Embed Size (px)
Citation preview

PROPERTIES OF THE MILK FAT GLOBULE MEMBRANE
DERIVED FROM BUTTERMILKS FROM DIFFERENT SOURCES
A Thesis
Presen ted to
The Faculty of Graduate Studies
of
University of Guelph
by
MILENA CORREDIG
ln partial hilfilment of requirements
for the degree of
Doctor of Philosophy
January, 1998
O Milena Corredig, 1998

National Library I * m of Canada Bibliothèque nationale du Canada
Acquisitions and Acquisitions et Bibliographie Services services bibliographiques
395 Wellington Street 395, nie Wellington OtoiwaON K1AûN4 OttawaON KtAON4 Canada canada
The author has granted a non- L'auteur a accordé une licence non exclusive licence allowing the exclusive permettant à la National Library of Canada to Bibliothèque nationale du Canada de reproduce, loan, distribute or sell reproduire, prêter, distribuer ou copies of this thesis in microform, vendre des copies de cette thèse sous paper or electronic formats. la forme de microfiche/nIm, de
reproduction sur papier ou sur format électronique.
The author retains ownership of the L'auteur conserve la propriété du copyright in this thesis. Neither the droit d'auteur qui protège cette thèse. thesis nor substantial extracts ffom it Ni la thèse ni des extraits substantiels may be printed or otherwise de celle-ci ne doivent être imprimés reproduced without the author's ou autrement reproduits sans son permission. autorisation.

ABSTRACT
PROPERTIES OF THE MILK FAT GLOBULE MEMBRANE DERIVED FROM BUTTERMILKS FROM DWFFlRENT SOURCES
Milena Corredig University of Guelph, 1998
Advisor Professor D. G. Dalgleish
The objective of this research was to study the composition of industrial
buttermilk and its use as an ingredient in oil-in-water ernulsions. Industrial buttermïlks
were found to contain large arnounts of caseins and whey proteins and also material
denved from the milk fat globule membrane (MFGM). When buttermilk was used to
prepare oil-in-water emulsions, the behaviour of the final product was mainly determined
by the caseins, adsorbed in the highest amount at the interface. In these buttrnnilk
emulsions, skim m i k proteins did not show cornpetitive adsorption with the MFGM
matenai. To determine the role played by the membrane fraction, MFGM isolates were
prepared, and their emulsifying properties studied. These isolates were obtained from
buttermilks from indusuial processes, and from pasteunzed or unheated creams. The heat
ueatment of the cream affected the emulsifying properties of the MFGM isolates. A
large amount of P-lactoglobulin was associated with MFGM obtained from industrial
buttermilk and from heated creams as a result of heat-induced interactions occumng
between whey proteins and MFGM during cream pasteurization. Emulsions prepared
with MFGM isolate from unheated cream had smaiier droplet size distributions than
those of emulsions prepared with isolates from creams heated at temperatures > 65'C.
Significant differences in properties of the MFGM such as solubility, colour, iron content.
amount of whey protein associated and erndsifying power, were found to be related to
the extent of heat treatrnent. Some of the differences between the emulsions prepared

with the various MFGM isolates were elucidated using transmission electron microscopy.
The instability of emulsions prepared with MFGM isolates obtained from indusrrial
butter* could be ascribed to bridging of membrane fragments to more than one oil
droplet, causing their flocculation and coalescence. These structures differed from those
of emulsions prepared with MFGM isolates from unheated cream. The poor emulsifying
properties of MFGM isolates were improved by proteolysis, which either created more
surface active polypeptides or disrupted the bridges between the droplets. This
improvement was considerable in MFGM isolates from unheated crearn, but even greater
for MFGM obtained frorn industrial butterrnilk.

ACKNOWCLEDGMENTS
I wish to thank the Ontario Dairy Council, the Ontario Ministry of Agriculture
and Foods and the Natural Sciences and Engineering Council for fmancial support
Sincere thanks to ail my friends and colieagues, who are too many to mention
and some of them already working on other continents. Special thanks to Dr. Yuan Fang
for her encouragement, she is and always WU be not only a great fnend but a talenteci
woman to be considered as role model.
1 acknowledge the support of my committee members, Dr. Hill, Dr. Goff and
Dr. Pouiiot. Their help improved the outcome of my thesis work.
Great appreciation goes to my Advisor, Dr. Douglas Dalgleish, who served as
mentor throughout these years. Thank you for believing in me and for giving me a
chance. You contributed greatiy not only to my professional but also persona1
development.
1 dedicate this thesis to my father, who always took joy in my successes and who,
1 believe, is always beside me, giving me the energy to overcome difficult moments. 1
know he would be very proud of his daughter, his joy having always derived of his
children's success. 1 regret the fact that he did not leave enough time for me to fulfiil
some of his dreams.
A special thanks goes to my family, to my mother, Davide, Sabrina and m y little
nephew Nicoio. Their love at distance is appreciated.
1 do not have enough words to thank my beloved husband, Hugh, for his suppon
He decided to spend the rest of his life beside me; being married to a bright scientist wiii
make my Life very interesting and successful.

Table of Contents
Table of Contents
List of Tables
List of Figures
List of Abbreviations
1. GENERAL INTRODUCTION AND STATEMENT OF
THE OBJECTIVES
2. INTRODUCTION
2.1 COMPOSITION OF BUTTERMILK
2.1.1 The Milk Fat Globule Membrane
2.1.1.1 Origin and Structure
2.1.1.2 MFGM Composition
2.1.1.2.1 Protein Composition
2.1.1.2.2 Lipid Composition
2.1.2 Skim Milk Proteins
2.1.2.1 Whey Proteins
2.1.2.2 Casein Micelles
2.2 PROTEIN STABILIZED OIL-IN-WATER EMULSIONS
2.2.1 Emulsion Formation
2.2.2 Emulsion Stability
2.2.3 Proteins at OiVWater In terfaces
3. METHODOLOGIES
3.1 B r n R M A K I N G
3.2 ELECTROPHORESIS (SDS-PAGE)
3.3 MICROFLUIDIZATION

INTEGRATED LIGHT SCAITERING 32
MEASUREMENT OF ZETA POTENTIAL 37
PHOSPHOLIPID ANALYSIS: IATROSCAN 40
TRANSMISSION ELECTRON MICROSCOPY 41
BUTTERMILK PROPERTIES IN EMULSIONS WITH SOYABEAN OIL
AS AFFECTED BY FAT GLOBULE MEMBRANE-DE-D
PROTEINS
INTRODUCTION
MATERIALS AND METHODS
Transmission Electron Microscopy
Emulsion Preparation
Determination of Particle Size
Electrophoresis (SDS-PAGE)
RESULTS AND DISCUSSION
Composition of the Material
Emulsions Fonned Using Buttermilk
CONCLUSIONS
STUDIES OF EMULSIONS STABILIZED BY SKLM MILK
ISOLATES FROM INDUSTRIAL BUTTERMILK: EMULSIFYING
PROPERTIES OF MATERIAL DERIVED FROM THE MILK FAT
GLOBULE MEMBRANE
INTRODUCTION
MATERIALS AND METHODS
Preparation of MFGM Isolate
Transmission Electron Microscopy
Phos pholipid De termination
Em ulsion Preparation

5.2.5 Determination of Particle Size Distribution and Protein Adsorption 7 5
5.2.6 Electrophoresis (SDS-PAGE) 75
5.3 RESULTS AND DISCUSSION 7 5
5.3.1 Production of Isolates from Buttermilk and Their Composition 7 5
5.3.2 Emulsions Prepared with Isolates from Buttermilk 8 1
5.4 CONCLUSIONS 86
6. CHARACTERUATION OF THE INTERFACE OF AN OIL-IN-WATER
EMULSION STABILIZED BY PROTEIN DERIVED FROM THE MILK
FAT GLOBULE MEMBRANE 89
6.1 INTRODUCTION 89
6.2 MATERIALS AND METHODS Y0
6.2.1 Preparation of the Membrane Isolate Y 0
6.2.2 Emulsion Preparation 9 1
6.2.3 Measurement of Particle Size and Stability of the Emulsions 9 1
6.2.4 SDS-PAGE 92
6.2.5 Measurements of c-Potential 92
6.2.6 Exchange Studies 94
6.3 RESULTS 94
6.3.1 Protein Composition of the Isolates 94
6.3.2 Puticle Size Distribution and Stability of the Emulsions Y7
6.3.3 Determination of c-Potential 1 03
6.3.4 Exchange Studies 106
6.4 DISCUSSION 1 1 1
7. THE EFFECT OF HEATING OF CREAM ON THE FUNCTIONAL
PROPERTIES OF MILK FAT GLOBULE MEMBRANE ISOLATES 1 13
7.1 INTRODUCTION 113
7.2 MATERIALS AND METHODS 115

7.2.1 Heat Treatment of the Cream 115
7.2-2 Production of Buttermilk 115
7.2.3 Preparation of the Isolates 116
7.2.4 Emulsion Preparation 116
7.2.5 SDS-PAGE 117
7.2.6 Fat Globule Size Distribution 117
7.2.7 Size Exclusion Chromatography 117
7.2.8 Solubility Studies 118
7.2.9 Iron Determination 119
7.2.10 Statistical Analysis 119
7.3 RESULTS 119
7.3.1 Studies on Lndustrid Cream and Buttennik 119
7.3.1.1 Emulsifying Properties of Isolates from Buttermik prepared with Crenm
Heated by HTST (76-85'C) 119
7.3. 1 -2 Size Exclusion Chromatograp hy 134
7.3.2 MFGM Isolates Denved from C r e m Heated at Temperatures of 60'-80'C 126
7.3.2-1 Solubility Studies 127
7.3.2.2 Emulsifying Properties of the MFGM Isolates 130
7.3.2.3 Whey Proteins Associated with the MFGM 132
7.2.2.4 Iron Content 135
7.4 CONCLUSIONS 137
8. STUDIES ON MEMBRANE MATERIAL DERIVING FROM
BUTTERMLLKS OBTADED FROM DIFF'ERENT BUTTER
PROCESSES
8.1 INTRODUCTION
8.2 MATERIALS AND METHODS
8.2.1 Andytical Methods

Transmission Electron Microscopy 142
RESULTS 142
Emulsifying Properties of Isolates from Different Buttermilks L44
CONCLUSIONS 153
STUDIES ON THE SUSCEPTIBILITY OF MEMBRANE-DEmD
PROTEINS TO PROTEOLYSIS AS RELATED TO CHANGES IN THEIR
EMULSIFYING PROPERTES 156
INTRODUCTION 156
MATEMALS AND METHODS 158
MFGM Isolation 158
Emulsion Preparation 158
Hydrol ysis 158
9.2.3.1 Hydrolysis in Solution
9.2.3.2 S urface-H ydrolysis
9.2.4 SDS-PAGE
9.2.5 Emulsion Droplet Size
9.3 RESULTS
9.3.1 Susceptibility of MFGM to Proteolysis
9.3.2 Proteolysis at the OiWater Interface - Surface Hydrolysis
9.3.3 Particle Size Distribution of MFGM Isolates Emulsions
9.4 CONCLUSIONS
10. GENERAL CONCLUSIONS
11. REFERENCES

List of Tables
Table 2.1: Composition of buttermik 5
Table 2.2: Estimated average composition of natural milk fat globule membranes 8
Table 4.1: Comparative table summarizing the major differences observed between skirn
milk and buttemilk emdsions 70

List of Figures
Figure 2.1: Mode1 of the mi& fat globule membrane after secretion and after
reorganization in the acinar lumen
Figure 2.2: Different types of instability in emulsions
Figure 3.1: The laboratory-size chum used to prepare buttermilk
Figure 32: The process of phase inversion during the production of anhydrous
mik fat
Figure 3.3: Schernatic representation of the Mastersizer X
Figure 3.4: Schematic representation of the Stem's mode1 of the elecuicai double
layer
Figure 4.1: Typical SDS-PAGE of proteins obtained frorn skim mik, buttermik and
whole milk
Figure 4.2: Scanned TEM image of buttermilk
Figure 4.3: Average droplet size of emulsions ( 10% oil), related to buttemilk
concentration
Figure 4.4: Size distribution of ernulsion droplets containing 10% soyabran oil
Figure 4.5: Protein % relative of the total protein rnigrated in 2 0 9 hornogcnrous gel
electrophoresis
Figure 4.6: Typical SDS-PAGE polypeptide pattern of the aqueous phase, sepanted by
centrifugation, of 10% oil emulsions with different concentrations of buttermilk 55
Figure 4.7: Size distribution of ernulsion droplets stabilized with 12% buttemilk 57
Figure 4.8: Protein surface concentration as a function of total solids for emulsions
containing 10% soyabean oil 60
Figure 4.9: Changes in droplet size distribution of ernulsions with 10% oil 6 1
Figure 4.10: Size distribution of ernulsion droplets containing 1 0 9 oil and 0.25%. 1%
and 8% skim milk solids 63
vii

Figure 4.11: Average droplet size of 10% oil-in-water emulsions as a function of skirn
miik solids 64
Figure 4.12: Caseins, 8 relative to ihc totd casein present in the emulsion. adsorbed
ont0 the oil droplers as a function of skim milk solids 65
Figure 4.13: Typical SDS-PAGE of s e m phase, oil droplets and original emulsions
prepared with different concentrations of skim mik solids 67
Figure 4.14: Droplet size distribution of emulsions prepared with 8% skim m i k proteins
and IO and 20% soyabean oil 69
Figure 5.1: Typical SDS-PAGE of freeze dried matenal frorn buttermilk 77
Figure 5.2: Transmission electron micrognph of pellet isolated by centrifugation of
buttermiik 79
Figure 5.3: Lipid composition (%) as determined by TLC (Iatroscan) 80
Figure 5.4: Typicai particle size distribution of 10% oil-in-water emulsions 82
Figure 5.5: Average droplet size of emulsions as a function of MFGM isolate
concentration 83
Figure 5.6: Typicd SDS-PAGE gel of proteins obtained from samplrs of 10% oil-in-
water emulsions and resuspended cream 85
Figure 5.7: Changes in particle size distribution of 10% oil-in-water emulsion prepared
with 2.5% MFGM isolate and 2.5% MFGM isolate + 0.5% caseinrite 87
Figure 6.1: SDS-PAGE of MFGM and buttermilk isolates from unheated crem. The
migration profiie is aiso shown 96
Figure 6.2: Typical particle size distribution of 1 0 8 oil-in-water emulsions 98
Fipre 6.3: Average droplet size of 5% and 10% oil-in-water emulsions as a function of
the MFGM concentration used during homogenization 99
Figure 6.4: Typical particle size distribution of 10% oil-in-water emulsions prepared
with 0.25%, 0.5% and 1 % MFGM isolate 1 0 1
. . , V l l l

Figure 6.5: Size distribution of 1 0 1 soyabean oil-in-water emulsion droplets in relation
to storage, heat treatrnent and changes in pH 102
Figure 6.6: 5-Potential of emulsions prepared with 1%. 1 - 5 6 MFGM and 1 % lysozyme
and 10% 02, as a function of pH 104
Figure 6.7: SDS-PAGE of LO% oil-in-water emulsion prepared with 1% of MFGM
isolate from unheated cream 107
Figure 6.8: SDS-PAGE of 10% oil-in-water emulsion prepared with 0.5% casein 109
Figure 6.9: SDS-PAGE of protein adsorbed on a MFGM-stabilized interface as a
function of other proteins added to the emulsion 110
Figure 7.1: Typical droplet size distribution of 10% oil-in-water emulsions prepared
with buttermilk isolates 131
Figure 7.2: Typical droplet size distribution of 10% oil-in-water emulsions prepared with
MFGM isolates 122
Figure 7.3: SDS-PAGE of MFGM and buttermilk isolates from HTST creams 123
Figure 7.4: Electrophoretic migration of the protein fractions eluted by gel prrmration
chromatography 125
Figure 7.5: SDS-PAGE of the soluble fractions of MFGM isolate suspensions in 4 M
urea buffer 125
Fi y r e 7.6: Concentration of soluble protein after centrifugation of suspensions of
different MFGM isolates as a function of temperature of heating of the cream 129
Figure 7.7: Typical droplet size distribution of 10% oil-in-water emulsions prepared with
MFGM from differently heated creams 13 i
Figure 7.8: Average diameter of oil-in-water emulsions containing 10% oil and 2%
MFGM isolate, as a function of temperature of heating of the cream 133
Figure 7.9: Amounts of a-lactalbumin and P-lactoglobulin as a function of temperature
of heating of the cream 133

Figure 7.10: Arnount of iron in the PAFGM isolates as a function of heating temperature
of the crearn 136
Figure 7.11: Amount of B-lac toglobulin present in the MFGM isolates and average
diameter of ernulsions plotted versus the amount of iron present in the MFGM and the
solubility of the MFGM isolates 138
Figure 8.1: SDS-PAGE of soluble phases after resuspension of the isolates in 1 M urea
and 5 m M 2-mercaptoethanol 143
Figure 8.2: Particle size distribution of emulsions prepared with MFGM isolatrs (rom
butterrnilk obtained from churning 135
Figure 8.3: Elecuon micrographs of oil-in-water emulsions prepared with MFGM
isolates from buttermilk obtained from continuous churning 146
Figure 8.4: Thin section of precipitate obtained by centrifugation of an emulsion
prepared with MFGM isolate 148
Figure 8.5: Transmission electron micrograph of a 3 9 (wh) MFGM isolate from
unheated crearn 150
Figure 8.6: Size distribution of ernulsions prepared with MFGM isolate from
buttermiks collected frorn the AMF process 15 1
Figure 8.7: Arnount of adsorbed whey protein and MFGM protein in emulsions
prepared from buttermilks collected from the AMF process 152
Figure 8.8: Transmission electron micrographs of emulsions prepared with 3 9 MFGM
isolate from buttermilk obtained from the AMF process 154
Figure 9.1: SDS-PAGE of MFGM isolates treated with trypsin and chymotrypsin 162
Figure 9.2: SDS-PAGE of MFGM isolates denved from industrial buttermik, treated
with trypsin 164
Figure 9.3: SDS-PAGE of emulsions prepared with 10% oil and I C/c MFGM isolate
obtained from unheated crem, treated with trypsin 166

Figure 9.4: SDS-PAGE of ernulsions prepared with MFGM isolate treated with trypsin
or chymotrypsin 167
Figure 9.5: SDS-PAGE of the oil droplets of an emulsion prepared with 3 6 MFGM
isolate from industrial buttermilk, marnent with trypsin 169
Figure 9.6: Typical droplet size distribution of an ernulsion prepared with 1% MFGM
isolate and same emulsion surface-hydrolyzed with trypsin and chymotrypsin 170
Figure 9.7: Typical droplet size distribution of 0.5% MFGM isolate ernulsion hydrolyzed
with trypsin and chymotrypsin 172
Figure 9.8: Typical droplet size distribution of 1 95 MFGM isolate emulsion hydrolyzed
with trypsin and chymotrypsin 173
Figure 9.9: Typical droplet size distribution of emulsions prepared with MFGM isolate
from industrial buttermilk, after hydrolysis in solution with trypsin 175
Figure 9.10: Typical droplet size distribution of emulsions prepared with 10% oil and
MFGM isolates from industrial buttermik hydrolyzed in solution or after
homogenization 177

List of Abbreviations
AMF
DLVO
EDTA
F m
FID
HTST
MFGM
MW
PAS
SDS-PAGE
TEM
n c
Anhydrous Milk Fat
Derjaguin-Landau-Verwey-Overbeek (theory)
Ethylene Diamine Teua-Acetic Acid
Flavine Adenine Dinucleotide
Flarne Ionkation Detector
High Temperature Short Time
Milk Fat Globule Membrane
Molecular Weight
Periodic Acid-Schiff (reagent)
Sodium Dodecyl Sulphate - Polyacrilmide Gel Electrophoresis
Transmission Elecuon Microscopy
Thin Layer Chromarognphy

CHAPTER 1
GENERAL INTRODUCTION AND STATEMENT OF THE OB JECTLVES
During the process of buttermaking, cream undergoes a mechanical treatrnent -
chuming, which involves agitation, in the presence of air, until the fat globules
aggregate and phase inversion occurs. During this process, the membrane which
surrounds the fat globules disrupts. Most of the water phase is released in what is calied
buttermilk (Campbell et aL , 1996). Buttermilk is rich in materiai denving from this
membrane and contains a number of components which make it unique among dm
produc ts.
According to Statistics Canada (1996), the butter production has averaged h
Canada 90 000 000 kg, and the production of buttermi& powder increased of 32%
relatively to the previous year (August-Jdy 1995 to 1996). The market value of
buttersnilk powder is about $4-5/kg, which is faûly low if compared to skim mik or
whole m3.k powder, which have a similar protein content Buttermilk powder contains
more lipids (5-6%) than non-fat dry milk (les than 1 %). The high phospholipid content
seems to limit the storage stability of buttexrnilk and its use in other food applications
(Malin et ai., 1 994).
The fuli potential of butterrnilk has not been exploited, perhaps because its
composition is commonly thought to be comparable to that of skim milk. Buttermilk
might have desirable properties because of the high content of phospholipid and protein
in the milk fat globule membrane (MFGM), which is released during churning.
Buttermilk has been widely used as an ingredient in ice cream, bakery and low
fat products as a source of milk solids. It has been reported that if buttemilk is used in
the manufacture of cheese, the resulting products may have a hypocholesterolemic effect
(Abou-Zeid, 1992). In the traditionai technology of low fat cheese making, when the fat
content of the milk is lowered, the amount of material derived from the MFGM which

surrounds the fat globules is also lowered. Among the numerous advantages of the use of
milk solids from buttemilk are not only a decrease in the amount of waste disposai and
improved economics, but also improved flavour and texture of the final product (Mayes
et aL , 1994; Schoenfiss & McGregor, 1997). Recently, attention has been given to
ultrafiitration of buttexmik (Ramachandra Rao et al., 1995) and its incorporation in
cheese formulations to improve its body and texture (Mistry et aï., 1996).
Certainiy, there has been Little interest in the fractionation of commercial
buttermillr cornpared to its widespread uses as an unfractionated mixture. Isolation of
individual proteins has recently gained more attention, with the advancement of
knowledge of how to adapt proteins and isolates to dietetic and technological
requirements of food products. The research described in this thesis was designed to
define buttermilk and describe its functional properties, and also to investigate the
possibiiity of fractionation of this byproduct to obtain isolates with unique properties.
The work described in this thesis had the following objectives:
L
IL
m.
IV.
To study the components present in commercial buttermilk. After a careful
analysis of the available literature on buttemilk, it was concluded that a better
understanding of its chemistry may suggest alternative uses for this byproduct.
To develop methodologies to prepare MFGM isolates from commercial
buttermilk, and to study the ernulsifying properties of these MFGM isolates.
To compare the adsorption behaviour and other properties of MFGM isolates
from fresh raw cream to those reported in the literature and to those of isolates
from industriai buttermilk.
To determine the effect of heat during processing of cream on the functionai
properties of MFGM, and to apply this knowledge to enhance understanding of

how different indusuial buttermaking processes affect the functionality of
MFGM.
V. To determine the susceptibility of the MFGM isolates to proteolysis and to
determine the emulsifying behaviour of the hydrolyzed matenal.
Chapter L provides insight on the rationale behind this research and why industrial
butte& was chosen for this study. Chapter 2 presents some background information
on the components present in buttermilk, with special focus on the composition of the
MFGM. Emulsion formation, stability and protein a t oil-water interfaces are also
subjects of the review enclosed in this chapter. More specific literature and
methodologies are cited in the 'Resulis' section. The main methodologies used in this
research are briefly summarized in chapter 3. The remaining chapters o u t h e the results
of the experirnental work. Chapter 4 describes the behaviour of emulsions made with
buttermilk and soybean oil. The emulsifying properties of butte& are compared to
those of skim milk. The contribution of the MFGM-material to emulsification is
assessed. Chap ter 5 describes the production of membrane isolates from indus trial
buttermilk and their behaviour when used to prepare oil-in-water emulsions. Chap ter 6
illustrates the results obtained after the isolation of MFGM from untreated butterrnik
(churning was canied out on fresh raw cream) and its use in emulsions with soyabean oil.
In chapter 7 and 8 the effect of heat treatment of the cream on the emulsifying behaviour
of the MFGM isolates is discussed. Particular attention is given in chapter 8 to the effect
of the different industrial processes on the functional properties of the MFGM isolates
derived. In this chapter the emulsifying properties of MFGM isolates from two industrial
processes will be descnbed: the production of butter by churning of cream and the
production of anhydrous m i k fat (W. Chapter 9 describes work carried out on the
proteolysis of MFGM isolates. Chapter 10 contains a general discussion and some
recommendations for future studies.

CHAPTER 2
INTRODUCTION
This chapter is a surnrnary of the information reported in the literature on MFGM
and its composition. Skim milk proteins are dso mentioned in this review, because of
their presence in high amounts in indusaial buttermillc and because. in this thesis, some
comparative studies have been carried out between MFGM proteins and skim milk
proteins and their behaviour at oiYwater interfaces. In the last section of this chapter a
brief review of emulsions, their stability and the behaviour of protein at oUwater
interfaces is given.
2.1 COMPOSITION OF BUTTERMILK
The cream used in buttermaking typicdy contains around 40% fat, and skim
milk constitutes the serum phase. During cream destabilization, this water phase
becomes e ~ c h e d with MFGM material. Buttennillc, the water phase released after
chuming, is often considered very sirnilar in composition to skim milk. The two products
have a similar solids content however, their composition differs substantially.
Butteriniik contains a higher amount of iipid (industrial buttermilk has an average fat
content -0.6%) and it contains materid denved from the MFGM, the membrane which
surrounds the milk fat globules in whole milk and cream. The average composition of
buttermilk is shown in table 2.1.

Table 2.1: Composition of butterrnilk (Ramachandra Rao et al., 1995).
Concentration Range [%]
Total Solids 1 8.5-9.0
Lactose
Fat
Ash
2.1.1 The Milk Fat Globule Membrane (MFGM)
4.0-4.2
0.6-0.9
6.0-6.5
Total Calcium
2.1.1.1 Origin and Structure
2.0-2.2 mM
The milk fat globules are synthesized in the endoplasmic reticulum and released
from the ceiI by envelopment in the plasma membrane. Therefore, the MFGM mainly
originates from the apical plasma membrane of mammary epithelid ceils. Evidence of
this has been provideci by imrnunomicroscopy (Franke et al., 198 1) and by biochernicd
observations (Keenan et al., 1983; Aoki et al., 1994). In more detail, the MFGM is a
tri-partite structure, originating not only from the apical plasma membrane, but also from
the endoplasmic reticulum and other cellular compartments (Keenan & Dylewski, 1995).
The portion derived from the apical plasma membrane has a typical bilayer appearance,
with some electron-dense material on the inner membrane side. Mather & Keenan
(1975) suggested that the tightly associated intemal coat is mainly derived from the

endoplasmic reticulum. 'Ibis intemal coat àifferentiates the MFGM from most other
biological membranes, and appem to be a rnonoiayer of proteins and polar lipids which
covers the lipid core of the globules in the cytoplasm. This dense coat matenal present
with the MFGM seems to be resistant to extensive washings with various salt buffers and
non-ionic detergents (Freudenstein et al., 1979). The constituents of this coat seem to
be involved in intracellular fusion of the globules, recognition and interaction of the fat
droplets with the plasma membrane. Transmission electron microscopy (TEM) studies
and themodynamic considerations suggest that the proteins of the mammary plasma
membrane undergo a structural rearrangement during and after the formation of the
MFGM. The cytoplasmic face of the plasma membrane undergoes an abrupt change of
environmen t during fat globule envelopmen t, from conditions predorninantly h ydro philic
to hydrophobie (Mather & Keenan, 1975). A structural reorganization seems to occur
after globule secretion (McPherson & Kitchen, 1983). Figure 2.1 shows a schematic
mode1 of the MFGM at secretion and after reorganization, as proposed by Kanno (1990).
After secretion, the MFGM structure is composed of an inner coat membrane (CM) and
an outer plasma membrane (PM). During reorganization, the outer plasma membrane
fuses with the inner coat membrane. The sandwiched components from the coat material
reorganize in such a way that the polar regions, such as the glycoprotein chains, are
exposed on the outer aqueous surface and the apolar moieties interact strongly with the
lipid regions of the membrane.
2.1.1.2 MFGM Composition
The MFGM is characterized by a complex mixture of proteins, phospholipids and
glycoproteins, and acts as a naturai emulsifier by covering the surface of the milk fat
globule (McPherson & Kitchen, 1983). The composition of MFGM, as reported by Goff
& Hill (1993), is given in table 2.2.

-tL lycoli pid protein from coat material
0 protein a phospholipid
CM Coat Membrane PM Plasma Membrane
Figure 2.1: Mode1 of the milk fat globule membrane after secretion and &ter
reorganization in the acinar lumen (Kanno, 1990).

