Embed Size (px)
Citation preview

Eur. J . Biochem. 72, 341-352 (1977)
Properties of the Penicillin-Binding Proteins of Escherichia coli K 12
Brian G. SPRATT
Department of Genetics, University of Leicester
(Received September 14 / October 30, 1976)
Ben~yl[ '~C] penicillin binds to six proteins with molecular weights of between 40000 and 91 000 in the inner membrane of Escherichia coli. Two additional binding proteins with molecular weights of 29000 and 32000 were sometimes detected. All proteins were accessible to ben~yl['~C]penicillin in whole cells. Proteins 5 and 6 released bound ben~yI['~C]penicillin with half times of 5 and 19 min at 30 'C but the other binding proteins showed less than 50% release during a 60-min period at 30 "C. The rate of release of bound penicillin from some of the proteins was greatly stimulated by 2-mei-captoethanol and neutral hydroxylamine. Release of ben~yI['~C]penicillin did not occur ~ t ' the binding proteins were denatured in anionic detergent and so was probably enzymic. No additional binding proteins were detected with two ['4C]cephalosporins. These p-lactams bound to either all or some of those proteins to which ben~yl['~C]penicillin bound. No binding proteins have been detected in the outer membrane of E. coli with any p-['4C]lactam. The binding of a range of unlabelled penicillins and cephalosporins were studied by measuring their competition for the binding of berl~yl['~C]penicillin to the six penicillin-binding proteins. These results, together with those obtained by direct binding experiments with P-[14C]lactams, showed that penicillins bind to all six proteins but that at least some cephalosporins fail to bind, or bind very slowly, to proteins 2, 5 and 6, although they bind to the other proteins. Since these cephalosporins inhibited cell division and caused cell lysis at concentrations where we could detect no binding to proteins 2, 5 and 6, we believe that these latter proteins are not the targets at which S-lactams bind to elicit the above physiological responses. The binding properties of proteins 1, 3 and 4 correlate reasonably well with those expected for the above killing targets.
The p-lactam group of antibiotics (penicillins and cephalosporins) kill bacteria by inhibiting the terminal stages of peptidoglycan metabolism [l - 31. The intro- duction of nascent precursor chains into the growing peptidoglycan appears to be the primary target of their action [4,5].
In recent years two complementary approaches have been taken to study the interaction of these antibiotics with bacterial cells. The first of these involves the identification and purification of enzymes which are inhibited by P-lactams. In Escherichia coli three enzymic reactions have been shown to be penicillin-sensitive : D-alanine carboxypeptidase 1, peptidoglycan trdnspeptidase and peptidoglycan endo- peptidase [6- lo]. In all bacteria that have been studied in any detail multiple penicillin-sensitive enzymes have been identified and in several cases purified enzymes have been shown to catalyse more than one of the above reactions [6,9, lo].
The second of these approaches depends on the old observation that penicillin irreversibly binds to bacterial membranes [I I ] and the recognition that this reflects the irreversible binding of ,8-lactams to the enzymes which they inhibit [2,6]. This implies that penicillin-sensitive enzymes and penicillin-binding proteins are synonymous, and that penicillin-sensitive enzymes can be detected as those proteins which bind penicillin. Multiple penicillin-binding proteins have been detected in the membranes of all bacteria that have been examined [6,12,13], and it is now clear that the interaction of p-lactams with bacterial cells is considerably more complex than once was thought.
The physiological and morphological response of E.coli to P-lactam antibiotics is also complex (e.g p-lactams can inhibit cell division, cause cell lysis, bulge formation or the formation of osmotically-stable ovoid cells) and we have recently been identifying the physjological targets to which P-lactams bind to

342 Penicillin-Binding Proteins of Escherichia coli
elicit the above responses [13,14]. For this purpose we have developed a method for detecting penicillin- binding proteins on dodecylsulphate-polyacrylamide slab gels and in this paper describe the properties of the E. coli penicillin-binding proteins.
MATERIALS AND METHODS
Organisms
E. coli strain KN126 was used in all experiments. It has the genotype F- trpE9829 tyr- ilv- sup-126 and was obtained from Dr T. Nagata.
Growth of Cells and Preparation of Membranes
Cells were routinely grown in 3-1 batch cultures in pennassay broth (Difco Antibiotic Medium No. 3) at 37 "C with vigorous aeration and were harvested in mid to late exponential growth phase. After cooling on ice, the cells were centrifuged at 6000 x g for 6 rnin and resuspended in 80 ml of ice-cold 10 mM sodium phosphate buffer (pH 7.0). 2-Mercaptoethanol was added (final concentration of 0.14 M) and the cells were broken open by three 30-s pulses of sonication with intervening 30-s periods of cooling. Unbroken cells were removed by centrifugation at 8000 x g for 20 min at 4 "C in the SS-34 rotor of a Sorvall RC2B centrifuge, and the cell membranes were pelleted out of the supernatant by centrifugation at 100000 x g for 40 rnin at 4 "C in the 50Ti rotor of a Beckman ultra- centrifuge. The membranes were resuspended in 20 ml of ice-cold phosphate buffer (as above), washed twice by centrifugation at 100000 x g, and finally stored in the above buffer at a concentration of approximately 40 mg protein/ml in a Revco -80 "C freezer. These washed cell membranes consist of inner (cytoplasmic) membrane, outer membrane and peptidoglycan but will be referred to as membranes for brevity.
Binding of Benzyl[ ''C]penicillin to E. coli Membranes
Membranes were thawed slowly and adjusted to 5 - 10 mg protein/ml and to 50 mM sodium phosphate (buffer pH 7.0). MgC12, which had been included in the assay mixture in earlier experiments [13], was omitted as it was found to be inessential for binding. For the standard assay of penicillin-binding proteins 20 pl of ben~yl['~C]penicillin was bound to 200 pl of membranes for 10 rnin at 30 "C and the reaction was terminated by the addition of 5 pl of non-radioactive benzylpenicillin (120 mg/ml) and 10 pl of 20% (w/v) sodium lauroyl sarcosinate (Sarkosyl NL-97). The addition of the anionic detergent sarkosyl has a dual
purpose. Firstly it denatures the penicillin-binding proteins and prevents the enzymic loss of bound penicillin (see Results), and secondly, it selectively and completely solubilises the proteins of the inner membrane whilst leaving the outer membrane in- soluble 1151.
After solubilisation of the inner membrane for 20 min at room temperature, the sarkosyl-insoluble outer membrane and peptidoglycan were removed by centrifugation at 100000 x g for 40 min at 10 "C in the 50Ti rotor of a Beckman ultracentrifuge using 10 ml Oak-Ridge-type polycarbonate screw-cap tubes. In recent experiments the removal of the sarkosyl- insoluble fraction by centrifugation at 40000 x g in the SM24 rotor of a Sorvall RC2B centrifuge has been used with identical results. 100 p1 of the sarkosyl- soluble supernatant were added to 50 p1 of gel sample buffer (0.2 M Tris-HC1, pH 6.8 ; 3% w/v sodium dodecylsulphate; 30'x vjv glycerol; 0.002 bromo- phenol blue).
To study binding to the outer membrane, the sarkosyl-insoluble fraction from above was re-ex- tracted with 2 ml of 1% sarkosyl for 5 min at room temperature, pelleted by centrifugation at 100000 x g for 40 rnin at 10 "C, and resuspended in 200 pl of distilled water. 100 pl was added to 50 p1 of gel sample buffer. All samples were then fractionated by dodecyl- sulphate-polyacrylamide slab gel electrophoresis as described below.
