Embed Size (px)
Citation preview

Primitive Stem Cells Derived from Bone MarrowExpress Glial and Neuronal Markers
and Support Revascularization in Injured RetinaExposed to Ischemic and Mechanical Damage
Nitza Goldenberg-Cohen,1–3,* Bat-Chen R. Avraham-Lubin,1,4,* Tamilla Sadikov,1,3
Ronald S. Goldstein,4 and Nadir Askenasy5
Ischemic or mechanical injury to the optic nerve is an irreversible cause of vision loss, associated with limitedregeneration and poor response to neuroprotective agents. The aim of this study was to assess the capacity ofadult bone marrow cells to participate in retinal regeneration following the induction of anterior ischemic opticneuropathy (AION) and optic nerve crush (ONC) in a rodent model. The small-sized subset of cells isolated byelutriation and lineage depletion (Fr25lin - ) was found to be negative for the neuroglial markers nestin and glialfibrillary acidic protein (GFAP). Syngeneic donor cells, identified by genomic marker in sex-mismatchedtransplants and green fluorescent protein, incorporated into the injured retina (AION and ONC) at a frequencyof 0.35%–0.45% after intravenous infusion and 1.8%–2% after intravitreous implantation. Perivascular cells withastrocytic morphology expressing GFAP and vimentin were of the predominant lineage that engrafted afterAION injury; 10%–18% of the donor cells incorporated in the retinal ganglion cell layer and expressed NeuN,Thy-1, neurofilament, and beta-tubulin III. The Fr25lin - cells displayed an excellent capacity to migrate to sitesof tissue disruption and developed coordinated site-specific morphological and phenotypic neural and glialmarkers. In addition to cellular reconstitution of the injured retinal layers, these cells contributed to endothelialrevascularization and apparently supported remodeling by secretion of insulin-like growth factor-1. Theseresults suggest that elutriated autologous adult bone marrow-derived stem cells may serve as an accessiblesource for cellular reconstitution of the retina following injury.
Introduction
Ischemic injury of the retina usually leads to the char-acteristic gliosis-type scarring and loss of vision, with little
chance of spontaneous recovery. Efficacy studies of neuro-protective agents have been widely performed in rodentmodels of anterior ischemic optic neuropathy (AION) andtraumatic optic nerve injury [optic nerve crush (ONC)] [1–3].However, vision preservation has been generally less thansatisfactory [4–6], because necrotic and apoptotic death of theretinal ganglion cells (RGCs) had already commenced prior todrug administration [7–11].
To overcome this problem, researchers are directing at-tention to stem cell therapy. Neuronal and retinal progeni-
tors, induced pluripotent cells, and embryonic stem cellshave been shown to incorporate into injured retina at vari-able frequencies [12,13]. The bone marrow is a potential ac-cessible source of autologous cells for retinal cell therapy,without the disadvantage of the immune barrier inherent toallogeneic cell transplants. Studies have suggested thatcommitted retinal progenitors residing in the bone marrowcompartment can be mobilized to contribute to retinal repair[14,15], particularly endothelial precursors and hemangio-blasts that support angiogenesis and neovascularization [16–18]. Accordingly, murine SCA-1 + lin - c-kit + and humanCD133 + hematopoietic precursors were able to regenerateretinal pigment epithelium and support revascularization[19–22] for extended periods [23]. In all instances, acute
1Krieger Eye Research Laboratory and 2Department Ophthalmology, Pediatric Unit, Schneider Children’s Medical Center of Israel,Petach Tikva, Israel.
3Sackler School of Medicine, Tel Aviv University, Tel Aviv, Israel.4Mina and Everard Goodman Faculty of Life Sciences, Bar Ilan University, Ramat Gan, Israel.5Frankel Laboratory, Center for Stem Cell Research, Schneider Children’s Medical Center of Israel, Petach Tikva, Israel.*Both authors contributed equally to this study as first authors.
STEM CELLS AND DEVELOPMENT
Volume 21, Number 9, 2012
� Mary Ann Liebert, Inc.
DOI: 10.1089/scd.2011.0366
1488
injury was an essential prerequisite for cell incorporation inthe retina [19–23]. In proof-of-concept studies of the capacityof adult bone marrow cells to adopt neural and glial phe-notypes in vivo [24–27], researchers reported a distinctmechanism of cell contribution to tissue repair in addition tosupport of endogenous recovery from injury [16,28]. Thesedata are consistent with incidental findings of neurons, as-trocytes, and microglia of donor origin in the brain of re-cipients of sex-mismatched bone marrow transplants [29].However, efforts to induce such differentiation in experi-mental models and in vitro have been largely inconclusive[30,31].
Bone marrow cells isolated by counterflow centrifugalelutriation and depleted of hematopoietic lineage markershave limited radioprotective activity but are endowed withindefinite hematopoietic reconstituting potential [32,33]. Thecells differentiate into hepatocytes in vitro and in vivo [34]and into endocrine pancreas and endothelial cells in vivo[35]. In addition, single cells are able to regenerate multipleepithelial tissues [36]. The aim of the present study was toassess the capacity of this subset of bone marrow cells toparticipate in neogenesis of neural and glial lineages inmurine models after induction of ischemic and mechanicalretinal injury. These models are particularly suited to eval-uation of cell therapies, because the injured retina can beuninterruptedly accessed through small and medium-sizedblood vessels [8–11].
Materials and Methods
Animal preparation and transplantation
Two mouse strains were purchased from Jackson Labora-tories (Bar Harbor, ME): C57BL/6J (B6, CD45.2), B6.SJL-Ptprca
Pepcb/BoyJ (CD45.1), and C57BL/6-TgN(ACTbEGFP)1Osb[green fluorescent protein (GFP), CD45.2]. The mice weremaintained and handled in accordance with the ARVOStatement for the Use of Animals in Ophthalmic and VisionResearch and the NIH guidelines, and the protocols wereapproved by the institutional Animal Care Committee.
