Embed Size (px)
Citation preview

Eur. J. Biochem. 220, 717-727 (1994) 0 FEBS 1994
Properties of multiple G A mismatches in stable oligonucleotide duplexes Andrew LANE', Susanne EBELZ and Tom BROWNZ ' National Institute for Medical Research, London, England
University of Edinburgh, Department of Chemistry, Scotland
(Received October 8IDecember 6, 1993) - EJB 93 1521/2
The solution structure of the deoxydecanucleotide [d(GAGTUACGA)] . [d(GAGTUACGA)] has been determined by NMR methods. This duplex, which contains six G . A mismatches and four Watson-Crick base pairs, is thermodynamically more stable than a decamer where T . A base pairs are substituted for the G . A mismatches, and is less stable than the duplex that contains G . C base pairs. Circular-dichroism spectroscopy indicates an overall B-like conformation for the decamer, but stronger than usual base staclung. 'H-NMR spectroscopy revealed that the N1H groups of the mismatched guanine residues are not hydrogen bonded, and 31P-NMR showed the presence of B,, phosphate conformations for the GpA steps. Detailed analysis of the NMR data showed that all nucleotides have anti glycosidic torsion angles and S type sugar puckers. The G . A mismatches pair in the amino form as originally proposed by Li et al. [Li, Y., Zon, G. & Wilson, W. D. (1991) Proc. Nut1 Acad. Sci. USA 88, 26-30], which results in extensive base-base stacking between the tandem G . A base pairs and their nearest neighbours. The terminal G . A base pairs are less stable than the central base pairs and show evidence of an equilibrium between two conformations, one involving B,, phosphate.
G . A mismatches vary extensively in their structure and this may explain why they frequently escape detection by DNA polymerase I11 (Fersht et al., 1982). A specific repair system has evolved to deal with these mismatches (Michaels et al., 1990). G . A base-pair conformation has been shown to depend on the base sequence around the mismatch site (Brown et al., 1990; Leonard et al., 1993) and several crystal structures of DNA duplexes containing G . A base pairs have been analysed in attempts to understand the nature of the influence of the surrounding bases. The G(anti) . A(syn) base pair has been found in [d(CGCGAATTAGCG)] . [d(CGC- - GAATTAGCG)] (Brown et al., 1986; Hunter et al., 1986) and in [d(CGCAAGCTGGCG)] . [d(CGCAAGCTGGCG)] at or above pH7 (Webster et al., 1990) and the G(syn) . AH+(anti) pair has been observed in crystals of [d(CGCA- AATTGGCG)] . [d(CGCAAATTGGCG)] at pH < 7 (Brown et al., 1989; Leonard et al., 1990a,b). The structures of G . A mismatches in solution have been investigated in a number of synthetic DNA duplexes. The G(anti) . A(anti) pair was found in the self-complementary duplex [d(CGAGAAT- TCGCG)] . [d(CGAGAATTCGCG)] (Patel et al., 1984) and in the [d(GCCACAAGCTC)] . [d(GAGCTGGTGGC)] du- plex (Carbonneaux et al:, 1991). The conversion of G(anti) . A(anti) to G(syn) . AH+(anti) at acidic pH has been demon- strated in the former sequence and the [d(GCCACAA- GCTC)] . [d(GCTGGTGGC)] duplex (Gao and Patel, 1988;
Correspondence to T. Brown, University of Edinburgh, Depart- ment of Chemistry, West Mains Road, Edinburgh, Scotland EH9 3JJ
Abbreviations. TOCSY, total correlation spectroscopy ; NOESY, nuclear-Overhauser-enhancement spectroscopy ; T,,,, melting temper- ature; C,, concentration of single strands.
Carbonnaux et al., 1991) and in [d(CGCAAATTGGCG)] . [d(CGCAAATTGGCG)] (Lane et al., 1991).
The structure of tandem 5' ... GA ... 3' mismatches has re- cently been investigated by several research groups. G(anti) . A(anti) tandem mismatches which adopt an imino-like base pairing (Fig. 1A) have been characterized in the [d(CCAA- ~ GATTGG)] . [d(CCAAGATTGG)] duplex, whereas G(anti) . A(anti) tandem mismatches of the amino type (Fig. 1B) have been found in the following sequences: [d(ATUGC- GA ATA)] . [ d( ATGAGCGA ATA)] ; [ d( GTGA ACTT)] . [ d( G- TEAACTT)] ; [d(CCAC&AGTGG)] . [d(CCACGAGTGG)] ; [d(GCGAATAAGCG)] . [d(GCGAATA&CG)] (Li et al., 1991; Chou et al., 1992a; Lane et al., 1992; Ebel et al., 1992; Maskos et al., 1993). These mismatches confer an unu- sually high stability on the duplex due to extensive cross- strand purine-purine base stacking between the mismatches and increased stacking of the 5' ... GpA ... 3' dimer with its nearest neighbours. 31P-NMR studies indicate an unusual backbone B,, geometry for the phosphates in the (GpA) block (Chou et al., 1992b). Investigations of a range of tandem G . A mismatches in different base-stacking environments have revealed that very stable amino pairs form when a tandem G . A mismatch is flanked by a pyrimidine base on the 5' side and a purine base on the 3' side (Ebel et al., 1992; Cheng et al., 1992).
Thermal melting studies of heteroduplexes have revealed that the multiple G . A mismatches are accomodated in a stacked helical structure (Dodgson and Wells, 1977). It ap- peared feasible to us that oligodeoxynucleotide sequences consisting largely or entirely of G . A mismatches might form stable duplexes and we now present the results of a

718
detailed investigation of the solution structure and thermal stability of multiple G . A mismatches in DNA.
MATERIALS AND METHODS Oligonucleotide synthesis
DNA phosphoramidite monomers and ancillary reagents were purchased from Applied Biosystems and RNA phos- phoramidites and ancillary reagents were obtained from Cruachem. Sephadex G-10 gel-filtration medium and pre- packed Sephadex G-25 (NAP) columns were purchased from Pharmacia. Purified water was obtained from a MilliRO/ MilliQ system (Millipore).
Oligonucleotide synthesis and purification Oligodeoxyribonucleotides were synthesised on an ABI
394 DNA synthesiser by the phosphoramidite method (Beau- cage and Caruthers, 1981) and purified by HPLC using a Brownlee aquapore reverse-phase octyl column (25 cm X 10 mm; Brown and Brown, 1991).
Ultraviolet melting studies Melting temperatures were determined in 100 mM NaC1,
10 mM NaH,PO,, 20 mM sodium cacodylate, 1 mM EDTA, pH 7.0, for all oligonucleotides except [d(GA),,], for which the sodium chloride concentration was increased to 1 .O M. Oligonucleotides were annealed slowly by cooling from 80 "C to 2 "C and absorbance/temperature melting curves were measured at 260 nm on a Perkin-Elmer Lambda 15 ul- traviolet spectrophotometer equipped with a Peltier block and controlled by an IBM PS2 microcomputer. A heating rate of 0.9"C . min-' was used and melting curves were measured in triplicate unless otherwise stated. Melting-curve data sets consisted of 300-530 data points. The data were processed using the PECSS2 software package (Perkin-Elmer). Con- centrations of single strands (C,) were determined spectro- photometrically at a temperature well above the T, (melting temperature; maximum point on the first derivative of the ultraviolet melting curve) and absorption coefficients were calculated (Puglisi and Tinoco, 1989). Thermodynamic parameters were calculated using the following equations (SantaLucia et al., 1991; Marky and Breslauer, 1987):
l/Tm = (R/AHD) . In(C,) + AS"/AHD,
dG"(298 K) = R T . In(C,) and AG'(298 K) = AW-TAP.
The data points were fitted to a van't Hoff plot of reciprocal melting temperature versus logarithm of the strand concen- tration (T,,-' versus In C,). All oligonucleotide duplexes in- vestigated were shown by the above relationship to form bimolecular duplexes.
