Embed Size (px)
Citation preview

Organization of the skeleton – structure and function The skeletal system is a highly specialized connective tissue system comprising poorly cellular matrix together with the cells that make the matrix. Broadly, it consists of bones, cartilage, joints and ligaments. The structure and function of these components is greatly influenced by the nature of the matrix and fibrous elements. Bone is highly organized at a macroscopic and microscopic level, to serve its functions. The bony skeleton is divided into the axial and appendiceal skeleton. The axial skeleton consists of vertebrae, ribs, pelvic bones and ribs. The appendiceal skeleton consists of the long bones of the upper and lower limbs, and also the small tubular bones of the hands and feet. Grossly, the two types of bones are long bones and flat bones. Examples of flat bones are ribs and pelvic bones. Examples of long bones are humerus and femur. Anatomically, long bones may be divided in to the epiphyses, metaphyses and diaphysis. After puberty and in adults, the epiphyses becomes fused with the metaphyses.
Constituents of bone Bone consists of mineralised bone matrix and bone cells. These cells are responsible for the development and maintenance of an adequate structure, through the processes of modeling and remodeling. Over 90% of bone is composed of matrix, which is predominantly Type 1 collagen together with other non-collagenous proteins. This is fully mineralised in
adult bone. Unmineralised bone matrix is called osteoid. The bone cells are osteoblasts, osteocytes and osteoclasts.
The function of bones is: to provide structural support eg axial skeleton
to protect vital internal organs eg skull, ribs
to enable locomotion eg long bones of upper and lower limbs
to store calcium, and participate in calcium homeostasis
