Embed Size (px)
Citation preview

Protein ChemistryBasics
• Protein function
• Protein structure– Primary
• Amino acids
• Linkage
• Protein conformation framework– Dihedral angles– Ramachandran plots
• Sequence similarity and variation

Protein Function in Cell
1. Enzymes • Catalyze biological reactions
2. Structural role• Cell wall
• Cell membrane
• Cytoplasm

Protein Structure

Protein Structure
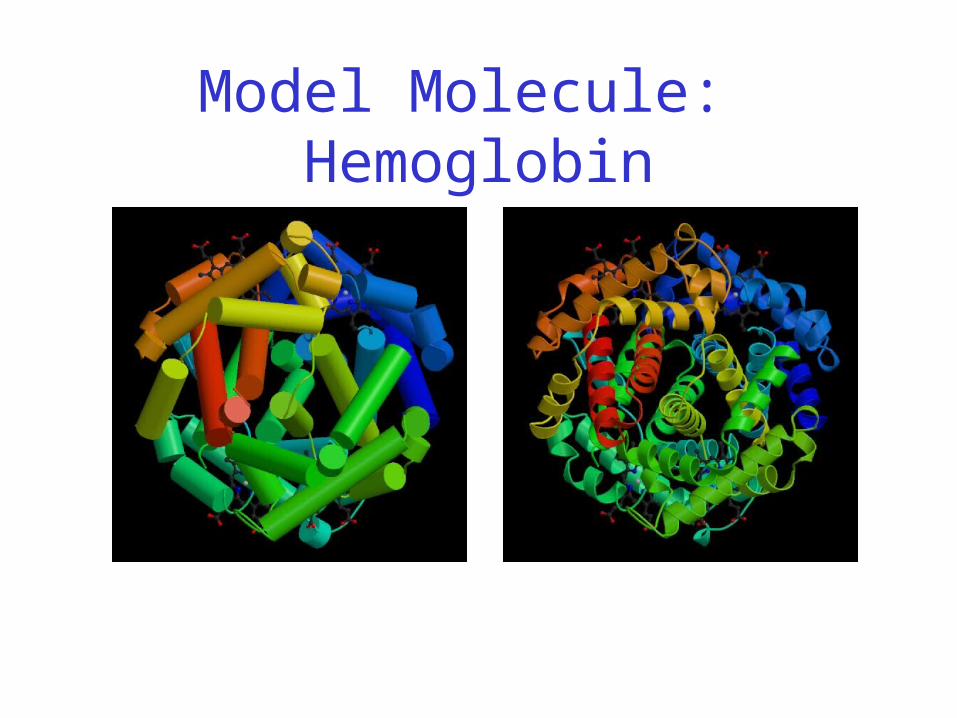
Model Molecule: Hemoglobin

Hemoglobin: Background
• Protein in red blood cells

Red Blood Cell (Erythrocyte)

Hemoglobin: Background
• Protein in red blood cells
• Composed of four subunits, each containing a heme group: a ring-like structure with a central iron atom that binds oxygen

Heme Groups in Hemoglobin

Hemoglobin: Background
• Protein in red blood cells
• Composed of four subunits, each containing a heme group: a ring-like structure with a central iron atom that binds oxygen
• Picks up oxygen in lungs, releases it in peripheral tissues (e.g. muscles)

Hemoglobin – Quaternary Structure
Two alpha subunits and two beta subunits(141 AA per alpha, 146 AA per beta)
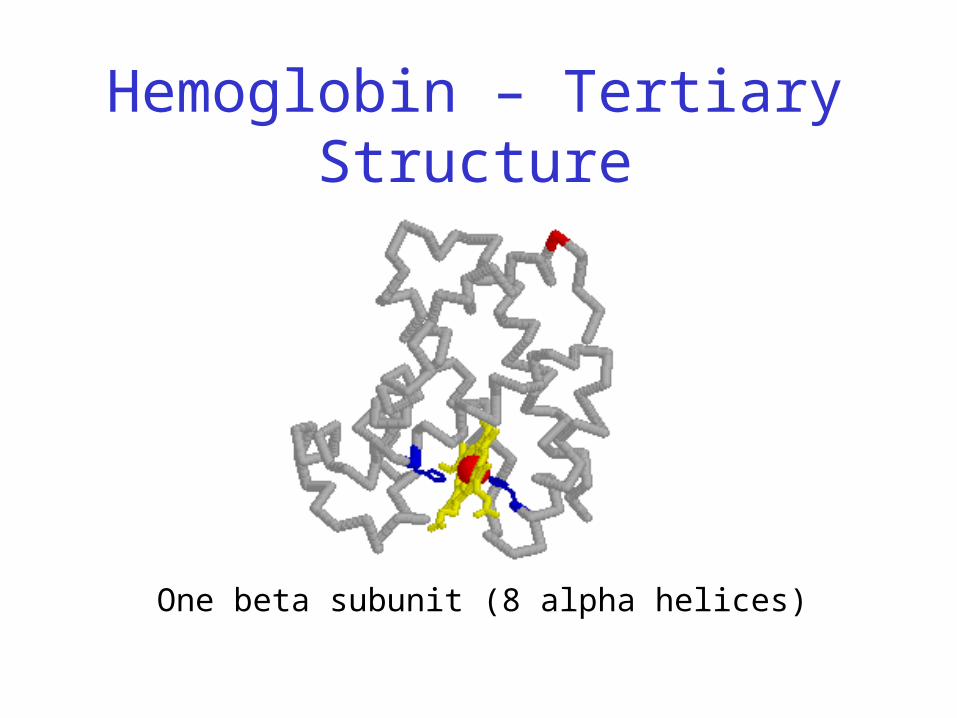
Hemoglobin – Tertiary Structure
One beta subunit (8 alpha helices)

Hemoglobin – Secondary Structure
alpha helix

Xin Zhan CS 882 course project 14
β-Hairpin Motif
• Simplest protein motif involving two beta strands [from Wikipedia]
– adjacent in primary sequence
– antiparallel
– linked by a short loop
• As isolated ribbon or part of beta sheet
• a special case of a turn
– direction of protein backbone reverses
– flanking secondary structure elements interact (hydrogen bonds)

Xin Zhan CS 882 course project 15
Types of Turns
• β-turn (most common)– donor and acceptor residues of hydrogen bonds are separated by 3
residues (i i +3 H-bonding)• δ-turn
– i i +1 H-bonding• γ-turn
– i i +2 H-bonding• α-turn
– i i +4 H-bonding• π-turn
– i i +5 H-bonding• ω-loop
– a longer loop with no internal hydrogen bonding

Structure Stabilizing Interactions
• Noncovalent– Van der Waals forces (transient, weak electrical
attraction of one atom for another)– Hydrophobic (clustering of nonpolar groups)– Hydrogen bonding

Hydrogen Bonding
• Involves three atoms: – Donor electronegative atom (D)
(Nitrogen or Oxygen in proteins)
– Hydrogen bound to donor (H)– Acceptor electronegative atom (A) in close
proximity
D – H A

D-H Interaction• Polarization due to electron withdrawal from
the hydrogen to D giving D partial negative charge and the H a partial positive charge
• Proximity of the Acceptor A causes further charge separation
D – H Aδ- δ+ δ-

D-H Interaction• Polarization due to electron withdrawal from the
hydrogen to D giving D partial negative charge and the H a partial positive charge
• Proximity of the Acceptor A causes further charge separation
• Result:– Closer approach of A to H– Higher interaction energy than a simple van der Waals
interaction
D – H Aδ- δ+ δ-

Hydrogen BondingAnd Secondary Structure
alpha-helix beta-sheet

Structure Stabilizing Interactions
• Noncovalent– Van der Waals forces (transient, weak electrical
attraction of one atom for another)– Hydrophobic (clustering of nonpolar groups)– Hydrogen bonding
• Covalent– Disulfide bonds
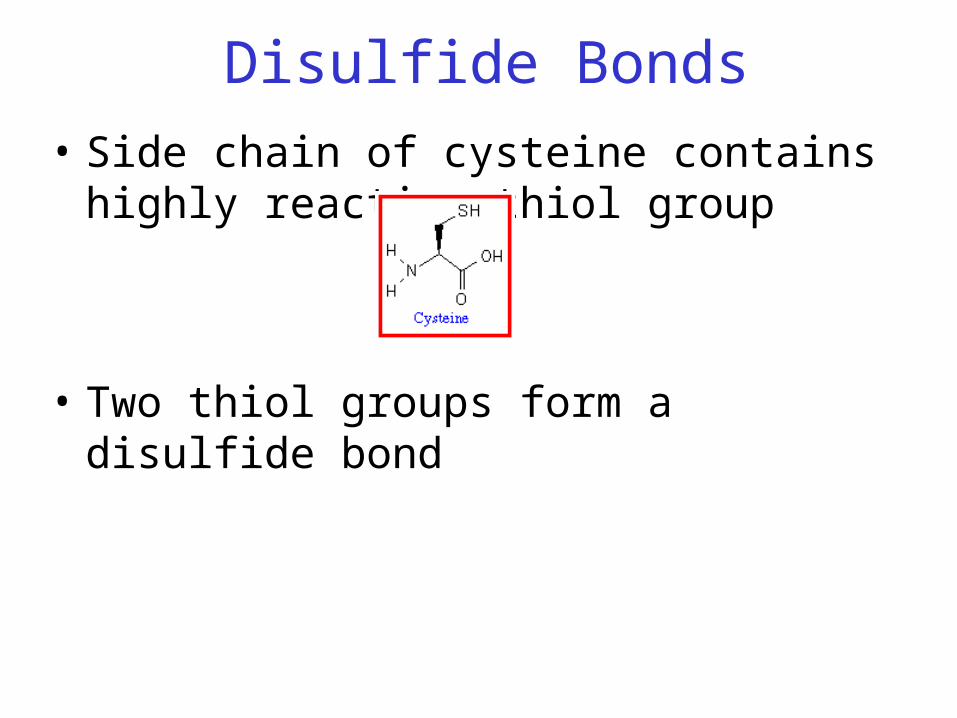
Disulfide Bonds
• Side chain of cysteine contains highly reactive thiol group
• Two thiol groups form a disulfide bond
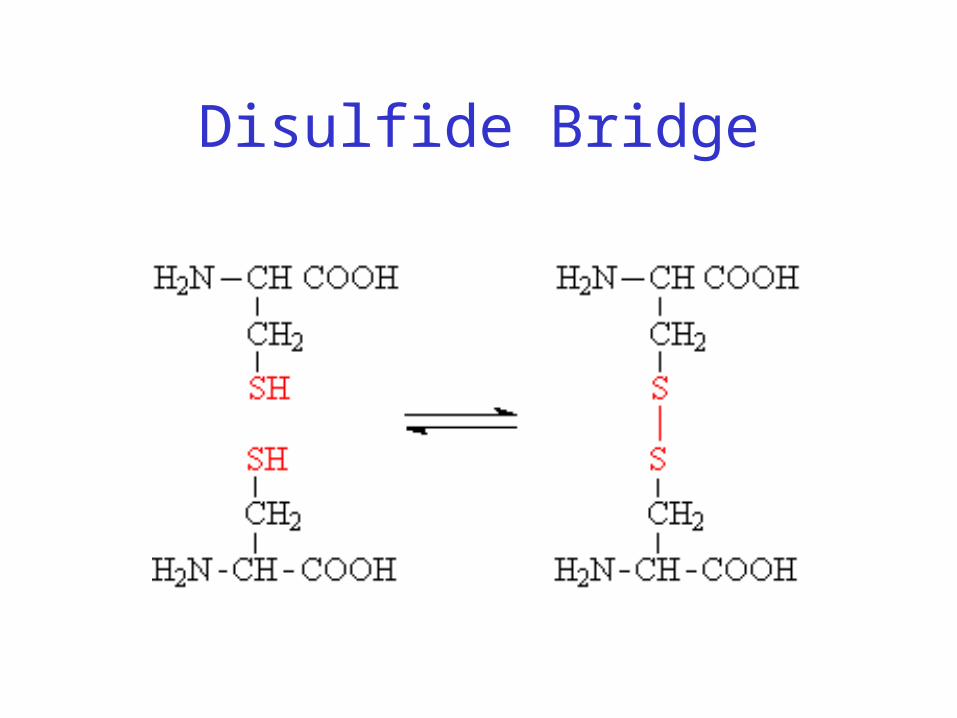
Disulfide Bridge

