar and Cotter, 2006). FHA was shownto function as a multi-functional attachment protein,able to mediate binding to several ligands. A hallmarkof FHA is that a significant amount of the protein isreleased from the bacterial surface by the OM proteaseSphB1. Bacteria deficient for SphB1 adhered signifi-cantly better to cultured epithelial cells but wereattenuated in a mouse model of lung colonization(Coutte et al., 2003). For FHA, immuno-modulatoryfunctions could be assigned to the secreted protein, forexample, by influencing the cytokine pattern secreted bydendritic cells (McGuirk et al., 2002). Release ofadhesins into the extracellular space seems to be acommon but poorly understood phenomenon.
Integral OM adhesins
Several b-barrel-containing OMPs have functions inadhesion (Niemann et al., 2004). OmpA plays animportant role in the crossing of the blood–brain barrierof E. coli causing meningitis in neonates. The receptor ofOmpA was identified as Ecgp, a 96-kDa glycoproteinpresent on the surface of human brain microvascularendothelial cells (Prasadarao, 2002). Neisseria meningi-
tidis possesses another Omp, OpcA, shown to mediatebacterial adhesion and invasion of human epithelial andendothelial cells (Virji et al., 1992). The recognitionpattern of OpcA includes heparan sulfate proteoglycans(de Vries et al., 1998) as well as integrins which arebound via the serum protein vitronectin (Virji et al.,1993). A recent study reported that OpcA binds sialicacid and is specific for pyranose saccharides (Mooreet al., 2005).
The Yersinia pseudotuberculosis Omp invasin binds tomembers of the b1-integrin family, which inducesformation of pseudopods leading to the uptake of thebacteria into M cells (Isberg and Leong, 1990). Invasinconsists of an N-terminal b-barrel membrane anchor,four domains (D1–D4), belonging to the Ig superfamily,and a C-terminal D5 domain. The D5 domainstructurally belongs to the C-type lectins (Kogelbergand Feizi, 2001). There are some overlapping functionsin the Yersinia adhesins invasin and YadA in triggeringb1-integrin-dependent signaling cascades (Hudson et al.,2005), but both adhesins are expressed under distinctenvironmental conditions and may not be co-expressedduring infection (Eitel and Dersch, 2002).
Intimin is another Omp found in EPEC, EHEC andCitrobacter spp. It is related to invasin in primarysequence as well as regarding structural features. TheN-terminal b-barrel membrane anchor is followed bydomains D1–3 (Ig fold) and a C-type lectin domain(D4). Domains D3 and D4 form together a rigidsuperdomain able to bind the cognate receptor translo-cated intimin receptor (Tir) (Batchelor et al., 2000). Tiris an effector molecule of a T3SS which is phosphory-lated by the host cell after translocation and subse-quently inserted into the host cell membrane (Kenny etal., 1997). All components of this adhesion system wherethe bacteria provide both the ligand and the receptor areencoded together within the locus of enterocyte efface-ment. Further members of the family of integral OMPsinclude OmpX of E. coli, Ail of Yersinia enterocolitica
and PagC of S. enterica.
Secreted adhesins
A comprehensive search for putative T1SS substrates ingenomes of Gram-negative bacteria revealed manyputative proteins with predicted adhesion function(Delepelaire, 2004). The criterion used in this screen was

ARTICLE IN PRESS
Table 1. The biofilm-associated proteins (BAP) family of large repetitive proteins involved in bacterial adhesion and/or biofilm
formation
Protein Species Function Reference
BapA Salmonella enterica Biofilm Latasa et al. (2005)
Bap Staphylococcus aureus Biofilm Cucarella et al. (2001)
Bap Burkholderia cepacia Biofilm Huber et al. (2002)
Bhp/Bap Staphylococcus epidermidis Biofilm Tormo et al. (2005)
Esp Enterococcus faecalis Biofilm Shankar et al. (1999)
Espfm Enterococcus faecium Adhesion Lund and Edlund (2003)
LapA Pseudomonas fluorescens Adhesion/biofilm Hinsa et al. (2003)
Lsp Lactobacillus reuteri Adhesion Walter et al. (2005)
R28 Streptococcus pyogenes Adhesion Stalhammar-Carlemalm et al. (1999)
Mus20 Pseudomonas putida Adhesion Espinosa-Urgel et al. (2000)
VAP1445 Vibrio parahaemolyticus Biofilm Enos-Berlage et al. (2005)
YeeJ Escherichia coli Adhesion Roux et al. (2005)
Modified from Lasa and Penades (2006).
R.G. Gerlach, M. Hensel / International Journal of Medical Microbiology 297 (2007) 401–415410
the presence of glycine-rich repeats (GGXGXDXXX),which are found in a majority of T1SS-secreted proteinsand presumably bind calcium ions (Baumann et al.,1993). Many of the predicted adhesins were extremelylarge and repetitive proteins, ranging from 1209 to 8682amino acids.
