Embed Size (px)
Citation preview

PRODUCTION OF MEDIUM-CHAIN-LENGTHPOLY(3-HYDROXYALKANOATES) FROMCRUDE FATTY ACIDS MIXTURE BYPSEUDOMONAS PUTIDA
M. S. M. Annuar1,�, I. K. P. Tan1, S. Ibrahim2 and K. B. Ramachandran3
1Institute of Biological Sciences, Faculty of Science.2Department of Civil Engineering, University of Malaya, Kuala Lumpur, Malaysia.3Department of Biotechnology, Indian Institute of Technology (IIT) Madras, Chennai, India.
Abstract: Microbial production of medium-chain-length poly(3-hydroxyalkanoates), PHAMCL
from crude fatty acids mixture was investigated. With ammonium as the growth limiting substrate,fatty acids mixture from saponified palm kernel oil (SPKO) supports good growth and PHAMCL
production of Pseudomonas putida PGA1. Growth of this microorganism on ammonium exhibitedsubstrate inhibition kinetics which can be described using Andrews model with the substrateinhibition constant, Ki ¼ 1.2 g L21. Concentration of SPKO in the aqueous medium should beat 10 g L21 or less, as higher concentrations can significantly reduced the volumetric oxygentransfer coefficient (KLa), biomass growth and PHAMCL production. Uptake of SPKO by theorganism follows a zero-order kinetics, indicating a mass transfer limitation of the free fattyacids by the P. putida PGA1 cells. In batch and fed-batch fermentations, PHAMCL accumulationis encouraged under ammonium-limited condition with SPKO as the sole carbon and energysource. The amount of PHAMCL accumulated and its specific production rate, qPHA were influ-enced by the residual ammonium concentration level in the culture medium. It was observedthat in both fermentation modes, when the residual ammonium becomes exhausted(,0.05 g L21), the PHAMCL accumulation and qPHA were significantly reduced. However, thiseffect can be reversed by feeding low amount of ammonium to the culture, resulting in signifi-cantly improved PHAMCL yield and productivity. It is concluded that the feeding of residualammonium concentration in the culture medium during the PHAMCL accumulation has a positiveeffect on sustaining the PHAMCL biosynthetic capability of the organism.
Keywords: poly-(3-hydroxyalkanoates); medium-chain-length; kinetics; ammonium;palm kernel oil.
INTRODUCTION
Poly(3-hydroxyalkanoates) (PHA) is a poly-mer of biological origin produced by a widevariety of microorganisms. The biopolymerhas similar characteristics as the petro-chemical derived plastics (Hocking andMarchessault, 1994). It has a huge potentialof being an alternative to the synthetic plasticsin numerous applications which is furthermade attractive by the fact that the PHA isreadily biodegradable. There are two majorgroups of PHAs: the short-chain-length PHA(PHASCL) with five or less carbon atoms,and medium-chain-length PHA (PHAMCL)with six or more carbon atoms. Only thePHASCL have been commercially producedup to 500 tons year21, which was manufac-tured by Monsanto (Kellerhals et al., 2000).
The PHAMCL is yet to make a significantimpact as a viable choice due to the factthat it is very expensive to produce this poly-mer in bulk amount even for material testingpurposes. To date, the final PHAMCL yieldand content obtained is relatively lower com-pared to those of PHASCL, which hampereddevelopment of its applications (Lee et al.,2000). PHAMCL copolymers can be producedusing a variety of substrates including plantoils. Due to their long carbon number, thesesubstrates have high energy content whichis excellent for good cell growth and energymetabolism. The variation in the compositionof plant oils directly influences the monomercomposition, which allows production ofPHAMCL with a wide range of material proper-ties. Preliminary studies on the production ofPHAMCL from renewable and cost-effective
104 Vol 85 (C2) 104–119
�Correspondence to:Dr M.S.M. Annuar, Instituteof Biological Sciences,Faculty of Science,University of Malaya,Kuala Lumpur 50603,Malaysia.E-mail: [email protected]
DOI: 10.1205/fbp06017
0960–3085/07/$30.00þ 0.00
Food and BioproductsProcessing
Trans IChemE,Part C, June 2007
# 2007 Institutionof Chemical Engineers

substrates like palm oil, palm kernel oil and their major fattyacids fractions by Pseudomonas putida have been reported(Tan et al., 1997).PHA accumulation by bacteria is a response to the imbal-
ance in growth environment, with excess carbon source andsimultaneous limitation of nutrients such as nitrogen, phos-phorus, oxygen, magnesium and so on, this physiologicalcondition can be exploited in the fermentation process toachieve high yields and productivity. Ammonium ion is usuallychosen as the limiting nutrient as it is relatively easier to makea bacterial culture ammonium-limited than other mineral ions;in addition to the growth of microorganisms is more depen-dent on nitrogen, it is also assimilated more rapidly than areother mineral ions (Suzuki et al., 1986a).
Several PHAMCL production strategies in the bioreactorsuch as batch and continuous (Durner et al., 2001; Junget al., 2001), fed-batch (Beom, 2002) and high-cell-densityprocess (Lee et al., 2000) under various cultivation conditionshave been described. Crude plant oils, which are renewableand economical, are excellent fermentation feedstock forbiomass growth and PHAMCL production by microorganisms,owing mainly to their highly reduced nature and structuralsimilarity to the PHAMCL itself. The work presented heredetails the investigation on the production of PHAMCL inbatch and fed-batch bioreactor using crude fatty acidsmixture from palm kernel oil, namely the saponified palmkernel oil (SPKO), as sole carbon and energy source. Import-ant aspects e.g., effect of SPKO concentration on the volu-metric oxygen transfer coefficient (KLa), effect of limitingsubstrate (ammonium) on growth, effect of carbon-to-nitrogen(C/N) ratio on PHAMCL production and kinetics of fatty acidsmixture utilization were studied to better understand the bio-process and to provide useful guidelines in a rational processdesign.
METHODS AND MATERIALS
Microorganism
Pseudomonas putida PGA1 strain was a gift from Pro-fessor G. Eggink of the Agrotechnological Research Institute,Wageningen, The Netherlands.
Medium Composition
Medium for both inoculum cultivation and bioreactorstudies was the mineral salt solution containing (in g L21):5.74 K2HPO4, 3.7 KH2PO4, 1.0 ml trace elements (MT) sol-ution (Lageveen et al., 1988), 10.0 ml 0.1 M MgSO4
. 7H2O.Saponified palm kernel oil (SPKO) was supplied as the solecarbon and energy source with NaNH4HPO4
.H2O as thelimiting nutrient. Saponification of palm kernel oil (PKO)was carried out according to Tan et al. (1997). PKO is theextract from the nut of the oil palm (Elaeis guineensisJacq.) fruit. The oil consists of a mixture of C6–C18 : 2 fattyacids with approximately 82% saturated fatty acids and18% unsaturated fractions (Table 1). The same medium com-position was used for inoculum and fermenter cultivations toreduce or eliminate lag period.To avoid precipitation during autoclaving, solutions of
magnesium salt, trace elements and SPKO (pH 7.0) weresterilized separately before adding to the rest of themedium components.
To grow significant amount of biomass without PHAMCL
production, rich medium was used, which consists of(g L21): 10.0 yeast extract, 15.0 nutrient broth and 5.0(NH4)2SO4 (Hori et al., 1994).
Inoculum
The inoculum was grown in shake-flasks at 308C and250 rpm rotation. During active growth at 18 h, the whole con-tent of the flask was used to seed the bioreactor. This corre-sponds to biomass dry weight of 0.10–0.14 (+14%) g L21.
Bioreactor
Fermenter experiments were performed in a BiostatwB3-litre fermenter (B. Braun Biotech International). A roundbottom culture vessel (Type B2) made of borosilicate glassand stainless steel equipped with an outer thermostatjacket was used. The main geometric characteristics of thefermenter are shown in Table 2. Temperature, pH and partialoxygen pressure (pO2) were measured, monitored and con-trolled by a digital system.
Table 1. Fatty acids composition of palm kernel oil (Elson, 1992).
Fatty acids Percentage (%)
C6:0 (caproic acid) 0.2C8:0 (caprylic acid) 3.0C10:0 (capric acid) 4.0C12:0 (lauric acid) 48.0C14:0 (myristic acid) 16.0C16:0 (palmitic acid) 8.0C18:0 (stearic acid) 3.0C18:1 (oleic acid) 15.4C18:2 (linoleic acid) 2.4C20:0 (arachinoic acid) 0.1SFAa 82.2
(SCFA)b 7.2(MCFA)c 64.0(LCFA)d 11.1
MUFAe 15.4PUFAf 2.5
aSaturated fatty acids; bshort chain SFA; cmedium chain SFA; dlongchain SFA; emonounsaturated fatty acids; fpolyunsaturated fattyacids.
Table 2. Dimensions of the fermenter and its components.
Design parameters Specifications
Total volume 3 litresDiameter of inner tank 130 mmHeight of tank 240 mmNumber of baffles 4Baffle width 10.5 mmType of impellers Rushton disc turbineNumber of impellers 2Distance between impellers 79.5 mmDistance of lower impeller from bottom plate 25 mmImpeller diameter of disc 53 mmNumber of blades 6Impeller blade width 10.5 mmImpeller blade length 14.5 mmDiameter of single ring sparger 48 mmNumber of holes 14Distance of ring sparger from bottom plate 20 mmDiameter of oxygen electrode 12 mm
Trans IChemE, Part C, Food and Bioproducts Processing, 2007, 85(C2): 104–119
MEDIUM-CHAIN-LENGTH POLY(3-HYDROXYALKANOATES) FROM CRUDE FATTY ACIDS 105