Injury models
The models were based on a series of preliminary studiesintended to optimize several variables in the experimentalapproach. In all experiments, injury was induced in 1 eyewith the contralateral eye serving as a control. The inductionof AION was performed as previously described [2]. Fifty-three mice were anesthetized with ketamine 80 mg/kg (FortDodge Laboratories, Fort Dodge, IA) and xylazine 4 mg/kg(Veterinary Medicines Directorate, Surrey, UK). A plasticcustom-designed fundus corneal contact lens was used fordirect in vivo visualization of the retina and optic nervehead. AION was induced by intravenous injection of 40mL of2.5 mM Rose Bengal dye (Sigma, St. Louis, MO) followed byselective photoactivation above the optic disk at 532 nm withan argon laser beam at the following specifications: 200mmspot size, 50 mW power, and constant duration (0.1 s) [2]. Inall mice, the right eye was injured and the fellow (healthy)eye served as an internal control. The injury was re-evaluatedby RGC count in tissue sections; a 35%–40% RGC loss waspreviously reported for this reproducible procedure [2,8].ONC was induced by 3 repeated applications of a forceps
for 7 s at *3.0 mm posterior to the eye globe [3]. Studieshave shown that crush of the optic nerve leads to *70%RGC loss [3].
Isolation of bone marrow cells
Nucleated bone marrow cells were harvested fromC57BL/6 mice constitutively expressing GFP. Femurs andtibia were loaded onto a Beckman counterflow elutriationcentrifuge (Fullerton, CA; www.beckmancoulter.com)equipped with a J-6 rotor [37]. Fractions were collected atincreasing medium flow rates and a constant rotor speed(3,000 rpm) in order to isolate the smallest subset of nucle-ated cells at a flow rate of 25 mL/min (Fr25) and large pro-genitors at rotor off (R/O) at rest. Cells expressing lineagemarkers were depleted using saturating amounts of anti-bodies against T cells (CD5), B lymphocytes (B220), granu-locytes (GR-1), and macrophages and monocytes (Mac-1),obtained from hybridoma cell line cultures (ATCC, Mana-ssas, VA; www.atcc.org), and erythroid cells (Ter-119,eBiosciences, San Diego, CA; www.ebioscience.com). Cellswere coated with monoclonal antibodies, washed twice withphosphate-buffered saline (PBS), and incubated with sheep-anti-rat IgG conjugated to M-450 Dynal magnetic beads at aratio of 4 beads per cell (Invitrogen, Oslo, Norway;www.invitrogen.com). Rosetted cells were precipitated byexposure to a magnetic field, and the supernatant containinglineage-negative (Fr25lin - ) cells was collected. The effec-tiveness of the isolation procedure was confirmed by flowcytometery (Vantage SE flow cytometer, Becton Dickinson,Franklin Lakes, NJ; www.bdbiosciences.com). The cells weresuspended in RBC lysis buffer, washed with PBS, and la-beled with monoclonal antibodies for 30 min at 4�C in thepresence of 1mL mouse serum [37].
Stem cell administration
The elutriated Fr25lin - subset harvested from the donormice was transplanted into the congenic C57BL/6 recipientsusing 2 routes of administration: 106 cells in 0.2 mL PBS in-fused intravenously and 2 · 105 cells in 3mL PBS inoculatedintravitreally (Table 1). Treatment was administered 24 hafter injury induction on the basis of preliminary findings ofbetter cell incorporation with this schedule than with treat-ment before or immediately after injury (Fig. 1A). Con-ditioning was eliminated for all transplantations, becauseour preliminary trials showed that sublethal irradiation
Table 1. Transplantation of Fr25Lin-
Cells
in Murine Models of Retinal Injury
Route of injection
Injurymodel
Cell typetransplanted
IV(n = 30)
IVT(n = 23)
Total(n = 53)
AION Fr25Lin - 16 10 26R/O 9 3 12
ONC Fr25Lin - 5 10 15
AION, anterior ischemic optic neuropathy; ONC, optic nervecrush; Fr25Lin - , fraction 25 lineage-negative; R/O, rotor off; IV,intravenous; IVT, intravitreal.
BONE MARROW CELLS DIFFERENTIATE IN INJURED RETINA 1489
(650 rad, 106 rad/min) using an X-ray irradiator (RadSource2000, Suwanee, GA; www.radsource.com) at 1 day prior totreatment improved hematopoietic engraftment but reducedquantitative cell incorporation in the retina.
Histology and immunohistochemistry
Mice were sacrificed at 30 days after transplantation, andthe tissues were analyzed by histology and immunohisto-chemistry. The eye bulb was enucleated, fixed in 4% para-formaldehyde overnight, and then transferred to 20% sucroseovernight at 4�C. Samples embedded in frozen OCT medium(Tissue-Tek, Sakura, Finerek, CA; www.sakura-americas.com)were cryosectioned at 6mm thickness using a cryotome(Thermo Shandon, Cheshire, UK; www.thermo.com). Sections