The largest predicted T1SS-secreted adhesin to date isLapA from the soil bacterium Pseudomonas fluorescens
with about 900 kDa. The protein is encoded within anoperon which harbors also genes coding for a putativeABC transporter (lapBCE). Bacteria deficient in one ofthe lap genes were impaired in adhesion to quartz sand(Hinsa et al., 2003). This adhesin was shown to beloosely surface-associated, a feature which is essentialfor its function and that partially depends on LapD, anIM protein (Hinsa and O’Toole, 2006). Furthermore,LapA seems to play a role in the early steps of biofilmformation, where the bacteria transit from reversible toirreversible attachment (Hinsa et al., 2003). LapA is amember of the BAP family of large, repetitive proteins(Table 1). BAP family members are predominantlyinvolved in biofilm formation but are also shown tomediate adhesion (reviewed in Lasa and Penades(2006)). A cluster of genes encoding a putative type Isecretion system has been identified also in the vicinityof Salmonella Typhimurium bapA, suggesting a com-mon secretion mechanism (Latasa et al., 2005). Re-cently, Gerlach et al. (2007) identified a T1SS encodedby S. enterica SPI4 that secretes SiiE, a highly repetitivenon-fimbrial adhesin of about 600 kDa. SiiE is onlyrequired for adhesion to polarized epithelial cells andco-regulated with the SPI1-encoded T3SS involved inhost cell invasion.
The V. cholerae GbpA adhesion factor is secreted in aT2SS-dependent manner (extracellular protein secretionsystem, EPS). This protein was shown to mediatebinding of bacteria to HT-29 epithelial cells and chitin
beads through interaction with N-acetylglucosamine(GlcNAc) residues (Kirn et al., 2005).
It is difficult to understand by which means a secretedprotein can mediate adhesion, and two alternativemodels have been proposed to explain this phenomenon:(i) The secreted adhesin could bind to its receptor andthen interact with a bacterial cell surface structureproviding a bridging function. (ii) The protein is presentin two forms: secreted and (minor) cell surface-associated, with the surface-bound form exerting theadhesion function or interacting with the secreted form(discussed in Kirn et al. (2005)). None of thesehypotheses are experimentally proven so far.
Adhesins associated with biofilm formation
The ability to form a biofilm can be of crucialimportance for the colonization of certain biotic orabiotic surfaces and can influence the virulence ofpathogenic bacteria. Tight adhesion mediated by specificinteractions of adhesins with receptor structures is aprerequisite of biofilm formation. Homotypic interac-tions between surface-associated adhesins can promotemicrobial aggregation and trigger biofilm formation(reviewed in Dunne (2002)).
There is one large family of adhesins that are presentin Gram-positive as well as in Gram-negative species.Bap of Staphylococcus spp. is the prototypical memberof this BAP family (Table 1). This surface-associatedprotein expressed by different Staphylococcus isolateswas described to mediate biofilm formation of Staphy-
lococcus aureus in a PIA-independent manner (Cucarellaet al., 2001). Bap consists of 2276 amino acids, has an N-terminal secretion signal, a long repetitive central moietyand a C-terminal cell wall-attachment region comprisingan LPXTG motif.

ARTICLE IN PRESSR.G. Gerlach, M. Hensel / International Journal of Medical Microbiology 297 (2007) 401–415 411
All BAP family members share a high molecularmass, a signal sequence for extracellular secretion and arepetitive structure (reviewed in Lasa and Penades(2006)). The number of repeats within BAP is variablenot only among different isolates of Enterococcus
faecalis or Staphylococcus spp. (Shankar et al., 1999;Tormo et al., 2005) but also during the course ofinfection with a single strain of Staphylococcus aureus
(Cucarella et al., 2004). Although size variants ofStaphylococcus aureus Bap did not differ in their abilityto mediate biofilm formation, it could rather be amechanism to evade immune responses by antigenicvariation (Cucarella et al., 2004). In Salmonella Typhi-murium, BapA was identified as a BAP family memberimportant for biofilm formation. Expression of BapAwas shown to depend on CsgD, the global regulator ofbiofilm formation in Salmonella strains (Latasa et al.,2005).
Conclusions and outlook
A remarkable variety of systems for secretion ofproteins into the extracellular space is found in Gram-negative bacteria. Starting from relatively simple T1SSand T5SS apparatuses that usually transport one singlesubstrate protein, secretion systems range to thecomplex T3SS and T4SS that translocate a set ofeffector proteins into eukaryotic host cells. In additionto the secretion of toxins and various other substrateproteins and the translocation of effector proteinsinto eukaryotic host cells, several secretion systemsare involved in the assembly of protein adhesins.Specific branches of the GSP have evolved for theassembly of fimbrial adhesins. In contrast, most non-fimbrial adhesins are secreted by T5SS, and veryrecently the role of T1SS in secretion of large non-fimbrial adhesins involved in biofilm formation andbinding to eukaryotic cells became evident. There is onlyone remarkable example for a T3SS involved inadhesion, i.e., the EPEC/EHEP T3SS that translocatesTir into host cells acting as receptor for the bacterialadhesin.
In most Gram-negative pathogens, copies of thevarious secretion systems can be found with functionsin different phases of the pathogenesis. There is alsohigh redundancy within one class of secretion system,for example, two T3SS or multiple T1SS are present inmany pathogens. This observation may indicate that therepetitive acquisition of gene clusters for secretionsystems by horizontal gene transfer allows a more rapidevolution and adaptation to new niches than theadaptation of existing secretion systems to new ormultiple functions. A similar situation seems to applyfor the large number of adhesins since many pathogenspossess multiple fimbrial adhesins as well as different
non-fimbrial adhesins. The improved understanding ofthe structure of non-fimbrial adhesins suggests thatthese proteins can functionally mimic the fimbrialadhesins. A single polypeptide contains domains thatbuild the head, the shaft and the membrane anchor ofthe adhesin. In fimbrial adhesins, the different subunitshave adapted to these different functions. Several of thenon-fimbrial adhesins are proteins of extremely largesize that are characterized by large numbers of domainrepeats that build the shaft-like part of the molecule.The number of repeats indicates the size constraints forthe non-fimbrial adhesins that are most likely given bythe O-antigen length of the LPS.