NMR studies
The decamer d(GAGTGAACG4) and the dodecamer d(CGAGTGAACmG) were each dissolved in 1 ml 100 mM KCI, 10 mM NaHPO,, 0.1 mM EDTA, pH 7.0, and annealed slowly from 80°C. The samples were lyophilised and dis- solved in 0.5 ml 100% D,O. Phase-sensitive nuclear-Over- hauser-enhancement spectroscopy (NOESY) measurements were recorded at 10°C at 14.1 T on a Varian Unity spectrom- eter using the method of States et al. (1982) and at 20°C
at 11.75 T on a Bruker AM 500 spectrometer using time- proportional phase incrementation (Marion and Wuthrich, 1983). A total correlation spectroscopy (TOCSY) analysis was performed at 11.75 T on a Bruker AM 500 spectrometer using an MLEV17 mixing sequence (Bax and Davis, 1985). Spectra in 'H,O were recorded at 11.75 T on a Bruker AM 500 spectrometer, using a 1331 composite pulse (Hore, 1983). 31P-NMR spectra were recorded at 4.7 T on a Bruker WB200 spectrometer as previously described (Lane et al., 1992). '*P shifts were referenced to external methylene di- phosphonate.
Circular dichroism Circular-dichroism spectra were recorded at 15 "C in
500 mM KCl, 20 mM sodium phosphate, pH 7.0, with ap- proximately 40 pM duplex as previously described (Lane et al., 1992).
Molecular modelling Models were generated using the restrained-molecular-
dynamics and energy-minimisation routines within the pro- gram Discover (Biosym) using the Amber force field. The Coulomb term was attenuated using a dielectric constant equal to the distance. 1,4 interactions were scaled by 0.5. Restraints were taken from analysis of the NMR data and included 60 intra-residue constraints, 30 inter-residue con- straints and cross-strand constraints, and 14 hydrogen-bond constraints. To prevent excessive buckling, the bases of the terminal pairs were fixed during the calculations. Also, to prevent bending of the bases, a large mass was applied to the out-of-plane term in the force field for the initial energy- minimisation and molecular-dynamics runs. Only the central six base-pairs, [d(GTGAAC)] . [d(GTGAAC)], were used for these studies, owing to conformational averaging of the terminal two mismatched base pairs (see below). The initial model was standard B-DNA, modified with the G . A mis- matches. 2000 steps of conjugate-gradient minimisation were run, followed by 10 ps of restrained molecular dynamics at 1200 K, with electrostatics, torsion potentials and non-bond interactions scaled to 0.1. Further cycles of energy minimisa- tion and restrained molecular dynamics were performed at 600 K and 300 K, with the scaling parameters increased to 0.3, 0.6 and finally 1.0. The final molecular-dynamics run at 300 K gave negative energies and no significant overall changes over a period of 15 ps. This structure was energy minimised with 2500 cycles of conjugate-gradient refine- ment, producing a final energy of -850 kJ . moV-' and no constraint violations.
RESULTS AND DISCUSSION Oligonucleotides containing multiple G - A mismatches
The decamer d('GAGTGAACGA'O) forms a self-com- plementary duplex of the type [d(GAGTGAACGA)] . [ d ( w G T m A C m ) ] containing six tandem G . A mis- matches and four standard Watson-Crick base pairs. It has been shown that the amino mode of G . A base pairing (Fig. 1B) is favoured in sequences where the mismatches are flanked by a pyrimidine (pyr) on the 5' side and a purine (pur) on the 3' side (Ebel et al., 1992; Lane et al., 1992). Hence, the tandem G . A mismatches in this case are ex- pected to be amino paired since the base stacking environ-
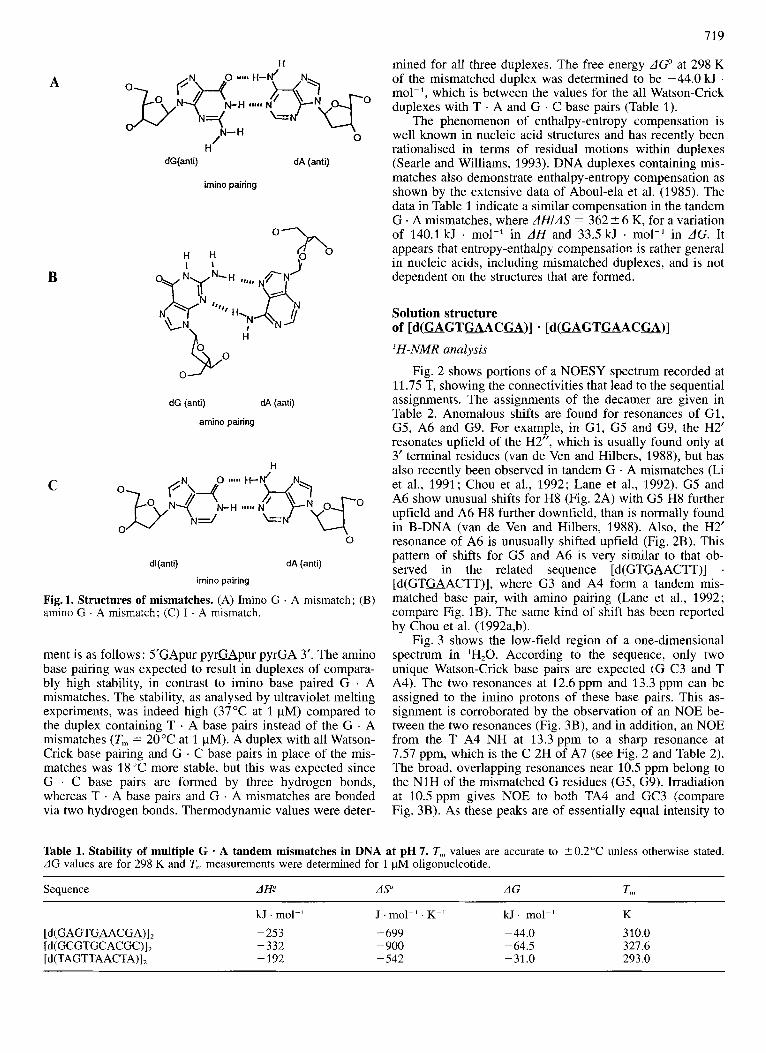
719
A
B
C
H
dG(anti) dA (anti)
imino pairing
dG (anti) dA (anti)
amino pairing
H
dl(anti) dA (anti)
imino pairing
Fig. 1. Structures of mismatches. (A) Imino G . A mismatch; (B) amino G . A mismatch; (C) I . A mismatch.
ment is as follows : 5'GApur py rGpur pyrKA 3'. The amino base pairing was expected to result in duplexes of compara- bly high stability, in contrast to imino base paired G . A mismatches. The stability, as analysed by ultraviolet melting experiments, was indeed high (37°C at 1 pM) compared to the duplex containing T . A base pairs instead of the G . A mismatches (Tn, = 20°C at 1 pM). A duplex with all Watson- Crick base pairing and G . C base pairs in place of the mis- matches was 18 "C more stable, but this was expected since G . C base pairs are formed by three hydrogen bonds, whereas T . A base pairs and G . A mismatches are bonded via two hydrogen bonds. Thermodynamic values were deter-
mined for all three duplexes. The free energy AGO at 298 K of the mismatched duplex was determined to be -44.0 kJ . mol-', which is between the values for the all Watson-Crick duplexes with T . A and G . C base pairs (Table 1).
The phenomenon of enthalpy-entropy compensation is well known in nucleic acid structures and has recently been rationalised in terms of residual motions within duplexes (Searle and Williams, 1993). DNA duplexes containing mis- matches also demonstrate enthalpy-entropy compensation as shown by the extensive data of Aboul-ela et al. (1985). The data in Table 1 indicate a similar compensation in the tandem G . A mismatches, where AH/AS = 362 2 6 K, for a variation of 140.1 kJ . mol-' in AH and 33.5 kJ . mol-' in A G . It appears that entropy-enthalpy compensation is rather general in nucleic acids, including mismatched duplexes, and is not dependent on the structures that are formed.