Experiments with other P-['4C]lactams were per- formed in a similar manner.
Slab Gel Electrophoresis and Detection of Penicillin-Binding Proteins
20 pl of 2-mercaptoethanol were added to each of the above samples immediately before they were heated for 3 rnin in a boiling water bath. 50-65 p1 of each sample (containing about 200 pg protein) were loaded into the gel slots of a 10% sodium dodecyl- sulphate polyacrylamide slab gel. The apparatus and the discontinuous buffer system have been described in detail by Laemmli and Favre [16]. Electrophoresis was performed at a constant current of 20 mA (initial voltage of about 60 V increasing to about 200 V at the end of electrophoresis) until the bromophenol blue tracking dye had migrated 11 cm into the sepa- rating gel. The gel was removed from the apparatus, stained for 1 h with 0.1% Coomassie brilliant blue R (Sigma) in 50% methanol 10% acetic acid, diffusion- destained with several changes of 5% methanol 10% acetic acid, and prepared for fluorography (scintil- lation autoradiography) by the incorporation of the scintillant PPO into the gel matrix as described by Bonner and Laskey [17]. Staining with Coomassie brilliant blue was used only to monitor the quality of

B. G. Spratt 343
the protein fractionation on the gel since the stained bands were not visible clearly after fluorography. The PPO-impregnated gel was dried under vacuum on to Whatman number 17 filter paper and placed on Kodak RP Royal X-Omat X-ray film at -80 "C. X-ray films were developed in Kodak liquid X-ray developer for 5 rnin at 19 "C without agitation, washed in 3% acetic acid and fixed for 5 min at 19 "C in Kodak X-ray fixer. Conventional autoradiography (without incorporation of PPO) was used in one experiment and in this case Ilford industrial G X-ray film was used.
The use of fluorography is essential to obtain the maximal sensitivity of 14C detection on the X-ray film and even with this technique the exposure times required are still inconveniently long. The binding proteins can be detected by eye after about 10 days fluorography but a considerably longer exposure is required to obtain image densities sufficient for accurate densitometry of the minor proteins. Fluoro- graphy probably introduces some errors into the quantitisation of the results since the image density on the X-ray films is not linear with the amount of radioactivity applied [18]. This problem can be eliminated by pre-fogging of the X-ray films [18].
A further substantial increase in sensitivity is obtained by the removal of the outer membrane proteins before applying samples to the gels. This is possible as we have failed to detect any penicillin- binding proteins in the E.coli outer membrane. Although the outer membrane constitutes only about 50% of the total protein of the membranes, the majority of the outer membrane consists of three proteins with almost identical mobilities on dodecyl- sulphate-polyacrylamide gels [ 191 and this results in overloading when relatively small amounts of outer membrane protein are applied to the gels. In contrast, the inner membrane consists of many proteins in approximately similar amounts, with the result that much larger amounts of protein can be applied before distortion of the gels due to over- loading occurs. Extraction of the inner membrane is complete under the conditions that we use, however, an increase in the concentration of sarkosyl to at least 2% can be used without interfering with the running of the gels or significantly extracting outer membrane proteins.
Apparent molecular weights of the penicillin- binding proteins were measured by comparison of their mobilities on dodecylsulphate-polyacrylamide slab gels with that of eight proteins of known molecular weights. The proteins and their molecular weights were : P-galactosidase (130000), phosphorylase A (lOOOOO), bovine serum albumin (68 000), pyruvate kinase (57 OW), ovalbumin (43 000), glyceraldehyde-3- phosphate dehydrogenase (36000), carbonic anhydrase (29000), and chymotrypsinogen (25 700).
Measurement of the Binding of P-Lactams by Competition with Benzyl[14C]penicillin
The binding of non-radioactive P-lactams was studied by measuring their competition for the binding of ben~yl['~C]penicillin to each of the proteins. 200 pl of membranes (5 - 10 mg protein/ml in 50 mM sodium phosphate buffer pH 7.0) were pre-incubated with 10 p1 of either distilled water or dilutions of the non-radioactive p-lactam for 10 rnin at 30 "C and then 20 pl of 50 pCi/ml ben~yl[~~C]penicillin were added for a further 10 min at 30 "C and the binding terminated by the addition of 5 p1 of non-radioactive benzylpenicillin (120 mg/ml) and 10 p1 of 20% (w/v) sarkosyl as described above. The final concentration of ben~yl['~C]penicillin (30 pg/ml) was approximately three times that required to obtain saturation of all binding proteins under the conditions used. Samples were fractionated by electrophoresis and the level of ben~yl['~C]penicillin bound to each protein measured as described above.
Release of Bound Benzyl[ ''C]penicilEin
To measure the spontaneous rate of release of the product of bound ben~yl['~C]penicillin (which is not benzylpenicillin itself but probably either benzyl- penicilloic acid [9] or phenacetylglycine and thia- zolidine ring fragmentation products [20, 21]), mem- branes (5 - 10 mg protein/ml in 50 mM sodium phos- phate buffer pH 7.0) were labelled with benzyl[14C] penicillin (16.5 pg/ml final concentration) for 10 rnin at 30 "C; a 1000-fold excess of non-radioactive benzylpenicillin was added, and samples of 200 p1 removed immediately and at intervals during con- tinued incubation at 30 "C into tubes containing 10 p1 of 20% (w/v) sarkosyl. Denaturation of the penicillin- binding proteins in sarkosyl prevented further release of ben~yl['~C]penicillin since release is enzymic (see Results). The samples were fractionated by electro- phoresis and fluorographs prepared as described above. The spontaneous release of other f i-[ '"C] lactams was measured in a similar fashion.
Various compounds (e. g. 2-mercaptoethanol, neu- tral hydroxylamine) were tested for their effect on the spontaneous rate of ben~yl['~C]penicillin release by adding them immediately after the excess non-radio- active benzylpenicillin and sampling as above into sarkosyl. In some experiments where the excess benzylpenicillin was known, or suspected, to react with the test compound, the further binding of ben~yl['~C]penicillin was prevented by the addition of 0.1 vol. of penicillinase (100000 units/ml) for 5 min at 30 "C. Penicillinase does not remove bound penicillin.

344 Penicillin-Binding Proteins of Escherichia coli
Quantit isat ion of Penicillin- Binding Proteins by Densitometry of X-Ray Films
X-ray films of sufficient image density for accurate quantitisation (but not so dense as to approach saturation of the film emulsion) were scanned with a Joyce-Loebl Mkl 1 lc microdensitometer. The peaks obtained were carefully cut out and weighed. Cor- rections were made, when necessary, for differences in widths of the gel slots.
Measurement of Minimal inhibitory Concentrations of ' p- Lac tams
Minimdl inhibitory concentrations weredetermined using E. coli KN126 growing exponentially at 37 "C in penassay broth with initial cell densities of approximately 2 x lo7 organisms/ml. The minimal inhibitory concentration was defined as the lowest concentration of the p-lactam which produced a clear alteration of the normal rod morphology of the cells (depending on the 8-lactam this could be the appear- ance of cell lysis, filamentation or ovoid cells).