Table 2.2: Estirnatecl average composition of natural milk fat globule membranes (Goff
& Hill, 1993).
Component
Protein
Copper 0.0 1 -
mg/100 g of fat globules
900
33
- - - - - - --
Phos pholipid
Cere brosides
Numerous analytical methods have been described for the preparation of the
MFGM (Kanno & Kim, 1990). In general, the MFGM material is extracted from fresh
cream separated from raw milk. The fat globules are then washed with water or buffered
solutions, and the washing procedure is repeated to reduce the presence of serum
constituents to a level acceptable for the patticular study. Basch et al. (1985) reviewed
the different washing procedures used in the isolation of MFGM and compared the
composition of the preparations obtained. The different methods for recovering MFGM
material show significant effects on the lipid and protein composition (Mather et al .,
1977; Basch et al., 1985; Keenan & Dylewski, 1995). After washing of the fat globules,
Percent of membrane
48
600
80 4

the membranes are usuaily released by physicai methods such as churning, freezing and
thawing, or treaûnent with detergents (McPherson & Kitchen, 1983).
2.1.1.2.1 Protein Composition
Although MFGM proteins represent only 1% of the total milk protein. they are
fundamental in the stability and integrity of the fat globule membrane in whole and
unhomogenized milk (McPherson & Kitchen, 1983). While very Linle is yet known about
the functionai properties of MFGM proteins, much progress has been made in their
isolation and purification. Most workers have observed strong association between
butyrophilin, xanthine oxidase and some minor MFGM proteins (Keenan & Dylewski,
1995). The proteins of the MFGM are known to be associated with Lipids mainly via
hydrophobic interactions (Shimizu etal., 1976). The specificity oflocalization of
butyrophilin and xanthine oxidase suggest that they may be involved in cellular
recognition and envelopment of fat globules.
The MFGM proteins are usually classified according to their sodium dodecyl
sulphate - polyacrylamide gel electrophoresis (SDS-PAGE) migration (McPherson &
Kitchen, 1983: Basch et al., 1985). Five major polypeptide bands are identifïed by
SDS-PAGE electrophoresis with Coomassie blue staining. Penodic acid-Schiff (PAS)
reagent can also be used as staining procedure, because of its specificity for
glycoproteins, which are abundant in the MFGM.
In the past, MFGM proteins have been isolated rnainly by preparative gel
electrophoresis (Basch et al , 1985). Recently, the understanding of the structure of
MFGM proteins has irnproved via the use of molecular cloning techniques.
Xanthine oxidase accounts for 8-10% of the protein of the MFGM (Mangino &
Bmnner, 1977). It is a complex metallo-flavoprotein whose biological function in the
MFGM is not yet fully understood. This enzyme has multifunctional activity: it oxidizes

aldehydes, purines. pyrimidines, and can produce superoxide radicals. The membrane
bound form of xanthine oxidase contains small amounts of fatty acids which may also
play a role in fat globule recognition and in the secretion process (Keenan et al., 1983).
It has been purified and studied by many authors (Mangino & Bninner, 1977:
Rajagopalan, 1985; Cheng et al., 1988; Berglund et aL , 1996). In native conditions
xanthine oxidase is present mainly in a dimer form. Each subunit has a molecular
weight of 150 kDa and contains four oxidatïon-reduction active sites, one Mo centre,
one flavin adenine dinucleotide (FAD) moiety and two iron-sulphur centers of the
ferredoxui type (Fez-S z), which ac t as electron reservoirs (Cheng et al. , 1988). The
iron-sulphur centres ac t as electron sink to keep Mo oxidized and flavîn reduced during
turnover. The amino acid sequence of xanthine oxidase has recently been determined
(Berglund et al., 1996); the monomer contains 1332 arnino acids and 38 cysteine
residues (Cheng et al., 1988). in the Fez-S 2 - ferredoxins four cysteines bind the iron
and sulphur cluster (Berglund et al. , 1996). One of the ferredoxin iron-sulphur centres
seems to be located in the N-terminal domain of xanthine oxidase (between cysteines 43,
48 and 5 l), where there are 1 1 cysteine residues which are fully conserved between
species (Berglund et al., 1996). Keenan et al. (1983) have identified four isoelectric
variants for bovine xanthine oxidase between pH 7.0 and 7.5. The subunits of the
enzyme seem to be independently active and limited proteolysis of the protein, shows no
effect on its ability to convert xanthine to uric acid (Cheng et al., 1988). The subunits of
the enzyme after proteolysis are held in the associated fom by hydrophobic and
disulphide bonds, and therefore, enzymatic func tion and antibody recognition are still
possible (Mangino & Brumer, 1977; Cheng et al., 1988).
Butyrophiiin is the major integral protein associated with the MFGM (Jack &
Mather, 1990). It has an approximate molecular weight of 66 kDa and contains
glycosylated and phosphorylated amino acid residues, and tightly bound fatty acids
(Freudenstein et al., 1979; Keenan et al ., 1982). The high specificity of its location on

the membrane suggests its involvernent in the envelopment of fat globules (Franke et al.
198 1). Butyrophilin is specific to mamrnary cells, is developmentally regulated and
expressed at maximum levek during lactation (Franke et al.. 198 1). It is a pro tein
highly conserveci across species (Neira & Mather, 1990) with a prirnary structure of 526
amino acids (Jack & Mather, 1990). At least four different variants have been identifieci
by isoelectric focusing in the pH range 4.7-5.3 (Mather et al., 1980; Keenan & Dylewski,
1995).
Butyrophilin tends to associate with other proteins in the MFGM and this supra-
molecular complex is essential to the interactions between the apical plasma membrane
and the lipid droplets fomed in the cytoplasm. Proteins that may associate with
butyrophilin are xanthine oxidase and several other glycoproteins with molecular weight
higher than 40 kDa (Jack & Mather, 1990). This supra-rnolecular cornplex is formed by
hydropho bic and hydrophilic interactions, and possibly disulphide bonds (Mather &
Jack, 1993). Human butyrophilin has recently been sequenced (Taylor et al., 1996) and
its sequence is 84% identical to the bovine forrn. Butyrophilin has a typical
transmembrane protein structure (Aoki et al. , 1994; Taylor et al. , 1996) with a
hydrophobic spanning moiety in the middle of the arnino acid sequence. In the apical
membrane, butyrophilin is oriented with the N-terminus in the exoplasmic space and the
C-terminus facing the cytoplasm. Its variable orientation in the MFGM suggests that
butyrophilin may have two distinct origins: part may derive from the apical surface and
part from iniracellular material. In fact, in spite of its trammembrane-nature, a fraction
of the protein appears to be associated entirely with the cytoplasmic side of the MFGM
(Mather & Jack, 1993). It has a significant hornology with another developmentally
regulated protein present in embryonic tissues. It has been suggested that butyrophilin
rnight function as a component of the immune system (Mather & Jack, 1993).
A senun glycoprotein, component PP3 (proteose peptone 3) has recently been
isolated (Sorensen & Petersen, 1993). Component PP3 is a phospho-glycoprotein with an

apparent molecular weight of 28 kDa, containhg 135 amino acid residues. Antibodies
against MFGM proteins recognize PP3, suggesting an association with the membrane or a
common ongin with the MFGM glycoproteins (Sorensen & Petersen, 1993; Girardet et
al., 1995). This glycoprotein seems to be characterized by strong surface activity
(Courthaudon et al., 1995). Component PP3 inhibits the activity of lipoprotein lipase by
competing with Lipase for the interface; this protein seems to decrease the surface tension
to a higher extent than lipases (Girardet et al. , 1993; Courthaudon et al. , 1995). These
results might be in agreement with previous literature: Shimizu et al. (1978) recognized
the association with the MFGM of a smaU quantity of low molecular weight components,
which can be easily extracted from the MFGM and therefore considered rnainly surface
oriented. These small components rnay originate from the milk serum, and be reversibly
adsorbed to the MFGM.
PAS-6 and PAS-7 are the most abundant glycoproteins in bovine MFGM after
butyrophilin, about 10- 15% of the total membrane proteins (Basch et al., 1985). These
proteins are not tightly associated with the membrane (Mather & Keenan, 1975; Kim et
al., 1992). Kim et al. ( 1992) selectively extracted these two glycoproteins (50 and 47
kDa, respectively) by gel fütration and affinity chromatography. PAS-6 and PAS-7
seem to be part of a Lipoprotein complex with properties resembling those of actin; they
interact with myosin and aggregate at high ionic strength (Keenan et al., 1977). Mather
et al. (1980) identified several isoelectric variants at a pH range between pH 4.2 and 7.1.
The isoelectric heterogeneity of these proteins seems to be caused mainiy by differences
in their sialic acid residues (Kim et al., 1992).
PAS-4 is a 78 kDa integral membrane glycoprotein and makes up 2-596 of the
total protein in the MFGM. It has been isolated from both human and bovine miik
(Mather et ai. , 1980; Greenvalt et al., 1990). The hydrophobie nature of PAS4 was
confimed by its amino acid composition: at least 50% of the residues are non polar and a
further 20% have side chains with uncharged polar groups at neutral pH. The protein is

characterized by a low level of cysteine and methionine and a high level of branched
chain amino acids such as valine, leucine and isoleucine. Approximately 5.34 of the
glycoprotein on a weight basis is carbohydrate, especially mannose, galactose and sialic
acid. Greenwalt & Mather (1985) resolved 4 variants by isoelectric focusing, with
apparent isoelectric points between 7.8 and 8.5. The same authors suggested that PAS
IV may play a role as component of the immune system. PAS IV is indeed present in
the capiiiay endothelial cells of many tissues and in exposed extemal epithelial surfaces
(rnarnmq ceils and bronchiolar epithelium).
2.1.12.2 Lipid Composition
The fat globule membrane in its natural state contains a small portion of
triglycendes. Although the presence of a considerable amount of triglycendes,
especially high-melting triglycerides, in MFGM preparations has been reporteci in the
past, these lipids seem to be denved from fat crystals, contaminating the membrane
during chuming (Walstra, 1974: Wooding & Kemp, 1975).
The phospholipids present in the MFGM account for approximately 50% of the
total milk phospholipids. The phosphoiipid composition in the MFGM has been
determined: sphingomyelin (22% of the total phospholipid), phosphatidyl choline (36%),
phosphatidyl ethanolamine (27%), phosphatidyl inositol (1 1 %) and phosphatidyl senne
(4%) (Kanno, 1990; Malrnsten et al ., 1994; Keenan & Dylewski, 1995). In the MFGM
there is a high ratio of sphingomyeiin to phosphatidyl choline. This distinctive
distribution pattern of phospholipids shared by the plasma membrane and the MFGM is
the strongest biochemicai evidence for the plasma origin of the MFGM (Keenan et aL,
1983). The properties of sphingomyelin from the MFGM have been recently investigated
(Malrnsten et aL , 1994). A source of phospholipids rich in sphingorn yelin has several
advantages over a source rich in phosphatidylcholine: sphingom yelins are less

susceptible to hydrolysis and in general they have a lower num ber of cis double bonds
compared to phosphatidyl choline.
Several different carbohydrate containhg sphingolipids have been identifid as
constituents of the MFGM, both neutral (cerebrosides) and acidic (gangliosides).
GIycosphingolipids are relatively minor constituents and have been the subject of great
interest because of their role in biological activities such as cell interaction,
differentiation and signaiing. Some glycosphingolipids from buttermilk have been
characterized and they appear to be iinked to cellular differentiation and development
(Ren et al. , 1992).
MFGM is a complex mixture of phospholipids and proteins. Its most important
function in whole milk and crearn is to protect and stabilize the fat globules by
surrounding them. The potential uses of this membrane matenal have not been studied
and their functional properties in food systems are largely unknown.
2.1.2 Skim Milk Proteins
A large fraction of the protein present in buttermilk (about 70% of the total
protein) is derived from skim milk. Skim mîlk proteins exhibit a range of important
functional properties; arnong those of most interest are curd and gel formation, foaming,
and emulsifying properties. Skim milk proteins can be divided into two main categories:
whey proteins ( a-lactalbumin and p-lac toglobulin) and caseins (asl -, u~ -, P- and K-
casein).
2.1.2.1 Whey Proteins
The major protein component of the whey fraction in miik is b-lactoglobulin.
This protein, which has high homology of structure with the retinol-binding protein,

seems to function in the transport of srnail hydrophobic substances Wrez & Calvo,
1995). Its primary sequence consists of 162 amino acid residues with a molecular
weight of 18400 Da (Hamblùig et al ., 1992). A detailed description of its structure is
given by Papiz et al. (1986). It has a constrained globular structure consisting of nine
strands of anti-parallel B-sheets, eight of which wrap around to create a conical barre1
closed at one end. A three turn a-helix is situated on the outer surface of the calyx.
Under physiologicd conditions fklactoglobulin exists predominantly as a dimer,
maintained largely by electrostatic forces. At a lower pH the protein associates
revenibly to form octamenc structures. Each monomer contains five cysteine residues,
four of which are constrained in interna1 disulphide bridges. The free thiol is located, in
the native molecule, near the surface in a hydrophobic channel, beside the a-helix. This
highly reactive cysteine 12 1 is involved in intramolecular and intermolecular disulphide
interchange and plays an important role in determining the hnctionality of
lactoglobulin.
The other major whey protein - 20% of the total whey proteins - a-lactalbumin, is
a meMo-protein binding one mole of calcium per mole of protein (Brew & Grobler,
1992). Crys tallographic studies of a -1actalbumin revealed a close homology of s tnicture
between this protein and lysozyme (Acharya et al., 1989). It is characterized by a
compact globular structure containing four disulphide bonds. These bonds are mainly
responsible for the revesible conformational changes occurring upon heat denaturation
( R e h et al ., 1992). Also, a-lactalbumin undergoes structural changes related to the
loss of Ca2+. The release of calcium has profound effects on the thermal s tability of a-
lactalbumin, as weli as on a number of other physico-chemical properties ( R e h et d ,
1992; Matsumura et al ., 1994).
Heat-induced denaturation and interactions of whey proteins have been studied in
various mi& systems Pamenberg & Kessler, 1988; Parris et al. , 199 1). Whey proieins
undergo conformational changes at temperatures between 6û and 80°C (de Wit &

Klarenbeek, 1981;Relkin etcil., 1992; Boyeetal., 1996). The conformationalchanges
are usually followed by protein-protein interactions resulting in the formation of gels by
crosslinking (Gezimati et al., 1997; Parris et al. , 1997) or complexes with other milk
components. In particular, whey proteins may interact with casein micelles (Corredig &
Dalgleish, 1996a; Law et al., 1994) and they may also bind to milk fat globules to an
extent that depends on the state of the MFGM (Daigleish & Banks, 199 1 ; S h m a &
Dalgleish, 1994; Comedig & Dalgleish, 1996b).
Caseins are the major components in bovine rnilk, making up more than 80% of
the total protein (Goff & Hill, 1993). Casein micelles are aggregates of four major
caseins -, as- P-and K-casein and they are polydisperse in size with a wide
distribution in diameter from 30 to 400 nm (Holt, 1992). The skeleton of the casein
micelles is mainiy constituted by aS1 - and ag- casein. They are the least susceptible
among the caseins to micellar dissociation upon cooling (Ono et al., 1990). A cluster of
phosphoseryl residues and three hydrophobic moieties characterize the structure of a,i -
casein; as2- is highly phosphorylated and is the most hydrophilic of the caseins
(Swaisgood, 1992). K-casein and &casein have high voluminosities as monomers and
they have a tendency to self associate (Swaisgood, 1992; Holt & Home, 1996). For fi-
casein this association is caused by a C-terminal hydrophobic region and a polar N-
terminus. By the same token, K-casein is characterized by two distinct domains - a
hydrophobic N-terminal region and the C-terminal macropeptide; its association may
also be characterized by disulphide bonds (Swaisgood, 1992). It adopts a largely
unfolded open conformation in solution and when adsorbed at interfaces. Because of
their molecular structure, both K- and B-casein play important roles as stabilizers in
colioid systems (Leaver & Dalgleish, 1992; Holt & Home, 1996). At

low temperature, as hydrophobic interactions become weaker, &casein is released from
the micelles (Law, 1996). Because of the revenibility of this dissociation, p-casein is
considered to be loosely associated in the micelles by hydrophobic bonds and to be the
least critical between the caseins for the structural framework of the casein micelles. On
the other hand, K-casein acts as a stabilizing colloid, and it is the main contributor to the
stenc stability of the miceiles. There is a strong correlation between the increase in
rnicellar K-casein and the decrease in size of the micelles (Dalgleish et al., 1989). The
surface of the micelles is mainly covered by K-casein, which dominates the behaviour of
the casein micelle in milk. The surface presents a hairy appearance, since the
hydrophilic tails of K-casein protmde into solution (Holt & Home, 1996). The
glycomacropeptide is therefore susceptible to hydrolysis with chymosin and its release
causes instability and aggregation of the casein miceiles (rennehg).
Colloidal phosphate is of considerable importance in maintaining micellar
integnty (Holt et al. , 1986). Calcium phosphate seems to form a centre to which casein
molecules attach via phosphoseryl residues (Holt & Home, 1996). The strength of
bindhg of the individual caseins within the miceiles is largely related to the number of
phosphoseryl residues present in the molecules: P- and K-casein dissociate to a greater
extent than a,l- and ag -caseins (Holt et al., 1986).
Casein micelies are a remarkably stable colloida1 system, however, their
dissociation can be induced by cooling, heating and acidification, alone or in
combination (Dalgleish & Law, 1988; Law, 1996; Anema & Kiostermeyer, 1997). A
more extensive dissociation cm be obtained by the addition of urea or calcium chelating
agents (Holt et al., 1986; Holt, 1992).

2.2. PROTEIN STABILIZED OIL-IN-WATER EMULSIONS
2 . 2 Emulsion Formation
An emulsion is a dispersion of one liquid in another irnmiscible liquid. Emulsion
formation requires an energy input and surface active agents. To break large droplets
into smaller ones, mechanical energy has to be applied to the system, to counterbalance
the Laplace pressure generated by the curved interface of the fat droplet The Laplace
pressure is proportional to y lr [y, surface tension; r, droplet radius]. The fmal size of
the fat droplets in the emulsion depends on the power density of the homogenization
system (it depends on the size of the turbulent eddies: the smaller the eddies, the larger
the pressure gradient). The greatest proportion of energy is dissipated into heat during
emulsification (Walstra, 1983).
The presence of a surfactant facilitates the formation of small droplets, because it
lowers the interfacial tension between the two phases. The solubility of the surfactant,
its concentration and the viscosity of the continuos phase affect the process of
emulsification and the nature of the emulsion formed. During emulsification, droplets are
defomed and broken, and the surfactant is adsorbed ont0 the newly formed interface. In
the manufacture of protein emulsions by homogenization, m a s transport of protein to the
newly fomed interface takes place mainly by turbulence-controlled convection and not
by diffusion (Walstra & Oortwijn, 1982; Walstra, 1993a). Under these conditions. the
rate of mass transport to the interface increases with the size of the adsorbing particles.
Walstra & Oortwijn (1982) showed that during hornogenization of dairy emulsions, large
casein micelles (400 nm) adsorb in preference to smaller aggregates of whey proteins (4-
5 nm).

At the moment of homogenization two phenornena occur simultaneously: particle
recoalescence and particle stabilization by surfactant (Walstra, 1993a). The surfactant
adsorbed at the interface wil l eventually settle to equilibrium after homogenization.
Therefore, the surfactant, on one hand, lowers the interfacial tension and facilitates the
formation of smaii droplets, and on the other hand, prevents recoalescence. Surfactants
may be roughly divided into small molecules (amphiphilic molecules such as lecithins,
detergents, monoacylglycerols) and large molecules such as proteins. Srnall molecule
surfactants give lower interfacial tension [y] than proteins. In generai, a protein-
stabilized emulsion resdts in larger ernulsion droplets than those of an emulsion
stabilized by small molecule surfactants. In a real food system, often both surfactants
and proteins are present and contribute to the stability and behaviour of emulsions.
2.2.2. Emulsion StabTty
Emulsions are thermodynamically unstable since the creation of an oil-water
interface results in a free çnergy increase. When surface active molecules are adsorbed
at the oiVwater interface, the stability of the emulsion droplets is dependent on both
electrostatic and steric interactions. The DLVO (Deryagin-Landau-Vervey-Overbeck)
theory is the classical theory of coiloidal stabilization often used as an approach to
understand emulsion stability. This theory tries to explain the stability of particles
against aggregation on the basis of van der Waals attraction and electrostatic repulsion.
At any distance, attraction and repulsion each are proportional to the particle diameter,
and are additive (Walsua, 1993b). The electrostatic repulsion caused by the presence of
the electrical double layer constitutes a positive potential while the van der Waals
attractive forces are responsible for the negative potential. There is, in general, an
energy banîer that the particles have to overcome before reaching the primas, minimum
potential, where it is assumed that emulsion droplets irreversibly aggregate. The

DLVO theory does not describe adequately the stabilization of emulsions; however, it is
a useful model in predicting when destabilization may occur (Walstra, 1993a).
Adsorbed proteins stabilize oil-in-water emulsions not only elec&rostatically, but
by a combination of charge repulsion and steric stabilization. The latter is quite important
in food emulsions; proteins stericaily prevent droplets from coming together, since this
would cornpress the tails of the macromolecules. resulting in a loss of configurational
entropy. Steric stabilization is lffected by solvent quality. Casein micelles are a good
example of steric stabilization: in milk the micelles are stabilized by the
glycomacropep tide of K-casein (Holt. 1992). Another model of s tenc s tabiliza tion can
be found in Bcasein: when oil-in-water emulsions are prepared with this protein, its
hydrophilic tail protrudes a few nanometers into solution prevenàng droplets from
coming together (Dalgleish & Leaver, 199 1; Fang & Dalgleish, 1993a). If
environmental conditions change, (for example, if ethanol is added (Agboola, 1996), or
the stabilizing tails are hydrolyzed by en y m e s (Holt, 1992; Leaver & Dalgleish, 1992),
destabilization of these sterically-stabilized colloids occurs.
During emulsion formation, if the surfactant is not sufficient to cover the entire
oil-water interface, the oil droplets will decrease their total surface area by coalescing.
In case of an insufficient presence of large molecules (protein aggregates), bndging
between the fat globules can aiso occur.
A schematic diagram of the various mechanisms which would Iead to
destabilization and creaming of an emulsion is shown in figure 2.2.
In oil-in-water emulsions. four main mechanisms of instability can be descnbed:
flocculation, coalescence, partial coaiescence and creaming. In flocculation and
aggregation, two or more droplets come together with no changes in the distribution of
their particle size. Flocculated particles usually maintain their shape and size with
defmed boundaries; this sometimes can be a reversible process. As already mentioned,
if Uisufficient surfactant is present and the macromolecules are adsorbed at the interface

1 Phase 1 1 Separation 1
Coalescence
t
Coalescence
t Rapid Creaming
/' - Slow Creaming
Emulsion
Figure 2.2: Different types of instability in emulsions (Walstra, 1983).

of two different droplets, bridging flocculation occurs. A third type of flocculation
mechanism is depletion flocculation. In this case, large macromolecules which are not
adsorbed at the interface result in high osmotic effect which causes aggregation of the oil
dropleu. In more complex systems. the role played by macromolecules may change
depending on their concentration in the emulsion. An example of depletion flocculation
has been reporteci recently for emulsions containhg an excess concentration of sodium
caseinate (Dickinson, 1997).
During coalescence, the intedacial layer ruptures as two globules corne together.
This generaily occurs after flocculation or creaming, when the droplets are in close
proximity to each other. It is enhanced by large droplet size and a high oil phase volume,
where the Brownian motion of the oil droplets is high. If partial coalescence occurs, the
oil droplets aggregate irreversibly, because of the formation of a crystal network between
them. Partial coalescence is often used to produce desirable texture
and structure in food products such as whipped toppings and ice cream (Goff et al.,
1989).
The kinetics of destabilization are central to the behaviour of food systems.
Emulsions are themodynarnicaily unstable and they might be stable only under a kinetic
point of view; therefore, it is of great interest to determine the optimal conditions to
extend their stability over their commercial Me-time. AU the destabilization mechanisms
outlined above eventually lead to creaming of the oil droplets and phase separation. This
kinetic event can be descnbed by the Stoke's Law:
where:
v = velocity of creaming

a = acceleration
Ap = difference in density between the two phases
q = "scosity of the continuos phase
d = particle diameter.
This explains why in food emulsions creaming can be reduced by reducing the
particle size, matching the densities between the dispersed and the continuous phase.
and by increasing the viscosity of the continuous phase.
2.2.3. Proteins at OiVWater Interfaces
Proteins as charged polymers are effective in stabilizing emulsions. Two different
mechanisms are involved: charge stabilization - by repulsion of droplets with the same
interfacial charge, and steric stabilization - by extension of part of the molecule several
nanometers into solution.
Adsorption of protein is often accompanied by a loss of configurationai and
hydration energy of the molecule. Hydrophobie groups are likely to interact with the
hydrophobic surface of the forming oil droplet. The adsorption behaviour of proteins at
surfaces is the net result of various types of interactions. In the case of a protein with a
strong interna1 coherence, a 'hard' protein, the adsorption would be primarily governed
by electrostatic interaction and dehydration of hydrophobic areas. On the other hand, in
'soft' proteins there is an additional driving force caused by stnictural rearrangements of
the molecule (Haynes & Norde, 1994). Many globular proteins change conformation
upon adsorption, losing rnost of their native tertiary structure whiie st i l l retaining their
secondary structure. The extent and the reversibility of the conformational change
depend on the elas ticity of the molecule (Steadman et al. , 1992; Corredig & Daigleis h,
1995). Four effects usualiy make a primary contribution to the adsorption behaviour:

rearrangements of the protein structure, dehydration of the sorbent surface, redis tribu tion
of charges at the interface, and protein surface polarity (Haynes & Norde, 1994).
Proteins such a s caseins are very flexible, spreading ont0 the interface to cover a
large part of the surface area when they are present at low concentration. If more protein
is present, then the molecules rnay pack and extend further from the interface (Fang &
Dalgleish, 1993a). Among milk proteins, the arnphiphilic charmer of pcasein makes it
ideal to be used as mode1 for protein adsorption (Leaver & Dalgleish, 1992; Nyiander &
Wahlgren, 1994: Dickinson, 1984). Its hydrophilic N-terminus is fundamental to its
function as a protective surfactant.
In any particular dairy system, the surface concentration of the rnilk proteins will
depend on the protein composition, the ratio between single proteins, the state of
aggregation of the proteins, the environmental conditions, and the presence of low
molecular weight surfactants. Cornpetitive adsorption between various proteins in dairy
emulsions has been descnbed by several workers (Dickinson. 1986; Hunt & Dalgleish,
1994a; Matsumura et ai. , 1994). For example, it is known that purified k a s e i n
cornpetes very efficiently with other skim miik proteins for the interface;
lactoglobulin and a-lactaiburnin adsorb equally at pH 7, while at lower pH, a-
lactalbumin adsorbs better than P -lac toglobulin (Matsumura et al., 1994).
If emulsions are prepared with mixtures of proteins and surfactants, the
composition of the interfacial layer depends on the competition between these surfactants
during homogenization and their behaviour after homogenization (Goff & Jordan, 1989;
Counhaudon et al., 199 l4b; Dickinson, 1984). Usually the interfacial layer is weakened
by protein displacement at the interface, and this rnay lead to flocculation or coalescence
of the emulsion droplets.
It is evident frorn a large number of studies that in complex food systems there
can be sirnultaneous competition between different proteins and different surfactants.
The environmental conditions and the previous history of the ingredients have to be

taken into consideration; the displacement of a protein by another surface active
molecuie is highly affected by factors such as exposure to the interface, the age of the
adsorbed layer (Counhaudon et al ., LW 1a,b; Dickinson & Matsumura, 199 1 ) and any
thermal or environmental denaniration of the protein (Matsumura et al., 1994; Hunt &
Dalgleis h, 1 995).
In snidying the largely unloiown ernulsZying properties of material derived from
buttermilk, a complex behaviour of the emulsions prepared is expected. Not only will
skim miik proteins contribute to the stability of the oil droplets, but the MFGM fraction
(constituted by a mixture of protein and phospholipid) wiii also affect the properties of
buttermilk emulsions.

CHAPTER 3
METHODOLOGIES
In this chapter theones and applications of the methodologies used in this thesis
are discussed. Details of the methods emplo yed in the various stages of the research will
be given in the chapters descnbing the results of the experiments.
3.1 BUTTERMAKING
During the process of buttermaking, cream, an oil-in-waier emulsion, is
converted to butter. After c o o h g the cream, the fat globules, surrounded by a
membrane composed of phospholipids, proteins and cholesterol, contain fat in a semi-
crystalline state. In the fust step of the buttermaking process, the fat globule membrane
ruptures and the oil-in-water emulsion destabilizes. As a result, fat globules clump and
cluster together. The process is carried out in the presence of air, because of the
inefficient rupture of the MFGM in the absence of air. The fat globules are then M e r
worked and butter, the water-in-oil emulsion, is obtained. The water phase released
from this phase inversion is commonly called 'buttermilk'.
The results of the work described in this thesis were obtained by using, as a
source of MFGM matenal, buttermilk from three different buttermaking processes.
In the laboratory this process was carried out with a bench-size chum as illustrateci in
figure 3.1. Fresh unpasteurized or heat-treated cream, with a fat content of about 408,
was kept at refrigeration temperature (4'C) overnight The cream was then chumed at
10'C until phase inversion was apparent The buttermillc was separated from the butter
granules by f îdte~g through a cheesecloth.

Figure 3.1: The laboratory-size chum used to prepare buttermik

In the conventional process of c o n ~ u o u s buttermaking, cream is pasteurized in a
plate pasteurizer at temperatures of at least 80°C for 16 seconds prior to churning. At the
dairy, phase inversion is induced by churning cream with 40% fat. at low temperature
( 10- 14'C). Rupture of the MFGM is achieved by foaming the cream in the presence of
air. A kneading system works the butter granules and then the water phase, buttermilk,
is released. We obtauied samples of this buttermilk from Gay Lea Foods, Guelph.
Ontario.
In the anhydrous rnilk fat (AMF) process, phase inversion is produced in the
absence of air. Figure 3.2 illustrates the two main steps of this process. The
temperature of the cream is raised to 55-60°C and then the cream, containing about 40%
fat, is fed to a concentrator, similar to a centrifuga1 separator (phase 1). The water phase
leaving this stage usually contains 1-2% fa t This serum is fed to a buttermilk separator
to recover the residud fat, which is reninied to the concentrator (Iilingworth & Bissel,
1994). This serum released during phase 1 is commonly calied 'skirn rndk'.
The cream leaving the concentrator after phase 1 contains between 70-80% fat and is fed
to an homogenizing device, where phase inversion occurs. At this stage, the fat globules
of the 'plastic crearn' are not able to withstand the high shear, and imrnediately rupture
(Illingwonh & Bissel, 1994). The semrn released in this phase is buttenniik. The
process of AMF starts with good quality cream, to avoid the problems of low yield and
stability of the final product caused by the use of fermented creams. In some cases,
sweet whey crearns (byproducts of cheesemaking) are used in the AMF process. We
obtained samples of this buttermillc from Ault Foods Ltd., New Dundee, Ontario. The
results of the emuls-g properties of MFGM obtained from buttermilk from this
process are descnbed in chapter 8.

I Phase 1 Phase II
1 40% Fat 70-80% Fat AMF
Fat globule + Skim Milk
Figure 3.2: The process of phase inversion during the production of anhydrous mi& fat
(Illingworth & Bissel, 1994).

3.2 ELECTROF'HORESIS (SDS-PAGE)
Electrophoresis is commonly used in protein analysis. Macromolecules are
forced by an electric field through a gel characterized by a certain porosity. Their relative
mo bility is related to their molecular charactenstics. Pol yacrylamide gel electrophoresis
(PAGE) is widely employed because the pore size can be fmely controlled by the use of
this synthetic polymer, and the average pore size is related to the acrylamide
concentration. The cationic detergent sodium dodecyl sulphate (SDS), is also commonly
used in PAGE to solubilize proteins and to neutralize their charge. SDS binds to proteins
giving an overail negative charge (the binding ratio is about 1 g to 1.4 g of protein)
(Strange et al., 1992). With SDS-PAGE the separation is achieved on the ba i s of
molecular weight.
In reducing conditions, when 2-mercaptoethanol is added to the protein sample,
disulphide bonds are dissociated, and the protein quatemary structure together with any
S-S complexes are disrupted. In contrat, in "nativew- PAGE (in the absence of SDS
and 2-mercaptoethanol) the proteins separate depending on their charge to size ratio.
SDS-PAGE has been widely used in the analysis of milk proteins and proteolysis
of cheese (Strange et al., 1992). More recently, the technique has been also used to
determine protein adsorption at oiUwater interfaces (Hunt & Dalgleish, 1994b; Corredig
& Dalgleish, 1996b).
The recent introduction of precast ultrathin minigels and an automated system for
performing migration, s taining and des taining (Phasuys tem, Pharmacia B iotech, B aié
d'Urfé, Quebec) has improved the analysis tirne. With 20% PAGE gels it is possible to
efficiently separate milk proteins (Strange et al., 1992). Malin et al. ( 1994) described a
method that, with the use of the Phastsystem, allows the detemination of buttermilk
powder when added to skim milk powder, by identification of three polypeptide bands
specifk to the MFGM.

In a typical analysis procedure for SDS-PAGE with the Phastsystern (Pharmacia
Biotech), protein samples were resuspended in electrophoresis buffer containing EDTA,
SDS and bromophenol blue. When analyses were performed in reducing conditions, 2-
mercaptoethanol was also added. The protein mixture was denatured by heating at
LOO°C. The samples were loaded ont0 a PAGE gel and run in the Phastsystem. After
electrophoretic separation, the gels were stained with Coomassie blue. After staining the
excess stain was removed by a solution containing 30 J methanol and 10% acetic acid,
and before drying the gel was treated with a preserving solution containing 10% glycerol
and 10% acetic acid. Details of the particular conditions used in this research are given
in the chapters describing the experiments.
Band identification is carried out by comparing the polypeptide migration to that
of protein standards. The amount of protein can also be quantified using a densitometric
scanner. In this work, a Sharp scanner was employed, dong with the Imagemaster,
software for image analysis (Pharmacia Biotech).
Microfluidization is a relatively new technique of hornogenization, patented by
Microfluidics Corporation (Newton, MA). The microfluidizer consists of an inlet
resewoir and an air-dnven intensifier pump, the pressure of which can be adjusted. The
mixture is fed into the reservoir and it is directed, with cons tant pressure, into the ceramic
interaction chamber. Inside the ceramic chamber the pressunzed Stream is direc ted
through micro-channels at very high velocities. Microstreams coliide against one
another at right angles, leading to an intense impact, shear and cavitation. This results in
2 very efficient emulsification. A recirculation valve conuols the output, and
recirculating the emulsion may result in a further reduction in particle sizes.

