Embed Size (px)
Citation preview

389
Production of Haploid and Doubled Haploid Plants from Anther-derived Callus of Lilium formosanum D.-S. Han and Y. Niimi Faculty of Agriculture, Niigata University 2-8050 Ikarashi, Niigata 950-2181 Japan Keywords: anther culture, haploid callus, lily, plant regeneration Abstract
Anther cultures of Lilium formosanum were made to obtain haploid and doubled haploid plants. Anthers containing microspores at the uninucleate stage were cultured on MS medium supplemented with picloram and zeatin. Five haploid callus lines were obtained and maintained long term by subculturing them at 2- to 3-month intervals on MS medium containing picloram. Haploid plants (2n = x = 12) were regenerated from the anther-derived callus lines and grown to flowering. These haploid plants showed poor growth and abnormal meiosis resulting in formation of sterile pollen. Doubled haploid plants were produced from a colchicine-treated haploid callus line, but pollen of the doubled haploid plants also was sterile. The ploidy level of the callus lines was determined by flow cytometry. Five anther-derived callus lines which had been maintained over 4 years showed a difference in ploidy: 2 lines remained at a haploid level, 2 lines had became diploid and one line was a mixoploid population consisting of haploid, diploid and triploid cells. INTRODUCTION
Lilies (Lilium spp.) are commercially important flower bulbs, and consist of about 130 species and numerous cultivars with different flower color and sizes. Since lilies are genetically heterozygous and usually propagated vegetatively, development of a breeding program using haploids will be useful for fixation of horticulturally important traits and production of novel cultivars. Haploid or doubled haploid (DH) plants have been produced via anther culture, but were limited to several Lilium species and cultivars containing L. longiflorum (Sharp et al., 1971; Arzate-Fernández et al., 1997), L. davidii (Gu and Cheng, 1982), L. x ‘Connecticut King’ (Han et al., 1997, 1999) and L. x ‘Pollyanna’ (Chu et al., 2001). The present paper describes haploid and doubled haploid production from anther-derived calluses of Lilium formosanum which takes only one to two years to flower from seed. MATERIALS AND METHODS Plant Material and Anther Culture
Lilium formosanum (2n = 2x = 24) was used in the present study. Flower buds (40 to 50 mm long) containing microspores at the uninucleate stage were harvested from plants grown under natural conditions in the field. The buds were stored at 4°C for 2 days, and then surface-disinfected with 70% ethanol for 1 min and with commercial bleach (sodium hypochlorite) containing 2% active chlorine for 20 min. The disinfected buds were rinsed three times with sterilized distilled water. Anthers were isolated from the buds and placed on MS medium (Murashige and Skoog, 1962) supplemented with 2 mg l-1 picloram, 2 mg l-1 zeatin and 6% (w/v) sucrose (Han et al., 1997). The culture medium was adjusted to pH 5.8 with 0.1 N NaOH before addition of 0.25% (w/v) gellan gum. Forty ml of dissolved medium was poured into each of the 100 ml Erlenmeyer flasks. The medium was autoclaved for 15 min at 121°C under a pressure of 1.2 kg cm-2. Twelve anthers were placed in each flask. Cultures were incubated in the dark at 25°C.
