Embed Size (px)
Citation preview

Protein conformational dynamics in themechanism of HIV-1 protease catalysisVladimir Yu. Torbeeva,b, H. Raghuramana,c, Donald Hamelbergd, Marco Tonellie, William M. Westlere,Eduardo Perozoa,c, and Stephen B. H. Kenta,b,c,1
aInstitute for Biophysical Dynamics, bDepartment of Chemistry, and cDepartment of Biochemistry and Molecular Biology, University of Chicago,Chicago, IL 60637; dDepartment of Chemistry, Georgia State University, Atlanta, GA 30302; and eNational Magnetic Resonance Facility at Madison,Madison, WI 53706
Edited by Adriaan Bax, National Institutes of Health, Bethesda, MD, and approved October 25, 2011 (received for review July 12, 2011)
We have used chemical protein synthesis and advanced physicalmethods to probe dynamics-function correlations for the HIV-1protease, an enzyme that has received considerable attention asa target for the treatment of AIDS. Chemical synthesis was usedto prepare a series of unique analogues of the HIV-1 protease inwhich the flexibility of the “flap” structures (residues 37–61 in eachmonomer of the homodimeric protein molecule) was systemati-cally varied. These analogue enzymes were further studied byX-ray crystallography, NMR relaxation, and pulse-EPR methods,in conjunction with molecular dynamics simulations. We show thatconformational isomerization in the flaps is correlated with struc-tural reorganization of residues in the active site, and that it ispreorganization of the active site that is a rate-limiting factor incatalysis.
enzyme catalysis ∣ HIV protease ∣ protein dynamics ∣ protein NMR
The dynamics of the HIV-1 protease protein molecule haveattracted a great deal of attention because of the critical role
that this enzyme plays in the maturation of the AIDS virus (1).NMR studies of the HIV-1 protease have identified regions ofthe protein molecule with enhanced mobility (2, 3), and attemptshave been made with the help of molecular dynamics (MD)simulations to utilize knowledge of the dynamic properties of theHIV-1 protease protein molecule for improved drug design andto rationalize drug resistant mutations (4–7). Particularly impor-tant are two highly mobile regions in the HIV-1 protease mole-cule, the pair of so-called “flaps” (Fig. 1A), which are seen toclose over the substrate or substrate-derived inhibitor in the crys-tal state (8–11). The conventional view is that the role of the flapsis to provide a gating mechanism for substrate binding and forproduct release, and to help to orient the substrate within theenzyme-substrate complex in a conformation suitable for catalysis(2–7). In the work reported here, we have reinvestigated the roleof the HIV-1 protease flaps in enzyme catalysis by employingadvanced biophysical methods in conjunction with a series ofunique analogue enzymes prepared by total chemical synthesis.
ResultsDesign and Chemical Synthesis of Flap Analogues of HIV-1 Protease. InX-ray structures of the HIV-1 protease complexed with a peptidesubstrate or substrate-derived inhibitor, the tips of the flaps adopttwo different β-turn conformations, type I or type II (Fig. 1 Band C) (8, 9). The differences between these two flap structuresare most pronounced for the highly conserved residues Gly51and Gly51′ in the middle of the β-turns, where the Gly residuein one flap adopts an L-amino acid conformation (φ ¼ −103°,ψ ¼ −3°) whereas the Gly residue in the other flap has aD-aminoacid conformation (φ ¼ 97°, ψ ¼ −13°). Previous NMR studies ofHIV-1 protease enzyme/inhibitor complexes have demonstratedthe isomerization of these two different β-turn conformations,on both subnanosecond and microsecond-to-millisecond timescales (2, 3). To evaluate whether such isomerizations have anyfunctional significance, we introduced conformational constraints
by preparing a series of protein analogues in which residuesGly51/Gly51′ were substituted with L-Ala, D-Ala, or Aib (α-ami-noisobutyric acid) in one or both of the flaps of the homodimericHIV-1 protease protein molecule. In addition, we synthesizedchemical analogues of the HIV-1 protease in which individualα-amino acid residues 50–52 at the tips of both flaps were system-atically substituted with isosteric α-hydroxy acids, replacing theamide N–H of the peptide bond with an ester moiety.
To enable asymmetric incorporation of different α-amino acidsor α-hydroxy acids in the flaps of a single HIV-1 protease enzymemolecule, the two 99-residue monomers were covalently joinedthrough a short linker (five amino acids). The asymmetric analo-gues were prepared by a fully convergent total synthesis of the203-amino acid residue protein, based on a combination of nativechemical ligation (12) and kinetically controlled ligation (13),giving a “covalent dimer” HIV-1 protease enzyme molecule withfull catalytic activity (14). The wild-type enzyme and nine enzymeanalogues prepared in this way are listed in Table 1.