Estimation of Volumetric Oxygen TransferCoefficient (KLa)
The estimation of volumetric oxygen transfer coefficient,KLa was carried out in a batch bioreactor using static gas-sing-out method. The aqueous phase consisted of actualcomposition of the fermentation medium in 1.0 litre distilledwater. Increasing SPKO concentrations i.e., (g L21): 1.0,5.0, 10.0, 15.0 and 20.0 were tested. As a control, KLavalue of distilled water was also determined under similaroperating conditions. The aqueous medium was first deoxy-genated by sparging gaseous nitrogen until all traces ofoxygen was stripped away. Then air was sparged into thebioreactor at 0.6 L min21 and at 600 rpm agitation rate. Theincrease in the oxygen partial pressure (%pO2) was recordedat regular intervals until readings become almost constantwhich indicated the saturation of the liquid medium withoxygen.The KLa was determined using the relationship:
dCL=dt ¼ KLa(C�L � CL) (1)
where CL�
is the %pO2 saturation value and CL is the %pO2 inthe aqueous medium. Upon integration, equation (1) trans-formed into
ln (C�L � CL) ¼ �KLa � t þ lnC�L (2)
Plotting of ln (CL�
2 CL) versus t yields a straight line withthe KLa value determined directly from the slope.
Electrode Response Time
The main disadvantage of the calculation of KLa usingstatic gassing out method is the slow response time of theelectrode changes of oxygen concentration in the liquid.Response time is defined as the time required by the elec-trode to measure 63% of the global value of the change inthe concentration of oxygen and it is related to the diffusionof oxygen through the membrane of the electrode. In orderfor the measured values to be sufficiently reliable, theresponse time of the electrode must be smaller or equal to1/KLa (Montes et al., 1999). Therefore, an estimation of theresponse time of the oxygen electrode (Mettler-Toledo12/200 A Type) used throughout the experiments wasmade by measuring the variation of oxygen concentrationafter the electrode was moved from a saturated aqueoussolution of nitrogen to a saturated oxygen solution. Theresults indicate that 63% of oxygen saturation was reachedat 31.9 (+2.2) s and 100% was reached at around 67.6(+1.5) s. Thus, only values of KLa less than 0.03125 s21
should be considered reliable when using this electrode.
Ammonium Ion Inhibition Studies
A rapid and simple batch experiment using shake-flasksbased on initial ammonium concentrations was carried out.Each ammonium ion concentration was prepared in triplicateshake-flasks with the SPKO supplied at 5.0 g L21 in themineral salt medium. In order to eliminate the lag periodupon cells transfer to the medium with the respectiveammonium concentrations, inoculum was grown using thesame medium composition with 0.4 g L21 ammonium and
10.0 g L21 SPKO. Three flasks containing the inocula wereincubated for 24 h. Cells were aseptically harvested by cen-trifugation and washed twice with saline solution to get ridof the residual ammonium. Cells pellet from the three flaskswas mixed and re-suspended in the mineral medium minusthe ammonium and SPKO. This cells suspension was usedto inoculate the flasks with different ammonium ion concen-trations. The ammonium ion concentrations used were(g L21): 0.008, 0.04, 0.08, 0.24, 0.40, 0.56 and 0.80 with0.5% (w/v) SPKO. The starting optical density of all culturesat 450 nm (OD450) was approximately 0.1206 (+0.001)units. Samples were withdrawn at regular intervals andcentrifuged. The collected cells were washed twice in salineprior to optical density determination at 450 nm (OD450)with saline as blanks. If necessary, appropriate dilution ofthe cells with saline was performed so that the OD450recorded is within the linear range of pre-determinedcalibration.Overall specific growth rate, m (h21) was calculated accord-
ing to Bitar and Underhill (1990), described in equation (3):
m ¼ ( lnOD450t2 � lnOD450t1)=(t2 � t1) (3)
where OD450t1 being the optical density at 450 nm at time t1,OD450t2 is the optical density at 450 nm at time t2, and t2 2 t1is the sampling interval.
Effect of Carbon-to-Nitrogen (C/N) Ratio
The effect of different initial C/N ratio on yield of biomassand PHAMCL, and on PHAMCL content of the cells wasstudied using shake-flasks. Cells were first grown in therich medium to obtain biomass without PHAMCL. After cultiva-tion for 24 h, cells were aseptically harvested by centrifu-gation at 48C. This was transferred aseptically into mineralmedium for PHAMCL production. All flasks were inoculatedwith 2.5 (+0.3) g L21 cells. The sole carbon and energysource was SPKO with ammonium as the nitrogen substrate.The cultivation was carried out for 36 h whereby the cellswere harvested by centrifugation at 48C. Pellets werewashed twice with saline, and then with methanol beforeoven-dried (�508C) until constant weight. PHAMCL wasobtained from dried biomass using solvent extraction(Tan et al., 1997).
Batch Fermentation
Inoculum was added at 10% v/v of the fermenter workingvolume. The temperature and the pH were maintained at 30(+0.5)8C and 7.0 (+0.05), respectively; with the agitationrate of 600 rpm and the aeration rate of 0.6 L min21 of filteredair. Silicone anti-foaming agent (BDH) was included in thefermenter at 1.0 g L21. Samples were withdrawn at regularintervals.
Fed-Batch Fermentation
The fermentation was carried out using fermenter setupas in the batch with the addition of the feeding routine.The fermentation was performed as batch start-upand at 9 h the feeding commenced. Feeding medium(0.16 g ammonium in 0.4 L) consisted of fixed ratio of
Trans IChemE, Part C, Food and Bioproducts Processing, 2007, 85(C2): 104–119
106 ANNUAR et al.

SPKO-to-ammonium (6.9 mole: 1 mole) in the mineral saltsolution. Feeding was carried out using Masterflex7521–35 peristaltic pump at a constant flow rate of0.14 L h21 (0.056 g ammonium h21). The contents of thefeed reservoir were continuously stirred using magneticstirrer to obtain a homogenous feed into the fermenter.
Analytical Methods
All measurements were performed in triplicates.Total biomass concentration was estimated by first spin-
ning down the cells in pre-weighed micro-centrifuge tubes.The cells were then washed twice with saline solution anddried at 908C till constant weight.The residual ammonium in the cell-free culture medium
was determined using spectrophotometric method (Solor-zano, 1969).The residual free fatty acids from SPKO in the culture
medium supernatant were estimated by solvent titration(Cocks and van Rede, 1966).To obtain PHAMCL production profile, 8.0 mg of dried cells
were subjected to acid-catalysed methanolysis and thePHA amount and composition were determined by gaschromatography (GC) using benzoic acid methyl ester asthe internal standard (Brandl et al., 1988). The gas chromato-graph used was Varian Star 3400CX equipped with a fusedsilica capillary column (30 m � 0.53 mm ID) (SupelcoSPBTM-608) and a flame ionization detector.For molecular weight determination, the PHA was first
extracted by refluxing 1.0 g cells (dry weight) in 200 mLchloroform for 4 h. The mixture was filtered to remove celldebris and the filtrate was concentrated by rotary evapor-ation. The concentrated chloroform was added slowly to arapidly stirred cold methanol to precipitate the PHA. Thefilm was re-dissolved in chloroform and precipitated againin cold methanol to purify the polymer. The PHA film obtainedwas air-dried.The average molecular mass and molecular mass distri-
bution of the extracted PHA were obtained by gel-permeationchromatography, using a TOSOH HLC-8020 Gel PermeationChromatograph (GPC) (Japan) instrument, equipped withtwo GMHXL m-styrogel columns (103–105) and a UV254 nm refractile index detector. Approximately 2 mg mL21
polymer sample was eluted by tetrahydrofuran at a flowrate of 1 mL min21 at 408C and injection volume in allcases was 200 mL. The instrument was calibrated usingpolystyrene standards (TSK Standard Polystyrene, TOSOHJapan) of low polydispersity. The molecular weight range is500–1 090 000.
Data Analysis and Calculations
The calculations for kinetics data were performed usingMATLAB 6.1 software (The MathWorks Inc.).Calculation for volumetric productivity of PHAMCL (F) was
according to Yamane (1992), as shown in equation (4).
F ¼ ½P�=(td þ tf ) (g L�1h�1) (4)
where [P] ¼ PHAMCL concentration (g L21); td ¼ down time (h)and tf ¼ fermentation time (h). In this case where thefermentations were at the bench-scale, td ¼ 0.
PHAMCL yield from the SPKO supplied (YP/C) for the batchfermentation was calculated as in equation (5):
YP=C ¼ D(PHAmaximum � PHAinitial)=SPKOinitial
(g PHA g�1SPKO) (5)
where PHAmaximum ¼ maximum mass of PHA (g) attained;PHAinitial ¼ mass of PHA (g) at the start of cultivation;SPKOinitial ¼ mass of SPKO (g) at the start of cultivation.Residual biomass yield from SPKO supplied (YR/C) for
batch fermentation was calculated as in equation (6):
YR=C ¼ D(Rmaximum � Rinitial)=SPKOinitial
(g residual biomass g�1SPKO) (6)
where residual biomass ¼ total biomass minus PHA mass(g); Rmaximum ¼ maximum mass of residual biomass (g)attained; Rinitial ¼ mass of residual biomass (g) at the startof cultivation. To calculate the residual biomass yield fromammonium (YR/N), the same equation was used exceptthat SPKOinitial term was replaced by initial ammoniummass, NH4initial
þ (g).In the case of fed-batch cultivation, the PHAMCL yield from
the SPKO (YP/C) was calculated as in equation (7):
YP=C ¼ D(PHAmaximum � PHAinitial)=(SPKOinitial
þ SPKOfed)(g PHA g�1SPKO) (7)
where SPKOinitialþ SPKOfed ¼ mass of SPKO (g) at the startof cultivation plus mass of SPKO added during feeding (g).Residual biomass yield from SPKO supplied (YR/C) was
calculated as in equation (8):
YR=C ¼ D(Rmaximum � Rinitial)=(SPKOinitial
þ SPKOfed)(g residual biomass g�1SPKO) (8)
To calculate the residual biomass yield from ammonium(YR/N), the same equation was used except that (SPKOinitial þ
SPKOfed) term was replaced by initial ammonium mass plusmass of ammonium added during feeding i.e., (NH4initial
þþ
NH4fedþ ) (g).
Numerical Calculations
A least-squares method using Marquardt–Levenberg’salgorithm was used for parameter optimization. The calcu-lations were performed using MATLAB 6.1 software (TheMathWorks Inc.).
RESULTS AND ANALYSES
Effect of SPKO Concentrations on the VolumetricOxygen Transfer Coefficient (KLa)
Since SPKO is a highly reduced substrate metabolizedthrough an aerobic b-oxidation pathway, adequate oxygensupply to the cells in the PHAMCL fermentation is importantto maximize yield and productivity. Due to the soap-like prop-erties of the SPKO, it would be reasonable to expect that itspresence in the liquid medium could affect the efficiency ofoxygen transfer to the fermentation system.
Trans IChemE, Part C, Food and Bioproducts Processing, 2007, 85(C2): 104–119
MEDIUM-CHAIN-LENGTH POLY(3-HYDROXYALKANOATES) FROM CRUDE FATTY ACIDS 107