Proc. IXth Intl. Symp. on Flower Bulbs Eds.: H. Okubo, W.B. Miller and G.A. Chastagner Acta Hort. 673, ISHS 2005

390
Maintenance of Anther-derived Calluses and Plant Regeneration Anther-derived calluses were transferred to 100 ml Erlenmeyer flasks containing
40 ml MS callus-proliferating medium containing 1 mg l-1 picloram, 3% (w/v) sucrose, and 0.25% (w/v) gellan gum. Cultures were maintained in the dark at 25°C and subcultured at 2- to 3-month intervals. To induce plant regeneration, calluses were transferred to 100 ml Erlenmeyer flasks containing 40 ml MS regenerating medium supplemented with 0.1 mg l-1 NAA, 0.01 mg l-1 BA, 3% (w/v) sucrose, and 0.25% (w/v) gellan gum, and then incubated in the dark at 25°C. Regenerated plantlets were transplanted in the greenhouse into pots (5 x 5 x 5 cm) containing vermiculite after cold treatment at 4°C for 10 weeks. They were then transplanted into a 24-cm clay pot containing mixed soil and grown in a plastic greenhouse. Ploidy Determinations
Ploidy levels of anther-derived calluses and plants were determined by flow cytometric analysis as previously described (Mishiba et al., 2000). Calluses and leaves of regenerated plantlets were used for the measurement of relative DNA contents by flow cytometer PA (Partec GmbH, Münster, Germany) for comparison with the DNA content in leaves of the mother plants. To isolate nuclei, samples were chopped with a razor blade in 0.5 ml of Solution A (High Resolution DNA kit, Type A; Partec GmbH). Nuclei were then stained for 10 min with 2 ml of Solution B (High Resolution DNA kit, Type P; Partec GmbH) containing 4,6-diamidino-2-phenylindole (DAPI) prior to flow cytometric analysis.
A squash technique was used for chromosome counts. Root-tips excised from anther-derived plantlets grown in vitro were fixed in an acetic acid-ethanol solution (absolute ethanol : glacial acetic acid (3:1, v/v)) at room temperature for at least 24 h, and hydrolyzed in 1 N HCl at 60°C for 10 min. Samples were then rinsed with distilled water, stained with a 2% aceto-orcein solution, and examined under a light microscope. Colchicine Treatment of Haploid Calluses