The rnicrofluidizer is more efficient in disrupting fat globules than a conventional
high-pressure valve homogenizer (McCrae, 1994). The microfluidization pmcess results
in a large population of very smalI emulsion droplets (McCrae. 1994: Strawbridge et al.,
1995; Robin et al. , 1993). It has been recently shown that when milk is homogenized
by microfluidization, not only is the size distribution different from that of conventionally
homogenized mi&, but also the microfluidized particles have a very unique structure
(Dalgleish et al. , 1996).
3.4 INTEGRATED LIGHT SCATTERING
Colloida1 particles, immersed in a fluid, scatter light. The scattering pattern - the
intensity of the scattered light as a function of the scattering angle (8) between the
incident and the scattered beam - depends very strongly on the particle size of the
coiioidal suspension, and on the wavelength (h) of the incident light (Hunter, 1993).
According to the Rayleigh theory (valid for particle diameter d d R O and for a
refractive index close to l), the Raleigh ratio Re (i-e. energy scattered per unit t h e into
a solid angle defmed by 8) is proportional to the number of particles present per unit
volume (N), the geometry of the system and the rnolar mass of the particle (M):
Where Is and Io are the intensity of the scattered Light and the incident light respectively,
k is a constant which accounts for the geometry of the system and r is the sample-
detector distance (Hunter, 1993). This theory is applied to the shidy of macromolecules
rather than colioids, because with the point scatterers assurn ption, the average diameter

has to be less than U20. An important condition is that no interference occurs between
the point scatterers. Under these assurnptions, the scattering pattem is independent of the
angie 8.
if the scatterer is a larger particle, the Rayleigh-Gans-Debye approximation is
used. The large particle is an assembly of point scatterers, the scattered photons from
which can interfere with each other. In this approximation. the refractive index does not
differ substantiaiiy from that of the surrounding medium and the largest particle diameter
is less than N2. The scattering pattern is now angle dependent, and the Raleigh ratio
becomes
P(8) and S(8) are the intraparticle and the interparticle scattering factors; both factors
depend on the scattering angle 8. This has very important consequences: when a particle
scatters light. it produces a unique iight intensity which is a function of the angle of
observation. The scattering factor P(8) is a consequence of the interference occming
from different points of the same particle; it contains information about the size, shape
and internal structure of the scatterer. It is possible to calculate P(0) for particles with
simple shapes. The interparticle scattering factor S(8) is a cornplicating factor
accounting for interferences occurring between particles. In very diluted systems
interparticle interactions can be neglected, and S(8) = 1.
When particles have sizes comparable with the h of the incident iight, the
scattering pattem is more complex, and the determination of the scattering factor P(0)
becomes more complex as well. This size range is referred to as the Mie region, after the
Lorenz-Mie theory, which describes the angular dependence of the scattering pattern

from spheres of size comparable to h. The P(8) is described as a complex function of the
size parameter x, the relative index of refraction rn and the angle of observation 8.
The relative index of refraction is the ratio between the refractive index of the particle
and that of the medium. The size parameter x for a sphere of diameter d is given by
Few colloidal systems conforrn to the assumptions necessary to apply the
Rayleigh theory (Le. d d ) . When the sizes of the scatterers are larger than k, the particles
diffract Light and this property c m be used for their size determination. The Fraunhofer
diffraction theory (as the Rayleigh theory) explains that the intensity of the scattered light
is proportional to the particle size. The size of the diffraction pattern (scattering angle
(0)) is inversely proportional to the size of the particle (Weiner, 1984). The intensity of
the Fraunhofer diffraction pattern is given by the Airy equation:
where f is a mathematical function and x is the size parameter

where s is the radial distance from the op tical axis in the detection plane, d is the particle
diameter and F is the focal length of the lens.
The Mastersizer X uses laser diffraction to determine the particle size distribution
of colloidal suspensions (Figure 3.3). The instrument is able to colleet data from 32
different angles in a very short time, by illurnina~g the sample with a laser beam and
collecting the diffracted Light with a special lens which directs it to an array of detectors.
In the Mastersizer X, the light scattered by the particles is incident on a receiver lens.
The detector consists of a series of 3 1 concentric annular sections and collects the
scattered Light From the Airy equation (5) it is possible to measure the intensity
distribution of the light over a senes of circles of radius S. In the Mastenizer two optical
configurations c m be use& the conventional Fourier optic and the reverse Fourier optic.
With the reverse Fourier optic, it is possible to measure a range of sizes down to O. I Pm.
The conventional Fourier configuration rneasures, using the diffraction theory, only
particles of diameter larger than 0.5 Pm.
In the rneasurements described in this thesis the reverse Fourier op tic
configuration was used. The data analysis in this particle size range is performed
employing the Mie theory, and it is perfonned automatically by the software provided
with the instruments.
With the Mastersizer, the disuibution of particle sizes in the emulsion can be
determined. Particles are divided into size classes and the number of particles on each
class is identified.
Various types of averages can be determined for any polydisperse system:

where Ni is the number of particles in the ith class per unit volume and d is the diarneter
in the ith class. In the work presented in this thesis, the average diameters which best
characterized the emulsions were d32 [a=3] , also caiIed volume-surface diameter, and
ci43 [a=4], called weight average diameter.
Smple Receiver Lens ceil
Beam expander
Detec tor plane
Figure 3.3: Schematic representation of the Mastersizer X.
Measuring microfluidized emulsions with the Mastersizer has some important
limitations. Microfluidization creates a large population of very smail particles

(Dalgleish et al., 1996) which can not be determined by the Mastersizer, which has a
lower b i t of 0.1 pm diameter. Strawbridge et aL (1995) showed that when the particle
size distribution of a microfluidized ernuision is measured with the Masterisizer, the
surface area c m be in some cases underestimated by as much as 50%. This has to be
taken into account as a limitation when the amount of protein adsorbed at the interface is
determined, or, more generally, when the specific surface area of the emulsion droplet is
the variable of interest.
3.5 MEASUREMENT OF ZETA POTENTIAL
Many of the properties of food colloids are determined by the charges which
characterize their interface. The 6-potential is often used as a parameter to describe the
electrostatic properties of a colloidal surface.
ln an electrolyte solution, a charged particle attracts ions of the opposite charge.
As a consequence, a Iayer of ions foms around the particle to fom a "diffuse double
layer". The electrical double layer cm be descnbed as divided into two regions (Figure
3.4). In the region closer to the particle the ions interact strongly with the charges on the
surface, while in the outer region the charges are more diffuse (Dickinson & Stansby,
1982). The electricai double Iayer determines the electrokinetic behaviour of colloid
particles.
The surface potential Y of the double layer has been descnbed by Goüy and
Chapman. In the simplifed mode1 proposed by Stem. the difference between the two
regions of the double layer, the close region of adsorbed ions (of thickness 8) and the
diffuse region, is taken into account The potential faUs from Y. at the surface to Y5 at
the Stem plane, and then it decays exponentially to zero in the diffuse region

Where K-1 represents the Debye Iength, the thickness of the double layer as shown in
figure 3.5, and is the surface charge. The thickness of the double layer depends
suongly on the ionic strength of the medium (Myers, 199 1).
The Stern potential, '3'8 can be approxirnated by the electrokinetic potential (6-
potential, which is the apparent charge at the surface of shear), determined by
measurements of electrophoretic mobility. In the research described in this thesis, the 6 -
potential of emulsion droplets was determined by laser Doppler electrophoresis with a
Malvem Zetasizer 5004 (Malvern Instruments Inc., Southboro, MA).
Figure 3.4: Schematic representation of Stern's mode1 of the electrical double layer. The
Debye length (K-l) and the Stem layer (6) are indicated.

In laser Doppler electrophoresis the technique of dynamic light scattering is used
to detemine the motion of particles affected by the electric field. Dynamic Light
scattering is based on the principle that iight is scattered by moving particles. The
heterodyne detection mode is used where the signal results from the interference of the
Doppler shiftzd light from the particles with the frequency of the unshifted Light, the
latter dominating the signal (Haliet, 1995). The fmal frequency is a function of the
particle motion (Hunter, 1993). The peak signal moves from zero in proportion to the
particle velocity .
The c-potential is based on an estimation of the electrophoretic rnobility (p) by
the Hückel equation (Hunter, 198 1)
where E is the dielectric constant and q is the viscosity of the medium. In the
determination of the c-potential of emulsion droplets at Iow ionic strength, the
electrophoretic mobility has to be adjusted by a Henry function Am)
where a is the particle radius and rl is the Debye length. The Henry function f ( ~ a )
depends on the particle shape; it is q u a i to 1 for small (m) and 3/2 for large ( ~ a )
(Hunter, 198 1).
The measurement has to be taken in the instrument's ceii at a point where the net
electrophoretic flow is zero, so that the rnobility of the particles is caused only by the
electric field acting on them. The flow velocity v is a function of 1 (the point at which
the rneasurernent is made), r (diameter of the celi) and L (length of the cell):

When 12=0.5r2, there is no net flow. For this reason, the rneasurements are perfomed in
the ZET 5 104 cell (Malvern Instruments) at 14.6% of the distance across the tube (from
the wall) where the net electro-osmotic flow equals O.
3.6 PHOSPHOLIPID ANALYSE: IATROSCAN
The phospholipid composition can be determined by the use of thin-Iayer
chromatography (TU3). The Iatroscan (Iatron Laboratories, Tokyo, Japan) is a TLC
apparatus set in a microscale arrangement A flame ionization detector (FID) is
employed for lipids quantification. The system has been recently described by Shanta
(1992) and Tvrzicka & Votruba (1994). The iatroscan consists of two independent units
- the Chromarods, which constitute the TU3 component, and the FID scanner unit.
The thin layer is a quarz rod 0.9 mm in diameter and 15 cm long, covered by
sintered silica (Chromarods, Iatron Laboratories). Ten rods are held together by a metal
frame. The quarz rods are spotted with a few pl of sample and the development is cmied
out with an appropnate solvent mixture. After migration, the chromatorods are scmned
with the FID, which consists of an hydrogen flame jet and an ion collector. The detection
system is based on the same pnnciple as for gas chromatography analysis; the sampk is
ionized in the flame and the detector signal reflects any changes in ionization current.
With the scanning system it is possible to determine the migration of the different lipid
components. To identify the compounds present in the sarnple, calibration with
standard mixtures of phospholipids, triglycerides and cholesterol has to be perfomed.

3.7 TRANSMISSION ELECTRON MICROSCOPY
Transmission electron microscopy (TEM) has been widely used to study the
structure of emulsions (Goff et al., 1987; 0ehlmann et al.. 1994; Ferragut & Chirait,
1996). Furthermore, TEM has been empbyed to observe the effects of homogenization
in milk (Robin et al.. 1992; Dalgleish et al ., 1996) and in the study of the structure of the
MFGM (Kenan et al ., 1977; Franke et al. , 198 1 ; Freudenstein et al ., 1979).
Electron microscopy allows the observation of biological specùnens at very high
resolution (2-3 A). Sample preparation is extremely critical: the sample has to conserve
its structure when transfened to a vacuum chamber and bornbarded with high energy
electrons. Problems such as electrostatic charging, melting and evaporation c m be
minimized by careful chemical fixation and dehydration.
In general, the sample undergoes a chemical fixation with glutaraldehyde and
osmium tetroxide (OsO4). Glutaraldehyde is a diaidehyde which, by crosslinking, acts
as a rapid fixative of pm tein s tnictures (Hayat, 1 993). The specimen is then s tained
with Os04 (MW 254), which acts as both fixative and stain. Osmium tetroxide is a
tetrahedrai non-polar molecule with very good penetration capability in biological tissues.
Imidazole and malachite green. added to the specimen with 0 ~ 0 4 , may improve the
sample staining. Angermuller & Fahimi (1982) used imidazole buffer (a weak organic
base) during the osmication step. With this procedure the staining of unsaturated lipids
was improved. The imidazole molecules form a coordination complex with OsO4. The
imidazole-osmium rnethod has been used as an effective approach for staining
unsaturated fatty acids, phospholipids and lipoproteins (Goff et al., 1987; Hayat, 1993).
In addition to imidazole, the use of malachite green has been suggested not only to main
and stain glutaraldehyde-soluble iipids but also to enhance the overali staining (Hayat,
1993). Malachite green is a monocationic dye, often used as a pH indicator, which
penetrates easily into the cells and into the core of lipid droplets. To be effective,

fiation with malachite green has to be accompanied by treatment with glutaraldehyde
and Os04 (Hayat, 1993).
After chemical fixation the sample undergoes a senes of dehydration steps and it
is embedded in plastic resin. The embedding medium must be thermo and vacuum
stable. After embedding, the min blocks are then cut in thin sections (10-100 nm), to
ensure sufficient penetration of the specimen by the electrons.
The thin sections are then suineci once more to irnprove contrast in the specimens.
Post staining increases the electron opacity of samples which have been aiready stainrd
during chemical fixation. With post staining, it is often possible to observe with more
detail structures which are not clearly defmed when only osmication is performed.
Uranyl acetate and lead citrate are the most widely used poststaining reagents (Hayat,
1993).

CHAPTER 4*
BUTTERMILK PROPERTIES IN EMULSIONS WITH SOYABEAN OIL AS
AFFECTED BY FAT GLOBULE MEMBRANE-DERIVED PROTEINS
This chapter describes a study of the emulsifying properties of buttermilk
proteins. At the end of the chapter, asection is included which descnbes results of
parailel experiments carried out on emulsions prepared, under the same conditions, with
skim milk powder. Because of the different composition of buttermilk and skim milk. it
seemed appropriate to compare the behaviour of the two emulsion systerns.
4.1 INTRODUCTION
When cream is churned during buttermaking, an aqueous phase (buttermillc) is
released. During chuming, the fat globules rupture and the miik fat globule membrane
(MFGM) is released into the aqueous phase. Thus, buttermilk contains several
componenü which are not found in other milk-denved fractions. The composition of
this byproduct is rather similar to that of skim mik, with the exception of considerable
amounts of material (phospholipids and proteins) derived from the MFGM (Malin et ai..
1994; Mistry et al., 1996). Although buttermilk is used in a varïety of food products as
a protein-nch ingredient, it tends not to be used for functional attributes, and the effects
of its individual constituents have not been elucidated.
Many studies have been conducted on isolated MFGM (Mather & Keenan, 1975;
McPherson & Kitchen, 1983; Keenan & Dylewski. 1995). In miik and cream, the
MFGM covers the fat globules and keeps them dispersed in the aqueous phase. Although
they constitute a very small portion of the total milk proteins, the MFGM proteins are
fund in fat globules (McPherson & Kitchen, 1983). A number of MFGM proteins have
been identified, mainly by their migration in SDS-PAGE electrophoresis (Mather &
*Subrnitted essentially in îhis form tu the Journal of Food Science. Accepted for publication, October 1997.

Keenan, 1975; Basch et al., 1985). The high hydrophobicity of certain regions of their
structure seems to play a role in stabilizing the milk fat globules (Basch etal., 1985;
Keenan et al,, 1983).
The manner in which the cream is handled, treated or stored can affect the
composition of the isolated MFGM (Keenan & Dylewski, 1995). Most reported studies
have been focused on examining the characteristics of the MFGM. In these studies, to
obtain valid analytical data the milk fat globules had to be pretreated to remove any
adsorbed semm constituents. Therefore, cream was abundantly washed with buffered
solutions pnor to the isolation of MFGM (Mather & Keenan 1975; Keenan et al.. 1983).
However, the water phase which results from the indusaial process of buttemaking
contains, together with the MFGM components, high levels of s W milk protein (Malin
et&, 1994). Therefore, results obtained in a laboratory usuig washed creams are not
necessarily representative of the MFGM-denved material from industrial buttemilk.
The feasibility of reconstituting fat globules with MFGM extracts in emulsions has been
widely demonstrated (Kanno et al ., 199 1 ; Oehlrnann et al ., 1994), and the presence of
MFGM proteins enhanced the emulsification properties of buttermiik (Kanno, 1989).
However, in those studies MFGM-derived protein was isolated from cream which had
not been heat treated, and after abundant washing.
The potential uses of MFGM-derived proteins as food ingredients for their
functional properties require additional study of mem brane-derived materials isolated
from industrial products. The difference in composition between industrial buttermiik
and aqueous phases containing pure MFGM extract, clearly shows the need for a careful
selection of the protein source. Our objective was to investigate the protein composition
of bumrrnillc from an industrial process. The emdsifying propefles of the proteins in
buttermilk were also studied, by reconstituting this product with vegetable oil, and the
stability of such emulsions and the protein adsorption at the olw interface were
determined

4.2 MATERIALS AND METHODS
Buttemïlk samples were collected from a local butter manufacturer using
noncultured cream, and sodium azide (0.02% w/v) was added. The buttermilk was
freeze dried and stored at -20°C. Protein concentration was deterrnined with a modified
Lowry procedure (Markweil et al., 1978), using bovine serum albumin as protein
standard. AU experiments were c&ed out at l e s t in tripkate.
4.2.1 Transmission Electron Microscopy
Butterrnik was acidified with HCI to pH 4.6 and then centrifuged at 3000 g for 1 O
min to compact the precipitate as reported by Kalab (1980). The specimen was h e d in
aqueous glutaraldehyde (2% w/v), post fmed in a 2% (w/v) O s 0 4 in veronal-acetate
buffer (0.05 M, pH 6.8), and dehydrated in a graded ethanol series. The samples were
then embedded in epoxyresin (Luft, 1961). Thin sections were stained with uranyl
acetate and lead citrate. The electron microscope (Hitachi H7 100 EM) was used at 75 kV
emission.
4-2.2 Emulsion Preparation
Freeze dned buttermilk was reconstituted with 20 m M Tns/HCl buffer at pH 7.00
in a range of concentration between 0.5 and 20% (w/w). Oil-in-water emulsions were
prepared using soyabean oil (Sigma Chernical Co., St. Louis, MO) (10 and 20% wlw) and
different concentrations of buttermi1.k solids. The mixtures were homogenized using a
Microfluidizer (mode1 1 los, Microfluidics Corp., Newton, MA) with an input pressure

of 56 MPa (Hunt & Dalgleish, 1994b). Each sarnplc was circulated for 10 strokes of the
pump and then coliected, and the procedure was repeated for 4 homogenization passes.
4.2.3 Determination of Partide Size
The diameter distribution of the fat globules and their specific surface area were
detemined by integrated iight scattering using a Mastersizer X (Malvem Instruments
Inc., Southboro, MA). The samples were diluted c a 1 500 in MilliQ water and rneasured
at arnbient temperature (25'C). The presentation factor used was 0303 corresponding to a
refractive index of 1-41 (Hunt & Daigleish, 1994b).
4.2.4 Electrophoresis (SDS-PAGE)
The protein composition of liquid buttermilk, emulsion, oil droplets and semm
phases were determined by SDS-PAGE electrophoresis. Aliquots (200N) of fresh
butterrnillc, whole milk and skirn milk were added to electrophoresis buffer ( 10 mM Tris,
1 m M EDTA, 20 mM imidazole, pH 8.0), 300 pl of SDS 20% (w/v), 100 pl of 2-
mercaptoethanol and 100 p1 of bromophenol blue (0.05% wlv). SDS-PAGE was also
performed to determine the adsorption on the oil droplets of the emulsions prepared. To
obtain cream and serurn phases, the emulsions were separated by ultracentnfugation
(Beckman, preparative ultracentrifuge L8-70M, Pa10 Alto, CA) at 42000 g for 40 min.
Sample preparation for SDS-PAGE was carried out as descnbed by Hunt & Dalgleish
(1994b). Each sample (1 pl) was loaded into a 20% homogeneous "Phastgel" (Pharmacia
Biotech, Baie d'Urf6, Quebec), and run in a rapid electrophoresis systern (Phastsystem,
Phamacia Biotech) at 15'C. The gels were stained with Coomassie blue (PhastGel
Blue R, Pharmacia Biotech). After the separation, scanning densitometry was performed
with a scanner (Sharp JX330, Pharmacia Biotech). The scanned images were evaluated

with the "ImageMastef' software (Pharrnacia Biotech). Sample migration was compared
to that of known samples of purifiecl individual milk proteins (hi, a,*, and K-caseins,
a-lactaiburnin, p-lactoglobulin and BSA) and to published results on components of
MFGM (Keenan et aL, 1983; Basch et al., 1985; Jack & Mather, 1990). From this, the
identities and molecular weights of the proteins were established.
4 3 RESULTS AND DISCUSSION
43.1 Composition of the Material
Samples of industrial buttermillc contained considerable arnounts of caseins and
whey proteins, and so this product was different in protein composition from the aqueous
phase obtained after churning of washed cream (Mather & Keenan, 1975). The protein
compositions of skim mi&, buttermilk and whole milk were analyzed by SDS-PAGE.
The presence of high molecular weight polypeptides differentiated buttermilk from skim
and whole milk (Figure 4.1). Of the total protein, around 25% was represented by
MFGM proteins. Band 1 was identifid as xanthine oxidase. a complex iron-
molybdenum flavoprotein abundantly present in the MFGM (Mangino & Brunner, 1977).
Band 2 had an apparent molecular weight of 70000 Da. This band corresponded to
butyrophilin, a membrane-spanning protein with high affiïty for the fat globule
membrane (Keenan et al., 1983). This protein constitutes more than 40% of the total
protein associateci with the MFGM (Jack & Mather, 1990). A band at about 45000 Da
was also present (identifid as Band 3, figure 4.1). This band corresponded to two
distinct proteins PAS-6 and PAS-7, as reported by Baxh et al. (1985). These proteins
were the most abundant glycoproteins in bovine MFGM after butyrophilin

Band 1 +
Migration direction
Figure 4.1 : Typical SDS-PAGE gel of proteins obtained from skim millc (lane 1);
buttermilk (lane 2); whole mi& (lane 3). Analysis performed on 20% homogeneous SDS
gel, in reducing conditions. In order of migration the caseins are ag-, asl -, p- and K-
casein.

The protein composition of buttermilk was M e r analyzed by scanning
densitometry. The amount of MFGM-derived proteins constituted around 25% of the
total protein which migrated in the gel (assurning that the MFGM proteins stained
similarly to the other milk proteins). Skun miIk proteins (caseins and whey proteins, fb
lactoglobulin and a-lactalbumin) constituted 75% of the total protein in buttermilk.
These results were confmed by electron microscopy of buttermilk (Figure 4.2). The
industriai product contained a large quantity of casein micelles. Numerous fragments of
MFGM were also observed in buttermilk samples. The membrane sheets were
characterized by densely stained coat material. This dense coat material has been
reported to be on the face which was originally onented toward the fat globule interior
(Keenan & Dylewski, 1995) and it is resistant to ueatments with various salt buffers and
non-ionic detergents (Freudenstein et ai., 1979).
4.3.2 Emulsions Formed Using Buttermilk
We next determined the functionality of the protein present in the commercial
butterrnilk. The emulsifying properties of bunermilk were investigated by reconstituting
buttermiik with soyabean oil. The stability of the oil-in-water ernulsions was determined
related to the concentration of buttermilk solids. The freeze dried buttermilk used for
emulsion preparation contained 26.8 f 0.8 % (wlw) protein. The average size D 3 2 [
of the emulsion droplets as a hnction of buttermilk solids in a 10% (wlw)
oil-in-water emulsion is shown in figure 4.3. A decrease in the droplet size from 0.4 to
0.25 pm was observed as the buttermilk solids in the emulsion increased from 1% to
15%; however, only 1% of solids was necessary to obtaui a fairly smail average droplet
size. The particle size distribution of an emulsion containing 0.5% (wlw) buttemillc
solids and 10% (w/w) soyabean oil was birnodal, but at higher buttermilk concentration

Figure 4.2: Scanned TEM image of buttennilk, precipitated by acidifcation at pH 4.6.
The specimen was fixed with glutaraldehyde and post-futed with OsO4. Bar=0.3 p.m.
CM, casein micelle; C, coat material; M. membrane sheets.

O 2 4 6 8 10 12 14 16 Buttermilk Solids [%]
Figure 4.3 : Average droplet size (D3.2) of emulsions (10% oil wlw), related to
buttermilk concentration. Analysis carried out by integrated light scattering. The results
are the average of three independent expenments. The standard deviation of the
emulsions was lower that 0.03 Pm.

the distribution was monomodal and the emulsions were stable, i.e. they did not
aggregate during storage over a 3 day penod (Figure 4.4).
Surface coverage studies carried out by SDS-PAGE electrophoresis
demonstrated that caseins made up about 50% of the total protein adsorbed to the
interface. However, MFGM proteins and whey proteins were also present, and their
relative concentrations at the interface did not differ from their original ratio in
butte&. The protein patterns of the crearn samples were compared to those of the
whole emulsions to determine the type of protein adsorbed and any changes that might
have occurred at the interface (Figure 4.5). No difference was found between the protein
ratio in the emulsion and the one of protein adsorbed ont0 the oillwater interface, for
either caseins or MFGM proteins. Moreover, the relative concentration of caseins and
MFGM proteins was not changed by increasing the concentration of buttermilk solids in
the emulsion. The MFGM-derived proteins adsorbed at the owwater surface according to
their concentration in buttennillr, and even when the protein was present in excess, at up
to 15% solids, there was no preferential adsorption of the caseins over membrane
pro teins (Figure 4.5).
SDS-PAGE of the semm phase of the emulsions after centrifugation showed that
with increasing concentration of solids a greater variety of proteins was found in the
semm phase (Figure 4.6). At low buttermilk solids, the serum phase of the emulsion
contained only p-casein and traces of whey proteins. When 8% ( w h ) buttermilk solids
was used to prepare the emulsion, the serum phase contained not only p-casein but also
a , ~ -casein, P-lactoglobulin, a-lactaibumin and traces of MFGM proteins. The release
of p-casein into solution, even at very low concentrations of buttermilk, appears in
contradiction with the high hydrophobicity of the protein (Swaisgood, 1992). Among the
caseins, pcasein is the most hydrophobie protein, and it adsorbs more strongly on
oil/water interfaces than astaseins but it also displaces as-caseins from the interface

0.01 O. 1 1 10 100 Particle Diameter [pm]
Figure 4.4: Typical size distribution of emulsion droplets containing 10% soyabean oil.
Solid line, 3% (wfv) buttermilk solids; broken line, 0.5% (wlv) buttermiUc solids.
Analyses c h e d out by integrated Light scattering.

O 2 4 6 8 10 12 14 16 Buttemi i l k Solids [%]
Figure 4.5: Protein (56 relative to the total protein) migrated in 20% homogeneous gel
electrophoresis. Caseins present in the original emulsion (m); caseins in the cream
phase (O); MFGM proteins present in the original emulsion (a); MFGM proteins in the
crûam phase (O). Peak integration was performed &ter densitometric scanning on 3
independent experiments. Error bars indicate standard deviations.

1 a-lactalbumin
. . - P-lactoglobulin
Migration direction
Figure 4.6: Typical SDS-PAGE polypeptide pattern of the aqueous phase. separated by
cenuifugation, of 10% (wfw) soyabean oil emulsions with difTerent concentration (wlv)
of buttennilk solids. Lanes 1,5: 3% butkrmilk; lanes 2,6: 5% buttemilk; lane 3: 8%
buttermilk; lane 4: 15% buttemilk. Electrophoresis perfomed in reducing conditions.

(Nylander & Wahlgren, 1994). This release couid be explained by the dissociation of fk
casein from the adsorbed micellar caseins occurring d u h g storage at low temperature
(Holt & Home, 1996). For this reason experiments were also c h e d out by keeping
emulsions at room temperature (25°C) until separation of oil droplets and serum phase
by centrifugation. In such samples, pcasein was absent in the serum phase at 3% (wlv)
buttermilk solids, but at higher concentrations the bands of p-casein and a,l -casein were
the first to appear in SDS-PAGE gel. The increasing amount of protein found in the
serum phase at high buttermilk concentration confieci that, up to 8% (wlw) solids and
10% (wfw) oil, there was some excess of protein in the emulsions. As mentioned, even
when protein was in excess in the emulsions, no notable changes in the relative amounts
of proteins adsorbed at the oiYwater interface were determined by SDS-PAGE
electrophoresis, even when low amounts of residual protein were observed in the serum
phase (Figure 4.5). At buttermilk soiids concentrations > 8% (wlv), a small pellet
formed after centrifugation of the emulsion. In the pelleted fraction, a-lactaibumin was
absent, while the composition of the other proteins reflected that of the emulsions.
The amount of protein present and the concentration of oil were critical in
determinhg the stability of the ernulsions. At higher soybean oil concentration (20%
w/w), the emulsions did not show a monornodal particle siu: distribution, even when
20% (wlw) butterrnilk solids were incorporated. The distribution of oil droplet size for
emulsions with 12% (wlv) buttermillc was compared at two different oil concentrations
(Figure 4.7). The emulsions containing 20% (w/w) oil showed aggregation, and their
viscosity was so high that funher analyses were not possible. The results illustrate that
factors other than the protein-to-oil ratio were also important, because the ratios which
gave stable emulsions with 10% (wlw) oil were ineffective at 20% (w/w) soyabean oil.
In experiments carried out on buttermiuc (concentriated by ultrafütration) and soybean oil,
a two times concentrate buttermilk microfluidized with 20% wlw oil formed a stable

0.01 o. 1 1 10 100
Particle Diameter [pm]
Figure 4.7: Size distribution of emulsion droplets stabilized with 12% (wfw) buttermilk
solids. Solid line 10% (w/w) oil; broken line 20% (w/w) oi1. Analyses carried out by
integrated light scattering.

emulsion with a monomodal distribution of droplet sizes (data not shown). The arnount
of protein was, therefore, fundamental to achieve a complete coverage of the oillwater
interface. However, in the case of reconstituted buitermilk, the presence of small
molecuiar weight components might have affecteci the instability of the oil-in-water
emulsion forrned.
The protein present in the emuision was determined, by considering the protein
concentration of the freeze dried buttermilk. When 10% (w/w) soybean oil emulsions
were made, emulsion stability was achieved at protein concentrations < 0.5% (wlv).
According to Fang & Dalgleish (1993a) a very low arnount of sodium caseinate is
necessary to stabilize a 20% oil in water emulsion. However, in the butterrnilk, the
caseins were stiil present in micellar form and therefore, a higher quantity of protein was
required to cover the interface. At low concentrations of buttemilk solids, casein
micelles were not present in ~ ~ c i e n t amount to cover the interface. Therefore, we
assumed that other protein components contributeci to the stability of ihe emulsion.
Moreover, previous studies have shown that the pmence of phospholipids decreased the
amount of protein necessary to cover the oiVwater interface (Fang & Dalgleish, 1993b).
In those studies, a synergistic effect of protein and phospholipid was described whereby
phosphoiipids adsorbed in the gaps present at the interface, caused by insufficient protein.
The release of $-casein in the serum phase of the emulsions, dissociation observed at low
buttermilk concentrations (Figure 4.6) may occur not oniy by dissociation of the protein
from the micelles, but also via an interaction between $sasein and certain phospholipids
(Fang & Dalgleish, 1996). A hydrophilic complex can be found between DOPC
(dioleylphosphatidylcholine) and P-casein, which leaves the interface for the serum
phase (Fang & Dalgleish, 1996).
We combined the specific surface area of the emulsion droplet, determined by
integrated light scattering, with the pro tein concentration of the buttermiik in the
emulsions. Thus we could determine the surface load in emulsions with concentrations <

8% (wlv) buttermilk (Figure 4.8). since most of the protein was adsorbed at the interface
(Figure 4.6). The surface load appeared to reach a plateau at about 8 mg/m2, which
wouid indicate the presence of protein aggregates such as casein micelles on the surface.
This contrasted with previous observations that a monolayer of spread caseinate required
a minimum surface load of around 1 rng/m2 and was saturated at about 3 mg/m2 (Fang &
Dalgleish, 1993a).
An excess of protein seemed necessary for the emulsions to be stable to heat
treatment (Figure 4.9). At 4% (wlv) buttermilk solids, 10% (wlw) soyabean oil
emulsions showed instability after heat treatment at 90mC for 3 min (Figure 4.9A).
Only at concentrations > 8% (w/v) buttemilk, was the monomodal distribution retained
after heating (Figure 4.9B, C). No notable changes in the relative amounts of caseins
and MFGM proteins adsorbed to the interface were found. The main cause of instability
would be the interaction between adsorbed molecuIes, which would cause bridging
between oil droplets, especially with no excess of protein in solution.
4.4 CONCLUSIONS
Pretreatment of cream highly affects the nature and the composition of the
MFGM isolated from it. Most reponed studies on MFGM proteins have been
characterized by careful sample preparation to elhinate any skim miik proteins. The
characteristics of isolated MFGM proteins are only in part applicable to the understanding
of the functional properties of proteins deriveci from buttennilk. Because of their origin,
MFGM proteins are expected to be good emulsifiers, but in our samples this was not
found when commercial buttermik was used as an ingredient to prepare oil-in-water
emulsions. The contributions of al1 of the different constituents must be considered, and
generally the use of buttermillc in foods depends heavily on the properties of the caseins.

O 1 2 3 4 5 6 7 8 Buttermilk Çolids [O/]
Figure 4.8: Protein surface concentration as a function of total solids for ernulsions
containing 10% (w/w) soyabean oil. Values obtauied by combining the protein
concentration in the emulsions with the specific surface area of the emulsion droplets
obtained by integrated light scattering. Values calculated up to 8% ( w h ) buttermilk
solids emulsions, concentration resulting in an excess of protein. Results are the average
of three indipendent experiments; error bars indicate standard deviations.

0.01 0.1 1 10 1 O0
Particle Diameter [pm]
Figure 4.9: Changes in droplet size distribution of emulsions with 10% (w/w) soyabean
oil. (A) emulsions containing 4% (wlv) buttermilk solids; (B) emulsions containing 8%
(w/v) buttermilk solids; (C) emulsions containhg 12% (wlv) buttermilk solids. Solid
lines, results obtained from fresh emulsions; broken lines. results obtained after heating
the ernulsions at 90°C for 3 min.

Additional studies are required to defme the individual behaviour of the membrane
components when used in food emulsions.
4.5 STUDIES OF EMULSIONS STABILIZED BY S K I M MlLK
These experiments aimed at comparing the emuls*ing properties of the pro tein
in buttemilk to those of the proteins in skim millc. Skim milk and butte& have very
sirnilar protein composition; however, they differ substantially in the amount of MFGM
matenal, which is extremely low in skirn milk. In this section, the behaviour of
emulsions prepared with skim milk is compared to that of emuisions prepared with
buttcrmillc.
Low temperature skim milk powder (New Dundee Crearnery, New Dundee, ON)
was resuspended in 20 mM TrisMc1 buffer (pH 7.0), in a range of concentrations
between 0.25 and 15% (wlv). The ernuls-g properties of skim milk proteins were
investigated by preparing emulsions with skim milk solids and soyabean oil (10 and 20
wlw). Experiments were carried out at least in duplicate. Figure 4.10 illustrates typical
droplet size distributions of emulsions prepared with 10% (wlw) oil and various
concentrations of skim mi& solids. The ernulsions were charactenzed by a monomodal
distribution of droplet size even at low concentration of skim milk solids. The average
droplet size decreased from 0.8 pm to 0.30 pm as the amount of skim rnilk solids in the
emulsion was increased (Figure 4.1 1). These results differed substantially from those
reported for ernulsions prepared with freeze dried buttermilk. Ln oil-in-water emulsions a
buttemilk solids concentration of 1% (wlv) was needed to achieve a monomodai droplet
size distribution (Figure 4.4).
At low concentrations of skim milk solids, most of the caseins in the emulsion
adsorbed ont0 the surface of the oil droplets (Figure 4.12). When the amount of skim

0.01 o. 1 1 10 1 00
Average Diameter [pm]
Figure 4.10: Size distribution of emulsion droplets containhg 10% (wfw) soyabean oil
and 0.25% (wh) (solid h e ) , 1% (w/v) (dotted line) and 8% (wfv) (broken line) skim
milk solids. Analysis camed out by integraied light scattering.

O 2 4 6 8 10 12 14 16 [% Total Solids]
Figure 4.11: Average droplet size of 10% (w/w) oil-in-water emulsions as a function of
skim mi& solids. Analyses carried out by integrated Light scattering. Resuits are the
average of 2 independent experirnents. Standard deviation was less than 0.04 Pm.