Types of bone:At a microanatomic level, there are two types of bone; cortical bone and cancellous bone. Bone in turn, also exist as two forms, woven bone and lamellar bone.Woven bone and lamellar bone are distinguished by the orientation of their collagen fibres. In woven bone, the collagen fibres are randomly arranged, and this can be seen under polarized light. This is an immature form of bone that is produced when bone is formed rapidly eg in the neonate or in the early stages of fracture repair. This is
subsequently remodelled into lamellar bone, which is stronger. Lamellar bone is composed of sheets or lamellae of parallel collagen. This can also be visualized under polarized light. Almost all bones in healthy adults consists of lamellar bone.
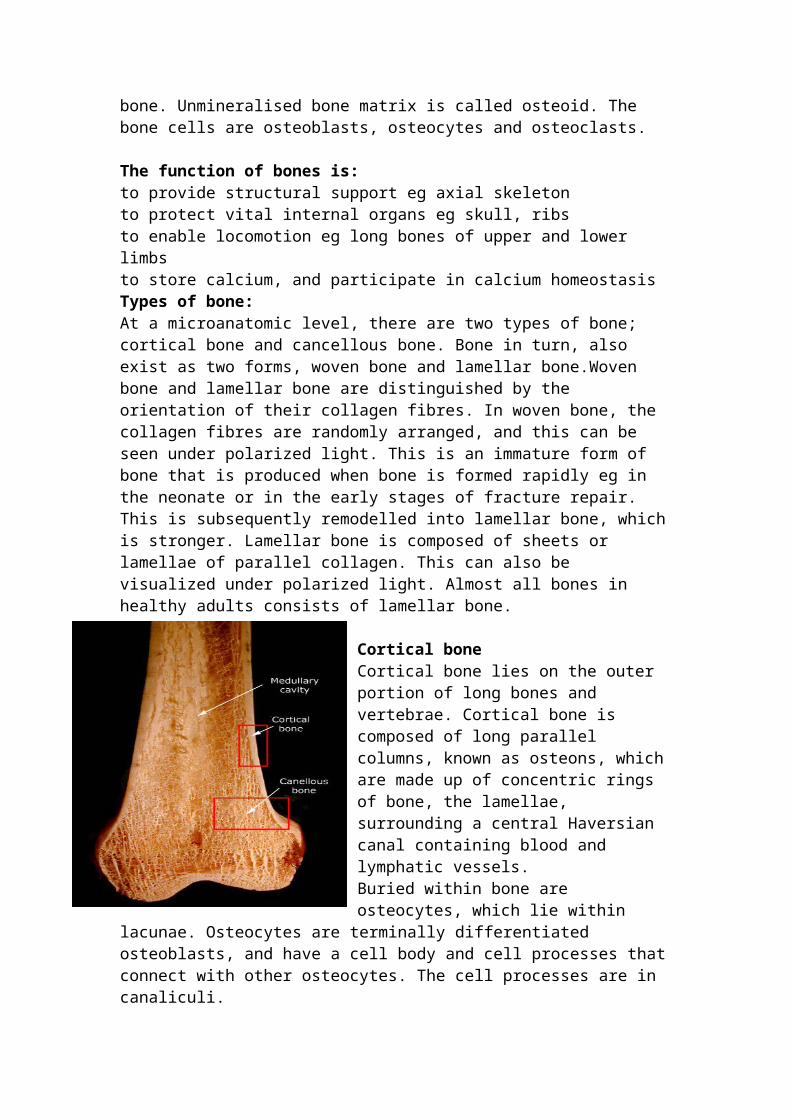
Cortical bone Cortical bone lies on the outer portion of long bones and vertebrae. Cortical bone is composed of long parallel columns, known as osteons, which are made up of concentric rings of bone, the lamellae, surrounding a central Haversian canal containing blood and lymphatic vessels. Buried within bone are osteocytes, which lie within lacunae. Osteocytes are terminally differentiated osteoblasts, and have a cell body and cell processes that connect with other osteocytes. The cell processes are in canaliculi. Cortical bone, in turn is covered by periosteum. This consists of fibrous connective tissue. The medullary cavity or center of bones is
composed of cancellous bone, with intervening marrow.
Cortical bone is composed of functional units called osteons. These consist of long parallel columns, each made up of concentric rings of bone, lamellae, around a central channel containing blood and lymphatic vessels. These channels are called the Haversian canals.
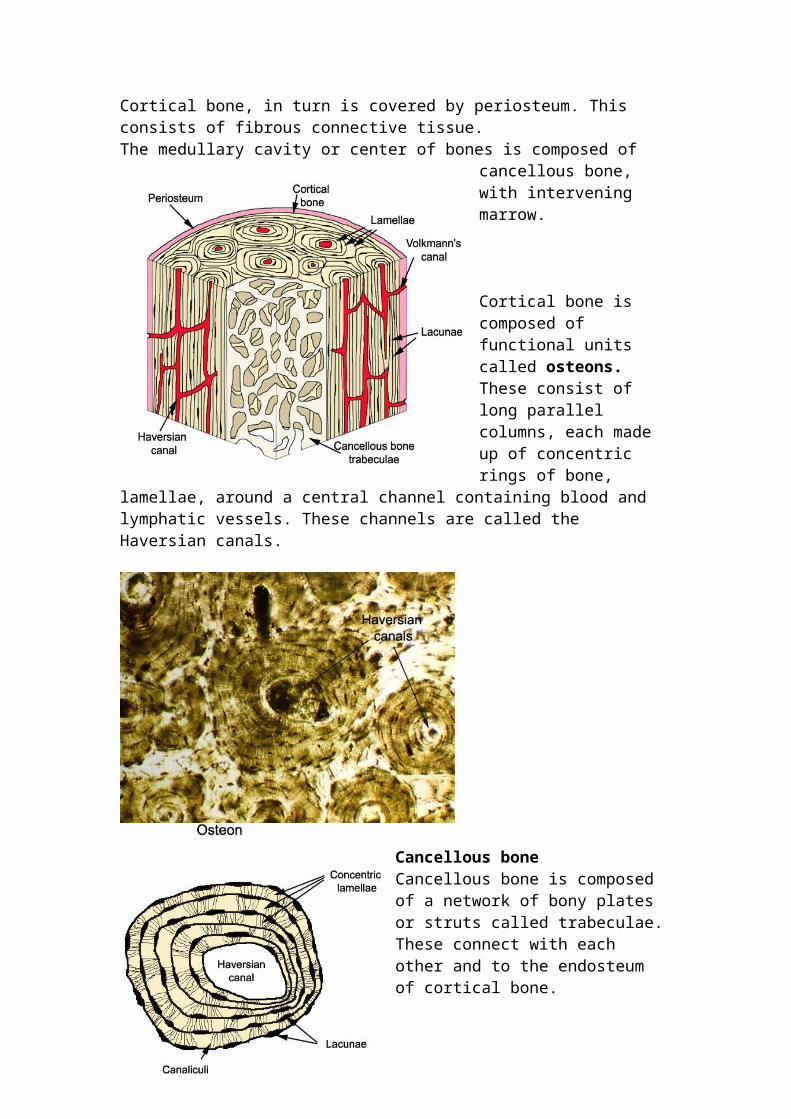
Cancellous bone Cancellous bone is composed of a network of bony plates or struts called trabeculae. These connect with each other and to the endosteum of cortical bone. In adults, the spaces between the trabeculae are filled with either haemopoetic bone marrow or adipose tissue.