To obtain doubled haploid plants, haploid calluses were transferred into a liquid callus-proliferating MS medium supplemented with 0.5 mM colchicine, incubated on a rotary shaker at 100 rpm in the dark at 25°C for 72 h, and then rinsed three times with sterilized distilled water (Han et al., 1999). Colchicine-treated calluses were transferred to the MS regenerating medium described above. RESULTS AND DISCUSSION Callus Formation and Plant Regeneration
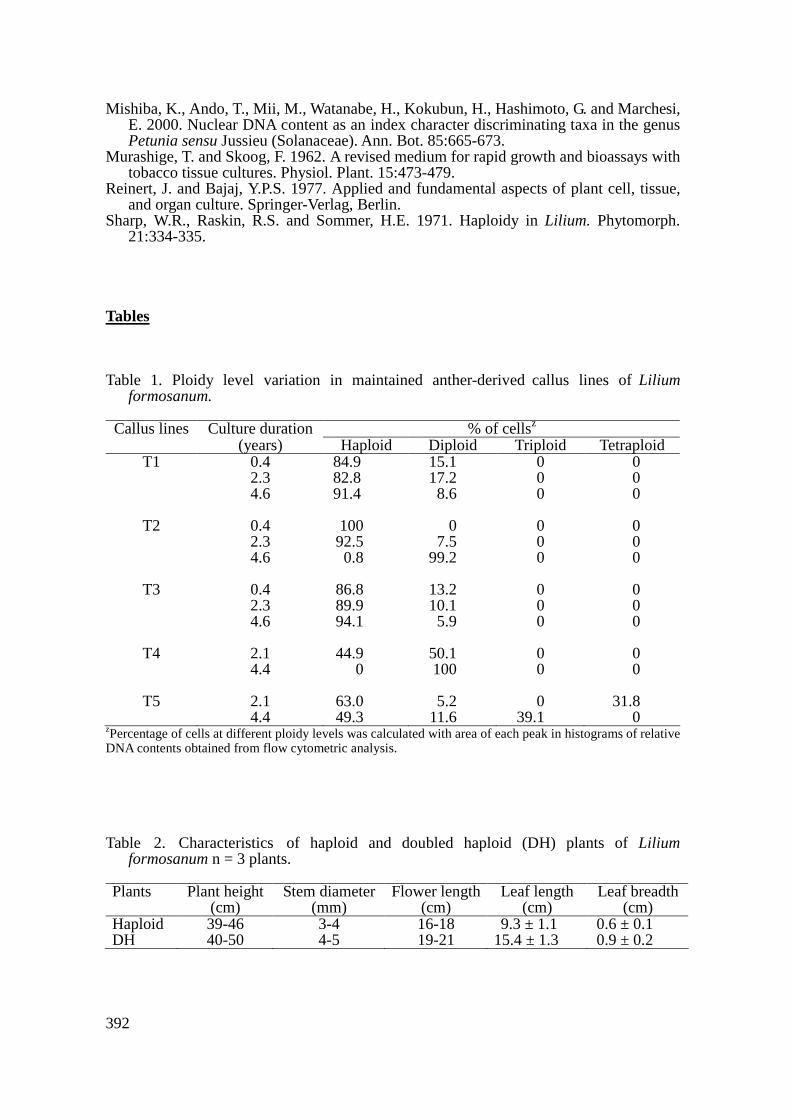
When anthers with uninucleate microspores were cultured for 3 to 4 months, two types of calluses were produced; nodular calluses developed within dehisced anthers, and white and compact ones formed from remnants of filaments. Since calluses from remnants of filaments were of somatic origin (Han et al., 1997), only calluses developed within dehisced anthers were used for callus proliferation and plant regeneration. Five callus lines developing within dehisced anthers were excised from the original explants and transferred to the callus-proliferating MS medium. Flow cytometric analysis showed that these callus lines had a high proportion (85 to 100%) of haploid cells, suggesting that they were of gametophytic origin. When haploid calluses were transferred to the MS regenerating medium, plantlets were regenerated 2 months after culture (Fig. 1A). Among 18 individual regenerants analyzed, fifteen were haploid (2n = x = 12; Fig. 1B), and three were diploid. Long-term Maintenance of Haploid Callus Lines
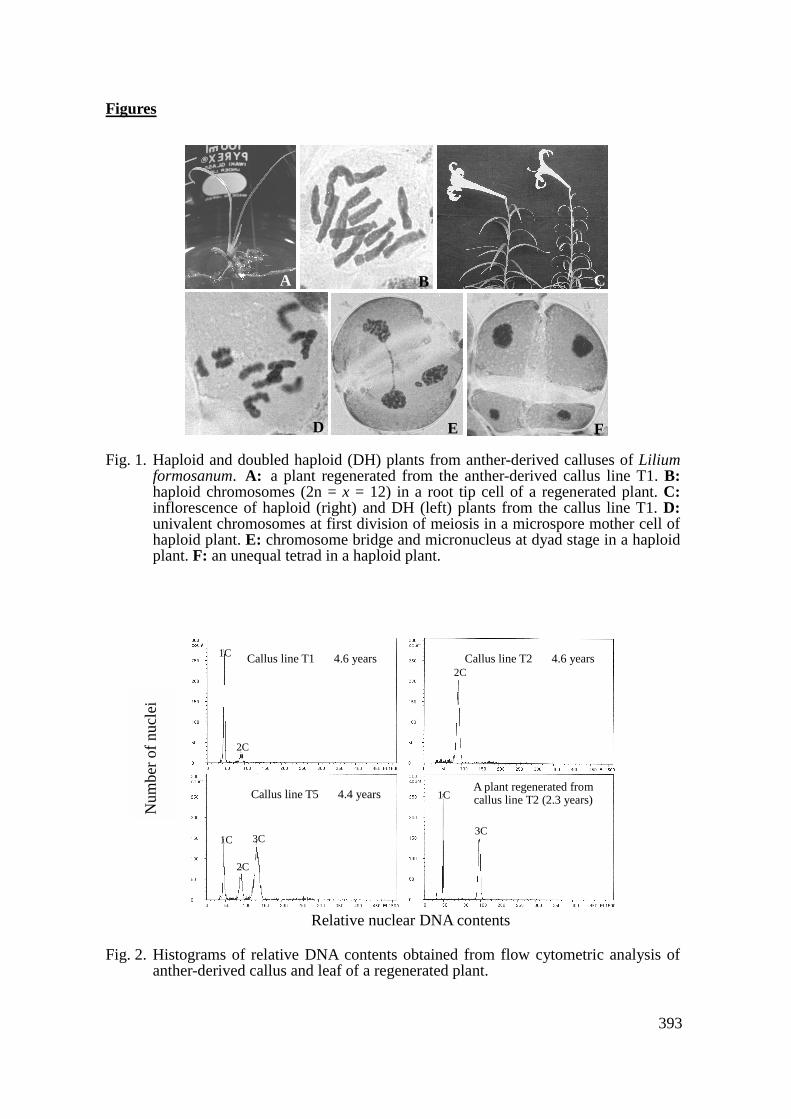
Haploid calluses were maintained on MS callus-proliferating medium in the dark at 25°C for more than 4 years. The calluses increased their volume 2 to 3 times each 2 to 3 months. Table 1 shows the changes in ploidy level of callus lines during subculture: The T1

391
and T3 lines generally remained at the haploid level, the T2 and T4 lines mostly changed into diploid, and the T5 line was mixoploid with haploid, diploid and triploid cells (Fig. 2). Our previous result showed that a haploid callus line of the Asiatic hybrid lily ‘Connecticut King’ could be maintained longer than 2 years (Han et al., 2000). Results of the present study also proved that anther-derived calluses of L. formosanum could be maintained long term at a haploid level, whereas a spontaneous ploidy variation of haploid calluses occurred during subculture. These culture systems will be useful materials for not only production of doubled haploid lily but also in vitro mutagenesis, genetic transformation and somatic hybridization (Reinert and Bajaj, 1977; Janick, 1979; Bhojwani and Razdan, 1983; Gleba and Shlumukov, 1990). The present study also showed that plantlets of haploid, diploid and mixoploid consisting of haploid and triploid cells have been regenerated from these callus lines maintained long term (Fig. 2). Characteristics of Haploid and Doubled Haploid Plants