were blocked for 15 min at room temperature with PBS con-taining 2% bovine serum albumin and 0.5% Triton X-100,followed by several washes. Every 10th slide from each eyewas selected for staining of various tissue markers by over-night incubation at 4�C: GFP (rabbit polyclonal, 1:100; SantaCruz Biotechnology, Santa Cruz, CA; www.scbt.com), glialfibrillary acidic protein (GFAP; mouse polyclonal, 1:250; Mil-lipore, Billerica, MA; www.millipore.com), vimentin (chickenpolyclonal, 1:500; Millipore), CD45 (rat, 1:50; Millipore), NeuN(mouse, 1:100; Millipore), beta-tubulin III (mouse, 1:100; Pro-mega, Madison, WI; www.promega.com), CD31 (rat, 1:100;Millipore), and Thy-1 (Mouse 1:100; Millipore). Primary an-tibodies were counterstained with NorthernLights� 493 (NL-493)-conjugated donkey anti-rabbit (1:200; R&D Systems,Minneapolis, MN; www.RnDSystems.com), NL557-conjugated
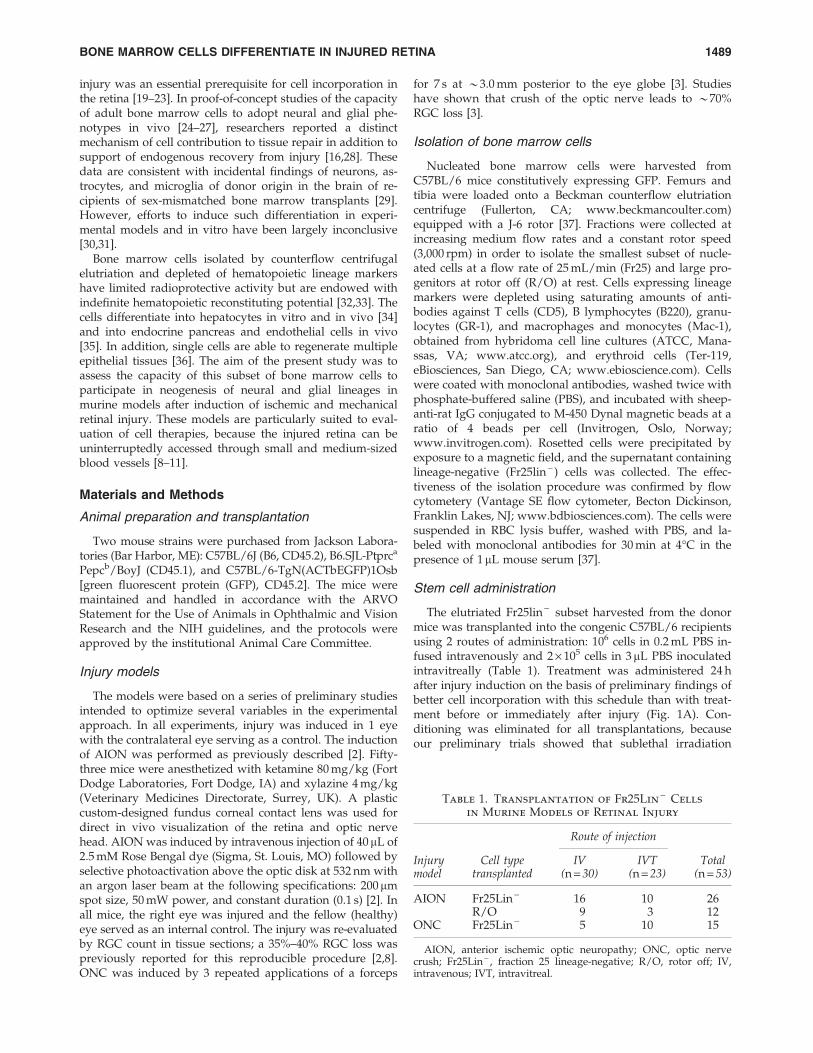
FIG. 1. Incorporation of do-nor cells in the injured opticnerve and retina. (A) The ex-perimental sequence includedinduction of ischemia [ante-rior ischemic optic neuropa-thy (AION)] or optic nervecrush (ONC) at 1 day beforecell transplant, with the con-tralateral eye serving as theinjury control. Enucleatedeyes were assessed by immu-nohistochemistry and fluores-cence in situ hybridization(FISH) at 1 month after trans-plantation. (B) Longitudinalsections of the optic nerve( · 40) showing massive gliosis(vimentin) and modest in-flammation (CD45) in theAION and ONC models. (C)Fr25lin - GFP + donor cells in-corporated in the anteriorvasculature (left panel) andretinal ganglion cell (RGC)layer (middle panel), adoptingthe characteristic morphology(scale bar: 20mm); note theirabsence in the control eyewithout injury (right panel).(D) Detection of donor cellsusing a genomic marker inmale-to-female transplants byFISH demonstrating XY malecells incorporated in the RGClayer (white arrows) along hostXX genotypes (scale bar:30 mm).
1490 GOLDENBERG-COHEN ET AL.

donkey anti-mouse (1:200; R&D Systems), NL557-conjugatedgoat anti-chicken (1:200; R&D Systems), and goat anti-ratmonoclonal antibodies (1:200; Molecular Probes, Eugene,OR; www.invitrogen.com).
Fluorescence in situ hybridization
Grafted cell incorporation into the vascular and RGClayers was validated by fluorescence in situ hybridization(FISH) in male-to-female transplants [38]. In brief, FISH wasperformed in sections washed in 0.09% saline and 2 · salinesodium citrate buffer (SSC) and digested by 0.5 mg/mLproteinase K. Sections were dehydrated at ascending ethanolconcentrations (70%–100%) and air-dried. Thereafter, 10mLof the probe solution labeled with PlatinumBright 495 and550 (Poseidon FISH Probes, Amsterdam, The Netherlands;www.kreatech.eu) were applied overnight at 37�C, followedby immersion in 0.4 · SSC and 0.3% octylphenyl-polyethyleneglycol (Igepal; Sigma, St. Louis, MO; www.sigmaaldrich.com) for 2 min at 72�C in 2 · SSC and 0.1% Igepal for 1 minat room temperature. They were then dehydrated again andmounted with antifade containing diamidino-2-phenylindole(Vectashield, Vector Laboratories, Southfield, MI; www.vectorlabs.com).
Microscopy
Images were acquired with an Axioplan 2 fluorescencemicroscope (C. Zeiss, Gottingen, Germany; www.zeiss.com)equipped with an Apotome using AxioVision 4.5 software.Narrow-band excitation and emission filters (rhodamine540/580 nm, FITC-GFP 485/530 nm, and DAPI-Hoechst360/455 nm) at 10 · –100 · magnifications were used. Theimages were pseudocolored (Adobe Photoshop) and re-constructed from RGB layers.
Quantitative assessment of cell incorporationin the injured retina
To estimate the number of labeled cells in each eye, 6–15nonoverlapping fields of view were assessed in every 10thslide, with each slide including 3 consecutive cryosections.The incorporation of transplanted cells was calculated bydividing the number of donor cells (GFP + ) by the number ofcells staining positive for tissue markers in the relevant layer.The total number of donor cells was calculated by multi-plying the number of cells per section by the number ofslides (10) and correcting for the spherical shape of the retina:Pr2/4r2 =P/4. The homing efficiency (in percent) was cal-culated by dividing the total number of GFP + cells by thenumber of infused cells.