O 2 4 6 8 10 12 14 16 [% Total Solids]
Figure 4.12: Caseins, % relative to the total casein present in the emulsion, adsorbed
onto the oil droplets as a function of skim milk solids. Results are the average of at least
two independent experiments. Error bars indicate standard deviations.

milk soiïds in the emulsions was increased, the casein adsorbed at the interface decreased
and the arnount (determined as percentage of the total caseins present) seemed to reach a
plateau around 40%. Since the average diameter of the emulsion dropleü and their size
distribution did not change when the concentration of skim miik solids was increased, it
was assumed that the surface load of the protein increased with more skim milk solids
present in the emulsion. It is known that caseins adsorb at interfaces in monolayers at
low concentrations (spreading of the flexible molecules can be observed), and fonn
multilayers at higher concentration, when more protein is available during
homogenization (Fang & Dalgleish, 1993a). The high amount of caseins adsorbed onto
the oil droplets could also be explained by the presence of casein micelles at the oiYwater
in terface.
Analysis of the protein composition of the original emulsion, the serum phase and
the oil droplets clearly showed that caseins adsorbed preferentially to whey proteins in
skim milk emulsions (Figure 4.13). The serum of emulsions prepared with 10% oil and
3 8 (w/v) skim milk solids contained whey protein and no casein. When emuisions were
prepared with 8% (w/v) buttemik, the semm phase contained p-casein, cql-casein. P-
lactoglobulin, a-lactalbumin and traces of MFGM proteins (Figure 4.6). The presence of
P-casein and c casein in was also observed in the serum phase of emulsions prepared
with skim n d k (Figure 4.13).
SDS-PAGE showed the difference in adsorption behaviour of skim miik and
butterma proteins. Up to 8% (w/v) buttermilk solids, no unadsorbed protein was found
in the serum phase of the emulsion. These results demonstrated that all the protein
adsorbed at the interface. On the other hand, in emulsions with only 3% (w/v) skim milk
solids, some whey protein was unadsorbed, as shown by SDS-PAGE of the serum
phases. Neither a-lactalbumin nor P-lactoglobulin adsorbed at the interface of skim milk
emulsions with < 15% (wlv) solids, as shown in figure 4.13.

Figure 4.13: Typical SDS-PAGE electrophoresis of semm phase, oil droplets and original emulsions prepared with different concentrations (wlv) skim milk solids and
10% (w/w) soyabean oil. Lane 1: emulsion 0.25%; Lane 2: semm 0.25%; Lane 3: emulsion 0.5%; Lane 4: s e m 0.5%; Lane 5: emulsion 3%; Lane 6: serum 3%; Lane
7: emulsion 6%; Lane 8: serum 6%; Lane 9: emulsion 8%; Lane 10: oil droplets 81;
Lane 1 1: emulsion 15%; Lane 12: oil droplet 15%. SDS-PAGE performed in reducing conditions.

Our results on the preferential adsorption of caseins a g r d with reported
information on protein adsorption in reconstituted or homogenid mik (Darling &
Butcher, 1978; Sharma & Dalgleish, 1993; Shaxma et al., 1996). Preferential
adsorption, as aiready suggested by Walstra & Oortwijin (1982), was caused durllig
homogenization by the turbulence-controlied mass trans fer whic h facilitated the
adsorption of large particles (casein miceiles) more than smail particles (whey proteins).
The absence of whey proteins on the oil droplet surface did not c o n f i what has been
suggested by Sharma et ol. (1996)- but was in cornplete agreement with S h m a &
Dalgleish (1993) who found no whey proteins at the fat globule surface of unheated
hornogenized milk.
The results of skim emulsions confirmeci what has already been found for
butterrnilk emulsions: the protein-to-oil ratio was not the only factor detemining the
stability of oil-in-water emulsions. Figure 4.14 illustrates the particle size distribution of
an emulsion prepared with 8% (wlv) skim milk solids and 10 or 20% (wlw) soyabean oil.
In this case also, the presence of smaU molecular weight components could explain, in
part, the instability of the emulsions. The presence of large aggregates (in s k h mi&
mostly casein micelles, and in buttermik also MFGM matenal) in the aqueous phase
rnight have caused an osmotic pressure gradient which led to aggregation of the emulsion
droplets by depletion flocculation; this rnechanism has been described for caseinate
emulsions @ickinson & Golding, 1997).
Skim milk stabilizeà emulsions presented a few differences from buttemiik
stabilized emulsions. A summary of the differences is given in table 4.1. An amount of
skim rnilk solids lower than that of buttemilk solids was necessary to achieve emulsion
stability. Preferential adsorption of caseins was also observai in skun milk ernulsions.
ButtermiUr emulsions did not have unadsorbed protein in the semm at concentration of
solids < 8% (wlv), whüe the serum phase of skim milk emulsions contained whey
protein at 3% (wfv).

0.01 0.1 1 10 1 O0
Average Diameter [pm]
Figure 4.14: Typical droplet size distribution of emulsions prepared with 8% (wlv)
skim milk proteins and 10% (solid h e ) or 20% (wlw) (dotted line) soyabean oil.
Analysis carried out by integrated light scattering.

The main differences observed could be attributed to two major causes: (i), skim
milk powder did not contain MFGM material, which in buttermilk showed very poor
emulsifying properties; (ii), skim rniik was obtained from a low temperature spray
drying process (which probably resulted in very iittle heat darnage to the protein), whiie
the freeze dried buttermilk was coliected after churning of cream pasteunzed at high
temperature (85°C for 16 s).
Table 4.1: Comparative table summarizing the major differences observed between skim
milk and buttermilk emulsions.
Concentration of soiids needed to achieve a monomodal distribution of sizes ( 10% oil-in-water ernulsion) (w/v) Preferential adsorption at the interface SoIids concentrations necessary to observe presence of unadsorbed pro tein (w/v) Type of protein present in the semm phase
Buttermik emulsions Skim m i k emulsions
pref. adsorption of caseins vs. whey proteins
p casein and CQ -casein B-lactoglobulin and a-Iactalbumin

CHAPTER 5*
ISOLATES FROM INDUSTRIAL BUTTERMILK: EMULSIFYING
PROPERTES OF MATERIAL DERIVED FROM THE MILK FAT
GLOBULE MEMBRANE
5 . 1 INTRODUCTION
Buttermiik, which is the aqueous phase released during the manufacture of butter,
contains not only skim milk proteins but material. mainly composed of proteins and
phospholipids, denved from the original milk fat globule membrane (MFGM). This
characteris tic composition makes buttermilk an interesting source of ingredients with
unique functionai properties arnong milk-derived products. The growing interest in the
dairy industry for new products has led to an increase in the number of studies on
buttermik (Ramachancira Rao et al., 1995; Mistry et al., 1996); however, the full poten tial
of this byproduct has yet to be exploited.
in untreated milk and cream, the MFGM is characterized by a complex mixture of
proteins, phospholipids and glycoproteins, and acts as a natural emulsifier (Keenan et al..
1983). Although littie is known about their functional properties, much progress has been
made in the purifïcation of the proteins from the membrane (McPherson & Kitchen, 1983;
Basch et al., 1985; Kim et al., 1992). The proteins from MFGM have been identitird
accordhg to their migration during SDS-PAGE (Keenan & Dylewski, 1995). Two
polypeptides, xanthine oxidase (MW 150 000 Da) and butyrophilin (MW about 67 000
Da) account for nearly 50% of the total protein in the MFGM material, as detected by
comassie blue stainùig of electrophoretic gels (Greenwalt & Mather, 1985).
Most studies on MFGM proteins have been carried out with membranes released
and coflected from washed, untreated milk fat globules (Keenan & Dylewski, 1995). It
*Published essentially in this f om as Corredig & Dalg leish, 1 997. Journal of Ag ricultural and Food Chemistry 45. 4595-4600.

has b e n reported thai., with preireaunent of the crearn, losses of membrane material may
occur during the washing steps (Mc Pherson & Kitchen, 1983; Bash et aL, 1985),
however, at present there appears to be no alternative procedure for obtaining MFGM
proteins free. from skim milk proteins. These MFGM play a very important role in
stabiiizing fat globules in cream and they have been used to monstitute milk fat in
emulsions (Kanno et al., 199 1 ; Oehlmann et al., 1994). It has been reported that MFGM
proteins contained in butîermilk enhance the func tional properties of this b yproduct
(Kanno, 1989; Mistry et al., 1996).
The increasing interest in MFGM proteins as food ingredienis and in their
fùnctional properties makes it important to study mem brane-derived material isolated from
indus triai butterm ilk rather than from bu t t e d producecl from laboratory procedures.
The research described here aimed at isolaàng rnembrane-denved matenal frorn an
industrial source of butte& and investigating the potentiai utilization of the byproduct as
an ingrdient of oil-in- water emuisions.
5.2 MATERIALS AND METHODS
Buttermilk was coilected from the butter production line of a local dairy and 0.02%
(wlv) sodium azide was added as a preservative. When freshly coiiected buttermilk was
centrifuged (Beckman, preparative ultracentrifuge, mode1 L8-70M) at 15°C for 50 min at
lûûûûûg, the sedirnented peliet contained an isolate ("buttermilk isolate") with a protein
composition identical to that of butterrnilk.

5.2.1 Preparaîion of MFGM Isolate
Calcium is essentiai for maintaining the structure and stability of casein micelles
(Holt et al., 1986), and consequently they can be dismpted by the addition of the calcium-
chelating agent sodium citrate. In buttemilk, citrate added at concentrations between 2 and
5% (wlv) provided the required breakdown of the miceDar süuc tue. The addition of
sodium citrate did not substantialiy alter the pH of buttermik (in most cases the increase in
pH was l e s than 0.5 unit-). In practice, 2% (wlv) tri-sodium citrate was added to the
buttermiik, and the mixture was incubateci ovemight at 4°C. The treated samples were
then centrifugeci at 1OOOOOg for 50 min at 15°C and the resulting smaii pellets were
collected and dned on a fdter paper (Whatman, no. 4), resuspended in a minimum amount
of MilliQ water with a tissue homogenizer (Polytron PT 2000, Kinematica AG,
Switzerland) and freeze dried. The freeze dried sarnples were stored at -20°C; these are the
MFGM isolates. By analysis of the serurn after centrifugation, it was established that
virtuaiiy all of the membrane material from the buttennilk was collected in the pellet.
5.2 .2 Transmission Electron Microscopy
The pellet fractions isolated by ultracentrifugation were dned on fdter papar
(Whatman no. 4) and treated overnight with 1.4% glutaraldehyde in 0.05 veronal acetate
buffer (pH 7.0). The preparations were then washed in buffer and fmed for 24 hours in
imidazole and Os04 solution at 0.5%. The samples were then dehydrateci in a graded
alcohol senes (Kalab, 1980) and ernbedded with epon resin (Luft, 196 1). Post staining
and was carried out as described in paragraph 4.2.1. The micrographs were produced by
a Hitachi transmission electron microscope with a measuring voltage of 75 kV.

5.2.3 Phospholipid Determination
Freeze dried samples (30 mg) were resuspended in 6 ml of 0.5% (wh) KCl
solution andextracte. with 30 ml of chloroform-methanol in a 2:l ratio. The two phases
were separated by low speed centrifugation (2000 g for 5 min) and the lipid was dried by
rotary evaporation. The dry sample was then resuspended in 0.5 ml of
chlorofomi/methanol(2: 1) and fütered with 0.2 pm fdter (Gelman Sci. Ann Arbor, Mi).
Sarnples were spotted at the bottom of chromatorods (Iatron Laboratones. Tokyo, Japan).
A two step solvent development was carried out: (I), chlorofonn/methanoVwater
(50:20:2.5 vfv) up to 5 cm migration; (2), exanddiethyl etherfacetic acid (65:5:0.5 v/v) for
a 10 cm migration. Mter the two development steps. the lipid migration was detennined
by scanning with Iatroscan (Iatron Laboratories, Tokio, Japan) equipped with a flame
ionization detector (Shantha, 1992; Tvnicka & Votmba, 1994). The lipids present in the
sample were identii5ed by cornparison with the migration of standard phosphoiipid sarnples
(Avanti polar lipids Inc., Alabaster, Al), and standard samples of triolein and cholesterol
(Sigma, St. Louis, Mo).
5.2.4 Emulsion Preparation
Aliquots of freeze dried protein isolate (either butiennilk or MFGM isolate),
soybean oil and buffer (20 mM TridHCI buffer at pH 7.00) were rnixed in different
proportions. The mixtures were homogenized using a Microfluidizer as described in detail
in paragraph 4.2.2. Experiments were conducted at least in triplicate.

5.2.5 Determination of Particle Size Distribution and Protein Adsorption
The diameter distribution of the fat globules in the emuision and their s p e ~ ~ c
surface areas were determined by integrated Light s c a t t e ~ g as described in paragraph
4.2.3.
5.2.6 Electrophoresis (SDS-PAGE)
The protein compositions of the isolates were deterrnined by SDS-PAGE. Aliquots
of freeze dried samples (approximately 3 mg) were resuspended in 200 jU of
electrophoresis buffer (10 mM Tris, 1 m M EDTA, 20 mM irnidmle, pH &O), rnixed with
300 pl of 20% (w/v) SDS and LOO pl of 2-mercaptoethanol and bromophenol blue (0.051
wfv). The emulsions prepared with the isolates and the corresponding creams and serum
phases were separated by ultracentrifugation (Beckman) at 15°C for 40 min at 10000 g and
analyzed by SDS-PAGE as described earlier (4.2.4).
5.3 RESULTS AND DISCUSSION
5.3.1 Production of Isolates from Buttermilk and Their Composition
Both MW;M isolate and buttermilk isolate contained approximately 40% lipid, in
agreement with the reports of other authors (Kanno & Kim, 1990). The moisture contents
of the freeze-dried isolates were low (-34% w/w). After high speed centrifugation of
buttermilk, SDS-PAGE showed that the pellet of buttermilk isolate had a protein
composition sirnilar to that of the whole buttemiillc. The isolate contahed the four caseins,
a-lactalburnin, P-lactoglobulin and proteins derived frorn the fat globule membrane, with
caseins making up approximately 50% of the total proteins. This agreed with previous

results; it has been reported that skim milk and buaermilk have very sirnilar protein
compositions (Chapter 4). In our study, the presence of skim milk proteins hindered the
detemination of the physical and chernical properties of MFGM proteins. In cream. most
of the skim mik proteins can be removed by pretrea~g the milk fat globules prior to
extraction of the MFGM proteins (Basch et al., 1985), but in the industrial buttermilk, the
presence of caseins and whey proteins was unavoidable. We therefore used the method
described above (5.2.1) to separate the MFGM fraction from the caseins.
Figure 5.1 shows the SDS-PAGE of the two different isolates. The pellet from the
buttermilk after treatment with citrate contained a large quantity of MFGM proteins, and in
this isolate casehs were present only in small amounts. The MFGM isolate differed
substantiaily in composition from the butte& isolate. It contained three major
polypeptides migrating in the high molecular weight range of the SDS-PAGE gel (MW >45
000): band 1, identifiable as xanthine oxidase (MW 15ûûûû), band II, at about 70000 Da,
butyrophilin, and band III, migrating at approxirnately 50000 Da (Keenan & Dylewski,
1995). Another band, of mobility higher than that of band III, was also detected, and has
not b e n identifed. The electrophoretic migration of the MFGM isolate was similar to that
reported for MFGM proteins derived from native fat globules (Kanno & Kim, 1990; Maiin
et aL, 1994). In addition, the MFGM isolate contained large amounu of B-lactoglobulin,
which appeared to be covalently linked, by disulphide bonds with the MFGM. This
association of P-lactoglobulin with MFGM proteins was almost certainiy the result of the
heat treatment of the cream pnor to buttermaking. Several studies have been canied out
on the interactions of whey proteins with fat globule membranes during heating of whole
milk and cream; however, these interactions are not W y understood (Dalgleish & Banks,
1991; Kim & Jimenez Flores, 1995; Corredig & Dalgleish, 1996b). The presence of P- lactoglobulin and the almost complete absence of a-lactalbumin in the MFGM-isolate from
industrial buttends confirmeci the hypothesis that a major interaction occurring during

. - 1 At 3 - 4.- 5: . 6 Migration _ _. ._ _ _
Buttermilk MFGM Buttermilk
Figure 5.1: Typical SDS-PAGE of freeze dried material from buttemik Alïquots (3 mg) of material were resuspended in 200 pi of electrophoresis buffer, 300 pl SDS and LOO pl of mercaptoethanol and bromophenol blue. Analysis performed on 20% homogeneous SDS gel. Lane 1: Butte&; lanes 2,3,4: MFGM isolate; lanes 5,6: buttermilk isolate.

heating of cream is the reaction between fblactoglobului and the MFGM proteins. There
were also in the MFGM isolate, quantities of aggregated proteins which did not enter the
gel. These are probably large complexes formed when the cream is heat-treated, which do
not break down in the presence of SDS and mercaptoethanol. These complexes were also
present, but to a srnaller extent, in the buttermiik isolate.
The two isolates (buttermilk isolate and MFGM isolate) produced from industrial
butîennilk were also analyzed by transmission elecuon microscopy (TEM). These
observations confirmed the results obtained by SDS-PAGE. The buttermiik isolate,
derived from unireated buttemi&, contained a large amount of casein miceiles (Figure 5.2
AB). The polydisperse population of micelles coexisteci with fragments of membrane
material, which were characterized by a size varying from 0.4 to 1.5 p. On the other
hand, the MFGM isolate contained a large nurnber of the sheet-like membrane fragments
and many fewer casein miceiles than did the buttermilk isolate (Figure 5.2 C,D). The
sections obtained from MFGM isolate were very sirnilar to those reportai in the literature
for purified MFGM material from washed native fat globules (Keenan et al.. 1977;
Keenan & Dylewski, 1995). The membranes were characterized by a dense-staining
material which was oriented toward the interior of the milk fat globule in native crearn
(Figure 5.2 CTD) (Keenan et al., 1977). Previous work had demonstrated that this coat
matenal retained its structure even after washing of the membrane fractions with salt and
various detergents (Freudenstein et al., 1979).
The lipid composition of the two protein isolates was determined by thin layer
chromatography (Figure 5.3). The buttermilk isolate contained many neutral iipids with a
smailer arnount of phospholipids; about 65% of the total lipids consisted of trïglicerides
and cholesterol with the remaining lipid fraction containing rnainly phosphatidyl
ethanolamine, phosphatidyl choline and sphingomyelin (Figure 5.3). In contrast, the
MFGM isolate contained a significantly lower amount (40%) of apolar lipids than the

Figure 5.2: Transmission electron micrograph of pellet isolated by centrifugation of
butterrnik. The specimen was fixed with glutaraldehyde and post fixed with OsO4. A,B
peiler buttemilk (protein isolate); C,D pellet from MFGM isolate. Bar size: 1.1 p (A.
C); 0.70 pm (B, D).

Figure 5.3: Lipid composition (%) as determined by TLC Oauoscan); samples of
buttermiik isolate (solid) and MFGM isolate (pattern). Values are the average of 3
independent expenments, error bars representing the standard deviation.

buttermik isolate. These results agreed with the reported phosphofipid composition of the
fat globule membrane (Kanno, 1990; Malmsten et al., 1994). The selective
centrifùg ation of membrane fragments occuming during the preparation of MFGM isolate
decreased the arnount of apolar Lipids presenk possibly because in the original buttermilk
fraction smaU fat globules were complexed with casein miceiles and CO-sedirnented with
them. In the MFGM isolate phospholipids constituted approximately 60% of the total
lipids with the foilowing composition: 33% phosphatidyl ethanolamine, 16% phosphatidyl
choline, 9% sphingomyeh.
5.3.2 Emulsions Prepared with Isolates from Buttermilk
Figure 5.4 illustrates the size distribution of emulsion droplets prepared with 10%
(w/w) soybean oil. When buttermilk isolate was used, the emulsions were characterized
by a monomodal size distribution, at fairly small concentration of protein (1-2% w/v of
protein isolate) (Figure 5.4A). On the other hand, the 10% oil-in-water emulsions
prepared with MFGM isolate showed aggregation behaviour and bimodd distribution of
droplet size up to 4% (w/v) of the isolate. Only by using a much higher concentration of
MFGM isolate was a monomodai distribution of droplet size achieved (Figure 5.4B).
Figure 5.5 illustrates the change in average droplet size, expressed as D3.2, with
increasing concentration of MFGM isolate in emulsions with 5 and 10% (w/w) soybean
oil. While at Ieast 2% (w/v) of MFGM isolate was needed to obtain an average droplet
size of 0.4 pm in 5% oil emulsions, 4.596 (w/v) of MFGM isolate was necessary to obtain
an average size of 0.25 pm in 10% oil emulsions (i-e., as the amount of emulsion surface
increased, the amount of protein necessary was also increased). The amount of pro tein
necessary to produce emulsions with smaU droplet size was higher than the 0.5- 1%
required for emulsions prepared with isolateci milk proteins (Fang & Dalgleish, 1993a;

Figure 5.4: Typical particle size distribution of 10% (w/w) so ybean oil-in-water
emulsions. Analysis carried out by integrated light scattering. A: Emulsions prepared
with buttermik isolate at 1% (wh) (solid line), and 2% (w/v) (broken line). B:
Emulsions prepared with MFGM isolate at 4% (wlv) (solid h e ) , and 8% (w/v) (broken
iine) .

0 1 2 3 4 5 6 7 8 9 1 0 MFGM lsolate [%]
Figure 5.5: Average droplet size of emulsions as a function of MFGM isolate
concentration. Emulsions prepared with (0) 5% (wlw) soybean oil and (a) 10%
(w/w) sûybean oil. Error bars represent standard deviation. Average of at least 3
measurements. Analyses canied out by integrated light scattering.

Hunt & Dalgleish, 1994b). The different behaviour observed between the whole ernulsion
made with buttermilk isolate and the MFGM isolate may be attributed to the difference in
protein composition between the two isolates and the structural organization of the
membrane material.
A complete coverage of the interface took place only at high concentrations of
MFGM isolate (i.e. > 5% w/v in 10% oïl-in-water emulsions), because of the large
aggregates of membrane matenal present in the MFGM isolate. In the emulsions prepared
with buttermi& isolate. a large amount of casein micelles was present, and the required
surface coverage could be achieved with a lower protein concentration than that of the
emulsions prepared with MFGM isolate. Figure 5.6 iilusuates the protein composition.
as determined by SDS-PAGE, of whole emulsions and oil droplets separated by
centrifugation. No preferential adsorption of MFGM proteins compared to caseins or whey
proteins seemed to occur at the interface, and the arnounts of the different proteins adsorbed
depended on their concentrations in the original ernulsion. in emulsions made with
buttermilk isolate, most of the proteins adsorbed at the interface were of skim milk origin:
caseins and whey proteins (Figure 5.6). In contrast, in the MFGM isolate emulsions very
few caseins were present at the oiVwater interface (shce the fraction contained littie casein).
In addition, a large amount of fblactoglobulin was present in the MFGM isolate emulsion
and its oil phase. A more detailed analysis by scanning densitometry of the three main
MFGM protein bands present in the MFGM isolate emulsions and oil phases suggested that
no preferential adsorption occurred among MFGM proteins.
We have noted that both of the isolates contain quantities of phospholipid, which is
generaüy known to be highly surface active and to at least enhance the capacity of proteins
to form emulsions (Fang & Dalgleish, 1993b), although the mechanisrn of the action is not
well understood, because their interaction with the interface in the presence of proteins may
be smaU (Fang & Dalgleish, 1993b; Courthaudon et al., 1991~). However, it is evident

t a-lactalbumin
t P-lactoglobulin
B M 2% 3%
em. cream , m m M m M em. em.
Figure 5.6: Typical SDS-PAGE gel of proteins obtained from sarnples of 10% oil-in-
water emulsions and resuspended cream (oil droplets resuspended in the same volume of
die original emulsion). Analysis perfomed on 204 homogeneous gel, in reducing
conditions. Lane 1: 1% (w/v) buttemilk isolate emulsion; Lane 2: oil phase from
emulsion in lane 1; Lane 3: 2% (w/v) MFGM isolate emulsion; Lane 4: oil phase from
emulsion in lane 3; Lane 5: 3% (w/v) MFGM isolate emulsion.

that in the isolates descnbed here, the phospholipids seem not to exert an enhancing effect
on the emuls+g capacity of proteins. This suggests that the effect of heat matment on
the MFGM material in the buttefmilk is not only on the proteins but also affects the
hinctional properties of the phospholipid part of the membrane isolates.
The poor emulsifying properties of MFGM proteins obtained from industrial
buttermilk were c o d i e d by addition of s m d quantities of sodium caseinate to emulsions
prepared with MFGM isolate. When 2.5% (wlv) MKiM isolate was used, the 10% oil-
in-water emulsions showed a bimodal size distribution (Figure 5.7). The same emulsion
prepared with the addition of 0.5% (w/v) caseinate was charac~rized by a monomodal
droplet size distribution. The instabiiity of emulsions prepared with low concentrations of
MFGM isolate was caused by incomplete coverage of the oiVwater interface by the large
clumps of membrane material, which formed bridges berneen the ernukion droplets (see
next chapters). We have shown above that if enough MFGM isolate is used, stable
emulsions cm be made, so that the initial instability was not caused by a tendency of the
MFGM itself to aggregate. The bndging was prevented by the addition of casein, which
ailows more complete coverage of the interface of the dropleis, so that a sharp monomodal
distribution of droplet size could be obtained (Figure 5.7).
5.4 CONCLUSIONS
Although the role of MFGM proteins in the stability of the native milk fat globule is
recognized (Keenan & Dylewski, 1995), the membrane matenal released in solution after
some buttermaking processes is characterized by poor emulsifying properties. The
process of heat treatment, churning and the release of membrane fragments to the water
phase causes a high degree of aggregation and changes in the functional properties of
membrane proteins and phospholipids. The microfluidization process did not allow the

0.01 O. 1 1 10 1 O0
Diameter [ym]
Figure 5.7: Changes in particle size distribution of 101 oil-in-water emulsion prepared
with 2.5% MFGM isolate (solid line) and with 2.5% MFGM isolate and 0.5% caseinate
(broken line).

MFGM proteins present i~ this membrane aggregate to be released into solution and
migrate to the interface to act as ernulsifers. It is possible that the ngidity of the MFGM
buttennilk was the result of heat treatment of cream at source. It is known that interactions
occur during heat treatment between skim milk proteins and MFGM proteins @algleish &
Banks, 199 1); and the large amounts of ~lactoglobulin in the MFGM isolate from
industrial buttennilk could be used as further evidence of this. The present study
demonstrates that industrial buttennilk rnay contain MFGM proteins with poor hinctional
properties. The complex struchue of the membranes in the MFGM isolates did not allow a
complete coverage of the ouwater interface, r e s u i ~ g in an unstable emuision at low
MFGM isolate concentration. These resuits are in strong disagreement with the data on
MFGM proteins and emulsions prepared with butteroi1 (Kanno et al., 199 1 ; Oehlmann et
al., 1994). However, previous authors used raw-cream buttemilk as the MFGM protein
source. The industrial origin of buttermilk used in this research makes the results difficult
to compare with the previous literature.
It is reportai that the fat globule membrane present in commercial buttermilk may
enhance the emulsifcation properties of cheese (Mistry et al., 1996). The large amount of
casein micelles may, however, overcome any effect deriving from the other component
present in a srnalier amount.

CHAPTER 6*
CHARACTERIZATION OF THE INTERFACE OF AN OIL-IN-WATER
EMULSION STABILIZED BY PROTEIN DERIVED FROM THE iMIJ.lK FAT
GLOBULE MEMBRANE
6.1 INTRODUCTION
The emulsifying properties of miik fractions have been widely studied (Singh et
al-. 1993; Dalgleish, 1995). While milk protein isolates, casehates and whey protein
concentrates are commonly used as functional ingredients in a wide variety of food
products, less attention has been given to the protein fraction derived from the mik fat
globule membrane (MFGM).
Although MFGM proteins represent a very small fraction of the total protein
present in milk, they are fundamentai for the stability and the integrity of the fat globule
membrane in whole milk and cream; the MFGM acts as a natural emulsifier, aiiowing
fat globules to remain dispersed in the water phase. The MFGM is mainly detived from
the apical plasma membrane of mammary secretory cells and it consists of a complex
mixture of phospholipids, glycolipids and proteins (McPherson & Kitchen, 1983). While
progress has been made on understanding the biological function of these proteins during
synthesis and secretion of fat globules, very little is known of their behaviour in
reconstituted systems such a s emulsions (Keenan & Dylewski, 1995).
Because of its origin, many authors have attributed to MFGM a high ernulsifying
capacity (Kanno, 1989; Oehlrnann et al., 1994). Recently some work has been camied
out on investigating the potential uses of MFGM as a stabilizing agent in d a q emulsions
and low-fat cheese (Kanno et al. , 199 1 ; Mistry et al ., 1996). The objective of our work
was to obtain more information on the surface properties of isolated MFGM. For this
purpose the MFGM, contaking protein and phospholipids, was extracted from fresh raw
*Accepted for publication essentially in this form by the Journal of Dairy Research, January 1998.

cream. Emulsions were prepared with soyabean oil using MFGM isolate as emulsifier.
The types of proteins adsorbed at the interface of the oil-in-water emulsions and their
changes in composition were studied. Dynamic iight scattering was combined with
particle electrophoresis to measure the apparent <-potenrials of the surfaces of the
emulsion droplets. In addition, the s tability of the emulsions after heating and
acidification was studied. The information derived from these results aliow a
cornparison to be made between the properties of emulsions prepared with MFGM
isolates and the propenies of the emulsions stabilized by the weU characterized skim milk
proteins.
6.2 MATERIALS AND METHODS
Soyabean oil, buffer salts, sodium citrate, 2-mercaptoethanol, Tween 20, Tween
80, Triton X- 100 were purchased from Sigma chernicals (St. Louis, MO). Lysozyme was
also purchased from Sigma. Whole casein was prepared in the laboratory by acid
precipitation from skim milk as descnbed previously (Fang & Dalgleish, 1993a) and f$-
lactogiobulin was purified by ion exchange chromatography on Sepharose Fast Row Q
(Pharrnacia Biotech, Baie D'Urfé, Quebec) from a sample obtained from Protose
Separations Inc. (Teeswater, ON).
6.2.1 Preparation of the Membrane Isolate
Fresh raw cream (about 40% fat) was coiiected from a local dairy, after
separation from whoie unpasteurimd rniik. M e r addition of 0.02% sodium azide as
preservative, the cream was kept at 4'C ovemight. The cream was churned at 10' C in a
laboratory-scale churn (1 litre capacity) until phase separation occurred. Buttermillc,
released from the butter granules, was fütered through a cheesecloth. The MFGM isolate

was prepared from butterrnilk as described in 5.2.1. Sodium citrate (2% wlv) was added
to buttermilk and the samples were centrihiged at 15'C for 50 min at lOOOOOg in a
preparative ultracentrifuge (Beckman). Buttermilk isolates were also prepared by
centrifugation of buttemilk without the addition of sodium citrate. ButtenniUc isolates
and MFGM isolates were composed of proteins and phosphoiipids, but this paper
describes m a d y the results obtained from observations of the protein fraction in the
isolates.
6.2.2 Emulsion Preparation
Aliquots of freeze dned isolates were resuspended in buffer (20 rnM TrisMCl) at
pH 7. Soyabean oil (10% wlw) was then added and the mixture was homogenized usine
a microfluidizer, as described in paragraph 4.2.2.
6.2.3 Measurement of Particle Size and S tability of the Emulsions
The diameter distribution of the oil droplets in the emulsions was determined by
integrated light scattering as described in detail in paragraph 4.2.3. The particle size
distribution and the volume-to-surface average diameter (d32) were used as indices to
characterize the different emulsions and their aggregation during storage, heating and
changes of pH.
Particle size rneasurements were made on emulsions soon afier they were made
and after storage in a refrigerator (4°C) for 10 days.
Aiiquots (10 ml) of fresh emulsions were heated at 90°C for 5 min in a water bath
and immediately cooled to room temperature in an ice bath. Their particie size
distribution was determined both immediately after heating and after 10 days of storage.

The pH of ihe emulsions was lowered by slow addition of 1 M HCI on freshly
made ernulsions, and their droplet size distribution was determined as the pH changed.
Repeated experiments showed that emulsions could be made with differences in
d32 of no more than 0.05 p m for d 32 less than 0.6 Pm, and no more than 0.15 p.m for
d32 higher than 0.6 Pm, between different preparations with the sarne composition.
6.2.4 SDS-PAGE
The protein composition of two isolates prepared from buttermilk was determined
by SDS-PAGE on freeze dried material. Approximately 3 mg of sample were suspended
in 200 pl of electrophoresis buffer (10 mM Tns, 1 m M EDTA, 20 rnM imidazole, pH 8).
300 p l of SDS (20%), 100 p l of bromophenol blue (0.05%) and 100 p l of 2-
mercap toethanol.
The composition of the protein adsorbed at the interface of the oil-in-water
ernulsion was detennined as described previously (paragraphs 4.2.4 and 5.2.6). The oil
phase of the emulsions was separated by ultracentrifugation (Beckman) at 10000 g for 40
min at 15'C. After carefully collecting the subnatant using a synnge with a long needle,
the oil droplets were draineci on füter paper (Whatrnan no. 4) and resuspended with
electrophoresis buffer to a fmal concentration of 10% w/v. By comparing the protein
composition of the original emulsions to that of the oil droplet and of the serum phase, it
was possible to determine any difference in the adsorption between the different
components of the MFGM isolate.
The c-potentials of the oil-in-water emulsions prepared with I and 1.5% (wh)
MFGM isolate and 10% (w/w) soyabean oil were determined using a Malvem Zetasizer

(Malvern Instruments Inc.) attached to a 7032 correlator. MFGM proteins in isolation
have isoelectric points ranging from 3.5 to 8.0 (Mather et al., 1980: Kim et al., 1992).
The changes in c-potentials of emulsion droplets stabilized by caseinate, which has an
isoelectric point of pH 4.6, have been studied (Dalgleish et al.. 1995). To be able to
compare the changes in 6-potentid as a function of pH of emulsions prepared with
MFGM with those of emulsions stabilized by protein with a high isoelectric point.
emulsions made using lysozyme were also studied. The behaviour of this protein, with
an isoelectric point of 1 1.5, both in solution and at solidlwater interfaces has been
reported (Norde & Favier, 1992). The c-potentials of an emulsion prepared with 10%
(wlw) soyabean oil and 1% (wlv) lysozyme were analyzed in the sarne range of pH used
to study the MFGM emdsions.
Measurernents were performed in the staûonary layer of a ZET 5 104 cell. The
6-potential was calculated from the electrophoretic mobility of the oil droplets using an
approximation of the Hückel equation (Darling & Dickson, 1979). Buffers at different
pH were prepared from a stock solution of citric acid (O. 1 M) and sodium phosphate
monobasic (0.2 M) and then they were diluted to approximately 0.8 mS of conductivity
to try to maintain constant ionic activity. The conductivity was measured with an Hi
8733 conductivity meter (HANNA Instruments, Fisher laboratory supplies, Unionville,
ON). After dilution the buffer was checked for any change of pH. Samples were
prepared by addition of 6 p l of emulsion to 5 ml of buffer and allowing them to
equilibrate for 5 min. Each sample was analyzed ten Urnes and the average was
calculated. Each emulsion was analyzed at least twice and an average between two
independent experiments was calculated.