Steady-State Kinetics of HIV-1 Protease Flap Analogues. Steady-stateproteolysis kinetics of the analogue enzymes were measured withthe fluorogenic substrate Abz-NF6*. The results are shown inTable 1. The values of kcat∕Km varied over a wide range, wherethe least active enzyme was analogue 8 containing Aib51/Aib51′(approximately 1,000 times lower activity than wild-type enzyme1). Interestingly, the introduction of L-Ala or D-Ala in eithersymmetric or asymmetric fashion had distinct effects on catalyticefficiencies as measured by steady-state kinetics. Substitution ofboth Gly51/51′ with L-Ala led to analogue 5 in which proteolyticactivity was found to be approximately 10 times lower than forwild-type enzyme 1, whereas insertion ofD-Ala at position 51∕510in both flaps (enzyme 7) gave an enzyme that had approximately100 times lower activity. Substituting Gly at position 51 in oneflap only with L-Ala (enzyme 3) enhanced the measured Kmalmost two times with no effect on kcat, whereas substituting withD-Ala in one flap only (enzyme 4) led both to a 2-fold higher Kmand an approximately 4-fold reduction of kcat. In contrast to theseresults, the asymmetric enzyme molecule 2 with L-Ala51 in oneflap andD-Ala51′ in the other flap had proteolytic activity similarto the wild-type enzyme 1, suggesting that the asymmetric confor-mations of the Gly51 and Gly51′ residues seen in the X-ray struc-ture of the wild-type enzyme may be functionally relevant.
Author contributions: V.Y.T. and S.B.H.K. designed research; V.Y.T., H.R., and D.H.performed research; M.T., W.M.W., and E.P. contributed new reagents/analytic tools;V.Y.T., H.R., D.H., and S.B.H.K. analyzed data; and V.Y.T. and S.B.H.K. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
Data deposition: The X-ray structures have been deposited in the Protein Data Bank,www.pdb.org (PDB ID codes 3HAU, 3FSM, 3KA2, 3HLO, 3HAW, 3IAW, 3HBO, 3HDK,3NXE, 3NWQ, 3NXN, 3NWX, and 3NYG)1To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1111202108/-/DCSupplemental.
20982–20987 ∣ PNAS ∣ December 27, 2011 ∣ vol. 108 ∣ no. 52 www.pnas.org/cgi/doi/10.1073/pnas.1111202108

High-Resolution X-Ray Structures of Chemical Analogues of HIV-1Protease.High-resolution X-ray structures of the HIV-1 proteaseprotein molecule and its chemical analogues were obtained incomplexes with the reduced isostere containing substrate-derivedinhibitor MVT-101 (SI Appendix, Tables S3–S7). The enzymeprotein crystals were all found to be isomorphous (space groupP212121), with the geometry and H-bond network at the protein-inhibitor interface highly preserved for different enzyme analo-gues (see SI Appendix, Table S2). In most of the structures,the flaps were observed to close over the inhibitor, adoptingasymmetric βI/βII turn conformations at the flap tips as seen inthe wild-type HIV-1 protease. Exceptions were the [L-Ala51/51′]HIV-1 protease where both flaps close over the MVT-101 inhi-bitor in the symmetric βII/βII conformation, and the [Gly51/Aib51′]HIV-1 protease-MVT-101 complex, where the asym-metric βI/βII conformation is switched to βII/βI relative to theN-to-C orientation of the MVT-101 substrate-derived inhibitor(SI Appendix, Fig. S11). In addition, in the case of the [Aib51/
51′]HIV-1 protease complex with MVT-101, one flap does notfully close over the inhibitor. It was not possible to rationalizethe very different enzyme kinetics observed for these analogueproteins by looking only at the structures of the flaps.
Interestingly, despite identical growth conditions (pH 6.0) andisomorphous crystal forms for all the protein-inhibitor complexes,the “O—O” distances for the carboxyl oxygens distal from theinhibitor in the catalytic residues Asp25 and Asp25′ were foundto vary over the range 2.34–2.55 Å for different chemical analo-gues (SI Appendix, Table S3). These O—O distances are signifi-cantly shorter than the sum of the van der Waal’s contact radii(approximately 3.0–3.1 Å), indicating that a proton must bepresent between the two oxygens in all these complexes. At thesame time, these distances are too short in order to accommodatetwo protons (i.e., the diprotonated state), as illustrated in thecrystal structure of the diprotonated wild-type HIV-1 proteasewith DMP-323 inhibitor (15) where the corresponding distancewas found to be 3.4 Å (16).
Interflap Distance Measurements. In order to measure how substi-tutions at position Gly51 affect large-scale motions of the flaps,we incorporated nitroxide spin labels at amino acid positions55∕550 and measured the nitroxide-to-nitroxide distance distribu-tion profiles for symmetric analogues using the pulse-EPR doubleelectron–electron resonance (DEER) method (Fig. 2 A and B)(17). The experiments were performed with approximately 45 μMsolutions of unliganded enzymes, frozen in the glass capillary to55 K with helium gas, by recording the dipolar spin-echo evolu-tion responses and converting the obtained data into distancesnapshots of all conformers present in a given sample at themoment of flash-freezing (18). As was reported earlier (18), threemajor conformers were observed for all samples—so-called“closed,” “semiopen,” and “open.” We found significant differ-ences in the nitroxide-to-nitroxide distance distributions for theD-Ala51/51′ symmetric analogue, which became much narrower,with the semiopen conformer being more populated here thanin all other analogues. This observed predominance of the semi-open conformer for the D-Ala51/51′ symmetric analogue is inagreement with the conformations previously observed in thecrystal structure of the unliganded form of HIV-1 protease, whereboth Gly51 and Gly51′ have a D-amino acid backbone confor-mation and the flaps adopt a semiopen conformation and aresymmetrically related via a twofold axis (19).