Solution structure of [d(GAGT=ACGA)] - [d(GAGTGAACM)] 'H-NMR analysis
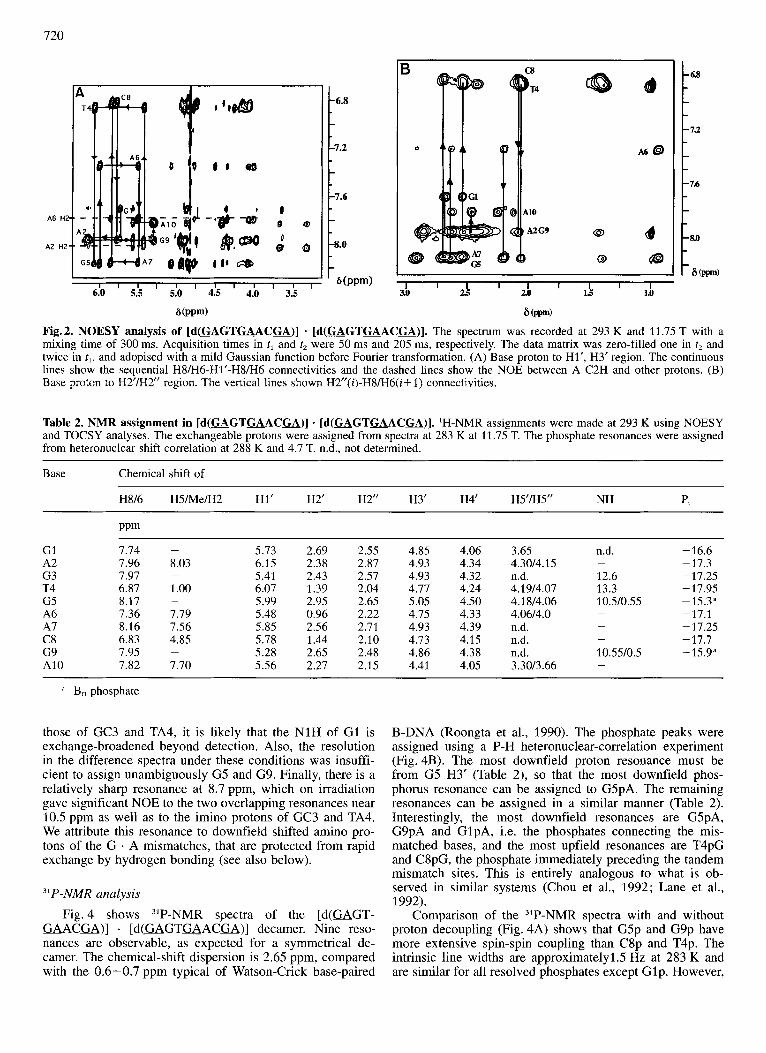
Fig. 2 shows portions of a NOESY spectrum recorded at 11.75 T, showing the connectivities that lead to the sequential assignments. The assignments of the decamer are given in Table 2. Anomalous shifts are found for resonances of G1, G5, A6 and G9. For example, in G1, G5 and G9, the H2' resonates upfield of the H2", which is usually found only at 3' terminal residues (van de Ven and Hilbers, 1988), but has also recently been observed in tandem G . A mismatches (Li et al., 1991 ; Chou et al., 1992; Lane et al., 1992). G5 and A6 show unusual shifts for H8 (Fig. 2A) with G5 H8 further upfield and A6 H8 further downfield, than is normally found in B-DNA (van de Ven and Hilbers, 1988). Also, the H2' resonance of A6 is unusually shifted upfield (Fig. 2B). This pattern of shifts for G5 and A6 is very similar to that ob- served in the related sequence [d(GTmACTT)] . [d(GTUACTT)], where G3 and A4 form a tandem mis- matched base pair, with amino pairing (Lane et al., 1992; compare Fig. 1B). The same kind of shift has been reported by Chou et al. (1992a,b).
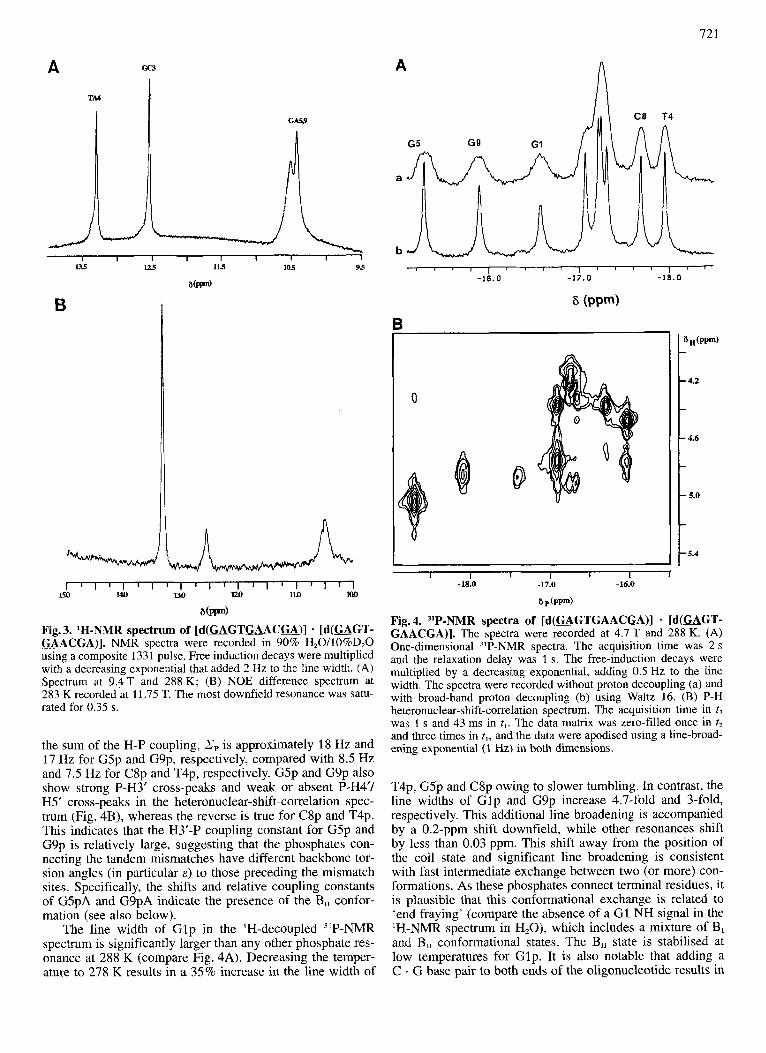
Fig. 3 shows the low-field region of a one-dimensional spectrum in 'H,O. According to the sequence, only two unique Watson-Crick base pairs are expected (G C3 and T A4). The two resonances at 12.6 ppm and 13.3 ppm can be assigned to the imino protons of these base pairs. This as- signment is corroborated by the observation of an NOE be- tween the two resonances (Fig. 3B), and in addition, an NOE from the T A4 NH at 13.3ppm to a sharp resonance at 7.57 ppm, which is the C 2H of A7 (see Fig. 2 and Table 2). The broad, overlapping resonances near 10.5 ppm belong to the N1H of the mismatched G residues (G5, G9). Irradiation at 10.5 ppm gives NOE to both TA4 and GC3 (compare Fig. 3B). As these peaks are of essentially equal intensity to
Table 1. Stability of multiple G - A tandem mismatches in DNA at pH 7. T, values are accurate to ? 0.2"C unless otherwise stated. AG values are for 298 K and T,, measurements were determined for 1 pM oligonucleotide.
Sequence AH: AS" AG
[ d ( c A G T ~ A C c A ) ] , [d(GCGTGCACGC)], [d(TAGTTAACTA)],
kJ . mol-' J . mol-' . K-' kJ. mol-' K -253 -332 -192
- 699 - 900 -542
-44.0 -64.5 -31.0
310.0 327.6 293.0
720
A6 HZ
A2 H2'
I 1 1 1 I I I I I I I I
6.0 5.5 I 5.0 4.5 4.0 3.5
G(ppm)
6.8
7.2
7.6
8.0
b(PPm) I I I I I 1 I I I 31) 25 20 15 1.0
Fig.2. NOESY analysis of [d(MGTGAACGA)] - [ d ( G A G T a A C u ) ] . The spectrum was recorded at 293 K and 11.75 T with a mixing time of 300 ms. Acquisition times in tl and t2 were 50 ms and 205 ms, respectively. The data matrix was zero-filled one in t2 and twice in tl, and adopised with a mild Gaussian function before Fourier transformation. (A) Base proton to Hl', H3' region. The continuous lines show the sequential H8/H6-H1'-H8/H6 connectivities and the dashed lines show the NOE between A C2H and other protons. (B) Base proton to H2'lH2" region. The vertical lines shown H2"(i)-H8/'H6(i+ 1) connectivities.
