Embed Size (px)
Citation preview

Production of Protein from Cloned Genes
OrR
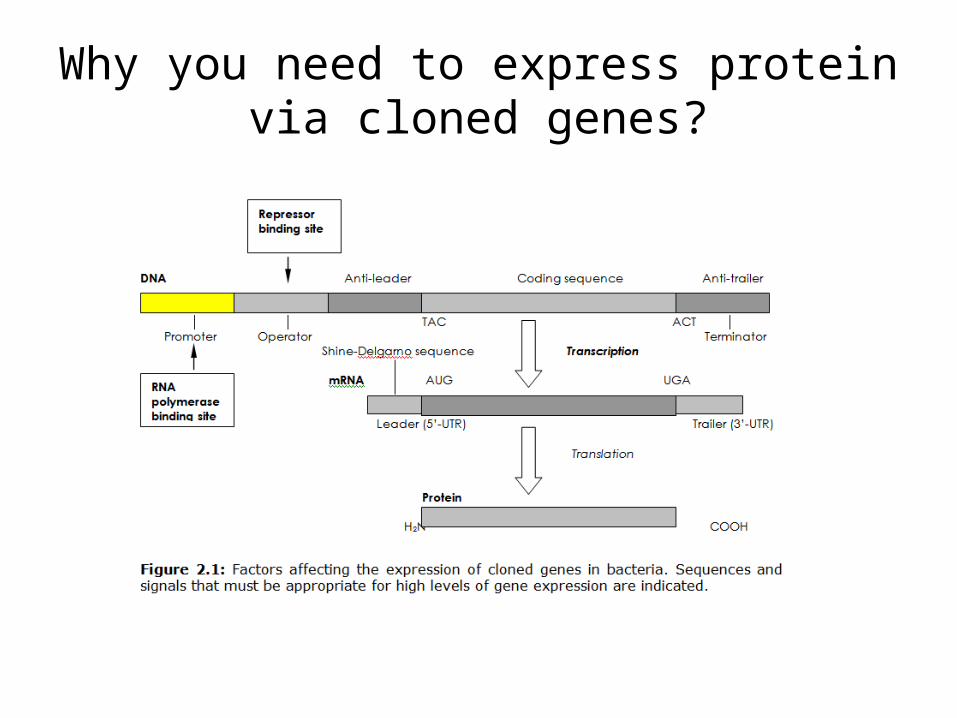
Why you need to express protein via cloned genes?

Regulation of Gene ExpressionReasons for expressing a gene all the time:• Some proteins that are of commercial interest
are toxic to the bacterial hosts. • In addition, some expression system, such as
that involving the T7 promoter, are so powerful that normal host genes cannot be expressed.
The ideal situation is to be able to grow the culture containing the expression vector until a large population of cells is obtained, each containing a large copy number of the vector, and then turn on expression in all copies simultaneously by manipulation of a regulatory switch.

Repressor-Operator System • repressor/operator system in regulating gene transcription
• A strong repressor can completely block the synthesis of the proteins under its control by binding to the operator region.
• Repressor function can be turned off at the chosen time by adding an inducer, allowing the transcription of the genes controlled by the operator.

This permits proper arrangement of the sequence of genetic elements:
promoteroperatorribosome-binding sitestructural gene
Lac operon in E. coliFunction - to produce enzymes which break down lactose (milk sugar)
lactose is not a common sugar, so there is not a great need for these enzymes when lactose is present, they turn on and produce enzymes
Two components - repressor genes and functional genes
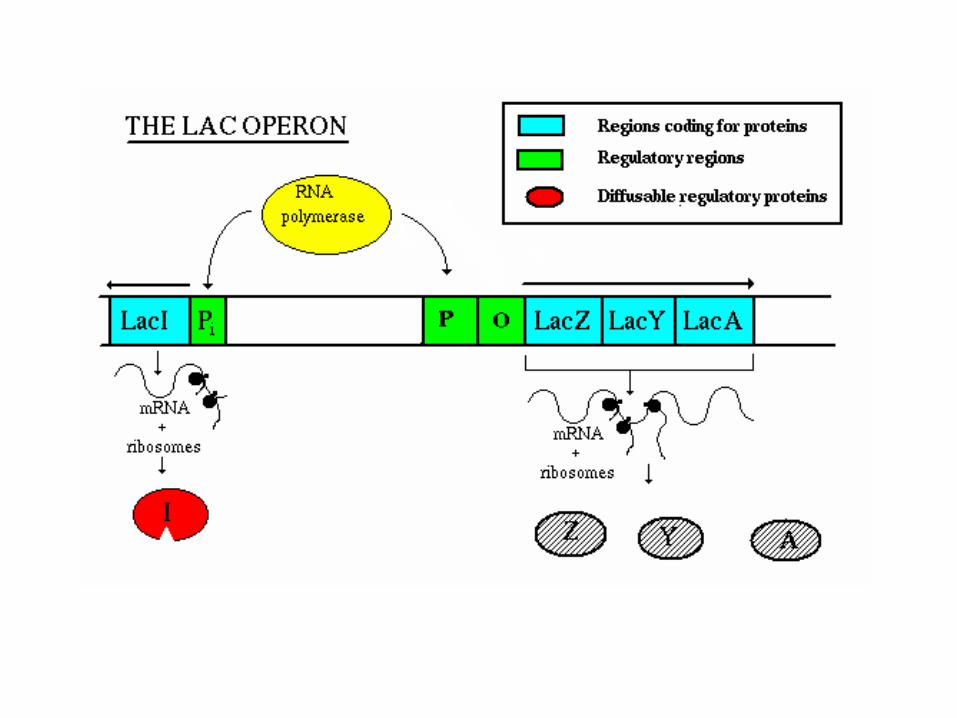