Haploid and doubled haploid plantlets were obtained from haploid and colchicine-treated calluses. These plantlets were acclimated in the greenhouse and grew into flowering plants. Haploid plants showed poor characteristics with smaller flowers and leaves than those of DH ones (Table 2; Fig. 1C). Meiotic abnormalities and sterile pollen were observed in haploid plants (Fig. 1D, E, F). The investigated doubled haploid plants had normal flowers and leaves, similar to diploid ones, but pollen grains were still sterile. It is unclear why doubled haploid plants of L. formosanum were sterile. Further experiments are necessary to understand the reason. CONCLUSION
Five haploid callus lines of Lilium formosanum were obtained by anther culture and maintained more than 4 years. Two anther-derived calluses lines were maintained long term at a haploid level, whereas three lines showed a spontaneous doubling during subculture. Haploid, diploid and mixoploid plants could be regenerated from the anther-derived callus lines. Haploid plants showed poor growth and abnormal meiosis resulting in formation of sterile pollen. Doubled haploid plants can be produced from a colchicine-treated haploid callus line, but they form sterile pollens. Literature Cited Arzate-Fernández, A.M., Nakazaki, T., Yamagata, H. and Tanisaka, T. 1997. Production of
doubled-haploid plants from Lilium longiflorum Thunb. anther culture. Plant Sci. 123:179-187.
Bhojwani, S.S. and Razdan, M.K. 1983. Plant tissue culture: theory and practice. Elsevier, Amsterdam.
Chu, Y., Chen, L., Huang, Y. and Zhang, Y. 2001. Studies on anther culture of lily. Acta Hort. Sin. 28:472-474.
Gleba, Y.Y. and Shlumukov, L.R. 1990. Somatic hybridization and cybridization. p.316-345. In: S.S. Bhojwani (ed.), Plant Tissue Culture: Applications and Limitations, Developments in Crop Science 19, Elsevier, Amsterdam.
Gu, Z.-P. and Cheng, K.-C. 1982. Studies on induction of pollen plantlets from the anther cultures of lily. Acta Bot. Sin. 24:28-32.