Disulfide Bonds• Side chain of cysteine contains highly reactive
thiol group
• Two thiol groups form a disulfide bond• Contribute to the stability of the folded state by
linking distant parts of the polypeptide chain
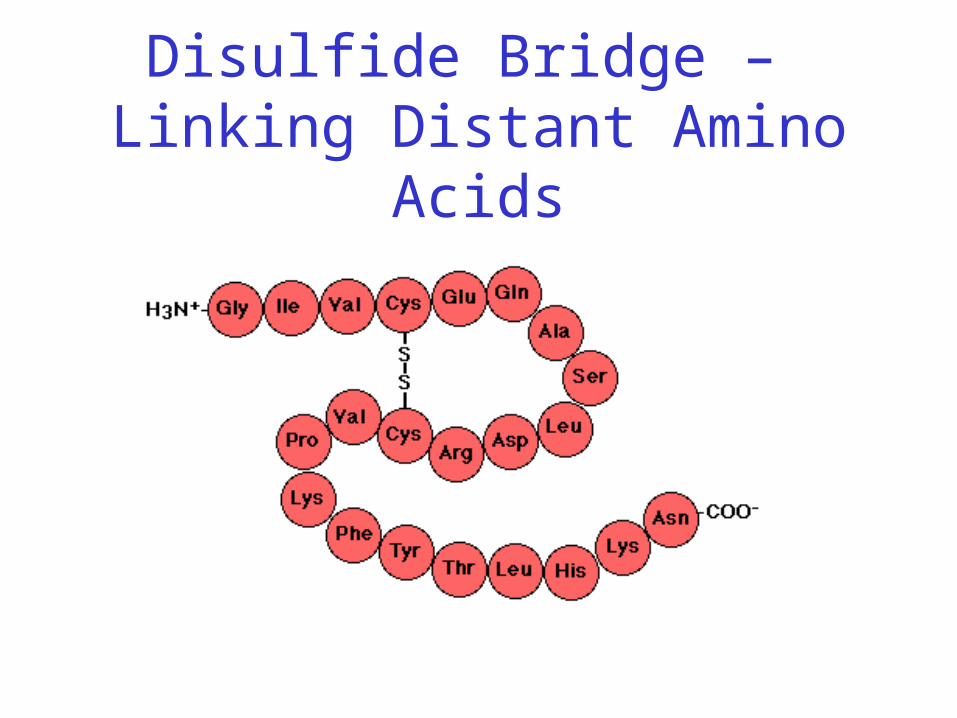
Disulfide Bridge – Linking Distant Amino Acids

Hemoglobin – Primary Structure
NH2-Val-His-Leu-Thr-Pro-Glu-Glu-Lys-Ser-Ala-Val-Thr-Ala-Leu-Trp-Gly-Lys-Val-Asn-Val-Asp-Glu-Val-Gly-Gly-Glu-…..
beta subunit amino acid sequence

Protein Structure - Primary
• Protein: chain of amino acids joined by peptide bonds

Protein Structure - Primary
• Protein: chain of amino acids joined by peptide bonds
• Amino Acid– Central carbon (Cα) attached to:
• Hydrogen (H)
• Amino group (-NH2)
• Carboxyl group (-COOH)
• Side chain (R)
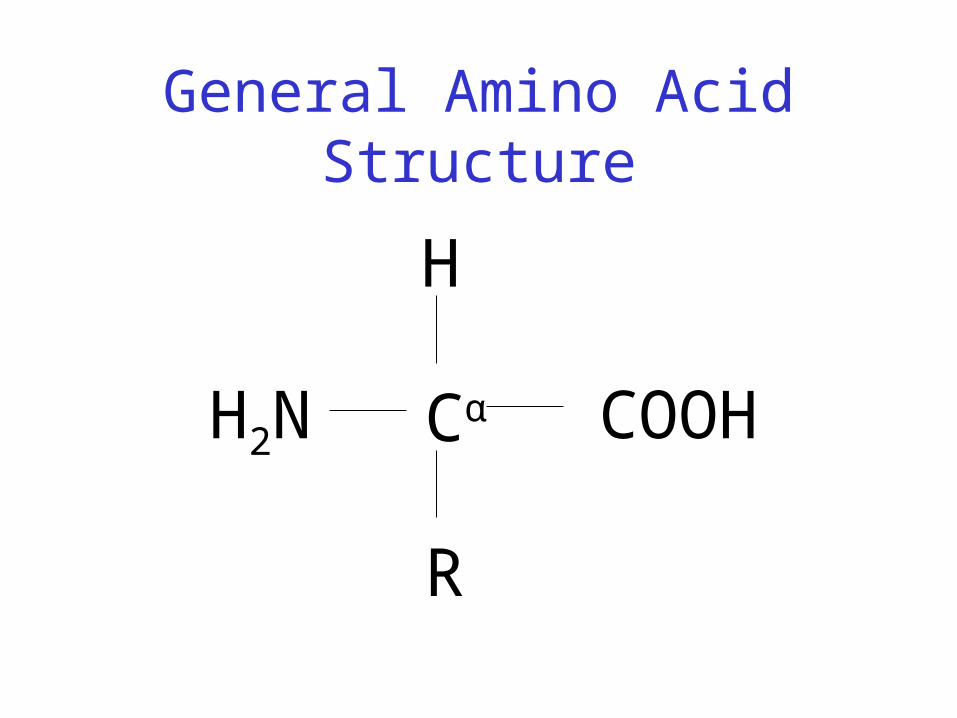
General Amino Acid Structure
Cα
H
R
COOHH2N
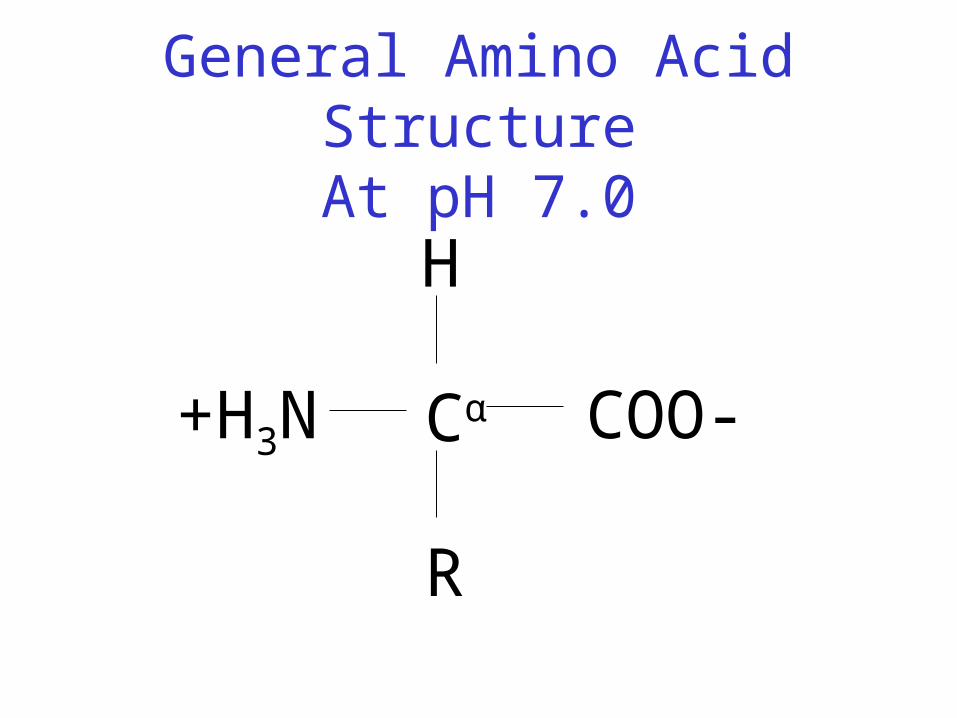
General Amino Acid StructureAt pH 7.0
Cα
H
R
COO-+H3N
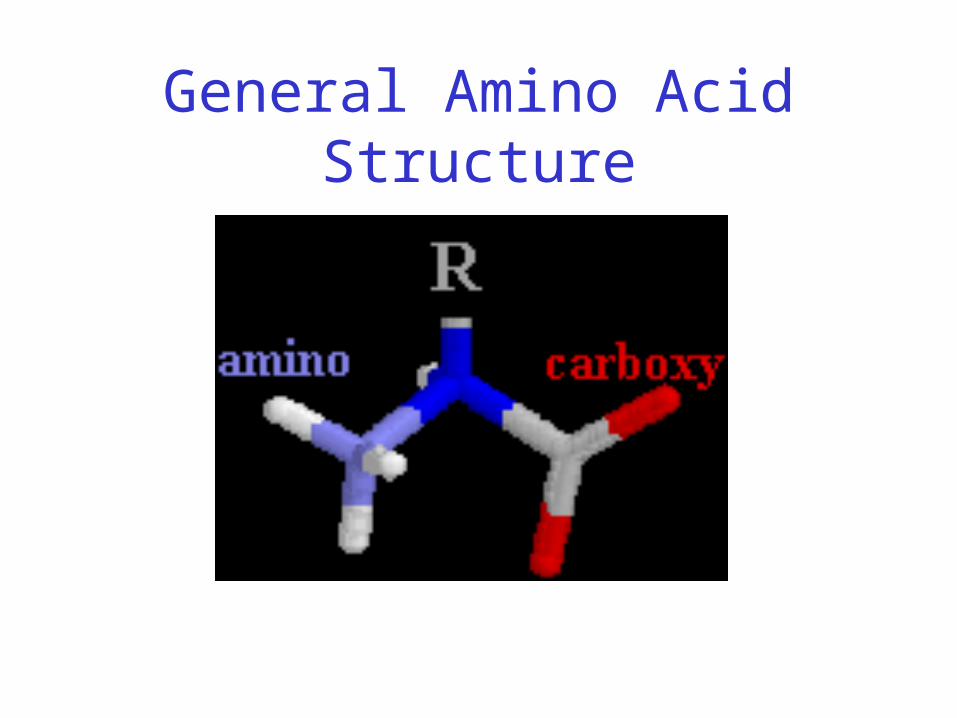
General Amino Acid Structure

Amino Acids
• Chiral
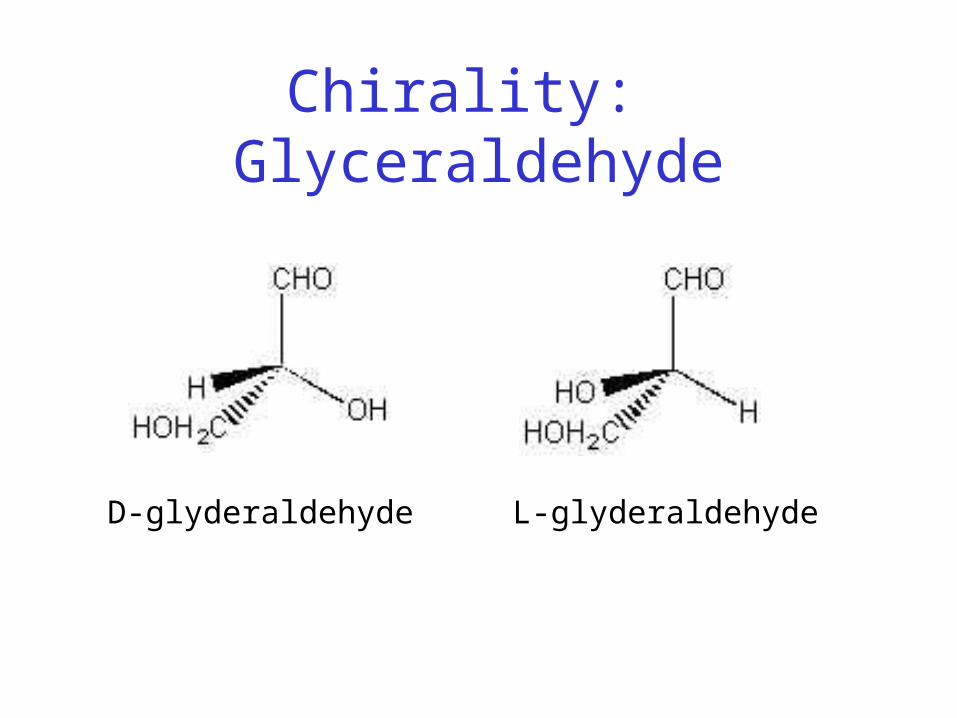
Chirality: Glyceraldehyde
L-glyderaldehydeD-glyderaldehyde