lac operon in E. coli
Three functional genes: lacZ produces B-galactosidase. This enzyme hydrolyzes the bond between the two sugars, glucose and galactose lacY produces permease. This enzyme spans the cell membrane and brings lactose into the cell from the outside environment. The membrane is otherwise essentially impermeable to lactose. lacA produces B-galactosidase transacetylase. The function of this enzyme is still not known.
Promoter (P) - aids in RNA polymerase binding Operator (O) - "on/off" switch - binding site for the repressor protein Repressor (lacI) gene
Repressor gene (lacI) - produces repressor protein w/ two binding sites, one for the operator and one for lactose
• The repressor protein is under allosteric control - when not bound to lactose, the repressor protein can bind to the operator
• When lactose is present, an isomer of lactose, allolactose, will also be present in small amounts. Allolactose binds to the allosteric site and changes the conformation of the repressor protein so that it is no longer capable of binding to the operator
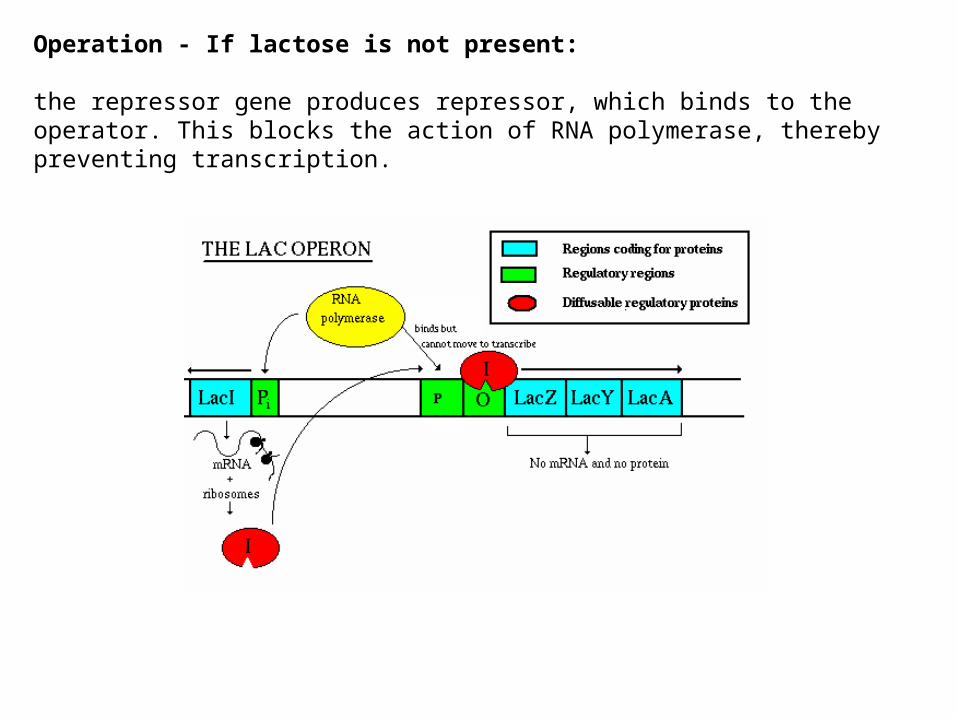
Operation - If lactose is not present:
the repressor gene produces repressor, which binds to the operator. This blocks the action of RNA polymerase, thereby preventing transcription.
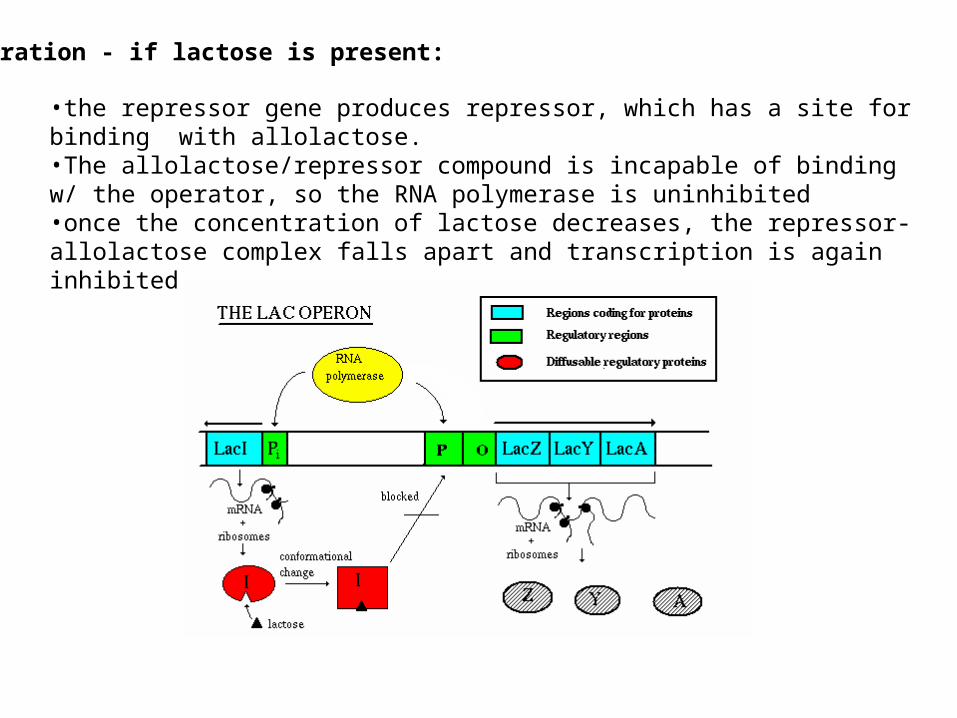
Operation - if lactose is present:
•the repressor gene produces repressor, which has a site for binding with allolactose. •The allolactose/repressor compound is incapable of binding w/ the operator, so the RNA polymerase is uninhibited •once the concentration of lactose decreases, the repressor-allolactose complex falls apart and transcription is again inhibited

The lac operon is an example of an inducible operon - it is normally off, but when a molecule called an inducer is present, the operon turns on.
The trp operon is an example of a repressible operon - it is normally on but when a molecule called a repressor is present the operon turns off.
Task for students:
Solve the trp operon function in details.

Factors for designing expression vectors:
1. Number of Copies of the Gene Per Cell
2. Strength of the Transcriptional Promoter
3. Presence of the Bacterial Ribosome-Binding Site
4. Proper Reading Frame
5. Codon Uses
6. Fate of the Protein after It is Produced

1. Number of Copies of the Gene Per Cell
•In general, more product is made if many copies of the gene are present.
•Vectors such as small plasmids (e.g., pBR322) are valuable because they can replicate to a high copy number.
•Sometimes, often for research purposes, it is desirable to have only a single copy of the cloned gene in the cell. For these cases, integrating vectors have been developed so that the gene can recombine into the host chromosome.

2. Strength of the Transcriptional Promoter
For bacteria, the DNA region around 10 and 35 nucleotides before the start of transcription (called the 10 and 35 region) is especially important in the promoter.
Many Escherichia coli genes are controlled by relatively weak promoters, and promoters from eukatotes and some other prokaryotes function poorly or not at all in E. coli.
Strong E. coli promoters that have been used in the construction of expression vectors include:
•lacuv 5 : which normally controls -galactosidase•try :which normally controls tryptophan synthetase•Tac :a synthetic hybrid of the 35 region of the try promoter and the 10 region of the lac promoter •lambda PL :which normally regulates lambda virus production•ompF :which regulates production of an outer membrane protein• T7 promoters

3. Presence of the Bacterial Ribosome-Binding Site
The transcribed mRNA must bind firmly to the ribosome if translation is to begin, and an early part of the transcript contains the ribosome-binding site (Shine-Dalgarno sequence).
Bacterial ribosome-binding sites are not found in eukaryotic genes, and it is thus essential that the bacterial region be present in the cloned gene if high levels of gene expression are to be obtained.
Part of the requirement for proper ribosome binding is the necessity for a proper distance between the ribosome-binding site and the translation initiation codon.
If these sites are too close or too far apart, the gene will be translated at low efficiency.

4. Proper Reading Frame
•The ribosome-binding site and even the initiation codon for the gene to be cloned are part of the expression vector.
•Because of the way the source DNA is fused into such a vector, three possible reading frames could be obtained, only one of which is satisfactory.
•One approach that can be used if the correct frame is not known is the use of three vectors, each having the restriction site into which new DNA will be inserted positioned such that the insert will be in a different reading frame.
•The gene fragment is inserted into all three vectors and the one which gives proper expression is selected by testing.

5. Codon Uses
6. Fate of the Protein after It is Produced
The skill of the genetic engineer is thus essential in the construction of an appropriate vector, which can be (1) efficiently incorporated into the proper host, (2) replicated to high copy number, (3) efficiently transcribed(4) efficiently translated.