Table 2. NMR assignment in [d(WGTGAAC=)] - [d(WGTGAACa)]. 'H-NMR assignments were made at 293 K using NOESY and TOCSY analyses. The exchangeable protons were assigned from spectra at 283 K at 11.75 T. The phosphate resonances were assigned from heteronuclear shift correlation at 288 K and 4.7 T. n.d., not determined.
Base Chemical shift of
H816 H5A4eIH2 HI' H2' H2" H3' H4' HS'M5" NH p,
GI A2 G3 T4 G5 A6 A7 C8 G9 A10
7.74 -
7.96 8.03 7.97 - 6.87 1.00 8.17 -
7.36 7.79 8.16 7.56 6.83 4.85 7.95 -
7.82 7.70
5.73 2.69 2.55 4.85 4.06 6.15 2.38 2.87 4.93 4.34 5.41 2.43 2.57 4.93 4.32 6.07 1.39 2.04 4.77 4.24 5.99 2.95 2.65 5.05 4.50 5.48 0.96 2.22 4.7.5 4.33 5.85 2.56 2.71 4.93 4.39 5.78 1.44 2.10 4.73 4.15 5.28 2.65 2.48 4.86 4.38 5.56 2.27 2.15 4.41 4.05
3.65 4.3014.15 n.d. 4.1914.07 4.1 814.06 4.0614.0 n.d. n.d. n.d. 3.3013.66
n.d.
12.6 13.3 10.510.55
-
-
10.55l0.5 -
-16.6 -17.3 -17.25 -17.95 -15.3" -17.1 - 17.25 -17.7 -15.9"
a B,, phosphate
those of GC3 and TA4, it is likely that the N1H of G1 is exchange-broadened beyond detection. Also, the resolution in the difference spectra under these conditions was insuffi- cient to assign unambiguously G5 and G9. Finally, there is a relatively sharp resonance at 8.7 ppm, which on irradiation gave significant NOE to the two overlapping resonances near 10.5 ppm as well as to the imino protons of GC3 and TA4. We attribute this resonance to downfield shifted amino pro- tons of the G . A mismatches, that are protected from rapid exchange by hydrogen bonding (see also below).
"P-NMR analysis Fig. 4 shows "P-NMR spectra of the [d(GAGT-
GAACGA)] . [d (GAGTGACG)] decamer. Nine reso- nances are observable, as expected for a symmetrical de- camer. The chemical-shift dispersion is 2.65 ppm, compared with the 0.6-0.7 ppm typical of Watson-Crick base-paired
B-DNA (Roongta et al., 1990). The phosphate peaks were assigned using a P-H heteronuclear-correlation experiment (Fig. 4B). The most downfield proton resonance must be from G5 H3' (Table 2), so that the most downfield phos- phorus resonance can be assigned to G5pA. The remaining resonances can be assigned in a similar manner (Table 2). Interestingly, the most downfield resonances are GSpA, G9pA and GlpA, i.e. the phosphates connecting the mis- matched bases, and the most upfield resonances are T4pG and C8pG, the phosphate immediately preceding the tandem mismatch sites. This is entirely analogous to what is ob- served in similar systems (Chou et al., 1992; Lane et al., 1992).
Comparison of the 3'P-NMR spectra with and without proton decoupling (Fig. 4A) shows that G5p and G9p have more extensive spin-spin coupling than C8p and T4p. The intrinsic line widths are approximatelyl.5 Hz at 283 K and are similar for all resolved phosphates except Glp. However,
A Gc3
TA4
A
721
I I I I I I I I 1 l3.5 125 115 105 95
B
l ~ l ~ l ~ ~ ~ l ~ l ~ l ~ l i l J l l l
S W )
15a 140 UD m 1Ul lIM
Fig. 3. 'H-NMR spectrum of [d(GAGTGAAC=)] - [d(=GT- U A C U ) ] . NMR spectra were recorded in 90% H20/10%D20 using a composite 1331 pulse. Free induction decays were multiplied with a decreasing exponential that added 2 Hz to the line width. (A) Spectrum at 9.4 T and 288 K; (B) NOE difference spectrum at 283 K recorded at 11.75 T. The most downfield resonance was satu- rated for 0.35 s.
the sum of the H-P coupling, Zp is approximately 18 Hz and 17 Hz for G5p and G9p, respectively, compared with 8.5 Hz and 7.5 Hz for C8p and T4p, respectively. G5p and G9p also show strong P-H3' cross-peaks and weak or absent P-H4'/ H5' cross-peaks in the heteronuclear-shift-correlation spec- trum (Fig. 4B), whereas the reverse is true for C8p and T4p. This indicates that the H3'-P coupling constant for G5p and G9p is relatively large, suggesting that the phosphates con- necting the tandem mismatches have different backbone tor- sion angles (in particular E ) to those preceding the mismatch sites. Specifically, the shifts and relative coupling constants of G5pA and G9pA indicate the presence of the B,, confor- mation (see also below).
The line width of G lp in the 'H-decoupled 31P-NMR spectrum is significantly larger than any other phosphate res- onance at 288 K (compare Fig. 4A). Decreasing the temper- ature to 278 K results in a 35% increase in the line width of
1 1 ' ' " 1 ' ' ' ' ~ ' '
-16.0 -17 .0 -18.0
6 (PPW
.4.2
.46
. 5.0
.5.4
I I 1 I L I t -18.0 -17.0 -16.0
6 p ( P e w
Fig. 4. "P-NMR spectra of [d(=GTGAACGA)] - [d(GAGT- GAACGA)]. The spectra were recorded at 4.7 T and 288 K. (A) One-dimensional 3'P-NMR spectra. The acquisition time was 2 s and the relaxation delay was 1 s . The free-induction decays were multiplied by a decreasing exponential, adding 0.5 Hz to the line width. The spectra were recorded without proton decoupling (a) and with broad-band proton decoupling (b) using Waltz 16. (B) P-H heteronuclear-shift-correlation spectrum. The acquisition time in t2 was 1 s and 43 ms in tl. The data matrix was zero-filled once in t2 and three times in t,, and the data were apodised using a line-broad- ening exponential (1 Hz) in both dimensions.
T4p, G5p and C8p owing to slower tumbling. In contrast, the line widths of Glp and G9p increase 4.7-fold and 3-fold, respectively. This additional line broadening is accompanied by a 0.2-ppm shift downfield, while other resonances shift by less than 0.03 ppm. This shift away from the position of the coil state and significant line broadening is consistent with fast intermediate exchange between two (or more) con- formations. As these phosphates connect terminal residues, it is plausible that this conformational exchange is related to 'end fraying' (compare the absence of a G1 NH signal in the 'H-NMR spectrum in H,O), which includes a mixture of B, and B,, conformational states. The B,, state is stabilised at low temperatures for Glp. It is also notable that adding a C . G base pair to both ends of the oligonucleotide results in
722
B
240 260 280 300
Wavelength (nm) Wavelength (nm)
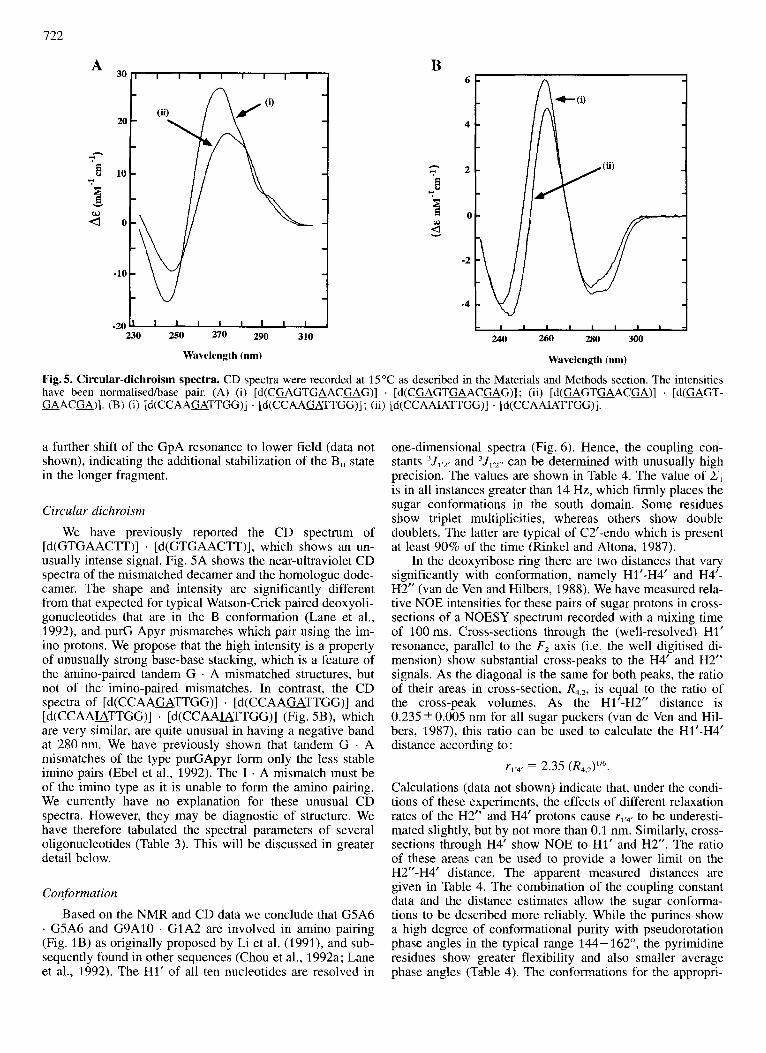
Fig. 5. Circular-dichroism spectra. CD spectra were recorded at 15 “C as described in the Materials and Methods section. The intensities have been normalisedlbase pair. (A) (i) [d(C=GTaACGAG)] . [ d ( C a G T u A C m G ) ] ; (ii) [ d ( G A G T m A C m ) ] . [d(=GT- GAACU)] . (B) (i) [d(CCAAaTTGG)] . [d(CCAAGATTGG)] ; (ii) [d(CCAA&TTGG)] . [d(CCAA&TTGG)].