Protein Dynamics on the Subnanosecond Time Scale. In order tocharacterize how substitutions at position 51∕510 of the HIV-1protease affect the conformational properties of the flaps in theseunique enzyme analogues, we performed NMR studies on theunliganded enzymes using site-specifically 15N-labeled protein
Fig. 1. Structural features of HIV-1 protease. (A) Homodimeric catalyticallyactive form of HIV-1 protease (2 × 99 amino acids) complexed with the pep-tidomimetic reduced isostere-containing MVT-101 inhibitor. (B) The β-turnstructures (residues 37–61) from each monomer, known as “flaps,” donatehydrogen bonds to the substrate (or inhibitor) through a structural water(water 301) molecule (in green). (C) Flaps with β-turn type I (Upper Left)and β-turn type II (Lower Right) conformations depicted separately for clarity.Residue Gly51 has D-amino acid conformation in the β-turn type I structureand L-amino acid conformation in the β-turn type II structure. (D) Flap X-raystructures in chemically synthesized [L-Ala51; D-Ala51′] covalent dimer HIV-1protease molecule; 2Fo − Fc electron density contoured in blue at 1.5σ levelfor the 1.6-Å resolution crystal structure [Protein Data Bank (PDB) ID code3FSM; see SI Appendix].
Table 1. Steady-state kinetics of proteolysis by chemically synthesized HIV-1 protease and its analogues
Flap Flap′ kcat, s−1; mean� SD Km, μM; mean� SD kcat∕Km; s−1 μM−1 PDB ID code
1* Gly51 Gly51 23.4 ± 0.4 25.1 ± 1.2 0.93 3HAU2† L-Ala51 D-Ala51 17.6 ± 0.3 26.1 ± 1.4 0.67 3FSM3 L-Ala51 Gly51 22.2 ± 0.6 47.4 ± 2.9 0.47 3KA24 Gly51 D-Ala51 6.2 ± 0.2 43.8 ± 2.8 0.14 3HLO5 L-Ala51 L-Ala51 3.7 ± 0.1 50.1 ± 2.6 0.073 3HAW6 Gly51 Aib51 4.04 104.6 0.042 3IAW7 D-Ala51 D-Ala51 4.9 434.2 0.011 3HBO8 Aib51 Aib51 n∕a n∕a 0.001 3HDK9 Ile50 O-Ile50 4.2 ± 0.2 41.2 ± 3.2 0.1 3NXE10 O-Ile50 O-Ile50 0.94 99.6 0.009 3NWQ
The PDB ID code of the corresponding X-ray structure with MVT-101 inhibitor is included in the rightmost column.*The wild-type enzyme 1 was prepared both as a homodimer and as a covalent dimer, and the catalytic properties were found to be identical withinexperimental uncertainty.
†In a control experiment, the 99-residue L-Ala51 HIV-1 protease polypeptide was folded by dialysis in the presence of an equimolar amount of the 99-residue D-Ala51 HIV-1 protease polypeptide. We observed kcat 9.1 s−1 (one half of the value for analogue 2) and Km 25.6 μM, which corresponded to theproperties of the statistical 0.5-molar ratio of (L-Ala51, D-Ala51′) heterodimer enzyme in the mixture; this result precluded the possibility that eitherdimerization equilibrium or the five-amino acid interdomain linker peptide sequence inserted in covalent dimers affected the observed enzyme kinetics.
Torbeev et al. PNAS ∣ December 27, 2011 ∣ vol. 108 ∣ no. 52 ∣ 20983
BIOCH
EMISTR
YCH
EMISTR
Y

molecules. Key residues in the flaps were site-specifically 15N-la-beled; in addition, the residue Gly40 in the “elbow” regions (seeFig. 1A) and residues Asp25 and Gly27 in the catalytic site werealso 15N-labeled (see Fig. 1 A and B and SI Appendix). For theseNMR studies, all measurements were performed with symmetri-cally substituted chemical analogues (homodimers) in order tosimplify interpretation of the data (Fig. 2C).
Order parameters (S2) for the protein backbone amides werederived from model-free analysis of measured R1 (spin-lattice re-laxation rate), R2 (spin-spin relaxation rate), and heteronuclear1H-15N NOE (nuclear Overhauser effect) values (20). The orderparameter (S2) represents the degree of spatial restriction ofinternal fluctuation of the amide bond on the subnanosecondtime scale, ranging from 0 (completely unrestricted motions) to1 (completely rigid). S2 values measured for unliganded enzymeswere found to be significantly higher for the D-Ala51-containingflap (“D-flap”) and Aib51-containing flap (“Aib-flap”) than forthe L-Ala51-containing flap (“L-flap”) (Fig. 2C, Inset). The high-er order parameters for the tips of the flaps (residues 48–52) ofD-flap and Aib-flap enzyme molecules indicate greater rigidity onthe subnanosecond time scale. Remarkably, the L-Ala51-contain-ing flap enzyme molecule had S2 values and hence subnanose-cond flexibility comparable to those of the corresponding wild-type Gly51-containing flaps (21). This is consistent with ourobservation that, of the symmetrically substituted enzyme analo-gues, the L-Ala51/51′ homodimer had proteolytic activity closestto that of the wild-type enzyme (Table 1; see entries 1, 5, 7, and 8,respectively).