Expression Vector
A cloning vector that has been constructed in such a way that, after insertion of a DNA molecule, its coding sequence is properly transcribed and the mRNA is translated.
The cloned gene is put under the control of a promoter sequence for the initiation of transcription, and often also has a transcription termination sequence at its end.
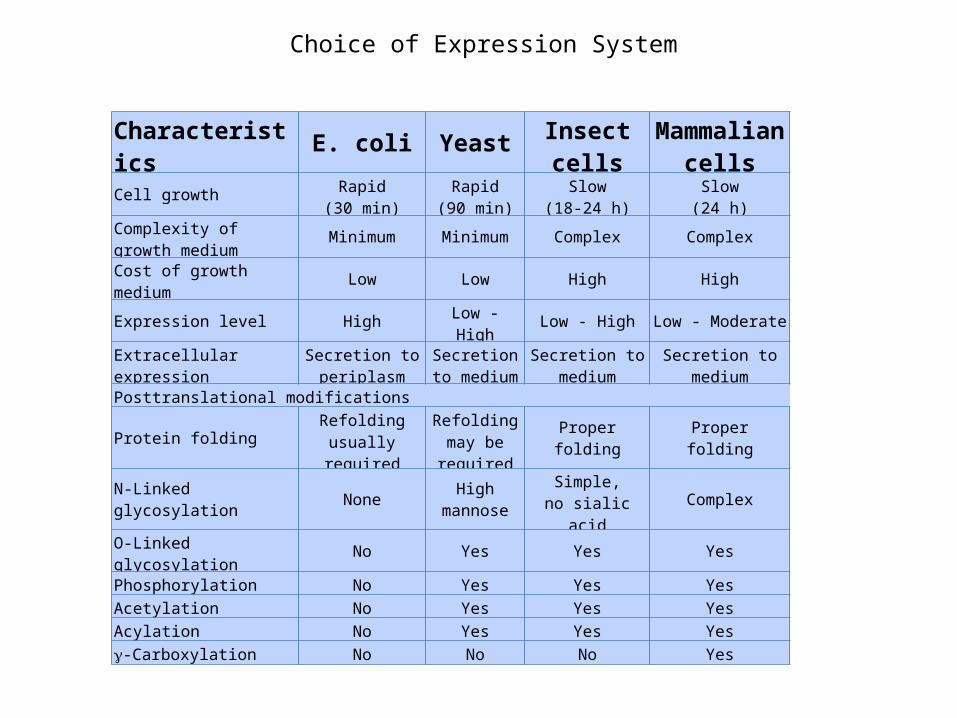
Choice of Expression System
Characteristics E. coli Yeast Insect cells Mammalian cells
Cell growth Rapid(30 min)
Rapid(90 min)
Slow(18-24 h)
Slow(24 h)
Complexity of growth medium Minimum Minimum Complex Complex
Cost of growth medium Low Low High HighExpression level High Low - High Low - High Low - Moderate
Extracellular expression Secretion to periplasm
Secretion to medium
Secretion to medium
Secretion to medium
Posttranslational modifications
Protein folding Refolding usually required
Refolding may be required
Properfolding
Properfolding
N-Linked glycosylation None High mannose Simple,no sialic acid Complex
O-Linked glycosylation No Yes Yes YesPhosphorylation No Yes Yes YesAcetylation No Yes Yes YesAcylation No Yes Yes Yes-Carboxylation No No No Yes

Choice of Expression System
1. What type of protein do I want to express?
2. Do I get soluble protein when I express in E. coli?
3. Does my protein need post-translational modifications for structure or activity?
4. What is the codon usage in my protein?
The following problems are often encountered:(i) Interrupted translation, which leads to a variety of truncated protein products,(ii) Frame shifting,(iii) Misincorporation of amino acids. For instance, lysine for arginine as a result of the AGA codon, and(iv) Inhibition of protein synthesis and cell growth

To increase the expression levels of proteins containing rare codons in E. coli, two main methods are available:
(i) Site-directed mutagenesis to replace the rare codons by more commonly used codons for the same residue; e.g. the rare argenines codons AGA and AGG by the E. coli preferred CGC codon.
(ii) Co-expression of the genes, which encode rare tRNAs. There are several commercial E. coli strains available that encode for a number of the rare codon genes.
Providing the protein with a C-terminal tag (e.g. His6-tag) will help you to purify only the full-length protein using affinity chromatography.
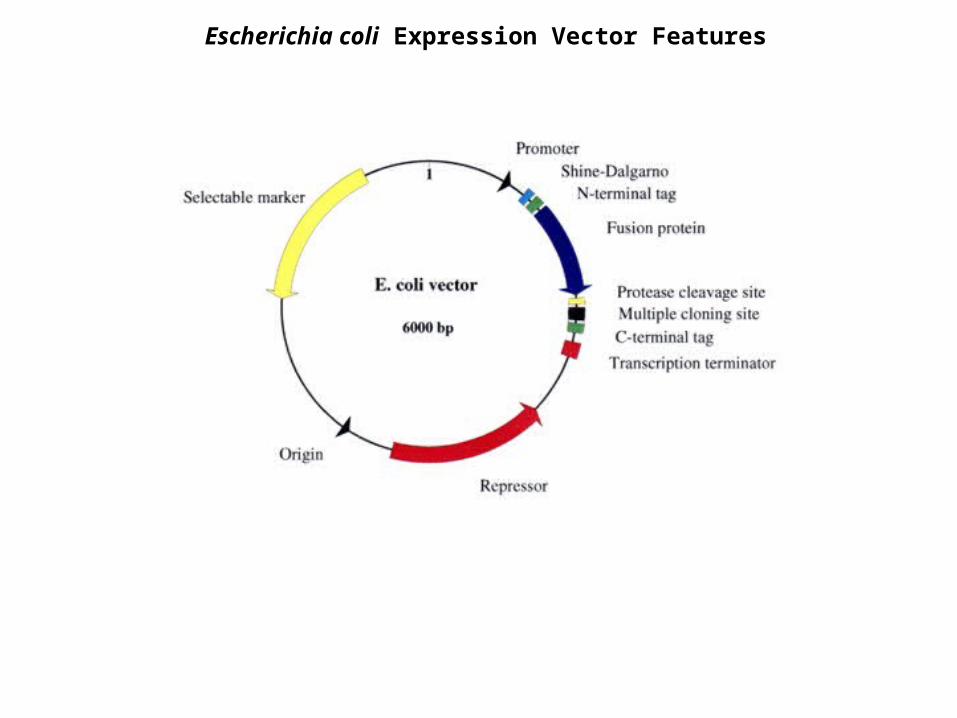
Escherichia coli Expression Vector Features
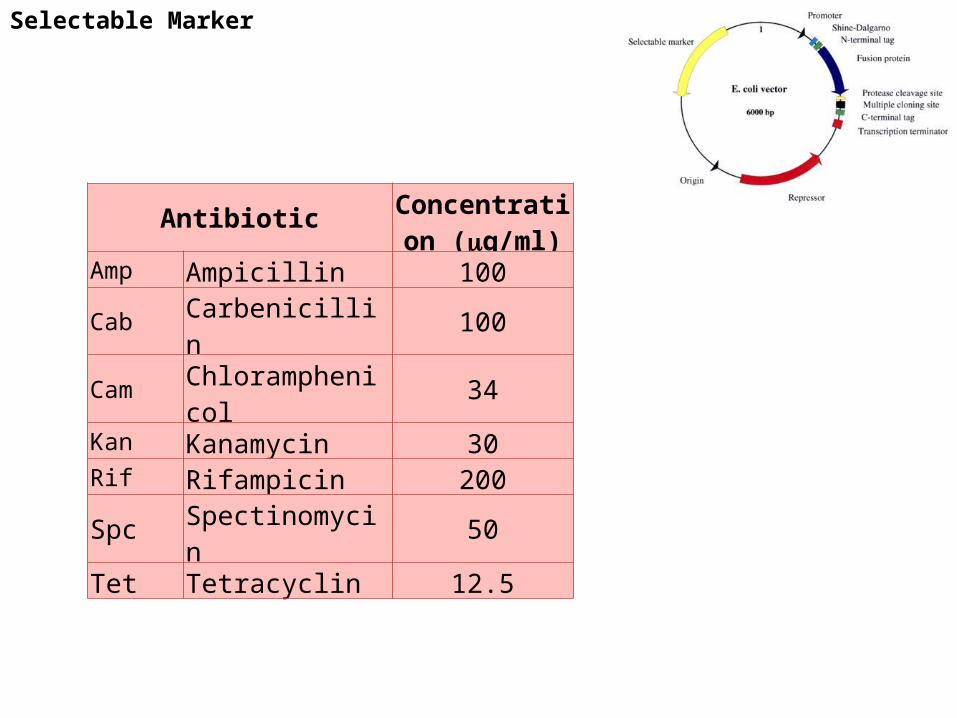
Selectable Marker
Antibiotic Concentration (g/ml)
Amp Ampicillin 100Cab Carbenicillin 100Cam Chloramphenicol 34Kan Kanamycin 30Rif Rifampicin 200Spc Spectinomycin 50Tet Tetracyclin 12.5

