Embed Size (px)
Citation preview

1
1
Progesterone RegulatesCardiac Repolarization Through
a Nongenomic PathwayAn In Vitro Patch-Clamp and Computational Modeling Study
Tokyo Medical Research Institute, Tokyo, Japan, and Institute for Computational Biomedicine, Weill Medical College, Cornell University, New York, NY.
Hiroaki Nakamura,Junko Kurokawa,
Chang-Xi Bai, et al.
December, 2007 2
Long QT syndrome (LQTS)
• A heart condition associated with prolongation of repolarization of the cardiac action potential.
• LQTS– Inherited as congenital mutations of ion channels carrying the
cardiac impulse/action potential – Acquired as a result of drugs that block these cardiac ion
currents• LQTS is associated with torsade de pointes (TdP), which
can deteriorate into ventricular fibrillation and ultimately death
• Delayed repolarization prolongs the inward flow of positive depolarizing currents, leading to early after depolarization (EAD), the triggering event for TdP.
3 4
Female sex is a risk factor for the development of TdP in both healthy and LQTS
(Makkar et al., 1993, Nakagawa M et al, 2006)
QT interval and TdP risk fluctuate dynamically during the menstrual cycle and pregnancy
(Nakagawa, Ooie, Takahashi, Taniguchi et al., 2006)
Repolarization duration is shorter in the luteal phase than in the follicular phase by 10 ms
(Nakagawa, Ooie, Takahashi et al., 2006)
In the follicular phase, drug-induced QT prolongation is more severe, and TdP risk is increased
5
Ibutilide (class III antiarrhythmic agent) induced QTc prolongation greatest during menses, intermediate in ovulation, and least in the luteal phase. (RodriguezI, Kilborn, Liu et al., 2001)
In congenital LQTS patients, TdP risk is low during pregnancy and suddenly increases postpartum when serum progesterone suddenly drops. (Seth, Moss, McNitt et al., 2007)
In postmenopausal womenhormone replacement therapy (estrogen) causes
prolongation of the QTc intervalcombinational hormone replacement therapy (estrogen +
progestin) consistently shortens QTc interval(Kadish, Greenland, Limacher et al., 2004) 6
Collectively, these clinical data suggest that the luteal hormone progesterone has protective
effects against long QT– associated arrhythmias;however, very few studies have investigated the effects of progesterone on cardiac repolarization.

2
7
Repolarization: Delayed rectifier K+ channel- IKs; Slowly activating delayed rectifier K+ channel - IKr; Rapidly activating delayed rectifier K+ channel 8
Testosterone acutely affects cardiac repolarization by modulating slowly activating delayed rectifier K+ current (IKs) and L-type Ca2+ current (ICa,L) through the nongenomic pathway of the androgen receptor.
(Bai, Kurokawa, Tamagawa et al., 2005)
Progesterone exhibits various actions through a nongenomic pathway in several cells and tissues,e.g. human T lymphocytes
(Ehring, Kerschbaum, Eder et al., 1998)
9 10
Objectives
• To study the acute effects of progesterone on – Action potential duration (APD)– Repolarizing membrane current: IKs & ICa,L– Basal state vs Sympathetic stimulation state
• To study the nongenomic pathway which mediated the progesterone effects
• Simulation studies in a cardiac cell model:– single cells & multicellular coupled cells– simulating progesterone effect on APD, QT, and
EAD (-> TdP)
11
Animal
• Single ventricular myocytes of female adult guinea pigs (n=112, white Hartrey)
• Staging of the estrus cycle : vaginal impedance (3-4 wks)
Statistical analyses– mean ± SEM– nonparametric test (Wilcoxon signed-rank test) using
for between baseline and drug – ANOVA followed by Bonferroni’s multiple-comparison
using for repeated measures between baseline and various drugs
– P < 0.05 taken as significant 12
Patch clamp
Sucrose density gradient fractionation,immunoblot analysis
Computational methods
Methods / Results / Discussion
3
13
Patch clamp
Sucrose density gradient fractionation,immunoblot analysis
Computational methods
Methods / Results / Discussion
14
• Recording action potentials and membrane currents
• Perforated patch / whole-cell configuration
• Patch perforation: Amphotericin B (0.6 mg/mL) in pipette solution
Patch clamp studies
15
Action potential recordingCurrent-clamping, whole-cell / perforated patch
Pulses : Suprathreshold current pulses, <2 ms, 1 Hz
Internal solution: (mmol/L)110 aspartic acid, 30 KCl, 1 CaCl2, 5 MgATP, 5 K2CrP, 5 HEPES,10 EGTA,pH 7.25 adjusted with KOH.