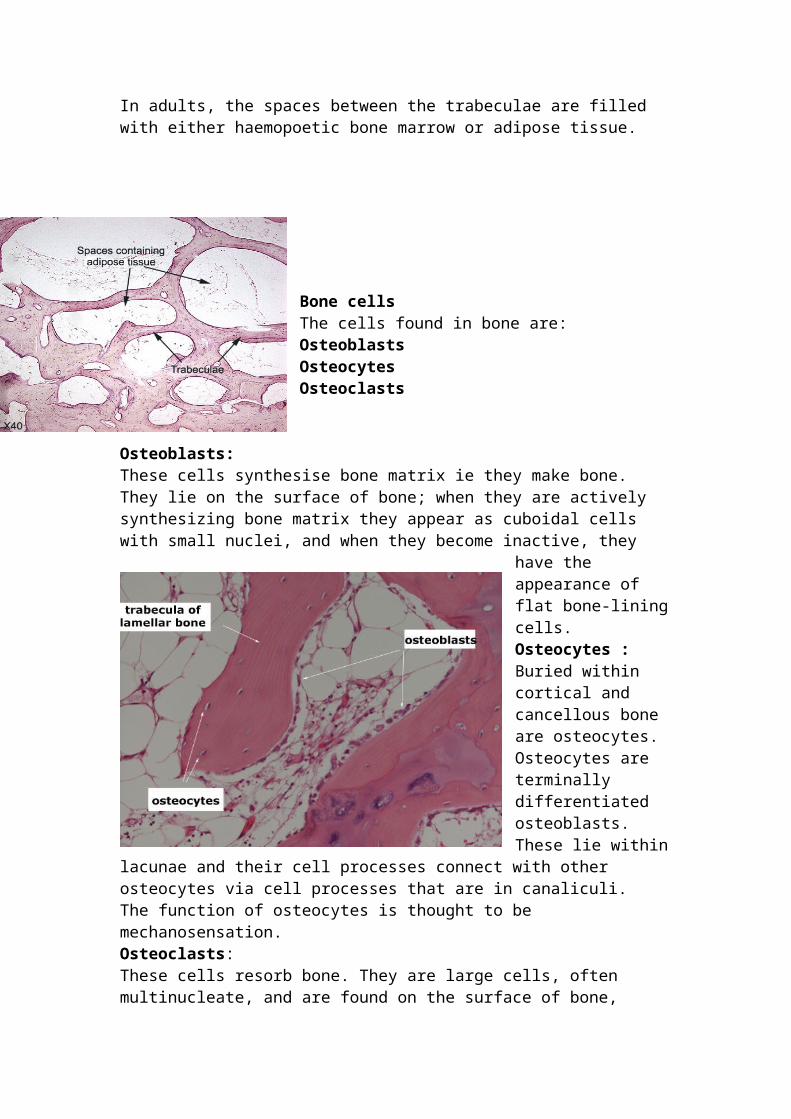
Bone cells The cells found in bone are: Osteoblasts
Osteocytes
Osteoclasts
Osteoblasts: These cells synthesise bone matrix ie they make bone. They lie on the surface of bone; when they are actively synthesizing bone matrix they appear as cuboidal cells with small nuclei, and when they become inactive, they have the appearance of flat
bone-lining cells. Osteocytes :Buried within cortical and cancellous bone are osteocytes. Osteocytes are terminally differentiated osteoblasts. These lie within lacunae and their cell processes connect with other osteocytes via cell processes that are in canaliculi. The function of osteocytes is thought to be mechanosensation.