a further shift of the GpA resonance to lower field (data not shown), indicating the additional stabilization of the B,, state in the longer fragment.
Circular dichroism
We have previously reported the CD spectrum of [d(GTGAACTT)] . [d(GTGAACTT)], which shows an un- usually intense signal. Fig. SA shows the near-ultraviolet CD spectra of the mismatched decamer and the homologue dode- camer. The shape and intensity are significantly different from that expected for typical Watson-Crick paired deoxyoli- gonucleotides that are in the B conformation (Lane et al., 1992), and purC Apyr mismatches which pair using the im- ino protons. We propose that the high intensity is a property of unusually strong base-base stacking, which is a feature of the amino-paired tandem G . A mismatched structures, but not of the. imino-paired mismatches. In contrast, the CD spectra of [d(CCAAGATTGG)] . [d(CCAAGATTGG)] and [d(CCAAUTTGG)] . [d(CCAAUTTGG)] (Fig. SB), which are very similar, are quite unusual in having a negative band at 280nm. We have previously shown that tandem G . A mismatches of the type purGApyr form only the less stable imino pairs (Ebel et al., 1992). The I . A mismatch must be of the imino type as it is unable to form the amino pairing. We currently have no explanation for these unusual CD spectra. However, they may be diagnostic of structure. We have therefore tabulated the spectral parameters of several oligonucleotides (Table 3). This will be discussed in greater detail below.
Conformation
Based on the NMR and CD data we conclude that GSA6 . GSA6 and G9A10 . G1A2 are involved in amino pairing (Fig. 1B) as originally proposed by Li et al. (1991), and sub- sequently found in other sequences (Chou et al., 1992a; Lane et al., 1992). The H1’ of all ten nucleotides are resolved in
one-dimensional spectra (Fig. 6). Hence, the coupling con- stants ’.I,,,. and 351.2” can be determined with unusually high precision. The values are shown in Table 4. The value of 2, is in all instances greater than 14 Hz, which firmly places the sugar conformations in the south domain. Some residues show triplet multiplicities, whereas others show double doublets. The latter are typical of C2’-endo which is present at least 90% of the time (Rinkel and Altona, 1987).
In the deoxyribose ring there are two distances that vary significantly with conformation, namely Hl’-H4’ and H4’- H2” (van de Ven and Hilbers, 1988). We have measured rela- tive NOE intensities for these pairs of sugar protons in cross- sections of a NOESY spectrum recorded with a mixing time of 100 ms. Cross-sections through the (well-resolved) H1‘ resonance, parallel to the Fz axis (i.e. the well digitised di- mension) show substantial cross-peaks to the H4’ and H2” signals. As the diagonal is the same for both peaks, the ratio of their areas in cross-section, R4,z, is equal to the ratio of the cross-peak volumes. As the Hl’-H2” distance is 0.235 2 0.005 nm for all sugar puckers (van de Ven and Hil- bers, 1987), this ratio can be used to calculate the Hl’-H4‘ distance according to:
r1.4, = 2.35 (R4,2)1’6.
Calculations (data not shown) indicate that, under the condi- tions of these experiments, the effects of different relaxation rates of the H2” and H4’ protons cause r,.< to be underesti- mated slightly, but by not more than 0.1 nm. Similarly, cross- sections through H4’ show NOE to H1’ and H2”. The ratio of these areas can be used to provide a lower limit on the H2”-H4’ distance. The apparent measured distances are given in Table 4. The combination of the coupling constant data and the distance estimates allow the sugar conforma- tions to be described more reliably. While the purines show a high degree of conformational purity with pseudorotation phase angles in the typical range 144-162”, the pyrimidine residues show greater flexibility and also smaller average phase angles (Table 4). The conformations for the appropri-

723
Table 3. Circular-dichroism analysis of oligonucleotides. CD spectra not from previous studies were recorded at 15°C for 300-400 pM base pairs in 500 mM KCl, 20 mM sodium phosphate, pH 7, using a 2-mm path-length cuvette. Four scans were added together in the computer memory and the solvent baseline was subtracted. The helix type is taken from the literature. LZW, Li, Zon, Wilson base pairing; PD, PrivC-Dickerson base pairing. The CD intensity has been normalisedhase pair.
Sequence A,,, & Helix Reference type
nm &-' . cm-'
B-DNA 278 245 -
Ivanov et al. (1974) 5 B 6
A-RNA 260 14.6 A 240 - 1 210 -14
247 -10
250 - 7.8
250 - 7.7
[d(GTGAACTT)] . [d(GTGAACTT)] 275 17.5 BLZW Lane et al. (1992)
Lane et al. (1992)
Lane et al. (1 992)
[d(CGCAAATTGGCG)] . [d(CGCAAATTGGCG)J (PH 5.0, G (~yit)] 280 2.1 B
[d(CGCAAATTGGCG)J . [d(CGCAAATTGGCG)J [PH 7.5, G (anti)] 282 3.1 B
[d(CCAAGATTGG)] . [d(CCAAmTTGG)]
[d(CCAAUTTGG)] . [d(CCAAUTTGG)]
[d(GAGTGAACGA)] . [d(GAGTGAACGA)]
[d(CGAGTGAACGAG)] . [d(CGAGTGAACGG)]
280 - 3.5 BPD this study 260 5 245 - 5 280 - 3.2 BED this study 260 5 245 - 5 270 17 BLZW this study 245 - 9 267 26 243 -15
BLZW this study
A[ A6
GI
I
/ " " 1 ' ~ ~ ~ l ' " ' I ' " ' I ' ' " / " " 1 " ' 1 ' ~ ' ' 1 ' ' ' ' I ' ' ' ' 6 . 2 6.1 6 . 0 5 . 9 5.8 5.7 5 . 6 5 . 5 5 . 4 5.3
6 (PPm)
Fig.6. 'H-NMR spectrum of the H1' region of [d(GAGTGAACGA)] * [d(GAGTuACGA)]. The spectrum was recorded at 14.1 T and 298 K with a digital resolution of 0.36 Hz/point. The free-induction decay was multiplied by a mild Gaussian function prior to Fourier transformation.
ate sugars derived for the present sequence agree well with those determined for the smaller analogue [d(GTWACTT)I2 (Lane et a]., 1992). Table 4 also indicates that A10 is subject to considerable averaging of the sugar conformation, which corroborates the 31P-NMR data (see above).