Han, D.-S., Niimi, Y. and Nakano, M. 1997. Regeneration of haploid plants from anther cultures of the Asiatic hybrid lily ‘Connecticut King’. Plant Cell, Tiss. Org. Cult. 47:152-158.
Han, D.-S., Niimi, Y. and Nakano, M. 1999. Production of doubled haploid plants through colchicines treatment of anther-derived haploid calli in the Asiatic hybrid lily ‘Connecticut King’. J. Japan. Soc. Hort. Sci. 47:979-983.
Han, D.-S., Niimi, Y. and Nakano, M. 2000. Long term maintenance of an anther-derived haploid callus line of the Asiatic hybrid lily ‘Connecticut King’. Plant Cell, Tiss. Org. Cult. 61:215-219.
Janick, J. 1979. Horticultural science. W.H. Freeman and Company, San Francisco.
392
Mishiba, K., Ando, T., Mii, M., Watanabe, H., Kokubun, H., Hashimoto, G. and Marchesi, E. 2000. Nuclear DNA content as an index character discriminating taxa in the genus Petunia sensu Jussieu (Solanaceae). Ann. Bot. 85:665-673.
Murashige, T. and Skoog, F. 1962. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 15:473-479.
Reinert, J. and Bajaj, Y.P.S. 1977. Applied and fundamental aspects of plant cell, tissue, and organ culture. Springer-Verlag, Berlin.
Sharp, W.R., Raskin, R.S. and Sommer, H.E. 1971. Haploidy in Lilium. Phytomorph. 21:334-335.
Tables Table 1. Ploidy level variation in maintained anther-derived callus lines of Lilium
formosanum.
Callus lines Culture duration % of cellsz (years) Haploid Diploid Triploid Tetraploid
T1 0.4 84.9 15.1 0 0 2.3 82.8 17.2 0 0 4.6 91.4 8.6 0 0
T2 0.4 100 0 0 0 2.3 92.5 7.5 0 0 4.6 0.8 99.2 0 0
T3 0.4 86.8 13.2 0 0 2.3 89.9 10.1 0 0 4.6 94.1 5.9 0 0
T4 2.1 44.9 50.1 0 0 4.4 0 100 0 0
T5 2.1 63.0 5.2 0 31.8 4.4 49.3 11.6 39.1 0
zPercentage of cells at different ploidy levels was calculated with area of each peak in histograms of relative DNA contents obtained from flow cytometric analysis. Table 2. Characteristics of haploid and doubled haploid (DH) plants of Lilium
formosanum n = 3 plants. Plants Plant height
(cm) Stem diameter
(mm) Flower length
(cm) Leaf length
(cm) Leaf breadth
(cm) Haploid 39-46 3-4 16-18 9.3 ± 1.1 0.6 ± 0.1 DH 40-50 4-5 19-21 15.4 ± 1.3 0.9 ± 0.2
393
Figures Fig. 1. Haploid and doubled haploid (DH) plants from anther-derived calluses of Lilium
formosanum. A: a plant regenerated from the anther-derived callus line T1. B: haploid chromosomes (2n = x = 12) in a root tip cell of a regenerated plant. C: inflorescence of haploid (right) and DH (left) plants from the callus line T1. D: univalent chromosomes at first division of meiosis in a microspore mother cell of haploid plant. E: chromosome bridge and micronucleus at dyad stage in a haploid plant. F: an unequal tetrad in a haploid plant.
Fig. 2. Histograms of relative DNA contents obtained from flow cytometric analysis of
anther-derived callus and leaf of a regenerated plant.
A B C
D F E
1C
2C
Callus line T1 �4.6 years� Callus line T2 �4.6 years�
2C
3C
Callus line T5 �4.4 years� A plant regenerated from callus line T2 (2.3 years)
Relative nuclear DNA contents
3C
1C
1C
2C
Num
ber o
f nuc
lei





