Amino Acids
• Chiral
• 20 naturally occuring; distinguishing side chain

20 Naturally-occurring Amino Acids

Amino Acids
• Chiral
• 20 naturally occuring; distinguishing side chain
• Classification: • Non-polar (hydrophobic)• Charged polar• Uncharged polar
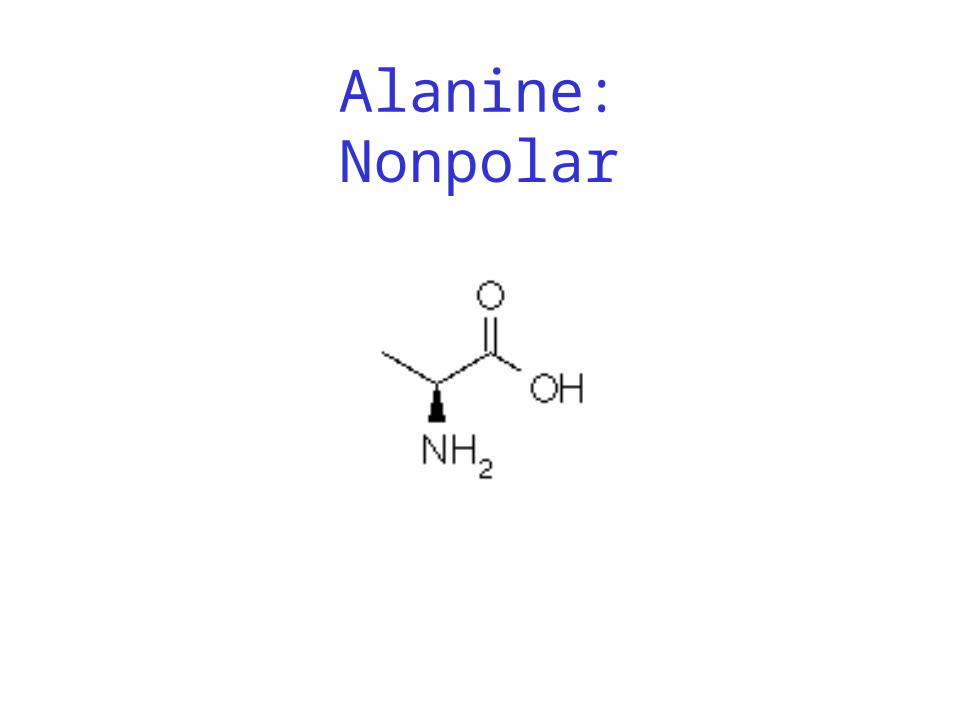
Alanine:Nonpolar

Serine:Uncharged Polar

Aspartic AcidCharged Polar
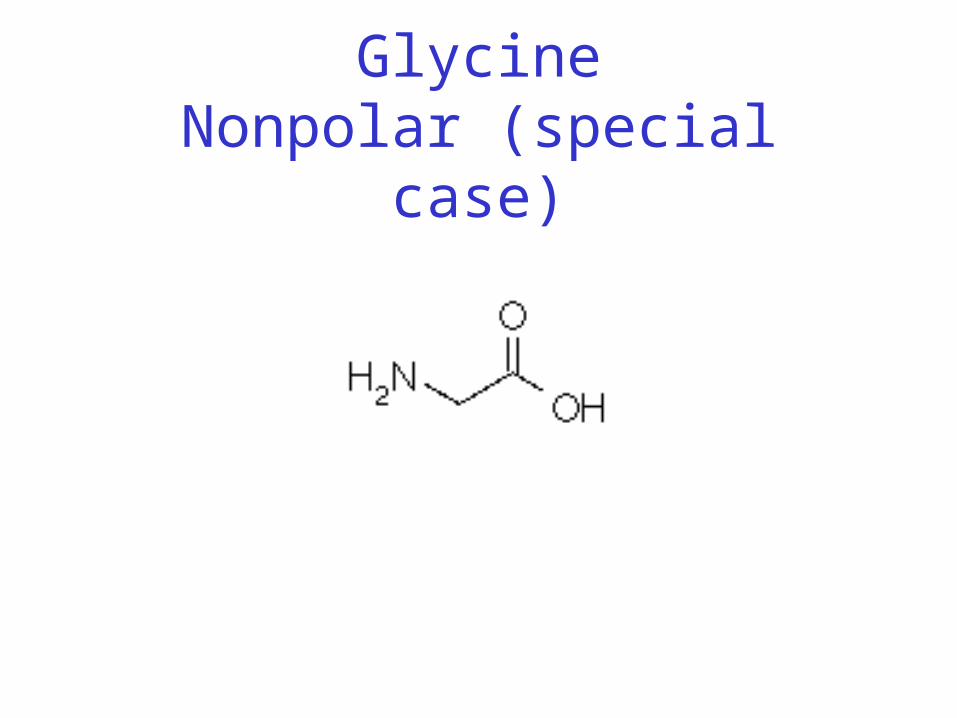
GlycineNonpolar (special case)

Peptide Bond
• Joins amino acids
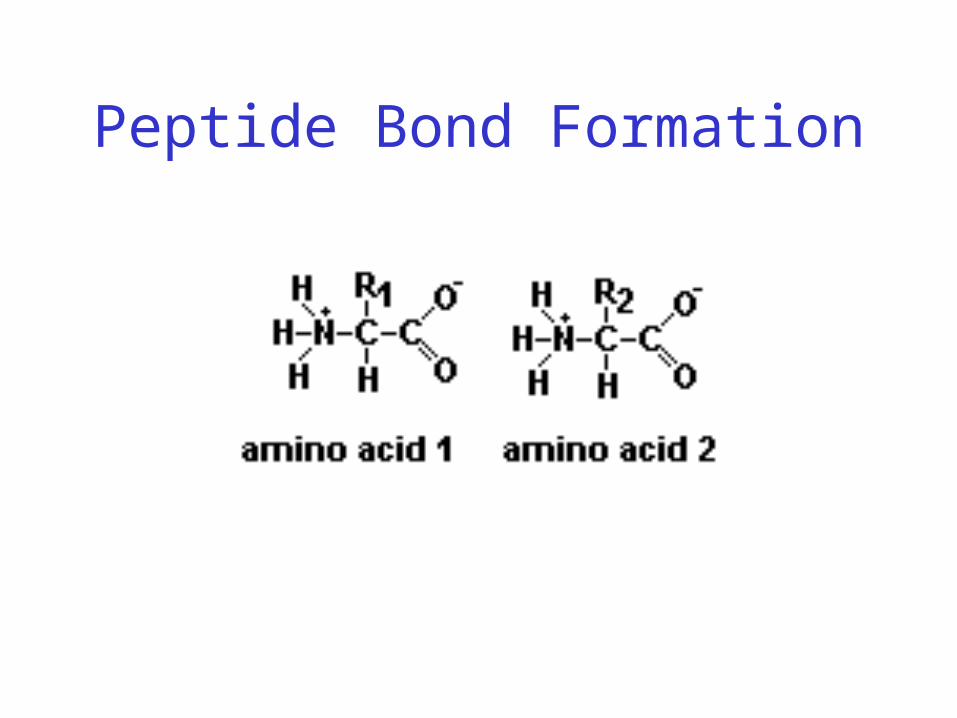
Peptide Bond Formation
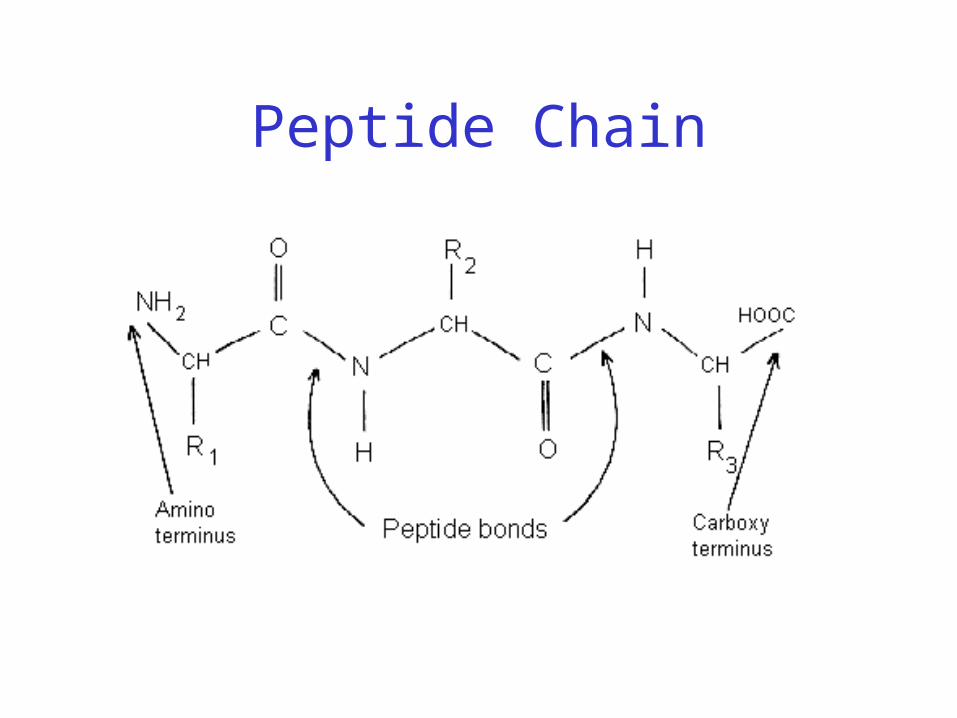
Peptide Chain

Peptide Bond
• Joins amino acids
• 40% double bond character– Caused by resonance
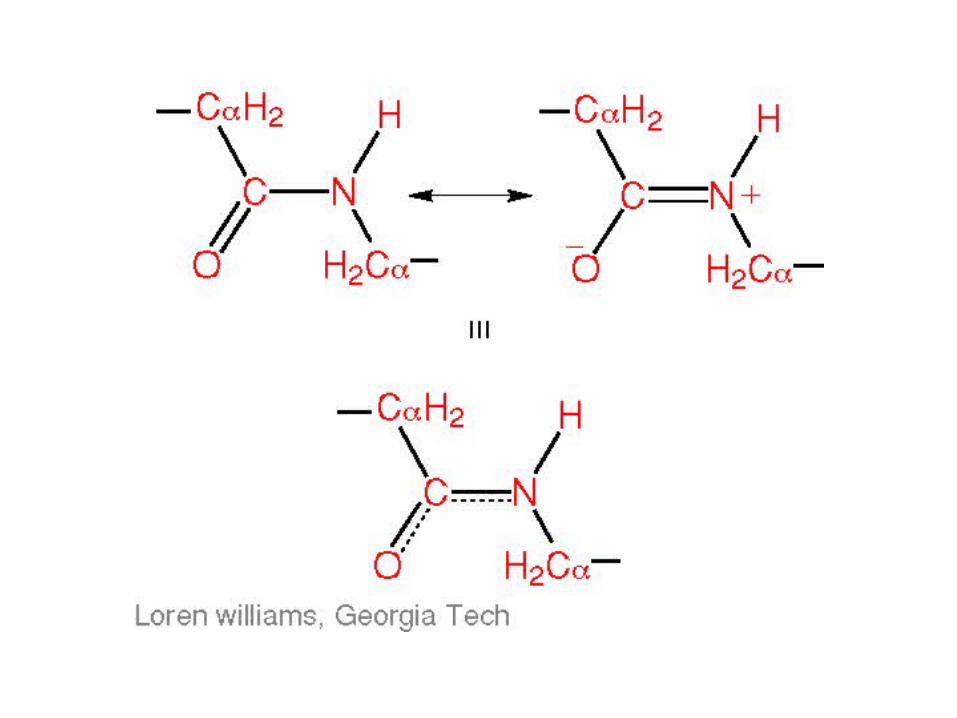