External solution (Tyrode’s): (mmol/L) 135 NaCl, 0.33 NaH2PO4, 5.4 KCl, 1.8 CaCl2, 0.53 MgCl2, 5.5 glucose, 5 HEPES, pH 7.4 adjusted with NaOH
16
IKs recordingVoltage-clamping, whole-cell / perforated patch
Internal solution: (mmol/L)110 aspartic acid, 30 KCl, 1 CaCl2, 5 MgATP, 5 K2CrP, 5 HEPES,10 EGTA,pH 7.25 adjusted with KOH.
External solution(Tyrode’s, excluding KCl): 135 NaCl, 0.33 NaH2PO4, 1.8 CaCl2, 0.53 MgCl2, 5.5 glucose, 5 HEPES, pH 7.4 adjusted with NaOH. Adding nisoldipine (3 mmol/L) and E-4031 (10 mmol/L)
Absence of [K+]o blocks IKr& enhances IKs
(Nakagawa et al.,2006)
Nisoldipine is ICa,L blockerE-4031 is IKr blocker
17
IKs
3.5 s
6.5 s-40
+50
0.1 Hz
Vt
Vh
mV
s
400 ms
600 ms-40
+50
1 Hz
Vt
Vhs
mV
Pulse patternsFor current recordings
18
IKs Pulse patternsFor I-V curve construction
400 ms
-40
+100
Vh
mV
s
Vt20 mV
4
19
ICa,L recording
External solution: (mmol/L) 135 NaCl, 5.4 CsCl, 0.53 MgCl2, 2 CaCl2,5.5 glucose, 5 HEPES, pH 7.4 adjusted with CsOH
Internal solution: (mmol/L)130 CsCl, 2 MgCl2, 5 Na2ATP,10 HEPES, 20 TEA-Cl, 10 EGTA pH 7.25 adjusted with CsOH
20
200 ms
100ms-40
0
1 Hz
Vt
Vh -80700 ms
ICa,L Pulse patternsFor current recordings
21
ICa,L Pulse patternsFor I-V curve construction
200 ms
-40
+70
Vh
mV
s
Vt
-80
100 ms
10 mV
22
• No significant difference in progesterone effects on APD between estrous and diestrous stages.
• Data obtained from guinea pigs at different stages of the estrous cycle were pooled
Effects of Progesterone UnderBasal Conditions
23
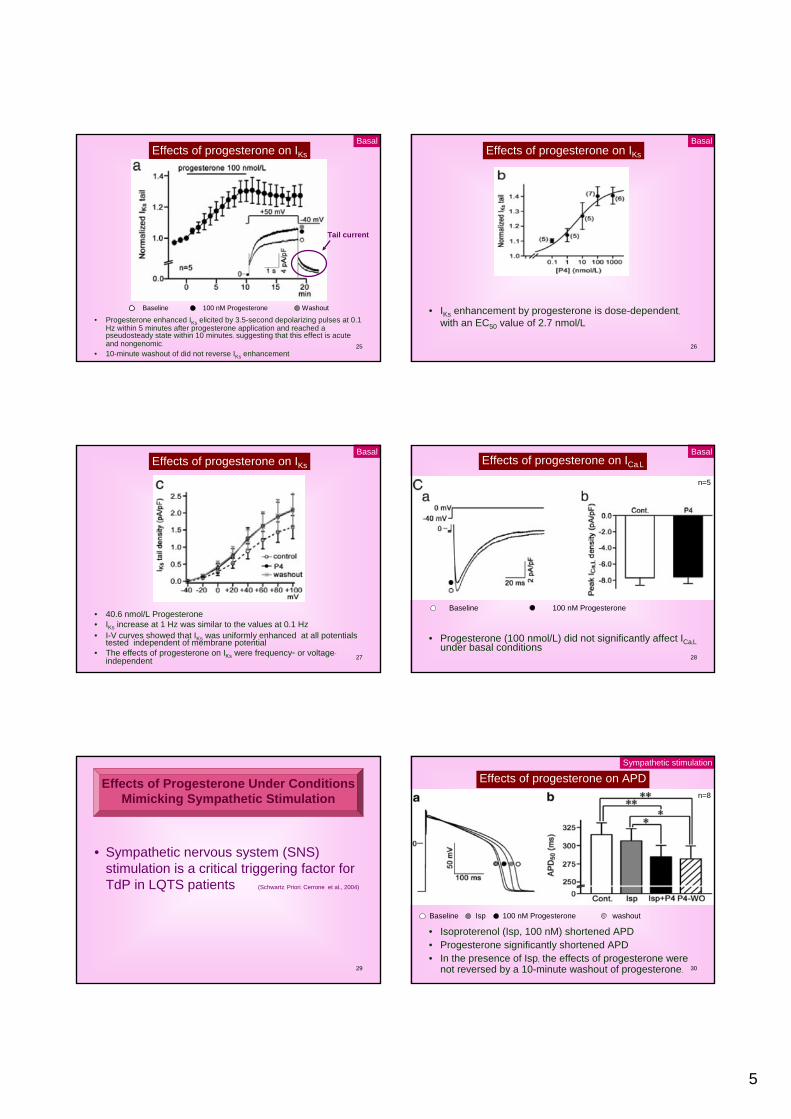
Effects of progesterone on APD
• Progesterone shortened APD• 10-minute washout did not reverse the effects of
progesterone• APD50 was decreased after progesterone