Osteoclasts: These cells resorb bone. They are large cells, often multinucleate, and are found on the surface of bone, often in eroded surfaces or depressions that they have created.
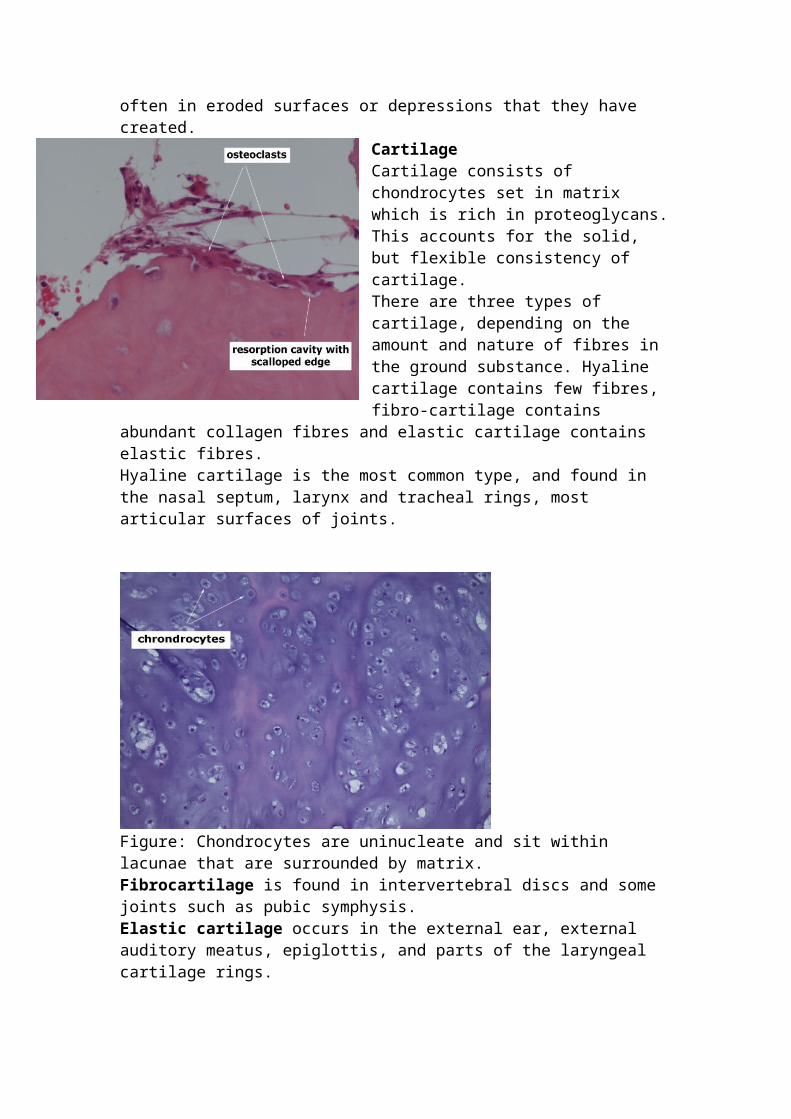
Cartilage Cartilage consists of chondrocytes set in matrix which is rich in proteoglycans. This accounts for the solid, but flexible consistency of cartilage. There are three types of cartilage, depending on the amount and nature of fibres in the ground substance. Hyaline cartilage contains few fibres, fibro-cartilage contains abundant collagen fibres and elastic cartilage contains elastic fibres. Hyaline cartilage is the most common type, and found in the nasal septum, larynx and tracheal rings, most articular surfaces of joints.