Peptide bond
• Joins amino acids
• 40% double bond character– Caused by resonance– Results in shorter bond length
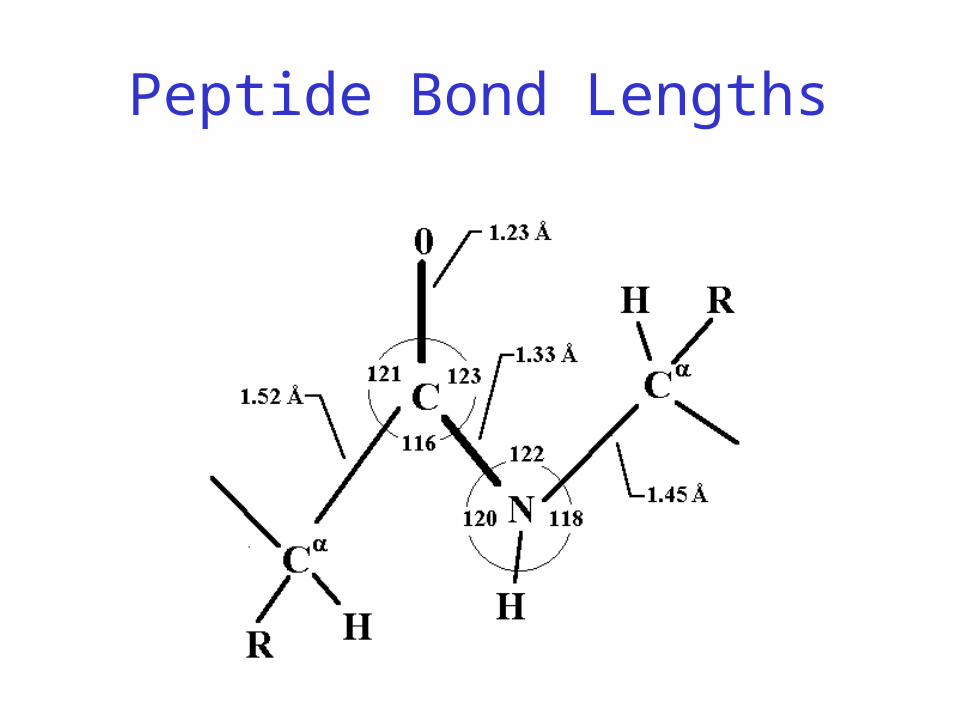
Peptide Bond Lengths

Peptide bond
• Joins amino acids
• 40% double bond character– Caused by resonance– Results in shorter bond length– Double bond disallows rotation

Protein Conformation Framework
• Bond rotation determines protein folding, 3D structure
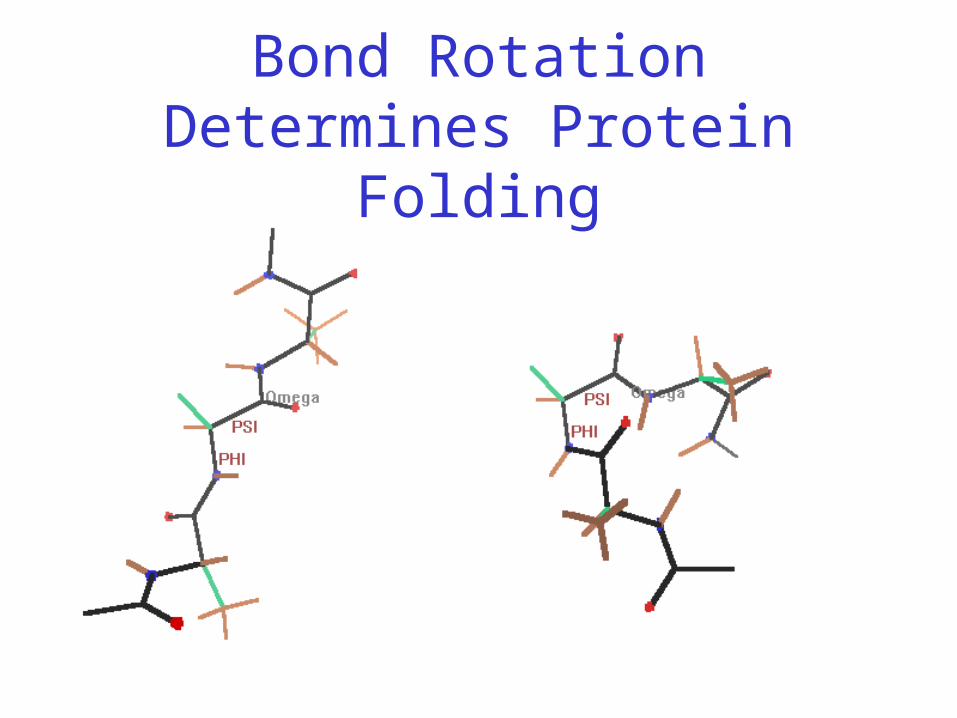
Bond Rotation Determines Protein Folding

Protein Conformation Framework
• Bond rotation determines protein folding, 3D structure
• Torsion angle (dihedral angle) τ– Measures orientation of four linked
atoms in a molecule: A, B, C, D
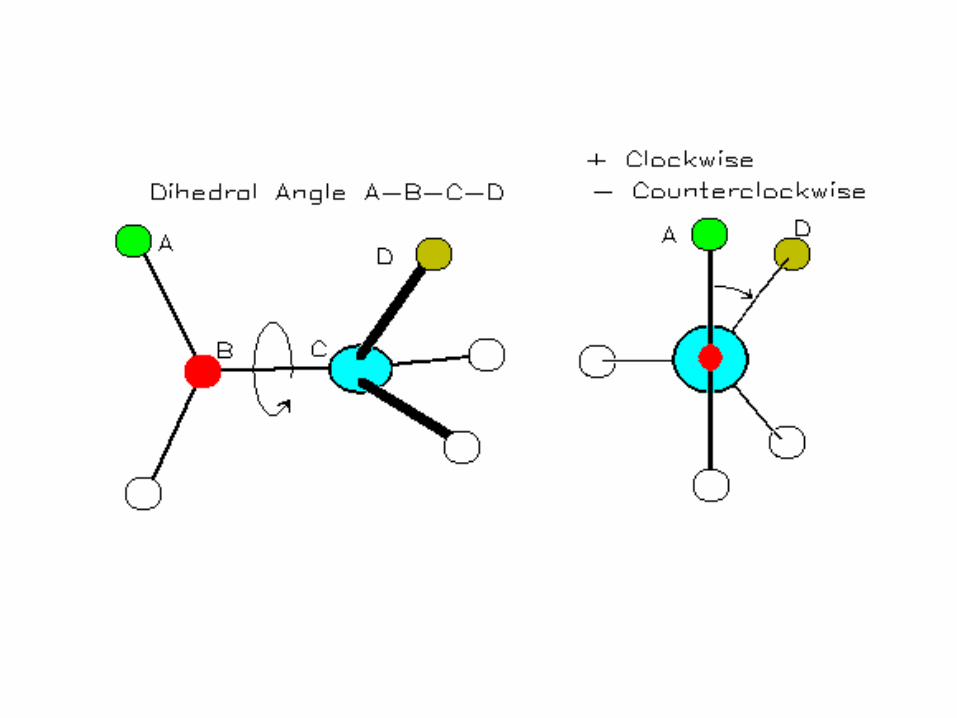

Protein Conformation Framework
• Bond rotation determines protein folding, 3D structure
• Torsion angle (dihedral angle) τ– Measures orientation of four linked atoms
in a molecule: A, B, C, D
– τABCD defined as the angle between the normal to the plane of atoms A-B-C and normal to the plane of atoms B-C-D
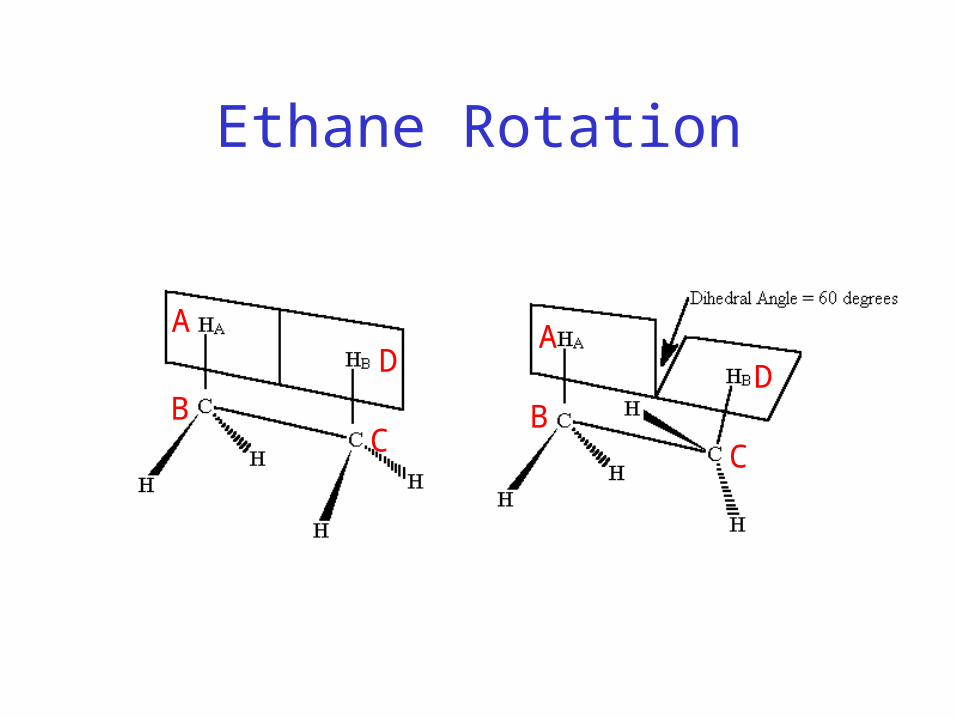
Ethane Rotation
A
CB
DA
BC
D

Protein Conformation Framework
• Bond rotation determines protein folding, 3D structure
• Torsion angle (dihedral angle) τ– Measures orientation of four linked atoms
in a molecule: A, B, C, D
– τABCD defined as the angle between the normal to the plane of atoms A-B-C and normal to the plane of atoms B-C-D
– Three repeating torsion angles along protein backbone: ω, φ, ψ
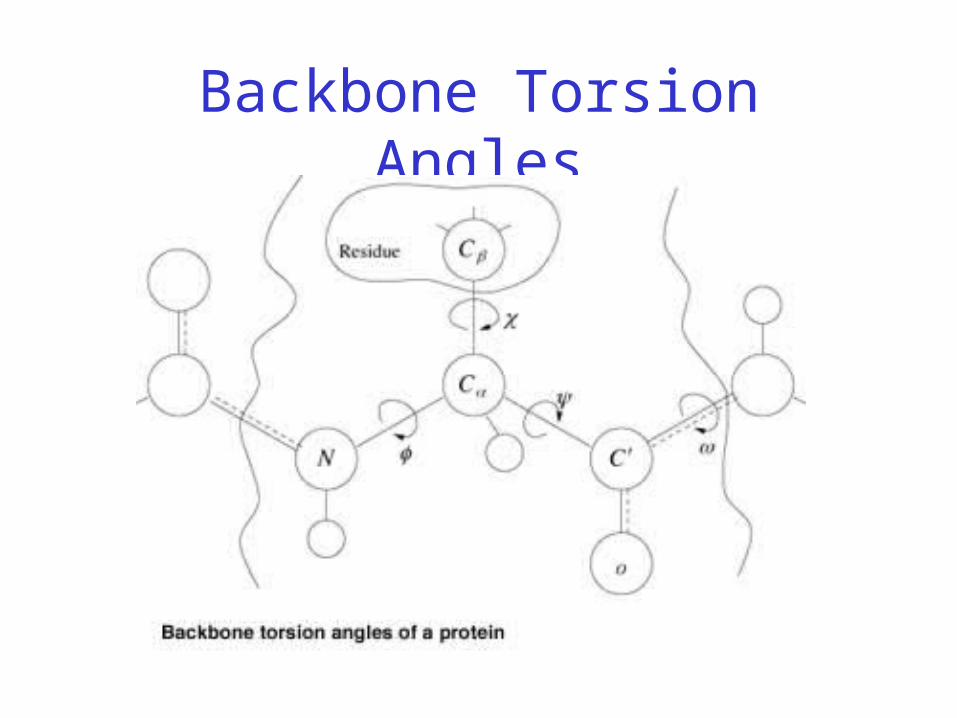
Backbone Torsion Angles