Protein Dynamics on the Microsecond–Millisecond Time Scale. Toelucidate the dynamics of the enzyme analogues in the microse-cond–millisecond time regime, 15N Carr–Purcell–Meiboom–Gill
(CPMG) relaxation dispersion measurements were performed onthe unliganded enzyme analogues (Fig. 2D and SI Appendix) (22).In the three symmetric analogues containing L-flaps, D-flaps, orAib-flaps, we observed a systematic decrease in mobility on themicrosecond–millisecond time scale; the previously observedhigher subnanosecond mobility of the L-flap enzyme analogue(above) was also reflected in faster microsecond–millisecondregime dynamics, and a greater subnanosecond rigidity corre-sponding to slower mobility on the microsecond–millisecond timescale was observed for theD-flap and Aib-flap enzyme analogues.In the L-flap and D-flap enzyme analogues, all labeled residuesshowed coherently matched chemical exchange rates in theseNMR measurements; after fitting the data on a residue-by-residue basis, they were further fitted globally with a three-siteexchange model (23), yielding kex (fast) 2;350� 100 s−1 (mean�SD), kex (slow) 38� 2 s−1 (mean� SD) for the L-flap enzyme,and kex (fast) 1;380� 60 s−1 (mean� SD), kex (slow) 80� 5 s−1(mean� SD) for the D-flap enzyme (see SI Appendix, Figs. S12and S13, Table S12, and Scheme S3). In the Aib-flap enzymeanalogue, however, it was not possible to perform global fittingbecause of substantial noncoherent variations in exchange ratesfor different residues, indicating either greater motional com-plexity or a shift in the principal isomerization rate constant forthe flaps as global structures. In fact, several residues actuallylost their R2 dispersion in the Aib-flap enzyme analogue (seeSI Appendix, Figs. S12 and S13).
The most remarkable differences observed for this seriesof flap analogues of the HIV-1 protease involved the active-site residues. Our NMR measurements on the flap analogue-containing unliganded enzyme molecules showed that the micro-second–millisecond dynamics of the catalytic residues Asp25/25′as well as the nearby Gly27/27′ residues were correlated with the
Fig. 2. Dynamic properties of chemically synthesized Gly51 analogues of HIV-1 protease. (A) “Snapshots” of the conformational states (open, semiopen, andclosed overlaid in B) of HIV-1 protease chemical analogues labeled with nitroxide-spin label, measured at 55 K using pulse-EPR spectroscopy. Populations ofconformers are depicted for Gly51, L-Ala51, D-Ala51 and Aib51 homodimers in green, blue, red, and black, respectively. In the case of the D-Ala51 homodimeranalogue, the semiopen conformer is overpopulated. (C) 15N-HSQC spectra overlaid for L-Ala51, D-Ala51, and Aib51 homodimers with the same color coding asinA. Peaks for the wild-type HIV-1 protease (in green) for corresponding residues were reconstructed from a previous study (21). Order parameters S2, obtainedby model-free analysis of R1, R2 and 1H-15N NOE values, versus residue number are depicted as Inset. (D) CPMG 15N relaxation dispersion data for catalyticresidue Asp25 in three HIV-1 protease symmetric homodimers obtained at twomagnetic fields (600 and 900MHz). Remarkably, the chemical exchange rates forthe three analogue enzyme molecules are drastically different, and are correlated with the dynamic properties of the flaps and with the catalytic rates forproteolysis (Table 1; entries 5, 7, and 8).
20984 ∣ www.pnas.org/cgi/doi/10.1073/pnas.1111202108 Torbeev et al.

dynamics of the flaps. For the L-flap enzyme analogue, bothGly27 and Asp25 in the catalytic region have microsecond–milli-second chemical exchange constants (kex 2;325� 200 s−1,mean� SD) similar to the fast exchange constants observedfor the flap region; for the D-flap enzyme analogue, the slowermicrosecond–millisecond dynamics observed in the flap werefound to be correlated to the reduced microsecond–millisecondmobility of the Asp25 and Gly27 residues (kex 420� 70 s−1,mean� SD). And, for the Aib-flap-containing enzyme, boththe catalytic Asp25 and residue Gly27 were found to be lackingR2 dispersion as was the case for some residues in the flaps inthis enzyme analogue (Fig. 2D, Bottom and SI Appendix, Figs. S12and S13), meaning that the time scale window probed with ourexperimental approach was not suitable for this particularchemical analogue.