Chemicals
Ben~yI['~C]penicillin was purchased from the Radiochemical Centre, Amersham, England (specific activity 53 - 54 Ci/mol). ['4C]Cefoxitin (2.8 Ci/mol) and ['4C]cephacetrile (4.6 Ci/nioI) were generously provided by Dr Edward Stapley (Merck Sharp and Dohme, Rahway, New Jersey, U.S.A.) and Dr W. Zimmermann (Ciba-Geigy, Basel, Switzerland) re- spectively. All three radioactive p-lactams were of high purity and were labelled in the R, sidechain (position 6 of penicillins ; position 7 of cephalosporins). Reference samples of 8-lactams were kindly provided by the following companies: ampicillin by Bristol Laboratories (Syracuse, New York, U.S.A.), cepha- cetrile and cephradine by Ciba-Geigy Ltd (Basel, Switzerland), mecillinam by Leo Laboratories, (Ballerup, Denmark), cephalexin, cephaloridine and cephalothin by Eli Lilly andCo. (Indianapolis, Indiana, U.S.A.), cefoxitin by Merck, Sharp and Dohme (Rahway, New Jersey, U.S.A.), benzylpenicillin, cephalosporin G and penicillin V by Squibb and Sons (Princeton, New Jersey, U.S.A.). /j-['"C]Lactams were stored as solids at -2O';C and were dissolved in 10 mM sodium phosphate buffer pH 7.0 immediately before use. Aqueous solutions were stored at - 80 "C and could be used for at least a week without the appearance of non-specific binding problems due to degradation products of the fl-lactams. Solutions of nonradioactive p-lactams were always prepared fresh immediately before each experiment. The solid com- pounds were stored desiccated at - 20 "C.
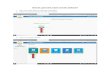
Protein
1
2 3
5 6
Fig. 1 . Detection uf rhe E. coli penicillin-binding proteins utzd their location in the inner memhrune. 40 p1 of ben~yl['~C]penicillin (31 pg/ml final concentration) were added to 400 pl of membranes for 10 min at 30 "C. To measure penicillin-binding proteins in the total membrane 100 pI of the assay mixture was added to 50 pl of gel sample buffer. A further 200 p1 were added to 10 pl of 20"/> (w/v) sarkosyl and the sarkosyl-soluble (Inner membrane) and sarkosyl-insoluble (outer membrane) fractions were obtained as described in Methods. All three samples were fractionated on a 10% polyacrylamide slab gel and a fluorograph prepared after exposure of the gel against Kodak RP Royal X-Omat X-ray film for 32 days at -80 'C. (A) Total membrane; (B) outer membrane; (C) inner membrane
Penicillinase was obtained from Wellcome Reagents Ltd (Beckenham, England). Sarkosyl NL-97 was purchased from Geigy Industria1 Chemicals (New York, U.S.A.).
RESULTS
All the penicillin-binding proteins are located in the E.coli inner membrane, defined as that fraction of the cell envelope which is solubilised by lo( sarkosyl at room temperature. Fig.1 shows the binding of ben~yl['~C]peniciIlin to total cell envelopes and to the sarkosyl-soluble and sarkosyl-insoluble fractions. No penicillin-binding proteins have been detected in the sarkosyl-insoluble (outer membrane) fraction with any of the P-['4C]lactams that we have studied. In all further experiments only the sarkosyl-soluble (inner membrane) fraction was analysed for penicillin- binding proteins. All the proteins are accessible to ben~yI[~~C]penicillin in whole cells since they could be detected in membranes prepared from whole cells which had been incubated with ben~yl['~C]penicillin.

B. G. Spratt 345
In this experiment mercaptoethanol was omitted during sonication as this compound releases bound penicillin (see below).
Since several of the chemicals which are used in dodecylsulphate-polyacrylamide gel electrophoresis (e. g . ammonium persulphate, 2-mercaptoethanol) have been reported to release bound penicillin [12], we have re-investigated the effects of these compounds on the recovery of the penicillin-binding proteins. 2-Mer- captoethanol released bound penicillin rapidly from proteins 5 and 6 if it was added to the binding assay mixture before the addition of sarkosyl (see below). However this release was clearly enzymic since it was completely prevented by denaturation of the proteins in 1 % sarkosyl. Once denatured, we could detect no differences in the levels of the penicillin- binding proteins in samples which had been boiled in gel sample buffer in the presence or absence of 10% (v/v) 2-mercaptoethanol (data not shown). Similarly we could find no effect of ammonium persulphate (0.1%) on the release of bound penicillin once the proteins had been denatured in sarkosyl.
In earlier experiments [13] we included 10 mM MgC1, in the penicillin-binding protein assay mixture but since we find no stimulation of binding by Mg2+, or any inhibition of binding by 1 mM EDTA, we now omit MgCl, from the assay. Pre-incubation of samples of membrane with N-ethylmaleimide (5 mM) or p - hydroxymercuribenzoate (1.5 mM) for 10 min at 30 "C did not prevent binding to any of the proteins.
The pattern of six proteins is extremely repro- ducible. In some gels additional minor penicillin- binding proteins were observed. These bands were in general non-reproducible, although two bands, proteins 7 and 8, with molecular weights 29000 and 32000 were apparent in several gels. An identical pattern of penicillin-binding proteins was found in all E.coli K12 strains that we have examined. E.coli B/r strains also showed an identical pattern although protein 7 was more prominent. The apparent molecular weights of the proteins were determined by comparing their mobility on a dodecylsulphate-polyacrylamide slab gel with that of eight proteins of known molecular weights. Table 1 shows the apparent molecular weights, and the relative abundance of each penicillin-binding protein, as well as an estimate of the number of molecules of each protein per bacterium (see below).
The penicillin-binding proteins are minor com- ponents of the E.coli inner membrane. Fig.2 shows the location of these proteins on a Coomassie-blue- stained gel of the inner membrane (note the absence of the characteristic major outer membrane proteins [19] with molecular weights 36000 on this gel of the sarkosyl-soluble membrane). The major proteins (5 and 6) can be tentatively identified as minor stained bands but the others are below the resolution of detection as stained bands.