We have fairly good understanding of the temporaland spatial control of the expression and function ofvarious protein secretion systems in bacterial pathogens.In contrast, such understanding is incomplete for thenumerous adhesins and future research has to reveal therole of various adhesins in different host species.
Acknowledgements
Work in our lab is supported by grants HE 1964 ofthe Deutsche Forschungsgemeinschaft. M. Hensel alsolikes to thank the Fonds der Chemischen Industrie forsupport. We thank the members of the Hensel lab forcritical comments on the manuscript.
References
Akeda, Y., Galan, J.E., 2005. Chaperone release and unfolding
of substrates in type III secretion. Nature 437, 911–915.
Backert, S., Meyer, T.F., 2006. Type IV secretion systems and
their effectors in bacterial pathogenesis. Curr. Opin.
Microbiol. 9, 207–217.
Batchelor, M., Prasannan, S., Daniell, S., Reece, S., Con-
nerton, I., Bloomberg, G., Dougan, G., Frankel, G.,
Matthews, S., 2000. Structural basis for recognition of the
translocated intimin receptor (Tir) by intimin from
enteropathogenic E. coli. EMBO J. 19, 2452–2464.
Baumann, U., Wu, S., Flaherty, K.M., McKay, D.B., 1993.
Three-dimensional structure of the alkaline protease of
Pseudomonas aeruginosa: a two-domain protein with a
calcium binding parallel beta roll motif. EMBO J. 12,
3357–3364.
Baumler, A.J., Tsolis, R.M., Heffron, F., 1996. Contribution
of fimbrial operons to attachment to and invasion of
epithelial cell lines by Salmonella typhimurium. Infect.
Immun. 64, 1862–1865.
Ben Nasr, A., Olsen, A., Sjobring, U., Muller-Esterl, W.,
Bjorck, L., 1996. Assembly of human contact phase
proteins and release of bradykinin at the surface of curli-
expressing E. coli. Mol. Microbiol. 20, 927–935.
Bendtsen, J.D., Binnewies, T.T., Hallin, P.F., Siche-
ritz-Ponten, T., Ussery, D.W., 2005. Genome update:

ARTICLE IN PRESSR.G. Gerlach, M. Hensel / International Journal of Medical Microbiology 297 (2007) 401–415412
prediction of secreted proteins in 225 bacterial proteomes.
Microbiology 151, 1725–1727.
Berger, C.N., Billker, O., Meyer, T.F., Servin, A.L., Kansau,
I., 2004. Differential recognition of members of the
carcinoembryonic antigen family by Afa/Dr adhesins of
diffusely adhering E. coli (Afa/Dr DAEC). Mol. Microbiol.
52, 963–983.
Bilge, S.S., Clausen, C.R., Lau, W., Moseley, S.L., 1989.
Molecular characterization of a fimbrial adhesin, F1845,
mediating diffuse adherence of diarrhea-associated E. coli
to HEp-2 cells. J. Bacteriol. 171, 4281–4289.
Binet, R., Wandersman, C., 1995. Protein secretion by hybrid
bacterial ABC-transporters: specific functions of the
membrane ATPase and the membrane fusion protein.
EMBO J. 14, 2298–2306.
Brombacher, E., Dorel, C., Zehnder, A.J., Landini, P., 2003.
The curli biosynthesis regulator CsgD co-ordinates the
expression of both positive and negative determinants for
biofilm formation in E. coli. Microbiology 149, 2847–2857.
Burns, D.L., 1999. Biochemistry of type IV secretion. Curr.
Opin. Microbiol. 2, 25–29.
Burrows, L.L., 2005. Weapons of mass retraction. Mol.
Microbiol. 57, 878–888.
Capitani, G., Eidam, O., Glockshuber, R., Grutter, M.G.,
2006. Structural and functional insights into the assembly
of type 1 pili from E. coli. Microbes Infect. 8, 2284–2290.
Cascales, E., Christie, P.J., 2003. The versatile bacterial type
IV secretion systems. Nat. Rev. Microbiol. 1, 137–149.
Chakravortty, D., Rohde, M., Jager, L., Deiwick, J., Hensel,
M., 2005. Formation of a novel surface structure encoded
by Salmonella pathogenicity island 2. EMBO J. 24,
2043–2052.
Christie, P.J., Vogel, J.P., 2000. Bacterial type IV secretion:
conjugation systems adapted to deliver effector molecules
to host cells. Trends Microbiol. 8, 354–360.
Cordes, F.S., Komoriya, K., Larquet, E., Yang, S., Egelman,
E.H., Blocker, A., Lea, S.M., 2003. Helical structure of the
needle of the type III secretion system of Shigella flexneri.
J. Biol. Chem. 278, 17103–17107.
Cornelis, G.R., 2006. The type III secretion injectisome. Nat.
Rev. Microbiol. 4, 811–825.
Coutte, L., Alonso, S., Reveneau, N., Willery, E., Quatannens,
B., Locht, C., Jacob-Dubuisson, F., 2003. Role of adhesin
release for mucosal colonization by a bacterial pathogen. J.
Exp. Med. 197, 735–742.
Craig, L., Pique, M.E., Tainer, J.A., 2004. Type IV pilus
structure and bacterial pathogenicity. Nat. Rev. Microbiol.
2, 363–378.
Cucarella, C., Solano, C., Valle, J., Amorena, B., Lasa, I.,
Penades, J.R., 2001. Bap, a Staphylococcus aureus surface
protein involved in biofilm formation. J. Bacteriol. 183,
2888–2896.