Baseline 100 nM Progesterone Washout
n=7
Basal
24
Effects of progesterone on APD
• Mifepristone (1 µmol/L) (specific progesterone receptor inhibitor) reversed APD shortening
n=6
Basal
5
25
Effects of progesterone on IKs
• Progesterone enhanced IKs elicited by 3.5-second depolarizing pulses at 0.1Hz within 5 minutes after progesterone application and reached a pseudosteady state within 10 minutes, suggesting that this effect is acute and nongenomic.
• 10-minute washout of did not reverse IKs enhancement
Baseline 100 nM Progesterone Washout
Basal
Tail current
26
Effects of progesterone on IKs
• IKs enhancement by progesterone is dose-dependent,with an EC50 value of 2.7 nmol/L
Basal
27
Effects of progesterone on IKs
• 40.6 nmol/L Progesterone• IKs increase at 1 Hz was similar to the values at 0.1 Hz • I-V curves showed that IKs was uniformly enhanced at all potentials
tested independent of membrane potential• The effects of progesterone on IKs were frequency- or voltage-
independent
Basal
28
Effects of progesterone on ICa,L
• Progesterone (100 nmol/L) did not significantly affect ICa,Lunder basal conditions
Baseline 100 nM Progesterone
n=5
Basal
29
• Sympathetic nervous system (SNS) stimulation is a critical triggering factor for TdP in LQTS patients (Schwartz, Priori, Cerrone et al., 2004)
Effects of Progesterone Under ConditionsMimicking Sympathetic Stimulation
30
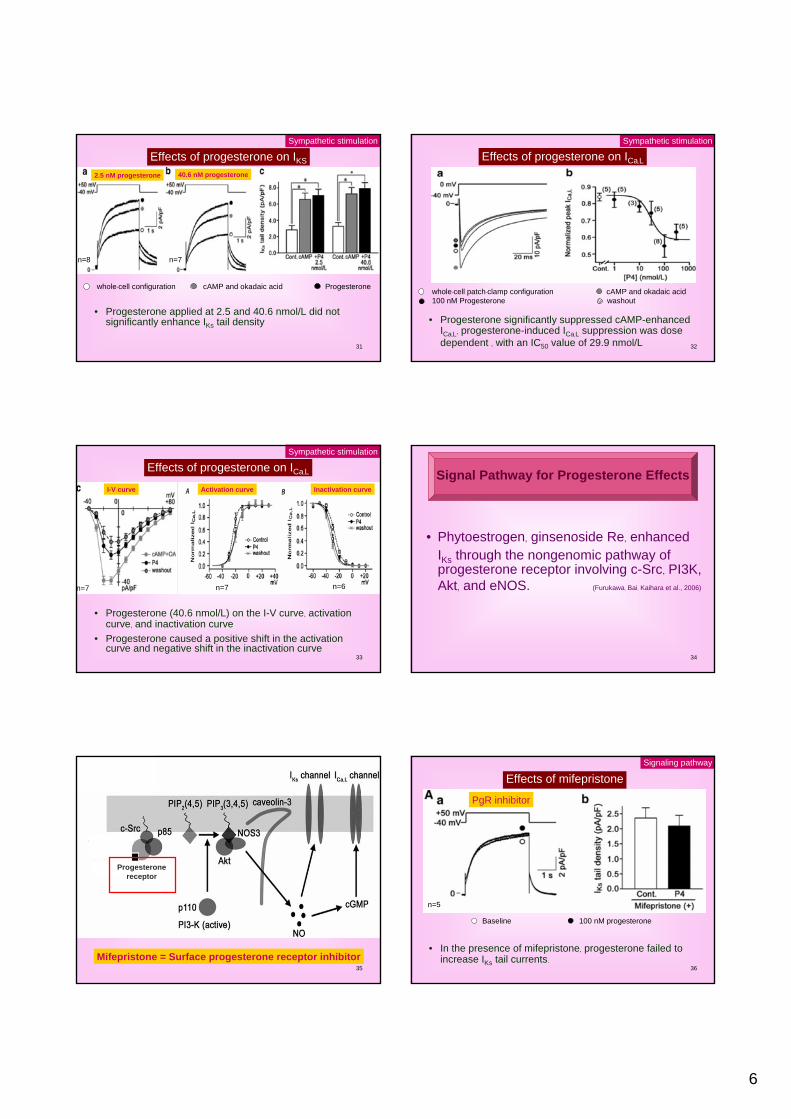
Effects of progesterone on APD
• Isoproterenol (Isp, 100 nM) shortened APD• Progesterone significantly shortened APD• In the presence of Isp, the effects of progesterone were
not reversed by a 10-minute washout of progesterone.