Table 1. MoJecufar weights and relative abundance of the penicillin- binding proteins
Protein Apparent Binding Molecules/cell" molecular of weight penicillin
%total
1 91 OOO 8.1 230 2 66 000 0.7 20 3 60000 1.9 50 4 49000 4.0 110 5 42000 64.7 1800 6 40000 20.6 570
a See Results for details
Kinetics of Binding of p-[ 14C]Lactams
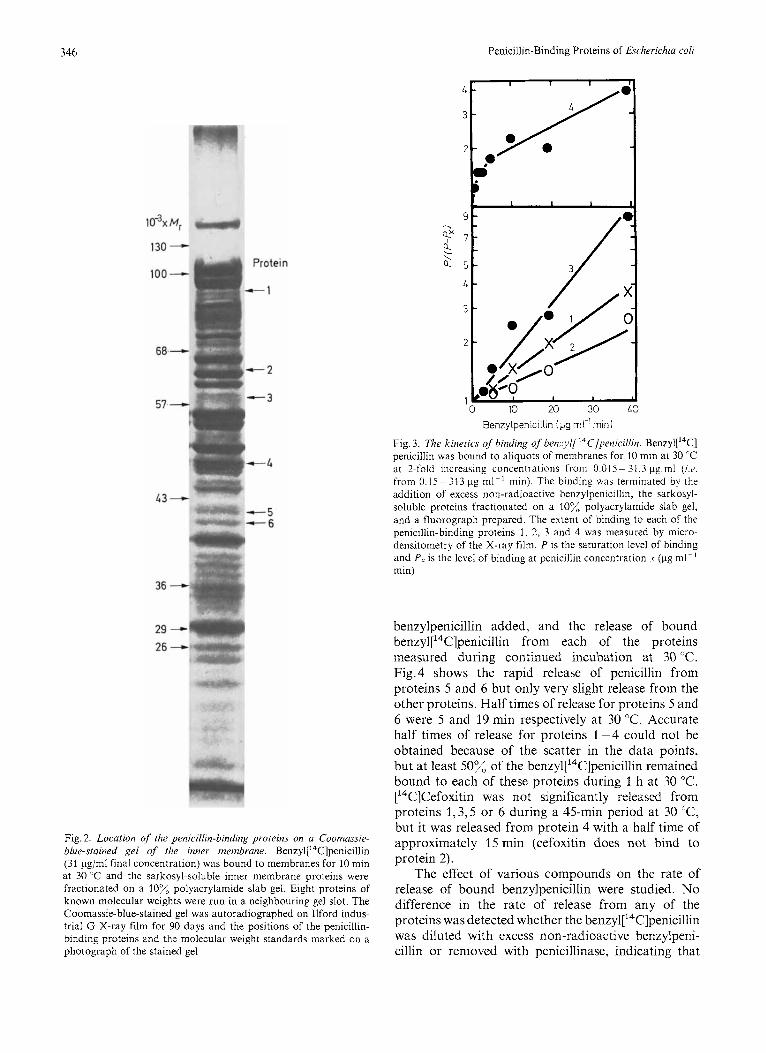
Benzyl[14C]penicillin was added in 2-fold in- creasing concentrations (final concentrations of 0.01 5 -31.3 pg/ml) to aliquots of membranes for 10 min at 30 "C and the extent of binding to each of the proteins measured as described in Methods. The binding of penicillin to proteins 1-4 was treated as a simple bimolecular irreversible reaction. For such a reaction the degree of acylation by penicillin should be proportional to the product of the penicillin concentration and the time of incubation [22]. Fig.3 shows that proteins 1 - 3 bind penicillin in accord with the above kinetics, but protein 4 shows more complex kinetics. Proteins 5 and 6 were not plotted in this way as they released bound penicillin with half lives which are of the same order as the incubation time of the assay (see below). Similar kinetics of ben~yl['~C]penicillin binding were obtained when a fixed concentration of penicillin was bound for increasing incubation times.
Saturation of all penicillin-binding proteins was essentially complete at 80 pg ml - ' min. The con- centration of ben~yl['~C]penicillin in our standard binding assay was over 3.5 times this level (313 pg ml-' min) to ensure saturation of all penicillin- binding proteins.
Although experiments with ['4C]cefoxitin and ['4C]cephacetrile were undertaken (see below) the specific activity of these materials was too low to obtain images on the X-ray films of sufficient density for accurate quantitisation. It was, however, observed that [14C]cephacetrile bound to the same six proteins as ben~yl['~C]penicillin and that ['4C]cefoxitin bound to all except protein 2. No additional penicillin- binding proteins were observed with any p-['"C] lactams.
Reversihility of the Binding of p-[ ''C']Lnctan~.s The penicillin-binding proteins were saturated
with benzyl['"C]penicillin, excess non-radioactive
346 Penicillin-Binding Proteins of Escherichia coli
1 M,
130-
100- Protein
-1
-2
- -3 57. --c
L3 - 4-5 4-6
36 - 29 - 26 -
Benzylpenicillin Ipg rn-lrnin)
Fig. 3. The kinetics of binding of benzyl[ ''C]penicillin. Ben~yl[ '~C] penicillin was bound to aliquots of membranes for 10 min at 30 "C at ?-fold increasing concentrations from 0.015- 31.3 pg in1 (w. from 0.15 - 31 3 pg m1-l min). The binding was terminated by the addition of excess non-radioactive benzylpenicillin, the sarkosyl- soluble proteins fractionated on a lox, polyacrylamide slab gel, and a fluorograph prepared. The extent of binding to each of the penicillin-binding proteins 1, 2, 3 and 4 was measured by micro- densitometry of the X-ray film. P is the saturation level of binding and P, is the level of binding at penicillin concentration .Y (pg ml- ' min)
benzylpenicillin added, and the release of bound ben~yl[~~C]penicillin from each of the proteins measured during continued incubation at 30 "C. Fig.4 shows the rapid release of penicillin from proteins 5 and 6 but only very slight release from the other proteins. Half times of release for proteins 5 and 6 were 5 and 19 min respectively at 30 "C. Accurate half times of release for proteins 1-4 could not be obtained because of the scatter in the data points, but at least 50% of the benzyl['4C]penicillin remained bound to each of these proteins during 1 h at 30 "C. [14C]Cefoxitin was not significantly released from proteins 1,3,5 or 6 during a 45-min period at 30 "C, but it was released from protein 4 with a half time of approximately 15 min (cefoxitin does not bind to protein 2).