Displacement studies were carried out on emuisions made with 1% (w/v) MFGM
isolate and 10% (wlw) soyabean oil. To aliquots (6 ml) of emulsions, 2% (wlv)
detergent was added. Tween 20, Tween 80 and Tnton X- 100 were used in this study
because of the different HLB number (9.5 for Tnton X- 100 and 16 and 20 for Tween 80
and Tween 20 respectively). The protein exchange occurrhg between molecules in the
soluble phase and those at the interface was aiso invesugated by adding to the soyabean
oil emulsion, sodium caseinate (0.5 and 1 % wlv) and ~lactoglobulin ( 1 % w/v). The
ueated emulsions were s h e d at room temperature (22'C) for 3 hours, and the size
disuibution of the emulsion droplets was then measured. Immediately after incubation,
the ernulsions were separated by centrifügation (Beckman) to detemine, by SDS-PAGE,
the effect of the addition of the surfactant on the protein composition of the oil/water
interface. The emulsions were kept at room temperature throughout the experiment.
6 3 RESULTS
6.3.1 Protein Composition of the Isolates
After separation of the soluble phase of buttermilk by centrifugation, the
sedimented pellet had an orange-brown colour, as described by Mather & Keenan
(1975). An analysis of the composition of the protein isolates obtained was carried out
by SDS-PAGE electrophoresis (Figure 6.1). When buttermik was centrifuged in the
absence of sodium citrate (buttermilk isolate, lane 1) the resulûng pellet had a protein
composition very different from the isolate produced with the addition of sodium citrate
(MFGM isolate, lane 2). Caseins (indicaced as the duster of bands shown as IV) were
present in large amounts in the buttermilk isoiate (iane 1). These results confirm those

reported in chapter 5 on isolates obtained from buttermilk of indusuial ongin. The
necessity of a method for separating membrane proteins directly from cream, without
perforrning abundant washes of the fat globules prior to the chuming process has been
reported previously. In fact, washing the fat globules led to changes in protein yields
and distributions depending on the extent of the pretreatment (Mather et al., 1977;
Keenan & Dylewski, 1995). By addition of sodium citrate to buttermilk it is possible to
obiain an isolate composed rnainly of MFGMderived proteins.
In both isolates whey proteins were absent; both ~lactoglobulin and a-
lactalbumin remained in the soluble phase after centrifugation of buttemilk. This was
different from what was found in protein isolates prepared in the same fashion. but from
industrial buttemulk (as described in chapter 3, where whey proteins are present in
considerable amounts in both isolates since the cream may undergo severe heat treatment
(chapter 4). Moreover, in emulsions prepared with MFGM isolates from industrial
buttermilk, a large amount of material is necessary to obtain stable emulsions. It has been
hypothesized that the heat treatrnent has profound effects on the emulsming properties of
the MFGM (chapter 5). The absence of whey proteins in the MFGM isolates from raw
cream demonstrated that the association of a-lactalbumin and p-lactoglobulin with the
membrane material was caused by heat treatment and not by the chuming of crearn. The
protein composition of the MFGM isolate shown in lane 2 agreed with that reported by
other authors (Oehlmann et al., 1994; Innocente et al., 1997). In lane 2, three main
bands were identifed as characteris tic of MFGM-denved pro teins: xanthine oxidase (1).
butyrophylin (II) and PAS-6 and PAS-7 proteins (III). These proteins have been
characterized by other authors (Kim et al., 1992; Jack & Mather, 1990; Keenan &
Dylewski, 1995).

Figure 6.1: SDS-PAGE electrophoresis (left) of protein isolates prepared by ultracentrifugation of buttermilk with no addition of sodium citrate (Lane 1) and with addition of 2% sodium citrate (Lane 2). The migration profile is also shown (rïght) as
detemined by scanning densitometry. Peaks are numbered following the direction of migration. The caseins are taken together as group N.

63.2 Particle Size Distribution and Stability of the Ernulsions
Figure 6.2 illustrates the particle size distribution of emulsions prepared with the
isolates obtained from fresh raw cream. There was no significant difference in the size
distribution of the oil droplets between emulsions made with 1% (w/v) buttemilk isoiate
and 1 or 2% (wlv) of MFGM isolate. This was in strong disagreement with what was
observed for emulsions prepared with buttermilk and MFGM isolates from industrial
buttermilk (chapter 5) , where higher amounts of MFGM isolate were needed to obtain
emulsions of size distributions sirnilar to those of the b u t t e r d isolates shown here.
These emulsions were stable over time and thek monomodal size distributions had an
average diameter of 0.35 Pm. MFGM isolates seemed, therefore, to be able to fom
emulsions with a smali average droplet size as already reported for the weU charactenzed
caseins (Fang & Dalgleish, 1993a). As shown by SDS-PAGE, the protein composition of
the buttermilk isolate was very different from that of the MFGM isolate (Figure 6.1). ln
the emulsion prepared with 1 % (wlv) buttermilk isolate more than 60% of the total
protein consisted of miceilar caseins, which are known to have very good emulsifying
properties. The sirnilar droplet size distribution between casein-containing ernulsions and
MFGM emulsions confmed reports by other authors, that MFGM material has good
ernulsifying properties, as might be expected from its origin on the surface of the fat
globules in milk (Kanno et al., 1991: Chazelas etal., 1995).
Figure 6.3 illustrates the effect of protein and oil concentration on the droplet size
of emulsions prepared with the MFGM isolates. With 10% (wfw) soyabean oil, 1%
(w/v) MFGM isolate was needed to Iower the droplet diarneter and produce a monomodal
distribution of droplet sizes. At low protein concentrations, during the homogenization
process the droplet size of the emulsion varied inversely with the amounts of surfactant
present, as already shown for other m i k protein emulsions (Dalgleish, 1995). At low

0.01 0.1 1 10 100
Average Diameter k m ]
Figure 6.2: Typical particle size distribution of 10% (wlw) soyabean oïl-in-water
emulsions measured by integrated iight scattering. Emulsions were prepared with
(dotied line) 1% (wlv) bunermiik isolate; (solid h e ) 1% (w/v) MFGM isolate; (broken
îine) 2% (wlv) MFGM isolate.

- 0 0.5 1 1.5 2 2.5 3 3.5
MFGM lsolate [%]
Figure 6.3: Average droplet size (d3,2) of (m) 5% and (a) 10% (w/w) oil-in-water
emulsions as a function of the MFGM concentration used during homogenization,
measured by integrated light scattering. Points shown are the average obtained from
measurements of at least two independent experiments.

concentrations, there was not enough MFGM material to cover the interface and the
emulsions contained large oil droplets as show by their bimodal size disuibution.
Figure 6.4 illustrates the oil droplet size distributions of 10% (w/w) soyabean oil
emulsions prepared with low concentrations of MFGM (c 1 % wlv). At low protein
concentration, insufficient coverage of the droplet surface resulted in a broad and
bimodal population of particles with diameter larger than 10 Pm.
Changes in the emulsion droplet distribution as a function of tirne of storage. heat
treatment and pH were also investigated (Figure 6.5). After storage for 1 O days, the
particle size distribution of an emulsion prepared with L0% (w/w) oil and L% (wh)
MFGM did not change from that of the freshly measured emulsion (Figure 6.5A).
It is known by the amino acid composition of MFGM proteins that their rnolecular
structures include a large number of cysteine residues (Keenan & Dylewski, 1995).
However, heating the emulsions prepared with MFGM proteins did not cause any
changes in their droplet size. The droplet diameter of the emulsions did not change after
heating at 90°C for 5 min (Figure 6.5b). These results were a clear indication that heat
induced protein-protein interactions and bridging did not occur at this heating
temperature. If this behaviour is compared with that of other dairy emulsions, casein
emulsions, for example, do not show heat coagulation; caseins do not have a specific
transition temperature and they are considered heat stable molecules. They are
characterized by a very flexible structure, and only K- and ~ 1 - casein contain cysteine
residues. On the other hand, emulsions stabilized by cysteine-containhg whey proteins
fonn gels when heated (Masson & Jost, 1986). The lack of reaction of MFGM proteins
to heat (90°C for 5 min) when adsorbed at an oiVwater interface was in contrast with the
observed low temperature (4O'C) necessary to induce reactions with membrane proteins
on native fat globules in milk (Corredig & Dalgleish, 1996b). It could be hypothesized

0.01 0.1 1 10 100
Diameter Size [pm]
Figure 6.4: Typical particle size distribution of 10% (wfw) soyabean oil-in-water
emulsions measured by integrated light scattering. Emulsions were prepared with 0.25%
(dotted iine), 0.5% (broken line), and 1% (solid line) MFGM isolate.

0.01 o. 1 1 10 100 Diameter Size [pm]
Figure 6.5: Size distribution of 10% (w/w) soyabean oil-in-water emulsion droplets. (A)
emulsion prepared with 1% MFGM isolate (solid h e ) and same ernulsion after 10 days of storage at 4'C (broken line); (B) emulsion prepared with 1 % MFGM (solid line) after
heating at 90°C for 5 min; (C) emulsion prepared with 1% MFGM at pH 6 (broken line),
pH 5 (dotted h e ) and pH 4 (solid line).

that in this latter case, whey proteins are the molecules initiating the heat-induced
interactions in these s ystems.
Figure 6.5C illustrates the droplet size distribution of an ernulsion prepared with
1% MFGM and 10% soyabean oil as a function of changes in pH. In general, the
average size of the ernulsion droplets was increased by decreasing the pH. When pH
was lowered to 6.0 a small population of large droplets formed, indicating destabilization
of the emulsion. At lower pH the emulsion destabilized extensively. Small droplets
were absent, substituted by a population of very large particles (i-e. diarneter > than 10
pm). The destabilization was not reversible. Protein analysis b y SDS-PAGE showed
that the coverage of the different MFGM proteins at the oiUwater interface was not
affected by pH changes.
In conclusion, MFGM protein emulsions seemed to have a stability behaviour
similar to that of emulsions made with whole caseinate. However, cornparhg the two
protein emulsions, more material (about 5 tirnes more) was needed to cover the oiVwater
interface for MFGM proteins than for casein emulsions. This observation was in
agreement with the higher surface load reported by Chazelas et al. (1995) for adsorbed
MFGM (about 7 mglrn-2) compared to adsorbed sodium caseinate (about 2 mglm-2).
63.3 Determination of c-Potential
The observed increase in particle size as pH decreased occurred at the same time
as changes in c-potential, Le., the apparent charge at the surface of shear, detemined in
emulsions prepared with 10% (w/w) soyabean oil and MFGM isolate (Figure 6.6). In
MFGM emulsions, lowering the pH from 7 to 2.5 increased the <-potential of the
emulsion droplets from -25 mV, through O mV at pH around 4, to +25 mV. The
reduction of charge on the droplet surface with decreasing pH wilI diminish the repulsive
elecirostatic interaction between droplets, resulting in their aggregation (Figure 6.5C and

Figure 6.6: 6-potential [mV] of emulsions prepared with 1 % (H) , 1.5% (a) MFGM,
and 1% lysozyme (0) and 10% (wlw) soyabean oil as a function of pH. Points are the
average of two independent experiments; standard deviation were less than 1.6 mV in the
MFGM emulsion and 3 mV in the lysozyme emulsion.

6.6). No difference was shown between the emulsions prepared with 1 % and L .5%
MFGM isolate. It has k e n reported that MFGM proteins in isolation have isoelectric
points ranging from pH 3.5 to 8.0. (Mather et al., 1980: Kim et al., 1992), but we found
that the major@ of charge occurred below a pH of 5.5. A similar pattern has been
observed for emulsions stabilized by p-lactoglobulin or caseinate (Agboola, 1996).
To compare the c-potentials of MFGM emulsions with those prepared using a
protein with high isoelectric point, the changes in <-potential of an oil-in-water emulsion
stabiiized by 1 % (wlv) lysozyme were also observed (Figure 6.6). This protein is
characterized by a basic isoelectric point (pH 1 1.5). Lysozyme adsorbs to an oiVwater
interface in a monomeric fonn rnaintaining its compact structure (Norde & Favier, 1992:
Corredig & Dalgleish, 1995). In the range of pH analyzed (pH 4 to 8) the 5-potential of
the lysozyme emulsion was close to zero. The interface of emulsion droplets stabilized
by lysozyme, a single protein with an isoelectric point of pH 1 1.5, had a different charge
distribution from that of the interface of MFGM stabilized emulsions. The differences in
c-potential between lysozyme and MFGM emulsions, together with the similarities
between the changes in &-potential between casein and MFGM emulsions, suggested an
isoelectric point for the droplet surface between pH 5 and 4. These results were in
agreement with the hypothesis drawn by Kanno & Kim (1990) of an isoelectric point for
the whole membrane of about 4.8. These authors reported that this pH seems to be
optimal for MFGM separation by acid precipitation. Furthemore. butyrophilin. one of
the most abundant MFGM proteins, is characterized by an isoelectric point of 4.96 (Jack
& Mather. 1990). This membrane-spanning protein might play a role in determinhg the
isoelectric point of the whole membrane. MFGM proteins might adsorb at the interface
no t as individual proteins, but maintainhg at leas t in part their "membrane-like"
structure, with strong interactions occuning between the adsorbed MFGM proteins.

63.4 Exchange studios
It has been established that proteins and small molecule emulsifiers compete for
the interface of oil-in-water emulsions (Courthaudon et al ., 199 La; Dalgleish, 1995). It
has also been reported that surfactants may enhance cornpetition between protein
molecules if more than one species is present (Dalgleish. 1995). Moreover, recent
results have shown that the presence of MFGM-derived material seems to affect the
adsorption of caseins at interfaces (Chazelas et al., 1995). To provide more information
on the adsorption behaviour of MFGM proteins, emulsions prepared with 10% (wlw)
soyabean oil and 1% (wlv) MFGM isolates were analyzed d e r the addition of other
surface active molecules (small molecule surfactants and skim rnilk proteins). Three
small molecule surfactants were used in this study, Tween 20 and Tween 80, which are
known to be effective in the destabilization of d a j r emulsions (Goff & Jordan, 1989),
and Triton X- 100, often used in the solubilization of MFGM proteins from the native
membrane (Mather et al., 1980). SDS-PAGE electrophoresis was perforrned on the oil
droplets and the soluble phase after centrifugation of the emulsions with added surfactant.
Small molecule surfactants did not effectively displace MFGM proteins from the
droplet interface. Figure 6.7 illustrates the electrophoretic profile of the proteins present
in the original emulsion, in the oil droplets and in the soluble phases after treaunent with
surfactant. The newly formed membrane on the emulsion droplets was not affected by
the addition of 2% (wlv) Tween 20, Tween 80 or Triton X- 100. The small amount of
protein present in the serum phase suggested that the MFGM protein adsorbed remained
at the interface after treatment with surfactant. This behaviour was different from that
observed for casein emulsions. It has previously been reported by Dalgleish et al. (1995)
that the properties of the surface of casein-stabilized emulsions change with the addition
of surfactant. The behaviour of MFGM ernulsions was also different from that reported
by Courthaudon et al. (1991 b) for P-lactoglobulin emulsions: the presence of Tween 20

Migration
; A-, -.a
Figure 6.7: SDS-PAGE of a 10% oil-in-water emulsion prepared with 1% (wlv) of
MFGM isolate. Lane 1: protein in the original emulsion; Lane 2: oil droplets of the
emulsion after treatment with Tween 20 (resuspended to 10% wlv); Lane 3: soluble
phase of the emulsion after treatment with Tween 20; Lane 4: oil droplets of the
emulsion after treatment with Triton X- 100; Lane 5: oil droplets of the emulsion after
treatment with Tween 80; Lane 6: soluble phase of the emulsion after treatment with
Tween 20; Lane 7: soluble phase of the emulsion after treatment with Tween 80; Lane
8: soluble phase of the emulsion after marnent with Triton X- 100.

leads to a complete displacement of P-lactoglobuh h m the oiVwater interface without
any loss in stability. To confinu our observations, the same experiments were carried out
on casein emulsions. Figure 6.8 illustrates a SDS-PAGE gel of caseinate emulsions
(0.5% (w/v) caseinate, 10% (w/w) soyabean oil) after addition of 2% (w1v)Tween 20,
Tween 80 and Triton X- 100, under the same conditions described for the MFGM
emulsions. After treatment with surfactant, the oil droplet surface did not contain any
more caseins. Thus, MFGM proteins, because of their membrane origin, appear to
adsorb with a mechanism different from that of the other milk proteins. It is known that
for emulsion droplets stabilized by ~lactoglobulin the amount of emulsifier required for
complete displacement increases with the age of the adsorbed layer. It is possible that
intermolecular interactions might characterize the structure of the MFGM protein-
stabilized interface, which would cause the observed resistance to displacement by srnall
molecule surfactants. In addition, we know that the MFGM material adsorbed at the
interface contaùis phospholipids, which might contribute to lowering the interfacial
tension so that exchange with detergent becomes less favorable. Indeed, Chazelas et al.
( 1995) have reported that MFGM-stabilized in terfaces have a lower interfaciai tension
than those stabilized by caseinate.
Exchange studies were carried out by adding whole casein or P-lactoglobulin after
a 1% MFGM emulsion was prepared. The proteins present in the serum phase after
mwng the emulsion are shown in figure 6.9. None of the added proteins was present in
the oil droplets after mixing the ernulsion (not shown) and die serum phases did not
contain any MFGM proteins. Furthemore, in the semm phase ail of the caseins and P- lactoglobulin originally added to the samples were present. The oiVwater interface
stabilized by MFGM proteins was afTected by neither the flexible structure of caseins nor
the small globular structure of ~lactoglobulin. As already shown by the smaU molecule
surfactant experimenu, the MFGM material adsorbed did not aUow further adsorption

Migration
Figure 6.8: SDS-PAGE of a 10% (wfw) oil-in-water emulsion prepared with 0.54% (wfv)
casein. Lane 1: protein in the original emulsion; Lane 2: oil droplets of the original
emulsion resuspended to a final concentration of 10% oil; Lane 3: oil droplets of the
emulsion after treatment with Tween 20 (20% Final concentration): Lane 4: oil droplets
of the emulsion after treatment with Tween 80 (20% finai concentration); Lane 5: oil
droplets of the emulsion after treatment with Triton X-100 (20% final concentration);
Lane 6: soluble phase of the emulsion after treaunent with Tween 20: Lane 7: soluble
phase of the emulsion after treatrnent with Tween 80; Lane 8: soluble phase of the
emulsion after treatment with Triton X- 100.

Figure 6.9: SDS-PAGE gel of protein adsorbed on a MFGM-stabiiized interface as a
function of other proteins added to the emulsion. Lane 1 : original emulsion ( 10%
soyabean oil and 1% MFGM isolate); Lane 2: original emulsion with 0.5% casein
added; Lane 3: original emulsion with 1% casein added; Lanes 4,6: soluble phase of
the emulsion with 0.5% casein added; Lane 5: soluble phase of the emulsion with 1%
casein added; Lane 7: original emulsion with 1% p-lactoglobulin added; Lane 8:
soluble phase of the emulsion with 1% B-lactoglobulin added.

and/or dis placement of pro tein. These results differentiated the behaviour of MFGM -
stabilized emulsions from that of the other skim milk protein emulsions.
6.4 DISCUSSION
The stabiiity of emulsions prepared with MFGM isolate had many sirnilarities
with that reported for caseinate emulsions. The emulsions were stable at pH 7 and
underwent aggregation at acidic pH. These results provided more evidence supporting
the hypothesis that the isoelectric point of the whole MFGM is about pH 4.5. in spite of
the different isoelectric point found for MFGM proteins when analyzed as single
monomers. Furthermore, the droplet sizes in emulsions prepared with MFGM were not
affected by heating at 90'C for 5 min, demonstrating the stability of this newly formed
oiYwater interface. It could be concluded that the proteins adsorbed at the interface were
characterized by strong intermolecular interactions. The behaviour of emulsions
stabilized by MFGM material from untreated cream was quite different from that
observed (chapter 5) in emulsions stabilized by MFGM isolated from industrial
buttermilk. MFGM from industrial buttermilk contained large arnounts of whe y protein,
as a result of the heat treatrnent, and showed poor emulsifying behaviour. On the
contrary, in untreated MFGM emulsions, less matenal was needed to achieve emulsion
stability. Furthermore, the newly formed membrane on the oil droplet was stable and less
susceptible to changes at the interface, such as protein rearrangements, which are
common reactions occurring on surfaces formed with skim milk proteins (Dalgleish,
1995). In emulsions prepared with MFGM, the absence of displacement with the
addition of surfactant, or of exchange with other proteins could be taken as evidence of a
different mechanism of adsorption at the oiUwater interface. We may conclude that the
phospholipids present in the MFGM material adsorbed at the interface lowered the
interfacial tension causing great resistance to further exchange of surfactant

These results lead to important considerations on the application of these protein
isolates to real food systems. It is weli known that surfactants weaken the mechanical
propenies of the interface and lead to destabilization (Goff & Jordan, 1989). The
difference in ernulsifjing properties between MFGM isolates from unheated and heated
cream (commercial buttermilk) raised some questions on the role played by buttermilk
during partial coalescence in ice cream. The accepted fact, never questioned, that
buttermik added to ice cream mixes reduces the need for other emulsifiers has to be
revised in light of these resuits. The buttermilk powder used in the production of ice
cream has certainly been subjected to high temperature pasteurization. The use of this
byproduct as functional ingredient in ice cream mixes is questionable since its
functionality seems strongly dependent on its themal history. Perhaps, the use of low-
heat buttermilk powder would affect partial coalescence differently than commercial
buttermilk powder. A comparison between emuisions prepared with low-heat buttemiilk,
Iow-heat skim miik and commercial buttermik, in ice cream mixes, might give us the
necessary information on the real role played by MFGM during partial coalescence.

CHAPTER 7*
THE EFFECT OF HEATING OF CREAM ON THE FUNCTIONAL
PROPERTEES OF MILK FAT GLOBULE MEMBRANE ISOLATES
7.1 INTRODUCTION
In whole milk, fat globules are surrounded by a membrane, originaily derived from
the apical plasma membrane of the milk fat globule (McPherson & Kitchen, 1983). This
miik fat globule membrane W G M ) is a complex mixture of proteins and phospholipids
and acts as a natural emulsifier, preventing codescence of the fat globules. In the
conventional process of buttermaking, after pasteurization, destabilization of the fat
globules occurs in cream during churning, and a water phase (buttermilk) is released. The
protein content of buttermilk resembles that of skim milk (around 3% protein). However,
buttermiIk contains not only skim milk proteins (caseins and whey proteins) but also a large
fraction of MFGM material.
MFGM proteins represent ody a smail portion of the total m i k proteins but they are
fundamental for the stability of the fat globules (McPherson & Kitchen, 1983; Keenan &
Dylewski, 1995). The major proteins present in the MFGM have been isolated and studied
(Greenwalt & Mather, 1985; Mather & Jack, 1993; Berglund et al., 1996). In a recent
review, Keenan & Dylewski (1995) described the diffîculty inherent in the isolation of the
individual proteins in the MFGM, because of the strong association between butyrophilin
(67000 Da) , xanthine oxidase (about 150000 Da) and other minor MFGM proteins.
Most MFGM proteins are characterized by high hydrophobicity and are covalently bound to
fatty acids and to carbohydraie residues (Shimizu et al., 1976; Mather & Jack, 1993).
Because of their origin and amphiphilic nature, MFGM proteins are expected to be
good emulsifying agents. Recently, the functionality of MFGM proteins in emulsions has
*Submmrtted for publication essentially in this fonn to the Journal of Agricultural and Food Chemisby

been investigated by homogenizing milk fat in the presence of MFGM (Kanno et al., 199 1;
Oehlmann et al., 1994). These studies did not clarify which components of the MFGM
play a role in the stabilization of the reconstituted fat globules. These emulsions were also
prepared with MFGM from non pasteurized fresh crearn (Kanno et al., 199 1; Oehhann et
al., 1994).
In iight of a possible utilization of these fractions from buttermik as a functional
ingreclient in foods. an understanding of the effect of heating of the cream on the MFGM is
needed, since in the manufacturing process of buttermaking an initial heat treatment of the
cream is generally employed. It has been s h o w that during heating of whole millc, even at
low temperatures (Le. 65'C) strong interactions occur between a-lactalbumin, &
lactoglobulin and MFGM (Corredig & Dalgleish, 1996b). These interactions result in a
change in the electrophoretic mobility with very liale change in the diarneter of the fat
globules (Dalgleish & Banks, 199 1). A new membrane, more resistant to coalescence,
develops around the fat globules (Dalgleish & Banks, 199 1 ; van Boekel & Folkerts. 199 1 ).
The precise nature of these interactions is not yet clear; various hypotheses have b e n
formuIated (Dalgleish & Banks, 199 1 ; van Boekel & Folkerts, 199 1 ), that: (i), skim
milk proteins rnight displace the original membrane material; (ii), proteins may fill in gaps
in the membrane surface, which become exposed after heating; or (iii) skirn milk
proteins rnay bind to the MFGM proteins via disulphide bridging andor hydrophobic
interactions. The latter mechanism agrees with the results of Kim and Jirnenez-Flores
(1995); the extent of reaction of whey proteins with the MFGM proteins is strongly
dependent on temperature.
Recently, we have studied the properties of MFGM material from indusaial
butter& (chapter 5). Tfiese MFGM isolates contained considerable amounts of
associated whey proteins and their emuls@ing properties were found to be poor compared
to those of MFGM material isolated from unpasteurized crearn (chapter 6). The observed
difference in functionality between the two MFGM isolates led to the present study, in

which we investigated the changes occurring in the MFGM when cream is heated. Protein
solubility, protein adsorption and emulseing properties of the MFGM isolates as
tiinctions of temperature were determined. The cream used was heated by either industtial
pasteurization m g h temperature short time (HTST)) or by batch heating at controlled
temperatures (60' to 8S°C).
7.2 MATERIALS AND METHODS
7.2.1 Heat Treatment of the Cream
Cream (about 40% w/w fat) was obtained from a local daiq after separation from
fresh skirn millc. The centrifuga1 separation was performed at 50°C so that our "untreated
cream" had actualiy been heated at this temperature. Cream was f d e r heat treated in the
laboratory by batch heating for 10 min at temperatures between 60" and 85'C in a water
bath in 250 ml aliquots. Sarnples were heated in a water bath at 95'C to reach the required
temperature in less than 1 min. Cream was ais0 collected at the dairy plant after different
HTST pasteurization steps (76-8S°C for 16 s). After heating, the crearns were cooled to
roorn temperature in an ice bath and then, after addition of sodium rtzide (0.02%), were
kept refngerated (4'C) ovemight Every heating experiment was repeated at lest twice with
indipendent batches of crearn.
7.2 .2 Production of Butterrnilk
To assess the effec t of heating temperature on the functionality of the MFGM
fractions present in buttermilk, cream destabilization was achieved by churning in a small
scale chum (1 Litre capacity), as already descnbed in 6.2.1. The samples were chumed at
10'C until phase separation occurred. Buaemilk was also collected at the dairy plant after

~hunùng of pastedzed creams (85°C for 16 s) and used for gel permeauon
chromatography experiments.
7.2.3 Preparation of the Isolates
Butteimilk was released from the butter granules and fdtered through a cheesecloth
and then through a qualitative P8 füter (Fisher Sci.). The MFGM isoiates were prepared as
described in paragraph 5.2.1. After addition of sodium citrate (2% wlv), buttermillc was
kept at refngeration temperature (4'C) overnight The samples were then centrifuged at
15'C for 50 min at 100,000 g in a preparative ultracentrifuge (Bechan). Buttermilk
isolates were ais0 prepared as control samples by centrifugation of buttermilk without
addition of sodium citrate. The sedirnented pellets of the buttermilk and MFGM isolates
were drained on filter paper (Whatman no. 4, Fisher Sci.), resuspended in MiiIiQ water
using a tissue homogenizer (Polytron PT 2000) and then freeze dried. The freeze dried
samples were stored at -20°C. The isolates contained MFGM material consisting of a
mixture of phospholipid and protein. The protein concentration of die freeze ciried MFGM
isolates was about 60% w/w.
7.2.4 Emulsion Preparation
Aliquots of freeze dried isolates were resuspended in buffer (20 m M TrisMC1) at
pH 7. Soyabean oil(lO% wlv) was added and the mixture was homogenized using a
microfluidizer as described in detail in paragraph 4.2.2.

7.2.5 SDS-PAGE
Sample preparation for electrophoresis was carried out as described in detail
elsewhere (paragraphs 4.2.4, 5.2.6, 6.2.4).
The samples were loaded ont0 a 20% acrylarnide homogeneous gel (Pharmacia
Biotech) and run in a rapid electrophoresis system (Phmacia Biotech). The protein
bands were fixed and stained using a solution of Coomassie blue (1% wfv) in methanol-
acetic acid-water. Destaining was achieved by washing the gel in the same mixture of
methanol-acetic acid-water. The dned gels were scanned with a Sharp M330 scanner.
The images were analyzed with Imagemaster software (Phamacia Biotech) for quantitative
analysis. The amounts of a-lacialbumin and ~lactoglobulin were quantified as percentages
of the total protein present in the sample.
7.2 .6 Fat Globule Size Distribution
The fat globule size distribution of the emuisions was determined by integnted light
scattering as described in 4.2.3. The droplet size distribution and the average diameter,
d3,2 [ Lnd3End21, of the emulsions were us& to characterize the emulsifying behaviour of
the different isolates.
7.2.7 Size Exclusion Chromatography
To obtain information on the aggregation of the MFGM in buttermilk from heated
cream, size exclusion chromatography was performed (Superose 12, Pharmacia Biotech)
on samples of industrial buttermilk. The separation range of Superose 12, as declared by
the manufacturer, is between 1 and 300 kDa Before analyses. samples were fütered
through a 0.45 pm füter (Millipore, Fisher Sci.). Two different elution buffers (pH 7.0)

were use& (i), a non-dissociating bufTer containing 20 mM irnidazole, 5 m M CaC12 and
50 m M NaCl; (ii),. adissociating buffer containing 6 M ureaand 20 m M irnidazole. A
Pharmacia optical unit UV- 1 (Phmacia Biotech) at 280 nm was employed to follow the
elution peaks. To detennine which aggregates were mauily held together by disulphide
bonds, 5 rnM Zmercaptoethanol was added pnor to chromatography. The elution patterns
of reduced samples were compared with those of the same samples, eluted in the absence
of Zmercaptoethanol. Experiments were perfomed at least in duplicate. After collection of
the fractions separated by chromatography, SDS-PAGE was cmied out to determine the
protein composition of the eluted peaks. To 0.5 ml of sample, SDS (200 pl), 2-
rnercaptoethanol(50 pl ) and bromophenol blue (100 pl) wexe added.
7.2.8 SolubiIity Studies
Freeze dned protein isolates (-60% wlw protein) obtained from differently heated
crearns were resuspended (0.8% wlv) in one of three buffers: (i), 20 m M Tris. pH 7.0
with or without added Triton X-100 (1 % w/v); (ii), LOO m M Na2C03, pH 1 1.5: or. (iii),
20 mM Tris 4 M Urea, pH 7.0. The non-ionic detergent Triton X-100 and the carbonate
buffer at high pH were chosen because of their common use in removing proteins from
membrane material Cyanagita & Kagawa, 1986; Fujiki et al., 1982). After continuous
stirring for 3 h, the samples were centrifuged at lOOOOg for 30 min. Protein analysis on
the soluble phase was carrieci out by a Lowry procedure rnodified b y Markwell et al.
(1978) for membrane protein and lipoproteh preparations. Bovine Serurn Albumin (Sigma
Chem., St Louis, MO) was used as the protein standard. The protein composition of the
soluble fractions was also determined by SDS-PAGE. The electrophoresis was performed
as descnbed above, with aliquots (200 pI) of soluble phase.