Baseline Isp 100 nM Progesterone washout
n=8
Sympathetic stimulation
6
31
Effects of progesterone on IKS
• Progesterone applied at 2.5 and 40.6 nmol/L did not significantly enhance IKs tail density
Sympathetic stimulation
whole-cell configuration cAMP and okadaic acid Progesterone
n=8 n=7
2.5 nM progesterone 40.6 nM progesterone
32
Effects of progesterone on ICa,L
whole-cell patch-clamp configuration cAMP and okadaic acid 100 nM Progesterone washout
• Progesterone significantly suppressed cAMP-enhanced ICa,L, progesterone-induced ICa,L suppression was dose dependent , with an IC50 value of 29.9 nmol/L
Sympathetic stimulation
33
Effects of progesterone on ICa,L
• Progesterone (40.6 nmol/L) on the I-V curve, activation curve, and inactivation curve
• Progesterone caused a positive shift in the activation curve and negative shift in the inactivation curve
Sympathetic stimulation
n=7 n=7 n=6
Inactivation curveI-V curve Activation curve
34
• Phytoestrogen, ginsenoside Re, enhanced IKs through the nongenomic pathway of progesterone receptor involving c-Src, PI3K, Akt, and eNOS. (Furukawa, Bai, Kaihara et al., 2006)
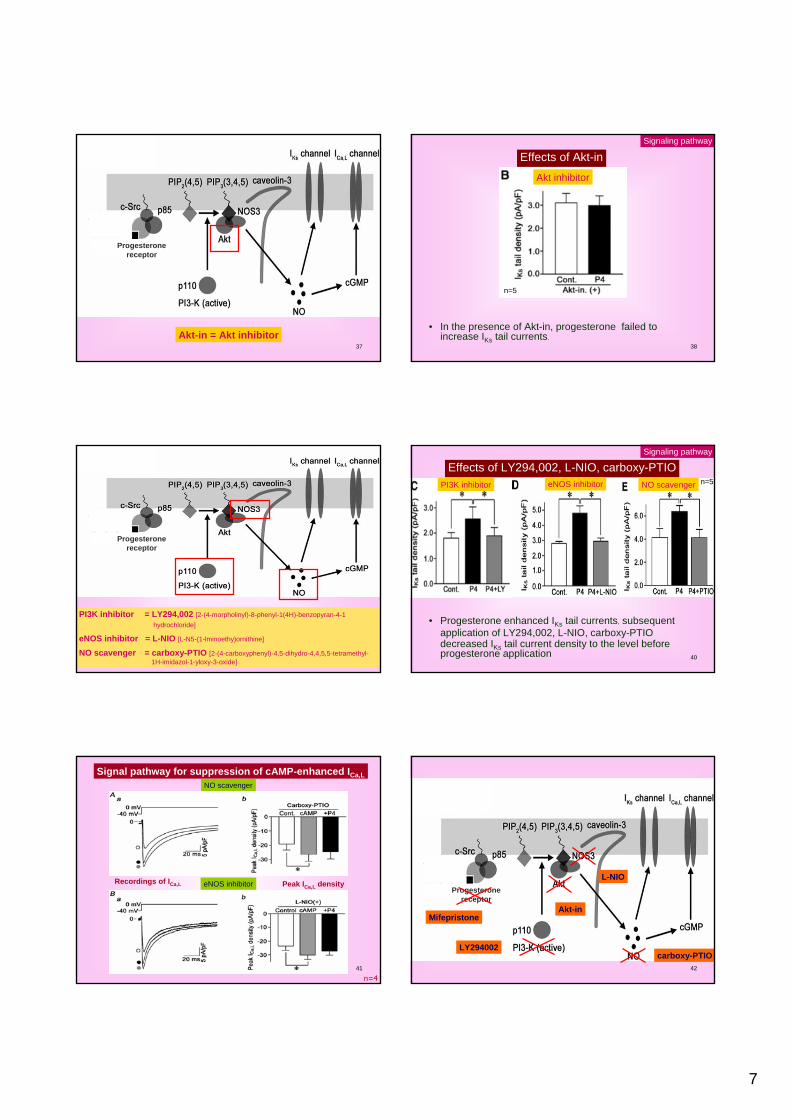
Signal Pathway for Progesterone Effects
35
Mifepristone = Surface progesterone receptor inhibitor
Progesteronereceptor
36
Effects of mifepristone
• In the presence of mifepristone, progesterone failed to increase IKs tail currents.