The effect of various compounds on the rate of release of bound benzylpenicillin were studied. NO difference in the rate of release from any of the proteins was detected whether the ben~yl['~C]penicillin was diluted with excess non-radioactive benzylpeni- cillin or removed with penicillinase, indicating that
Fig. 2. Location of the penicillin-binding proteins on a Coomassie- blue-stained gel of the inner membrane. Ben~yl['~C]penicillin (31 pg/ml final concentration) was bound to membranes for 10 min at 30 'C and the sarkosyl-soluble inner membrane proteins were fractionated on a 10% polyacrylamide slab gel. Eight proteins of known molecular weights were run in a neighbouring gel slot. The Coomassie-blue-stained gel was autoradiographed on Ilford indus- trial G X-ray film for 90 days and the positions of the penicillin- binding proteins and the molecular weight standards marked on a photograph of the stained gel
341
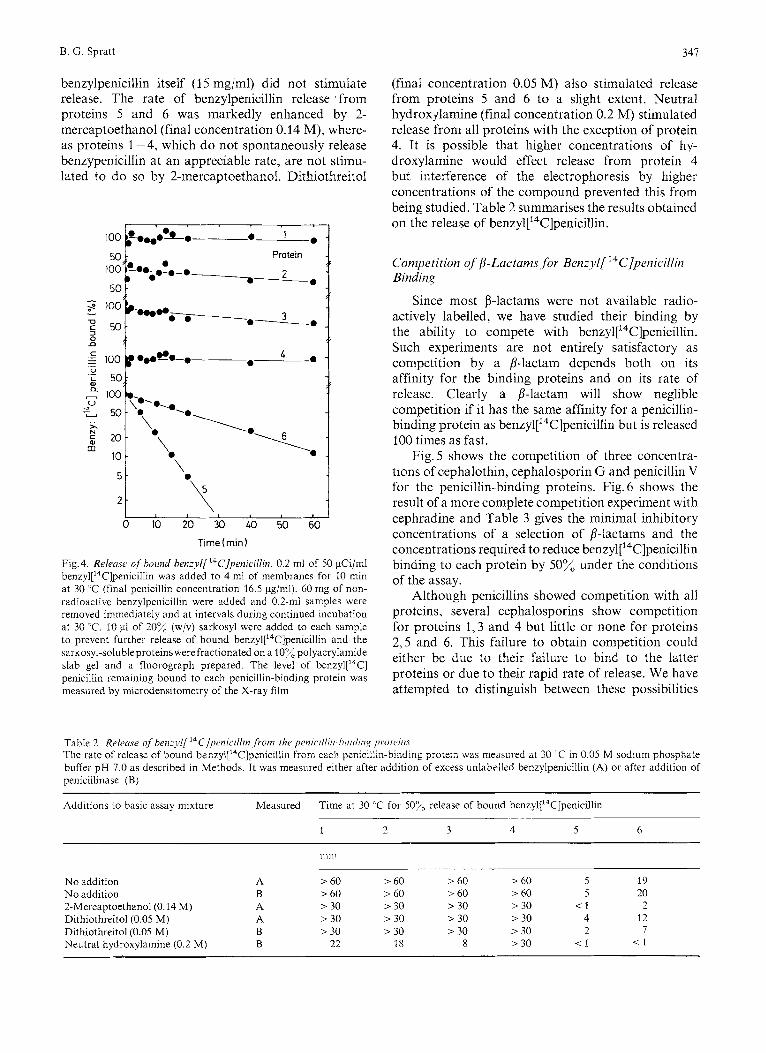
benzylpenicillin itself (1 5 mg/ml) did not stimulate release. The rate of benzylpenicillin release from proteins 5 and 6 was markedly enhanced by 2- mercaptoethanol (final concentration 0.14 M), where- as proteins 1-4, which do not spontaneously release benzypenicillin at an appreciable rate, are not stimu- lated to do so by 2-mercaptoethanol. Dithiothreitol
Protein 0
------a&. 5 0
$ 100 --0 0 0
1
0 - 0
V
x
0 \
Time(rnin1
Fig. 4. Releusr of hound henzyl[ ‘“C]penici/lin. 0.2 ml of 50 pCi/ml benzyl[14C]penicill~n was added to 4 ml of membranes for 10 min at 30 “C (final penicillin concentration 16.5 pg/ml). 60 mg of non- radioactive benzylpenicillin were added and 0.2-ml samples were removed immediately and at intervals during continued incubation at 30 “C. 10 pl of 20% (w/v) sarkosyl were added to each sample to prevent further release of bound benzyl[14C]penicillin and the sarkosyl-soluble proteinswerefractionated on a 10% polyacrylamide slab gel and a fluorograph prepared. The level of ben~yl [ ’~C] penicillin remaining bound to each penicillin-binding protein was measured by microdensitometry of the X-ray film
(final concentration 0.05 M) also stimulated release from proteins 5 and 6 to a slight extent. Neutral hydroxjlamine (final concentration 0.2 M) stimulated release from all proteins with the exception of protein 4. It is possible that higher concentrations of hy- droxylamine would effect release from protein 4 but interference of the electrophoresis by higher concentrations of the compound prevented this from being studied. Table 2 summarises the results obtained on the release of ben~yl[~~C]penicillin.
Competition of’/3-Lactams,for Benzyl[ ‘‘C]penicillin Binding
Since most B-lactams were not available radio- actively labelled, we have studied their binding by the ability to compete with ben~yl[’~C]penicillin. Such experiments are not entirely satisfactory as competition by a 8-lactam depends both on its affinity for the binding proteins and on its rate of release. Clearly a p-lactam will show neglible competition if it has the same affinity for a penicillin- binding protein as ben~yl[‘~C]penicillin but is released 100 times as fast.
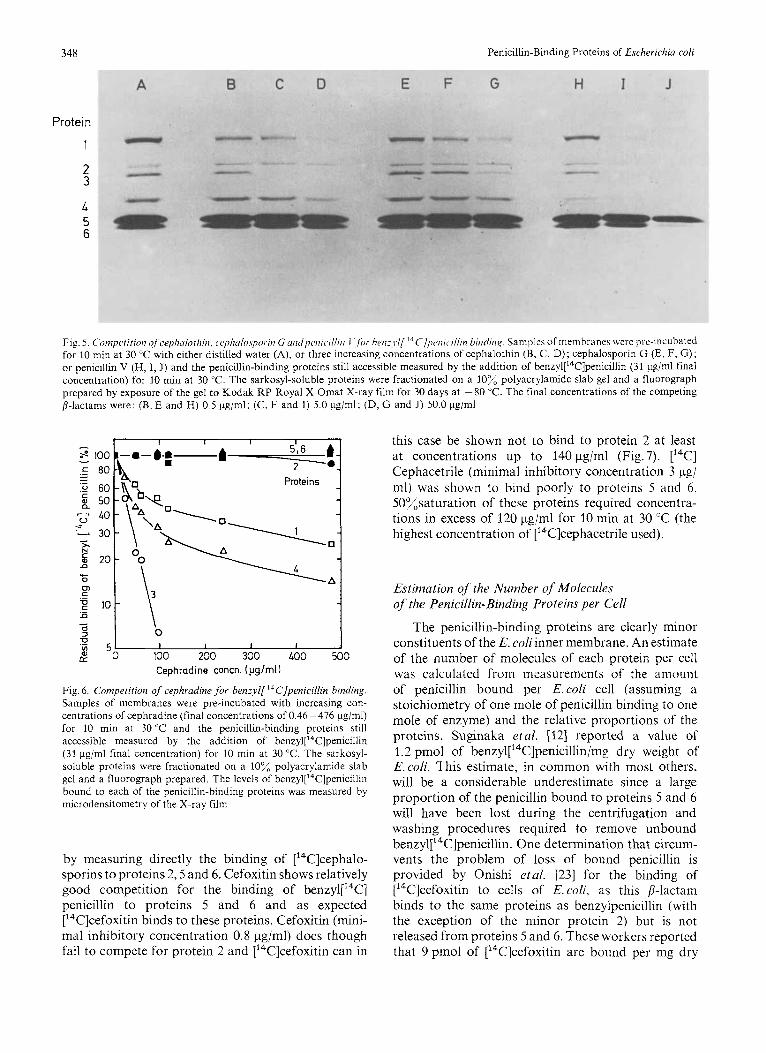
Fig.5 shows the competition of three concentra- tions of cephalothin, cephalosporin G and penicillin V for the penicillin-binding proteins. Fig. 6 shows the result of a more complete competition experiment with cephradine and Table 3 gives the minimal inhibitory concentrations of a selection of 8-lactams and the concentrations required to reduce ben~yl[’~C]penicilIin binding to each protein by 50% under the conditions of the assay.