Only experimental results from SPKO concentrations of1.0, 5.0 and 10.0 g L21 were obtained due to the problemof heavy foaming that occurred for the SPKO concentrationsabove 10.0 g L21. The foaming was extremely heavy duringthe KLa determination at SPKO concentration more than10.0 g L21 and hence the whole procedure had to be termi-nated. This was due to the foam leaking through the gasexhaust at the top of the fermenter steel flange. As theexhaust is connected to a water reservoir via siliconetubing, it can be immediately seen that the foaming depositedout significant amount of the carbon source in the reservoir.The estimated KLa values for the three SPKO concentrationsstudied are shown in Table 3.The KLa value for distilled water was the highest at 0.0313
(+3%) s21. The KLa values for SPKO concentrations of1.0 and 5.0 g L21 are very close to each other i.e., 0.025(+4%) s21 and 0.028 (+4%) s21, respectively. However,an increase in SPKO concentration from 5.0 to 10.0 g L21
in the liquid medium caused a reduction in KLa value by50%. It is suggested that further increased in the SPKO con-centration would have caused the KLa to decline even further.It has been reported that surface active agents e.g., sodiumlaurate sulfate at 10 m g L21 can reduce the KLa in the fer-menter by 50% (Aiba et al., 1973). Structurally similar fattyacids are an example of organic compound that could actas surface active agent. In practice, this information isvaluable as the range of SPKO concentrations used in thisstudy for the subsequent bioreactor experiments should bebetween 1.0–10.0 g L21.
Effect of Ammonium Concentration on theSpecific Growth Rate (m)
The effect of ammonium ion concentrations on the specificgrowth rate (m) of the cells was studied using shake-flasks.The calculated m at different ammonium concentrations areshown in Figure 1. It is obvious that m increased with theincrease in ammonium ion concentrations and reaching itsmaximum value around 0.1 g L21 of ammonium ion concen-tration. When the ammonium ion concentrations were furtherincreased, the m started to decline indicating inhibition bysubstrate.Suzuki et al. (1986a) tested six types of ammonium salt
compounds (NH4Cl, NaNH4HPO . 4H2O, NH4H2PO4,(NH4)2HPO4, NH4HCO3 and NH4NO3) for the growth ofPseudomonas sp. K on methanol. They found that all theabove ammonium compounds show similar substrateinhibition effects. In every case, cell growth was optimal at0.2 g NH4
þ L21, and was severely inhibited when NH4þ
exceeded 1.0 g L21. They attributed this effect to the toxiceffect of excess ammonium ion on the microorganism.In this study, the increasingm values were obtained between
0.04–0.10 g L21 ammonium ion concentrations and this is in
agreement with the reported values by Mulchandani et al.(1989), for Wautersia eutropha ATCC17697 (formerly knownas Alcaligenes eutrophus). They observed m was the highestwhen ammonium ion (in sulphate salt form) concentration aslimiting substrate was between 0.08–0.11 g L21. This concen-tration range where m is maximal in this study (0.1 g L21) isslightly lower than reported by Belfares et al. (1995) i.e.,0.4 g L21 for W. eutropha DSM545, when ammonium ion (insulphate salt form) was used as nitrogen source.They also reported that in their shake-flask experiments
using glucose as a carbon source, increasing amount ofthe ammonium ion concentrations caused a linear inhibitionof growth above this threshold concentration (0.4 g L21).The value of m obtained in this study also decreased in analmost similar manner with increasing ammonium ionconcentrations above 0.2 g L21 (Figure 1). Beaulieu et al.(1995) also reported that significantly less biomass of W.eutropha DSM545 was obtained in 3% (w/v) glucose wheninitial ammonium ion concentrations were increased above0.5g L21, indicating that growth was probably impaired athigher ammonium ion concentrations.
Growth Model and its Kinetic ParametersEstimation
It is hypothesized that the growth of the organism could bedescribed using a substrate-inhibition form. The modelequation chosen to describe growth took the form ofAndrew’s equation
m ¼ mmax � S=(Ks þ Sþ S2=Ki) with Ki .. Ks (9)
where mmax is the maximum specific growth rate (h21), Sis the limiting substrate i.e., ammonium (g L21), Ks isthe substrate saturation constant (g L21), and Ki is thesubstrate inhibition constant (g L21). Equation (9) is a
Table 3. Estimated KLa values at different SPKO concentrations.
SPKO concentration (g L21)
Water 1.0 5.0 10.0
KLa(s21) 0.0313
(+3%)0.025(+4%)
0.028(+4%)
0.014(+7%)
Figure 1. Specific growth rate, m (h21) versus ammonium ion concen-trations (g L21) and fitting of Andrews and Monod models to the data.(B experimental data; —— Andrews model; – – – Monod model).
Trans IChemE, Part C, Food and Bioproducts Processing, 2007, 85(C2): 104–119
108 ANNUAR et al.