Cucarella, C., Tormo, M.A., Ubeda, C., Trotonda, M.P.,
Monzon, M., Peris, C., Amorena, B., Lasa, I., Penades,
J.R., 2004. Role of biofilm-associated protein bap in the
pathogenesis of bovine Staphylococcus aureus. Infect.
Immun. 72, 2177–2185.
de Vries, F., Cole, R., Dankert, J., Frosch, M., van Putten, J.,
1998. Neisseria meningitidis producing the Opc adhesin
binds epithelial cell proteoglycan receptors. Mol. Micro-
biol. 27, 1203–1212.
Delepelaire, P., 2004. Type I secretion in Gram-negative
bacteria. Biochim. Biophys. Acta 1694, 149–161.
Dorsey, C.W., Laarakker, M.C., Humphries, A.D., Weening,
E.H., Baumler, A.J., 2005. Salmonella enterica serotype
Typhimurium MisL is an intestinal colonization factor that
binds fibronectin. Mol. Microbiol. 57, 196–211.
Duncan, M.J., Mann, E.L., Cohen, M.S., Ofek, I., Sharon, N.,
Abraham, S.N., 2005. The distinct binding specificities
exhibited by enterobacterial type 1 fimbriae are determined
by their fimbrial shafts. J. Biol. Chem. 280, 37707–37716.
Dunne Jr., W.M., 2002. Bacterial adhesion: seen any good
biofilms lately? Clin. Microbiol. Rev. 15, 155–166.
Duong, F., Wickner, W., 1997. The SecDFyajC domain of
preprotein translocase controls preprotein movement by
regulating SecA membrane cycling. EMBO J. 16,
4871–4879.
Eitel, J., Dersch, P., 2002. The YadA protein of Yersinia
pseudotuberculosis mediates high-efficiency uptake into
human cells under environmental conditions in which
invasin is repressed. Infect. Immun. 70, 4880–4891.
Enos-Berlage, J.L., Guvener, Z.T., Keenan, C.E., McCarter,
L.L., 2005. Genetic determinants of biofilm development of
opaque and translucent Vibrio parahaemolyticus. Mol.
Microbiol. 55, 1160–1182.
Eser, M., Ehrmann, M., 2003. SecA-dependent quality control
of intracellular protein localization. Proc. Natl. Acad. Sci.
USA 100, 13231–13234.
Espinosa-Urgel, M., Salido, A., Ramos, J.L., 2000. Genetic
analysis of functions involved in adhesion of Pseudomonas
putida to seeds. J. Bacteriol. 182, 2363–2369.
Feria, C., Machado, J., Duarte Correia, J., Goncalves, J.,
Gaastra, W., 2001. Distribution of papG alleles among
uropathogenic E. coli isolated from different species. FEMS
Microbiol. Lett. 202, 205–208.
Garcia, M.I., Jouve, M., Nataro, J.P., Gounon, P., Le
Bouguenec, C., 2000. Characterization of the AfaD-like
family of invasins encoded by pathogenic E. coli associated
with intestinal and extra-intestinal infections. FEBS Lett.
479, 111–117.
Gerlach, R.G., Jackel, D., Stecher, B., Wagner, C., Lupas, L.,
Hardt, W.D., Hensel, M., 2007. Salmonella pathogenicity
island 4 encodes a giant non-fimbrial adhesin and the
cognate type 1 secretion system. Cell. Microbiol. [E-pub
ahead of print, doi:10.1111/j.1462-5822.2007.00919.x].
Ghosh, P., 2004. Process of protein transport by the
type III secretion system. Microbiol. Mol. Biol. Rev. 68,
771–795.
Guignot, J., Peiffer, I., Bernet-Camard, M.F., Lublin, D.M.,
Carnoy, C., Moseley, S.L., Servin, A.L., 2000. Recruitment
of CD55 and CD66e brush border-associated glycosylphos-
phatidylinositol-anchored proteins by members of the Afa/
Dr diffusely adhering family of E. coli that infect the human
polarized intestinal Caco-2/TC7 cells. Infect. Immun. 68,
3554–3563.
Hammar, M., Arnqvist, A., Bian, Z., Olsen, A., Normark, S.,
1995. Expression of two csg operons is required for
production of fibronectin- and Congo red-binding curli
polymers in E. coli K-12. Mol. Microbiol. 18, 661–670.

ARTICLE IN PRESSR.G. Gerlach, M. Hensel / International Journal of Medical Microbiology 297 (2007) 401–415 413
Hammar, M., Bian, Z., Normark, S., 1996. Nucleator-
dependent intercellular assembly of adhesive curli organ-
elles in E. coli. Proc. Natl. Acad. Sci. USA 93, 6562–6566.
Hansen, J.K., Forest, K.T., 2006. Type IV pilin structures:
insights on shared architecture, fiber assembly, receptor
binding and type II secretion. J. Mol. Microbiol. Biotech-
nol. 11, 192–207.
Heise, T., Dersch, P., 2006. Identification of a domain in
Yersinia virulence factor YadA that is crucial for extra-
cellular matrix-specific cell adhesion and uptake. Proc.
Natl. Acad. Sci. USA 103, 3375–3380.
Hinsa, S.M., O’Toole, G.A., 2006. Biofilm formation by
Pseudomonas fluorescens WCS365: a role for LapD.
Microbiology 152, 1375–1383.