Figure: Chondrocytes are uninucleate and sit within lacunae that are surrounded by matrix. Fibrocartilage is found in intervertebral discs and some joints such as pubic symphysis. Elastic cartilage occurs in the external ear, external auditory meatus, epiglottis, and parts of the laryngeal cartilage rings.
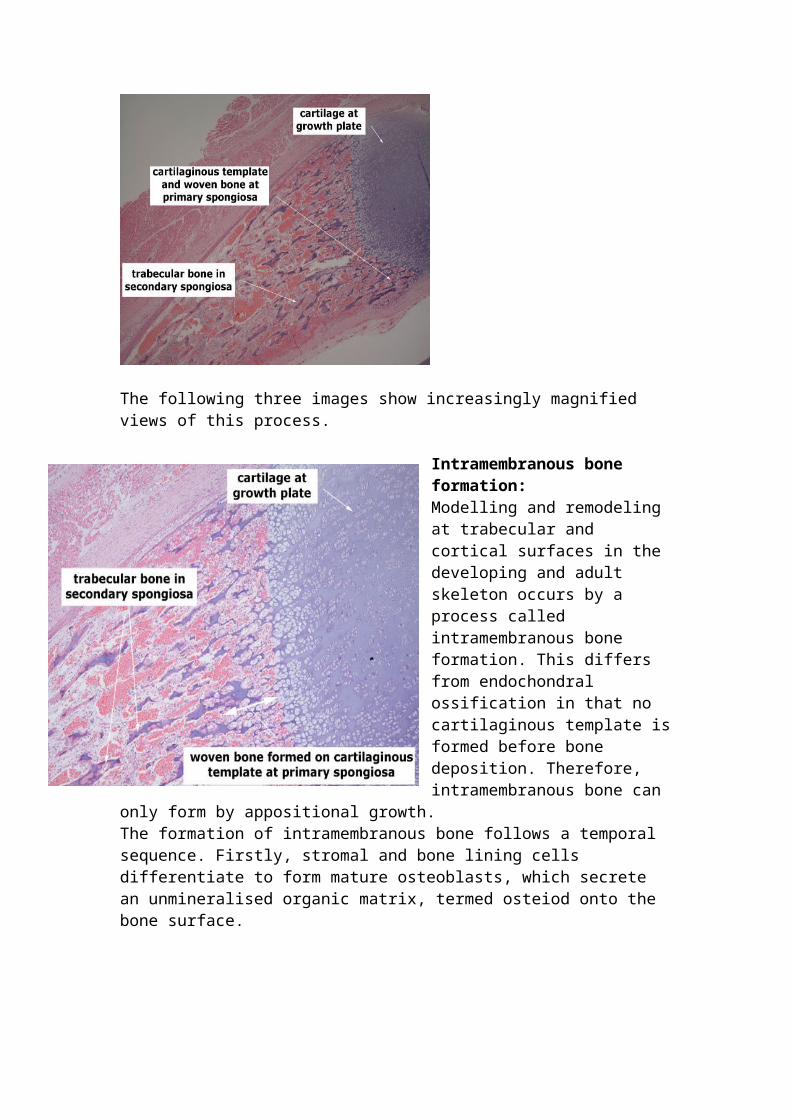
Bone development, bone formation and bone remodelling Endochondral ossification: Long bones elongate during development by a process called endochondral ossification, whereby a partially mineralized cartilaginous matrix formed at the growth plate becomes resorbed and replaced by woven bone initially, which is then subsequently remodelled into lamellar bone. Cartilage formation at the growth plate follows a sequential pattern, involving appositional and interstitial growth. The growth plate enlarges by appositional growth, as mesenchymal cells differentiate into matrix-secreting chondrocytes. These continue to divide within the proliferative zone, thereby further increasing the cartilage mass by interstitial growth. The matrix produced by chondrocytes in the proliferative zone contains poorly organized type II and type IX collagen fibrils, and several non-collagenous proteins including proteoglycans, chondrocalcin, osteonectin and matrix
gla protein. The proliferative chondrocytes progressively enlarge to form hypertrophic chondrocytes, which produce a matrix permissive for calcification in the hypertrophic zone by degrading type II collagen and increasing the synthesis of type X collagen and osteopontin. Once calcified, cartilage is partly resorbed by osteoclasts in the zone of vascular invasion and then replaced by woven bone to form the primary spongiosa. This is subsequently remodeled and replaced by lamellar bone forming the secondary spongiosa. This is a section showing the development of a fetal rib, which occurs by endochondral ossification.