modification of Monod growth expression which took intoaccount the inhibitory effect of certain substrates towardsmicrobial growth when present at high concentrations.For comparison, the Monod expression would also be fitted
to the experimental data, which is in the form of
m ¼ mmax � S=(Ks þ S) (10)
The equations (9) and (10) were fitted using the data fromshake-flasks experiment. The values of kinetic parametersfor equation (9) (mmax, Ks, Ki) and equation (10) (mmax, Ks)were estimated by fitting each model to the experimentaldata using a nonlinear least squares optimization techniqueby iterative methods that started with an initial guess of theunknown parameters. MATLAB 6.1 software was used forthis purpose which uses the Gauss–Newton algorithm withLevenberg–Marquardt modifications for global convergence.Similar method was used successfully by Mulchandani et al.(1989) to estimate kinetic parameters for growth expression(m) of PHB-producing W. eutropha ATCC 17697, which sub-sequently used in modelling its growth and polymer pro-duction. In modelling the growth and PHB production by W.eutropha DSM545, Belfares et al. (1995) estimated the par-ameters of the specific growth rate expression using batchexperiments with different initial ammonium concentrations,the result also showed substrate inhibition by ammonium.The graphical fitting of each models to the experimentaldata are shown in Figure 1.The estimated parameters value returned by the fitting
function is shown in Table 4 for both models. The experimen-tal data showed a satisfactory fit to the Andrews model withthe residual sum of squares of 0.0003 as compared toMonod’s, where the residual sum of squares was 0.0026.This was also evident from visual inspection of the plots inFigure 1. The estimated parameter values agreed well withthose obtained by other published works (Table 5). Hence,the Andrews model was chosen to describe the organism’sgrowth. For the subsequent fermentation experiments, theeffective ammonium ion concentration incorporated into themedium fell within 0–1.2 g L21.
Effect of Carbon-to-Nitrogen (C/N) Ratio
The C/N ratio experiment served as a basis for the designof an ammonium-limited medium (in which the free fatty acidsare in excess) to be fed during the fed-batch cultivations later.This approach has been successfully used to increase PHAproduction in various organisms and cultivation modesusing different carbon substrates (Suzuki et al., 1986b;Ramsay et al., 1989, 1992; Huijberts and Eggink 1996;
Durner et al., 2001). The optimal ratio was taken as thecarbon and ammonium composition that gave the highestPHA yield (g L21) and/or highest cellular PHAMCL content(gram PHAMCL per gram dry weight of total biomass). Thisoptimum ratio would be used in formulating the feedingmedium, which contains SPKO and ammonium, for the sub-sequent fed-batch experiment. Carbon and nitrogen sub-strates used were SPKO and ammonium, respectively. TheC/N ratio for the experiment is shown in Table 6. The resultsof the experiment are shown in Figure 2(a)–(c). In Figure 2(a)–(c), when cell dry weight (g L21), PHAMCL yield (g L21)and PHAMCL content (% of cells dry weight) were plotted asa function of C/N molar ratio, no significant differenceswere observed between C/N ratio from 3.4 : 1 to 8.6 : 1. It isnoted that within the C/N range tested, the cell dry weight,PHAMCL yield and PHAMCL content were approximately simi-lar in their respective values. C/N molar ratio of 6.9 : 1 wasarbitrarily selected for use in the feeding medium of thesubsequent fed-batch fermentation. At 10.3 : 1 molar ratio,however, poor results were obtained for all variables.Higher C/N molar ratios (or relatively lower amount of
ammonium) seemed to encourage PHAMCL biosynthesispossibly via the effect of nitrogen limitation. An even higherinitial C/N ratio significantly reduced the PHA yield and con-tent once the maximum is reached. The reason for this was
Table 4. Estimated parameters value of Andrews and Monod growthmodels.
Model ParametersParametervalues
95% confidenceintervals for thefitted values
Andrews mmax (h21) 0.2299 0.1920–0.2678
Ks (g L21) 0.0186 0.0089–0.0283Ki (g L21) 1.1938 0.4989–1.8888
Monod mmax (h21) 0.1687 0.1431–0.1943
Ks (g L21) 0.0083 20.0015–0.0181
Table 5. Comparison of Andrews and Monod models kineticparameters with other published works.
ParametersAndrewsform
Monodform
Publishedvalues
Organism/carbonsubstrate/nitrogen
source
mmax (h21) 0.23 0.17 0.29a P. putida
KT2442/octanoicacid/ammoniumsulphate
0.26a P. putidaKT2442/vegetablefatty acids/ammoniumsulphate
Ks (g L21) 0.02 0.01 0.1b W. eutrophaH16/gaseousCO2/ammoniumsulphate
0.41c W. eutrophaATCC17697/fructose/ammoniumsulphate
Ki (g L21) 1.19 — — —
aKellerhals et al. (2000).bHeinzle and Lafferty (1980).cMulchandani et al. (1989).
Table 6. Ratio of carbon (SPKO) to nitrogen (ammonium) for PHAMCL
production by P. putida PGA1.
SPKO-to-ammoniumin mass (g)
SPKO-to-ammoniumin mole
SPKO concentrationsequivalence (g L21)
0.67 : 0.016 3.4 : 1 3.351.01 : 0.016 5.2 : 1 5.051.35 : 0.016 6.9 : 1 6.751.68 : 0.016 8.6 : 1 8.402.02 : 0.016 10.3 : 1 10.1
�SPKO molecular weight: 217 (Cocks and van Rede 1996),ammonium molecular weight: 18.
Trans IChemE, Part C, Food and Bioproducts Processing, 2007, 85(C2): 104–119
MEDIUM-CHAIN-LENGTH POLY(3-HYDROXYALKANOATES) FROM CRUDE FATTY ACIDS 109

possibly due to the poor oxygen transfer to the aqueousmedium and/or toxic effect of free fatty acids mixture fromSPKO at high concentrations. The C/N ratios used in thisexperiment represented the SPKO concentrations rangingfrom 3.35 to 10.1 g L21. It was clear from Figure 2(a)–(c)that the SPKO concentration at 10.0 g L21 did not supportgood growth and PHAMCL production. Thus, effective SPKOconcentration to be included in the fermentation medium iswithin the range of 1.0–8.0 g L21.
Batch Fermentation
The initial amount of SPKO and ammonium provided were6.8 g L21 and 0.4 g L21, respectively. SPKO was the solecarbon and energy source which was in excess for bothgrowth and PHAMCL production, and ammonium was the lim-iting nutrient in the medium. Growth immediately commencedafter inoculation and continued until 12 h where a final0.9 g L21concentration of total biomass was obtained[Figure 3(a)]. The increase in total biomass is due to bothcellular growth and PHAMCL accumulation inside the cells.The pattern of linear growth observed is typical of a
Figure 3. Total biomass (a), ammonium (b), PHAMCL concentration (c) and intracellular content (d) profiles during batch fermentation of P. putidaPGA1.
Figure 2. Effect of different C : N ratio on biomass growth (g L21),PHAMCL concentration (g L21) and content (% biomass dry weight).
Trans IChemE, Part C, Food and Bioproducts Processing, 2007, 85(C2): 104–119
110 ANNUAR et al.

microorganism’s growth under substrate-limitation. Theprofile of ammonium utilization reflected the growth trend[Figure 3(b)]. mmax during the growth phase was approxi-mately 0.10 h21. The increase in PHAMCL concentrationand its intracellular content followed closely the trend ofgrowth, indicating that under the present conditionsPHAMCL accumulation was growth associated [Figure 3(c)and (d)]. Total biomass and PHAMCL concentration reachedtheir maxima at 12 h. Beyond 12 h, a decrease of totalbiomass was observed until the end of fermentation.The time period where a decline in total biomass and active
degradation of stored PHA coincided with the time period ofammonium exhaustion (from approximately 15–48 h) whereits residual concentration in the culture broth was lowerthan 0.05 g L21, which indicated a severe exhaustion andremained so until the end. The residual concentration offree fatty acids in the medium within the cultivation periodremained above 2.5 g L21, which was still in excess. Nooxygen limitation was observed as its partial pressure wasnever less than 50% of air saturation.
Kinetics of PHAMCL Biosynthesis in BatchFermentation
In this batch culture, ammonium-limitation condition wascharacterized by active PHAMCL accumulation whereasammonium-exhaustion condition was represented by activeinternal degradation of stored PHAMCL.The basis of specific PHA production rate, qPHA (g PHAMCL
produced g21 residual biomass h21) calculation was accord-ing to Suzuki et al. (1986b) and Yamane (1992) which is asfollows: when the microorganism produces an intracellularstorage material such as PHAMCL, the total biomass [X](g L21) consists of two main parts, namely PHAMCL [P](g L21) and residual biomass [R] (g L21), where [R] is calcu-lated as the difference between the total biomass and themass of PHAMCL i.e., [R] ¼ [X] 2 [P]. Then the total biomassin the culture is given by equation (11):
½X� � Vbroth ¼ ½P� � Vbroth þ ½R� � Vbroth (11)
where Vbroth is the working volume of the medium. [R] . Vbroth
can be considered as the anabolically active biomass includ-ing proteins and nucleic acids. Therefore the qPHA based onthe unit of residual biomass is described as follows:
qPHA ¼ 1=(½R�t � Vbroth)� d(½P� � Vbroth)=dt
¼ 1=((½X�t � ½P�t) � Vbroth)� d(½P� � Vbroth)=dt (12)
The PHAMCL accumulation data from batch fermenter[Figure 4(c)] was fitted using 4th degree polynomial[Figure 4(a)]. The polynomial form was as follows:
½P�(t) ¼ �3:5(10�8)t4 þ 8:5(10�6)t3
� 0:00056t2 þ 0:012t þ 0:0081 (13)
where [P](t) represents PHAMCL concentration data as a func-tion of time t. The polynomial function was differentiated andsubsequently evaluated with respect to the residual biomassconcentration at a particular time [R]t. This gave the qPHA (gPHAMCL produced g21 R h21) in the batch fermenter.It was clear that residual ammonium exhaustion in the
liquid medium [Figure 4(b)] was not a conducive condition
for effective PHAMCL production, because the PHAMCL bio-synthetic activities, as shown by the rapid decline of theorganism’s specific PHAMCL biosynthesis rate, qPHA[Figure 4(c)], were considerably reduced under this condition.The qPHA calculated at the time when maximum PHAMCL hasbeen accumulated intracellular (11.9% from the total drybiomass) was approximately 0.0062 g PHAMCL producedg21 R h21. The volumetric productivity of PHAMCL wascalculated to be 0.0028 g L21 h21.Suzukiet al. (1986b) calculated the specificPHASCL formation
rate, qPHA (g PHASCL formed g21 residual biomass h21)of Protomonas extorquens to be below 0.01 when theammonium concentration in the liquid medium decreased toalmost 0 ppm. Methanol was continuously supplied as asole carbon and energy source in their cultivations. Similareffect was observed by Bitar and Underhill (1990) in theirbatch culture of W. eutropha. They noted that without theammonium supplementation at the beginning of PHASCL
accumulation phase, the PHASCL accumulation rate (g PHAL21 h21) was below 0.1 g L21 h21. This is nearly 90% lessthan the maximum PHASCL accumulation rate attainable intheir batch culture studies.
Fed-Batch Fermentation
The cultivation was started in a batch mode (1.0 L) beforeswitching to a fed-batch via initiation of feeding. The initialamount of SPKO and ammonium in the fermenter were6.8 g L21 and 0.16 g L21, respectively.In the batch start-up phase, the cellular growth and intra-
cellular PHAMCL accumulation contributed to the increase inthe concentration of total biomass [Figure 5(a)]. The growth
Figure 4. Fitting of PHAMCL concentration time profile with 4th-degreepolynomial (a), the residual ammonium concentration with indicatedlimiting- and exhausted region (b), and the specific PHAMCL
production rate (c) in batch fermentation.
Trans IChemE, Part C, Food and Bioproducts Processing, 2007, 85(C2): 104–119
MEDIUM-CHAIN-LENGTH POLY(3-HYDROXYALKANOATES) FROM CRUDE FATTY ACIDS 111