Analysis of the NOE intensities, in combination with the coupling constant data and using the program NUCFIT (Lane, 1990), showed that the glycosidic torsion angles are all in the range 110-125", i.e. the normal anti range found in deoxyoligonucleotides. The relatively detailed information about the nucleotide conformations can be used for deriving more tightly constrained distances and torsion angles for re- strained molecular-dynamics calculations (see below).
The internucleotide NOE intensities for the step G3T4/ A7C8 are typical for right-handed B-DNA. In contrast, the NOE intensities for the GpA steps are significantly different. Further, there are other NOE resonances not usually observed in Watson-Crick base-paired B-DNA. These include strong cross-strand NOE A2 H2 to A10 H2, A10 H', and H10H4' and the analogous ones for A6 H4' , A6 H5'/H5", and A6 Hl'. These latter NOE can be assigned to the cross-strand interaction because the intraresidue NOE are very small. For A6 with a C2'-endo sugar conformation and anti glycosidic torsion angle, the minimum distance between H2 and the H1' of the same nucleotide is 0.46 nm, which gives an NOE intensity at least an order of magnitude smaller than observed
724
Table 4. Spin-spin couplings and proton-proton distances within deoxyriboses. The coupling constants were determined from resolution- enhanced one-dimensional NMR spectra. Distances were determined from cross-sections parallel to Fz in the NOESY experiment recorded with a mixing time of 100 ms as described in the Materials and Methods section. Ps is the pseudosymmetric phase angle of the dominant (S) confomer andfs i s the fraction of that confomer assuming a two-state N-S equilibrium.
Base NMR parameter
XI, 3J1~2 3J,,2 TI ry,. Ps f s
Hz nm degrees
3.1 3.3 162 0.9 G1 15 S 0.75 A2 14.2
G3 15.1 9.8 5.3 2.9 3.2 126 0.9 T4 15.1 10.1 5.0 2.9 3.5 144 0.90 G5 15.8 10.4 5.4 3.0 3.4 162 1 .o
162 0.79 A6 14.4 162 0.95 A7 15.4
C8 15.5 10.2 5.3 2.7 3.0 126 0.95 G9 15.0 10.3 4.7 3.0 162 0.89 A10 13
- -
- - - -
- 3.2 3.1
- -
- - -
-
162 0.6 - 3.3 - -
A B
Fig. 7. Structure of [d(GAGTUACGA)] * [d(GAGTUACU)]. The structure was generated using NUCFIT as described in the text. This structure (A) is idealised regarding the flexibility of the termi- nal base-pairs. The structure of standard B-DNA is shown for com- parison (B).
in the NOESY spectrum, and in one-dimensional NOE ex- periments performed with a longer recycle time. We con- clude that at least 90% of the NOE intensity arises from the cross-strand interaction, just as for the A2 H2 to A10 H1' NOE. These cross-strand interactions are analogous to those observed by Chou et al. (1992a).
Molecular modelling
Initial models of the decamer were built using the facili- ties with NUCFIT, which has been modified to incorporate arbitrary numbers of mismatched base pairs as previously described (Lane et al., 1992). For these purposes, the sugars were set to unique' conformations with P, = 162" for the purines and 144" or 126" for the pyrimidines (compare Table 4). This amounts to modelling the single most probable structure. The glycosidic torsion angles were set to -110" for the purine and -120" for the pyrimidines. The helical rise was set to 0.34 nm, base-pair inclinations to zero and base-pair roll angles to zero. The tandem mismatches were incorporated into the helix by appropriate translations (Dx and Dy) and rotations (helical twist), as described by Lane et al. (1992), so as to produce a structure that is symmetric
Table 5. Additional NOE for G5 A6 and G7 for the tandem mismatch in [ d ( U G T U A C M ) ] - [d(GAGTGAACGA)]. NOE were identified in NOESY spectra and in one-dimensional NOE analyses. Other intranucleotide and internucleotide NOE were iden- tified and treated as described in the text. w, weak; m, medium; s , strong.
NOE
from to Strength r (upper)
G5 H8 T4 H1' T4 H2' T4 H2" T4 H3'
A6 H8 G5 H1' G5 H2"
A7 H8 A6 HI' A6 H2' A6 H2" A6 H3'
A6 H2 A7 H2 A7 HI' A6' HI' A6' H4' A6' H5' A6' H5"
W
m S W
m W
m m m m
W W
S S W W
5 4 3 5 4 5 4 4 4 4 5 5 3 3 5 5
with respect to the two strands, has a favourable Lennard- Jones energy, allows the phosphate backbone to be placed and accounts qualitatively for the observed NOE. The helical twist was defined for these purposes through C8 (C6)-Nl/ N3H as in a Watson-Crick base pair. However, if the twists are defined between the axes formed by successive Cl'-Cl' for each base pair, the initial model has values of 68, 21, 36 and 23" for the first four steps (and the same again by sym- metry for the remainder). These values are very similar to those obtained by Chou et al. (1992a). This initial structure gave a Lennard-Jones energy of -705 kJ . mol-', compared with a standard B-DNA duplex of -868kJ . mol-'. The C3'(i)-C5'(i+ 1) distances were within reasonable limits such that the phosphate group can adopt stereochemically reason- able positions. Improved energies were obtained by allowing

725
A
B
Fig.8. Structure of the [d (GTuAC)] - [d(GTuAC)] . The structure was generated by restrained molecular dynamics and en- ergy minimisation as described in the text. (A) The view of the major groove showing the buckling of the mismatched base pairs. (B) The base-pair overlaps: T2 . A5/G3 . A4 (left); G3 . A41A4 . G3 (middle); A4 . G3/A5 . T2 (right).
the mismatched G . A base pairs to adopt a large propeller twist (compare Chou et al., 1992a); using a propellor twist of -20" for both of the mismatched G . A base pairs decreases the energy to -750 kJ . mol-'. This initial structure predicts relatively strong cross strand NOE from A H2 to A H2 and Hl'. However, the cross strand NOE H2-H4' is predicted to be weak (Y value approximately 0.5 nm). This NOE becomes greater by allowing the base pairs to tilt away from the per- pendicular. All the other NOE are also qualitatively ac- counted for by this model, whereas a structure built with mismatched base pairs having the imino pairing (compare PrivC et al., 1987) does not. Quantitative agreement with the experimental data is not expected because of the conforma- tional exchange affecting the first two and last two base pairs (see above), and because the structure is idealised. Fig. 7 shows the decamer having the amino-paired mismatches in comparison with standard B-DNA.
A second set of modelling experiments was performed using the Discover package on the hexanucleotide G T W A C
. G T W A C (which corresponds to the central six base pairs of the decamer) with the Watson-Crick paired bases fixed as described in the Materials and Methods section. Distance constraints were applied for the nucleotide units according to the NOE data described above. Only the central hexamer was treated because of the conformational averaging of the terminal mismatches (see above) which cannot be treated ad- equately by molecular-dynamics calculations. Additional in- ter-residue constraints were obtained from analysis of the NOESY spectra, with the aid of the initial models built with NUCFIT. Also, one-dimensional NOE time courses were used to obtain more precise estimates on critical cross-strand interactions from A H2. Nevertheless, as the nucleotide units themselves are averaging, very narrow distance limits were not used (Table 5). The Discover calculation serves to pro- vide a family of related solutions that are stereochemically reasonable. Internucleotide distances derived from NOE data as given in Table 5 were used. After several rounds of conju- gate-gradient energy minimisation and molecular-dynamics runs, a stable structure was obtained in which the root-mean- square energy gradient was less than 0.1 kJ . mol-' and the mean energy was -1472 W . mol-I. The final energy-mini- mised structure appeared symmetric on analysis of the nucle- otide conformations and distances between critical protons. The mean glycosidic torsion angle was -117t7". The gly- cosidic torsion angle for A4 was -121", compared with -108" for G3. The backbone angle 6 was on average 139 ? 8", which is typical of the range Cl'-exo-C2'-endo.
The conformations of the mismatched base pairs are rela- tively well determined owing to the presence of numerous cross-strand NOES. Fig. 8A shows a view of the central hex- amer in which the bases in each mispair incline in opposite directions (i.e. buckle). The phosphodiesters connecting the two base pairs are in the B,, conformation. The relative base- base stacking of the mismatched base-pairs is shown in Fig. 8B. As previously reported (Lane et al., 1992), the pu- rine residues of the mismatch stack extensively on the purine residues of the neighbouring mismatch on the opposite strand, forming a well-stacked block of two base pairs (mid- dle structure). Probably of equal importance concerning the overall thermodynamic stability of such tandem mismatches is the interaction of the tandem G . A block with the 5' and 3' neighbouring base pairs. As the left and right structures of Fig. 8B show, the tandem G . A block stacks extensively with both neighbours, considerably more so than the stacking of Watson-Crick base pairs in B-DNA.