Regulatory Gene (Repressor)
Many promoters show leakiness in their expression i.e. gene products are expressed at low level before the addition of the inducer. This becomes a problem when the gene product is toxic for the host
The lac-derived promoters are especially leaky.
These promoters can be controlled by the insertion of a lac-operator sequence downstream the promoter and the expression of the lac-repressor by host strains carrying the lacIq allele (for medium copy number plasmids) or from the same or a helper plasmid (for higher copy number plasmids).
Alternatively, repression can be achieved by the addition of 1% glucose to the culture medium.

Origin of Replication
The origin of replication controls the plasmid copy number.
Promoter
positioned 10-100 nucleotides upstream of the ribosome-binding site.
The ideal promoter exhibits several desirable features: (i) It is strong enough to allow product accumulation up to 50% of the total cellular protein. (ii) It has a low basal expression level (i.e., it is tightly regulated to prevent product toxicity). (iii) It is easy to induce.
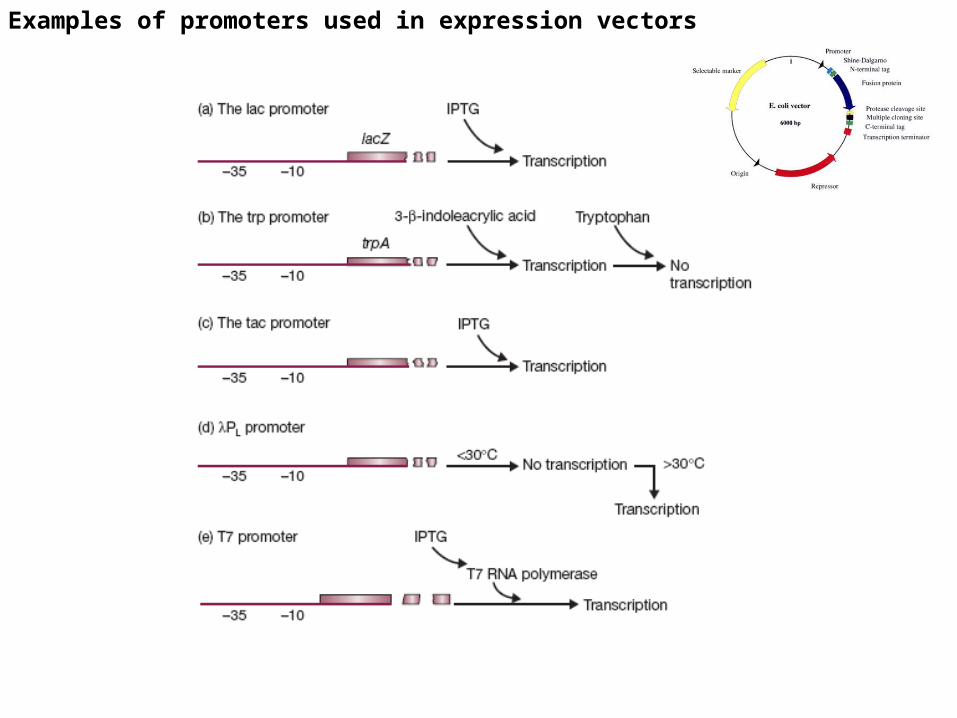
Examples of promoters used in expression vectors