Molecular Basis of a Protein Dynamics Feedback Catalytic Mechanism.To explore the role of flap backbone hydrogen bonds in catalysis,we synthesized both a monoester [Ile50, O-Ile50′]HIV-1 protease(enzyme 9) and the corresponding diester [O-Ile50,O-Ile50′]HIV-1 protease (enzyme 10), and obtained high-resolution X-raystructures for their complexes with substrate-based inhibitors[for catalytic activities see Table 1; values obtained were generallycomparable with previous experimental data (24, 25)]. Interest-ingly, in the X-ray structures of the ester analogue enzymescomplexed with the reduced isostere MVT-101 inhibitor, electrondensity for the nonnucleophilic water molecule (water 301) brid-ging the flaps of the enzyme and the carbonyls of the inhibitor wasfound to be much less intense (Fig. 3A); moreover, for the com-plex of diester enzyme 10 with MVT-101, we could not locateclear electron density for water 301 (Fig. 3B). This was to be ex-pected, because at least one hydrogen-bond-donating amide—NH—is replaced by an ester oxygen atom in these enzyme analogues,thus knocking out the possibility for a water 301 H2O—HN-Ile50hydrogen bond. However, the structure of the MVT-101 inhibitorin these complexes did not deviate significantly from the complexwith the wild-type enzyme molecule (for enzyme 9, rmsd backbone0.13 Å, rmsd all atoms 0.44 Å; for enzyme 10, rmsd backbone0.13 Å, rmsd all atoms 0.84 Å), and both flaps adopted a closedconformation rather similar to that found in the wild-type HIV-1protease complexed with the same inhibitor.
In the X-ray structures of these ester analogue enzyme mole-cules with the ketomethylene isostere KVS-1 inhibitor (whichclosely mimics a tetrahedral intermediate) (9), the occupancy ofwater 301 was not so drastically affected as in the case of com-plexes with MVT-101 inhibitor (Fig. 3 C and D). In the complexof the monoester enzyme 9 with KVS-1 inhibitor, water 301occupancy (see Fig. 3C) was comparable to that observed for thewild-type HIV-1 protease in the same complex (9). However, inthe case of the diester enzyme 10, the H-bonding network wassignificantly perturbed, and an additional water molecule hasbeen located at the flaps-inhibitor interface (Fig. 3D).
These observations suggest to us that the hydrogen-bondingnetwork at the flaps—inhibitor interface mediated by the non-nucleophilic water molecule may be responsible for a proteindynamics feedback mechanism in HIV-1 protease catalysis. In thecase of the ester analogues, the mediating role of the amide hy-drogen bond(s) present in the native NH-Ile50 is eliminated, thusdestabilizing the rigid closed flaps conformational state, whichthen would lead to higher flexibility of the corresponding flapstructure. Thus, the dynamics of the catalytic site could neveradjust to achieve the structure necessary for efficient electrostaticstabilization of the transition state of the catalyzed reaction ashappens in the wild-type enzyme. In the wild-type HIV-1 pro-tease, such stabilization is achieved by hydrogen bonds mediatedby water 301 that rigidify the flap structures, lock the substratein a productive conformation, and contribute to preorganizationof the structure of the catalytic site. The flaps and water 301 serve
to sense the progress of the proteolytic reaction through hydro-gen bonding to two carbonyl groups of the substrate (from theamide bond preceding and the amide bond following the scissileamide bond in the polypeptide substrate sequence, respectively),and to thus properly attenuate the dynamics of the catalytic as-partic acid residues. The distance between the aforementionedsubstrate carbonyl groups increases with the progress of theamide bond cleavage (formation of tetrahedral intermediate,breakdown of tetrahedral intermediate concomitant with productformation and stepwise protein/products dissociation) as wasshown by recent crystallographic work (10, 11).
The Active-Site Structure of the HIV-1 Protease Is Preorganized forCatalysis and Is Asymmetric. In silico experiments were per-formed to get a better picture of how the dynamics of the flapstructures correlate with the state of the catalytic residues in theHIV-1 protease-substrate complex. All-atom explicit water MDsimulations of symmetric L-flap and D-flap enzyme analoguesgave results in agreement with interflap distance distributionsobserved by pulse-EPR measurements (SI Appendix, Fig. S15).In the active site of these two analogue enzymes, MD simulationsshowed a symmetric cyclic structure consisting of two catalyticAsp25 and Asp25′ residues and the nucleophilic water moleculeas the most predominant conformer (Fig. 4B). Because protontransfer between the two catalytic aspartates Asp25 and Asp25′cannot be treated adequately by the classical molecular me-chanics approach employed in the MD simulation, for the case ofthe asymmetric [L-Ala51; D-Ala51] heterodimer, we performedcalculations on two boundary states, with either the L-flap-con-taining domain Asp25 or the D-flap-containing domain Asp25′
Fig. 3. X-ray structures of HIV-1 protease and its ester chemical analogues 9and 10 complexed with substrate-based inhibitors. (A and B) In complexes ofenzymes 9 and 10 with MVT-101 inhibitor, the electron density for structuralwater 301 molecule is significantly diffused—most strikingly in the enzyme10 complex (see B), where we could not locate clear density for a watermolecule at the flaps/inhibitor interface. (C and D) Interestingly, with the me-chanistically based hydrated ketomethylene inhibitor KVS-1 (9), water 301 iswell-populated in structures of enzymes 9 and 10. Moreover, in the complexof enzyme 10, there is second structural water molecule. [In all structures,side chains are deleted for clarity except the residues of interest. The2Fo − Fcelectron density map was contoured at 1.0 σ level (in magenta) forselected flap residues and water molecules, and at a level of 3σ (in green) forresidues Asp25 and Asp25′. Distances between distal oxygens of two catalyticAsp25 and Asp25′ and PDB codes are specified for each structure].