Although penicillins showed competition with all proteins, several cephalosporins show competition for proteins 1,3 and 4 but little or none for proteins 2,5 and 6. This failure to obtain competition could either be due to their failure to bind to the latter proteins or due to their rapid rate of release. We have attempted to distinguish between these possibilities
Table 2. Relrme ofhenzyl[ ’‘C]penicillin ,from the pmic~illiii-hiiitliii,~ prorc’iii.r The rate of release of bound ben~yl[’~C]penicillin from each penicillin-binding protein was measured at 30 ’C in 0.05 M sodium phosphate buffer pH 7.0 as described in Methods. It was measured either after addition of excess unlabelled benzylpenicillin (A) or after addition of penicillinase (B)
Additions to basic assay mixture Measured Time at 30 ‘C for 50% release of bound ben~yl[’~C]penicillin ~~
1 2 3 4 5 6
No addition A > 60 > 60 > 60 > 60 5 19 No addition B > 60 > 60 > 60 > 60 5 20 2-Mercaptoethanol(O.14 M) A > 30 > 30 > 30 > 30 < 1 2 Dithiothreitol(O.05 M) A > 30 > 30 > 30 > 30 4 12 Dithiothreitol(O.05 M) B > 30 > 30 > 30 > 30 2 7 Neutral hydroxylamine (0.2 M) B 22 18 8 > 30 < l < I
348 Penicillin-Binding Proteins of Escherichia coli
Protein
1
2 3
4 5 6
Fig. 5 C'omperition of cephalothin, cc~phalospor.itr G untlpiwic~illiri I hw:l.l/ ' 'C/p~wr~.~l l in bindiii,q. Samples o f mein hi-anes wei-e pre-incubated for 10 min at 30 "C with either distilled water (A), or three increasing concentrations of cephdlothin (B, C, D); cephalosporin G (E, F, G); 01- penicillin V (H, I, J) and the penicillin-binding proteins still accessible measured by the addition of ben~yl['~C]penicilIin (31 pg/ml final concentration) for 10 min at 30 "C. The sarkosyl-soluble proteins were fractionated on a 10% polyacrylamide slab gel and a fluorograph prepared by exposure of the gel to Kodak RP Royal X-Omat X-ray film for 30 days at -80 "C. The final concentrations of the competing 8-lactams were: (B, E and H) 0.5 pg/ml; (C, F and I ) 5.0 pg/ml; (D, G and J) 50.0 pg/ml
- 0
z I 1 1 I
fY % 50 100 200 300 400 500 Cephradine concn. (pg/ml)
Fig. 6. Competition of cephradine for benzyl['4C]pmicillin binding. Samples of membranes were pre-incubated with increasing con- centrations of cephradine (final concentrations of 0.46 -476 pg/ml) for 10 min at 30 'C and the penicillin-binding proteins still accessible measured by the addition of benzyl["T]penicillin (31 pg/ml final concentration) for 10 min at 30 "C. The sarkosyl- soluble proteins were fractionated on a 10% polyacrylamide slab gel and a fluorograph prepared. The levels of ben~yl['~C]penicillin bound to each of the penicillin-binding proteins was measured by microdensitometry of the X-ray film
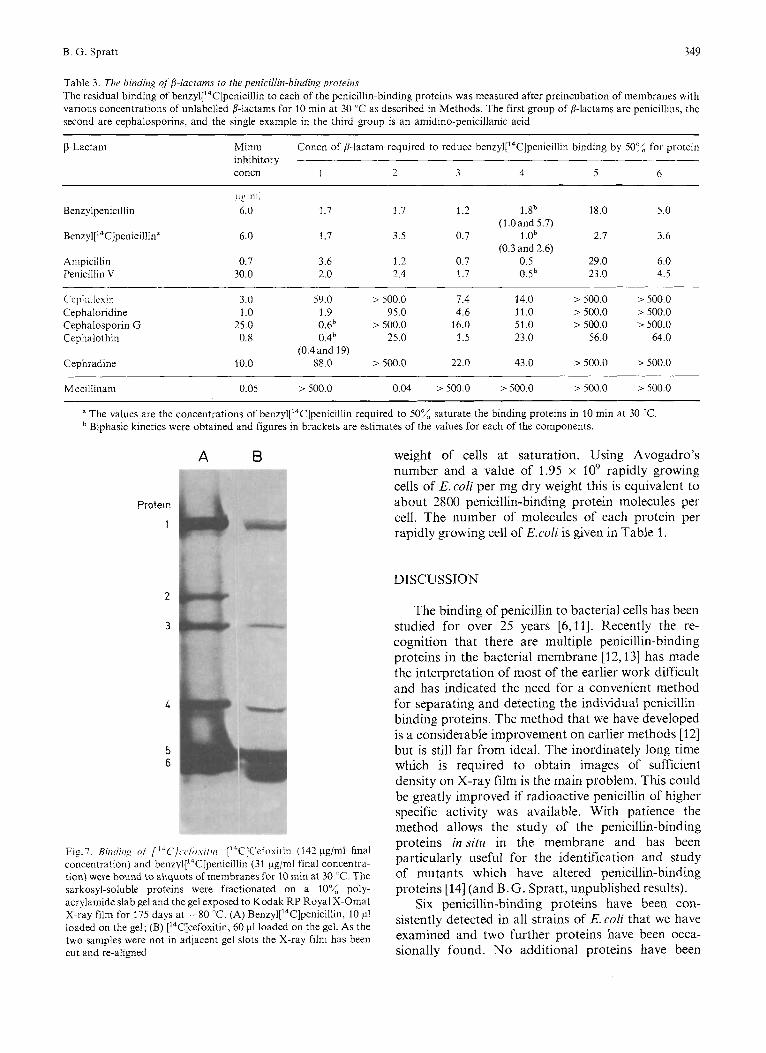
this case be shown not to bind to protein 2 at least at concentrations up to 140 pg/ml (Fig.7). ['"C] Cephacetrile (minimal inhibitory concentration 3 pg/ ml) was shown to bind poorly to proteins 5 and 6. 50%saturation of these proteins required concentra- tions in excess of 120 pg/nil for 10 min at 30 "C (the highest concentration of [14C]cephacetrile used).
Estimation of the Number c?fMolecules of the Penicillin-Binding Proteins per Cell
The penicillin-binding proteins are clearly minor constituents of the E. coliinner membrane. An estimate of the number of molecules of each protein per cell was calculated from measurements of the amount of penicillin bound per E.coli cell (assuming a stoichiometry of one mole of penicillin binding to one mole of enzyme) and the relative proportions of the proteins. Suginaka etal. [12] reported a value of 1.2 pmol of benzyl['4C]penicillin/mg dry weight of E. coli. This estimate, in common with most others, will be a considerable underestimate since a large proportion of the penicillin bound to proteins 5 and 6 will have been lost during the centrifugation and washing procedures required to remove unbound ben~yl['~C]penicillin. One determination that circum-
by measuring directly the binding of [14C]cephalo- vents the problem of loss of bound penicillin is sporins to proteins 2,5 and 6. Cefoxitin shows relatively provided by Onishi etal. 1231 for the binding of good competition for the binding of benzyl[14C] ['4C]cefoxitin to ceils of E.coli, as this /I-Iactam penicillin to proteins 5 and 6 and as expected binds to the same proteins as benzylpenicillin (with ['4C]cefoxitin binds to these proteins. Cefoxitin (mini- the exception of the minor protein 2) but is not ma1 inhibitory concentration 0.8 pg/ml) does though released from proteins 5 and 6. These workers reported fail to compete for protein 2 and ['"Clcefoxitin can in that 9 pmol of ['"Clcefoxitin are bound per mg dry
B. G. Spratt 349
Table 3 . Tlv binding of 8-luctarns to the penicillin-binding proteins The residual binding of benzyl['4C]penicillin to each of the penicillin-binding proteins was measured after preincubation of membranes with various concentrations of unlabelled fl-lactams for 10 rnin at 30 'T as described in Methods. The first group of B-lactams are penicillins, the second are cephalosporins, and the single example in the third group is an amidino-penicillanic acid
~~~~ ~
Minm inhibitory concn 1 - 3 4 5 6
Concn of 8-lactam required to reduce benzyl['4C]penicillin binding by 50% for protein
7
pg 1111
Benzylpenicillin 6.0 1.7 1.7 1.2 l.Sb 18.0 5.0
Ben~yl['~C]penicilIin" 6.0 1.7 3.5 0.7 1 .Ob 2.7 3.6
Ainpicillin 0.7 3.6 1.2 0.7 0.5 29.0 6.0 Penicillin V 30.0 2.0 2.4 1.7 0.5b 23.0 4.5
(1 .O and 5.7)
(0.3 and 2.6)
~ 'cpl l ' l lc~l l l Cephaloridine Cephalosporin G Cephalothin
Cephradine
> 500.0 > 500.0 3.0 59.0 > 500.0 7.4 14.0 > 500.0 > 500.0 1 .o 1.9 95.0 4.6 11.0 > 500.0 > 500.0 25.0 0.6b > 500.0 16.0 51.0
0.8 0.4b 25.0 1.5 23.0 56.0 64.0
> 500.0 > 500.0 (0.4and 19)
10.0 88.0 > 500.0 22.0 43.0
Mecillinam 0.05 > 500.0 0.04 > 500.0 >500.0 > 500.0 > 500.0
a The values are the concentrations of ben~yl['~C]penicillin required to 50% saturate the binding proteins in 10 min at 30 'C. Biphasic kinetics were obtained and figures in brackets are estimates of the values for each of the components.