The 31P-NMR spectrum indicates the presence of the B,, conformation for the GpA step (Table 6). Although explicit
Table 6. NMR parameters for oligonucleotides containing G * X mismatches, NH type, hydrogen-bonding mode of G N1H; A"(P,), total chemical shift dispersion; 'Gp, phosphate chemical shift for G3'p; P type phosphodiester conformation at mismatch site.
Molecule 'GNlH NH type 'Gp A&P,) P type Reference
PPm PPm
[d(GTaACTT)] . [d(GTaACTT)] 10.9 NHO -15.8 2.7 B ti Lane et al. (1992) [d(GAGTGAACGA)] . [d(GAGTGAACGA)] 10.5110.6 NHO -15.31 2.65 BIl this study
[d(CGCAAATTGGCG)] [d(CGCA_AATTGGCG)] ll.O(syrz) NHO >-17 0.6 BI Lane et al. (1991)
[d(CGCGAATTGGCG)] . [d(CGCGAATTGGCG)] 10.8 NHO -16.9 0.69 B," Borden et al. (1992) [d( AT-GC-ATA)] . [d( AT-GCaATA)] 10.3 NHO 2.8 Bii Li et al. (1991);
- 15.9
12.2(unti) NHN > - 17.3 0.6 B,
Chou et al. (1992)
* May also have B,, structure at low temperature.

726
constraints on the backbone angles E and 5 were not used in the modelling, the large conformational variation at the mismatch sites is expected to require rearrangement in the backbone. We found that in d(GTWAC),, a that Glp, T2p, A4p and A5p E and 5 were -170-+10” and -100-+21”, respectively, which are typical of the B, conformation. In contrast, E and < for G3p were -85” and -200°, respective- ly, which are similar to the values reported by PrivC et al. (1987) for phosphate in the B,, onformation and by Chou et al. (1992a,b) in a tandem G . A mismatched oligonucleotide.
Conclusions We have demonstrated that tandem G . A mismatches are
thermodynamically stable in the contexts pyrGApur, and also occur at the ends of duplexes such as 5’GApur and pyrGA3’. Even a decamer, containing six G . A mismatches forms a stable duplex. Based on the results presented in this study and elsewhere, we can state the probable structural properties of such mismatches and their signatures in terms of ultravio- let melting, circular dichroism and NMR spectroscopy. An overview of the characteristics of G . A mismatches in dif- ferent oligocleoxynucleotides is given in Tables 3 and 6. The amino-type G . A mismatch has essentially identical thermodynamic stability to a Watson-Crick base-paired T . A analogue in certain environments and the stability decreases markedly on substitution of inosine for guanine, which indi- cates the involvement of the G2 NH, in the base pairing. The phosphate backbone is in the B,, conformation connecting the tandem GpA and is B, elsewhere in the structure. This is manifest in a strongly downfield-shifted phosphate resonance and also by an upfield-shifted resonance which arises from the preceding phosphate (i.e. pGA).
The hydrogen bonding pattern of G . A mismatches in [d(GAGTGAACGA)] . d[GAGTGAACGA)] has been ana- lysed by ‘H-NMR. The N1H of the mismatched guanine is not involved in hydrogen bonding to DNA, only to water, and therefore resonates 1-2 ppm upfield of the Watson- Crick hydrogen-bonded G N1H. In addition, hydrogen bond- ing between the guanine amino group and A N7 would be expected to cause a downfield shift of the amino resonance and diminish the rotation about the CN bond. This would account for the observation of amino protons resonating near 8.7 ppm.
Uncommon base-stacking arrangements are manifest in unusual shifts of some proton resonances. The H2’ resonates downfield of the H2” in the guanine residue, and the H2’ of the adenine shifts upfield by approximately 2 ppm. The H8 of the mismatched guanine shifts downfield and the H8 of the mismatched adenine shifts upfield (approximately 0.5 ppm). Further, strong cross-strand NOE are observed be- tween the A H2 on one strand and the H4’ and H5/H5” of the other mismatched adenine residue on the complementary strand, showing that the two purine residues stack on one another. Both mismatched nucleotides are in the anti confor- mation and have sugar puckers in the S domain. The circular- dichroism spectra are non-conservative, with high intensity compared with ordinary B-form DNA, and extrema at 270- 275 nm (maximum) and 245 nm (minimum). This increased intensity is probably related to the much increased base stacking that occurs in the amino-paired tandem G . A mis- matches, not only between the tandem base pairs, but also with their nearest neighbours. It is notable that the intensity of the CD of the decamer containing six mismatches greatly increases on adding a base at each end. Apart from stabilising
the fraying, it also forces the terminal G . A mismatches to adopt the amino-base pairing fully (see above) and therefore increases the overall stacking. However, the presence of a 5’ base pair is not essential for the formation of this structure. Similarly, removal of the 3’ base pair greatly destabilises this structure.
This study was supported by the UK Medical Research Council and the Molecular Recognition Initiative of the UK Science and Engineering Research Council.
REFERENCES Aboul-ela, F., Koh, D., Tinoco, I. & Martin, F. H. (1985) Base-
base mismatches. Thermodynamics of double helix formation for d(CA,XA,G) and d(CT,YT,G) (X, Y = A, C, G, T), Nucleic Acids Res. 13, 4811 -4824.
Bax, A. & Davis, D. G. (1985) Mlev-17-based two-dimensional ho- monuclear magnetization transfer spectroscopy, J. Magn. Reson. 65, 355-360.
Beaucage, S. L. & Caruthers, M. H. (1981) Deoxynucleotide phos- phoramidites. A new class of key intermediates for deoxypoly- nucleotide synthesis, Tetrahedron Lett. 22, 1859- 1862.
Beijer, B., Sulston, I., Sproat, B. S., Rider, P., Lamond, A. & Neuner, P. (1990) Synthesis and applications of oligoribonucleotides with selected 2’-0-[1-(2-fluorophenyl)-4-methoxypiperidin-4-yl] pro- tecting group, Nucleic Acids Res. 17, 5143-5151.
Borden, K. L. B., Jenkins, T. C., Skelly, J. V., Brown, T. & Lane, A. N. (1992) Conformational properties of the G . G mismatch in [d(CGCGAATTGGCG)] . [d(CGCGAATTGGCG)] deter- mined by NMR, Biochemistry 31, 5411 -5422.
Brown, T., Leonard, G. A., Booth, E. D. & Kneale, G. (1990) Influ- ence of pH on the conformation and stability of mismatch base- pairs in DNA, J. Mol. Biol. 212, 437-440.
Brown, T. & Brown, D. J. S. (1991) Modem machine-aided methods of solid-phase oligonucleotide synthesis, in Oligonucleotides and analogues, a practical approach (Eckstein, F., ed.) pp. 1-23, IRL Press.
Carbonneaux, C., van der Marel, G. A., van Boom, J. H., Guschl- bauer, W. & Fazakerley, G. V. (1991) Solution structure of an oncogenic duplex containing a G . A mismatch, Biochemistry
Cheng, J.-W., Chou, S.-H. & Reid, B. (1992) Solution structure of [d(GCGTATACGC),, J . Mol. Biol. 228, 1037- 1041.
Chou, S.-H., Cheng, J.-W. & Reid, B. R. (1992a) Solution structure of [d(ATGAGCGAATA)],. Adjacent G . A mismatches stabilised by cross-strand base-stacking and B(I1)-phosphate groups, J. Mol. Biol. 228, 138-155.
Chou, S.-H., Cheng, J.-W., Fedoroff, 0. Y., Chuprina, V. P. & Reid, B. R. (1992b) Adjacent G . A mismatched base-pairs contain B(I1) phosphodiesters in solution, J. Am. Chem. SOC. 114, 3114- 3115.