Transcription Terminator
The terminator, which marks the point at the end of the gene where transcription should stop.
A terminator is usually a nucleotide sequence that can base pair with itself to form a stem–loop structure.
The transcription terminator reduces unwanted transcription and increases plasmid and mRNA stability.
Shine-Delgarno Sequence
The Shine-Dalgarno (SD) sequence is required for translation initiation and is complementary to the 3'-end of the 16S ribosomal RNA.
The concensus sequence is: 5'-TAAGGAGG-3'. It is positioned 4-14 nucleotides upstream the start codon with the optimal spacing being 8 nucleotides.
To avoid formation of secondary structures (which reduces expression levels) this region should be rich in A residues.
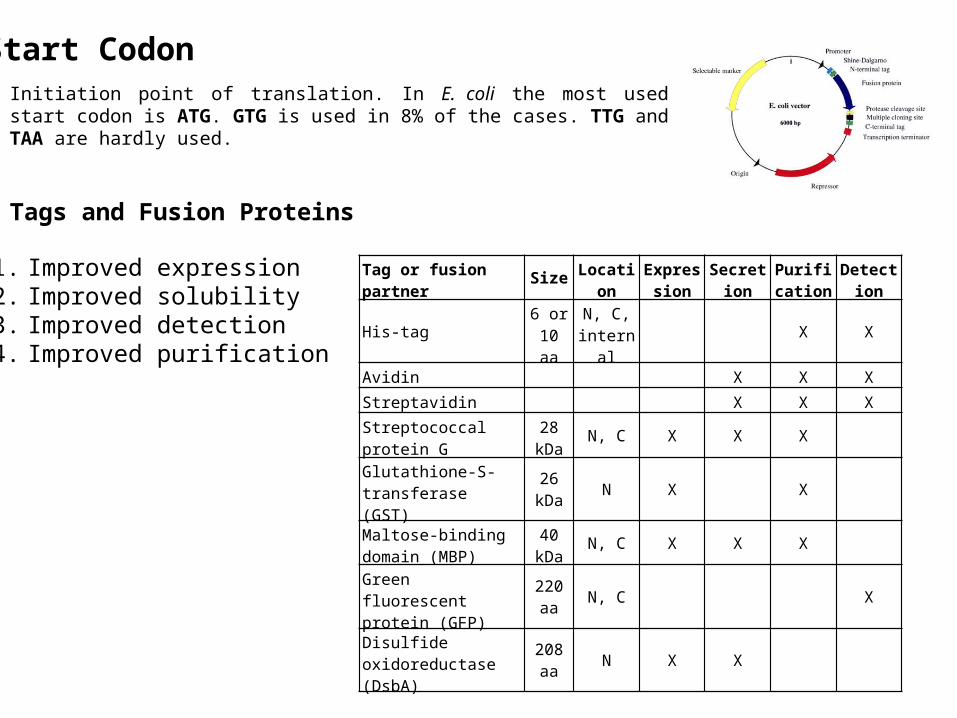
Start Codon
Initiation point of translation. In E. coli the most used start codon is ATG. GTG is used in 8% of the cases. TTG and TAA are hardly used.
Tags and Fusion Proteins
1. Improved expression2. Improved solubility3. Improved detection4. Improved purification
Tag or fusion partner Size Location Expression
Secretion
Purification
Detection
His-tag 6 or10 aa
N, C, internal X X
Avidin X X X
Streptavidin X X X
Streptococcal protein G 28 kDa N, C X X X
Glutathione-S-transferase (GST)
26 kDa N X X
Maltose-binding domain (MBP)
40 kDa N, C X X X
Green fluorescent protein (GFP) 220 aa N, C X
Disulfide oxidoreductase (DsbA) 208 aa N X X
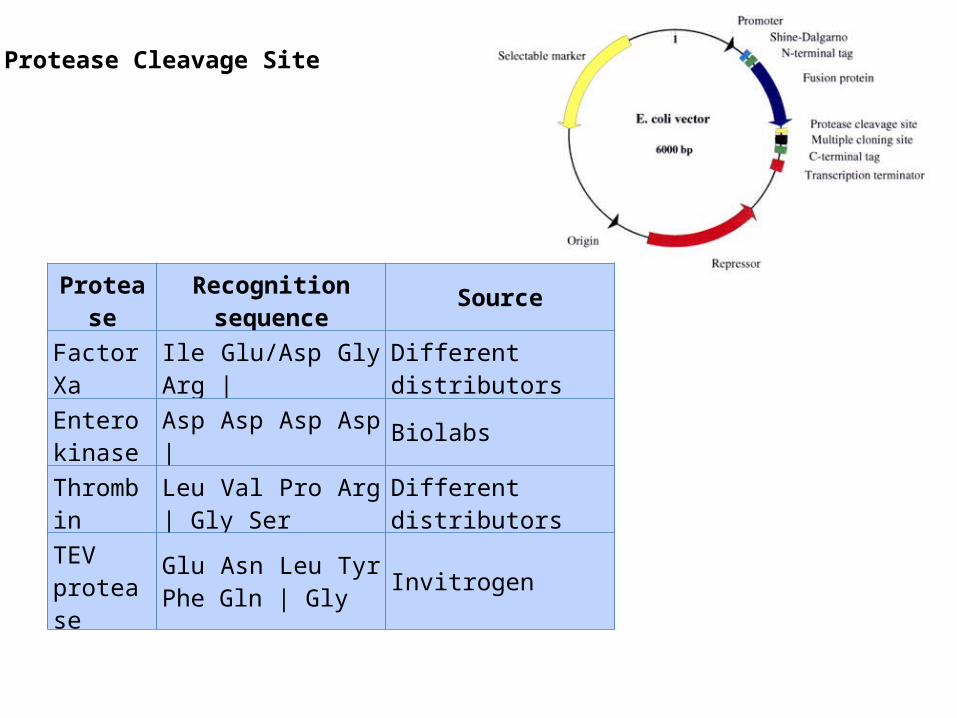
Protease Cleavage Site
Protease Recognition sequence Source
Factor Xa Ile Glu/Asp Gly Arg | Different distributors
Enterokinase Asp Asp Asp Asp | Biolabs
Thrombin Leu Val Pro Arg | Gly Ser Different distributors
TEV protease
Glu Asn Leu Tyr Phe Gln | Gly Invitrogen

Multiple Cloning Sites
A series of unique restriction sites that enables you to clone your gene of interest into the vector.
Stop Codon
Termination of translation. There are 3 possible stop codons but TAA is preferred because it is less prone to read-through than TAG and TGA. The efficiency of termination is increased by using 2 or 3 stop codons in series.
p E G1 634 75 bp
Te tR
A m p Rte tA '
o m p A 'S tre p ta g
ttp lpp
R B S te tA
p A C Y C 1 8 4 or i
-1 0Te tP -3 5
Bam H I (1211)
Eco R I (119 0 )
H ind III (1271)
Pst I (1233)
Sma I (120 8 )
X ma I (120 6 )
Acc 6 5I (120 2)Afe I (1239 )
Bsa I (1177)
Bsp MI (1236)
Eco ICR I (119 8 )
H a eI (1179)
K pnI (120 6 )
Sa c I (120 0 )
Sa l I (1223)
Sbf I (1233)
Sci I (1219)
Stu I (1179)
UthSI (120 7)
X ba I (10 9 8 )
X hoI (1217)