Backbone Torsion Angles
• Dihedral angle ω : rotation about the peptide bond, namely Cα
1-{C-N}- Cα2
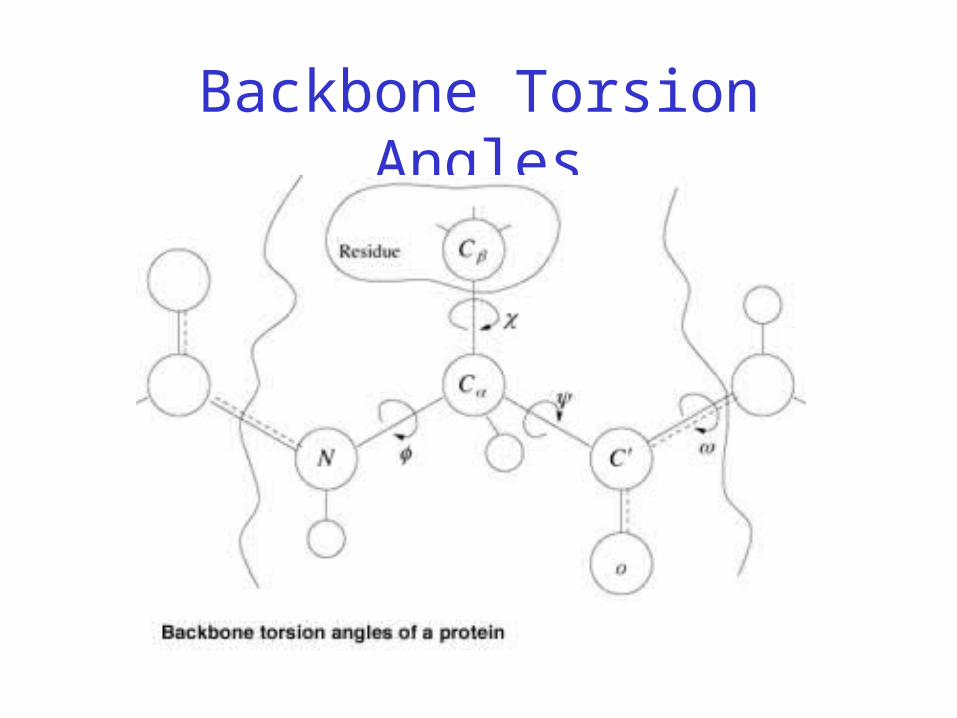
Backbone Torsion Angles

Backbone Torsion Angles
• Dihedral angle ω : rotation about the peptide bond, namely Cα
1-{C-N}- Cα2
• Dihedral angle φ : rotation about the bond between N and Cα
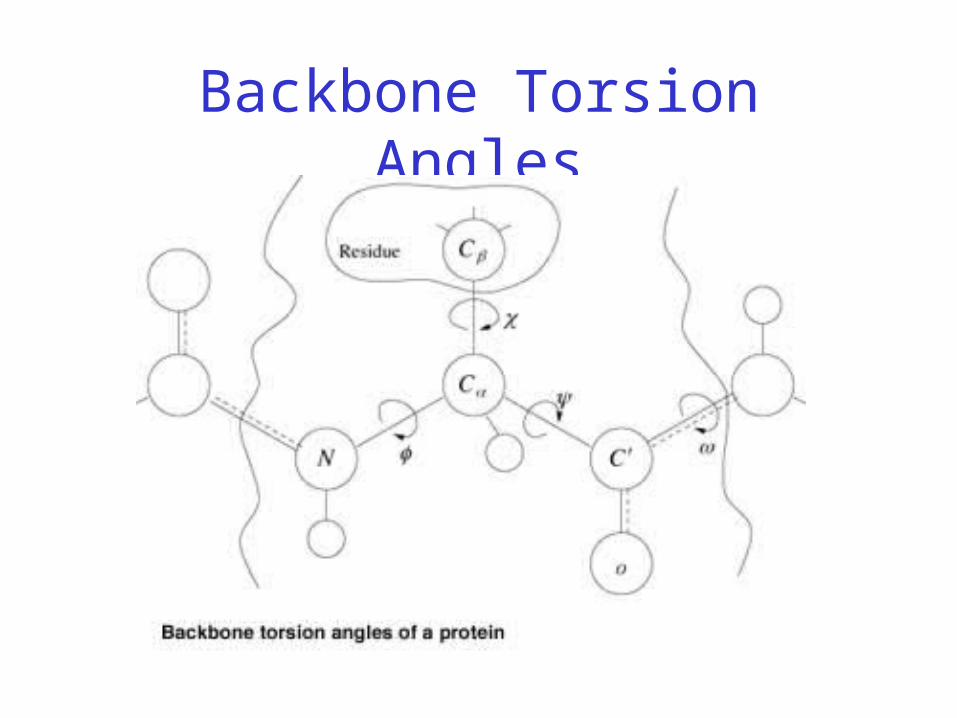
Backbone Torsion Angles

Backbone Torsion Angles
• Dihedral angle ω : rotation about the peptide bond, namely Cα
1-{C-N}- Cα2
• Dihedral angle φ : rotation about the bond between N and Cα
• Dihedral angle ψ : rotation about the bond between Cα and the carbonyl carbon
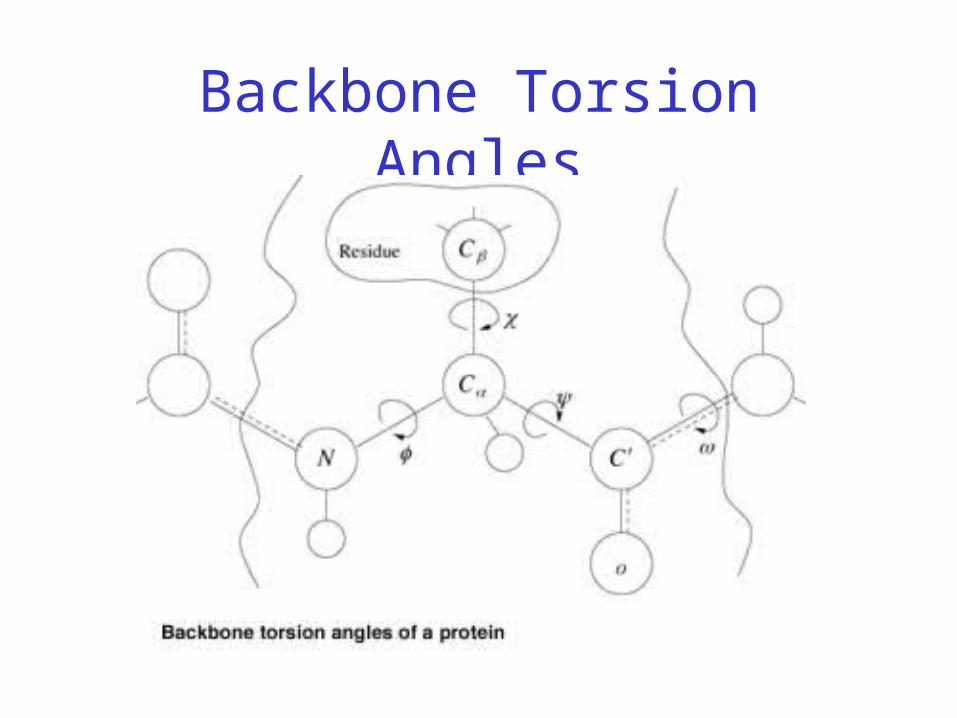
Backbone Torsion Angles

Backbone Torsion Angles
• ω angle tends to be planar (0º - cis, or 180 º - trans) due to delocalization of carbonyl π electrons and nitrogen lone pair
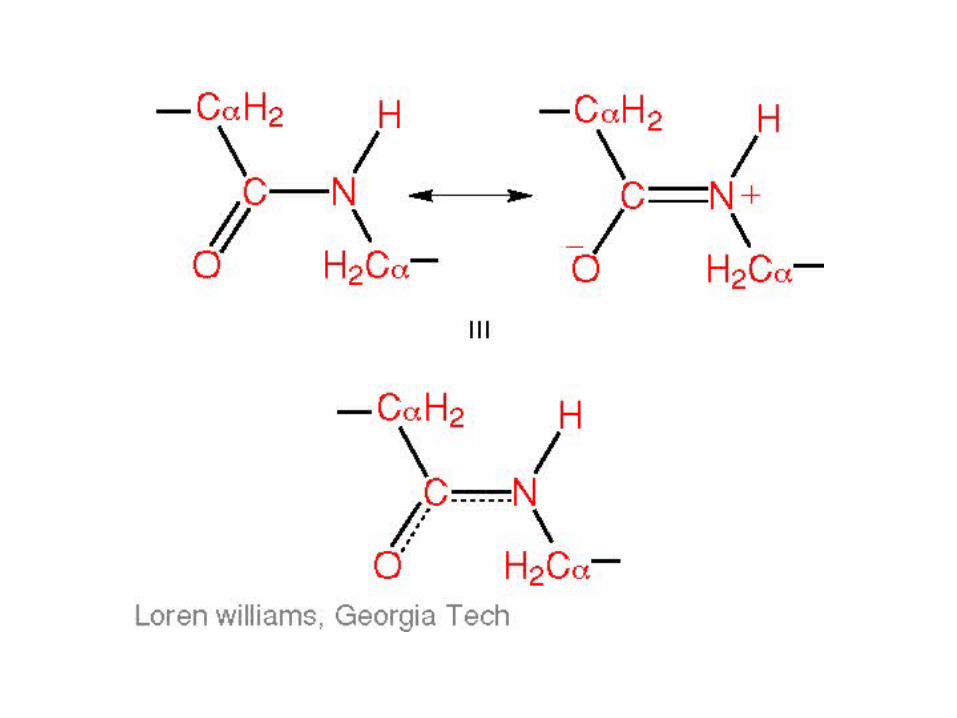

Backbone Torsion Angles
• ω angle tends to be planar (0º - cis, or 180 º - trans) due to delocalization of carbonyl pi electrons and nitrogen lone pair
• φ and ψ are flexible, therefore rotation occurs here

Backbone Torsion Angles

Backbone Torsion Angles
• ω angle tends to be planar (0º - cis, or 180 º - trans) due to delocalization of carbonyl pi electrons and nitrogen lone pair
• φ and ψ are flexible, therefore rotation occurs here• However, φ and ψ of a given amino acid residue
are limited due to steric hindrance

Steric Hindrance
• Interference to rotation caused by spatial arrangement of atoms within molecule
• Atoms cannot overlap
• Atom size defined by van der Waals radii
• Electron clouds repel each other

Backbone Torsion Angles
• ω angle tends to be planar (0º - cis, or 180 º - trans) due to delocalization of carbonyl pi electrons and nitrogen lone pair
• φ and ψ are flexible, therefore rotation occurs here• However, φ and ψ of a given amino acid residue
are limited due to steric hindrance• Only 10% of the {φ, ψ} combinations are
generally observed for proteins• First noticed by G.N. Ramachandran

G.N. Ramachandran
• Used computer models of small polypeptides to systematically vary φ and ψ with the objective of finding stable conformations
• For each conformation, the structure was examined for close contacts between atoms
• Atoms were treated as hard spheres with dimensions corresponding to their van der Waals radii
• Therefore, φ and ψ angles which cause spheres to collide correspond to sterically disallowed conformations of the polypeptide backbone