Hinsa, S.M., Espinosa-Urgel, M., Ramos, J.L., O’Toole, G.A.,
2003. Transition from reversible to irreversible attachment
during biofilm formation by Pseudomonas fluorescens
WCS365 requires an ABC transporter and a large secreted
protein. Mol. Microbiol. 49, 905–918.
Hoiczyk, E., Roggenkamp, A., Reichenbecher, M., Lupas, A.,
Heesemann, J., 2000. Structure and sequence analysis of
Yersinia YadA and Moraxella UspAs reveal a novel class of
adhesins. EMBO J. 19, 5989–5999.
Huber, B., Riedel, K., Kothe, M., Givskov, M., Molin, S.,
Eberl, L., 2002. Genetic analysis of functions involved in
the late stages of biofilm development in Burkholderia
cepacia H111. Mol. Microbiol. 46, 411–426.
Hudson, K.J., Bliska, J.B., Bouton, A.H., 2005. Distinct
mechanisms of integrin binding by Yersinia pseudotubercu-
losis adhesins determine the phagocytic response of host
macrophages. Cell. Microbiol. 7, 1474–1489.
Humphries, A., Deridder, S., Baumler, A.J., 2005. Salmonella
enterica serotype Typhimurium fimbrial proteins serve as
antigens during infection of mice. Infect. Immun. 73,
5329–5338.
Ide, T., Laarmann, S., Greune, L., Schillers, H., Oberleithner,
H., Schmidt, M.A., 2001. Characterization of translocation
pores inserted into plasma membranes by type III-secreted
Esp proteins of enteropathogenic E. coli. Cell. Microbiol. 3,
669–679.
Isberg, R.R., Leong, J.M., 1990. Multiple beta 1 chain
integrins are receptors for invasin, a protein that promotes
bacterial penetration into mammalian cells. Cell 60,
861–871.
Jacob-Dubuisson, F., Fernandez, R., Coutte, L., 2004. Protein
secretion through autotransporter and two-partner path-
ways. Biochim. Biophys. Acta 1694, 235–257.
Journet, L., Agrain, C., Broz, P., Cornelis, G.R., 2003. The
needle length of bacterial injectisomes is determined by a
molecular ruler. Science 302, 1757–1760.
Kansau, I., Berger, C., Hospital, M., Amsellem, R., Nicolas,
V., Servin, A.L., Bernet-Camard, M.F., 2004. Zipper-like
internalization of Dr-positive E. coli by epithelial cells is
preceded by an adhesin-induced mobilization of raft-
associated molecules in the initial step of adhesion. Infect.
Immun. 72, 3733–3742.
Kenny, B., DeVinney, R., Stein, M., Reinscheid, D.J., Frey,
E.A., Finlay, B.B., 1997. Enteropathogenic E. coli (EPEC)
transfers its receptor for intimate adherence into mamma-
lian cells. Cell 91, 511–520.
Kimbrough, T.G., Miller, S.I., 2000. Contribution of Salmo-
nella typhimurium type III secretion components to needle
complex formation. Proc. Natl. Acad. Sci. USA
11008–11013.
Kingsley, R.A., Santos, R.L., Keestra, A.M., Adams, L.G.,
Baumler, A.J., 2002. Salmonella enterica serotype Typhi-
murium ShdA is an outer membrane fibronectin-binding
protein that is expressed in the intestine. Mol. Microbiol.
43, 895–905.
Kirn, T.J., Jude, B.A., Taylor, R.K., 2005. A colonization
factor links Vibrio cholerae environmental survival and
human infection. Nature 438, 863–866.
Kogelberg, H., Feizi, T., 2001. New structural insights into
lectin-type proteins of the immune system. Curr. Opin.
Struct. Biol. 11, 635–643.
Koretke, K.K., Szczesny, P., Gruber, M., Lupas, A.N., 2006.
Model structure of the prototypical non-fimbrial adhesin
YadA of Yersinia enterocolitica. J. Struct. Biol. 155,
154–161.
Koronakis, V., Sharff, A., Koronakis, E., Luisi, B., Hughes,
C., 2000. Crystal structure of the bacterial membrane
protein TolC central to multidrug efflux and protein export.
Nature 405, 914–919.
Koronakis, V., Eswaran, J., Hughes, C., 2004. Structure and
function of TolC: the bacterial exit duct for proteins and
drugs. Annu. Rev. Biochem. 73, 467–489.
Kostakioti, M., Newman, C.L., Thanassi, D.G., Stathopoulos,
C., 2005. Mechanisms of protein export across the bacterial
outer membrane. J. Bacteriol. 187, 4306–4314.
Krivan, H.C., Ginsburg, V., Roberts, D.D., 1988. Pseudomo-
nas aeruginosa and Pseudomonas cepacia isolated from
cystic fibrosis patients bind specifically to gangliotetraosyl-
ceramide (asialo GM1) and gangliotriaosylceramide (asialo
GM2). Arch. Biochem. Biophys. 260, 493–496.
Lario, P.I., Pfuetzner, R.A., Frey, E.A., Creagh, L., Haynes,
C., Maurelli, A.T., Strynadka, N.C., 2005. Structure and
biochemical analysis of a secretin pilot protein. EMBO J.
24, 1111–1121.
Lasa, I., Penades, J.R., 2006. Bap: a family of surface
proteins involved in biofilm formation. Res. Microbiol.
157, 99–107.
Latasa, C., Roux, A., Toledo-Arana, A., Ghigo, J.M.,
Gamazo, C., Penades, J.R., Lasa, I., 2005. BapA, a large
secreted protein required for biofilm formation and host
colonization of Salmonella enterica serovar Enteritidis.
