Embed Size (px)
Citation preview

B I O T E C H N O L O G Y L E T T E R S Volume 15 No.lO (0ct.1993) pp.997-1000 Received as revised 22nd September
PRODUCTION OF AN INDOLIZIDINE ALKALOID, SWAINSONINE BY THE
FILAMENTOUS FUNGUS, M E T A R H I Z I U M ANISOPLIAE.
M Patrick, M.W. Adlard and T. Keshavarz*
School of Biological and Health Sciences, Faculty of Engineering and Science,
University of Westminster, 115 New Cavendish Street London WIM 8JS
Summary The physiology ofMetarhi~um anisophae has been studied with a view to improving swainsonine producUon. In prelimina.~" work, the stability of pure s~ainsonine in aqueous solution has been investigated under varying conditions of pH and temperature and in the presence of oxidizing agents. The effects of o.~.gen on the growth and morpholoKv of M.anisopliae and the production of swainsomne were also examined Shaken flask fermentations have been carried out in order to develop a medium suitable for production of swainsonine.
Introduction Swainsonine, an indolizidine alkaloid was first isolated from small pea-like plants (e.g. Swainsona sp. and Astragalus sp.), and was shown to be the agent responsible for livestock poisoning. Poisoning symptoms were found to be similar to hereditary marmosidosis (Fellows et al., 1990). Swainsonine, a sugar analogue,is composed of a fused piperidine and pyrrolidine ring system
OH H OH (Figure 1). It is a powerful and highly specific inhibitor of plant ~ O and animal a-mannosidases (Dorling et al., 1980 and Tulsiani et
H al., 1985). Glycosidase inhibitors may be of therapeutic importance in the future. Hino et al. (1985) reported that administration of swainsonine in tumour-bearing mice reduced the size of the tumour by 50% and prevented metastasis. Two species of fungi which produce swainsonine have been
Figure 1. Swainsonine isolated. The proposed swainsonine biosynthetic pathway (See Molecular Structure Figure 2, Harris et al., 1980) is based on what has been established for Rhizoctonia leguminicola (Guengrich et al., 1973). Metarhizium cmisopliae was also reported to produce swalnsonine (Hino et al., 1985). However, little is known of the physiology of M.anisopliae in relation to swainsonine production and so far there has been no reported work on improving swainsonine production in wild-type strains.
H2NCHCOOH H2NCHCOO H O = C --COOH I pH >9.0 ,
! CH 2 , H CH 2 ~ S~cck~e CH 2 COOH ~ C O O H I ÷ CH 2 ~ i + CH 2 CH 2 ÷ NAD(P) "--'~"-~ i 1 [
CH2OOH I a,,.i ~ d ~ , H20 ~ NH CH2, C~!t2 ClH2
AI~I~ CH2NH2 ~ . ~ Agid CH2NHCHCOOH L-Ly~g
OA~ ~ ~ " ~ QI!
H2N
Figure 2. PrOposed Pathway for Biosynthesis of Swainsonine in Kleguminicola
997

Msterials and Methods Strains A swainsonine-producing strain of M.anisopliae was kindly provided by the Jodrell Laboratory, Royal Botanical Gardens, Kew. Stock cultures were maintained on oat agar (OA). Media and Reagents Modified Hino's production medium - The medium was similar to that of Hino (Hino et al., 1985), with the exception that 0.4%(w/v) corn gluten meal and 0.2%(w/v) cottonseed hydrolysate were replaced with 0.6% (w/v) Pharmamedia. Oat meal extract medium - Oat meal (Health food outlets) was soaked in distilled water, filtered through a fine muslin bag and the extract autoclaved for 15 rain at 121 °C, 15 psi. Where necessary, the following additional ingredients were added to the above media in shaken flasks (the figures in brackets indicate the final concentrations);- L-lysine (30 mg/L); COC13(4 mg/L); D-lysine (30 mg/L); DL-lysine (30 mg/L). Unless otherwise specified, all reagents and chemicals utlized were obtained fi'om Sigma Chemical Co Ltd., Poole, Dorset BH17 7BIL UK. pH and Temperature Stability Studies The pH of duplicate 1 ml aliquots of aqueous pure swainsonlne (10 rag/L) were adjusted over the range 2-10 before incubation at 20 °C. Additional duplicate aliquots at pH 6.5 were incubated at temperatures 0-100 °C. Each aliquot was sampled every 24 hours and swainsonine levels monitored every 7 days. Oxidation of Swainsonine Pure swainsonine was dissolved in water at final concentrations of 10 mg/L and I ml were aliquots dispensed into Eppendofftubes. Air was bubbled through one aliquot at a rate of 0.2 rrd/min for 5 minutes, the Eppendorf tube was then sealed and incubated at room temperature. 200111 of0.1M H20 2 was added to the other aliquot, before sealing the tube and incubating at room temperature. Swainsonlne levels in duplicate samples of each aliquot were monitored every 24 hours over a 7 day period. Preparation of Spore Suspension Spore suspensions were prepared using sterile distilled water/glass beads to scrape spores off slopes and were vortexed for 3-4 minutes with sterile glass beads to disperse spore chains. Effect of Different Dissolved Oxygen Concentrations Different volumes of 2%(w/v) OMX medium were added to 250 rnl conical flasks in duplicate,to produce 10°6, 20°6, 50o6 and 70°6 v/v of medium in the flasks. Each shaken flask containing the medium was autoclaved for 15 min. at 121 °C, 15 psi, inoculated with 200 !~1 of spore suspension containing 107-108 spores/ml and was incubated at 26°C in an orbital shaker (New Brunswick Scientific, Edison, N.J., U.S.A.) at 200 rpm. Media Studies using Different Supplement(s) All supplements were filter sterilized prior to addition into 250 ml conical flasks containing 50°6 v/v sterilized medium (2% OMX). The same inoculation and culturing procedure as above was adopted. Quantitative Swainsonine Analysis High performance cation exchange chromatography with post-column addition of NaOH and pulsed amperometric detection, (Dionex UK Ltd., Camberley, Surrey.) was used for analysis of swainsonine (Donaldson et al., 1990). Results and Discussion Fermentation profiles (Patrick et ai., unpublished results) of M.anisopliae show that after approximately 250 hours of growth swainsonine titres in the broth decline. To ascertain possible reasons for this decline, stability tests on pure aqueous swainsonine were carried out. Investigations on the effect ofa pH range between 2.0-10.0 on swainsonine standards showed that it was stable within this pH range over a time period of 7 days and at temperatures
998
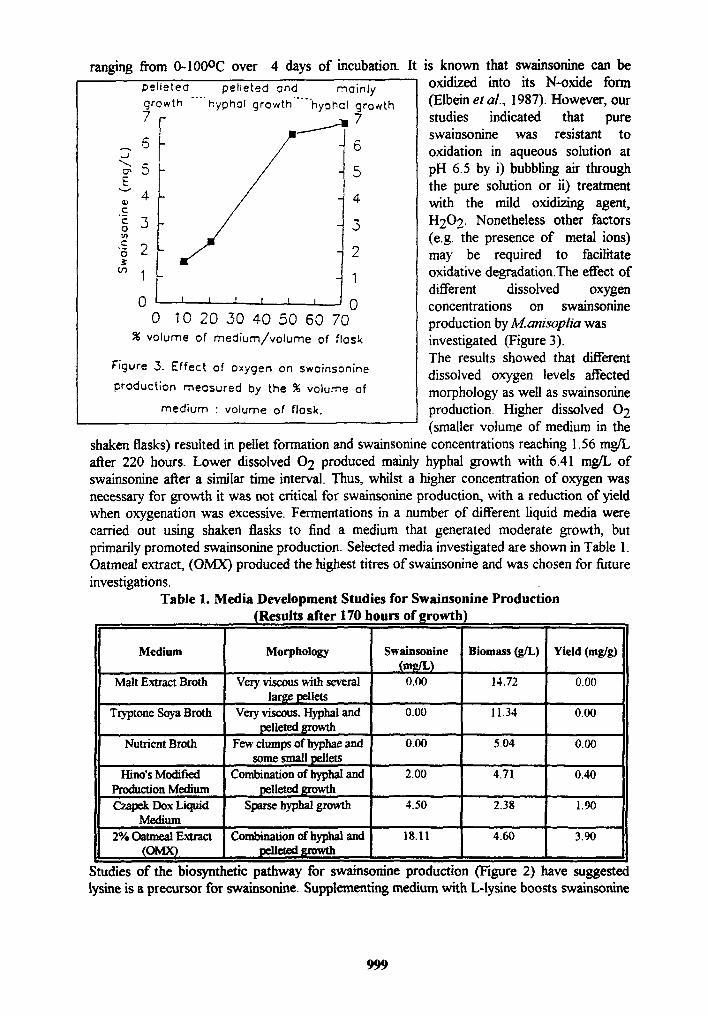
ranging from 0-100°C over 4 days of incubation. It is known that swainsonine can be pelleted pelleted and mainly oxidized into its N-oxide form growth .... hyphoI growth ..... hyol~oI growth (Elbeinetal., 1987). However, our 7 / l 7 studies indicated that pure
swainsonine was resistant to ~, 6 6 oxidation in aqueous solution at
5 5 pH 6.5 by i) bubbling air through E the pure solution or it) treatment
"~" 4 4 with the mild oxidizing agent, C
~ 3 .3 H202. Nonetheless other factors O
~' (e.g. the presence of metal ions)
J "6 2 2 may be required to facilitate ~n 1 1 oxidative degradation.The effect of
different dissolved oxygen 0 ' ' ' ~ f J 0 concentrations on swainsonine
0 10 2 0 3 0 4 0 5 0 6 0 70 production byM.anisopliawas % volume of medium/volume of flask investigated (Figure 3).