Ramachandran Plot
• Plot of φ vs. ψ
• The computed angles which are sterically allowed fall on certain regions of plot
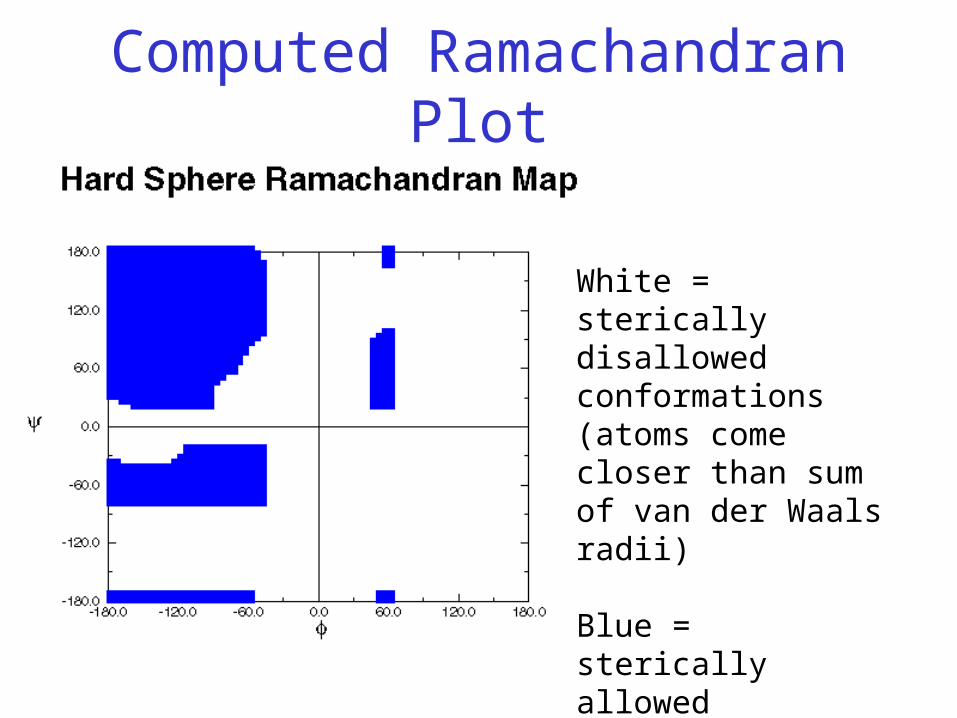
Computed Ramachandran Plot
White = sterically disallowed conformations (atoms come closer than sum of van der Waals radii)
Blue = sterically allowed conformations

Ramachandran Plot
• Plot of φ vs. ψ
• Computed sterically allowed angles fall on certain regions of plot
• Experimentally determined angles fall on same regions
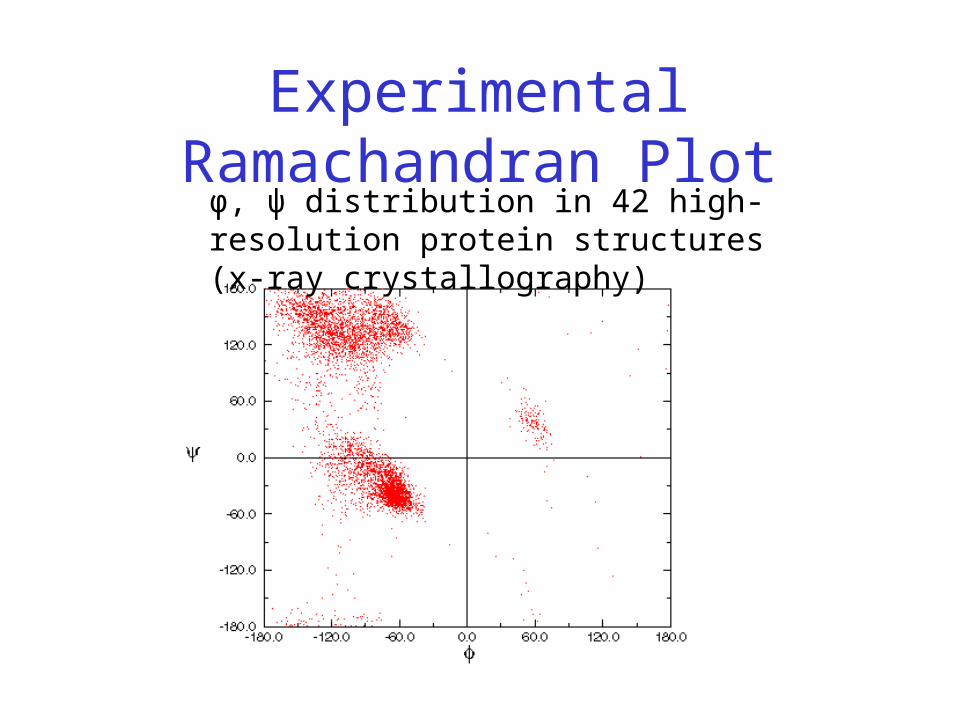
Experimental Ramachandran Plotφ, ψ distribution in 42 high-resolution protein structures (x-ray crystallography)

Ramachandran PlotAnd Secondary Structure
• Repeating values of φ and ψ along the chain result in regular structure
• For example, repeating values of φ ~ -57° and ψ ~ -47° give a right-handed helical fold (the alpha-helix)
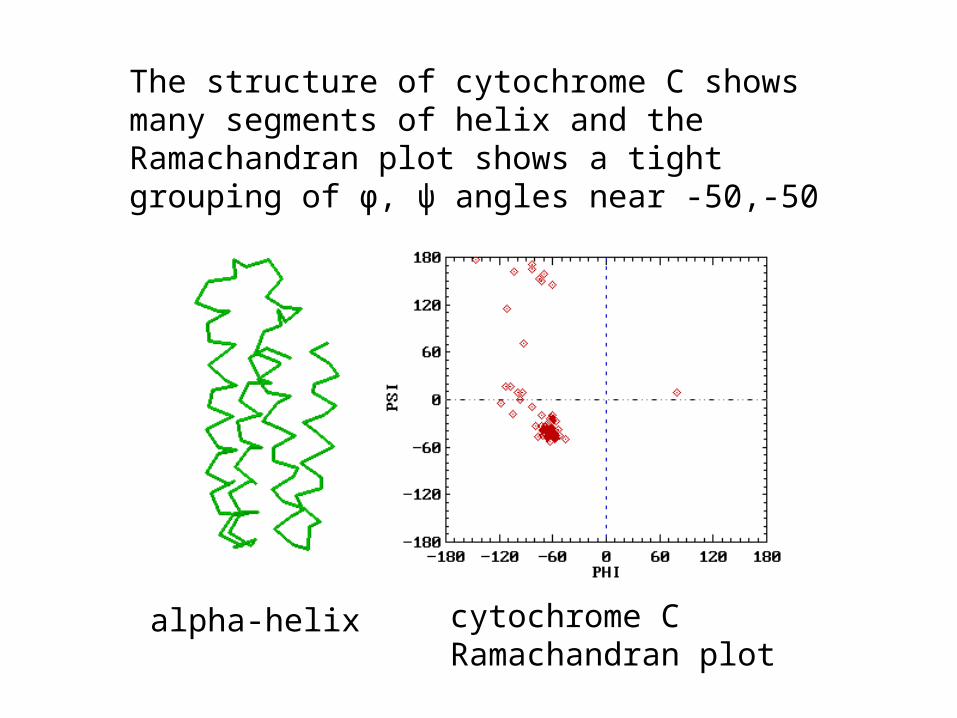
The structure of cytochrome C shows many segments of helix and the Ramachandran plot shows a tight grouping of φ, ψ angles near -50,-50
alpha-helix cytochrome CRamachandran plot
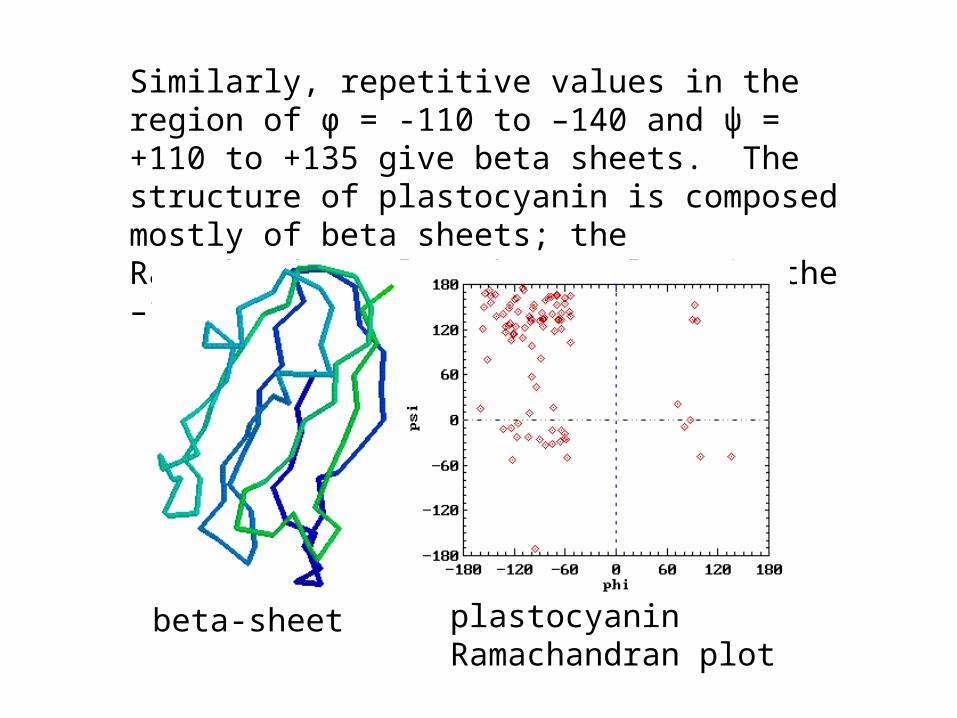
Similarly, repetitive values in the region of φ = -110 to –140 and ψ = +110 to +135 give beta sheets. The structure of plastocyanin is composed mostly of beta sheets; the Ramachandran plot shows values in the –110, +130 region:
beta-sheet plastocyanin Ramachandran plot
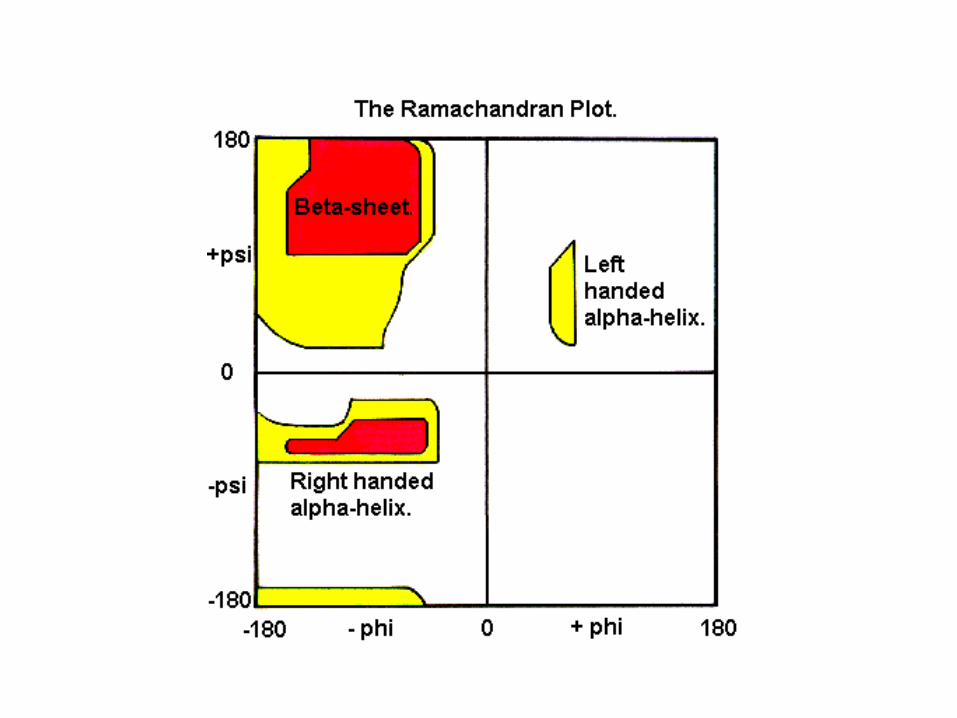