General Problems with the Production of Recombinant Protein in E. coli
1. Problems Resulting from the Sequence of the Foreign Gene
2. Problems Caused by E. coli
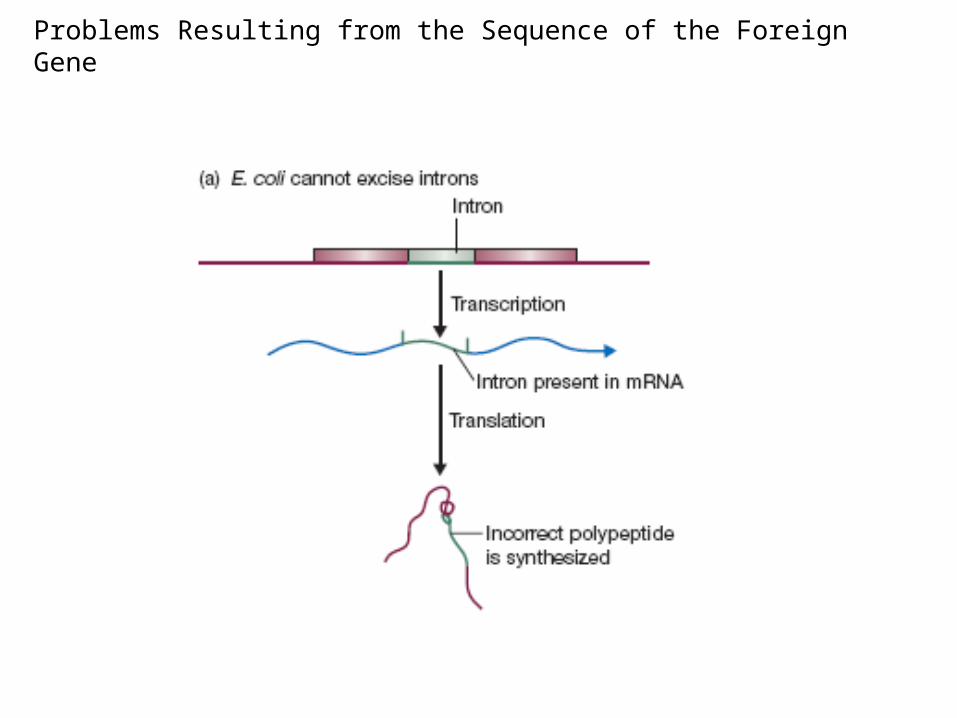
Problems Resulting from the Sequence of the Foreign Gene
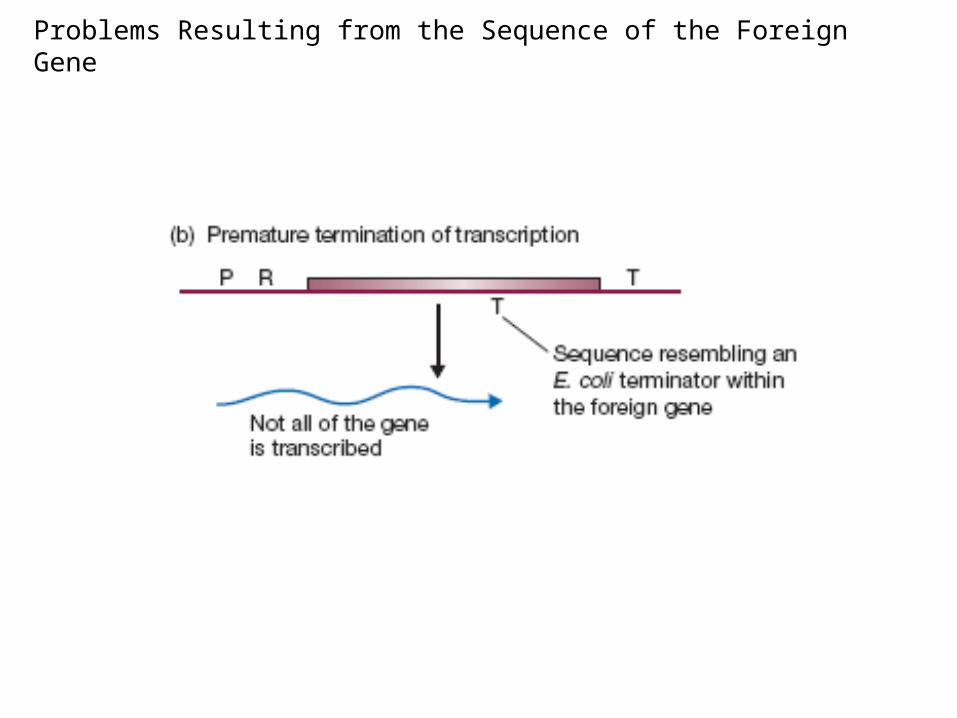
Problems Resulting from the Sequence of the Foreign Gene
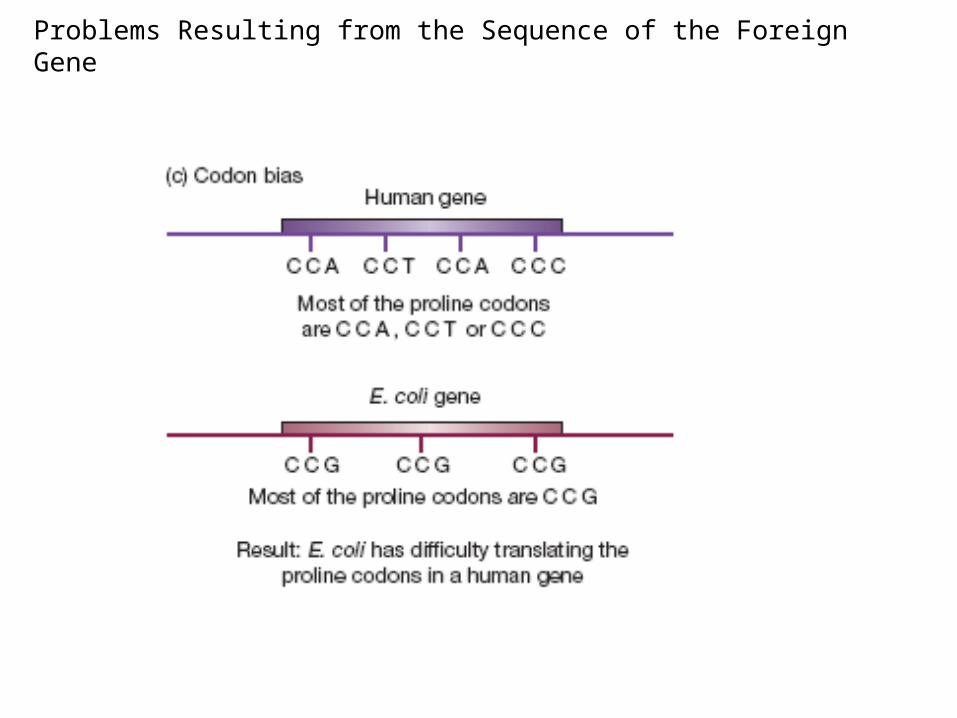
Problems Resulting from the Sequence of the Foreign Gene

Problems Caused by E. coli
1. E. coli might not process the recombinant protein correctly.
The proteins of most organisms are processed after translation, by chemical modification of amino acids within the polypeptide.
2. E. coli might not fold the recombinant protein correctly, and generally is unable to synthesize the disulphide bonds present in many animal proteins.
If the protein does not take up its correctly folded tertiary structure, then usually it is insoluble and forms an inclusion body within the bacterium
3. E. coli might degrade the recombinant protein.

In Vitro Translation
Application of In Vitro Translation
The in vitro synthesis of proteins in cell-free extracts is an important tool for molecular biologists and has a variety of applications, including the
(1)rapid identification of gene products (e.g., proteomics),
(2)localization of mutations through synthesis of truncated gene products,
(3)protein folding studies, and
(4)incorporation of modified or unnatural amino acids for functional studies.

In Vitro Translation
Advantages of in vitro Translation
The use of in vitro translation systems can have advantages over in vivo gene expression:
(1) when the over-expressed product is toxic to the host cell,
(2) when the product is insoluble or forms inclusion bodies, or
(3) when the protein undergoes rapid proteolytic degradation by intracellular proteases.

Cell-Free Expression Systems
The most frequently used cell-free translation systems consist of extracts from:
(1)Rabbit reticulocytes,
(2)Wheat germ and
(3)Escherichia coli.
In vitro protein synthesis has two approaches based on starting materials:
1. Standard translation system: start with RNA template, eg 1 and 22. Coupled and linked system: start with DNA template, eg. 3

Rabbit Reticulocyte Lysate
•In vivo, reticulocytes are highly specialized cells primarily responsible for the synthesis of haemoglobin, which represents more than 90% of the protein made in the reticulocyte.
•These immature red cells have already lost their nuclei, but contain adequate mRNA, as well as complete translation machinery, for extensive globin synthesis.
•The endogenous globin mRNA can be eliminated by incubation with Ca2+-dependent micrococcal nuclease, which is later inactivated by chelation of the Ca2+ by ethyleneglycol tetraacetic acid (EGTA).
•Ambion offers a nuclease-treated reticulocyte lysate. This type of lysate is the most widely used RNA-dependent cell-free system because of its low background and its efficient utilization of exogenous RNAs even at low concentrations
•Exogenous proteins are synthesized at a rate close to that observed in intact reticulocyte cells.
•This type of lysate is typically used for studying the translation machinery, e.g. studying the effects of inhibitors on globin translation.
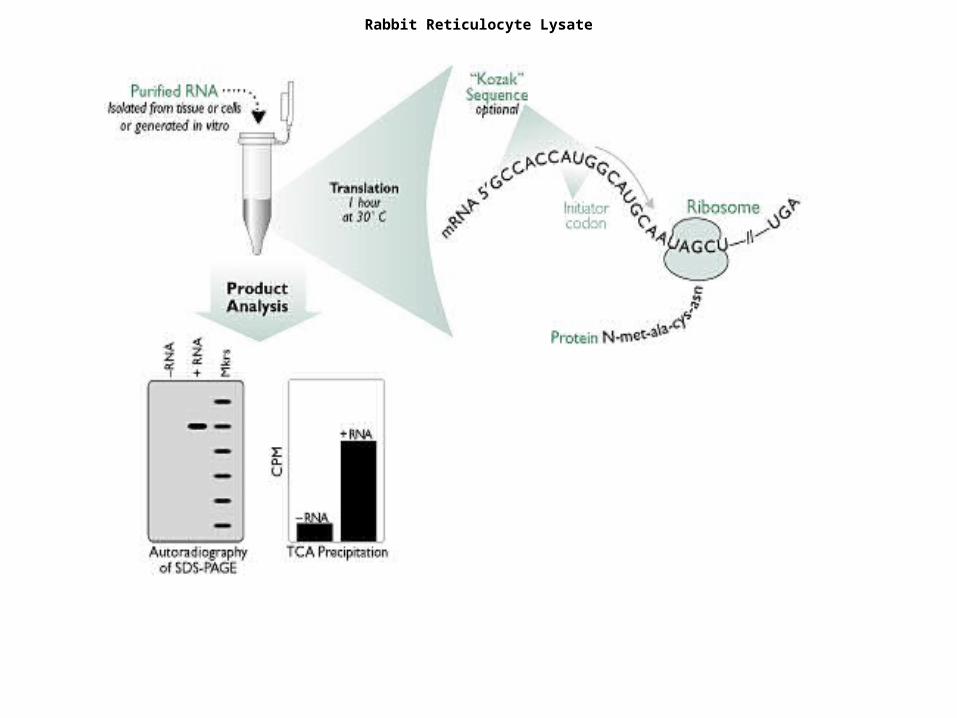
Rabbit Reticulocyte Lysate