Torbeev et al. PNAS ∣ December 27, 2011 ∣ vol. 108 ∣ no. 52 ∣ 20985
BIOCH
EMISTR
YCH
EMISTR
Y

ionized and the other aspartate in the same molecule protonated[note: the monoprotonated state for the two catalytic aspartatesis invoked for general acid-general base catalysis (26, 27)]. Forthe case in which the L-flap-containing domain has a chargedAsp25 carboxylate andD-flap-containing domain Asp25′ has a pro-tonated side chain, we observed the same cyclic hydrogen-bondedstructure that we found as the most populous state in symmetrichomodimers. Strikingly, if we reversed protonation states (i.e., withthe L-flap-containing domain Asp25 now being protonated andthe D-flap-containing domain Asp25′ being charged), MD simula-tions showed an enhanced population (approximately 100-fold) ofthe structure composed of Asp25, Asp25′ residues, and the nucleo-philic water molecule hydrogen-bonded asymmetrically and with ageometry preorganized for catalysis (Fig. 4A) (28).
The computational prediction of a greater concentrationof conformers preorganized for catalysis being present in theunliganded [L-Ala51, D-Ala51′] heterodimeric enzyme moleculewas supported by surface plasmon resonance (SPR) bindingexperiments (see SI Appendix, Fig. S16). In the SPR measure-ments on the HIV-1 protease flap analogues, using a reducedisostere inhibitor to mimic the earlier transition state of the en-zyme-catalyzed proteolysis, the [L-Ala51; D-Ala51′] asymmetricenzyme analogue showed tighter binding affinity toward inhibitorthan any other studied enzyme analogues, including the wild-typeHIV-1 protease (Kd value of 100 nM, four times lower than theKd value of 420 nM observed for the wild-type enzyme).
DiscussionThe accepted chemical mechanism for the aspartyl proteasesinvolves general acid–general base catalysis, where one catalyticaspartate side chain carboxylate (COO−) acts as a general baseto remove a proton from the water molecule nucleophile, whileanother aspartic acid side chain carboxyl (COOH) general aciddonates a proton to the carbonyl oxygen atom of the scissile pep-tide bond (26, 27). Our results are in agreement with the generalacid–general base mechanism, with a nucleophilic water moleculepreorganized for catalysis in the asymmetric environment ofthe two aspartates, one being protonated and one being ionized(Fig. 4C). Thus, our data suggest that in the case of the HIV-1protease, the so-called “L domain” (which contains the L-Gly51conformation flap) is more mobile and bears the general acid at
the Asp25 COOH, whereas the less mobile “D domain” (contain-ing the D-Gly51 flap) is more rigid and bears the general baseAsp25 COO− in the active site.
We recently reported that flap structures in HIV-1 protease—complexed with three different inhibitors that mimic, respec-tively, an early transition state, the tetrahedral intermediate, anda late transition state—display significantly different equilibriumpopulations of conformers; the flaps are least mobile at the ear-lier stages of the reaction, and attain more flexibility in the courseof the reaction en route to product release (18). The resultsobtained from the combination of experiments used in the workreported here show that the glycine residue at position 51 in eachmonomer serves as a surrogate for both the L- andD-amino acidsrequired at that position in each domain of the homodimericHIV-1 protease molecule preorganized for catalysis. Our datasuggest that catalysis is not rate-limited by opening and closingevents of flaps, but rather by the emergence of catalytically pre-organized asymmetric β-turn type I/β-turn type II conformers inthe Gly51/51′-containing wild-type HIV-1 protease. If substratebinding/product release were rate-limiting, we would expect the[L-Ala51, D-Ala51′] covalent dimer enzyme to posses catalyticefficiency lower than the more flexible [L-Ala51, L-Ala51′] homo-dimer and greater than more rigid [D-Ala51, D-Ala51′] homo-dimer. However, as illustrated in Fig. 5, the [L-Ala51, D-Ala51′]heterodimer enzyme is approximately 10 times and 100 times, re-spectively, more efficient as a catalyst than the above mentionedtwo homodimer enzymes (see also Table 1, entries 2, 5, and 7).
These results provide strong experimental support for achemical mechanism in HIV-1 protease in which the contributionof both flaps to catalysis is critical. First, flap β-turn isomerizationattenuates the dynamics of the whole protein molecule, resultingin correlation of the dynamics of the flaps and the catalyticresidues; then, hydrogen-bonding interactions of HN-Ile50 andHN-Ile50′ at the tips of the flaps to the carbonyls of the substrateon either side of the scissile bond are mediated by a (nonnucleo-philic) water molecule and lock the substrate and enzyme mole-cule in a productive catalytic conformation. In contrast to theHIV-1 protease and related viral aspartyl proteases, the cell-encoded aspartyl protease enzyme molecules have a single poly-peptide chain that folds to form two domains, and a single flapthat in crystal structures is seen to form direct hydrogen bonds to
Fig. 4. Active-site structures revealed by MD simulations, and a scheme for the mechanism of HIV-1 protease catalysis taking into account the dynamics ofthe flaps. (A) An asymmetric structure of catalytic Asp25 and Asp25′ and the nucleophilic water molecule dominated in the MD trajectory for the asymmetric[L-Ala51; D-Ala51′] chemical analogue of HIV-1 protease with the L-domain having a protonated Asp25 and the D-domain having a charged Asp25′ side chain.(B) Symmetric structure for catalytic residues and nucleophilic water was observed as the most populous state in the MD simulation for the symmetric [L-Ala51;L-Ala51′] and [D-Ala51; D-Ala51′] chemical analogues. (C) Scheme for the mechanism of HIV-1 protease catalysis. Asymmetric conformers are preorganized forcatalysis. Conformational isomerizations are depicted by blue arrows, and electron rearrangements are depicted by red arrows. TS1 and TS2 are earlier andlater transition states, respectively, and E.I is the enzyme complex with the tetrahedral intermediate.