profile was linear during the batch start-up phase, which ischaracteristic of substrate-limited growth. Final total biomassobtained was 2.1 (+0.1) g L21. The consumption ofammonium followed the growth trend until the commence-ment of feeding [Figure 5(b)]. mmax during the batch start-upgrowth phase was approximately 0.27 h21, which is higherthan in batch fermentation. Higher ammonium ion concen-tration in batch fermentation could have caused growth tobe slightly inhibited.The mass of R remained essentially constant from the start
of feeding (�0.83+ 0.04 g) until the end of the experiment(�0.84+ 0.02 g). This shows that the increase in the totalbiomass was largely due to the increase in the PHAMCL
amount being accumulated intracellular by the cells. Between0–4 h, an increase in PHAMCL concentrations were observed[Figure 5(c)]. However, a slight decline occurred after 4 h untilthe beginning of feeding, where the PHAMCL concentrationsstarted to increase again. A similar trend was observed forthe PHAMCL content of the cells [Figure 5(d)]. The residualammonium concentration in the culture medium during thefeeding period increased temporarily, and approaching0.02 g L21 at 10 h. After that, it decreased to near zero atthe end of fermentation. The decline of PHAMCL concen-tration and its content coincided with the exhaustion ofammonium; with the residual concentration in the culturebroth ,0.05 g L21 [Figure 5(b)]. Similarly, this was observed
in the batch fermentation i.e., when residual ammoniumconcentration ,0.05 g L21 in culture broth, confirming theammonium exhaustion limits (,0.05 g L21). This is regard-less of the initial ammonium concentrations used in thisstudy, which was 0.4 g L21 and 0.16 g L21 for the batchand fed-batch fermentations, respectively. The decline ofPHAMCL concentrations and content is ascribed to theaction of intracellular PHAMCL depolymerase. The free fattyacids concentrations were estimated to be no less than4.0 g L21 throughout the fermentation with final total biomassof 2.1 (+0.1) g L21. Throughout the cultivation, the partialoxygen pressure was never less than 50%.
Kinetics of PHAMCL Biosynthesis in Fed-BatchFermentation
The nitrogen source feeding (0.056 g ammonium h21)together with SPKO was to supply low concentration ofammonium and excess carbon source in the form of freefatty acids during the PHAMCL biosynthesis stage.The calculation for the qPHA was the same as described for
the batch cultivation. In addition, the amounts of culture broth(Vbroth) in a fed-batch culture is time variable. The PHAMCL
accumulation data from fed-batch fermenter was fittedusing 4th degree polynomial [Figure 6(a)]. The polynomial
Figure 5. Total biomass (a), ammonium (b), PHAMCL concentration (c) and intracellular content (d) profiles during fed-batch fermentation ofP. putida PGA1 (dotted line indicated the start of feeding).
Trans IChemE, Part C, Food and Bioproducts Processing, 2007, 85(C2): 104–119
112 ANNUAR et al.

form was as follows:
½P�(t)¼�0:00018t4þ0:0067t3þ0:062t2þ0:22t
þ0:068 (14)
From Figure 6(b), it can be seen that during the batch start-up phase of the cultivation, ammonium concentrations in theculture medium gradually declined to become almost zero.This resulted in a rapid reduction in the specific PHAMCL bio-synthesis rate, qPHA [Figure 6(c)]. However, when the feedingbegins, certain level of residual ammonium was present inthe culture liquid rather than completely exhausted, and theqPHA started to recover and reached the maximum rate ofapproximately 0.6 g PHAMCL produced g21 R h21; with onlya slight decrease until the termination of the experiment at12 h. The volumetric productivity of PHAMCL for the fed-batch fermentation is 0.07 g L21 h21. Jung et al. (2001)obtained similar qPHA i.e., 0.67 g PHAMCL g21 residualbiomass h21 in the chemostat cultivation of Pseudomonasoleovorans using n-octane as carbon source.In fed-batch cultures using fixed-rate ammonia feeding,
60% PHASCL content of the Protomonas extorquens wasobtained faster at 58 h with constant ammonia feeding rateat 0.08 g h21, as compared to nearly 100 h for the samePHASCL content in the case without ammonia feeding(Suzuki et al., 1986b). Improved volumetric PHASCL syn-thesis rate from 0.1 to 0.99 g PHASCL produced L21 h21
was recorded by Bitar and Underhill (1990) during theperiod of ammonium supplementation to the batch cultureof W. eutropha. A similar effect was seen with regards tothe maximal specific rate of monomer production ofPHASCL (0.125 g 3-hydroxyalkanoates monomer producedg21 residual biomass h21), where the rate was maintained
for longer periods in nitrogen fed cultures than in the nitrogenexhausted culture (Aragao et al., 1996).The results obtained here are also comparable to that of pH-
stat fed-batch fermentation using carbon-to-nitrogen (C :N)ratio of 20 g octanoic acid g21 ammonium nitrate (Beom,2002). Using P. oleovorans, highest PHAMCL content of 75%was obtained at this C :N ratio with 0.63 g L21 h21 productivity.In this study, the deterioration in the specific PHAMCL
production rate by P. putida PGA1 was successfully reversedby feeding mineral salt solution which contains fixed ratio ofSPKO-to-ammonium. Furthermore, the feeding enhancesthe biosynthesis of PHA as seen by the concomitant increasein both the PHAMCL concentration and content. The feeding ofresidual ammonium concentrations in the medium has apositive effect on the culture viability and in boosting thePHAMCL accumulation.
Kinetics of SPKO Utilization in Batch andFed-Batch Fermentations
An important aspect to be investigated in this study was theutilization kinetics of free fatty acids mixture, as they supplythe sole carbon and energy substrate for growth andPHAMCL production. The utilization kinetics was studiedduring the growth period of P. putida PGA1 in batch andfed-batch fermentations. From Figure 7, it is shown that theutilization of free fatty acids mixture from SPKO followed azero-order type of reaction kinetics for both batch and fed-batch fermentations. Hence, an increase in the effective con-centration of SPKO in the medium may not have much effectin encouraging better uptake of SPKO by the cells. This alsoindicated that the utilization of the free fatty acids from theaqueous medium into the cells was subjected to mass
Figure 6. Fitting of PHAMCL concentration time profile with 4th-degreepolynomial (a), the residual ammonium concentration profile (b), andthe specific PHAMCL production rate (c) in fed-batch fermentation.
Figure 7. Zero-order uptake kinetics of free fatty acids from SPKO(O: fed-batch; A: batch; S.E.: standard error).
Trans IChemE, Part C, Food and Bioproducts Processing, 2007, 85(C2): 104–119
MEDIUM-CHAIN-LENGTH POLY(3-HYDROXYALKANOATES) FROM CRUDE FATTY ACIDS 113