The results showed that different Figure 3. Effect of oxygen on swainsonine dissolved oxygen levels affected production measured by the % volume of morphology as well as swainsortine
medium • volume of flask, production. Higher dissolved 0 2 (smaller volume of medium in the
shaken flasks) resulted in pellet formation and swainsonine concentrations reaching 1.56 mg/L after 220 hours. Lower dissolved 0 2 produced mainly hyphal growth with 6.41 mg/L of swainsonine after a similar time interval. Thus, whilst a higher concentration of oxygen was necessary for growth it was not critical for swainsonine production, with a reduction of yield when oxygenation was excessive. Fermentations in a number of different liquid media were carried out using shaken flasks to find a medium that generated moderate growth, but primarily promoted swainsonine production. Selected media investigated are shown in Table 1. Oatmeal extract, (OMX) produced the highest titres of swainsonine and was chosen for future investigations.
Table 1. Media Development Studies for Swainsonine Production (Results after 170 hour, of growth)
Medium
Malt Extract Broth
Tryptone Soya Broth
Nutrient Broth
'Hino's Modified Production Medium Czapek Dox Liquid
Medium 2% Oatmeal Extract
(OMX)
Morphology
• Very viscous with several large pellets
Very viscous. Hyphal and l~ileted growth,,
Few clumps of hyphae and some small pellets
Combination of hyphal and .... pelleted ~,owth
Sparse hyphal growth
C~4.ombination of hyphal and pelleted growth,,,
Swainsonine (rag/L)
0.00
Biomass (g/L)
14.72
Yield (mg/g) ,==
0.00
0.00 11.34 0.00
0.00 5.04 0.00
2,00 4.?i 0.40
2.38 1.90 4.50
m ,
18.11 4.60 3.90
Studies o f the biosynthetic pathway for swainsonine production (Figure 2) have suggested lysine is a precursor for swainsonine. Supplementing medium with L-lysine boosts swainsonine
999
production (Table 2). There are conflicting reports on the preferred enantiomeric form of lysine for incorporation into pipecolic acid, a precursor in the swainsonine biosynthetic pathway (Harris et al., 1988). Studies on R.legummicola (Wickwire et al., 1990) indicated that L-lysine was the preferred substrate, although some L-pipecolate was also produced from D-lysine. The results of supplementing the 2% OMX medium with equal concentrations olD-, DL- and L-iysine are shown in Table 2. A 34% yield increase was achieved by supplementing with L-lysine whereas a 62% increase resulted after adding the D-isomer.
Table 2. Effects of Different Lysine Isomers on Swainsonine Production (Results after 190 hours growth)
Supplement
Control
L-I~ine
DL-lvsine
D-lvsine
Swainsonine (mg/L)
18.92
27.42
3067
46.24
Biomass (g/L) Yield (rag/g)
4.60 3.90
4.67 5.90
4.49
4.55
6.80
10.20
The L-isomer of lysine is the commonly available form in the intracellular pool of amino acids and this, not the D-isomer is readily used by the organism for protein synthesis and subsequent growth processes. Uptake of L-lysine in the medium by the cell could result in feedback inhibition of lysine biosynthesis..Since D-lysine may not cause feedback inhibition, the organism may continue to synthesize L-lysine, resulting in more lysine being available for swainsonine production. Guengerich and Broquist (1973) reported the presence of a lysine racemase in Klegummicola and established that L-pipeeolic acid is formed via L-lysine in this fungus. However, there have been no studies on which enantiomer oflysine is preferentially taken up by M.anisopliae for L- pipecolate and hence swainsonine biosynthesis. Referenc~
1. Donaldson, M.J., Broby,H., Adlard, M.W. & Bucke, C. (1990). PhytochemicalAnalysis 1: 18-21. 2. Dorling, P.R., Huxtable, C.R. and Colegate, S.M. (1980). Biochem.J. 191: 649-651. 3. EIlgin, A.D. & Molyneux, R_J. (1987). Alkaloids 5: 1-54. 4. Fellows, L.E. & Nash, ILJ. (1990). Sc/. Progres~ 2(294): 245-255. 5. Guengerich, F.P. & Broquist, H.P. (1973). Biochemistry 12(21): 4270-4274. 6. Guengerich, F.P., Dirnari, S.J. and Broquist, H.P. (1973). JAm. Chem. Soc. 95(6): 2055-2056. 7. Harris, C.M., Schneider, M.J., Ungemach, F.S., Hill, J.E. and Harris, T.M. (1988). JAm. Chem. Soc.
110. 940-949. 8. Hino, M., Nakayama, O., Tsmam~ Y., Adachi, K., Shibata, T., Terano, H., Kohsaka, M., Aoki, H. &
Imanaka, H. (1985). J.Ant~biot. 38: 926-935. 9. Tulsiani, D.R.P., Broqui~ H.P. & Tous'ter, O. (1985). Archives ofBiochem. & Biophys. 236(1): 427-
434. 10. Wickwire, B.M., Wagner, C. & Broquist, H.P. (1990). d.Biol. Chem. 265(25):14748-14753.
1000

![1 Synthesis of Saturated Five-Membered Nitrogen Heterocycles via … · 2010. 8. 24. · in the synthesis of the natural product indolizidine 209D [29], and asymmetric variants have](https://img.pdfslide.us/doc/110x75/60e1bd2b4c23e3589a77c569/1-synthesis-of-saturated-five-membered-nitrogen-heterocycles-via-2010-8-24.jpg)



