20986 ∣ www.pnas.org/cgi/doi/10.1073/pnas.1111202108 Torbeev et al.

the substrate or substrate-derived inhibitor (27, 29). We predictthat the dynamic properties of the two domains in eukaryoticaspartic protease molecules will be distinct from one another andreminiscent of those reported here for the artificial [L-Ala51;D-Ala51′] covalent dimer HIV-1 protease prepared by totalchemical synthesis.
Materials and MethodsThe 99-residue polypeptide chains of symmetric homodimers and the 203-residue polypeptides for asymmetric heterodimers were prepared by totalchemical synthesis, folded by dialysis and assayed as described (9, 14).
X-Ray Crystallography. Crystals were grown at 20 °C by the hanging dropvapor diffusion method within 1–30 d and were frozen in liquid nitrogenusing mineral oil as cryoprotectant. Data collection was performed at 100 Kat the Advanced Photon Source, Argonne National Laboratory.
Pulse-EPR Experiments. Dipolar spin-echo evolution measurements werecarried out using a constant time version of the four-pulse DEER sequenceat a temperature of 55 K. Data were processed and analyzed usingDeerAnalysis 2008 software.
NMR Experiments. R1, R2, and heteronuclear NOE values measured usingstandard pulse sequences on a 600-MHz spectrometer were used to calculateS2 order parameters. Relaxation dispersion spectra were recorded on 600-and 900-MHz spectrometers using room-temperature probes. Dispersioncurves were fitted with either two-state or three-state exchange models.
MD Simulations. All-atom MD simulations were carried out on the symmetric“L-Ala51/51′-flaps” and “D-Ala51/51′ flaps” analogues of the HIV-1 protease,and on the asymmetric “L-Ala51 flap” and “D-Ala51′ flap” analogues, eachfor 300 ns. All of the simulations were carried out in a truncated octahedronperiodic box of explicit water and neutralizing chloride counterions.
ACKNOWLEDGMENTS. We acknowledge D.M. Korzhnev and L.E. Kay (Univer-sity of Toronto, Toronto, ON, Canada) for providing computer software usedfor fitting NMR relaxation-dispersion data. This work was supported by theOffice of Science, Biological and Environmental Research, US Department ofEnergy (DE-FG02-07ER64501 to S.B.H.K.). This study made use of the NationalMagnetic Resonance Facility at Madison, which is supported by NationalInstitute of Health Grants P41RR02301 and P41GM66326. Use of the beam-lines 14BM, 22BM, 23ID, and 24ID of the Advanced Photon Source (ArgonneNational Laboratory, Argonne, IL) was supported by the US Departmentof Energy (W-31-109-Eng-38, DE-AC02-06CH11357), the National CancerInstitute (Y1-CO-1020), the National Institute of General Medical Sciences(Y1-GM-1104), and the National Center for Research Resources at theNational Institutes of Health (RR-15301 and RR007707). D.H. is supportedin part by the National Science Foundation CAREER MCB-0953061 andGeorgia Cancer Coalition. The MD simulations were carried out using theIBM System p5 supercomputer at Georgia State University.
1. Pomerantz RJ, Horn DL (2003) Twenty years of therapy for HIV-1 infection. Nat Med9:867–873.
2. Nicholson LK, et al. (1995) Flexibility and function in HIV-1 protease. Nat Struct Biol2:274–280.
3. Ishima R, Louis JM (2008) A diverse view of protein dynamics from NMR studies ofHIV-1 protease flaps. Proteins 70:1408–1415.
4. Perryman AL, Lin J-H, McCammon JA (2004) HIV-1 protease molecular dynamics ofa wild-type and of the V82F/I84V mutant: Possible contributions to drug resistanceand a potential new target site for drugs. Protein Sci 12:1108–1123.
5. Hornak V, Simmerling C (2007) Targeting structural flexibility in HIV-1 protease inhi-bitor binding. Drug Discov Today 12:132–138.
6. Piana S, Carloni P, Rothlisberger U (2002) Drug resistance in HIV-1 protease: Flexibility-assisted mechanism of compensatory mutations. Protein Sci 11:2393–2402.
7. Chang C-E, Shen T, Trylska J, Tozzini V, McCammon JA (2006) Gated binding of ligandsto HIV-1 protease: Brownian dynamics simulations in a coarse-grained model. BiophysJ 90:3880–3885.
8. Miller M, et al. (1989) Structure of a complex of synthetic HIV-1 protease with asubstrate-based inhibitor at 2.3 Å resolution. Science 246:1149–1152.