Wheat Germ Extract
•alternative to the rabbit reticulocyte lysate cell-free system
•Wheat germ lysate efficiently translates exogenous RNA from a variety of different organisms, from viruses and yeast to higher plants and mammals.
•When using RNA synthesized in vitro, the presence of a 5' cap structure may enhance translational activity.
•Typically, translation by wheat germ extracts is more cap-dependent than translation by retic extracts.

E. coli Cell-Free System
•E. coli cell-free systems consist of a crude extract that is rich in endogenous mRNA.
• The extract is incubated during preparation so that this endogenous mRNA is translated and subsequently degraded.
•Because the levels of endogenous mRNA in the prepared lysate is low, the exogenous product is easily identified.
•In comparison to eukaryotic systems, the E.coli extract has a relatively simple translational apparatus with less complicated control at the initiation level, allowing this system to be very efficient in protein synthesis.
• E. coli extracts are ideal for coupled transcription : translation from DNA templates.

Linked and Coupled Transcription:Translation Systems
•In standard translation reactions, purified RNA is used as a template for translation, “linked” and “coupled” systems, on the other hand, use DNA as a template.
• DNA templates for transcription:translation reactions may be cloned into plasmid vectors or generated by PCR.
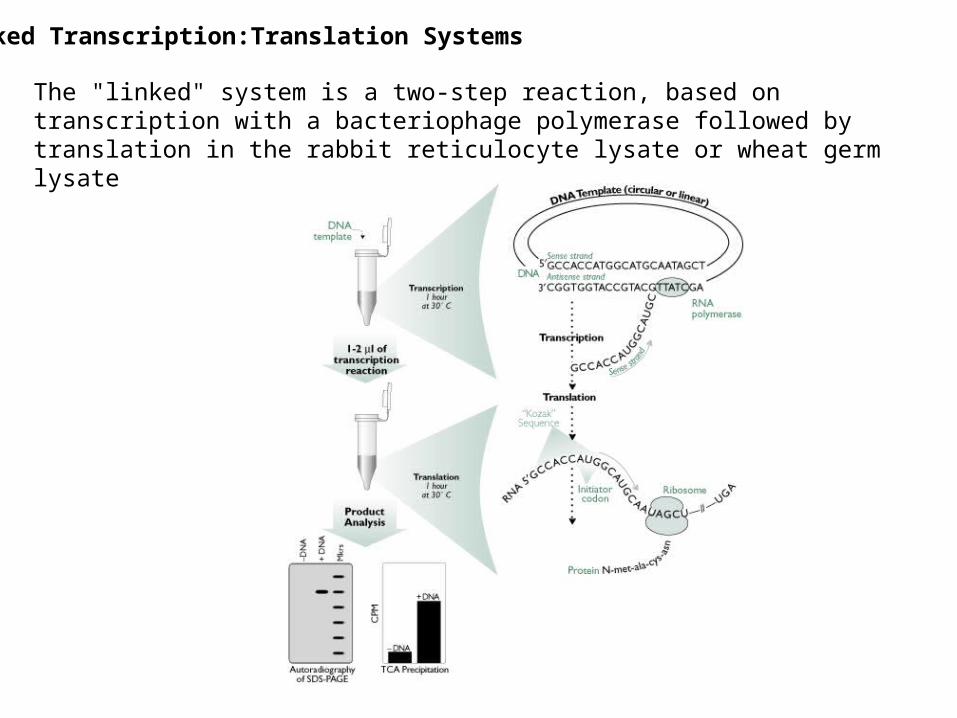
Linked Transcription:Translation Systems
The "linked" system is a two-step reaction, based on transcription with a bacteriophage polymerase followed by translation in the rabbit reticulocyte lysate or wheat germ lysate
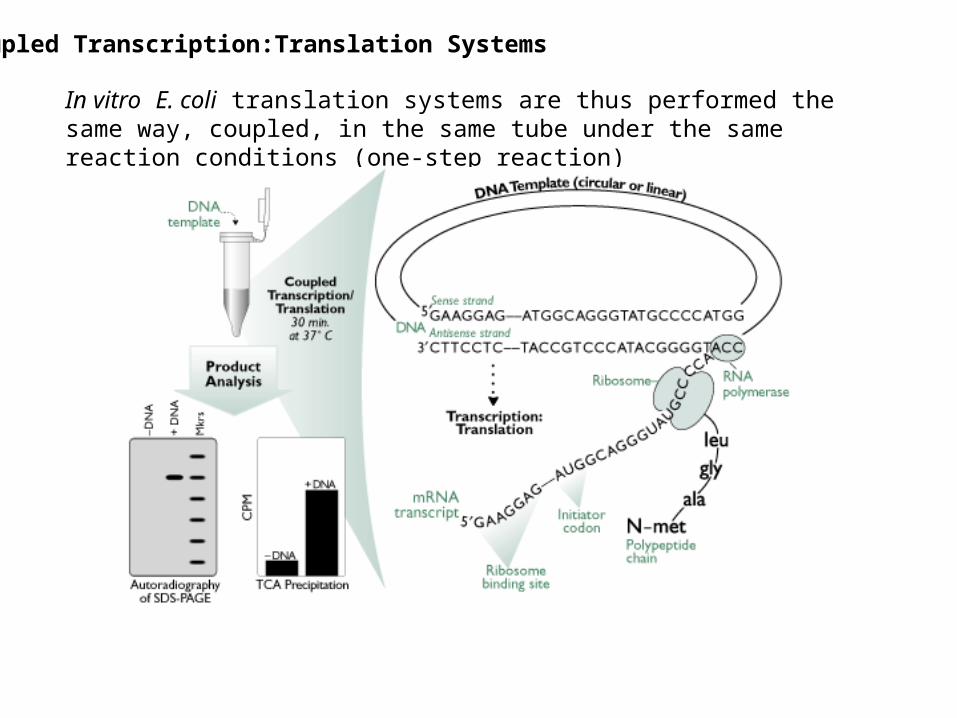
Coupled Transcription:Translation Systems
In vitro E. coli translation systems are thus performed the same way, coupled, in the same tube under the same reaction conditions (one-step reaction)

Important Elements for TranslationThere are some significant differences between prokaryotic and eukaryotic mRNA transcripts.
Eukaryotic mRNA Transcripts Prokaryotic mRNA Transcripts
Typically, eukaryotic mRNAs are characterized by two post-transcriptional modifications: (i) a 5'-7-methyl-GTP cap (ii) (ii) a 3'-poly(A) tail. (iii) Both modifications contribute to the
stability of the mRNA by preventing degradation.
In bacteria, the ribosome is guided to the AUG initiation site by a purine-rich region called the Shine-Dalgarno (SD) sequence.
This sequence is complementary to the 3' end of the 16s rRNA in the 30S ribosomal subunit.
The 5' cap structure enhances the translation of mRNA by helping to bind the eukaryotic ribosome and assuring recognition of the proper AUG initiator codon.
Upstream from the initiation AUG codon, the SD region has the consensus sequence 5'-UAAGGAGGUGA-3'.