transfer limitation. In practical sense, it is better in future thatthe SPKO be supplied to the fermentation via controlled feed-ing as this can avoid the toxic effect of the free fatty acids mix-ture to growth. It will also reduce the risk of heavy foamingand deterioration in KLa values. Furthermore the high batchwise supply of SPKO does not seem to improve its rate ofutilization by the cells. It appears that the fed-batch culturehad a better utilization rate of free fatty acids as comparedto the batch culture i.e., SPKO utilization rates of 20.348(+0.019) h21 and 20.067 (+0.005) h21, respectively. Theamount of residual biomass present during this uptakeperiod however was approximately 0.1–0.8 g L21 for bothfermentations. Hence, the difference in the utilization rate offree fatty acids between batch and fed-batch modes couldbe related to the higher specific growth rate in fed-batch(0.27 h21) as compared to batch (0.10 h21).
Comparison Between Batch and Fed-BatchCultivations
The performance between the two cultivation modes wasevaluated in terms of their volumetric and specific productiv-ities, yield, biomass concentration, product concentration andPHAMCL content (Table 7). It is clear that the fed-batch culti-vation mode is superior to batch cultivation. In terms of volu-metric productivity,F, the fed-batch is at least 25 timeshigher than the batch. The calculated productivity of0.07 g L21 h21 and 71.4 (+2.0)% PHAMCL content usingthe fed-batch mode in this study is higher than reported forthe PHA production using an industrial scale fermenter of20 000 L (Chen et al., 2001), where Aeromonas hydrophila4AK4 was used to produce PHA from lauric acid underammonium-limited conditions with the overall PHA pro-ductivity of 0.04 g L21 h21 and PHA content of 22% fromthe total dry biomass.
Better PHAMCL yield from SPKO (YP/C) was also obtainedusing the fed-batch compared to batch cultivation (Table 7).Lower residual biomass yield on ammonium (YR/N) as thegrowth limiting substrate in the batch cultivation indicatedthat the organism’s growth was slightly affected by highammonium concentration, as compared to fed-batch withlower ammonium concentration. Improved qPHA, final totalbiomass and product concentrations were obtained usingfed-batch mode (Table 7). This was evident from the finalPHAMCL content obtained i.e., 71.4 (+2.0)% from total drybiomass weight. This represents the highest reported valuefor this particular strain using SPKO as the sole carbon andenergy source. Furthermore, this high PHAMCL content wasachieved within 12 h in the fed-batch cultivation, which isrelatively a short period of time. These data supported theearlier suggestion that slight ammonium feeding during theactive PHAMCL biosynthesis stage was preferable for main-taining the anabolic activity to accumulate intracellularPHAMCL under ammonium-limited condition.Further comparison with other published data is shown in
Table 8. From Table 8, it can be seen that free fatty acids mix-ture from SPKO can be an excellent substrate for PHAMCL pro-duction in the fermenter in place of the generally moreexpensive pure fatty acids. The YP/C from free fatty acids mix-ture compared quite well with the yields from pure acids. In thecase of YR/C, most of the carbon from the pure acids seemedto be directed for growth rather than for PHAMCL biosynthesiswhich was indicated by higherYR/C values. In terms of specificPHAMCL productivity, the use of free fatty acidsmixture yields asuperior rate relative to the pure fatty acids fermentations. Thisis important if the volumetric PHAMCL productivity of thefermentation and cellular PHAMCL content were to beimproved, which is evident from the higher P/X value obtainedin the cultivation using free fatty acids mixture (Table 8).Bitar and Underhill (1990) reported higher PHASCL content
approximately 70% was achieved by ammonium
Table 7. Comparison between the batch and fed-batch fermentation modes.
Mode X (g L21) P (g L21) P/X (%) mmax (h21) YR/N (g g21) YP/C (g g21) F (g L21h21) qPHA
a (g P g21 R h21)
Batch 0.90+ 0.01 0.09+ 0.0 11.9+ 0.0 0.10 2.0 0.02 0.0028 0.0062Fed-batch 2.1 + 0.1 1.49+ 0.04 71.4+ 2.0 0.27 2.6 0.11 0.07 0.6
Keys: maximum total biomass concentration, X; maximum PHA concentration, P; maximum PHA content, P/X; maximum specific growth rate,mmax; residual biomass yield on ammonium, YR/N; PHAMCL yield on SPKO, YP/C; volumetric productivity, F; specific PHA production rate, qPHA.aThe maximum values calculated during the active PHA accumulation phase under ammonium-limitation.
Table 8. Comparison of culture parameters obtained from this study to the fermentation which employed pure acids as carbon substrates.
Carbon substrateParameter SPKO� Citric acid Hexanoic acid Heptanoic acid Octanoic acid Nonanoic acid
YP/C 0.11 — 0.004 0.12 0.23 0.19YR/C 0.04 0.73 0.95 0.75 0.80 0.88qPHA 0.6 — 0.001 0.074 0.147 0.091P/X 71.4 0 0.6 28.0 42.0 31.0
N.B. Data for the comparison was taken from P. oleovorans cultivation on different carbon sources. Parameter values are calculated fromammonium-limited condition in the chemostat with a constant dilution rate of 0.2 h21 (Durner et al., 2001).�data taken from fed-batch cultivation in this study.Symbols: YP/C (g g21): PHA yield coefficient on carbon source.YR/C (g g21): carbon growth yield coefficient.qPHA (g P g21 residual biomass h21): specific PHA production rate.P/X (%): PHA content (% total dry biomass weight).
Trans IChemE, Part C, Food and Bioproducts Processing, 2007, 85(C2): 104–119
114 ANNUAR et al.

supplementation to the batch culture of W. eutropha as com-pared to 59% without ammonium supply. In addition, PHASCL
yield from glucose was 0.2 g PHASCL g21 glucose in normalbatch culture compared with 0.447 g PHASCL g21 glucose inthe ammonium supplemented culture.It is clear that feeding with a small quantity of ammonium
during the PHAMCL accumulation phase improved the qPHAof the P. putida PGA1 under ammonium-limited condition,resulting in higher PHAMCL concentration and content. Onthe other hand, cultivation without ammonium feeding yieldedsignificantly reduced qPHA value with low PHAMCL concen-tration and content.
Monomer Compositions and MolecularWeight of PHAMCL
Monomer compositions of the PHAMCL produced duringbatch and fed-batch fermentations were also studied. Theconstituent monomers of PHAMCL synthesized by P. putidaPGA1 when SPKO is the sole carbon and energysource are 3-hydroxyoctanoate (C8-), 3-hydroxydecanoate(C10-), 3-hydroxydodecanoate (C12-), 3-hydroxytetradecano-ate (C14-) and 3-hydroxyhexadecanoate (C16-). In batchfermentation, the relative molar fractions of these monomersare almost constant up to 12 h of cultivation time at whichpoint PHAMCL accumulation is maximum [Figure 8(a)].Beyond 12 h, strong degradation of stored PHAMCL by thecells and total biomass decline can be observed.In fed-batch fermentation, there was a gradual increase in
the C8-, C10- and C12-molar fractions of the PHAMCL in thefirst 4 h of cultivation. At the same time, the C14- and C16-molar fractions started to decrease in relative proportion tothe increase in the C8-, C10- and C12-molar fractions[Figure 8(b)]. After 4 h, all the molar fractions of the constitu-ent monomers remained relatively constant until the end offermentation.In both cultivations, C8-fraction was the pre-dominant
monomer with molar fraction as high as 55 mole%; theother monomer constituents were present at varying relativeamount i.e., C10 15–30 mole%; C12 5–10 mole%; C14 5–20mole% and C16 15–30 mole%.The molecular weight (Mw), molecular number (Mn) and
polydispersity (Mw/Mn) of the PHAMCL produced in batchand fed-batch fermentations are shown in Table 9. ThePHAMCL obtained from both cultivations have high molecularweight and similar polydispersity. These values are similar tothe PHAMCL molecular masses reported by Tan et al. (1997)in their shake-flasks study when SPKO was used as carbonsubstrate for PHAMCL production by P. putida PGA1 (i.e.,124 000 Mw, 58 800 Mn and 2.1 Mw/Mn).
Simulation of Growth and PHAMCL Productionin a Two-Stage Chemostat System
The possibility of using a two-stage chemostat system forthe process is also examined. The growth and product for-mation steps need to be separated since optimal conditionsfor each step are different, hence justify the use of a two-stage system. Optimal growth is favoured under balancedcultivation conditions. For product formation as in the caseof PHAMCL fermentation, the conditions are essentially imbal-anced. The singular difference here is the optimal C/N ratiofor PHAMCL production as was shown in the fed-batch
fermentation. It is proposed that the first-stage chemostat isused for primarily residual biomass (R) growth; with thecarbon, nitrogen and other minerals supplied in stoichio-metric proportion for optimal growth. The second-stagechemostat is used for the PHAMCL production, (P) with theoptimal C/N ratio medium composition as the main charac-teristics. Growth in this stage involved intrinsically unba-lanced growth, even though it is steady-state growth. Newcells entering this stage are continuously adapting to thenew conditions.The fermentation profiles obtained from the fed-batch
fermentation were used for the graphical approach on thetwo-stage chemostat design, as proposed by Shuler andKargi (1992). However, the transfer of the information frombatch growth curve to predictions of the two-stage systemstill requires the assumption of balance growth. The growthdata as a function of time, t was fitted with the polynomialform as shown in equation (15):
R(t) ¼ �0:0019t3 þ 0:027t2 � 0:0097t þ 0:096 (15)
The PHAMCL production data was fitted with the polynomialform as follows:
P(t) ¼ �0:012t3 þ 0:37t2 � 3:4t þ 10 (16)
This approach required plots of dR/dt versus Rt and dP/dtversus Pt which was accomplished by using MATLAB 6.1software. Rt and Pt each represents residual biomass con-centration and PHAMCL concentration at time t, respectively.The intersection of the reaction curve with a line from massbalance equation of the form
rR,n(Rn, Sn) ¼ Dn(Rn � Rn�1) (17)
where n represents nth stage; r is the reaction rate; S is thesubstrate; and D is the dilution rate; determined the exitconcentration of R or P.With the mean residence time, u (¼1/D) simulated at
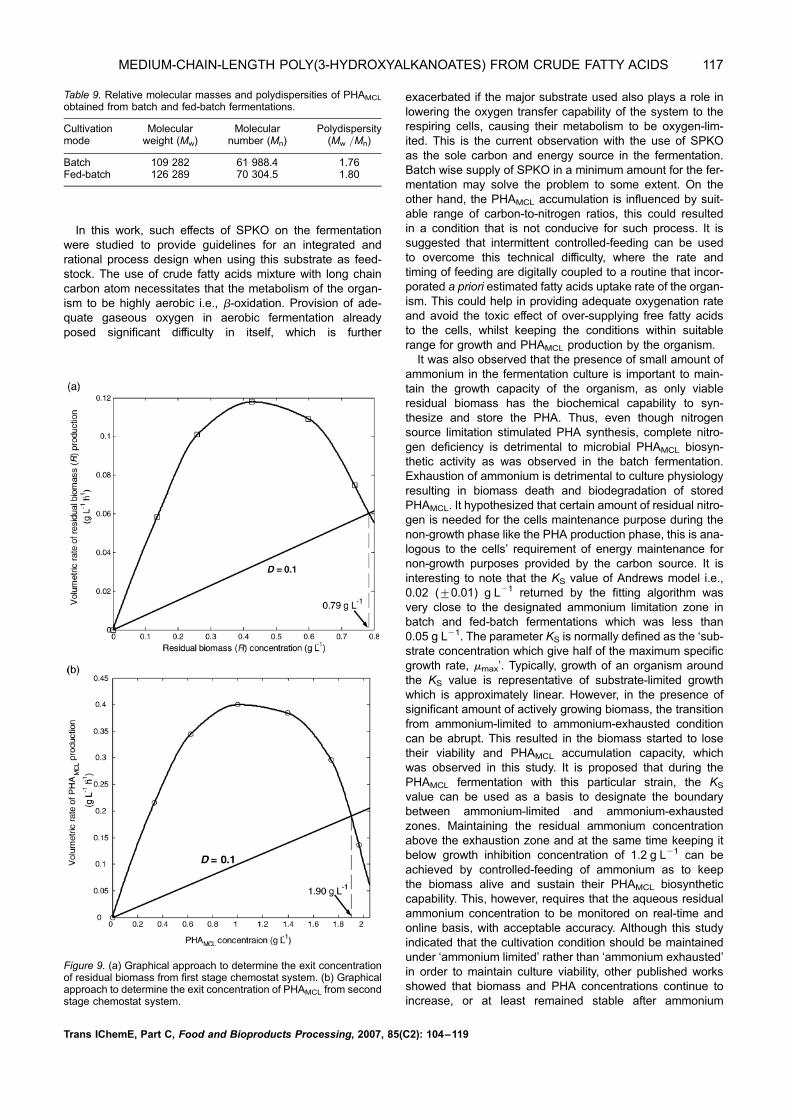
10 h, it was estimated that the exit concentration of R fromthe first-stage chemostat was approximately 0.79 g L21
[Figure 9(a)]. The value of D ¼ 0.1 h21 was chosen to rep-resent the lower m (which is important for PHA accumulation)than the mmax ¼ 0.23 h21 estimated in this study. The exitconcentration of P from the second-stage chemostat wasapproximately 1.90 g L21 [Figure 9(b)]. This gives PHAproductivity of approximately 0.2 g L21 h21. Assuming thatthe working volume of the fermenter was 700-L, thiscorresponds to 1.4 kg mass of PHA. It is immediately obviousthat the use of two-stage chemostat system theoreticallydoes not guarantee a significantly higher PHAMCL productionthan the fed-batch system for the process to be economicallyviable. The better choice in terms of cost would be the use ofthe fed-batch fermentation system.
DISCUSSION AND CONCLUSIONS
Crude fatty acids mixture has a high potential to be used asfeedstock in a large-scale fermentation processes. Therefore,it is imperative to elucidate aspects related to the use of thesesubstrates in fermentation. These include physical and bio-chemical related effects particularly those that have importantimplications for process improvement and scale-up.
Trans IChemE, Part C, Food and Bioproducts Processing, 2007, 85(C2): 104–119
MEDIUM-CHAIN-LENGTH POLY(3-HYDROXYALKANOATES) FROM CRUDE FATTY ACIDS 115