9. Torbeev VY, Mandal K, Terechko VA, Kent SBH (2008) Crystal structure of chemicallysynthesized HIV-1 protease and a ketomethylene isostere inhibitor based on the p2/NCcleavage site. Bioorg Med Chem Lett 18:4554–4557.
10. Das A, et al. (2006) Crystal structure of HIV-1 protease in situ product complex andobservation of a low-barrier hydrogen bond between catalytic aspartates. Proc NatlAcad Sci USA 103:18464–18469.
11. Das A, et al. (2010) X-ray snapshot of HIV-1 protease in action: Observation of tetra-hedral intermediate and short ionic hydrogen bond SIHBwith catalytic aspartate. J AmChem Soc 132:6366–6373.
12. Dawson PE, Muir TW, Clark-Lewis I, Kent SBH (1994) Synthesis of proteins by nativechemical ligation. Science 266:776–779.
13. Bang D, Pentelute BL, Kent SBH (2006) Kinetically controlled ligation for the conver-gent chemical synthesis of proteins. Angew Chem Int Ed Engl 45:3985–3988.
14. Torbeev VY, Kent SBH (2007) Convergent chemical synthesis and crystal structure ofa 203 amino acid ‘covalent dimer’ HIV-1 protease enzyme molecule. Angew ChemInt Ed Engl 46:1667–1670.
15. Yamazaki T, et al. (1994) NMR and X-ray evidence that the HIV protease catalyticaspartyl groups are protonated in the complex formed by the protease and a non-peptide cyclic urea-based inhibitor. J Am Chem Soc 116:10791–10792.
16. Lam PY, et al. (1996) Cyclic HIV protease inhibitors: Synthesis, conformational analysis,P2/P2′ structure-activity relationship, and molecular recognition of cyclic ureas. J MedChem 39:3514–3525.
17. Pannier M, Veit S, Godt A, Jeschke G, Spiess HW (2000) Dead-time free measurementof dipole-dipole interactions between electron spins. J Magn Reson 142:331–340.
18. Torbeev VY, et al. (2009) Dynamics of “flap” structures in three HIV-1 protease/inhibitor complexes probed by total chemical synthesis and pulse-EPR spectroscopy.J Am Chem Soc 131:884–885.
19. Wlodawer A, et al. (1989) Conserved folding in retroviral proteases: Crystal structureof a synthetic HIV-1 protease. Science 245:616–621.
20. Jarymowycz VA, Stone MJ (2006) Fast time scale dynamics of protein backbones:NMR relaxation methods, applications, and functional consequences. Chem Rev106:1624–1671.
21. Freedberg DI, et al. (2002) Rapid structural fluctuations of the free HIV proteaseflaps in solution: Relationship to crystal structures and comparison with predictionsof dynamic calculations. Protein Sci 11:221–232.
22. Palmer AG, Kroenke CD, Loria JP (2001) Nuclear magnetic resonance methodsfor quantifying microsecond-to-millisecond motions in biological macromolecules.Methods Enzymol 339:204–238.
23. Korzhnev DM, et al. (2004) Low-populated holding intermediates of Fyn SH3characterized by relaxation dispersion NMR. Nature 430:586–590.
24. Baca M, Kent SBH (1993) Catalytic contribution of flap-substrate hydrogen bondsin HIV-1 protease explored by chemical synthesis. Proc Natl Acad Sci USA90:11638–11642.
25. Baca M, Kent SBH (2000) Protein backbone engineering through total chemicalsynthesis: New insight into the mechanism of HIV-1 protease catalysis. Tetrahedron56:9503–9513.
26. Suguna K, Padlan EA, Smith CW, Carlson WD, Davies DR (1987) Binding of a reducedpeptide inhibitor to the aspartic proteinase from Rhizopus chinensis: Implications for amechanism of action. Proc Natl Acad Sci USA 84:7009–7013.
27. Davies DR (1990) The structure and function of the aspartic proteinases. Annu RevBiophys Biophys Chem 19:189–215.
28. Piana S, Bucher D, Carloni P, Rothlisberger U (2004) Reaction mechanism of HIV-1protease by hybrid Car-Parrinello/classical MD simulations. J Phys Chem B108:11139–11149.
29. Dunn BM (2002) Structure and mechanism of the pepsin-like family of asparticpeptidases. Chem Rev 102:4431–4458.
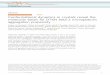
Fig. 5. “Activity–flexibility” relationship for a series of chemical analoguesof HIV-1 protease. The “activity” axis contains relative values of kcat andkcat∕Km for enzyme analogues 1–8 (see Table 1) normalized to those ofthe wild-type enzyme 1. The “flexibility” axis is built based on the assumptionthat the flap structure of the enzyme molecule containing Gly at position51 is most flexible, followed by molecules containing L-Ala, D-Ala, andAib (α-aminoisobutyric acid) replacing Gly. Most of the data follow a generaltrend where higher flexibility leads to higher activity. The exception is en-zyme analogue 2 with one flap containing L-Ala51 and another D-Ala51′.
Torbeev et al. PNAS ∣ December 27, 2011 ∣ vol. 108 ∣ no. 52 ∣ 20987
BIOCH
EMISTR
YCH
EMISTR
Y