A B
Protein
1
2
3
5 6
Fig.7. Riritii/i,g of [ ' * C ] w / o \ - / f r / i . ['*C]C'ei",ui~in ( 142 pg/ml final concentration) and ben~yl['~C]penicillin (31 pg/ml final concentra- tion) were bound to aliquots of membranes for 10 min at 30 ' C. The sarkosyl-soluble proteins were fractionated on a 10% poly- acrylamide slab gel and the gel exposed to Kodak RP Royal X-Omat X-ray film for 175 days at -80 "C. (A) Ben~yl['~C]penicillin, 10 pi loaded on the gel; (B) ['4C]cefoxitin, 60 pl loaded on the gel. As the two samples were not in adjacent gel slots the X-ray film has been cut and re-aligned
weight of cells at saturation. Using Avogadro's number and a value of 1.95 x lo9 rapidly growing cells of E. coli per mg dry weight this is equivalent to about 2800 penicillin-binding protein molecules per cell. The number of molecules of each protein per rapidly growing cell of E.coli is given in Table 1.
DISCUSSION
The binding of penicillin to bacterial cells has been studied for over 25 years [6,11]. Recently the re- cognition that there are multiple penicillin-binding proteins in the bacterial membrane [I 2,131 has made the interpretation of most of the earlier work difficult and has indicated the need for a convenient method for separating and detecting the individual penicillin- binding proteins. The method that we have developed is a considerable improvement on earlier methods [12] but is still far from ideal. The inordinately long time which is required to obtain images of sufficient density on X-ray film is the main problem. This could be greatly improved if radioactive penicillin of higher specific activity was available. With patience the method allows the study of the penicillin-binding proteins insitu in the membrane and has been particularly useful for the identification and study of mutants which have altered penicillin-binding proteins [14] (and B.G. Spratt, unpublished results).
Six penicillin-binding proteins have been con- sistently detected in all strains of E. coli that we have examined and two further proteins have been occa- sionally found. No additional proteins have been

350 Penicillin-Binding Proteins of Escherichia coli
detected with other @-['4C]lactams ; these compounds bound to either all or some of those proteins which bound ben~yl['~C]penicillin. The inner membrane location of the proteins is to be expected for enzymes catalysing the terminal stages of peptidoglycan meta- bolism.
Some correlation between penicillin-sensitive enzymes and penicillin-binding proteins can be made in E. coli. D-Alanine carboxypeptidase 1 A [9] has been shown to be equivalent to proteins 5 and 6 [24]. The purified enzyme forms a doublet on dodecylsulphate- polyacrylamide gels (the two components having identical mobilities to proteins 5 and 6) and exhibits peptidoglycan transpeptidase activity as well as D- alanine carboxypeptidase 1 activity. It is not known whether the two components of the purified enzyme are separate polypeptides (perhaps one having trans- peptidase activity and the other carboxypeptidase activity) or if they differ only by minor modifications of the same polypeptide. The two components (proteins 5 and 6) are distinguishable in some properties, e.g. they have different rates of benzyl ['4C]penicillin release. A second penicillin-sensitive enzyme has been extensively purified (D-alanine carboxypeptidase 1B) and is unusual in that it apparently fails to bind penicillin [9] and would not therefore be detected as a penicillin-binding protein. Recently protein 4 has been shown to corre- spond to a penicillin-sensitive enzyme [25] but the enzymes which presumably correspond to proteins 1, 2 and 3 have not been identified yet.
In some of our experiments protein 1 appears to resolve into two proteins; this may be an artefact due to the overloading of the gels which was required to obtain maximal sensitivity of detection of the penicillin-binding proteins. The binding of some p- lactams (measured in competition experiments) to this protein was markedly biphasic and the possibility exists that protein 1 may be two proteins with very similar electrophoretic mobilities on our gels. Protein 4 also showed biphasic binding of ben~yl['~C]penicillin but this is probably not due to the existence of two distinct proteins with similar mobilities since a mutant has been obtained which fails to bind any benzyl [14C]penicillin to protein 4 [25]. Clearly such mutants would not be obtained readily if there were two distinct proteins at the position of protein 4 on the gel. The reason for the biphasic kinetics is not known.
Considerable efforts have been made to correlate the binding of penicillins and cephalosporins to their killing action [6,11,22,26,27]. The usual approach has been to compare the minimal inhibitory concentra- tions of the B-lactams with those required to obtain 50% binding to bacterial cells. Much of this work was done before it was recognised that bacteria have multiple penicillin-binding proteins and is difficult, and often impossible, to interpret. Even when the
individual penicillin-binding proteins are studied this approach has several problems and limitations. One difficulty arises from the comparison of minimal inhibitory concentration values, measured with cells growing at low densities, with the 50% binding values obtained using concentrated membrane suspensions. Clearly the two situations are not strictly comparable, especially since permeability barriers to the @-lactams exist in the former, but presumably not in the latter case. Furthermore, does inhibition of bacterial growth result when an essential protein is inhibited by 10% or by 90%? A second equally serious problem occurs in the measurement of the 50% binding values. The binding of a p-lactam to a protein depends on both its concentration and the length of the binding assay [22]; quite different values could be obtained if this latter parameter was altered. Also the rate of release of most B-lactams from the proteins is not known and this would lead to overestimates of the 50% binding values, obtained by competition for ben~yl[ '~C] penicillin binding, if the half time of release was short in relation to the duration of the assay. This effect explains the discrepancy between the value obtained for the direct binding of ben~yl['~C]penicillin to protein 5 with that obtained by competition (benzylpenicillin is rapidly released from protein 5) .
In spite of these limitations to the measurement of the binding of 6-lactams by competition, our results strongly suggest that the penicillins which we examined bind to all the binding proteins but that at least some cephalosporins show little or no binding to proteins 2,5 and 6, although they bind to the remaining proteins. This result was directly confirmed by the failure of [14C]cefoxitin to bind to protein 2 and by the low rate of binding of ['4C]cephacetrile to proteins 5 and 6. These cephalosporins show the lysis and inhibition of cell division common to most 8-Iactams, at concentrations where little or no binding to proteins 2,5 and 6 occurs. For this reason we believe that these penicillin-binding proteins are not the targets at which @-lactams bind to elicit these physiological effects. This does not necessarily imply that these proteins are inessential for cell growth and protein 2 has been demonstrated to be required for the growth of E.coli as a rod-shaped cell and to be the killing site for amidinopenicillanic acids [13,14]. The concentrations of 6-lactams which produce 50% competition for ben~yl['~C]penicillin binding to proteins 1,3 and 4 correlate reasonably well with the minimal inhibitory concentrations of these compounds, and the killing sites for typical a-lactams are probably amongst these penicillin-binding proteins.