7.2 .9 Iron Determination
The amount of Fe contained in the MFGM isolates denved from crearn heated at
various t e m p e m m was quantifed by flame absorption spectrometry. Approximately
0.25 g of freeze dried MFGM isolate was digested with nitric acid, and analyzed with a
Perkin Elmer 5 100 ZL atomic absorption spectrometer. The analyses were performed by
the Laboratory Services Division of the University of Guelph (Guelph, Ontario).
7.2.10 Statistical Analysis
Analysis of variance of the effect of heating temperature on the arnount of whey
protein associated with the MFGM fraction, the average diameter of the emulsions prepared
and the solubility of the MFGM isolates was performed (SASBTAT software, SAS
Institute, NC). Results, average of at l e s t three independent expenments, were
considered significant for p<0.05.
7 . 3 RESULTS
7.3 .1 Studies on Industrial Cream and Buttermilk
7.3.1.1 Emulsifying Properties of Isolates frorn Buttermilk prepared with
Cream Heated by HTST (76-85'C)
Two different isolates were used to prepare oil-in-water emulsions: buttermilk
isolates and MFGM isolates. Butterrnilk isolates contained considerable amounts of skim
mik proteins, as they were prepared simply by cennifuging buttermilk. Figure 7.1
illustrates the droplet size distribution of 10% (w/v) soyabean oil-in-water emulsions

prepared with buuermilk isolates originating from creams pasteuriled by HTST at different
temperatUres. The diameter distribution of emulsions prepared with 2% (w/v) buttermilk
isolate from unpasteurized cream was not m e r e n t from that of emulsions prepared with
the same arnounts of butterniilk isolates from crems heated at 76", 78" or 85°C. Caseins,
present in the buttexmilk isolates, were the main proteins adsorbed to the oil-water
interface, and determined the behaviour of the emulsions. These results were in agreement
with those previously reported for isolates denved from industrial buttermilk (chapter 5 ) .
To study the effects of temperature of HTST pasteurization of the cream on the
emulsifjhg properties of the MFGM material, we determined the droplet size
distributions in emdsions prepared with MFGM isolats. These isolates contained a much
higher concentration of MFGM material than buttennilk isolates, and much s m d e r
amounts of skim milk proteins. The droplet size distributions of MFGM isolate
emdsions contahing 10% (w/w) soyabean oil are shown in figure 7.2. AU of the MFGM
isolates from creams heated by HTST at temperatures between 76' and 85°C showed
emulsifying properties which were much worse than those of MFGM isolates from
unpasteurized cream. Emulsions prepared with 1% (wiv) of MFGM isolate from unheated
cream had a narrow distribution of droplet sizes, with an average diarneter of about 0.4
p. AU of the emulsions made from the MFGM of HTST creams contained a large
nurnkr of droplets with diameter > 10 pm and relatively smail arnounts with diarneter c 1
pm (Figure 7.2).
When resuspended in Trislurea buffer, the MFGM isolates from HTST cream-
buttermilk were not soluble. In addition, SDS-PAGE indicated large amounts of whey
protein in these isolates. Figure 7.3 illustrates the electrophoretic migration of MFGM
isolates derived from cream heated at 76" and 78°C and also from industrial buttermilk. In
the industrial buttermillc, cream destabilization occurred in a continuous churn after
pasteurization at 85'C for 16 S. The presence of whey protein in the h4FGM of ail of these

0.01 o. 1 1 10 100
Average Diameter [pm]
Figure 7.1: Typical droplet size distribution of 10% (wfv) soyabean oil-in-water
emulsions measured by integrated light scattering. Ernulsions prepared with 2% (wlv)
buttermilk isolate denved from (a) untreated cream; (b-d) cream heated by HTST for 16s
at temperatures of (b) 76'C; (c) 78'C; (d) 85'C.

0.01 O. 1 1 10 1 O0
Average Diameter [ lm]
Figure 7.2: Typical droplet size distribution of 10% (wlw) soyabean oil-in-water
emulsions measured by integrated light s c a t t e ~ g . Emuisions prepared with 1 % (wlv) of
MFGM isolate from buttermilk denved from (a) untreated crearn; (b-e) cream heated b y
HTST for 16 s at temperatures of (b) 76'C; (c) 78'C; (d) 80°C; (e) 85'C.

A
Migration
1 Caseins
Figure 73: SDS-PAGE of MFGM and buttermilk isolates prepared by ultracentrifugation of buttermilk from HTST crearns. Lane 1: MFGM isolate from cream
heated at 78°C for 16 s; Lane 2: MFGM isolate from cream heated at 76°C for 16 s; Lane 3: MFGM isolate denved from industrial b u t t e d ; Lane 4: buttemdk isolate denved from unheated cream; Lane 5: buttermiik isolate denved from cream heated at 78'C for 16
s; Lane 6: buttermilk isolate derived from cream heated at 76'C for 16 s; Lane 7: buaermilk isolate derived from industriai buttermillc; Lane 8: MFGM isolate derived from
unheated crearn.

fractions was caused b y kat-induced interactions between a-lactaibumin, fblactog 10 bulin
and the membrane of the fat globules ofcurring in cream during heating (Dalgleish &
Banks, 1991; Corredig & Dalgleish, 1996b). Figure 7.3 aiso shows the electrophoretic
separation of butîemiillc isolates. Whey proteins were absent only in the isolates (both
buttennillc and MFGM) which had been obtained from non-pasteurized cream.
7.3.1.2 Size Exciusion Chromatography
The association of skim milk proteins with the MFGM was investigated by gel
permeation chromatography using two different buffers. Buttermilk was coilected after
industrial chuming of cream pasteurized at 8S°C for 16 S. When these buttermdk samples
were elutcd on a column of Superose 12, most protein was found in the excluded volume
(molecular size > 300 kDa). With Tris buffer, in the absence of 2-mercaptoethanol,
three fractions could be separated one at the exclusion limit of the column, and two others.
Large MFGM aggregates were eluted together with casein micelles in the excluded volume
(fraction 1, Figure 7.4A); however, fraction 1 also contained j3-lactoglobulin in
appreciable amount (about 50% of the total ~lactoglobulin). The remaining (3-lactoglobulin
was isolated in the second chromatographic peak. The third peak eluted from the column
contained only a-lactalburnin, which was absent in the other two peaks. Revious
research on heating of whole milk has shown that both a-lactalbumin and Plactoglobulin
interact with the membranes of fat globules (Dalgleish & Banks, 199 L: Corredig &
Dalgleish, 1996b). In the aggregates smaller than 0.45 p.m present in buttermilk (the
samples were fdtered before chromatography), only f!blactoglobulin was associated with
the MFGM fraction. Therefore, the reactions between proteins in heated cream and whole
milk may differ.
Urea buffer disrupts the interactions which hold together casein miceiles and
aggregates. Chromatography of the buttemilk samples in urea gave 5 peaks, whose

Figure 7.4: Electrophoretic migration of the protein fractions eluted by gel permeation chromatography. A) Industrial buttermilk eluted with irnidazole buffer; chromatography fractions 1 - 3. B) Chromatography fractions 1-5 from industrial buttermilk eluted with irnidazoldurea buffer, without (lanes 1-5) and with 5 mM 2-mercaptoethanol (lanes 6-10). SDS-PAGE was performed in reducing conditions.

protein composition is shown in figure 7.4B. The fraction eluting at the void volume
contained MFGM proteins, iccasein and plactoglobulin. Rogressive elution provided
fractions c o n t a k g less MFGM and more caseins, but also arnounts of ~lactoglobulin and
a-laccilbumin (Figure 7.4B, Lanes 1-4). The fîth peak in the elution pattern contained no
protein. When 2-mercaptoethanol was added to the sample, the ~iactogiobulin and K-
casein were lost from the void-volume peak and appeared at longer elution times, but the
membrane proteins remained in this fraction.
These results suggested that there are interactions be tween cys teine-containing
proteins of milk and those of MFGM. The Plactogiobulin and K-casein in the void volume
fraction must be there because they are linked by disulphide bonds to other proteins, but
these experiments do not permit us to detemine whether, for example, the K-casein is
specifically linked to plactoglobulin, or membrane proteins, or if it is simply
polymenc. Since the MFGM isolate dws not appear to contain K-casein (Figure 7.3), it is
iikely that it is present in the void volume peak because of its polymeric nature (Yaguchi et
al., 1967). At least part of the &lactoglobulin in the void volume peak is, however, bound
to the MFGM.
7.3.2 MFGM Isolates Derived from Cream Heated at Temperatures
(60-85" C )
No differences were observed in the e m u l s ~ g properties of MFGM isolates
from creams heated by HTST at 76' or 85'C. Al1 of the emulsions prepared with these
MFGM isolates showed a bimodal droplet size distribution with a large proportion of
droplets bigger than 10 Fm. On the other hand, the emulsions prepared with MFGM
isolates from buttermilk obtained from non-pasteurized crearn were stable; they were
characterized by a monomodal droplet size distribution with an average diameter of - 0.4
Pm-

Therefore, we studied the effect of heating at lower temperature than those used for
HTST on the properties of the ?.!FGM. Crearn was heated by batch heating for LO min at
tempemaires ranging between 60 and 8S°C. Unpasteurized cream was collectecl after
centrifuga1 separation at 50°C. MFGM isolates were prepared from ail of the heated
creams.
7.3.2.1 Solubiiity Studies
Solubility is an important index for the use of proteins in food systems. MFGM
isolates, obtained from creams heated at different temperatures, were resuspended in
various buffers (0.846 wh), and after centrifugation, the soluble phases were analyzed for
protein concentration as weLl as protein composition (Figure 7.5 and 7.6). When MFGM
isolates were resuspended in Trislurea buffer, the solubility was affected by temperature
of heating of the crearn (Figure 7.5). The three major bands which characterize the MFGM
proteins in SDS-PAGE were ideniifid in samples from untreated creams (heated at W C ,
during centrifuga1 separation at the dairy) and from creams heated at 60°, 62" and 65°C.
MFGM isolates of crearn heated at 70'C (Lane 4) contained in their soluble fraction only
one of these three MFGM protein bands. These residual proteins have been identified as
PAS4 and PAS-7, which are known to be on the surface of the membrane, not tightly
associated, and soluble in urea (Kim et al., 1992). MFGM isolate suspensions derived
from crearn heated at temperatures higher than 70'C were not soluble in 4 M urea buffer.
In addition, no whey protein was present in the soluble material. In the MFGM isolates
derived from cream heated at temperatures > 70°C, there was a large amount of whey
protein, but it was covalently linked to the membrane material and was insoluble.
The solubility of the MFGM fraction was strongly affected by the temperature of
heating of the cream, not only in urea buffer, but also in Tris/Triton X- 100 or sodium

ion
Figure 7.5: SDS-PAGE of the soluble fractions (after centrifugation) of MFGM isolate suspensions (0.8% w/v) in 4 M urea buffer. Lane 1: MFGM isolate from buttermilk
derived from unheated (50°C) cream; Ianes 2-6: MFGM isolates from creams heated at
temperatures of (lane 2), 60°C; (lane 3), 65°C; ( h e 4), at 70°C; (lane S) , 75'C; (lane 6). 80°C. SDS-PAGE was performed in reducing conditions.

40 45 50 55 60 65 70 75 80 85 90
Heating Temperature ['Cl
Figure 7.6: Concentration of soluble protein (% of total isolate weight) after
cenuihigation of suspensions (0.8% w/v) of different MFGM isolates (60% wlw protein),
as a function of temperanire of heating of the cream. (H) MFGM resuspended in 20 m M
Tris buffer, pH 7.0; (0) MFGM resuspended in 20 m M Tris buffer and 1% Triton X-
100, pH 7.0; (a) MFGM resuspended in 0.1 M NatC03 buffer, pH 1 1 S. Results are
the average of three experiments, error bars represent standard deviations.

carbonate buffer (Figure 7.6). The MFGM isolates were not soluble in Tris buffer,
while with the addition of Triton X- 100, their solubility was increased but depended
signifcantly (p<0.05) on the temperature of heating. The amount of protein in the soluble
phase of Tris/Triton X-LOO bufier differed from that present in the soluble phase of Tris
buffer for temperatures c 70'C. This effective solubilization of some MFGM protein with
Triton X- 100 agreed with the resdts reponed by Houlihan et al. ( 1992) for MFGM frorn
pasteurized cream and rnilk. Sodium carbonate (pH 1 1.5) solubilized larger arnounts of
protein than Triton X- 100, in samples of MFGM isolates from cream heated at
temperatures lower than 65'C. In this case also, there was a significant (pd.05) effect of
temperature of heating on the amount of residual soluble protein. However, the protein
solubilized was never > 50% of the total.
7.3.2.2 Emulsifying Properties of the MFGM isolates
Emulsions were prepared with MFGM isolates from buttermik obtained from
differently heated creams. Whiie 1 % (w/v) MFGM from unheated cream had good
emuls@ing ability (Figure 7.2), emulsions with 1% (wlv) MFGM isolates from heated
creams contained large particles, and for crearn heated at 75'-85'C. obvious phase
separation was observed &ter homogenization. The droplet size distribution of 10% (wlw)
oil-in-water emulsions prepared with 2 and 3% (wlv) MFGM isolates is shown in figure
7.7. Emulsions prepared with MFGM isolates from creams heated at 60' and 62'C had
monornodal distributions similar to emulsions prepared with MFGM isolates from unheatcd
crearn. When MFGM isolates obtained from crearns heated at higher temperatures were
used, the emuision droplets increased in size and the nurnber of large droplets seemed to
depend on the heating temperature of the cream. The overail size distribution improved
when emulsions were prepared with larger arnounts of MFGM isolates (3% w/v) (Figure

0.01 0.1 1 10 1 O0
Average Diameter @TI]
Figure 7.7: Typical droplet size distribution of 10% (w/w) soyabean oil-in-water
emulsion measured by integrated light scattering. Emulsions prepared with 2% (A) and 3%
(wfv) (B) MFGM isolate from butîermiik obtained from cream heated for 10 min at (a)
60°C: (b) 65°C; (c) 70°C; (d) 80°C.

7.7B), but even at this concentration a signiricant shift of the average droplet size to a larger
diameter occurred as MFGM from increasingly-heated cream was used.
Analysis of variance showed a significant (p4.05) effect of temperature on the
average diameter of the dropleü for emulsions prepared with 2 1 and 3% (wh) MFGM
isolates. Figure 7.8 shows the increase in d3,z for 2% (wlv) MFGM isolate in 10% (w/w)
soyabean oil emulsions, as a function of the heating temperature. In this case also, as
already shown for the solubility of the different MFGM isolates, up to 65°C there was no
significant difference in the average diameter sue.
7.3.2.3 Whey Proteins Associated with the MFGM
The protein compositions of the MFGM isolates prepared from creams heated at
different temperatures were analyzed by SDS-PAGE. The arnount of whey protein
associated with the MFGM increased with increasing temperature. These results were in
agreement with those reported for MFGM obtained from HTST crearns (Figure 7.3). The
association of ~lactoglobulin increased at temperatures higher than 65'C. SDS-PAGE gels
were analyzed by image analysis and the arnounts of a-lactalbumin and P-lactoglobulin
were quantifïed as percent of the total protein present (Figure 7.9). For temperatures up to
85'C, the amount of a-lactalbumin was not ~ i ~ c a n t l y affected by temperature. On the
other hand, the amount of P-lactoglobuh associated with the MFGM increased
simcantly (pcû.05) with temperature of heating especialiy above 65'C. The presence of
signitiicant amounts of Blactoglobulin and very Little a-lactalbumin in the heated
membranes was in agreement with the findings reported by McPherson et al. (1984) on
pasteurized crearns. Moreover, the higher arnounts of ~lactoglobuiin associated with the
MFGM at high temperature agreed with the ciifferences reported b y Kim and Jimenez-

40 45 50 55 60 65 70 75 80 85 90
Heating Temperature ['Cl
Figure 7.8: Average diameter as determined by integrated Light scattering of oil-in-water
emulsions containing 10% (wlw) soyabean oil and 2% (wlv) MFGM isolate. Values of
d3,2 are plotted versus temperature of heating of the cream. Results are average of three
independent experirnents, error bars represent standard deviations.

40 45 50 55 60 65 70 75 80 85 90
Heating Temperature ['Cl
Figure 7.9: Arnounts of (W) a-lactalburnin and (e) P-lactoglobulin (% of the total
MFGM protein in SDS-PAGE) as a function of temperature of heating of the crearn.
Results are the averages of at least 3 independent experiments. Error bars represent
standard deviations .

Flores (1995) in the binding of ~ l a c t o g l o b u h with proteins from the MFGM at different
temperatureS. A temperature higher than 65°C seemed to be necessary for ~lactoglobulin
to react with the MFGM. Jang and Swaisgood (1990) reponed very linle disulphide
in terchange between ~lactoglobulin and ic-casein in skim mdk during h e a ~ g at
temperatures lower than 75°C. Non covalent protein-protein interactions seem to play an
important role in s k h milk under those conditions. Recently, the formation of disulphide
aggregates at 65°C in Plactoglobulin in solution has k e n reported (Hoffmann and van
Mil, 1997). At the lowest temperature of heating of the cream. the formation of &
Iactoglobulin complexes might occur via both non-covalent interactions and disulphide
interc hange; these reac tions might occur simultaneousl y or sequen tially (Hoffmann and
van Mil, 1997).
7.3.2.4 Iron Content
The arnount of Ùon found in the MFGM isolates decreased with increasing
temperature of heating, from 110 ppm in the MFGM from unheated cream (5OoC), to 20
ppm in MFGM from crearn heated at 80'C (Figure 7.10). A change in the colour of the
MFGM isolates with temperature of heating was also observed, from brown-red in the
unheated isolates to white-yellow in the matenal denved from cream heated at 8S0C. In
contrast with the other observations we made, there did not appear to be a threshold
temperature in the region of 65'C. It was evident that loss of iron was sigmficant even at
62°C.
One of the major proteins in the MFGM (8- 10% of the total protein in the
membrane), xanthine oxidase, is a complex metao-flavoprotein (Mangino & Brunner.
1977). This enzyme is present in a dehydrogenase and an oxidase form. Conversion of
the dehydrogenase to the oxidase form cm be achieved by proteolysis, heating or
incubation with sulphydryl rnodming reagents (Coughian, 1980). The decrease in the

50 62 65 70 75 80 Heating Temperature ['Cl
Figure 7.10: Arnount of iron (ppm) in the MFGM isolates as a function of heating
temperature of the cream. Analyses performed by atomic absorption spectroscopy.

arnount of iron in the MFGM with increased heating temperature of the crearn might be
related to a heat-induced release of metal from xanthine oxidase. In fact, four cysteines
bind the iron-sulphur cluster of xanthine oxidase (Berglund et al., 1996). The structural
change of the protein and the release of iron wodd render the cysteines of the membrane
protein available for disulphide exchange with other cysteine-containing proteins present in
the crearn (Le. P-lactoglobulin, k-ca~ein).
7.4 CONCLUSIONS
Two main effects occurring during heating of fat globules in milk and cream have
been previously reported: the loss of membrane-constituent material and the formation of a
new protein-polymerized surface (Dalgleish & Banks, 199 1 ; van Boekel & Foikerts,
1994; Houlihan et al., 1992). With heating, an increasing amount of ~lactoglobulin
associated with MFGM. This was in agreement with the reported disulphide interaction
occurring between P-lactoglobulin and some MFGM proteins (Kim & Jimenez-Flores,
1995).
Heat treatment of the cream not only affected the formation of MFGM aggregates
with the cysteine-containing proteins present in skim mik , but also affected the iron
content, the solubility and the emulsifying properties of the MFGM isolates. Analysis of
variance of the dan showed a significant effect of temperature on ( i ) , the solubility of the
MFGM with urea, Tris/Triton X- 100 or sodium carbonate, (ii), the average droplet
diameter of 2 and 3% (w/v) MFGM isolate emulsions, and (iii), the arnount of P lactoglobulin associated with the MFGM. In general, apaxt from loss of iron, temperatures
up to 65'C did not result in signifcant changes of the functional properties of the MFGM
isolates.
Figure 7.11 shows a synthesis of our results, by relating the amount of j3-
lactoglobulin associated with the MFGM and the emdsifjing properties of the MFGM

01- - l l . - 1 1 I I - t . - . O i
O 0.2 0.4 0.6 0.8 1 1.2 1.4 1.6 Soluble Protein [mg/ml]
Figure 7.11: Amount of Plactoglobulin present in the MFGM isolates (W and ieft hand
scaie) and average diameter of emulsions prepared with 2% (wlv) MFGM isolates and 10%
(wlw) oil (e and right hand scale) plotted versus the arnount of iron present in the MFGM
(A) and the solubility of the MFGM isolates in Triflriton X- 100 buffer (B).

fractions (d3,2 of a 2% (w/v) MFGM isolate and 10% (wlv) oil emulsion) as functions of
the amount of ion and soluble protein in Tris/Triton X- 100 buffer. It is evident that
signif~cant comlations exist between all of these properties, and this demonstrates the
importance of h a h g on structure, composition and functionality of the membrane isolate.
Heating of cream before buttamaking is therefore a critical s tep w hich limits the
utilization of MFGM fractions isolated from b u t t e d . High temperatures caused
extensive denaturation of the MFGM isolates and interaction with skirn millc proteins.
Pasteurization of the cream affects the composition of the complex MFGM isolates by a
combination of effects of heat treatment on the MFGM itself and association of skirn milk
pmteins with the membrane. Thus, in considering the hinctional properties of the MFGM
fraction in buttermilk. it is necessary to know the heat matment given to (i), the milk kfore
cream separation; (ii), the crearn before chuming; and (iii), the butterrnillr after separation.

CHAPTER 8
STUDIES ON MEmRANE MATERIAL DERIVPNG FROM BUTTERMKKS
OBTAINED FROM DIE'FERENT BUTTER PROCESSES
8.1 INTRODUCTION
The milk fat globule membrane (MFGM) surrounds native fat globules and is
composed of phospholipid and protein. This intact membrane seems to be separated from
the iipid core by material which is densely stained in electromicrographs (Freudenstein et
al., 1979). Electron microscopy has also been perfomeci on MFGM isolates from
bovine milk, and it appears mostly in sheets with heavily stained compounds
(Freudenstein et al. , 1979; Franke et al., 198 1).
Increasing interest has been shown in the use of MFGM obtained from native fat
globules in the reconstitution of milk fat and in the stabilization of oil-in-water emulsions
(Kanno et al. , 199 1 ; Oehlmann et al. , 199 1 ; Chazelas et al., 1995). Previous results
descnbed in this thesis (chapters 6 and 7) have demonstrated the importance of
temperature of heating of the native fat globules of cream in determinhg changes in the
functional properties of the MFGM.
Heat and mechanical treatments lead to modifications of the fat globule
membrane composition and structure. For this reason, the functionality of the MFGM
material obtained from buttermillc from two different industrial processes was studied and
is describecl in ti-iis chapter. Buttermilk derived from two processes for the production of
butter: (i), the common production of butter by continuous churning, where crearn is
heated at high temperature and chunied in the presence of air, (ii), the process which
leads to anhydrous milk fat (AMF), where cream is heated at low temperature (-60'C)
and undergoes a centrifugai concentration (80% fat), and fat destabilization is obtained
by applying mechanical stress to the concentrated cream. In this research, the properties

of emulsions prepared with MFGM isolates obtained from different buttermilks were
studied. The emulsifying properties of these isolates were also studied in relation to the
microstructural characteristics of the emulsions, as indicated by transmission electron
microscopy observations.
8.2 MATERIALS AND METHODS
Buttermillcs were obtained from two local dairy plants. In the traditional churning
process, crearn was pasteurized at 85°C for 16 s and buttzrmilk was collected after
churning. In the AMF production, two sarnples were collected: the aqueous phase
O btained from the centrifugai concentration of the cream ("firs t buttermilk- AMF"), and
that obtained after mechanicai des tabilization, buttermilk. Details of the different
processes are described in 3.1. Sodium a i d e was added as preservative (0.026 (wfv)).
MFGM isolates and buttermilk isolates were prepared as described earlier (5.2.1 ).
MFGM isolates prepared frorn first buttemdk-AMF and from buttermilk from the AMF
process had a very similar composition, when analyzed by SDS-PAGE (data not shown).
8.2.1 Anaiytical Methods
Emulsions were prepared as described in 5.2.4. The droplet size disuibution of
the emulsions was detemined by integrated light scattering as previously outlined
(4.2.3). Solubility studies (7.2.8) were aiso performed on the MFGM isolates, as well as
SDS-PAGE on the isolates and the emulsions (5.2.6).

8.2.2 Transmission Electron Microscopy
The method of preparation of the samples for transmission electron microscopy
foliowed closely that of Goff et al. (1987). Ernukions were combined with 2% solution
of ultra low gelling agarose (FMC SeaPrep Agarose, Mandel Scientific) at 20°C at a
rate of 3 parts of agarose with 1 part of sample. The mix was solidified ovemight in the
refrigerator, fixed with 2% glutaraldehyde and 1 % malachite green as sugges ted by
Hayat (1993) in O.1M phosphate buffer (pH 7.0) for 24 h, and postfixed with 1% Os& in
O. 1 M irnidazolelphosphate buffer (Angemulier & Fahimi, 1982). After rinsing several
times with the buffer, the samples were dehydrated in graded concentrations of ethanol
and embedded in Spurr resin. Thin sections were poststained and viewed using a Hitachi
(H7 100 EM) microscope as descnbed in 4.2.1. Typically, magnification factors of
3 0 0 , 5OOOO and 80000 were used.
8.3 RESULTS
The isolates obtained from the two butter production lines showed very different
solubility, and they both differed from the MFGM isolates prepared frorn unheated
cream. MFGM isolates obtained from the AMF process seemed to be more soluble than
those obtained from buttermilk from traditional churning. Figure 8.1 illusuates the
protein composition of the soluble Fraction of MFGM isolates resuspended in 4M urea
and 5 mM rnercaptoethanol. Samples obtained from unheated cream were the most
soluble, and the four major MFGM proteins, xanthine oxidase, butyrophilin, PAS-6
and PAS-7, were present in the soluble phase (lanes 1 and 8, Figure 8.1). In the isolates
from unheated cream no whey protein was present. The amount of soluble protein in
the MFGM isolates from industrial buttemilks was lower than in the MFGM from

--_..- 4
t u-lactalbumin
1 Caseins
Figure 8.1 : SDS-PAGE of soluble phases after resuspension of the isolates with 4 M
urea and 5 mM 2mercaptoethanol. Lanes 1,8: MFGM isolate from unheated cream:
Lane 2: MFGM isolate from buttermilk obtained by churning process; Lane 3: MFGM isolate from buttermilk obtained from AMF process; Lane 4: buttermilk isolate from
unheated cream; Lane 5: buttermik isolate from churning process; Lane 6: buttermiik
isolate from AMF process; Lane 7: MFGM isolate from first buttermiik-AMF collected in the AMF process. SDS-PAGE performed in nducing conditions.

unheated crearn. MFGM isolates from buttermilk ob tained frorn the AMF process were
more soluble than those obtained from buttemilk from continuous churning. As already
shown in chapter 7, the solubility of ail MFGM isolates was fairly low.
ButtermiUr isolates (lanes 4-6, Figure 8.1) contained high amounts of caseins,
which were found in the soluble phase of samples treated with 4 M urea and 2-
mercaptoethanol. The samples obtained from buttermillcs collected from the indusinal
processes contained a considerable amount of whey proteins. Buttermilk isolaies
obtained from the AMF process contained more soluble a-laetalbumin than buttermilk
isolates from continuous chuming.
8.3.1 Ernulsi€jhg Properties of Isolates From Dinerent Butterrnilks
Emulsions prepared with MFGM isolate from buttermifi obtained by churning of
the crearn had a wide droplet size distribution, with a considerable number of large (> 5
pn) particles (Figure 8.2). Oil-in-water emulsions with 10% (wfv) soyabean oil showed
a monomodal distribution of sizes when prepared with 8% (w/v) MFGM isolate. At
lower concentrations (4%,6% w/v of MFGM isolate) the emulsions showed visible signs
of instability. These results agreed with those described in chapter 5.
The results obtained by integrated light scattering were compared with
observations from electron microscopy (Figure 8.3). The ernulsion droplets were
surrounded by a thin membrane and a considerable arnount of aggregates (darker
material, more heavily stained) seemed to be adsorbed at the interface. Srnall droplets
(<100 nm diameter) seemed to be attached to the surface of larger emulsion droplets.
Figure 8.3A represents a thin section of an emulsion prepared with 5% (w/w) oil and 3%
(wlv) MFGM isolate from bunermilk obtained from the chuming process. Integrated
light scattering measurements (Figure 8.2) showed that the particle size distribution of

0.01 O. 1 1 1 O 100
Average Diameter [pm]
Figure 8.2: Typical particle size distribution of emulsions prepared with 10% (w/w)
soyabean oil, 4% (w/v) MFGM isolate (bmken line), and 6% (wlv) MFGM isolate
(dotted line) and with 5% (wlw) soyabean oil and 3% (wlv) MFGM isolate (solid line).
MFGM isolates were prepared from buttermilk ob tained from continuous chuming.
Analysis perfonned by integrated iight scattering.

Figure 8.3: Electron micrographs of oil-in-water emulsions prepared with MFGM
isolates from buttermik obtained from the continuous churning process. (A) 5% (w/w)
oil and 3% (w/v) MFGM isolate emulsion; (B) 10% (w/w) oil and 4% (w/v) MFGM
isolate emulsion; (bar= 1.1 lm).

this emulsion was better than that of an emulsion prepared with 10% (wlw) oil and 4%
(w/v) MFGM isolate (represented in Figure 8.3B). While in figure 8.3A most of the
droplets were < 0.7 pm, in figure 8.3B the emulsion clearly showed particles > L Pm.
Even aUowing for some droplet destabilization during sample preparation for thin
sectioning, the emulsion prepared with 10% (wlw) oil and only 4% (wlv) MFGM isolate
had a large number of droplets > 5 pm (Figure 8.2). Fragments of MFGM were present,
mainly adsorbed ont0 the droplet surface. When insuficient material to cover the
interface was used in the emulsion, as is the case in figure 8.3B, ihe size of the emuision
droplets ranged from 10 nm up to > 1 Pm. Some large fragments of MFGM material
were shared by more than one individual droplet, causing bridging between them.
Droplets attached to one another through heavily stained material were clearly visible.
This heavily stained material probably consisted of protein and phospolipid aggregates,
residue of the original MFGM.
When centrifugation was carried out on emulsions prepared with an amount of
MFGM isolates in excess to that necessary to achieve complete coverage of the droplet
surface. not only a cream layer, but also a thick precipitate was separated. The interface
formed during rnicrofluidization was covered by an insoluble film, isolated on the
surface of the oil droplet T 'us , when a high concentration of MFGM isolate was used to
prepare the emulsion. the unadsorbed material was insoluble in the aqueous phase and
the excess formed a precipitate. Figure 8.4 represents a thin section of the precipitate.
Heavily s tained MFGM fragments denved from the isolate and partly disrup ted by
microfluidization constituted most of the pellet of these emulsions. Small oil droplets
were also present, with a considerable number of vescicles enveloped by MFGM
material.
Emulsions prepared with MFGM isolate from the continuous c h u m g process
showed very different structural features than did emulsions prepared with MFGM isolate

Figure 8.4: Thin section of precipitate obtained by centrifugation of an emulsion with
10% (wlw) oil and 8% (wlv) MFGM isolate. Bar = 1.1 Pm.

obtained from unheated cream (Figure 8.5). These emulsions contained an excess of
MFGM materiai at the interface (results described in chapter 6 have shown that 1 % (w/v)
MFGM isolate produced an emulsion with a monomodal size distribution). Each droplet
had a distinct membrane, iightly stained. On the thin membrane of the oil droplets there
were also extra aggregates of protein, with clusters of very small oiI droplets. These
emulsions, prepared with MFGM isolates from unheated cream, showed rnany
sirnilari ties with reporteci observations on micro fluidized milk. Micro fluidization creates
a large population of very small emulsion droplets (< 50 nm) (Strawbridge et al.. 1995;
Dalgleish et al., 1996). Those particles could not be disthguished with the integrated
light scattering equipment used in this research (Malvem Mastersizer X).
MFGM isolates from fxst buttennik and buttermilk obtained from the production
of AMF had very different emulsifying properties than those of MFGM isolates obtiiincd
from the traditional churning process. As shown in figure 8.6. srnaiier arnounts of
MFGM isolates were necessary to produce emulsions with a monomodal droplet size
distribution- When 10% (w/w) oil-in-water emulsions were prepared with 3 2 (w/v)
MFGM isolate. the distribution of particle size was monomodal and the emulsions had
an average diarneter of - 0.35 Pm. No difference in the emulsifying behaviour was
shown between the MFGM isolates obtained frorn first-buttermilk (obtained from the
first step of the AMF process) or buttermilk. Electrophoretic analysis showed that the
proteins (whey proteins and MFGM proteins) at the interface did not adsorb preferentially
(Figure 8.7).
The similar adsorption behaviour between whey proteins and MFGM proteins
observed in the emulsions prepared with MFGM isolate from the AMF process,
confirmed the hypothesis formulated previously (chapters 4,s) on MFGM isolates
adsorption: the oillwater interface is covered by aggregate material, and a tight
association exists between whey proteins and MFGM when the isolates are obtained from

Figure 8.5: Transmission electron micrograph of a 3% (w/v) MFGM isolate from
unheated cream and 5% (w/w) oil emulsion. Bar = 0.7pm.

0.0 1 o. 1 1 10 1 O0
Average Diameter [pm]
Figure 8.6: Size distribution of 10% (wlw) oil-in-water emulsions prepared with
MFGM isolates from f ~ s t buttermilk-AMF (A) and buttermilk (B) collected from the
AMF process. 1 % (wlv) (solid line); 3% (w/v) (broken line); 5 % (wlv) (do tted line).