The following three images show increasingly magnified views of this process.
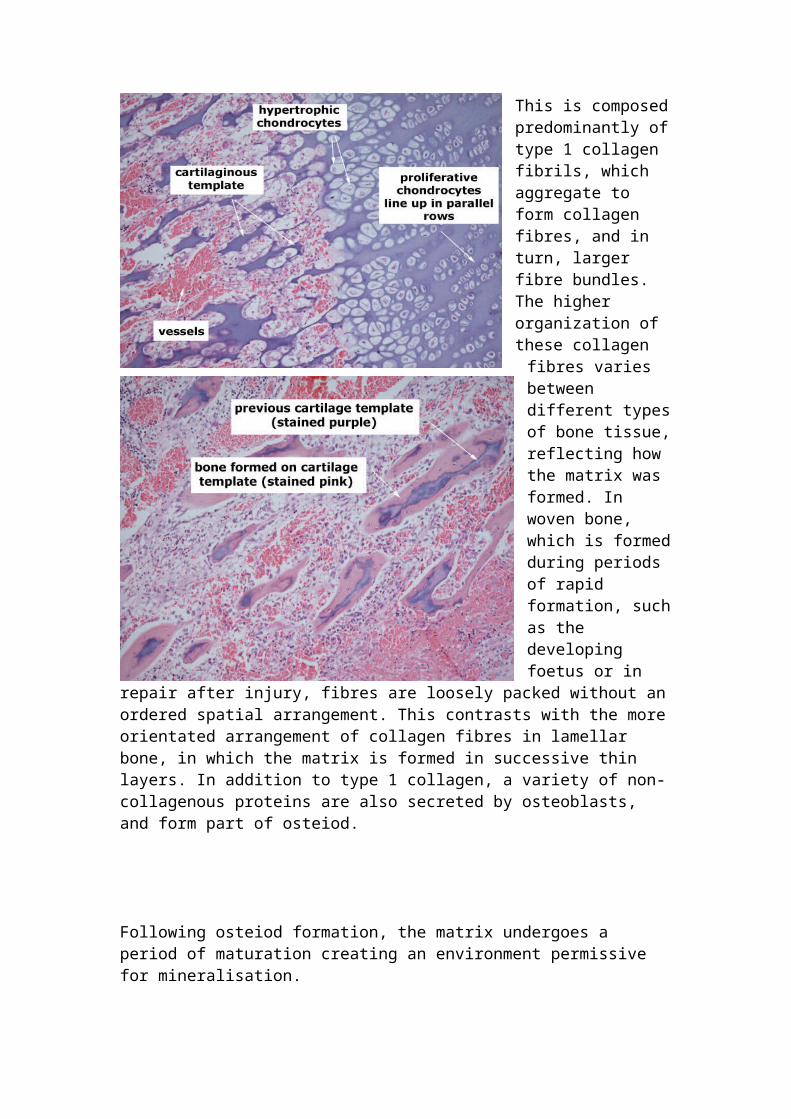
Intramembranous bone formation: Modelling and remodeling at trabecular and cortical surfaces in the developing and adult skeleton occurs by a process called intramembranous bone formation. This differs from endochondral ossification in that no cartilaginous template is formed before bone deposition. Therefore, intramembranous bone can only form by appositional growth. The formation of intramembranous bone follows a temporal sequence. Firstly, stromal and bone lining cells differentiate to form mature osteoblasts, which secrete an unmineralised organic matrix, termed osteiod onto the bone surface.