Signaling pathway
Baseline 100 nM progesterone
n=5
PgR inhibitor
7
37
Progesteronereceptor
Akt-in = Akt inhibitor38
Effects of Akt-in
• In the presence of Akt-in, progesterone failed to increase IKs tail currents.
Signaling pathway
n=5
Akt inhibitor
39
Progesteronereceptor
PI3K inhibitor = LY294,002 [2-(4-morpholinyl)-8-phenyl-1(4H)-benzopyran-4-1 hydrochloride]
eNOS inhibitor = L-NIO [L-N5-(1-lminoethy)ornithine]
NO scavenger = carboxy-PTIO [2-(4-carboxyphenyl)-4,5-dihydro-4,4,5,5-tetramethyl-1H-imidazol-1-yloxy-3-oxide]
40
Effects of LY294,002, L-NIO, carboxy-PTIO
• Progesterone enhanced IKs tail currents, subsequent application of LY294,002, L-NIO, carboxy-PTIO decreased IKs tail current density to the level before progesterone application
Signaling pathway
n=5PI3K inhibitor eNOS inhibitor NO scavenger
41
Signal pathway for suppression of cAMP-enhanced ICa,L
n=4
eNOS inhibitor
NO scavenger
Recordings of ICa,L Peak ICa,L density
42
Progesteronereceptor
LY294002
Akt-in
L-NIO
carboxy-PTIO
Mifepristone

8
43
• The present study to be the first to demonstrate nongenomic actions of progesterone in cardiac myocytes.
Progesterone exhibits its effects via NO produced through the nongenomic pathway
involving PI3-kinase and eNOS
Discussion
44
Under basal conditions progesterone affected only IKs
Under cAMP stimulated conditionsprogesterone clearly reversed cAMP-induced enhancement of ICa,L
This difference could be explained by differential mechanisms of NO to modulate IKs and ICa,L
NO-induced ICa,L suppression was attributable to cGMP-dependent pathwayNO-induced IKs enhancement was via a cGMP-independent pathway
(Bai, Takahashi, Masumiya et al., 2004)
The former agrees with the previous finding that cGMP counteracts cAMP-induced ICa,L enhancement.
(Fischmeister, Castro, Abi-Gerges et al., 2005)
45
Patch clamp
Sucrose density gradient fractionation,immunoblot analysis
Computational methods
Methods / Results / Discussion
46
• Using sucrose density gradient fractionation to determine the localization of molecules involved in the nongenomic pathway of progesterone
Caveolae Localization of molecules involved in The Nongenomic Pathway of Progesterone
47
Nuclei & cell debris
4
Cytosolic fraction
Membrane fraction
48
Transferred to PVDF membrane
Incubation with HRP-conjugated anti-rabbit IgG, anti-mouse IgG, or anti-goat IgG
The proteins were detected with an advanced enhanced chemiluminescencesystem using a lumino-image analyzer
anti-caveolin-3 antibodyanti-KCNQ1 antibodyanti-KCNE1 antibodyanti-Cav1.2 antibody
c-Src antibody Akt antibodyp85 antibody
anti-eNOS antibody anti-progesterone receptor antibody

9
49
n=5
Progesterone-free condition
50
n=4
Progesterone-treated condition
51
• The sucrose density gradient fractionation experiments demonstrate that substantial fractions of c-Src, Akt, eNOS, PR78, PR54, KCNQ1, KCNE1, and CaV1.2 are co-localized in the caveolae fraction.
Discussion
52
• Molecular identity for the PgR responsible for the nongenomic action is currently undetermined, but several candidates have been reported.