Important Elements for TranslationThere are some significant differences between prokaryotic and eukaryotic mRNA transcripts.
Eukaryotic mRNA Transcripts Prokaryotic mRNA Transcripts
The consensus sequence 5'-GCCACCAUGG-3', also known as the "Kozak" sequence, is considered to be the strongest ribosomal binding signal in eukaryotic mRNA.
Specific mRNAs vary considerably in the number of nucleotides that complement the anti-Shine-Dalgarno sequence of 16S rRNA, ranging from as few as two to nine or more.
For efficient translation initiation, the key elements are the G residue at the +1 position and the A residue at the -3 position.
An mRNA that lacks the Kozak consensus sequence may be translated efficiently in eukaryotic cell-free systems if it possesses a moderately long 5'-untranslated region (UTR) that lacks stable secondary structure.
The position of the ribosome-binding site (RBS) in relation to the AUG initiator is very important for efficiency of translation (usually from -6 to -10 relative to the A of the initiation site).

Production of Recombinant Protein by Eukaryotic Cells
Saccharomyces cerevisiae as the Host for Recombinant Protein Synthesis
•The yeast Saccharomyces cerevisiae is currently the most popular microbial eukaryote for recombinant protein production.
•Cloned genes are often placed under the control of the GAL promoter which is normally upstream of the gene coding for galactose epimerase, an enzyme involved in the metabolism of galactose.
•Other useful promoters are PHO5, which is regulated by the phosphate level in the growth medium, and CUP1, which is induced by copper.
•Most yeast expression vectors also carry a termination sequence from an S. cerevisiae gene, because animal termination signals do not work effectively in yeast.

Production of Recombinant Protein by Eukaryotic Cells
Saccharomyces cerevisiae as the Host for Recombinant Protein Synthesis
Yields of recombinant protein are relatively high, but
(1) S. cerevisiae is unable to glycosylate animal proteins correctly, often adding too many sugar units (“hyperglycosylation”), although this can be prevented or at least reduced by using a mutant host strain.
(2) S. cerevisiae also lacks an efficient system for secreting proteins into the growth medium. In the absence of secretion, recombinant proteins are retained in the cell and consequently are less easy to purify.
(3) Codon bias can also be a problem.

Using Animal Cells for Recombinant Protein Production
A problem with some animal cell lines is that they require a solid surface on which to grow, adding complications to the design of the culture vessels.
(1) One solution is to fill the inside of the vessel with plates, providing a large surface area, but this has the disadvantage that complete and continuous mixing of the medium within the vessel becomes very difficult.
(2) A second possibility is to use a standard vessel but to provide the cells with small inert particles (e.g., cellulose beads) on which to grow.
Protein Production in Mammalian Cells

Using Animal Cells for Recombinant Protein Production
•Insect cells provide an alternative to mammalian cells for animal protein production.
•Insect cells do not behave in culture any differently to mammalian cells but they have the great advantage that, thanks to a natural expression system, they can provide high yields of recombinant protein.
Protein Production in Insect Cells

•The expression system is based on the baculoviruses, a group of viruses that are common in insects but do not normally infect vertebrates.
•The baculovirus genome includes the polyhedrin gene, whose product accumulates in the insect cell as large nuclear inclusion bodies toward the end of the infection cycle
The Expression System Based on Baculoviruses
•The product of this single gene frequently makes up over 50% of the total cell protein.
•Similar levels of protein production also occur if the normal gene is replaced by a foreign one.
• Baculovirus vectors have been successfully used in production of a number of mammalian proteins, but unfortunately the resulting proteins are not glycosylated correctly.
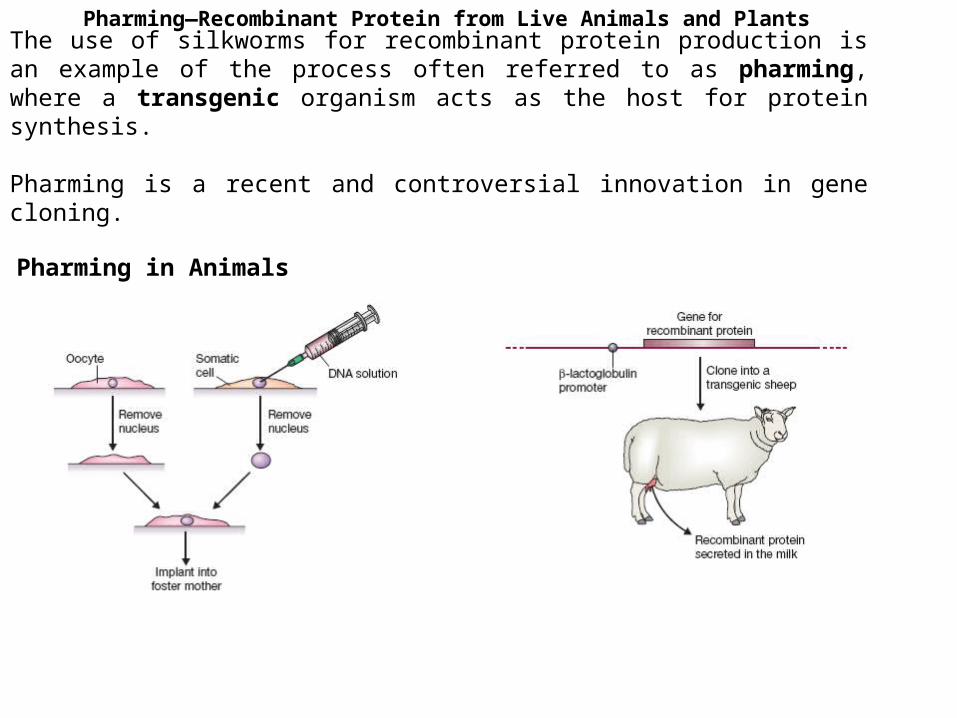
Pharming—Recombinant Protein from Live Animals and Plants
The use of silkworms for recombinant protein production is an example of the process often referred to as pharming, where a transgenic organism acts as the host for protein synthesis.
Pharming is a recent and controversial innovation in gene cloning.
Pharming in Animals

Pharming—Recombinant Protein from Live Animals and Plants
Recombinant Proteins from Plants
•Plants provide the final possibility for production of recombinant protein.
•Plants and animals have similar protein processing activities, although there are slight differences in the glycosylation pathways
•Plant cell culture is a well established technology that is already used in the commercial synthesis of natural plant products.
•Alternatively, intact plants can be grown to a high density in fields.













![The 60-Kilodalton Protein Encoded by orf2 in the cry19A ...operon in pHT315 (A. Delécluse, Institut Pasteur, Paris, France [19]). Cloned genes and recombinant plasmid constructs were](https://img.pdfslide.us/doc/110x75/5e3399390bb9fc1ac711d68d/the-60-kilodalton-protein-encoded-by-orf2-in-the-cry19a-operon-in-pht315-a.jpg)