Figure 8. (a) Relative molar fractions of 3-hydroxyalkanoate monomers in PHAMCL during batch cultivation (data for each monomer constituentwas an average of three to four replicates with standard deviation of 4% to 10%). (b) Relative molar fractions of 3-hydroxyalkanoate monomersin PHAMCL during fed-batch cultivation (data for each monomer constituent was an average of three to four replicates with standard deviation of6% to 12%).
Trans IChemE, Part C, Food and Bioproducts Processing, 2007, 85(C2): 104–119
116 ANNUAR et al.
In this work, such effects of SPKO on the fermentationwere studied to provide guidelines for an integrated andrational process design when using this substrate as feed-stock. The use of crude fatty acids mixture with long chaincarbon atom necessitates that the metabolism of the organ-ism to be highly aerobic i.e., b-oxidation. Provision of ade-quate gaseous oxygen in aerobic fermentation alreadyposed significant difficulty in itself, which is further
exacerbated if the major substrate used also plays a role inlowering the oxygen transfer capability of the system to therespiring cells, causing their metabolism to be oxygen-lim-ited. This is the current observation with the use of SPKOas the sole carbon and energy source in the fermentation.Batch wise supply of SPKO in a minimum amount for the fer-mentation may solve the problem to some extent. On theother hand, the PHAMCL accumulation is influenced by suit-able range of carbon-to-nitrogen ratios, this could resultedin a condition that is not conducive for such process. It issuggested that intermittent controlled-feeding can be usedto overcome this technical difficulty, where the rate andtiming of feeding are digitally coupled to a routine that incor-porated a priori estimated fatty acids uptake rate of the organ-ism. This could help in providing adequate oxygenation rateand avoid the toxic effect of over-supplying free fatty acidsto the cells, whilst keeping the conditions within suitablerange for growth and PHAMCL production by the organism.It was also observed that the presence of small amount of
ammonium in the fermentation culture is important to main-tain the growth capacity of the organism, as only viableresidual biomass has the biochemical capability to syn-thesize and store the PHA. Thus, even though nitrogensource limitation stimulated PHA synthesis, complete nitro-gen deficiency is detrimental to microbial PHAMCL biosyn-thetic activity as was observed in the batch fermentation.Exhaustion of ammonium is detrimental to culture physiologyresulting in biomass death and biodegradation of storedPHAMCL. It hypothesized that certain amount of residual nitro-gen is needed for the cells maintenance purpose during thenon-growth phase like the PHA production phase, this is ana-logous to the cells’ requirement of energy maintenance fornon-growth purposes provided by the carbon source. It isinteresting to note that the KS value of Andrews model i.e.,0.02 (+0.01) g L21 returned by the fitting algorithm wasvery close to the designated ammonium limitation zone inbatch and fed-batch fermentations which was less than0.05 g L21. The parameter KS is normally defined as the ‘sub-strate concentration which give half of the maximum specificgrowth rate, mmax’. Typically, growth of an organism aroundthe KS value is representative of substrate-limited growthwhich is approximately linear. However, in the presence ofsignificant amount of actively growing biomass, the transitionfrom ammonium-limited to ammonium-exhausted conditioncan be abrupt. This resulted in the biomass started to losetheir viability and PHAMCL accumulation capacity, whichwas observed in this study. It is proposed that during thePHAMCL fermentation with this particular strain, the KS
value can be used as a basis to designate the boundarybetween ammonium-limited and ammonium-exhaustedzones. Maintaining the residual ammonium concentrationabove the exhaustion zone and at the same time keeping itbelow growth inhibition concentration of 1.2 g L21 can beachieved by controlled-feeding of ammonium as to keepthe biomass alive and sustain their PHAMCL biosyntheticcapability. This, however, requires that the aqueous residualammonium concentration to be monitored on real-time andonline basis, with acceptable accuracy. Although this studyindicated that the cultivation condition should be maintainedunder ‘ammonium limited’ rather than ‘ammonium exhausted’in order to maintain culture viability, other published worksshowed that biomass and PHA concentrations continue toincrease, or at least remained stable after ammonium
Figure 9. (a) Graphical approach to determine the exit concentrationof residual biomass from first stage chemostat system. (b) Graphicalapproach to determine the exit concentration of PHAMCL from secondstage chemostat system.
Table 9. Relative molecular masses and polydispersities of PHAMCL
obtained from batch and fed-batch fermentations.
Cultivationmode
Molecularweight (Mw)
Molecularnumber (Mn)
Polydispersity(Mw /Mn)
Batch 109 282 61 988.4 1.76Fed-batch 126 289 70 304.5 1.80
Trans IChemE, Part C, Food and Bioproducts Processing, 2007, 85(C2): 104–119
MEDIUM-CHAIN-LENGTH POLY(3-HYDROXYALKANOATES) FROM CRUDE FATTY ACIDS 117