Mol. Microbiol. 58, 1322–1339.
Letzelter, M., Sorg, I., Mota, L.J., Meyer, S., Stalder, J.,
Feldman, M., Kuhn, M., Callebaut, I., Cornelis, G.R.,
2006. The discovery of SycO highlights a new function for
type III secretion effector chaperones. EMBO J. 25,
3223–3233.
Lindahl, M., Faris, A., Wadstrom, T., Hjerten, S., 1981. A new
test based on ‘salting out’ to measure relative surface
hydrophobicity of bacterial cells. Biochim. Biophys. Acta
677, 471–476.
Linke, D., Riess, T., Autenrieth, I.B., Lupas, A., Kempf, V.A.,
2006. Trimeric autotransporter adhesins: variable structure,
common function. Trends Microbiol. 14, 264–270.
Llosa, M., Zunzunegui, S., de la Cruz, F., 2003. Conjugative
coupling proteins interact with cognate and heterologous

ARTICLE IN PRESSR.G. Gerlach, M. Hensel / International Journal of Medical Microbiology 297 (2007) 401–415414
VirB10-like proteins while exhibiting specificity for
cognate relaxosomes. Proc. Natl. Acad. Sci. USA 100,
10465–10470.
Luirink, J., von Heijne, G., Houben, E., de Gier, J.W., 2005.
Biogenesis of inner membrane proteins in E. coli. Annu.
Rev. Microbiol. 59, 329–355.
Lund, B., Edlund, C., 2003. Bloodstream isolates of Entero-
coccus faecium enriched with the enterococcal surface
protein gene, esp, show increased adhesion to eukaryotic
cells. J. Clin. Microbiol. 41, 5183–5185.
Lyczak, J.B., Pier, G.B., 2002. Salmonella enterica serovar
Typhi modulates cell surface expression of its receptor, the
cystic fibrosis transmembrane conductance regulator, on
the intestinal epithelium. Infect. Immun. 70, 6416–6423.
Marlovits, T.C., Kubori, T., Sukhan, A., Thomas, D.R.,
Galan, J.E., Unger, V.M., 2004. Structural insights into the
assembly of the type III secretion needle complex. Science
306, 1040–1042.
Marlovits, T.C., Kubori, T., Lara-Tejero, M., Thomas, D.,
Unger, V.M., Galan, J.E., 2006. Assembly of the inner rod
determines needle length in the type III secretion injecti-
some. Nature 441, 637–640.
Martinez, J.J., Mulvey, M.A., Schilling, J.D., Pinkner, J.S.,
Hultgren, S.J., 2000. Type 1 pilus-mediated bacterial
invasion of bladder epithelial cells. EMBO J. 19,
2803–2812.
Mazar, J., Cotter, P.A., 2006. Topology and maturation of
filamentous haemagglutinin suggest a new model for two-
partner secretion. Mol. Microbiol. 62, 641–654.
McGuirk, P., McCann, C., Mills, K.H., 2002. Pathogen-
specific T regulatory 1 cells induced in the respiratory tract
by a bacterial molecule that stimulates interleukin 10
production by dendritic cells: a novel strategy for evasion
of protective T helper type 1 responses by Bordetella
pertussis. J. Exp. Med. 195, 221–231.
Moore, J., Bailey, S.E., Benmechernene, Z., Tzitzilonis, C.,
Griffiths, N.J., Virji, M., Derrick, J.P., 2005. Recognition
of saccharides by the OpcA, OpaD, and OpaB outer
membrane proteins from Neisseria meningitidis. J. Biol.
Chem. 280, 31489–31497.
Mota, L.J., Journet, L., Sorg, I., Agrain, C., Cornelis, G.R.,
2005. Bacterial injectisomes: needle length does matter.
Science 307, 1278.
Mougous, J.D., Cuff, M.E., Raunser, S., Shen, A., Zhou, M.,
Gifford, C.A., Goodman, A.L., Joachimiak, G., Ordonez,
C.L., Lory, S., Walz, T., Joachimiak, A., Mekalanos, J.J.,
2006. A virulence locus of Pseudomonas aeruginosa encodes
a protein secretion apparatus. Science 312, 1526–1530.
Mueller, C.A., Broz, P., Muller, S.A., Ringler, P., Erne-Brand,
F., Sorg, I., Kuhn, M., Engel, A., Cornelis, G.R., 2005. The
V-antigen of Yersinia forms a distinct structure at the tip of
injectisome needles. Science 310, 674–676.
Muller, S.A., Pozidis, C., Stone, R., Meesters, C., Chami, M.,
Engel, A., Economou, A., Stahlberg, H., 2006. Double
hexameric ring assembly of the type III protein translocase
ATPase HrcN. Mol. Microbiol. 61, 119–125.
Nagai, H., Kagan, J.C., Zhu, X., Kahn, R.A., Roy, C.R.,
2002. A bacterial guanine nucleotide exchange factor
activates ARF on Legionella phagosomes. Science 295,
679–682.
Niemann, H.H., Schubert, W.D., Heinz, D.W., 2004. Adhesins
and invasins of pathogenic bacteria: a structural view.
Microbes Infect. 6, 101–112.
Nouwen, N., Ranson, N., Saibil, H., Wolpensingers, B.,
Engels, A., Ghazi, A., Pugsley, A.P., 1999. Secretin PulD:
Association with pilot PulS, structure, and ion-conducting
channel formation. Proc. Natl. Acad. Sci. USA 96,
8173–8177.