Polymerase chain reaction
Real-time quantitative reverse transcription–polymerasechain reaction (qRT-PCR) was used to study the expressionsof nestin, GFAP, insulin-like growth factor-1 (IGF-1), fibro-blast growth factor-2 (FGF-2), transforming growth factor-b(TGF-b), and tumor necrosis factor-a (TNF-a). Elutriatedfractions were frozen in liquid nitrogen. Total RNA was ex-tracted with TRIzol (Invitrogen), followed by reverse tran-scription into cDNA using random hexamers (AmershamBiosciences, Cardiff, UK; www.amershambiosciences.biz) and
Moloney murine leukemia virus reverse transcriptase (Pro-mega). cDNA was analyzed with a Sequence Detection Sys-tem (ABI Prism 7000; Applied Biosystems, Foster City, CA;www.appliedbiosystems.com). Gene expression was mea-sured by normalizing cDNA input levels against mouse betaactin [39]. Reactions were performed for forward and reverseprimers in Master Mix buffer (SYBR Green I; Applied Bio-systems). Duplicate transcriptase-based quantitative PCRswere tested for each gene to minimize individual samplevariability and were normalized against standard curves. Theresults were quantified by the comparative 2 - ddCt method,using the following formula: ddCt = dCt (sample) – dCt (ref-erence gene). The following primers were used: b-actin(ACTB): sense TAG GCA CCA GGG TGT GAT GGT, anti-sense CAT GTC GTC CCA GTT GGT AAC A; nestin: senseAAC TGG CAC ACC TCA AGA TGT, antisense TCA AGGGTA TTA GGC AAG GGG; GFAP: sense CCA GCT TCGAGC CAA GGA, antisense GAA GCT CCG CCT GGT AGACA; IGF-1: sense AGA GAC CCT TTG CGG GGC, antisenseCGG ATA GAG CGG GCT GCT T; FGF-2: sense AAG CGGCTC TAC TGC AAG AAC G, antisense CCC AGT TCG TTTCAG TGC CAC; TGF-b: sense ATG ACA TGA ACC GGCCCT TC, antisense ACT TCC AAC CCA GGT CCT TC; TNF-a: sense TCT CAA AAT TCG AGT GAC AAG C, antisenseACT CCA GCT GCT CCT CCA C.
Statistics
All data are expressed as mean – standard error. The re-sults were statistically analyzed with SPSS for Windows,version 15.0.1 (SPSS, Inc., Chicago, IL). Two-way ANOVAwith repeated measures was used to analyze within subjectfactor type (route of injection, differentiation). A P valueof less than 0.05 was considered significant. Because ofthe relatively small size of some groups, we applied thenonparametric Mann–Whitney test for 2 independentsamples.
Results
Incorporation of donor cells in injured retina
Without cell transplantation, both AION and ONC led tocell loss in the retinal ganglion layer, of approximately 30%and 70%, respectively (Fig. 1).
Both AION and ONC caused massive gliosis but onlymodest inflammation (Fig. 1B). FISH study validated thepredominant incorporation of the elutriated Fr25lin - subsetinto the anterior vascular and RGC layers in the male-to-female transplants (Fig. 1D). In the particular area of theretina presented in Fig. 1D, the X chromosome was detectedin 85% of the RGC nuclei; 6 cells (30%) displayed the male X-Y genotype, in addition to cells incorporated in the innernuclear layer (GFP + cells; Fig. 1C, left and middle). Thecontrol eyes showed virtually no cell incorporation in theabsence of injury (Fig. 1C, right).
Differentiation of Fr25lin - cells in the injured retina
The subset of elutriated Fr25lin - cells used in this studyexpressed the panhematopoietic marker CD45 at variablelevels; however, the majority of cells that incorporated inthe RGC layer were CD45 negative (Figs. 2A and Table 1).
BONE MARROW CELLS DIFFERENTIATE IN INJURED RETINA 1491
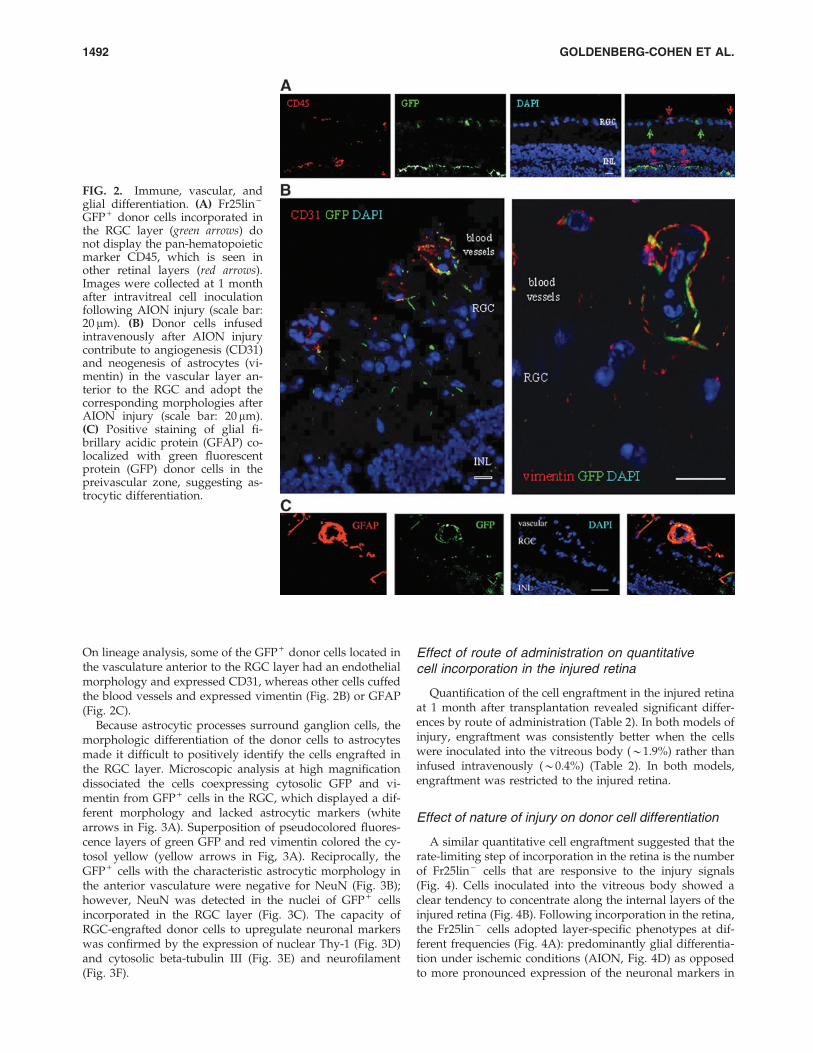
On lineage analysis, some of the GFP + donor cells located inthe vasculature anterior to the RGC layer had an endothelialmorphology and expressed CD31, whereas other cells cuffedthe blood vessels and expressed vimentin (Fig. 2B) or GFAP(Fig. 2C).