concentration dropped to extremely low values in batchfermentation (Sonnleitner et al., 1979; Heinzle and Lafferty,1980; Mulchandani et al., 1989; Belfares et al., 1995;Durner et al., 2001). Bacterial strains used in these studieswere W. eutropha and P. oleovorans. Thus, it can be seenthat the effect of ammonium exhaustion on PHA biosynthesismay be different from one strain to another.Kim et al. (1997) suggested that there would exist some
optimal concentrations of ammonium and dissolved oxygento maintain cell viability high enough to accumulatePHAMCL since its concentration depends on both the cell con-centration and its cellular content; therefore, a minimal supplyof these are necessary to achieve a high PHA productivity.This was based on their results from the two-step fed-batchfermentation of P. putida BM01 using combined glucoseand octanoate as carbon sources, where ammonium wasused as the limiting nutrient and supplied in a fixed ratio tooctanoate in feeding solution during the PHAMCL accumu-lation phase.The mode of cultivation has little influence on the proper-
ties of PHAMCL produced by P. putida PGA1 from SPKO.The polymers obtained from the different fermentationmodes exhibited high molecular masses and relatively similarmonomer compositions with 3-hydroxyoctanoate (C8) as thepre-dominant monomer.It is concluded that PHAMCL production can be improved by
feeding residual ammonium concentrations in the cultureliquid. This is supported by the comparison of the PHAMCL
production kinetics from the batch and fed-batch fermenta-tions as shown in this study.
NOMENCLATUREF volumetric productivity of PHAMCL, g L21 h21
u mean residence time, hm specific growth rate, h21
mmax maximum specific growth rate, h21
D dilution rate, h21
KS Monod constant, g L21
Ki substrate inhibition constant, g L21
NH4initialþ mass of ammonium at the start of cultivation, g
NH4fedþ mass of ammonium added during feeding, g
PHAinitial mass of PHAMCL at the start of cultivation, gPHAmaximum maximum mass of PHAMCL, gqPHA specific PHAMCL production rate, g PHAMCL g21
residual biomass h21
R residual biomass, gRinitial mass of residual biomass at the start of
cultivation, gRmaximum maximum mass of residual biomass attained, gs secondsS growth limiting substrateSPKOinitial mass of SPKO at the start of cultivation, gSPKOfed mass of SPKO added during feeding, gtd fermenter downtime, htf fermentation time, hVbroth working volume of the fermenter, LYP/C PHAMCL yield from SPKO, g PHAMCL g21 SPKOYR/C residual biomass yield from SPKO, g R g21 SPKOYR/N residual biomass yield from ammonium, g R g21
ammonium[P] PHAMCL concentration, g L21
[R] residual biomass concentration, g L21
[X] total biomass concentration, g L21
Subscriptst time, hn nth stage
REFERENCES
Aiba, S., Humphrey, A.E. and Millis, N.F., 1973, Biochemical Engin-eering, 2nd edition, 184–185 (University of Tokyo Press).
Aragao, G.M.F., Lindley, N.D., Uribelarrea, J.L. and Pareilleux, A.,1996, Maintaining a controlled residual growth capacity increasesthe production of polyhydroxyalkanoate copolymers by Alcaligeneseutrophus, Biotech Lett, 18: 937–942.
Beaulieu, M., Beaulieu, Y., Melinard, J., Pandian, S. and Goulet, J.,1995, Influence of ammonium salts and cane molasses ongrowth of Alcaligenes eutrophus and production of polyhydroxy-butyrate, Appl Environ Microbiol, 61: 165–169.
Belfares, L., Perrier, M., Ramsay, B.A., Ramsay, J.A., Jolicoeur, M.and Chavarie, C., 1995, Multi-inhibition kinetic model for thegrowth of Alcaligenes eutrophus, Can J Microbiol, 41: 249–256.
Beom, S.K., 2002, Production of medium chain length polyhydrox-yalkanoates by fed-batch culture of Pseudomonas oleovorans,Biotech Lett, 24: 125–130.
Bitar, A. and Underhill, S., 1990, Effect of ammonium supplemen-tation on production of poly-b-hydroxybutyric acid by Alcaligeneseutrophus in batch culture, Biotech Lett, 12: 563–568.
Brandl, H., Gross, R.A., Lenz, R.W. and Fuller, R.C., 1988, Pseudo-monas oleovorans as a source of poly(b-hydroxyalkanoates) forpotential applications as biodegradable polyesters, Appl EnvironMicrobiol, 54: 1977–1982.
Chen, G.Q., Zhang, G., Park, S.J. and Lee, S.Y., 2001, Industrialscale production of poly(3-hydroxybutyrate-co-3-hydroxyhexano-ate), Appl Microbiol Biotechnol, 57: 50–55.
Cocks, L.V. and van Rede, C., 1966, Laboratory Handbook for Oiland Fat Analysts, 113–117 (Academic Press, London, UK).
Durner, R., Zinn, M., Witholt, B. and Egli, T., 2001, Accumulation ofpoly[(R)-3-hydroxyalkanoates] in Pseudomonas oleovoransduring growth in batch and chemostat culture with differentcarbon sources, Biotechnol Bioengng, 72: 278–288.
Elson, C.E., 1992, Tropical oils: nutritional and scientific issues, CritRev Food Sci Nutr, 31(1/2): 79–102.
Heinzle, E. and Lafferty, R.M., 1980, A kinetic model for growth andsynthesis of poly-b-hydroxybutyric acid (PHB) in Alcaligenes eutro-phus H16, Eur J Appl Microbiol Biotechnol, 11: 8–16.
Hocking, P.J. and Marchessault, R.H., 1994, Biopolyesters, in Tech-nology of Biodegradable Polymers, Chemistry and Griffin, G.S.L.(ed.). 48–96 (Blackie Academic and Professional, Chapman &Hall, London, UK).
Hori, K., Soga, K. and Doi, Y., 1994, Effects of culture conditions onmolecular weights of poly(3-hydroxyalkanoates) produced byPseudomonas putida from octanoate, Biotech Lett, 16: 709–714.
Huijberts, G.N.M. and Eggink, G., 1996, Production ofpoly(3-hydroxyalkanoates) by Pseudomonas putida KT2442 incontinuous cultures, Appl Microbiol Biotechnol, 46: 233–239.
Jung, K., Hazenberg, W., Prieto, M. and Witholt, B., 2001, Two-stagecontinuous process development for the production of medium-chain-length poly(3-hydroxyalkanoates), Biotechnol Bioengng,72: 19–24.
Kellerhals, M.B., Kessler, B., Witholt, B., Tchouboukov, A. andBrandl, H., 2000, Renewable long-chain fatty acids for theproduction of biodegradable medium-chain-length polyhydroxyalk-anoates (mcl-PHAs) at laboratory and pilot plant scales,Macromol-ecules, 3: 4690–4698.
Kim, G.J., Lee, I.Y., Yoon, S.C., Shin, Y.C. and Prak, Y.H., 1997,Enhanced yield and high production of medium-chain-lengthpoly(3-hydroxyalkanoates) in a two-step fed-batch cultivation ofPseudomonas putida by combined use of glucose and octanoate,Enzyme Microbial Technol, 20: 500–505.
Lageveen, R.G., Huisman, G.W., Preusting, H., Ketelaar, P.,Eggink, G. and Witholt, B., 1988, Formation of polyesters by Pseu-domonas oleovorans: effect of substrates on formation andcomposition of poly-(R)-3-hydroxyalkanoates and poly-(R)-3-hydroxyalkenoates, Appl Environ Microbiol, 54: 2924–2932.
Lee, S.Y., Wong, H.H., Choi, J.I., Lee, S.H., Lee, S.C. and Han, C.S.,2000, Production of medium-chain-length polyhydroxyalkanoatesby high-cell-density cultivation of Pseudomonas putida underphosphorus limitation, Biotechnol Bioengng, 68: 466–470.
Montes, F.J., Catalan, J. and Galan, M.A., 1999, Prediction of KLa inyeast broths, Process Biochem, 34: 549–555.
Mulchandani, A., Luong, J.H.T. andGroom,C., 1989, Substrate inhibitionkinetics for microbial growth and synthesis of poly-b-hydroxybutyric
Trans IChemE, Part C, Food and Bioproducts Processing, 2007, 85(C2): 104–119
118 ANNUAR et al.

acid by Alcaligenes eutrophus ATCC 17697, Appl MicrobiolBiotechnol, 30: 11–17.
Ramsay, B.A., Ramsay, J.A. and Cooper, D.G., 1989, Production ofpoly-b-hydroxyalkanoic acid by Pseudomonas cepacia, ApplEnviron Microbiol, 55: 584–589.
Ramsay, B.A., Saracovan, I., Ramsay, J.A. and Marchessault, R.H.,1992, Effect of nitrogen limitation on long-side-chain poly-b-hydro-xyalkanoate synthesis by Pseudomonas resinovorans, ApplEnviron Microbiol, 58: 744–746.
Shuler, M.L. and Kargi, F., 1992, Bioprocess Engineering: BasicConcepts, 199–209 (Prentice-Hall International, Inc., NewJersey, USA).
Solorzano, L., 1969, Determination of ammonia in natural waters bythe phenolhypochlorite method, Limnol Oceanogr, 14: 799–801.
Sonnleitner, B., Heinzle, E., Braunegg, G. and Lafferty, R.M., 1979,Formal kinetics of poly-b-hydroxybutyric acid (PHB) production inAlcaligenes eutrophus H16 and Mycoplana rubra R14 with respectto the dissolved oxygen tension in ammonium-limited batchculture, Eur J Appl Microbiol Biotechnol, 7: 1–10.
Suzuki, T., Yamane, T. and Shimizu, S., 1986a, Mass production ofpoly-b-hydroxybutyric acid by fully automatic fed-batch cultureof methylotroph, Appl Microbiol Biotechnol, 23: 322–329.
Suzuki, T., Yamane, T. and Shimizu, S., 1986b, Kinetics and effect ofnitrogen source feeding on production of poly-b-hydroxybutyricacid by fed-batch culture, Appl Microbiol Biotechnol, 24: 366–369.
Tan, I.K.P., Sudesh Kumar, K., Theanmalar, M., Gan, S.N. andGordon III, B., 1997, Saponified palm kernel oil and its majorfree fatty acids as carbon substrates for the production of polyhy-droxyalkanoates in Pseudomonas putida PGA1, Appl MicrobiolBiotechnol, 47: 207–211.
Yamane, T., 1992, Cultivation engineering of microbial bioplasticsproduction, FEMS Microbiol Rev, 103: 257–264.
ACKNOWLEDGEMENTThis research was funded by the University of Malaya (grant no.
Vot F0156/2001A).
The manuscript was received 4 April 2006 and accepted forpublication after revision 4 January 2007. The paper was publishedonline ahead of print 18 May 2007.
Trans IChemE, Part C, Food and Bioproducts Processing, 2007, 85(C2): 104–119
MEDIUM-CHAIN-LENGTH POLY(3-HYDROXYALKANOATES) FROM CRUDE FATTY ACIDS 119






![Linear position sensors with Ethernet/IP interface FAST ...€¦ · M18×1.5/O-ring, Rod diameter 10.2 mm Z Thread 3/4"–16 UNF/O-ring, Rod diameter 10.2 mm Nominal length [4-digit]](https://img.pdfslide.us/doc/110x75/605c0d56b832773227584c0d/linear-position-sensors-with-ethernetip-interface-fast-m1815o-ring-rod.jpg)