Nowicki, B., Moulds, J., Hull, R., Hull, S., 1988. A
hemagglutinin of uropathogenic E. coli recognizes the Dr
blood group antigen. Infect. Immun. 56, 1057–1060.
Nummelin, H., Merckel, M.C., Leo, J.C., Lankinen, H.,
Skurnik, M., Goldman, A., 2004. The Yersinia adhesin
YadA collagen-binding domain structure is a novel left-
handed parallel beta-roll. EMBO J. 23, 701–711.
Olsen, A., Wick, M.J., Morgelin, M., Bjorck, L., 1998. Curli,
fibrous surface proteins of E. coli, interact with major
histocompatibility complex class I molecules. Infect.
Immun. 66, 944–949.
Pier, G.B., Grout, M., Zaidi, T., Meluleni, G., Mueschenborn,
S.S., Banting, G., Ratcliff, R., Evans, M.J., Colledge,
W.H., 1998. Salmonella typhi uses CFTR to enter intestinal
epithelial cells. Nature 393, 79–82.
Prasadarao, N.V., 2002. Identification of E. coli outer
membrane protein A receptor on human brain microvas-
cular endothelial cells. Infect. Immun. 70, 4556–4563.
Pugsley, A.P., Francetic, O., Possot, O.M., Sauvonnet, N.,
Hardie, K.R., 1997. Recent progress and future directions
in studies of the main terminal branch of the general
secretory pathway in Gram-negative bacteria – a review.
Gene 192, 13–19.
Pukatzki, S., Ma, A.T., Sturtevant, D., Krastins, B., Sarraci-
no, D., Nelson, W.C., Heidelberg, J.F., Mekalanos, J.J.,
2006. Identification of a conserved bacterial protein
secretion system in Vibrio cholerae using the Dictyostelium
host model system. Proc. Natl. Acad. Sci. USA 103,
1528–1533.
Riess, T., Andersson, S.G., Lupas, A., Schaller, M., Schafer,
A., Kyme, P., Martin, J., Walzlein, J.H., Ehehalt, U.,
Lindroos, H., Schirle, M., Nordheim, A., Autenrieth, I.B.,
Kempf, V.A., 2004. Bartonella adhesin A mediates a
proangiogenic host cell response. J. Exp. Med. 200,
1267–1278.
Robinson, L.S., Ashman, E.M., Hultgren, S.J., Chapman,
M.R., 2006. Secretion of curli fibre subunits is mediated by
the outer membrane-localized CsgG protein. Mol. Micro-
biol. 59, 870–881.
Roggenkamp, A., Ackermann, N., Jacobi, C.A., Truelzsch,
K., Hoffmann, H., Heesemann, J., 2003. Molecular analysis
of transport and oligomerization of the Yersinia enteroco-
litica adhesin YadA. J. Bacteriol. 185, 3735–3744.
Romling, U., Bian, Z., Hammar, M., Sierralta, W.D.,
Normark, S., 1998. Curli fibers are highly conserved
between Salmonella typhimurium and E. coli with respect
to operon structure and regulation. J. Bacteriol. 180,
722–731.
Roux, A., Beloin, C., Ghigo, J.M., 2005. Combined inactiva-
tion and expression strategy to study gene function under
physiological conditions: application to identification of
new E. coli adhesins. J. Bacteriol. 187, 1001–1013.

ARTICLE IN PRESSR.G. Gerlach, M. Hensel / International Journal of Medical Microbiology 297 (2007) 401–415 415
Rusch, S.L., Kendall, D.A., 2007. Oligomeric states of the
SecA and SecYEG core components of the bacterial Sec
translocon. Biochim. Biophys. Acta 1768, 5–12.
Sakellaris, H., Munson, G.P., Scott, J.R., 1999. A conserved
residue in the tip proteins of CS1 and CFA/I pili of
enterotoxigenic E. coli that is essential for adherence. Proc.
Natl. Acad. Sci. USA 96, 12828–12832.
Sandkvist, M., Bagdasarian, M., Howard, S.P., DiRita, V.J.,
1995. Interaction between the autokinase EpsE and EpsL in
the cytoplasmic membrane is required for extracellular
secretion in Vibrio cholerae. EMBO J. 14, 1664–1673.
Sargent, F., Berks, B.C., Palmer, T., 2006. Pathfinders and
trailblazers: a prokaryotic targeting system for transport of
folded proteins. FEMS Microbiol. Lett. 254, 198–207.
Saulino, E.T., Bullitt, E., Hultgren, S.J., 2000. Snapshots of
usher-mediated protein secretion and ordered pilus assem-
bly. Proc. Natl. Acad. Sci. USA 97, 9240–9245.
Schmitt, L., Benabdelhak, H., Blight, M.A., Holland, I.B.,
Stubbs, M.T., 2003. Crystal structure of the nucleotide-
binding domain of the ABC-transporter haemolysin B:
identification of a variable region within ABC helical
domains. J. Mol. Biol. 330, 333–342.
Schoehn, G., Di Guilmi, A.M., Lemaire, D., Attree, I.,
Weissenhorn, W., Dessen, A., 2003. Oligomerization of
type III secretion proteins PopB and PopD precedes pore
formation in Pseudomonas. EMBO J. 22, 4957–4967.
Schulein, R., Gentschev, I., Mollenkopf, H.J., Goebel, W.,
1992. A topological model for the haemolysin translocator
protein HlyD. Mol. Gen. Genet. 234, 155–163.
Segal, G., Feldman, M., Zusman, T., 2005. The Icm/Dot type-
IV secretion systems of Legionella pneumophila and
Coxiella burnetii. FEMS Microbiol. Rev. 29, 65–81.