This is composed predominantly of type 1 collagen fibrils, which aggregate to form collagen fibres, and in turn, larger fibre bundles. The higher
organization of these collagen fibres varies between different types of bone tissue, reflecting how the matrix was formed. In woven bone, which is formed during periods of rapid formation, such as the developing foetus or in repair after injury, fibres are loosely packed without an ordered spatial arrangement. This contrasts with the more orientated
arrangement of collagen fibres in lamellar bone, in which the matrix is formed in successive thin layers. In addition to type 1 collagen, a variety of non-collagenous proteins are also secreted by osteoblasts, and form part of osteiod.
Following osteiod formation, the matrix undergoes a period of maturation creating an environment permissive for mineralisation. Calcium and phosphate is deposited within the organic matrix in the form of hydroxyapatite. In decalcified H&E sections, both calcified bone and osteiod appears pink, because calcium has been removed during histological processing. Focal amounts of calcium that remains renders mineralized bone a dark blue-purple colour. In undecalcified sections, special stains may be used to distinguish unmineralised matrix (osteiod) and fully mineralized bone.
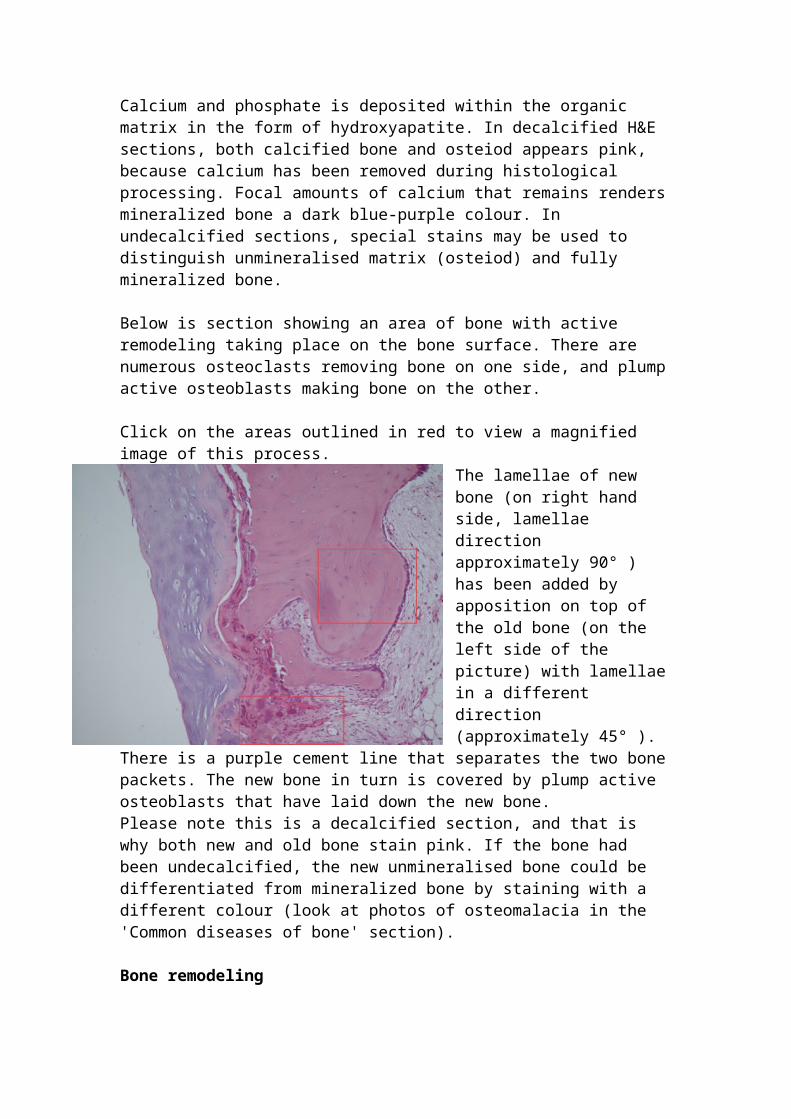
Below is section showing an area of bone with active remodeling taking place on the bone surface. There are numerous osteoclasts removing bone on one side, and plump active osteoblasts making bone on the other.