• One candidate is PR78 because injection of PR78 mRNA into oocytes rapidly accelerates progesterone-induced mitogen-activated protein kinase activation and oocyte maturation. (Bayaa, Booth, Sheng et al., 2000)
53
• Sucrose gradient experiments also showed that progesterone application shifted the localization of PR78 and PR54 to broad membrane fractions but not to the cytosolic fraction.
• Immunocytochemical experiments showed that progesterone did not induce translocation of PgR into nucleus.
• These findings are in line with the idea that PR78 is responsible for nongenomic actions, at least in cardiomyocytes.
54
• Further studies are certainly required– to determine whether PR78 and/or newly
identified PR54 is the receptor specific for the nongenomic pathway in cardiac myocytes
– to clarify the physiological implication of ligand-induced translocation of PR78 and PR54 within plasma membrane.
10
55
Patch clamp
Sucrose density gradient fractionation,immunoblot analysis
Computational model
Methods / Results / Discussion
56
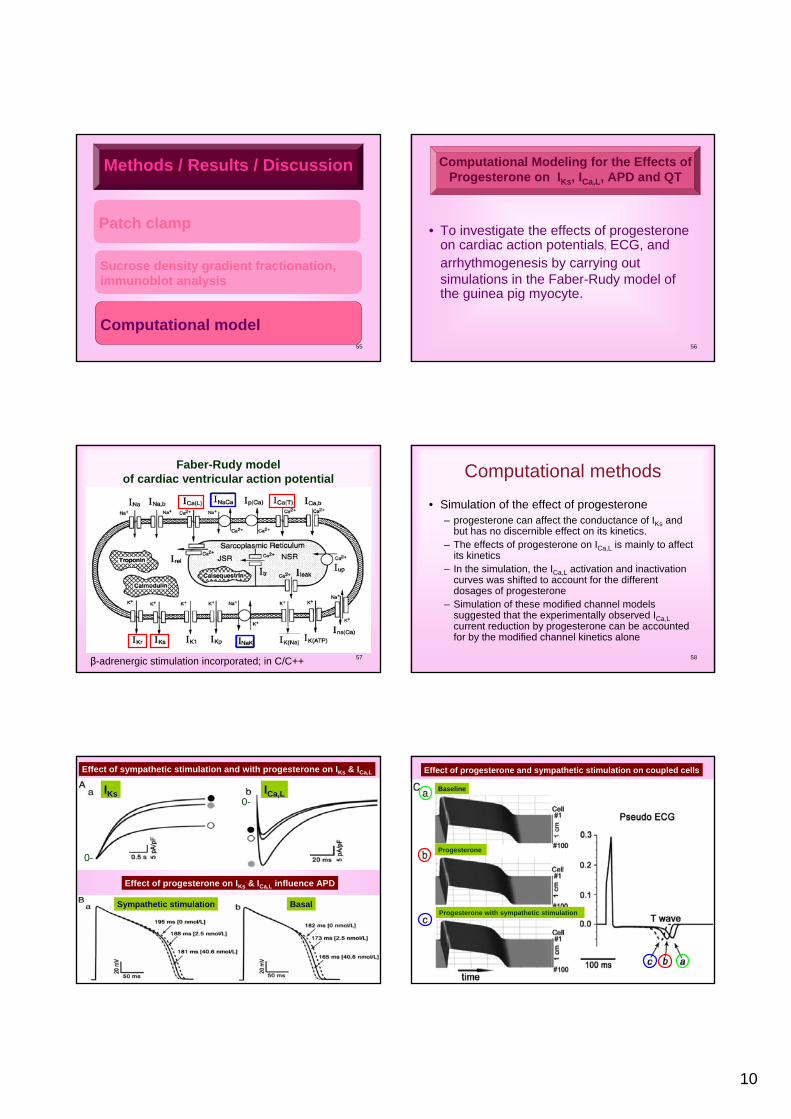
• To investigate the effects of progesterone on cardiac action potentials, ECG, and arrhythmogenesis by carrying out simulations in the Faber-Rudy model of the guinea pig myocyte.