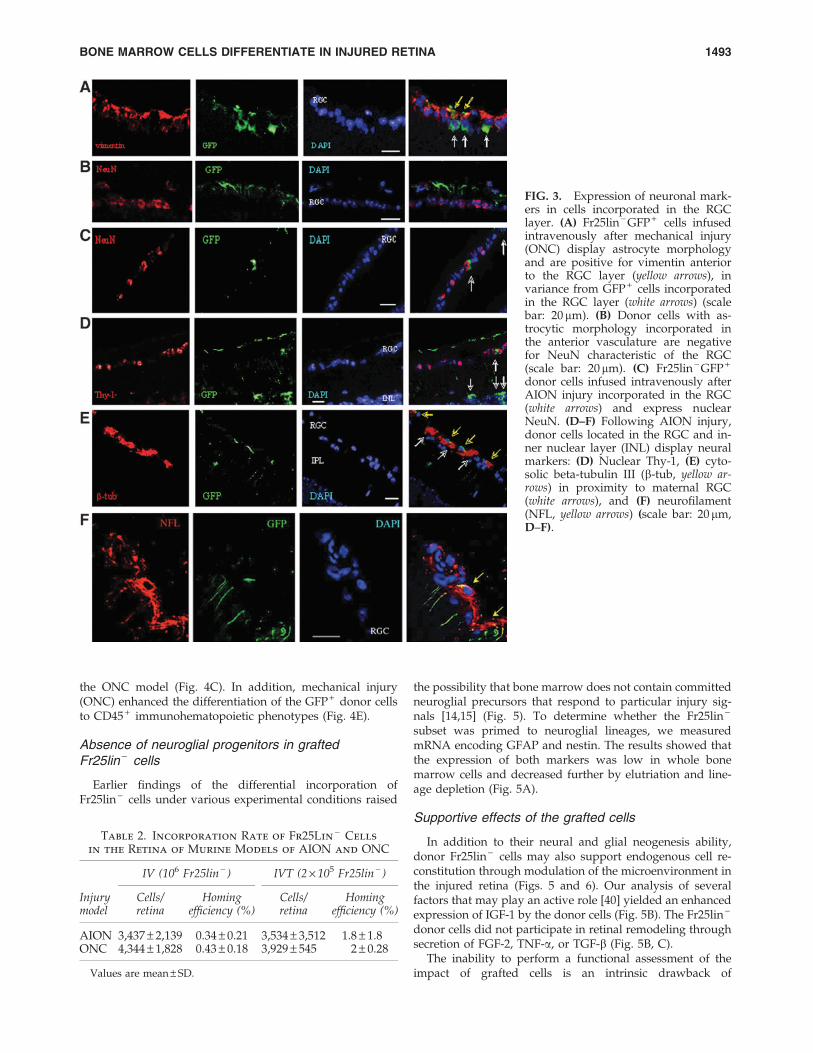
Because astrocytic processes surround ganglion cells, themorphologic differentiation of the donor cells to astrocytesmade it difficult to positively identify the cells engrafted inthe RGC layer. Microscopic analysis at high magnificationdissociated the cells coexpressing cytosolic GFP and vi-mentin from GFP + cells in the RGC, which displayed a dif-ferent morphology and lacked astrocytic markers (whitearrows in Fig. 3A). Superposition of pseudocolored fluores-cence layers of green GFP and red vimentin colored the cy-tosol yellow (yellow arrows in Fig, 3A). Reciprocally, theGFP + cells with the characteristic astrocytic morphology inthe anterior vasculature were negative for NeuN (Fig. 3B);however, NeuN was detected in the nuclei of GFP + cellsincorporated in the RGC layer (Fig. 3C). The capacity ofRGC-engrafted donor cells to upregulate neuronal markerswas confirmed by the expression of nuclear Thy-1 (Fig. 3D)and cytosolic beta-tubulin III (Fig. 3E) and neurofilament(Fig. 3F).
Effect of route of administration on quantitativecell incorporation in the injured retina
Quantification of the cell engraftment in the injured retinaat 1 month after transplantation revealed significant differ-ences by route of administration (Table 2). In both models ofinjury, engraftment was consistently better when the cellswere inoculated into the vitreous body (*1.9%) rather thaninfused intravenously (*0.4%) (Table 2). In both models,engraftment was restricted to the injured retina.