Click on the areas outlined in red to view a magnified image of this process.
The lamellae of new bone (on right hand side, lamellae direction approximately 90° ) has been added by apposition on top of the old bone (on the left side of the picture) with lamellae in a different direction (approximately 45° ). There is a purple cement line that separates the two bone packets. The new bone in turn is covered by plump active osteoblasts that have laid down the new bone. Please note this is a decalcified section, and that is why both new and old bone stain pink. If the
bone had been undecalcified, the new unmineralised bone could be differentiated from mineralized bone by staining with a different colour (look at photos of osteomalacia in the 'Common diseases of bone' section).
Bone remodeling This is a specialized form of intramembranous bone formation that takes place throughout life in the adult skeleton. This serves to remodel bone so that it remains structurally adapted to the mechanical needs of the body, in addition to participating in calcium homeostasis. It follows a strict temporal and spatial sequence. Bone is removed in small packets by osteoclasts, and osteoid is laid down by osteoblasts in the same location, thereby filling the resorption cavity and replacing the bone.
Common diseases of bone osteoporosis osteomalacia
Osteoporosis In osteoporosis, the total amount of bone is reduced, but the bone is fully and normally mineralised. This results in thinner trabeculae which are poorly connected with each other. This, in turn, causes the structural and mechanical integrity of bone to be diminished, such that bone fractures on minimal trauma.
Osteomalacia In osteomalacia, , the total amount of bone is preserved, but the amount of mineralized bone is reduced, and there is an increased amount of osteoid or unmineralised bone matrix.
Brief introduction to the organisation of the skeleton. This term, in order to start to understand how different organ systems interact within the body, you need to grasp some basics of each system. This DSL provides a grounding which will be built on next term. The skeletal system consists of bones, cartilage, joints and ligaments. There is sufficient detail in week 1 to cover all of these except bone
Bones have a role in: Support e.g. axial skeleton
Protection of internal organs e.g. skull and ribs
Enabling of locomotion e.g. long bones of the legs. Storage of calcium and participation in maintaining calcium balance in the blood.
Bone, like other specialised tissues, is made up of extracellular matrix containing specialised cells wrapped up in a covering - in this case thin fibrous tissue (periosteum), with a mechanism to control function. They have a rich blood supply and nerves. In mature bone, the extracellular matrix comprises 90% of the bone, is made up largely of type I collagen and is calcified. There are three specialised bone cell types: Osteoblasts: make osteoid a specialised extracellular matrix which is then calcified to form bone Osteoclasts: remove/resorb bone This recycling of bone (synthesis and resorption) is governed by hormones, vitamins and available calcium. The third specialised cell type is the osteocyte. These cells are derived from osteoblasts and sit deep within the bones. Osteocytes are fascinating as, despite the
nuclei being spaced far apart, the osteocytes are linked up by thin processes of cytoplasm connected by cell junctions. It is thought this web of osteocytes may have a role in sensation.
When looking at bone (without a microscope) you can see two appearances: closely packed ‘compact’ or ‘cortical’ bone, or bone looking like a calcified sponge so called spongy or cancellous bone. In some bones such as the femur the spaces in spongy bone contain bone marrow.
Microscopically, cortical bone has a very orderly arrangement with concentric cylinders of bone laid down around central blood vessels containing osteocytes (derived from osteoblasts trapped in the bone as it was made). More blood vessels connect the central vessels horizontally to form a complex mesh of small arteries and veins. These orderly cylindrical structures are known as Haversian systems or ‘osteons’. Osteons are packed closely together to form cortical bone and arranged as irregular plates in spongy bone. A less structured appearance is seen in bone as it is newly made and this is known as ‘woven’ bone.Common diseases of bone Osteoporosis: In osteoporosis, the total amount of bone is reduced, but the bone is fully and normally mineralised. This results in thinner trabeculae which are poorly connected with each other. This, in turn, causes the structural and mechanical integrity of bone to be diminished, such that bone fractures on minimal trauma.