Computational Modeling for the Effects ofProgesterone on IKs, ICa,L, APD and QT
57
Faber-Rudy model of cardiac ventricular action potential
β-adrenergic stimulation incorporated; in C/C++ 58
Computational methods
• Simulation of the effect of progesterone– progesterone can affect the conductance of IKs and
but has no discernible effect on its kinetics. – The effects of progesterone on ICa,L is mainly to affect
its kinetics – In the simulation, the ICa,L activation and inactivation
curves was shifted to account for the different dosages of progesterone
– Simulation of these modified channel models suggested that the experimentally observed ICa,Lcurrent reduction by progesterone can be accounted for by the modified channel kinetics alone
59
Effect of sympathetic stimulation and with progesterone on IKs & ICa,L
Effect of progesterone on IKs & ICa,L influence APD
IKs ICa,L
Sympathetic stimulation Basal
0-
0-
60
Effect of progesterone and sympathetic stimulation on coupled cells
Baseline
Progesterone
Progesterone with sympathetic stimulation

11
61
• To predict the effects of progesterone on LQTS-associated arrhythmia susceptibility.
• To examine the effects on SNS-induced arrhythmias– Used the D76N KCNE1 mutation linked to
congenital LQTS5; reduces the current and renders the IKs channel insensitive toadrenergic stimulation, readily develop TdP with SNS stimulation.
Protective Effects Against Arrhythmia in a Computer Modeling
62
Effect of progesterone on arrhythmic rhythms in congenital LQTS
Sympathetic stimulation
63
Effect of progesterone on drug-induced arrthymias
64
• The effects of progesterone on IKs and ICa,L were incorporated in the Faber-Rudy model of the guinea pig ventricular myocyte.
• The model predicts that 40.6 nmol/L progesterone, luteal phases; shortens APD by 3.7% under basal conditions and 4.6% under SNS-stimulated conditions compared with APD at 2.5 nmol/L progesterone, follicular phase.
• Clinically observed QT intervals are shorter by 2.4% to 2.8% in the luteal phase than in follicular phase, so the APD shortening predicted in the model (3.7% to 4.6%) fits well with the observed fluctuation in QT interval during the menstrual cycle.
Discussion
65
• The effects of progesterone and SNS stimulation in simulated coupled tissue were also investigated and the virtual electrograms computed.
• Simulations suggest that during the luteal phase when progesterone is 40.6 nmol/L, maximal SNS stimulation may additionally shorten the QT interval by 12.2%.
• These simulations support the notion that progesterone may exert protective QT shortening effects under conditions of SNS stimulation.
66
• In LQTS1 and LQTS5, IKs channel disturbance results in dysfunction of action potential adaptation to rapid heart rates and response to SNS stimulation.
• Drug-induced TdP is believed to occur by blockade of the human ether-a-go-go related gene (hERG) channel by drugs.
• Progesterone does not have apparent effects on IKr (data not shown); thus, the predicted protection against drug-induced early afterdepolarizations may be attributed to an increase in repolarization reserve by IKsenhancement.

12
67
• 1. progesterone modulates IKs and ICa,L through a nongenomic pathway involving c-Src/PI3K/Akt–dependent eNOS activation
• 2. progesterone enhances IKs under basal condition,whereas it inhibits ICa,L only when ICa,L is enhanced by cAMP
• 3. substantial fractions of molecules involved in the nongenomic pathway of progesterone colocalize in the caveolae fraction
4 findings regarding the effects ofprogesterone on cardiac repolarization
68
• 4. incorporating the effects of progesterone into a simulated myocyte and a coupled-tissue strand – To observed fluctuations of cardiac repolarization
duration during the menstrual cycle in women – To predicts the protective effects of progesterone
against rhythm disturbance in congenital and drug-induced LQTS
69
Thank you
70
71SCN4B, subunit of the voltage-gated sodium channelLQT10
Caveolin3, mutations in the membrane structural proteinLQT9
alpha subunit of the calcium channel Cav1.2 encoded by the gene CACNA1c.LQT8
potassium channel KCNJ2 (or Kir2.1)LQT7beta subunit MiRP1 (or KCNE2)LQT6beta subunit MinK (or KCNE1)LQT5anchor protein Ankyrin BLQT4alpha subunit of the sodium channel (SCN5A)LQT3
alpha subunit of the rapid delayed rectifier potassium channel (hERG)LQT2
alpha subunit of the slow delayed rectifier potassium channel (KCNQ1)LQT1
MutationType
72
Effects of progesterone on APD in two different estrous stages
estrous diestrous
• Progesterone at 100 nmol/L significantly shortened APD.
• There was no significant difference in fractional shortening of APD between at the estrous stage and at the diestrous stage
• conclude that effects of progesterone on APD showed little or nodifference depending on the stage of the estrous cycle of femaleguinea pigs.