O 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5
MFGM isolate concentration [%]
Figure 8.7: Arnount of adsorbed whey protein (H) and MFGM protein (a) (4% of the
total present in the emulsion) as detemined by SDS-PAGE electrophoresis and scanning
densitometry. Emulsions prepared with MFGM isolate from fit-buttemilk- AMF (A)
and buttermilk (B) collecteci from the AMF process. Results are the average of 3
independent expenments.

indusirial buttennilk. The arnount of protein adsorbed at the interface decreased as the
concentration of the MFGM isolate in the emuision was increased. There was an excess
of protein when 3% (wlv) MFGM isolate was used in a 10% (wlw) oil emulsion.
When diese emulsions were analyzed by elecvon microscopy, the structure was
sirnilar to that of ernulsions prepared with MFGM isolates from unheated cream (Figure
8.8). Most of the droplets were surrounded by a thin membrane (lightly stained) and
other aggregates were, at h e s , adsorbed on the surface. A few protein aggregates
(heavily stained) were aiso present and very small oil droplets were embedded in them.
In agreement with Iight scattering results, these emulsions did not contain droplets > 0.7
m . In the emulsions prepared with MFGM isolates obtained from the AMF process, as
well as with MFGM isolate obtained from unheated crem, no large membrane sheets
were apparent in the thin sections.
8.4 CONCLUSIONS
The results outlined in this chapter showed that the emulsifying properties of
MFGM isolates from buttermik, and from the aqueous phase from centrifugation (first
buttemik-AMF) obtained from the production of AMF, were better than those of
isolates from butte& collected from a traditional churning process. These results
confmed the hypothesis put forth in chapter 7, that heat treatrnent has a strong effect on
the emulsifying properties of the MFGM fraction isolated from buttermilk. In the
continuous churning process, the cream was pasteunzed at high temperature (85'C for 16
S) this treatment caused changes in the structure of the MFGM, and denatured the
MFGM proteins, causing whey proteins, especially plactoglobulin, to associate with the
MFGM (chapter 7). Heating seemed also to affect the susceptibility to disruption of
the MFGM during high pressure homogenization; the emulsions prepared with MFGM

Figure 8.8: Transmission electron micrographs of 10% (wlw) oil-in-water emulsions
prepared with 3% (w/v) MFGM isolate from buttermilk obtained from the AMF process;
Bar = 0.4 ym (A) and 0.7 pm (B).

isolate from buttermilk coilected after the traditional churning process still contained high
amounts of membrane sheets and fragments of MFGM. On the other hand, the MFGM
isolates from unheated cream did not show membrane-like structures after high pressure
homogenization.
These prelirninary studies on different industrial buttermilks brought more
evidence that the fimctionality of the MFGM isolates stnctiy depends on the processing
history of the buttermillc. Furthemore, not only the process but also the source of the
cream affected the functionality of the MFGM isolates. For example, the isolates
prepared from buttemilks from the AMF process showed substantial differences in
relation to the cream used to produce AMF, fresh cream or whey cream (byproduct of
cheesemaking) (data not shown).
A better understanding of the effects of processing on the functional properties of
the MFGM could be of great advantage in choosing ingredients from buttermilk
depending on the functionality required in food products.

CHAPTER 9*
STUDIES ON THE SUSCEPTIBILITY OF MEMBRANEDERlVED PROTEINS
TO PROTEOLYSIS AS RELATED TO
CHANGES IN THELR EMULSIFYING PROPERTIES
9.1 INTRODUCTION
In milk and cream, the milk fat globule membrane (MFGM) surrounds the fat
globules and prevents them from coalescing. The origin and composition of the MFGM
has been recently reviewed by Keenan & Dylewsky (1995). Because of i a origh
MFGM, which is a cornplex mixture of protein and phospholipid, is thought to have
good emulsifying properties, and it has been used to prepare oil-in-water emulsions
(Kanno et al. , 199 1 ; Chazelas et al. , 1995). The properties of the MFGM as a functional
ingredient in food emulsions strongly depend on the treatments the crearn undergoes
before isolation of the MFGM. During buttermaking crearn is subjected to high
temperature pasteurization, and the MFGM denved from the aqueous phase of this
process (buttermik) has poorer emulsifying properties than those of the MFGM frorn
unheated cream (chapter 7).
Enzymatic hydrolysis is a common method to modify the structure of proteins to
improve their functional properties. Hydrolysis has also been used as a tool to study
protein confornation, as weli as to determine asymmetry of protein organization in
membrane systems (Mather & Keenan, 1975; Dalgleish & Leaver, 199 1). Numerous
studies have shown that, depending on the protein conformation, there is a different
susceptibility to hydrolysis; for example, different peptides are formed when hydrolysis
is carried out in native solution, after heat treatrnent or with the protein adsorbed at
interfaces (Guo et al., 1995; Agboola & Dalgleish, 1996b). Caseins, which are
characterized by a flexible structure, seem more susceptible to proteolysis than globular
*Accepted for publication in this form by Food Research International. January 1998.

proteins ssuch as P-lactoglobulin, which is resistant to proteolytic digestion. If fi-
lactoglobulin undergoes heat matment or is subjected to changes which affect its
globular structure, proteolysis occurs at a faster rate than in the native solution (Schmidt
& Poll, 1991; Singh & Creamer, 1993; Guo et al., 1995). Hydrolysis of not only skirn
milk proteins, but also of some MFGM proteins has been studied in isolation (Amaya rt
al., 1990) or in the membrane of washed fat globules (Mather & Keenan, 1975)
The properties of proteins at interfaces are affected by conformational factors
such as molecular flexibility, amphiphilicity and hydrophobicity, charge and molecular
size. A minimum molecularsize and clustering of hydrophobic and hydrophilic amino
acids in distinct parts of the structure are essential to the functionality of hydrolyzed
peptides in ernulsions (Turgeon et al., 1992; Huang et al.. 1996). Agboola & Dalgleish
(1996b) studied the emulsifying properùes of hydrolyzed milk proteins and reported that
caseins seem to have better emulsifying capacity in their native fonn than when
hydrolyzed. Moreover, complete hydrolysis of P-lactoglobulin by tx-ypsin decreases its
emulsifying properties; this decrease might be caused by the formation of srnail peptides
with little amphiphilic character (Agboola & Dalgleish, 1996b).
This work aimed at describing the susceptibility to proteolytic enzymes of another
important group of proteins in mik, the MFGM proteins. They represent only 1% of the
total protein in milk, but they are fundamental for the stability of nanual fat globules.
Changes in the emulsifying properties of MFGM isolates occuning after proteolysis were
also determined. Two enzymes wirh very different specificity have been used, trypsin
and chymotrypsin. Trypsin acts on positively charged amino acids (lysine and arginine)
and chymotrypsin is specific for the peptide bond adjacent to hydrophobic amino acids
(Blow, 1971; Kei1, 1971).
This research provided more information on the potential utiiization of these
isolates as functional ingredients in food emulsions. MFGM is present in high amounts

in buttermilk; it codd be extracted from ihis byproduct and used as a functional
ingredient with high nuvitional value.
9.2 MATERIALS AND METHODS
9.2.1 MFGM Isolation
Butterrnillc (containing -3% wlw protein) and crearn (-40% wfw fat) were
obtained from a local da*. Sodium azide (0.02% w/v) was added to the samples as
preservative. Cream was kept at refrigeration temperature (4'C) ovemight and churned at
1O0C with a small s a l e churn (1 litre capacity) to obtain buttermilk. The aqueous phase,
buttemiik, was separated from butter granules with a cheesecloth. MFGM isolates were
prepared from buttemilk as descnbed in chapter 5 (5.2.1). The MFGM isolates
contained a mixture of phospholipid and protein, with 60% (w/w) average protein
concentration.
9.2.2 Emulsion Preparation
ALiquots of freeze dried isolates were resuspended in 20 m M phosphate buffer,
pH 7.0. Soyabean oil(10 or 20% w/w) was added and the mixture was homogenized at
56MPa using a microfluidizer as described in 4.2.2.
Trypsin, chymotrypsin and uypsin inhibitor (type II - T, inhibits both
chymotrypsin and trypsin in a ratio enzyme:inhibitor 1: 1) were purchased from Sigma
Chernicais (St. Louis, MO).

93.3-1 Hydrolysis in solution
MFGM isolates (1% wlv) were suspended in 20 m M phosphate buffer, pH 7.0.
After equilibration at 37'C, aliquots of trypsin or chyrnotrypsin (O. 1% wlv) were added
to give an enzyme-to-isolate ratio of 150; 1 500 or 1 : 1000. After preliminary
experiments the 1:500 enzyme-to-isolate ratio was chosen as optimal to study the
hydrolysis. During proteolysis at 37'C, aliquotî (250 pl) of MFGM isolate were
withdrawn at specified times, and added to the electrophoresis buffer to end the reaction.
To h ydroly ze MFGM isolates for homogenization, the sarn ples were res uspended in
phosphate buffer to a concentration corresponding to that of the final oil-in-water
emulsion, and incubated at 37'C with trypsin or chymotrypsin (1500 ratio). Afier 1
hour at 37°C the reaction was terminated by addition of an aliquot of 1 % (wlv) trypsin
inhibitor, enough to achieve a 1: 1 ratio with the added enzyme. The hydrolyzed solutions
were immediately homogenized with soyabean oil.
9.2.3.2 Surface-Hydrolysis
Emulsions prepared with different concentrations of MFGM isolates and 10-20%
(wlw) soyabean oil underwent treatment with uypsin or chymotrypsin (1 500 ratio). To
terminate the reaction, at various rimes, aiiquots (250 pl) of the surface-hydrolyzed
emulsions were withdrawn and added to ~ h e SDS-2-mercaptoethanol mixture. When
emulsions were prepared for integrated light scattering measurements, hydrolysis was
stopped by the addition of trypsin inhibitor ( 1 : 1 enzyme-to-inhibitor ratio).

92.4 SDS-PAGE
During the hydrolysis, aliquots of sample ( 2 5 0 ~ 1 ) were added to a mixture of 300
pl of SDS (20% wlv), 50 pl of bromophenol blue (0.05% wfv) and 100 pl of 2-
mercaptoethanol. Differences in protein composition at the oiYwater interface were
detennined by SDS-PAGE, after separating the oil droplets h m the serum phase as
descnbed in paragraphs 5.2.6 and 6.2.4. Emulsions were centrifuged at l O 0 g for 50
min (Beckman, mode1 L8-70 M). After collecting the oil droplets and drying them on a
füter paper, they were resuspended in 20 m M phosphate buffer, pH 7.0, to a fmal
concentration correspondhg to that of the original oil-in water emulsion. Samples of
emulsion and resuspended cream (200 pl) were then added to the SDS-electrophoresis
buffer. The samples were then denatured and loaded ont0 a 20% homogeneous Phast gei
(Phamacia Biotech) as descnbed previously (4.2.4).
9.2.5 Emulsion Droplet Size
The droplet size distribution of the unhydrolyzed and hydrolyzed emulsions was
measured by integrated light scattering using a Mastersizer X (Malvern Instruments.
Southboro. MA). The samples were dispersed in Milli-Q water with a dilution factor of
approximately 1:200 (4.2.3). The droplet size distribution was measured within two
hours from homogenization and also after 15 days of storage at refrigeration temperature
(4°C).

93.1 Susceptibility of MFGM to Proteolysis
As a result of their specifïc binding pockets, trypsin and chymotrypsin act on
different peptide bonds; trypsin can hydrolyze bonds adjacent to positively charged
arnino acids (argine, lysine) and chymotrypsin to bulky aromatic side chahs (tryptophan,
tyrosine and phenyialanine). When hydrolysis was performed with trypsin or
chymotrypsin on MFGM isolates from unheated crearn, SDS-PAGE revealed two
distinct polypeptide patterns (Figure 9.1). Aliquots of digested MFGEA isolates were
analyzed at various times, and intact protein was SM present in the hydrolyzed MFGM
samples after 3 h. In the control sample (lane 1) three major protein bands were
identified: xanthine oxidase (MW 150000 Da), butyrophilin (6700 Da) and PAS-6 and
PAS-7 (49000 Da) (Basch et al. , 1985; Mather & Jack, 1993; Berglund et al. , 1996).
Xanthine oxidase, which accounts for 8-10% of the total MFGM proteins, was
still present in considerable arnounts afier hydrolysis with trypsin; on the other hand,
butyrophilin, which represents 40% of the total protein associated with the MFGM, was
promptly hydrolyzed after a few min of incubation (Figure 9.1 A). Moreover, the
addition of trypsin to MFGM isolates produced some high MW polypeptides. These
were hydrolysis products of the large aggregates present in the MFGM. The primary
structure of butyrophilin, a membrane spanning protein, includes a large number of
lysine and arginine residues (Mather & Jack, 1993). This protein was highly susceptible
to proteolysis with trypsin. In contrast, the arginine and lysine residues of xanthine
oxidase were embedded in the membrane structure, making them inaccessible to trypsin.
Xanthine oxidase seemed to have a slightly lower MW in the hydrolyzed than in the
unhydroiyzed sample, in agreement with what was reported by Mihalyi (1978).

Figure 9.1: SDS-PAGE of MFGM isolates 1% wlv in phosphate buffer, treated with
trypsin (A) and chymotrypsin (B) in 1:500 enzyme-to-isolate ratio. Lanes correspond to different times of incubation at 37'C. Lane 1 : control (no enzyme added); Lane 2: 10 min; Lane 3: 20 min; Lane 4: 30 min; Lane 5: 60 min; Lane 6: 90 min; Lane 7: 120
min; Lane 8: 180 min. SDS-PAGE perfomed in reducing conditions.

When chymotrypsin was added to MFGM isolates, the polypeptide composition
of the hydrolyzates was quite different from that of the MFGM isolates treated with
trypsin. After 20 min of incubation, none of the original MFGM protein was identifiable
in the elecuophoretic pattern, and large polypeptides ( > SOOOO Da) were produced
(Figure 9.1B). In MFGM isolates from unheated cream, xanthine oxidase was more
susceptible to proteolysis when treated with chymotrypsin than with trypsin. These
results indicated that xanthine oxidase had more hydrophobic sites exposed than lysine
and arginine residues. Chymotrypsin is in fact specific for the peptide bonds adjacent to
hydrophobic amino acids, which in general are more buried within the structure of
proieins in solution. These results agreed with other literature data on xanthine oxidase,
which in isolation seems to form a product of 130000 Da when hydrolyzed by trypsin,
and fragments of 92000 and 42000 Da when hydrolyzed by chymotrypsin (Mihalyi,
1978; Amaya etal., 1990).
A very different susceptibility to proteolysis was found when MFGM isolates
were obtained from industrial buttermilk. Figure 9.2 illustrates the hydrolysis products of
these MFGM isolates after treatrnent with trypsin. The proteins were completely
hydrolyzed within the fiisst 10 min of incubation. By comparing the SDS-PAGE of
hydrolyzates of MFGM from unheated cream (Figure 9.1) to that of MFGM from
industrial buttermilk (Figure 9.2), it was concluded that the susceptibility of MFGM
proteuis to trypsin was related to the treatrnent the cream underwent before isolation of
the MFGM. No xanthine oxidase was found in these MFGM isolates after hydrolysis
with trypsin. The MFGM isolates from industriai buttermik containeci large arnounts of
P-lactoglobulin. This association of Plactoglobulin with the MFGM originated from the
heat-induced interactions between whey proteins and fat globules occurring during
pasteurization of the cream (chapter 7). Trypsin rapidly hydrolyzed the ~lactoglobuiin
present in the MFGM isolates. It is known that P-lactoglobulin is more susceptible to
proteolysis after heat-denaturation (Guo et al., 1995).

MFGM ( m e proteins j
5 6 7 g Migration
Figure 9.2: SDS-PAGE of 1% (wlv) MFGM isolates derived from industrial buttermilk.
treated with trypsin (enzyme-to-isolate ratio 1:500). Lanes correspond to different times of incubation at 37°C. . Lane 1: control (no trypsin); Lane 2: 10 min; Lane 3: 20 min;
Lane 4: 30 min; Lane 5: 60 min; Lane 6: 90 min; Lane 7: 120 min; Lane 8: 180 min. SDS-PAGE performed in reducing conditions.

A similar behaviour was observed when chymotrypsin was added to these MFGM
isolates. The differences in enzyme accessibility between the MFGM isolates derived
from unheated cream and those from industrial buttermilk were probably caused by a
combined effect of the heat treatment on changes in the conformation of the MFGM
proteins and on the overall composition of the membrane. This might result in a more
relaxed structure of the MFGM, making sites which were previously buried in the native
membrane available to hydrolysis.
9.3.2 Proteolysis at the oivwater interface - Surface Hydrolysis
When hydrolysis was carrieci out on emulsions prepared with MFGM isolates, the
polypeptide composition determined by SDS-PAGE did not show differences from that
of hydrolyzed MFGM in solution (Figure 9.3). Even if the same enzyme-to-isolate ratio
(1500) used in the MFGM isolate solutions was used for MFGM emulsions, no change
in the rate of hydrolysis was indicated by SDS-PAGE results. Xanthine oxidase was still
present in the hydrolyzed emulsion 3 h after the addition of trypsin (Figure 9.3). These
results showed that MFGM isolates, constituted by a mixture of protein and
phospholipid, adsorbed at the interface in aggregate fom, as suggested in previous
chapters. Changes in conformation often accompany the adsorption of pure proteins at
interfaces (Norde & Favier, 1992; Corredig & Dalgleish, 1995). In general, adsorption
ont0 oiUwater interfaces increases the protein sites which are accessible for enzyme
hydrolysis; for example, P-lactoglobulin is hydrolyzed at a much fas ter rate when
adsorbed at the interface than in solution (Agboola & Dalgleish, 1996b).
In MFGM isolates adsorbed onto the surface of oil droplets, no changes in the
susceptibility to proteolysis were apparent. Hydrolysis in solution and surface-hydrolysis
resulted in a very similar SDS-PAGE migration (Figure 9.4). A similar polypeptide
pattern was also observed when the composition of the emulsion droplets was analyzed

lint -a.
Butyrophilin >*la
Xanthine Oxidase 4 .<+-
_-. -- IC-- - - . - - . -
Migration
Figure 9.3 : SDS-PAGE of emulsions prepared with 108 (w/w) soyabean oil and 1 %
(wlv) MFGM isolate obtained from unheated cream, treated with trypsin (enzyme-to- isolate ratio 1 500). Lanes correspond to different times of hydrolysis. Lane 1 : control
(no enzyme added); Lane 2: 10 min; Lane 3: 20 min; Lane 4: 30 min; Lane 5: 60 min;
Lane 6: 90 min; Lane 7: 120 min; Lane 8: 180 min. SDS-PAGE perfonned in reducing conditions.

Figure 9.4: SDS-PAGE of emulsions prepared with 1 % (wfv) MFGM isolate from
unheated crearn and 10% (lanes 1-5) or 20% (lanes 6-8) (wlw) soyabean oil. Emulsions were treated with 1500 enzyme to isolate ratio for 1 h at 37'C. Lanes 1,6: control emulsion (no enzyme added); Lane 2: hydrolysis in solution widi trypsùi; Lane 3: surface-hydrolysis with trypsin; Lane 4: hydrolysis in solution with chymotrypsin; Lane 5: surface-hydrolysis with chymotrypsin; Lane 7: hydrolysis in solution with
chymotrypsin; Lane 8: hydrolysis in solution with trypsin. SDS-PAGE performed in reducing conditions.

by SDS-PAGE. However, the SDS-PAGE was limited to the determination of products
of hydrolysis with a size > 10000 Da.
A different polypeptide pattern was observed in emulsions prepared with MFGM
isolates from indusuial butterrniik (Figure 9.5). As in the experiments of hydrolysis in
solution, MFGM isolates from industrial buttermilk adsorbed at the interface were much
more susceptible to proteolysis than the MFGM obtained from unheated cream. Figure
9.5 illustrates the polypeptide composition of emulsions prepared with 3% (w/v) MFGM
isolate from industrial buttermilk and 10% (wlw) soyabean oil, before and after 1 h of
incubation with trypsin. Even if the original emulsion contained a large amount of
protein, especiaily if compared to emulsions prepared with MFGM from unheated cream,
the 3% (w/v) MFGM isolate from industrial buttermilk ernulsion was characterized by
visible instability. While the oil droplets separated from the control emulsion contaùied
di the major MFGM proteins, samples treated with trypsin did not show any residual
protein in the gel. A decreased amount of the large aggregates retained at the bonom of
the gel was dso observed. As already shown in the MFGM isolates from unheated
crearn, no differences could be O bsenred between emulsions wi th pro tein surface-
hydrolyzed and hydrolyzed in solution.
9.3.3 Particle Size Distribution of MFGM Isolates Emulsions
The particle size distribution of emulsions prepared with MFGM isolates. treated
with enzymes before or after homogenization, was cornpared to that of emulsions
prepared wiîh non hydrolyzed samples. Figure 9.6 iilustrates the typical droplet size
distribution of an emulsion prepared with 1% (wh) MFGM isolate from unheated crearn
and 10% (w/w) soyabean oil &ter 1 h of incubation with trypsin or chymotrypsin. In
this case, the MFGM isolate was present in sufficient amount to cover the newly formed

- -. - proteins
rilLC- - . --- . . -- -.-- - - Migration
Figure 9.5: SDS-PAGE of the oil droplets of emulsions prepared with 3% (w/v) MFGM
isolate from industrial buttermillc and 10% (w/w) soyabean oïl, treated with trypsin (ratio
1500). Lane 1: control emulsion (no trypsin added); Lanes 2,8: oil droplets control
emulsion (no trypsin); Lane 3: ernulsion prepared with MFGM hydrolyzed in solution; Lanes 4,7: oil droplets of the emulsion with MFGM hydrolyzed in solution; Lane 5: surface-hydrolyzed emulsion; Lane 6: oil droplets of a surface-hydrolyzed emulsion.
SDS-PAGE perforrned in reducing conditions.

0.01 0.1 1 10 100
Average Diameter [pm]
Figure 9.6 : Typical droplet size distribution of an emulsion prepared with 1% (wlv)
MFGM isolate from unheated cream and 10% (w/w) soyabean oil (solid Line); and same
emulsion surface-hydrolyzed for 1 h with trypsin (dotted Line) and chyrnotrypsin (broken
line). Analyses carried out with integrated light scattering.

interface, and these emulsions were characterized by a monomodal distribution of particle
size with an average diameter of 0.35 p.
No signifxcant changes from the distribution of the unhydrolyzed MFGM
emulsion were also observed when emulsions were prepared with MFGM hydrolyzed by
chymotrypsin or trypsin in solution. These results were quite different from those
reported by Agboola and Dalgleish ( 1999) for emulsions stabihzed by skim milk 1
proteins. Emulsions formed with P -1actoglobulin hydrolyzed in solution are
characterized by a considerable number of large (> 1 pm) particles. Emulsions prepared
with caseinate hydrolyzed in solution maintain their monomodal distribution but their
/ average diameter increased with hydrolysis (Agboola & Dalgleish, 1999 .
The decrease in the emulsifjmg properties of peptides formed by hydrolysis of
skirn milk proteins might be caused by the production of peptides which adsorb at the
interface, but are not large enough to effectively prevent droplet flocculation or
coalescence. Our results showed that some high MW polypeptides were still present
after hydrolysis of MFGM isolates from unheated crearn. SDS-PAGE electrophoresis in
non reducing conditions also revealed the formation of large polypeptides in MFGM
isolates. In these samples, among the products of hydrolysis, some amphiphilic
polypeptides might be present. This could improve the amulsifying properties of the
isolates, by lowering the interfacial tension. The coexistence of large aggregates with
small peptides could be highly effective for maintaining interfacial stability and
preventing coalescence.
When lower amounts of MFGM isolate (0.5% w/v MFGM, 10% wlw oil) or
higher amounts of soyabean oil (1% w/v MFGM, 20% w/w oil) were used to prepare oil-
in-water emulsions, their droplet size distribution was bimodai, with a considerable
number of droplets > 5 p m (Figures 9.7,9.8). In these cases, it was clearly observed
that hydrolysis improved the emulsming properties of MFGM isolates from unheated
cream. Both surface and solution-hydrolysis with trypsin im proved the droplet size

0.01 o. 1 1 10 1 O 0 Average Diameter
Figure 9.7: Typical droplet size distribution of 0.5% (w/v) MFGM isolate from unheated
cream and 10% (wfw) soyabean oil emulsion hydrolyzed with trypsin (A) and
chymotrypsin (B). Control emulsion (gray line); hydrolyzed in solution (solid line):
surface-hydrol y zed (broken line) .

0.01 o. 1 1 10 1 O0
Average Diameter [pm]
Figure 9.8: Typical droplet size distribution of 1% (w/v) MFGM isolate from unheated
cream and 20% (w/w) soyabean oil emulsion hydrolyzed with trypsin (A) and
chymotrypsin (B). Control emulsion (gray line); hydrolyzed in solution, before
homogenization (soiid lines); surface-hydrolyzed, after homogenization (broken lines).

distribution from bimodal to monomodal. Trypsinolysis disrupted the large particles and
the average diameter decreased to -0.6 Pm. When hydrolysis was canied out after
homogenization, the population of large droplets disappeared. It is known that, in the
presence of insufficient amounts of surface active material, large molecules adsorb onto
two distinct droplets causïng bndging flocculation (Dalgleish, 1995). Our resuits
indicated that in MFGM isolate emulsions, these bridges were broken by the addition of
trypsin and the size distribution shifted to a smailer average diameter (< 1 pm). It was
aiso hypothesized that the smaii peptides resulting from the trypsin hydrolysis contributed
to f i g in the gaps created by insufficient amount of MFGM material.
When chymottypsin was added to MFGM isolate solutions before
homogenization, the o v e r d size distribution did not improve (Figure 9.7B and 9.8B).
This differed substantially from the observations of hydrolysis in solution of MFGM
isolates carried out with trypsin. As already shown in figure 9.1, the products of
hydrolysis were very different for the two enzymes, and chymotrypsin seemed to
hydrolyze to a larger extent the MFGM proteins in the isolates. On the other hand, if
chymotrypsin was added after homogenization the size distribution of the emulsions
im pro ved.
Proteolysis carried out on MFGM isolates from industrial buttermilk improved the
emulsifying properties to a greater extent than for MFGM isolates derived from unheated
cream. Figure 9.9 illustrates the particle size disuibution of emulsions prepared with
different amounts of MFGM isolates from indusuial buttermillc hydrolyzed in solution by
trypsin. Momodal distributions of droplet sizes were observed with 3 and 6 % (w/v)
MFGM isolate, concentrations which did not produce good emulsions when
unhydrolyzed MFGM isolates were used. However, even with the addition of trypsin,
the amount of MFGM isolate which was needed to prepare emulsions with a small
average diameter was higher than that needed when MFGM isolates from unheated cream

o. 1 1 10
Average Diameter [pm]
Figure 9.9: Typical droplet size distribution of 10% soyabean-oil-in-water emulsions
prepared with MFGM hydrolyzed in solution by trypsin, containhg 1% (dotted line), 3 9
(broken line) and 6% (wlv) (solid h e ) of MFGM isolate from industrial buttermilk.

were used. A 1% (wlw) hydrolyzed MFGM isolate from industrial buttermi&
homogenized with 10% (w/w) soyabean oil resulted in an emulsion with a considerable
number of large droplets (> 50 pm) (Figure 9.9).
MFGM isolate from industrial buttermillc (3% w/v) produced an emulsion with a
monomodal distribution when trypsin was added before homogenization; however,
when surface-hydrolysis was carried out, large droplets were still present in considerable
amounts (Figure 9.10A). The MFGM isolates obtained from industrial buttermilk were
more susceptible to both solution and surface-hydrolysis (Figures 9.2.9.5). The absence
of large residual peptides with good amphiphilicity in the MFGM isolates was probably
the main cause of ihis instability. The overall distribution of droplet sizes improved
when emulsions were prepared with larger amounts of isolates (6% wfv). Trypsin, added
either before or after hornogenization, irnproved the droplet size distribution of the
emulsions (Figure 9.10B).
9.4 CONCLUSIONS
The susceptibiiity of MFGM isolates to hydrolysis was strongly affected by the
pretreatment of the crearn. While MFGM isolates from unheated crearn still contained
large polypeptides after trypsin or chymotrypsin hydrolysis, in MFGM isolates from
industrial buttermilk no polypeptides migrated in the SDS-PAGE. No changes could be
observed in the electrophoresis of MFGM isolates after surface-hydrolysis or hydrolysis
in solution.
When adsorbed ont0 oiljwater interfaces, the MFGM isolates from industrial
butte& were much more accessible to trypsin and chymotrypsin than the MFGM
isolates obtained from unheated crearn. It has been reported that reduction of disulphide

0.01 O* 1 1 10 1 O0 Average Diameter [prn]
Figure 9.10: Typical droplet size distribution of emulsions prepared with 10% (w/w) oil
and 3% (wfv) (A) and 6% (wfv) ( B ) MFGM isolates from indusuial buttermik. Control
emulsion (solid line); hydrolyzed in solution, before homogenization (broken iine);
surface-hydrolyzed, after homogenization (dotted line).

bonds caused by heat treatment rnay loosen the protein structure resulting in an increased
susceptibility to proteolysis. Structural changes might have occurred in the MFGM
during heating of the cream, causing exposure of buried sites and irnproving enzyme
accessi bility .
Emulsifying properties of MFGM isolates improved by addition of chymotrypsin
or trypsin. This differed from reported data on emulsions prepared with P-lactoglobulin
and caseins emulsions hydrolyzed in solution (Agboola & Dalgleish, 199&. In I n G M
isolats the concentration of large, arnphiphilic peptides was cntical to achieve the
desirable functionality in emulsions.
Trypsin hydrolysis improved the ernulsifying properties of MFGM isolates when
added either in solution or &ter homogenization. On the other hand, chymouypsin
improved the particle size distribution of the emulsions more effectiveiy when added after
emulsifkation. Chymoüypsin hydrolysis of MFGM isolates in solution produced
peptides with poorer emulsifying properties than those produced by trypsin. This agreed
with reported work on whey proteins, which suggested that tryptic peptides have better
emulsifying properties than chymotryptic peptides flurgeon et al. , 1992). After
homogenization, the addition of chymotrypsin and trypsin disrupted the bridges which
caused bndging flocculation in the MFGM isolate ernulsions, resulting in emulsions with
a monornodal distribution of droplet sizes with a small average diameter.
These results have important implications for the potential utilization of MFGM
isolates from buttemiilk. Hydrolysis of MFGM isolates may lead to ingredients with
improved functional properties and high nutritional value, because of their ongin from
the native fat globule membrane of milk.

CHAPTER 10
GENERAL CONCLUSIONS
The work presînted in this thesis questions the common perception that
buttermilk, when used as an ingredient in food products, imparts certain desirable
properties because of the presence of a phospholipid and pmtein mixture (the MFGM).
Previous work reported in the literature has suggested that the utilization of buttermilk
during the manufacturing of low fat cheese can improve cheese quality and increase
yields. The potential use of industrial buttermilk, concentrated by ultrafiltration, as a
milk supplement has also been studied by other authors. Our research focussed on the
isolation of the MFGM material present in buttermilk, to determine to what extent it
affects the functional properties of this byproduct. It was demonstrated that most of the
desirable properties atvibuted to burtermiik, when it is used as an ingredient in food
products, are in fact due to the casein fraction. rather than to the membrane matenal.
Thus, the real contribution of industrial buttermilk and its MFGM material, when used as
ingredient in food products, should be reevaluated.
Some of the questions outlined among the objectives of this thesis have been
answered: however, this research established the basis for further work. especially with
respect to improving understanding of how different industrial processes affect the
functionai properties of buttermilk.
Isolation of proteins from this byproduct cm be of great interest. MFGM
materid was isolated from buttermilk to study its properties. The method used can not be
industrially scaled, because of cost and low yields. Membrane filtration may present an
alternative method to separate the MFGM from the skim milk components. The source
of the buttemik might affect the feasibility of the separation andlor the fimal yield.

Results of research conducted on membrane material from washed fat globules
and fresh, unheated cream are only in part applicable to understanding the properties of
MFGM isolates obtained from industrial buttermilk. The types of membrane protein
present and their state seem to be different Thus, isolation of MFGM proteins might be
more successful if unheated cream or buttermilk from mild processes (low temperature
heating) are used than if indusuial buttermilk is used.
MFGM isolates s howed unique emulsifying be haviour, compared to other miik
proteins. Thus, MFGM may present an alternative ing~dient in food products.
Emulsions prepared with MFGM isolates had characteristics ranging from being similar
to caseinate emulsions, to being unstable, highly viscous or very prone to destabilization,
depending on the source of the buttermilk used. Further snidies on the effect of indusuial
processes on the final properties of the MFGM emuisions are required.
It is h o w n that the processing history of milk influences the functional properties
of skim milk proteins. Similarly, Our results indicated that the functionality of the
MFGM in butterrnilk is related to the processing history of rnilk. cream and buttermilk.
Heat treatment is the main deterrnining effect, and at temperatures as Iow as 65'C
changes in the MFGM isolates could be observed. This knowledge should be transferred
to those who formulate and manufacture food products containing buttermilk. Even if
heat seems to be the main factor in determining changes to the MFGM, the effect of
mechanical churning should be further investigated.
Cream pasteurization is a necessary step for product preservation in the dairy
industry. This thermal process must result in the elimination of pathogens and the
inactivation of enzymes. The tirne-temperature combination used is a compromise
between meeting safety criteria and preservation of nutritional and functional properties
of the components present The high sensitivit. to heat treatment of MFGM proteins
present in the cream might suggest an optimization of the process by the introduction of a

combination of membrane-based microfiltration with a low-heat treatment. which rnipht
produce a non heat-treated cream, which is pathogen free and suitable for chuming.
Our research mainly focused on the protein component of the MFGM isolate
(-60%), however, the phospholipid component (mainly composed of phosphatidyl
ethanolamine, phosphatidyl choline and sphingomyelin) may play an important role in
the stabilization of MFGM isolate emulsions, in the adsorption of the MFGM at the
oivwater interface, and may participate in the competition of the MFGM material with
the other buttermik components for adsorption sites.
The emulsifying properties of MFGM isolates cm be irnproved by addition of
trypsin and chymotrypsin. The susceptibility of MFGM proteins to hydrolysis differs
from that of skim milk proteins. Further studies on the functional propenies of these
polypeptides, together or in isolation. should be carried out. In addition. hydrolysis
may be used as means of determining what changes occur to the protein in the membrane.
and how they are related to changes in the structure of the MFGM.