A second, and much more promising, method for identifying the killing sites for @-lactam antibiotics involves theisolation ofmutants with altered penicillin- binding proteins. Using this method we have shown that protein 2 is involved in the production of ovoid

B. G. Spratt 351
cells by amidinopenicillanic acids and that protein 3 is the target at which P-lactams bind to inhibit cell division and cause filamentation [ 141. Recently mutants have been reported which lack protein4 and, since these mutants grow normally, it strongly suggests that this protein is not a killing site [25]. Only protein 1 remains as the site at which P-lactams bind to inhibit cell elongation and cause lysis. This assignment is in agreement with earlier work [14] but the isolation of a temperature-sensitive cell lysis mutant with a thermo- labile protein 1 would be the definitive test of this hypothesis. The possibility that penicillin-binding protein 1 may be two proteins complicates the issue. A further possibility is that lysis is the result of the inhibition of a penicillin-sensitive enzyme which does not bind penicillin covalently and so would not be detected as a penicillin-binding protein.
The finding that [I4C]cefoxitin and benzyl[14C] penicillin bind to precisely the same proteins (with the exception of protein 2) contradicts the failure of earlier experiments [23] to show competition of benzyl- penicillin for the binding of [I4C]cefoxitin to mem- branes of E.coli. This is probably explained by the inability of the rapidly released benzylpenicillin to compete effectively for binding to proteins 5 and 6 with the irreversibly bound [14C]cefoxitin and emphasises the dangers of making inferences about the binding of 8-lactams in experiments where total binding to membranes is measured.
Experiments on total binding of p-lactams, or measurements of total penicillin-sensitive activities, give little information on the mechanism of action of these antibiotics since their killings sites in E.coli and B. subtilis [22] are minor penicillin-binding proteins which presumably correspond to minor penicillin- sensitive enzymes.
The nature of the binding of penicillin to the proteins is unknown. Binding as a thioester has recently been favoured over earlier suggestions of an amide or ester linkage because of the release of bound penicillin under conditions where the former, but not the latter, would be cleaved [28]. The finding that release is enzymic [6] and does not occur with neutral hydroxylamine (or 2-mercaptoethanol) if the proteins are denatured by heat or anionic detergents decreases the strength of this argument. We have failed to prevent binding of penicillin with sulphydryl- blocking reagents (p-hydroxymercuribenzoate and N - ethylmaleimide) and similar results have been obtained for D-alanine carboxypeptidase 1 A (proteins 5 and 6) [9]. Whereas p-hydroxymercuribenzoate did not inhibit binding of penicillin to this enzyme, it did inhibit release 191. Sulphydryl groups may therefore be involved in the enzymic release of bound penicillin ; the stimulation of release by 2-mercaptoethanol, dithiothreitol and neutral hydroxylamine is consis- tent with this view. Whether penicillin binds directly
to a protein sulphydryl group is still an open question.
The product of benzylpenicillin release from D- alanine carboxypeptidase 1 A appears to be benzyl- penicilloic acid [9] and this enzyme therefore hdS true (but very weak) penicillinase activity. It is interesting that the p-lactam ase-resistan t derivative, cefoxi tin, is not released from this enzyme (proteins 5 and 6), as there has been speculation as to whether this enzyme could be an evolutionary precursor of P-lactamases ~ 2 , 91.
Part of this work was initiated in the laboratory of Dr Arthur B. Pardee at Princeton University (funded by U.S. Public Health Service Grant CA11595) and I am very grateful for his support and encouragement. Work carried out at Leicester University was funded by a project grant from the Medical Research Council. The gifts of ['4C]cefoxitin from Dr Edward 0. Stapley, [14C]cephacetrile from Dr W. Zimmermann and the technical assistance of Vinay Jobdnputra are gratefully acknowledged.
REFERENCES
1.
2.
3.
4.
5. 6.
7.
8.
9.
10.
11. 12.
13.
14. 15.
16. 17.
18
19 20.
21
22
Wise, E. M. & Park, J.T. (1965) Proc. Nut1 Acad. Sci. U.S.A. 54,
Tipper,D. J. & Strominger, J. L. (1965) Proc. Nut1 Acad. Sci.
Izaki,K., Matsuhashi, M. & Strominger,J. L. (1966) Proc. Natl
Mirelman, D., Bracha, R. & Sharon,N. (1974) Ann. N. Y . Acad
Ward,J.B. & Perkins,H.R. (1974) Biochem. J . 139, 781-784. Blumberg,P.M. & Strominger,J.L. (1974) Bacteriol. Rev. 38,
thgdanovsky. D., Bricas, E. & Dezelee. P. (1969) C.K Ilchd. Siances Acud. Sci. Ser. D, Sci. Nat. (Paris) 269, 390-393.
Hartmann,R., Holtje,J.-V. & Schwarz, U. (1972) Nature (Land.) 235,426-429.
Tamura.T., Imae,Y. & Strominger,J. L. (1976) J . Biol. Chem.
Nguyen-Disteche,M., Pollock, J. J., Ghuysen, J-M., Puig, J., Reynolds,P. E., Perkins,H. R., Coyette, J., & Salton, M. R. J. (1974) Eur. J . Biochem. 41, 457-463.
75-81.
U.S.A. 54, 1133-1141.
Acad. Sci. U.S.A. 55, 656-663.
Sci. 235,326 - 347.
291 -335.
251,414-423.
Cooper, P. D. (1956) Bacreriol. Rev. 20, 28 -48. Suginaka,H., Blumberg,P.M. & Strominger,J.L. (1972) 1.
Spratt,B.G. & Pardee,A.B. (1975) Nature (Lond.) 254,
Spratt,B.G. (1975) Proc. NutlAcad. Sci. U.S.A. 72,2999-3003. Filip.C., Fletcher,G., Wulff, J. L. & Earhart,C. F. (1973) J .
Laemmli,U.K. & Favre,M. (1973) J . Mol. Bzol. 80, 575-599. Bonner,W.M. & Laskey,R.A. (1974) Eur. J . Biochetn. 46,
Laskey,R.A. & Mills,A.D. (1975) Eur. J . Biochem. 56,
Hancock,R.E. W. & Braun,V. (1976) FEBSLett. 65,208-210. Hammarstrom,S. & Str0minger.J. L. (1975) P r r . Nofl Acad.
S5.i. U S . A . 72, 3463 - 3467. Frere, J-M., Ghuysen, J-M., Vanderhaeghe,H., Adriaens,P.,
Degelaen,T. & De Graeve,J. (1976) Nature (Lond.) 260, 451 -454.
Blumberg,P.M. & Strominger,J.L. (1972) J . Bid. Chern. 247, 8107 - 81 13.
Bid. Chern. 247, 5279-5288.
516-517.
Bacterial. 115. 71 7 - 722.
83 -88.
335 - 341.

352 B. G. Spratt : Penicillin-Binding Proteins of Escherichia coli
23. Onishi,H.R., Zimmermann,S.B. & Stapley,E.O. (1974) Ann.
24. Spratt,B.G. & Strominger,J.L. (1976) J . Bacteriol. 217, Sci. 235,300- 309.
26. Rogers,H.J. (1967) Biochem. J. 103,90-102. 27. Park, J.T., Edwards, J. R . & Wise, E.M. (1974) Ann. N . Y. Acad.
28. Lawrence,P.J. & Strominger,J.L. (1970) J . Biol. Chem. 245,
N . Y. Acud. Sci. 235.406-425.
660 - 663.
Suzuki,S., Nishiniura,Y., Ugino,U. & Hirota,Y. (1976) Proc. Narl Acad. Sci. U.S.A. in press.
25. Matsuhashi, M., Takagaki, Y ., Maruyama, 1. N., Tamaki,S., 3653 -3659.
B.G. Spratt, Department of Genetics, University of Leicester, University Road, Leicester, Great Britain, LE1 7RH