13
73
Indirect immunofluorescent analysis
Absence of progesterone (100 nmol/L)
Presence of progesterone (100 nmol/L)
Negative control without primary antibodies
74
5 main classes of antiarrhythmic agents
• Class I agents interfere with the sodium(Na+) channel (Na+ channel blocker)
• Class II agents are anti-sympathetic nervous system agents (Beta blockers)
• Class III agents affect potassium(K+) efflux (K+
blocker)• Class IV agents affect calcium channels and the
AV node (Ca2+ blocker)• Class V agents work by other or unknown
mechanisms (increases vagal activity )
75
•decrease myocardial infarction mortality•prevent recurrence of tachyarrhythmias
Beta blocking•propranolol•timolol•metoprolol•sotalol•atenolol
II
•In Wolff-Parkinson-White syndrome•(sotalol:) ventricular tachycardias and atrialfibrillation
K+ channel block•amiodarone•sotalol•bretylium
III
•prevent recurrence of paroxysmal supraventricular tachycardia•reduce ventricular rate in patients with atrialfibrillation
Ca2+ channel block•verapamil•diltiazem
IV
Increases vagal activity via its central action on the central nervous system, thus decreasing the conduction of electrical impulses through the AV node
digoxin and adenosine
v
•prevents paroxysmal atrial fibrillation•treats recurrent tachyarrhythmias of abnormal conduction system.
Na+ channel block block (slow association/dissociation)
•flecainide•propafenone
Ic
•treatment and prevention during and immediately after myocardial infarction
•ventricular tachycardia•atrial fibrillation
Na+ channel block (fast association/dissociation)
•lidocaine•phenytoin
Ib
•Ventricular arrhythmias•prevention of paroxysmal recurrent atrialfibrillation (triggered by vagal overactivity)
Na+ channel block (intermediate association/dissociation)
•disopyramide•procainamide•quinidine
Ia
Clinical usesMechanismExamplesClass
76
Indirect immunofluorescenceLeft ventricular myocytes plated onto laminin-coated glass bottom dishes,
allowing attachment
Incubated with vehicle or progesterone (100 nmol/L) for 10 min
Fixed and permeabilized using 99.5 % ethanol (15 min for -20 °C)
Incubated with 3 % bovine serum albumin in phosphate-buffered saline for 30 min to reduce non-specific binding
77
Overnight (4 °C) incubation with primary antibodies (anti-progesterone receptor antibody, and anti-caveolin-3 antibody)
Secondary antibodies (Alexa Fluor 488-conjugated anti-rabbit IgG& Alexa Fluor 647-conjugatged anti-mouse IgG); 2 h, room temperature
Confocal laser-scanning microscopy(Alexa 647, excited with 633-nm He-Ne laser beam, observed at 650-700 nm;
Alexa 488, excited with 488-nm Ar laser beam, observed at 505-520 nm)
78
Reagents• Stock solutions for E-4031 (10 mmol/L), L-NIO (1
mmol/L), cAMP (2 mmol/L), Isp (10 mmol/L), Akt-in (10 mmol/L), and okadaic acid (0.1 mmol/L) were prepared in distilled water
• progesterone (10 mmol/L) and mifepristone (10 mmol/L) in ethanol
• nisoldipine (10 mmol/L), carboxy-PTIO (200 mmol/L), and LY-294,002 (60 mmol/L) in dimethylsulfoxide(DMSO)
• The final concentration of DMSO (0.05 %[vol/vol]) or ethanol (0.01 %[vol/vol]) was confirmed to have no significant effects on action potentials or membrane currents
14
79
Reagents• E-4031• progesterone and tetrodotoxin• mifepristone• Nisoldipine• 2-(4-carboxyphenyl)-4,5-dihydro-4,4,5,5-tetramethyl-
1H-imidazol-1-yloxy-3-oxide (carboxy-PTIO)• L-N5-(1-lminoethy)ornithine (L-NIO)• adenosine 3′,5′-cyclic monophosphate (cAMP)• (-)-isoproterenol hydrochloride (Isp)• 2-(4-morpholinyl)-8-phenyl-1(4H)-benzopyran-4-1
hydrochloride (LY-294,002)• TAT-Akt-inhibitor peptide (Akt-in)• okadaic acid 80
81
•vasodilation•endotheliumNOS3Endothelial NOS(eNOS or NOS3 orcNOS)
•immunedefence against pathogens
•immune system•cardiovascular system
NOS2A, NOS2B, NOS2C
Inducible NOS(iNOS or NOS2)
•cell communication
•nervous tissue
NOS1Neuronal NOS(nNOS or NOS1)
FunctionLocationGene(s)Name





