Cognet, J. A. H., Gabarro-Arpa, J., Le Bret, M., van der Marel, G. A,, van Boom, J. H. & Fazakerley, G. V. (1991) The solution structure of a DNA hairpin containing a loop of 3 thymidines determined by nuclear-magnetic resonance and molecular me- chanics, Nucleic Acids Res. 24, 6771 -6779.
Brown, T., Hunter, W. N., Kneale, G. G. & Kennard, 0. (1986) Molecular structure of the G . A base pair in DNA and its implications for the mechanism of transversion mutations, Proc. Natl Acad. Sci USA 83, 2401 -2406.
Brown, T., Leonard, G. A., Booth, E. D. & Chambers, J. (1989) Crystal structure and stability of a DNA duplex containing A(anti) . G(syn) base pairs, Mol. Biol. 207, 455-457.
Dodgson, J. B. & Wells, R. D. (1977) Synthesis and thermal melting behaviour of oligomer-polymer complexes containing defined lengths of mismatched dA . dG and dG . dG nucleotides, Bio- chemistry 11, 2367-2374.
Ebel, S., Lane, A. N. & Brown, T. (1992) Very stable mismatch duplexes. Structural and thermodynamic studies on tandem G . A mismatches in DNA, Biochemistry 31, 12083-12086.
30, 5449-5458.

727
Fersht, A. R., Knill-Jones, J. W. & Tsui, W. C. (1982) Kinetic basis of spontaneous mutations. Misinsertion frequencies, proofread- ing specificity and cost of proofreading by DNA polymerases of E. coli, J. Mol. Biol. 156, 37-51.
Gao, X. & Patel, D. J. (1988) Gfsyn) . A(anti) mismatch formation in DNA dodecamers at acidic pH. pH dependent conformational transition of G . A mispairs detected by proton NMR, J. Am. Chem. Soc. 110, 5178-5182.
Hore, P. J. (1983) Solvent suppression in Fourier-transform nuclear- magnetic resonance, J. Magn. Reson. 55, 283-300.
Hunter, W. N., Brown, T. & Kennard, 0. (1986) Structural features and hydration of d(CGCGAATTAGCG). A double helix contain- ing 2 G . A mispairs, J. Biomol. Struct. & Dyn. 4, 173-191.
Ivanov, V. I., Minchenkova, L. E., Minyat, E. E., Frank-Kamentskii, M. D. & Schyolkina, A. K. (1974) The B to A transition of DNA in solution, J. Mol. Biol. 87, 817-833.
Lane, A. N. (1990) The determination of the conformational proper- ties of nucleic acids in solution from NMR data, Biochim. Bio- phys. Acta 1049, 189-204.
Lane, A. N., Jenkins, T. C., Brown, D. J. S. & Brown, T. (1991) NMR determination of the solution conformation and dynamics of the A . G mismatch in the [d(GCAAATTGGCG)] . [d(GCAAATTGGCG)] dodecamer, Biochem. J. 279, 269-281.
Lane, A. N. (1991) Solution conformation and dynamics of the octa- nucleotide [d(CACTAGTG)] . [d(CACTAGTG)] : a rnultinuclear NMR study, Carbohydl: Res. 221, 123-144.
Lane, A. N., Martin, S. R., Ebel, S. & Brown, T. (1992) Solution conformation of a deoxynucleotide containing tandem G . A mis- matched base-pairs and 3’-overhanging ends in [d(GTGAAC- TT)] . [d(GTGAACTT)], Biochemistry 31, 12087-12095.
Leonard, G. A., Booth, E. D. & Brown, T. (1990a) Structural and thermodynamic studies on the adenine guanine mismatch in B- DNA, Nucleic Acids Res. 18, 5617-5623.
Leonard, G. A., Thomson, J., Watson, W. P. & Brown, T. (1990b) High-resolution structure of a mutagenic lesion in DNA, Proc. Nut1 Acad. Sci USA 87, 9573-9576.
Leonard, G. A., Hunter, W. N. & Brown, T. (1993) Mismatches in DNA duplexes, Chem. in BI: 29,484-488.
Li, Y., Zon, G. & Wilson, W. D. (1991) Thermodynamics of DNA duplexes with adjacent G . A mismatches, Proc. Natl Acad. Sci USA 88, 26-30.
Marion, D. & Wuthrich, K. (1983) Application of phase-sensitive two dimensional correlated spectroscopy (COSY) for measure- ments of 1H-1 H spin-spin coupling constants in proteins, Biochem. Biophys. Res. Commun. 124, 774-783.
Marky, L. A. & Breslauer, K. J. (1987) Calculating thermodynamic data for transitions of any molecularity, Biopolymers 26, 1601 - 1620.
Maskos, K., Gunn, B. M., LeBlanc, D. A. & Morden, K. M. (1993) NMR study of G . A and A . A pairing in [(dGCGAATAAGCG)] . [(dGCGAATAAGCG)], Biochemistry 32, 3583 - 3595.
Michaels, M. L., Pharn. L., Ngheim, Y., Cruz, C. & Miller, J. H. (1990) Mut Y, an adenine glycosylase active on G-A mispairs,
has homology to endonuclease 111, Nucleic Acids Res. 18, 3841 - 3845.
Moe, J. G. & Russu, I. M. (1992) Kinetics and energetics of base pair opening in 5’-d(CGCGAATTCGCG)-3’ and a substituted dodecamer containing G . T mismatches, Biochemistry 31, 8421 -8428.
Patel, D. J., Koslowski, S. A,, Ikuta, S. & Itakura, K. (1984) Deoxy- guanosine-deoxyadenosine pairing in the [d(CGAGAATTCG- CC)] . [d(CGAGAATTCGCG)] duplex : conformation and dy- namics adjacent to the dG . dA mismatch site, Biochemistry 23, 3207-3217.
PrivC, G. G., Heinemann, U., Chandrasegaran, S., Kan, L.-S., Kopka, M. L. & Dickerson, R. E. (1987) Helix geometry, hydra- tion, and G . A mismatch in a B-DNA decamer, Science 238, 498-504.
Puglisi, J. D. & Tinoco, I. Jr. (1989) Methods Enzymol. 180, 304- 325.
Rinkel, L. J. & Altona, C. J. (1987) Conformational analysis of the deoxyribofuranose ring in DNA by means of sums of proton- proton coupling constants : a graphical method, Biomol. Struct. Dyn. 4, 621-649.
Roongta, V. A,, Jones, C. R. & Gorenstein, D. G. (1990) Effect of distortions in the deoxyribose phosphate backbone conformation of duplex oligodeoxyribonucleotide dodecamers containing GT, GG, GA, AA and GU base-pair mismatches on ”P-NMR spectra, Biochemistry 29, 524555258,
SantaLucia, J., Kierzek, R. & Turner, D. H. (1991) Stabilities of consecutive A . C, C . C, G . G, U . C, and U . U mismatches in RNA internal loops : evidence for stable hydrogen-bonded U . U and C . C pairs, Biochemistry 30, 8242-8251.
Searle, M. S. & Williams, D. H. (1993) On the stability of nucleic acid structures in solution : enthalpy-entropy compensations, in- ternal rotations and reversibility, Nucleic Acids Res. 21, 2051 - 2056.
States, D. J., Haberkorn, R. A. & Ruben, D. J. (1982) A two-dimen- sional nuclear Overhauser experiment with pure absorption phase in four quadrants, J. Magn. Reson. 48, 286-292.
van de Ven, F. J. M. & Hilbers, C. W. (1987) Nucleic acids and nuclear magnetic resonance, Eul: J. Biochem. 178, 1-38.
van de Ven, F. J. M. & Hilbers, C. W. (1988) Resonance assignments on non-exchangeable protons in B type DNA oligomers, an over- view, Nucleic Acids Res. 13, 5713-5726.
Webster, G. D., Sanderson, M. R., Skelly, J. V., Neidle, S., Swann, P. F., Li, B. F. & Tickle, I. J. (1990) Proc. Natl Acad. Sci USA 87, 6693-6697.
Westhofer, G. D., Sanderson, M. R., Skelly, J. V., Swan, P. F., Li, B. F. & Tickle, I. J. (1990) Crystal structure and sequence-depen- dent conformation of the A . G mispaired oligonucleotide d(CGCAAGCTGGCG), Proc. Natl Acad. Sci USA 87, 6693- 6697.





