Effect of nature of injury on donor cell differentiation
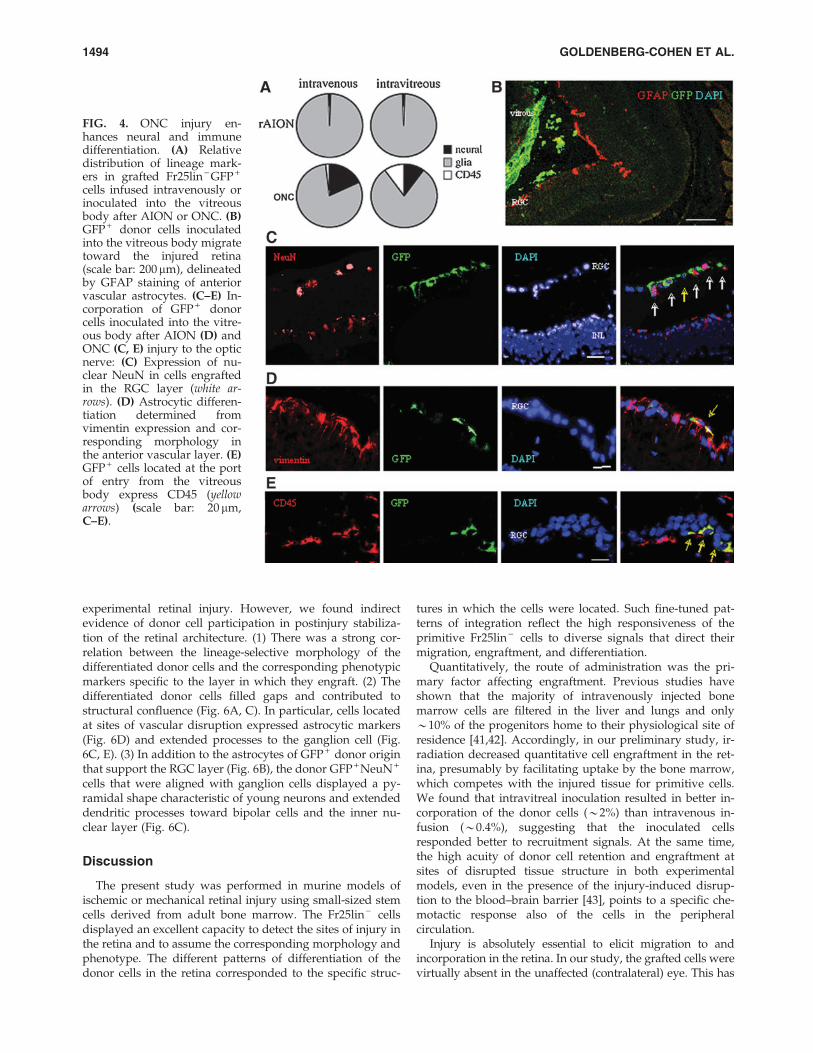
A similar quantitative cell engraftment suggested that therate-limiting step of incorporation in the retina is the numberof Fr25lin - cells that are responsive to the injury signals(Fig. 4). Cells inoculated into the vitreous body showed aclear tendency to concentrate along the internal layers of theinjured retina (Fig. 4B). Following incorporation in the retina,the Fr25lin - cells adopted layer-specific phenotypes at dif-ferent frequencies (Fig. 4A): predominantly glial differentia-tion under ischemic conditions (AION, Fig. 4D) as opposedto more pronounced expression of the neuronal markers in
FIG. 2. Immune, vascular, andglial differentiation. (A) Fr25lin -
GFP + donor cells incorporated inthe RGC layer (green arrows) donot display the pan-hematopoieticmarker CD45, which is seen inother retinal layers (red arrows).Images were collected at 1 monthafter intravitreal cell inoculationfollowing AION injury (scale bar:20 mm). (B) Donor cells infusedintravenously after AION injurycontribute to angiogenesis (CD31)and neogenesis of astrocytes (vi-mentin) in the vascular layer an-terior to the RGC and adopt thecorresponding morphologies afterAION injury (scale bar: 20 mm).(C) Positive staining of glial fi-brillary acidic protein (GFAP) co-localized with green fluorescentprotein (GFP) donor cells in thepreivascular zone, suggesting as-trocytic differentiation.
1492 GOLDENBERG-COHEN ET AL.
the ONC model (Fig. 4C). In addition, mechanical injury(ONC) enhanced the differentiation of the GFP + donor cellsto CD45 + immunohematopoietic phenotypes (Fig. 4E).
Absence of neuroglial progenitors in graftedFr25lin - cells
Earlier findings of the differential incorporation ofFr25lin - cells under various experimental conditions raised
the possibility that bone marrow does not contain committedneuroglial precursors that respond to particular injury sig-nals [14,15] (Fig. 5). To determine whether the Fr25lin -
subset was primed to neuroglial lineages, we measuredmRNA encoding GFAP and nestin. The results showed thatthe expression of both markers was low in whole bonemarrow cells and decreased further by elutriation and line-age depletion (Fig. 5A).
Supportive effects of the grafted cells
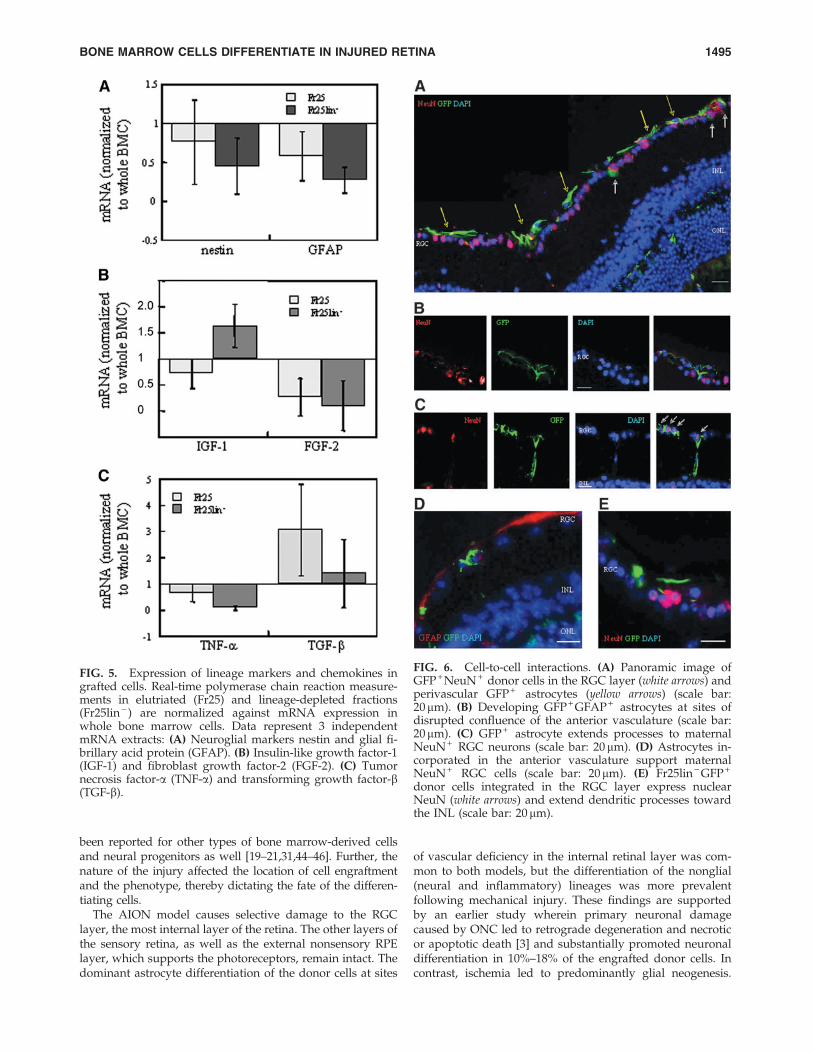
In addition to their neural and glial neogenesis ability,donor Fr25lin - cells may also support endogenous cell re-constitution through modulation of the microenvironment inthe injured retina (Figs. 5 and 6). Our analysis of severalfactors that may play an active role [40] yielded an enhancedexpression of IGF-1 by the donor cells (Fig. 5B). The Fr25lin -
donor cells did not participate in retinal remodeling throughsecretion of FGF-2, TNF-a, or TGF-b (Fig. 5B, C).