CHAPTER 11
REFERENCES
Abou-Zeid, N.A. 1992. New type of domiati cheese of potential benefit to people with
high blood cholesterol. J. Dairy Res. 59 89-94.
Acharya, KR., Stuart, D.I., Walker, N.P.C., Lewis, M., Phillips, D.C. 1989. Refmed
structure of baboon a-lactalburnin at 1.7 A resolution. J. Mol. Biol. UNI 99- 127.
Agboola, S.O. 1996. Studies on the stability of oil-water emulsions formed using milk
proteins. Ph.D. Thesis. University of Guelph.
Agboola, S.O., Dalgleish, D.G. 1996a Effects of pH and ethanol on the kinetics of
destabilisation of oil-in-water emulsions containing milk proteins. J. Sci. Food Agric.
72 448-454.
Agboola, S.O., Dalgleish, D.G. 1996b. Enzymatic hydrolysis of milk proteins used for
emulsion formation. 1. Kinetics of protein breakdown and storage stability of the
emulsions. J. Agric. Food Chem. 44 363 1-3636.
Agiculture Canada. 1996. Dairy Market Review. Pp. 84-85.
Amaya Y., Yamazaki, K., Sato, M., Noda, K., Nishino, T., Nishino, T. 1990.
Proteolytic conversion of xanthine dehydrogenase from NAD-dependent to the 02-
dependent type. Biol. Chem 265 14 170- 14 175.
Anema, S.G., Klos termeyer. H. 1997. Heat-induced pH-dependent dissociation of
casein micelles on heating reconstituted skirn milk at temperatures below lOO0C. J.
Agric. and Food Chem 45 1 108- 1 1 15.
Angermüller, S., Fahimi, H.D. 1982. Imidazole-buffered osmium teuoxide: an excellent
stain for visualization of lipids in transmission electron microscopy. BiochemicaI J.
14 823-835.

Aoki, N., Kuroda, H., Urabe, M., Taniguchi, Y., Adachi, T., Nakamura, R., Matsuda, T.
1994. Production and characterization of monoclonal antibodies directed against
bovine millc fat globule membrane (MFGM). Biochim Biophys. Acrn. 1199 87-95.
Basch, J.J., Greenberg, R., Farrell, H.M. 1985. Identification of the milk fat globule
membrane proteins. II. Isolation of major proteins from electrophoretic gels and
cornparison of their aminoacid compositions. Biochim Biophys. Acta. 830 127- 135.
Berglund, L., Rasa ussen, J.T., Andersen, M.D., Rasmussen, M.S., Petersen, T.E. 1996.
P-cation of xanthine oxidoreductase from milk fat globule membranes and cloning
of complementary deoxyribonucleic acid. J. Dairy Sci. 79 198-204.
Blow, D.H. 197 1. The structure of chymotrypsin. In: n e enzymes. Ed. Boyer,
Academic Press, Oxford, pp. 185-2 12.
Brew, K., Grobler, J.A. 1992. a-lactalbumin. In: Advanced Dairy C h e m i s ~ - 1:
Proteins. Ed. P . Fox, Elsevier Applied Science, London, pp. 19 1-229.
Boye, J.L, Ismail, A-A-, AUi, 1. 1996. Effects of physicochemical factors on the
secondary structure of b-lactoglobulin. J. Dairy Res. 63 97- 109.
Campbell, L, Norton, I., Morley, W. 1996. Factors controlling the phase inversion of oil-
in-water emulsions. Neth. Milk & Dairy J. 50 167- i 82.
Chazelas, S., Razafindralarnbo, H.. De Chassart, Q.D., Paquot, M. 1995. Surface
Properties of the milk fat globule membrane: cornpetition between casein and
membrane material. In: Food Macromolecules and Colloidi, pp. 95-98
Cheng, S.G., Koch, U., Brunner, J.R. 1988. Characteristics of purified cows' miik
xanthine oxidase and its submolecular characteristics. J. Dairy Sci 71 90 1-9 16.
Corredig, M., Dalgleish, D.G. 1995. A differential microcalorimevic study of whey
proteins and their behaviour in oil-in-water emulsions. Colloids and surfaces B . 4
41 1-422.
Corredig, M., Dalgleish, D.G. 1996a. Effect of tempenture and pH on the interactions
of whey proteins with casein micelles in skim milk. Food Res. Int. 29 49-55.

Corredig, M., Dalgleish, D.G. 1996b. Effect of different heat treatments on the strong
binding interactions between whey proteins and miUç fat globules in whole mik. J.
Dairy Res. 63 44 1-449.
Coughlan, M. 1980. Aldehyde Oxidase, Xanthine Oxidase and Xanthine Dehydrogenase:
hydroxylases containing molybdenum. iron-sulphur and flavin. In: Molybdenum and
rnolybdenum-containing errzymes . Ed. Coughlan, M. Pergamon Press, Oxford. pp.
1 19-186.
Courthaudon, LL., Dickinson, E., Dalgleish, D.G. 199 la. Competitive adsorption of f&
casein and nonionic surfactants in oil-in-water emulsions. J. Colloid und interjacr
Sci. 145 390-395.
Courthaudon, J-L., Dickinson, E., Matsumura, Y., Clark, D.C. 199 1 b. Competitive
adsorption of p-lactoglobulin + Tween 20 at the oil-water interface. Colloidr and
SurJaces . 56 293-300.
Courthaudon, J.-L., Dickinson, E., Christie, W. W. 199 1c. Cornpetitive adsorption of
lecithin and P-casein in oil-in-water emulsions. J. Agric. Food Chem. 39 1365-
1368.
Courthaudon, J.-L., Girardet, J.-M., Chapal, C.. Lorient, D., Linden, G. 1995. Surface
activity and cornpetitive adsorption of milk component 3 and porcine pancreatic
lipase at the dodecane-water interface. In: Food Macromoleculrs and Colloidr . Ed.
Dickinson & Lorient. Royal Society of Chemistry, Cambridge, pp. 58-70.
Dalgleish, D.G. 1995. Structures and properties of adsorbed layers in emulsions
containing milk proteins. In: Food Macromlecules and ColloidF. Ed. Dickinson &
Lorient, The Royal Society of Chemistry, Cambridge, U.K, pp 23-33.
Dalgleish, D.G., Banks, J.M. 199 1. The formation of complexes between serum proteins
and fat globules during heatirig of whole m a . Milchwiss. 46 75-78.
Dalgleish, D.G., Law, A.J.R. 1988. pH-induced dissociation of bovine casein micelies.
1. Andysis of liberated caseins. J. Dairy Res. 55 529-538.

Dalgleish, D.G., Home, D.S., Law, A.J.R. 1989. Site related differences in bovine casein
miceiies. Bioch im Biophys. Acta. 991 383-387.
Dalgleish, D.G., Leaver, J. 199 1. The possible conformations of milk proteins adsorbed
on oillwater interfaces. J. of Colloid and Inteeace Sci. 141 288-294.
Dalgleish, D.G., Srinivasan, M., Singh, H. 1995. Surface properties of oil-in-water
emulsion droplets containing casein and Tween 60. J. Agric. Food Chem 43 235 1 -
2355
Dalgleish, D.G., Tosh, S.M., West, S. 1996. Beyond homogenization: the formation of
very srnail emulsion droplets during the processing of milk by a microfluidizer. Neth.
M i l k & D a i r y J . 50 135-148.
Dannenberg, F., Kessler, H.G. 1988. Reaction kinetics of the denaturation of whey
proteins in milk. J. Food Sci. 53 258-263.
Darling, D.F., Butcher, D.W. 1978. MiJ.k-fat globule membrane in homogenized crem.
J. Dairy Res. 45 197-202.
Darling, D.F, Dickson, J. 1979. Electrophoretic mobility of casein micelles. J. Dai-
R a . 46 441-45 1
deWit, J.N., Klarenbeek, G. 198 1. A differential scanning calorimetric study of the
thermal behaviour of bovine p-lactoglobulin at temperatures up to 16O0C. J. Dairy
Res. 48 293-302.
Dickinson, E. 1984. Protein-stabiLized emulsions. J. Food Eng. 22 59-74.
Dickinson, E. 1986. Mixed proteinaceous ernulsifiers: review of corn petitive pro tein
adsorption and the relationship to food coiioid stabilization. Food Hydrocoll. 1 3-23.
Dickinson, E., Stainsby, G. 1982. Structure and stability of electrocratic colloids. In
Colloids in food. Applied Sci., London, pp. 33-66.
Dickinson, E., Matsumura, Y. 199 1. Time-dependent polymerization of P -1actoglobulin
through disulphide bonds at the oil-water interface in emulsions. Int. J. Biol.
Macromol. 13 26-30.

Dickinson, E., Golding, M. 1997. Depletion flocculation of emulsions containing
unadsorbed sodium caseinate. Food Hydrocoll. 11 13- 18.
Fang, Y., Dalgleish, D.G. 1993% Dimensions of the adsorbed layers in oil-in-water
emulsions stabilized by caseins. J. Colloid and InterJace Sci. 156 329-334.
Fang, Y., Dalgleish, D.G. 1993b. Casein adsorption on the surfaces of oil-in-water
emulsions rnodified by lecithin. Colloih and Surfaces B. 1 357-364.
Fang, Y ., Dalgleish, D.G., 1996. Competitive adsorption between
dioleoylphosphatidylcholine and sodium caseinate on oil-water interfaces. J. Agric.
Food Chem 4459-64.
Ferragut, V., Chiralt, A. 1996. Microstructure of oil-in-water low-fat ernulsions
containing skim milk powder and locus bean gum. Lebensm - Wiss. u. -Technul. 29
648-653.
Franke, W.W., Heid, H.W., Grund, C., Winter, S., Freudenstein, C., Schmid, E., Jarasch,
E-D, Keenan, T.W. 198 1. Antibodies to the major insoluble milk fat globule
membrane-associated protein: specific location in apical regions of lactating epithelial
cells. J. of Ce11 Biol. 89 485-494.
Freudenstein, C., Keenan, T.W., Eigel, W.N., Sasaki, M., Stadler, J., Franke, W.W.
1979. Preparation and characterization of the inner coat material associated with fat
globule membranes from bovine and human milk. Erp. Cell Res. 118 277-294.
Fujiki, Y., Hubbard, A.L., Fowler, S., Lazarow, P.B. 1982. Isolation of intracellular
membranes by means of sodium carbonate treatment: application to endoplasmic
reticulurn. J. Cell Biol. 93 97- 102.
Gezirnati, J., Creamer, L.K., Singh, H. 1997. Heat-Induced interactions and gelation of
mixtures of B-lactoglobulin and a-lactalbumin. J. Agric. Food Chem. 45 1130- 1136.
Girardet, J.-M., Linden, G., Loye, S., Courthaudon, J.-L., Lorient, D. 1993. Study of the
rnechanism of Lipolysis inhibition by bovine miik proteose-peptone component 3. J.
Dairy Sci. 76 2156-2163.

Girardet, J.-M., CoddeviUe, B., Pancke, Y., Strecker, G., Campagna, S., Spik, G., Linden,
G. 1995. Structure of glycopeptides isolated from bovine milk component PP3. Eur.
J. Biochem. 234 939-946.
Goff, H.D., Liboff, M., Jordan, W.K., Kinsella, J.E. 1987. The effects of polysorbate 80
on the fat emulsion in ice cream mix: evidence from transmission electron
microscopy snidies. Food Microsrr~cture 6 193- 198.
Goff, H.D., Jordan, W.K. 1989. Action of emulsifiers in promoting fat destabilization
during the manufacture of ice cream. J. Dairy Sci. 72 1 8-29.
Goff, H.D., Kinselia, J.E., Jordan, W.K. 1989. Influence of various milk protein isolates
on ice cream emulsion stability. J. Dairy Sci. 72 385-397.
Goff, H.D., Hill, A.R. 1993. Chemistry and Physics. In: Dairy Science and Technology
Handbook I . PrincipEes and Properties. Ed. Hui, VCH Publishers, New York, pp.
1-81.
Greenwalt, D.E., Mather, I.H. 1985. Characterization of an apically derived epithelial
membrane glycoprotein from bovine mi&, which is expressed in capillary endothelia
in diverse tissues. J. Cell Biol. 100 397-408.
Greenwalt, D.E., Watt, K.W. K., So, O.Y., Jiwani, N. 1990. PAS IV, an integral
membrane protein of mammary epithelial cells, is related to platelet and endothelid
cell CD36 (GP N) . Biochernis~. 29 7054-7059.
Guo. M.R., Fox, P.F., Flynn, A., Kindstedt, P.S. 1995. Susceptibility of B-lactoglobulin
and sodium caseinate to proteolysis by pepsin and trypsin. J. Dairy Sci. 78 2336-
2344.
Hambling, S.G., McAlpine, A.S., Sawyer, L. 1992. ~lactoglobuiin. In: Advanced
Dairy Chemistry. Vol. I. Proteins. Ed. P.F. Fox, Elsevier Appl. Science London,
pp. 141-190.
Hayat, M.A. 1993. Staining and Related reagents. In: Stains und cytochemical methuds.
Plenum press, New York, pp. 7 1-369.

Haynes, C.A., Norde, W. 1994. Globular proteins at solidlliquid interfaces. Colloids
and Surjiuces B. 2 5 17-566.
Haiiet, R. 1995. An introduction to light scattering. Lecture Notes, University of
Guelph, Ontario.
Hoffmann, M.A.M., van Mil, P.J.J.M. 1997. Heat-induced aggregation of j%
lactoglobuiin: role of the free thiol group and disulphide bonds. J. A g k . Food Chem.
45 2942-2948.
Holt, C. 1992. Structure and stability of bovine casein micelles. Advances in Protein
Chemisv 43 63- 15 1.
Holt, C., Davies, D.T., Law, A.J.R. 1986. Effects of colloidal calcium phosphate content
and free calcium ion concentration in the milk serum on the dissociation of bovine
casein micelies. J. Dairy Res. 53 557-572.
Holt, C., Home, D.S. 1996. The hairy casein micelle: evolution of the concept and its
implications for dairy technology. Neth. Milk & Dairy 1. 50 85- 1 1 1.
Houlihan, A. V., Goddard, P.N., Kitchen, B. J., Masters, C. J. 1992. Changes in structure
of the bovine milk fat globule membrane on heating whole miik. J. Da ig Rrs. 59
324-329.
Huang, X.L., Catignani, G.L., Swaisgood, H.E. 1996. hproved emulsifying properties
of P-barre1 domain peptides obtained by membrane-fractionation of a limited tryptic
hydrolysate of P-lac toglobulin. J. Agric. Food Chem 44 3437-3443.
Hunt, LA., Dalgleish, D.G. 1994a. Effect of pH on the stability and surface composition
of emulsions made with whey protein isolate. J. Agric. Food Chem. 42 2 13 1-2 135.
Hunt, J.A., Dalgleish, D.G. 1994b. Adsorption behaviour of whey protein isolate and
caseinate in soya oil-in-water emulsions. Food Hydrocoll. 8 175- 1 82.
Hunt, J.A., Ddgleish, DG. 1995. Heat stability of oil-in-water emulsions containing
miik proteins: effect of ionic strength and pH. J. Food Sci. 60 1088- 109 1.

Hunter, R.J. 198 1. The calculation of zeta poten tid. In: Zeta Potenrial in Colloid
Science. Principles m d Applicahons. Academic Press. London, pp. 59- 1 24.
Hunter, R.J. 1993. Detemination of particle size. In: Introduction to modem colloid
science. Oxford University Press, Oxford, U.K, pp. 58-96.
Ulingworth, D., Bissel, T.G. 1994. Anhydrous milk fat products and applications in
recombination. In: Fats in food producrs. Ed. Moran & Rahjah, Blackie Academic
and Professional, Chaprnan & Hall, Oxford, pp. 1 1 1 - 1 54.
Innocente, N., Blecker, C. Deroanne, C.. Paquot, M. 1997. Langmuir W m balance
study of the surface properties of a soluble fraction of milk fat-globule membrane.
Agric. Food C h e m 45 1559- 1563
Jack, L.J.W., Mather. I.H. 1990. Cloning and analysis of cDNA encoding bovine
butyrophilin, an apical glycoprotein expressed in rnarnmary tissue and secreted in
association with the milk-fat globule membrane during lactation. J. Biol. Chem 24
1448 1 - 14486.
Jang, H.D.. Swaisgood, H.E. 1990. Disulfide bond formation between thermally
denatured P-lactoglobulin and casein in in casein micelles. J. Dairy Sci. 73 900-904.
Kalab M. 1980. Possibilities of an electron-rnicroscopic detection of buttemilk made
from sweet cream in adulterated skim milk. Scanning Elecnon Microscopy 3 645-
65 1.
Kanno, C. 1989. Emulsifying properties of bovine milk fat globule membrane in milk fat
emulsion: conditions for the reconstitution of milk fat globules. J. Food Sci 54 1534-
1539.
Kanno, C. 1990. Secretory membranes of the Iactating mamrnary gland. Protoplasma
159 184-208.
Kanno, C., Kim, D.-H. 1990. A simple procedure for the preparation of bovine milk fat
globule membrane and a cornparison of its composition, enzymatic activities and

electrophoretic properties with those prepared with other methods. Biol. Chem. 54
2845-2854.
Kamo, C., Shimomura, Y., Takano, E. 199 1. Physicochemical properties of m i k fat
emulsions stabilized with bovine milk fat globule membrane. J. Food Sci. 56 1219-
1223.
Keenan, T.W., Freudenstein, C., Franke, W.W. 1977. Membranes of mammary gland.
Xm. A Lipoprotein cornplex derived from bovine mi& fat globule membrane with
some preparative characteristics resembling those of actin. Cytobiologie . 14 259-
278.
Keenan, T.W., Heid, H.W., Stadler, J., Jarasch, E.-D., Franke, W.W. 1982. Tight
attachent of fatty acids to proteins associated with milk lipid globule membrane.
Eur. J. Cell Biol. 26 270-276.
Keenan. T.W., Dylewski, D.P., Woodford, T.A., Ford, R.H. 1983. Origin of miik fat
globules and the nature of the m i k fat globule membrane. In: Developmenfs in Dai-
Chemistry - 2. Lipids. Ed. P.Fox, Applied Science Publishers, London, pp. 83- 1 18.
Keenan, T.W., Dylewski, D.P. 1995. Intracellular ongin of milk lipid globules and the
nature and structure of the milk lipid globule membrane. In: Advanced Dairy
Chemistqt Volume 2: Lipidr. Ed. P.F. Fox, Chapman & Hall, London, U.K, pp.
89- 130.
Keil, B. 197 1. Trypsin. In: me enqmes. Ed. Boyer, Academic Press, Oxford, pp. 250 -
275.
Kim, D.H., Kanno, C., Mizokami, Y. 1992. Purification and characterization of major
glycoproteins, PAS-6 and PAS-7, from bovine milk fat globule membrane. Biochim
Biophys. Acta 1122 203-211.
Kim, H.-H,Y ., Jirnenez-Florés, R. 1995. Heat induced interactions between the proteins
of m i k fat globule membrane and skim milk. J. Dairy Sci. 78 24-35.

Law, A.J.R. 1996. Effects of heat treatment and acidification on the dissociation of
bovine casein micelles. J. Dairy Res. 63 35-45.
Law, A.J.R.. Home, D.S., Banks, J.M., Leaver, J. 1994. Heat-induced changes in the
whey proteins and caseins. Milchwiss. 49 125- 129.
Leaver, J., Dalgleish, D.G. 1992. Variations in the binding of p-casein to 011-water
interfaces detected by trypsin-catalysed hydrolysis. I. Colloid and In te~ace Sci. 149
49-55.
LuA, J.H. 196 1. Improvements in epoxy resin cmbedding methods. J. Biophvs. Biochrrn
Cytology. 9 409-4 14.
Malin, E.L., Basch, J.J., Shieh. J.J., Sullivan, B.C., Holsinger, V.H. 1994. Detection of
adulteration of buttermilk powder by gel elecrophoresis. J. Dairy Sci 77 2 199-
2206.
Malmsten, M., Bergenstahl, B., Nyberg, L., Odham, G. 1994. Sphingomyeiin from milk
- charactenzation of Liquid crystalline, liposome and emulsion properties. JAOCS
71 102 1- 1026.
Mangino. M.E., Brunner, I.R. 1977. Isolation and partial characterization of xanthine
oxidase associated with the milk fat globule membrane of cows' milk. J. D u i ~ Sci.
60 841-850.
Markwell, M.A.K., Haas, S.M., Bieber, L.L., Tolbert, N.E. 1978. A modification of the
lowry procedure to simplify protein determination in membrane and lipoprotein
sarnples. Anal. Biochem 87 206-2 10.
Masson, G., Jost, R. 1986. A study of oil-in-water emulsions stabilized by whey
proteins. Colloid Polymer Sci. 264 63 1-638
Mather, LH., Keenan, T.W. 1975. Studies on the structure of milk fat globule
membrane. J. Membrane Biol. 21 65-85.

Mather, I.H., Weber, K., Keenan, T.W. 1977. Membranes of mammary gland. XII.
Loosely associated proteins and compositionai heterogeneity of bovine miik fat
globule membrane J. Dairy Sci . 60 394-402.
Mather, LH., Tampiin, C.B., Inring, M.G. 1980. Separation of the proteins of bovine
milk-fat-globule-membrane by electrofocusing with retention of enzymatic and
immunological activity. J. B i o c h e m i s ~ 110 327-336.
Mather, I.H., Jack L.J.W. 1993. A review of the molecular and cellular biology of
butyrophilin, the major protein of bovine milk fat globule membrane. ' Dairy Sci.
76 3832-3850.
Matsumura, Y., Mitsui, S., Dickinson, E., Mori, T. 1994. Cornpetitive adsorption of a-
lactalbumin in the molten globule state. Food Hydrocoll. 8 555-566.
Mayes, J.J., Urbach, G., Sutherland, B.J. 1994. Does addition of buttermilk affect the
organoleptic properties of low-fat cheddar cheese? Australiun J. of Dairy Technology
49 39-41.
McCrae, C.H. 1994. Homogenization of milk ernulsions: use of microfluidizer. J. Sociey of Dairy Tech. 47 28-3 1.
McPherson, A.V., Kitchen, B.J. 1983. Reviews of the progress of dairy science: the
bovine milk fat globule membrane - its formation, composition, structure and
behaviour in milk and dairy products. J. Daity Res. 50 107- 133.
McPherson, A.V., Dash, M.. Kitchen, B.J. 1984. Isolation and composition of milk fat
globule membrane material. 1. From paçteurized milks and creams. J. Dairy Res. 51
279-287.
Mihalyi, E. 1978. Effect of endopeptidases on native proteins. Artificial limited
proteolysis reactions. In: Application of proteolytic enzymes to protein structure
studies. Vol. II . CRC Press, Rorida, pp. 1-45.
Mistry, V.V., Metzger, L.E., Maubois, J.L. 1996. Use of ultrafiltered sweet buttermilk in
the manufacture of reduced fat cheddar cheese. J. Dairy Sei. 79 I 137- 1 145.

Myers, D. 199 1. Electrostatic forces and the electrical double layer. In: Su@aces.
Interfaces und CulluidS. Prïnciplrs and application. VCH Publ., New York, pp. 69-
85.
Neira, LM., Mather, I.H. 1990. Biochemical and immunological cornparison of bovine
butyrophilin with a butyrophilin-like glycoprotein in guinea pig milk-fat-globule
membrane. Protuphsrna. 159 168- 178.
Norde, W., Favier, J.P. 1992. Structure of adsorbed and desorbed proteins. Colloids
Surfaces. 64 87-93.
Nylander, T., Wahlgren, N.M. 1994. Corn petitive and sequential adsorption of h a s e i n
and P-lactoglobulin on hydrophobic surfaces and the interfacial structure of pcaîein.
J. Colbid Interface Sci. 162 15 1 - 162.
Oehlmann, SM., Duncan, S.E., Keenan, T.W. 1994. Butteroil emulsification with mi&-
derived membrane and pro tein fractions. J. Food Sn'. 59 53-56,66.
Ono, T., Murayama, T., Kaketa, S., Odagiri, S. 1990. Changes in the protein composition
and size distribution of bovine casein micelles induced by cooling. Agric. Biol.
Chem 54 1385-1392.
Papiz, M.Z., Sawyer. L., Eliopoulos, E.E., North, A.C.T., Findlay, J.B.C.,
Sivaprasadarao, R., Jones, T.A., Newcorner, M.E., Kraulis, P.J. 1986. The structure
of P-Iactoglobulin and its sirnilarity to plasma retinol-binding protein. Nature. 324
383-384.
Parris, N., Purcell, J.M., Ptashkin, S.M. 199 1. Thermal denaturation of whey proteins in
skim milk. J. Agric. Food Chern. 39 2167-2 170.
Parris, N., Hollar, C.M., Hsieh, A.,Cockley, K.D. 1997. Thermalstabilityof whey
protein concentrate mixtures: aggregate formation. Dairy Sci. 80 19-28.
Pérez, M.D., Calvo, M. 1995. Interaction of -lactoglobulin with retinol and fatty acids
and its role as a possible biological function for thjs protein: a review. J. Dairy Sci.
78 978-988.

Rajagopalan, K.V. 1985. Purification of bovine milk xanthine oxidase. In: CRC
Handbook of methods for uqgen radical research. Ed. Greenwald, CRC press, Inc.
Rorida, pp. 2 1-23.
Ramachandm Rao, H.G., Lewis, M.J., Grandison, AS. 1995. Effect of pH on flux
during ultrafiltration of sweet whey and buttermilk. J. Dairy Res. 62 44 1-449.
Rellcin, P., Eynard, L., Launay, B. 1992. Themodynamic parameters of P-lactoglobulin
and a-lactalbumin. A DSC study of denaturation by heating. Thermochimica Acta
204 111-121.
Ren, S., Scandale, LN., Ariga, T., Zhang, Y., Klein, R.A., Hartmann, R.. Kushi. Y..
E g p , H., Yu, R.K. 1992. O-Acetylated gangliosides in bovine buttermilk. J. Biol.
Chem. 267 12632- 12638.
Robin, O., Remillard, N., Paquin, P. 1993. Influence of major process and formulation
parameters on microfluidized fat globule size distribution and example of practical
consequence. Colloids and S i ~ ~ u c e s : Phÿsicocheinical and Engineering Aspects. 80
2 1 1-222.
Schmidt, D.G.. Poll, J.K. 199 1. Enzymatic hydrolysis of whey proteins. Hydrolyis of a-
lactalbumin and P-lactoglobulin in buffer solutions by proteolytic enzymes. Nrth.
Milk. Dai- J. 45 225-240.
Schoenfuss. TC., McGregor, J.U. 1997. Improving the flavour of a low fat chcrdar
cheese with homogenized cream and commercial buttermilk powder. J. Dain Sci.
80 Suppl. 1: 100.
Shantha, N.C. 1992. Thin-layer chromatography-flame ionization detection Iatroscan
system. J. Chromatography. 624 21-35.
Sharma, S.K., Dalgleish, D.G. 1993. Interactions between milk serurn proteins and
synthetic fat globule membrane during heating of homogenized wholc mik. J. Agric.
Food Chem 41 1407-1412.

Sharma, S.K., Dalgleish, D.G. 1994. Effect of heat treatrnenü on the incorporation of
milk serum proteins into the fat globule membrane of homogenized milk. J. Dain
Res. 61 375-384.
Sharma, R., Singh, H., Taylor, M.W. 1996. Composition and structure of fat globule
surface layers in recombined mik. J. Food Sci 61 28-32.
Shimizu, M., Kanno, C., Yamauchi, K. 1976. Dissociation of the soluble glycoprotein of
bovine milk fat globule membrane by sodium dodecyl sulfate. Agric. Biol. Chem. 10
171 1-1716.
Shimizu, M., Kanno, C., Yamauchi, K. 1978. Selective solubilization of bovine miik fat
globule membrane proteins with guanidine hydroclondr and disposition of some
proteins. AgNc. Biol. Chrm. 42 2309-2314.
Singh, H., Creamer, L.K. 1993. In vitro digestibility of whey proteinlw-casein
complexes isolated from heated concentrated mik. J. Food Sri. 58 299-302.306.
Singh, H., Fox, P.F., Cuddigan, M. 1993. Emulsifying properties of protein fractions
prepared from heated milk. Food Chrin. 47 1-6
Sorensen, E.S., Petersen, T.E., 1993. Phosphorylation, glycosilation and mino acid
sequence of component PP3 form proteose peptone fraction of bovine milk. J. D a i g
Res. 60 535-542.
Steadrnan, B.L., Thornpson, K.C., Middaugh, C.R., Matsuno. K.. Vrona. S., Lawson.
E.Q., Lewis, R-V. 1992. The effects of surface adsorption on the thermal stability of
proteins. Biotech. Bioeng. 40 8- 1 5.
Suange, E., Malin, E.L., van Hekken, D.L., Basch, J.J. 1992. Chromatographie and
electrophoretic methods used for analysis of milk proteins. J. Chroimtograph. 624
8 1- 102.
Strawbridge, K.B ., Ray, E., Hallet, F.R., Tosh, S.M., Ddgleish, D.G. 1995.
Measurement of particle size distributions in milk homogenized by a microfluidizrr:

estimation of populations of particles with radii less than 100 nrn. J. Colloid
InterfBce Sci. 171 392-398.
Swaisgood, H.E. 1992. Chemistry of the caseins. In: Advanced Dairy Chemistry - 1:
Proteins. Ed. P . Fox, Elsevier Applied Science. London, pp. 63- 1 10.
Taylor, M.R., Peterson, J.A., Ceriani, R.L., Couto, J.R. 1996. Cloning and sequence
analysis of human butyrophilin reveals a potential receptor function. Biochim.
Biophvs. Acta 1306 1-4.
Turgeon, S.L., Gauthier, S.F., Molle D., Léonil, 1. 1992. Interfacial properties of tryptic
peptides of B-Iactoglobulin. J. Agric. Food Chem. 40 669-675.
Tvrzicka, E., Votruba, M. 1994. Thin-Layer Chromatography with Flame-Ionization
Detection. In: Lipid Chromatographie Analysis . Ed. Shibamoto, Dekker, New York.
pp. 5 1-73.
van Boekel, M.A.J.S., Foikerts, T. 199 1 . Effect of heat treatment on the stability of rnilk
fat globules. Milchiviss. 46 758-765.
Walstra, P. 1974. High-melting uiglyceridrs in the fat globule membrane: on rirtrfiic~:'
Neth. Milk Dairy J. 28 3-9.
Walstra. P. 1983. Physical chemistry of m i k fat globules. In: Developments in d a i p
chemistry - 2. Lipids. P.Fox ed.. Applied Science Publishers, London, pp. 1 19- 158.
Walstra, P. 1993a. Principles of emulsion formation. Chem Engineering Sci. 48 333-
349.
Walsua, P. 1993b. Introduction to aggregation phenomena in food colloids. In: Food
colloids and polyrners: stabiliw and mechanical properties. Ed. Dickinson &
Walsua, Royal Society of Chemistry, Special Publ. 1 13, Cambridge, pp. 3- 15.
Walstra, P.. Oortwijn, H. 1982. The membranes of recombined fat globules. 3. Mode of
formation. Neth. Milk Dairy J. 36 103- 1 13.

Weiner. B.B. L984. Partide and droplet sizing using Fraunhofer diffraction. In: Motlrm
Methods of Particle Size Analysis. Ed. Barth, John Wiley & Sons, New York. pp.
135- 172.
Wooding, F. B.P., Kemp, P. 1975. High-melting-point triglycendes and the milk-fat
globule membrane. J. Dairy Res. 42 4 19-426.
Yaguchi, M., Davies, D.T., Kim. Y.K. 1967. Preparation of K-casein by gel filtration. J.
Dairy Sci. 51 473-477.
Yanagita, Y.; Kagawa, Y. 1986. Solubilization and purification of membrane proteins.
In: Techniques for the analysis of membrane protrins. Ed. Ragan & Cherry.
Chapman & Hall. London, pp. 6 1-76.

IMAGE EVALUATION TEST TARGET (QA-3)
APPLIED - 4 IMAGE . lnc = 1653 Eaçt Main Street - -. - Rochester, NY 14609 USA -- --= Phone: 71 6/482-0300 -- - Fax: 71 6/28&5989
O 1993. Appiied Image. Inc. All Rights Reserved










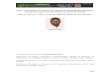


















![Benefits of Lactoferrin, Osteopontin and Milk Fat Globule ... · especially -casein [8,9], while endogenous peptide concentration in human milk is between 10 and 20 mg/L [10], which](https://img.pdfslide.us/doc/110x75/5f75caba52fa9d3bae18b223/benefits-of-lactoferrin-osteopontin-and-milk-fat-globule-especially-casein.jpg)