Servin, A.L., 2005. Pathogenesis of Afa/Dr diffusely adhering
E. coli. Clin. Microbiol. Rev. 18, 264–292.
Shankar, V., Baghdayan, A.S., Huycke, M.M., Lindahl, G.,
Gilmore, M.S., 1999. Infection-derived Enterococcus faeca-
lis strains are enriched in esp, a gene encoding a novel
surface protein. Infect. Immun. 67, 193–200.
Soto, G.E., Hultgren, S.J., 1999. Bacterial adhesins: common
themes and variations in architecture and assembly. J.
Bacteriol. 181, 1059–1071.
Stalhammar-Carlemalm, M., Areschoug, T., Larsson, C.,
Lindahl, G., 1999. The R28 protein of Streptococcus
pyogenes is related to several group B streptococcal surface
proteins, confers protective immunity and promotes bind-
ing to human epithelial cells. Mol. Microbiol. 33, 208–219.
Stanley, P., Koronakis, V., Hughes, C., 1991. Mutational
analysis supports a role for multiple structural features in
the C-terminal secretion signal of E. coli haemolysin. Mol.
Microbiol. 5, 2391–2403.
Starks, A.M., Froehlich, B.J., Jones, T.N., Scott, J.R., 2006.
Assembly of CS1 pili: the role of specific residues of the
major pilin, CooA. J. Bacteriol. 188, 231–239.
Thanabalu, T., Koronakis, E., Hughes, C., Koronakis, V.,
1998. Substrate-induced assembly of a contiguous channel
for protein export from E. coli: reversible bridging of an
inner-membrane translocase to an outer membrane exit
pore. EMBO J. 17, 6487–6496.
Thanassi, D.G., Hultgren, S.J., 2000. Multiple pathways allow
protein secretion across the bacterial outer membrane.
Curr. Opin. Cell Biol. 12, 420–430.
Tormo, M.A., Knecht, E., Gotz, F., Lasa, I., Penades, J.R.,
2005. Bap-dependent biofilm formation by pathogenic
species of Staphylococcus: evidence of horizontal gene
transfer? Microbiology 151, 2465–2475.
Vetsch, M., Puorger, C., Spirig, T., Grauschopf, U., Weber-
Ban, E.U., Glockshuber, R., 2004. Pilus chaperones
represent a new type of protein-folding catalyst. Nature
431, 329–333.
Viboud, G.I., Bliska, J.B., 2005. Yersinia outer proteins: role in
modulation of host cell signaling responses and pathogen-
esis. Annu. Rev. Microbiol. 59, 69–89.
Virji, M., Makepeace, K., Ferguson, D.J., Achtman, M.,
Sarkari, J., Moxon, E.R., 1992. Expression of the Opc
protein correlates with invasion of epithelial and endothe-
lial cells by Neisseria meningitidis. Mol. Microbiol. 6,
2785–2795.
Virji, M., Makepeace, K., Ferguson, D.J., Achtman, M.,
Moxon, E.R., 1993. Meningococcal Opa and Opc proteins:
their role in colonization and invasion of human epithelial
and endothelial cells. Mol. Microbiol. 10, 499–510.
Vogel, J.P., Isberg, R.R., 1999. Cell biology of Legionella
pneumophila. Curr. Opin. Microbiol. 2, 30–34.
Walter, J., Chagnaud, P., Tannock, G.W., Loach, D.M., Dal
Bello, F., Jenkinson, H.F., Hammes, W.P., Hertel, C.,
2005. A high-molecular-mass surface protein (Lsp)
and methionine sulfoxide reductase B (MsrB) contr-
ibute to the ecological performance of Lactobacillus
reuteri in the murine gut. Appl. Environ. Microbiol. 71,
979–986.
Ward, D.V., Draper, O., Zupan, J.R., Zambryski, P.C., 2002.
Peptide linkage mapping of the Agrobacterium tumefaciens
vir-encoded type IV secretion system reveals protein
subassemblies. Proc. Natl. Acad. Sci. USA 99,
11493–11500.
West, N.P., Sansonetti, P., Mounier, J., Exley, R.M., Parsot,
C., Guadagnini, S., Prevost, M.C., Prochnicka-Chalufour,
A., Delepierre, M., Tanguy, M., Tang, C.M., 2005.
Optimization of virulence functions through glucosylation
of Shigella LPS. Science 307, 1313–1317.
Woodbury, R.L., Hardy, S.J., Randall, L.L., 2002. Complex
behavior in solution of homodimeric SecA. Protein Sci. 11,
875–882.
Yahr, T.L., Wickner, W.T., 2000. Evaluating the oligomeric
state of SecYEG in preprotein translocase. EMBO J. 19,
4393–4401.
Zhang, X.L., Tsui, I.S., Yip, C.M., Fung, A.W., Wong, D.K.,
Dai, X., Yang, Y., Hackett, J., Morris, C., 2000. Salmonella
enterica serovar Typhi uses type IVB pili to enter human
intestinal epithelial cells. Infect. Immun. 68, 3067–3073.

















![The Giant Adhesin SiiE of Salmonella enterica · 2017. 6. 19. · and a type II secretion system related mechanism for type IV pili [10]. Fimbrial adhesins are characterized by their](https://img.pdfslide.us/doc/110x75/60ca8916ca11d329b82d2654/the-giant-adhesin-siie-of-salmonella-enterica-2017-6-19-and-a-type-ii-secretion.jpg)