The inability to perform a functional assessment of theimpact of grafted cells is an intrinsic drawback of
FIG. 3. Expression of neuronal mark-ers in cells incorporated in the RGClayer. (A) Fr25lin - GFP + cells infusedintravenously after mechanical injury(ONC) display astrocyte morphologyand are positive for vimentin anteriorto the RGC layer (yellow arrows), invariance from GFP + cells incorporatedin the RGC layer (white arrows) (scalebar: 20mm). (B) Donor cells with as-trocytic morphology incorporated inthe anterior vasculature are negativefor NeuN characteristic of the RGC(scale bar: 20mm). (C) Fr25lin - GFP +
donor cells infused intravenously afterAION injury incorporated in the RGC(white arrows) and express nuclearNeuN. (D–F) Following AION injury,donor cells located in the RGC and in-ner nuclear layer (INL) display neuralmarkers: (D) Nuclear Thy-1, (E) cyto-solic beta-tubulin III (b-tub, yellow ar-rows) in proximity to maternal RGC(white arrows), and (F) neurofilament(NFL, yellow arrows) (scale bar: 20mm,D–F).
Table 2. Incorporation Rate of Fr25Lin-
Cells
in the Retina of Murine Models of AION and ONC
IV (106 Fr25lin - ) IVT (2 · 105 Fr25lin - )
Injurymodel
Cells/retina
Homingefficiency (%)
Cells/retina
Homingefficiency (%)
AION 3,437 – 2,139 0.34 – 0.21 3,534 – 3,512 1.8 – 1.8ONC 4,344 – 1,828 0.43 – 0.18 3,929 – 545 2 – 0.28
Values are mean – SD.
BONE MARROW CELLS DIFFERENTIATE IN INJURED RETINA 1493
experimental retinal injury. However, we found indirectevidence of donor cell participation in postinjury stabiliza-tion of the retinal architecture. (1) There was a strong cor-relation between the lineage-selective morphology of thedifferentiated donor cells and the corresponding phenotypicmarkers specific to the layer in which they engraft. (2) Thedifferentiated donor cells filled gaps and contributed tostructural confluence (Fig. 6A, C). In particular, cells locatedat sites of vascular disruption expressed astrocytic markers(Fig. 6D) and extended processes to the ganglion cell (Fig.6C, E). (3) In addition to the astrocytes of GFP + donor originthat support the RGC layer (Fig. 6B), the donor GFP + NeuN +
cells that were aligned with ganglion cells displayed a py-ramidal shape characteristic of young neurons and extendeddendritic processes toward bipolar cells and the inner nu-clear layer (Fig. 6C).
Discussion
The present study was performed in murine models ofischemic or mechanical retinal injury using small-sized stemcells derived from adult bone marrow. The Fr25lin - cellsdisplayed an excellent capacity to detect the sites of injury inthe retina and to assume the corresponding morphology andphenotype. The different patterns of differentiation of thedonor cells in the retina corresponded to the specific struc-
tures in which the cells were located. Such fine-tuned pat-terns of integration reflect the high responsiveness of theprimitive Fr25lin - cells to diverse signals that direct theirmigration, engraftment, and differentiation.
Quantitatively, the route of administration was the pri-mary factor affecting engraftment. Previous studies haveshown that the majority of intravenously injected bonemarrow cells are filtered in the liver and lungs and only*10% of the progenitors home to their physiological site ofresidence [41,42]. Accordingly, in our preliminary study, ir-radiation decreased quantitative cell engraftment in the ret-ina, presumably by facilitating uptake by the bone marrow,which competes with the injured tissue for primitive cells.We found that intravitreal inoculation resulted in better in-corporation of the donor cells (*2%) than intravenous in-fusion (*0.4%), suggesting that the inoculated cellsresponded better to recruitment signals. At the same time,the high acuity of donor cell retention and engraftment atsites of disrupted tissue structure in both experimentalmodels, even in the presence of the injury-induced disrup-tion to the blood–brain barrier [43], points to a specific che-motactic response also of the cells in the peripheralcirculation.
Injury is absolutely essential to elicit migration to andincorporation in the retina. In our study, the grafted cells werevirtually absent in the unaffected (contralateral) eye. This has
FIG. 4. ONC injury en-hances neural and immunedifferentiation. (A) Relativedistribution of lineage mark-ers in grafted Fr25lin - GFP +
cells infused intravenously orinoculated into the vitreousbody after AION or ONC. (B)GFP + donor cells inoculatedinto the vitreous body migratetoward the injured retina(scale bar: 200mm), delineatedby GFAP staining of anteriorvascular astrocytes. (C–E) In-corporation of GFP + donorcells inoculated into the vitre-ous body after AION (D) andONC (C, E) injury to the opticnerve: (C) Expression of nu-clear NeuN in cells engraftedin the RGC layer (white ar-rows). (D) Astrocytic differen-tiation determined fromvimentin expression and cor-responding morphology inthe anterior vascular layer. (E)GFP + cells located at the portof entry from the vitreousbody express CD45 (yellowarrows) (scale bar: 20 mm,C–E).
1494 GOLDENBERG-COHEN ET AL.
been reported for other types of bone marrow-derived cellsand neural progenitors as well [19–21,31,44–46]. Further, thenature of the injury affected the location of cell engraftmentand the phenotype, thereby dictating the fate of the differen-tiating cells.
The AION model causes selective damage to the RGClayer, the most internal layer of the retina. The other layers ofthe sensory retina, as well as the external nonsensory RPElayer, which supports the photoreceptors, remain intact. Thedominant astrocyte differentiation of the donor cells at sites
of vascular deficiency in the internal retinal layer was com-mon to both models, but the differentiation of the nonglial(neural and inflammatory) lineages was more prevalentfollowing mechanical injury. These findings are supportedby an earlier study wherein primary neuronal damagecaused by ONC led to retrograde degeneration and necroticor apoptotic death [3] and substantially promoted neuronaldifferentiation in 10%–18% of the engrafted donor cells. Incontrast, ischemia led to predominantly glial neogenesis.