Ramachandran PlotAnd Secondary Structure
• White = sterically disallowed conformations
• Red = sterically allowed regions if strict (greater) radii are used (namely right-handed alpha helix and beta sheet)
• Yellow = sterically allowed if shorter radii are used (i.e. atoms allowed closer together; brings out left-handed helix)
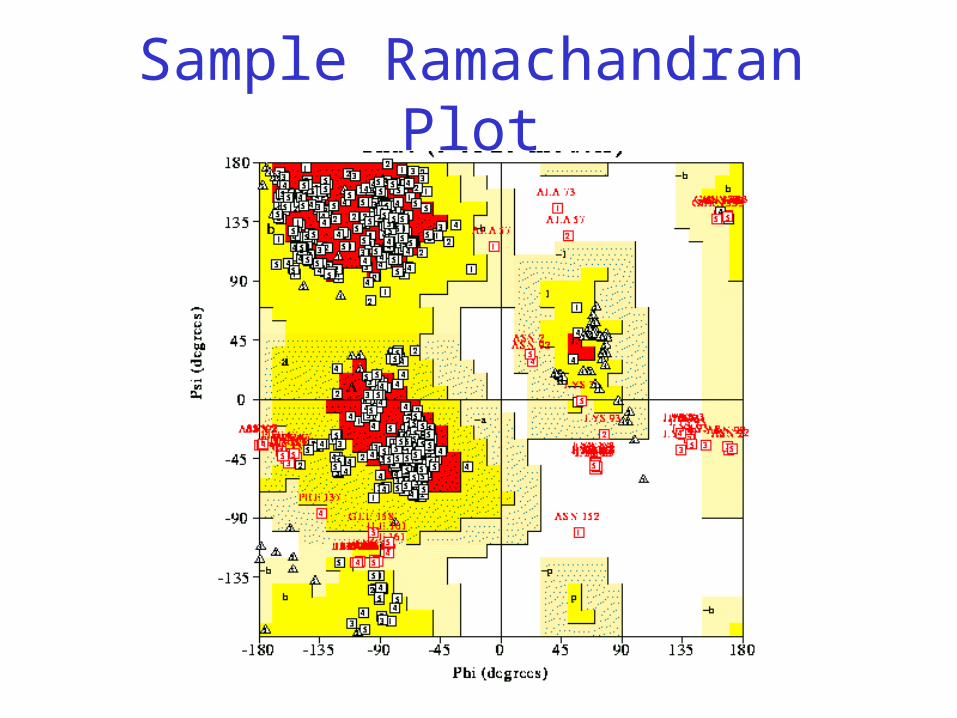
Sample Ramachandran Plot
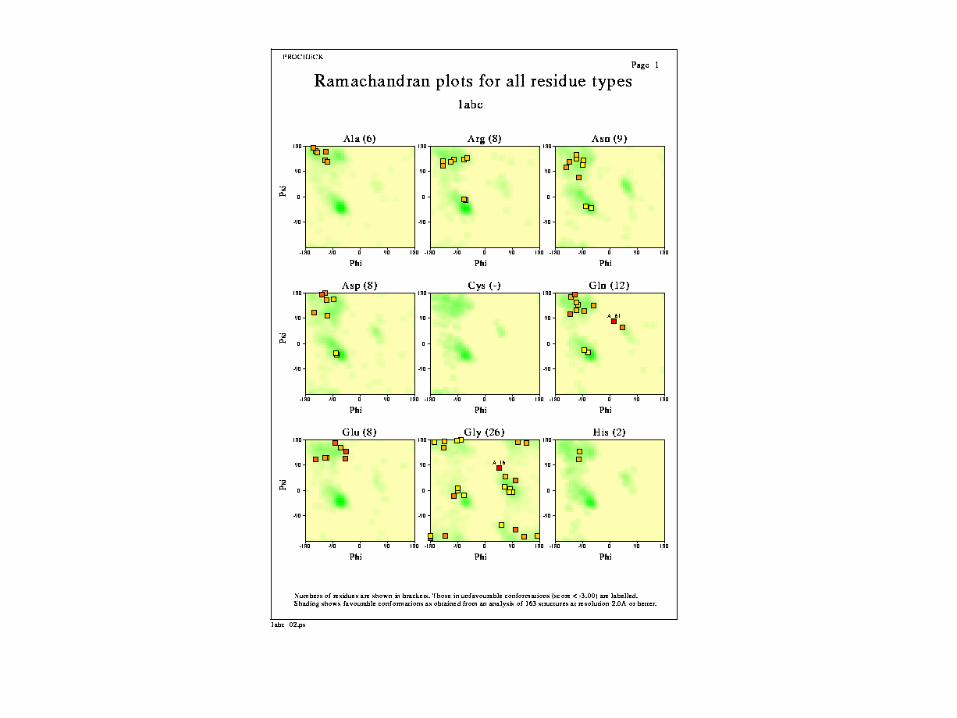
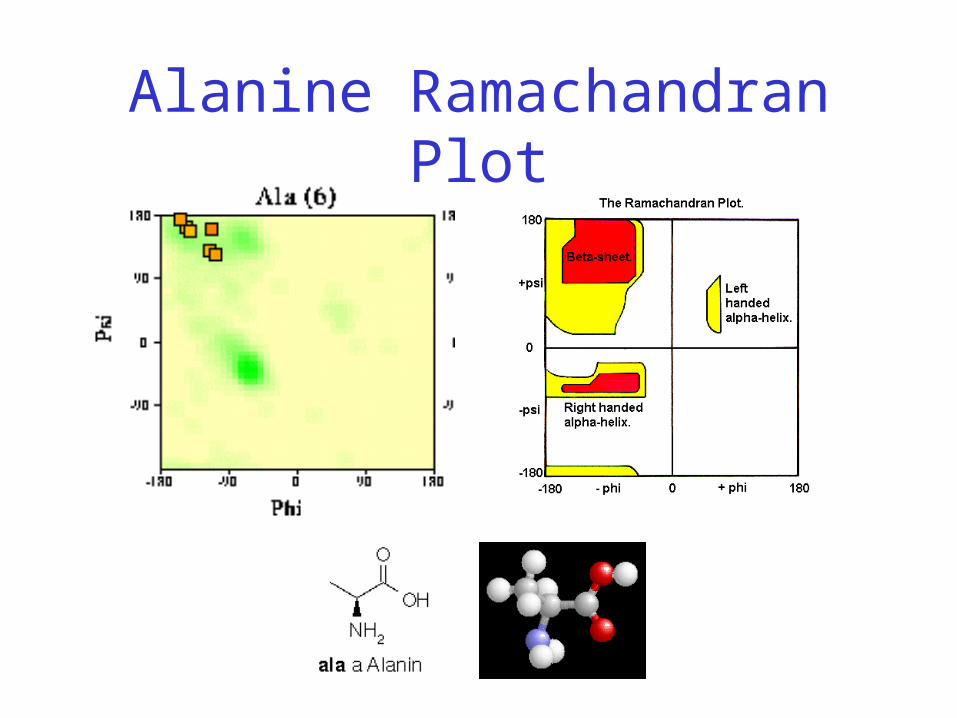
Alanine Ramachandran Plot

Arginine Ramachandran Plot
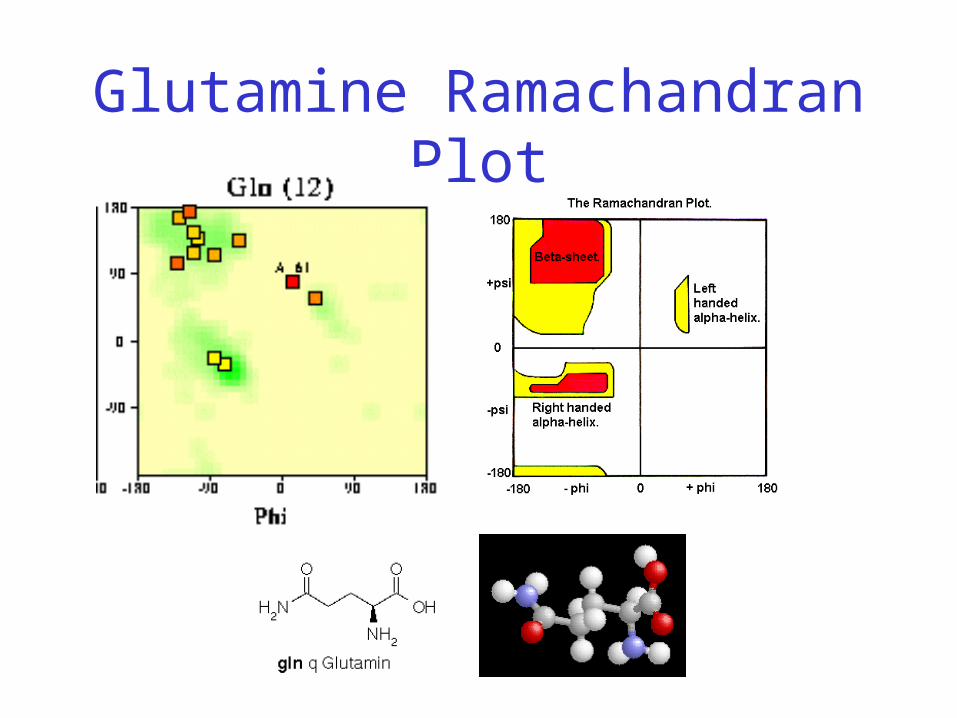
Glutamine Ramachandran Plot
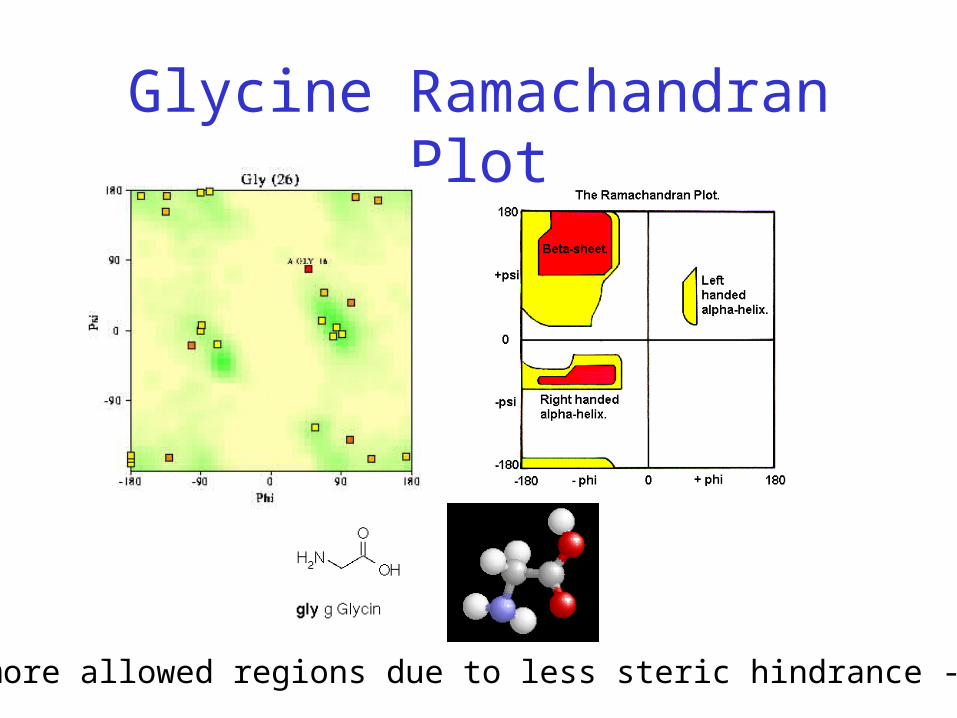
Glycine Ramachandran Plot
Note more allowed regions due to less steric hindrance - Turns
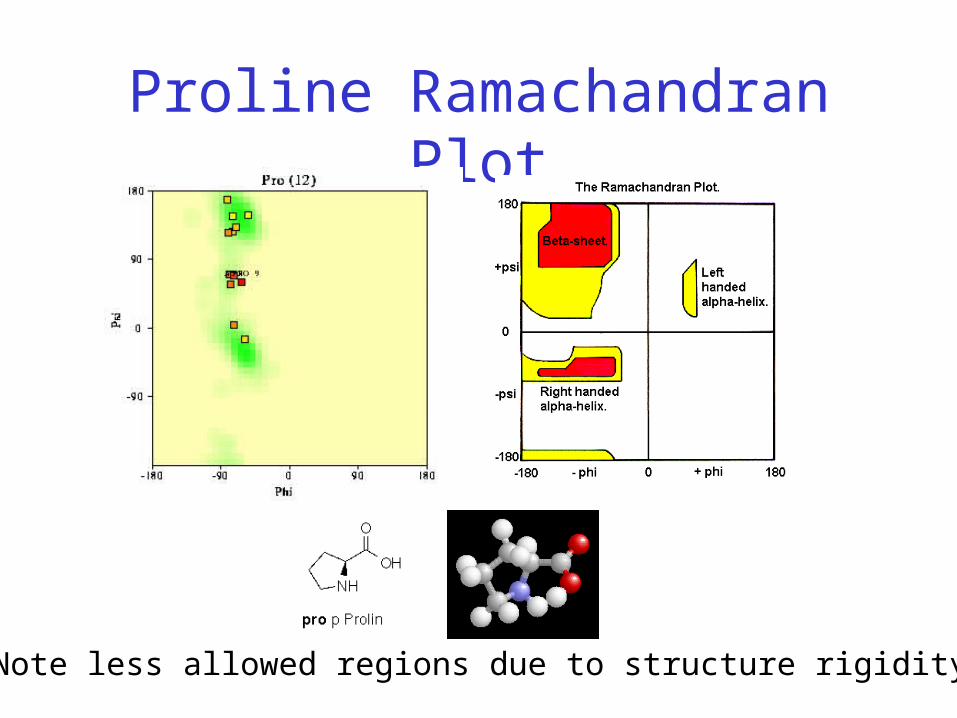
Proline Ramachandran Plot
Note less allowed regions due to structure rigidity

φ, ψ and Secondary Structure
Name φ ψ Structure ------------------- ------- ------- ---------------------------------alpha-L 57 47 left-handed alpha helix3-10 Helix -49 -26 right-handed.π helix -57 -80 right-handed.Type II helices -79 150 left-handed helices formed by polyglycine and polyproline.Collagen -51 153 right-handed coil formed of three left handed helicies.

Sequence Similarity
• Sequence similarity implies structural, functional, and evolutionary commonality

Homologous Proteins:Enterotoxin and Cholera toxin
Enterotoxin Cholera toxin
80% homology

Sequence Similarity
• Sequence similarity implies structural, functional, and evolutionary commonality
• Low sequence similarity implies little structural similarity
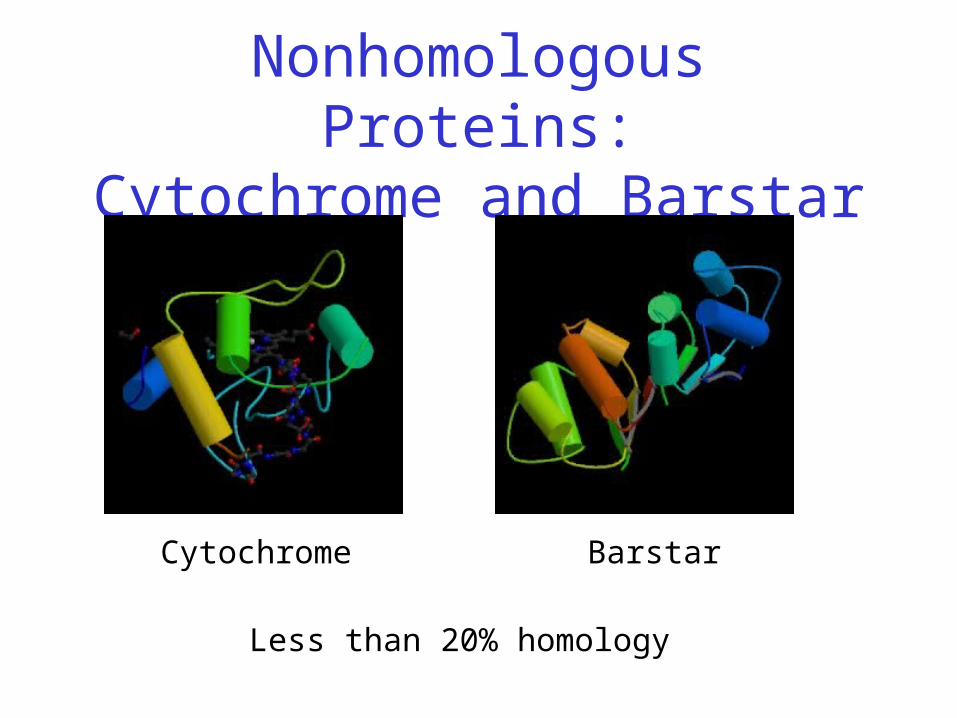
Nonhomologous Proteins:Cytochrome and Barstar
Cytochrome Barstar
Less than 20% homology

Sequence Similarity
• Sequence similarity implies structural, functional, and evolutionary commonality
• Low sequence similarity implies little structural similarity
• Small mutations generally well-tolerated by native structure – with exceptions!

Sequence Similarity Exception
• Sickle-cell anemia resulting from one residue change in hemoglobin protein
• Replace highly polar (hydrophilic) glutamate with nonpolar (hydrophobic) valine
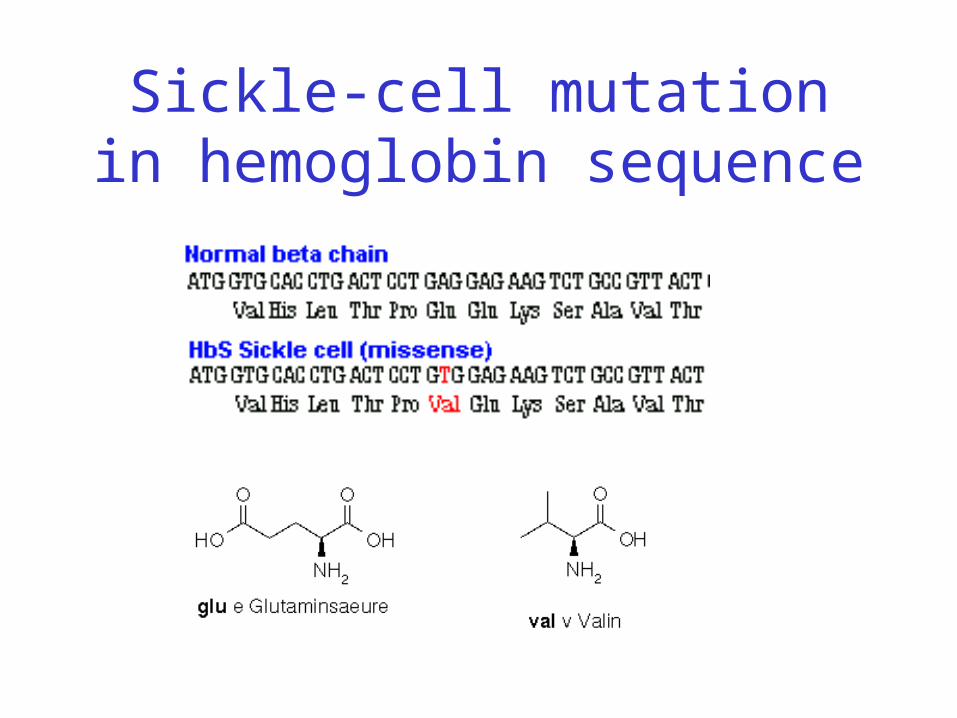
Sickle-cell mutation in hemoglobin sequence

Normal Trait• Hemoglobin molecules exist as single,
isolated units in RBC, whether oxygen bound or not
• Cells maintain basic disc shape, whether transporting oxygen or not

Sickle-cell Trait
• Oxy-hemoglobin is isolated, but de-oxyhemoglobin sticks together in polymers, distorting RBC
• Some cells take on “sickle” shape

Sickle-cell

RBC Distortion• Hydrophobic valine replaces hydrophilic glutamate• Causes hemoglobin molecules to repel water and be
attracted to one another• Leads to the formation of long hemoglobin filaments

Hemoglobin Polymerization
Normal
Mutant

RBC Distortion• Hydrophobic valine replaces hydrophilic glutamate• Causes hemoglobin molecules to repel water and be
attracted to one another• Leads to the formation of long hemoglobin filaments • Filaments distort the shape of red blood cells
(analogy: icicle in a water balloon)• Rigid structure of sickle cells blocks capillaries and
prevents red blood cells from delivering oxygen
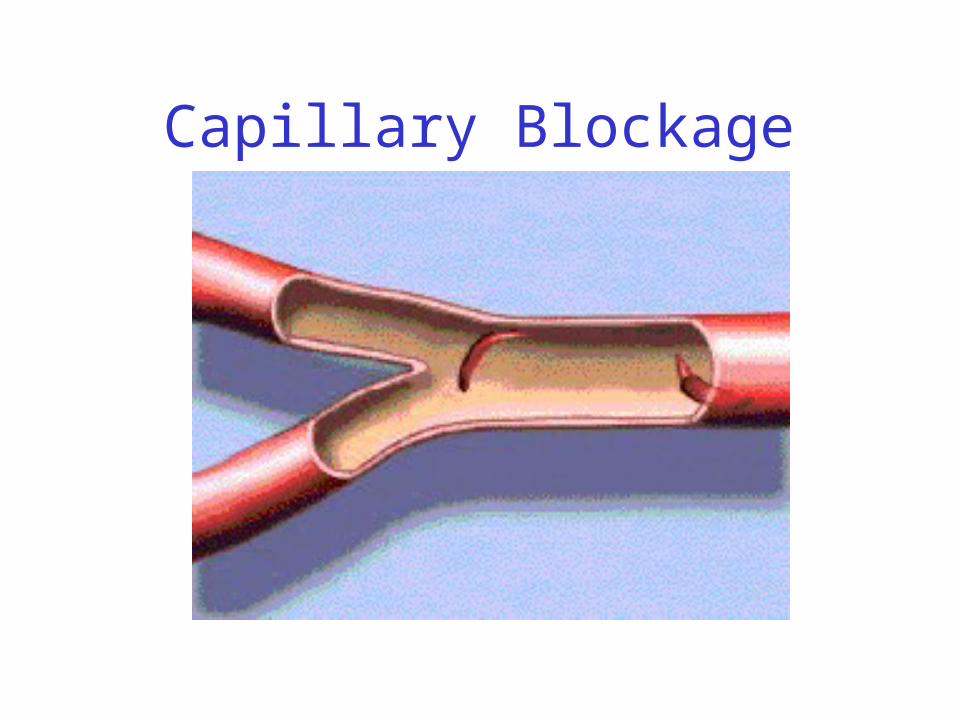
Capillary Blockage

Sickle-cell Trait
• Oxy-hemoglobin is isolated, but de-oxyhemoglobin sticks together in polymers, distorting RBC
• Some cells take on “sickle” shape
• When hemoglobin again binds oxygen, again becomes isolated
• Cyclic alteration damages hemoglobin and ultimately RBC itself

Protein: The Machinery of Life
“Life is the mode of existence of proteins, and this mode of existence essentially consists in the constant self-renewal of the chemical constituents of these substances.”
Friedrich Engles, 1878
NH2-Val-His-Leu-Thr-Pro-Glu-Glu-Lys-Ser-Ala-Val-Thr-Ala-Leu-Trp-Gly-Lys-Val-Asn-Val-Asp-Glu-Val-Gly-Gly-Glu-…..





