FIG. 5. Expression of lineage markers and chemokines ingrafted cells. Real-time polymerase chain reaction measure-ments in elutriated (Fr25) and lineage-depleted fractions(Fr25lin - ) are normalized against mRNA expression inwhole bone marrow cells. Data represent 3 independentmRNA extracts: (A) Neuroglial markers nestin and glial fi-brillary acid protein (GFAP). (B) Insulin-like growth factor-1(IGF-1) and fibroblast growth factor-2 (FGF-2). (C) Tumornecrosis factor-a (TNF-a) and transforming growth factor-b(TGF-b).
FIG. 6. Cell-to-cell interactions. (A) Panoramic image ofGFP + NeuN + donor cells in the RGC layer (white arrows) andperivascular GFP + astrocytes (yellow arrows) (scale bar:20mm). (B) Developing GFP + GFAP + astrocytes at sites ofdisrupted confluence of the anterior vasculature (scale bar:20mm). (C) GFP + astrocyte extends processes to maternalNeuN + RGC neurons (scale bar: 20 mm). (D) Astrocytes in-corporated in the anterior vasculature support maternalNeuN + RGC cells (scale bar: 20mm). (E) Fr25lin - GFP +
donor cells integrated in the RGC layer express nuclearNeuN (white arrows) and extend dendritic processes towardthe INL (scale bar: 20 mm).
BONE MARROW CELLS DIFFERENTIATE IN INJURED RETINA 1495

Others observed that in both ischemic and mechanical injurymodels, disruption of the blood–brain barrier abolished theeye’s immune privilege [47], but only mechanical injury wasfavorable to induction of CD45 immunohematopoietic cells.The marked variability in morphology and phenotype in thepresent study, together with the absence of hematopoieticcell engraftment in the absence of conditioning, suggest thatthe engrafted donor cells adopted a lineage-specific differ-entiation in response to local cues in situ. Even if a fewprogenitors reach the bone marrow, functional engraftmentof Fr25lin - cells usually appears later than 1 month post-transplantation [32,33,37], when the present analysis wasperformed.
In a recent study in a retinal laser burn model, Singh et al.[48] delivered Lin-negative bone marrow cells intravenouslyand intravitreously. The intravenous route was found to besuperior in terms of cell mobilization. However, their studydiffered from ours in several important aspects. (1) Singhet al. [48] used a whole bone marrow cell lineage markedwith chemical labeling (carboxyfluorescein diacetate succi-nimydal ester), whereas we used elutriated small fraction(Fr25) lineage-depleted cells of bone marrow extracted fromtransgenic GFP + mice. (2) Singh et al. [48] applied a laserinjury (burn) to the retina, which destroyed both the pho-toreceptors and the RPE, the most external layers of theretina. In contrast, we selectively damaged the RGC by in-ducing AION or ONC, which affected only the internalneuronal layer. The near location of the injured layer andvascular choroids relative to the distance of these layers fromthe vitreous may explain the better transplantation outcomefor the intravenous route in the study by Singh et al. [48]. (3)Singh et al. [48] injected the cells immediately after inductionof injury, whereas we injected them 1 day after injury, duringthe maximal point of damage (based on our preliminarydata). (4) Singh et al. [48] injected significantly fewer and lessselective cells, 50,000 and 100,000 cells intravitreously, withno incorporation at the higher dose because of clumping.Intravenously, they injected 100,000 cells compared with1 · 106 in our study, with better results. This finding maybe related to the different types of injury. Overall, thedifferent layers of the eye injured may account for thediscrepant results.
The bone marrow is a heterogeneous compartment thathosts multiple cell types and precursors. The manner inwhich it contributes to revascularization is still controversial.Some researchers attributed this contribution to the he-mangioblasts and endothelial progenitors committed toangiogenic differentiation that reside in the bone marrow[16–21,49,50]. This led to the assumption that committed retinalprogenitors from the marrow compartment can be recruitedto replenish the retinal pigment epithelium layer [14,15].Alternatively, the adult bone marrow may contain subsets ofpluripotent cells that can respond to multiple signals andadopt versatile differentiation traits. In the present study, inline with previous reports that elutriated Fr25 cells do notexpress the hemangioblast and hematopoietic stem cellmarkers CD31, CD34, SCA-1, and c-kit [51–53], we found noevidence of nestin or GFAP expression in these cells. Thus, itis unlikely that vascular, glial, and neural conversion can beattributed to tissue progenitors sequestered in the bonemarrow, even if the commitment of precursors is limited topriming at the transcriptional level [54,55]. Nestin is a pro-
miscuous marker of ectodermal and gastrointestinal pre-cursors and mature lineages [56,57]. However, it frequentlyaccompanies GFAP, an early marker of dual neural and glialdifferentiation of astrocytes and neural progenitors [58,59],during astrocyte differentiation [60]. Further, we recentlyreported that endocrine differentiation of Fr25lin - cells is notrestricted to the expression of progenitor markers SCA-1, c-kit, and Thy1 [38]. This finding is consistent with the minoradvantage of prospectively isolating SCA-1 + and Thy-1 + inbone marrow cells for the in vitro generation of nestin-positive neurospheres [61]. Although the neuroglial and he-matopoietic markers are considered to be involved in neuraldifferentiation of mesenchymal stroma cells [62,63], it re-mains to be determined whether the small-sized subset ofelutriated cells contains sufficient numbers of mesenchymalstromal cells to account for the observed engraftment in theinjured retina. Nevertheless, evidence points to a differentialcapacity of various bone marrow-derived cell types to repairthe retinal pigment epithelium, reduce photoreceptor de-generation, induce angiogenesis under various experimentalconditions, and mediate choroidal revascularization follow-ing laser-induced injury affecting the outer retinal layers andBruch’s membrane [14–22,31,46,49,50].
On the basis of the present findings, we suggest that aputative subset of pluripotent cells, such as the one used inthis study, migrates to various layers of the retina and, inresponse to specific local signals of injury, differentiates intoendothelium (CD31), immune cells (CD45), astrocytes (GFAPor vimentin), or neural cells (NeuN, Thy-1, beta-tubulin III).This interpretation complements earlier findings that thesecells regenerate the endocrine pancreas, liver, and multipleepithelial tissues [34–36] and that the various subsets of adultbone marrow cells adopt neural and glial phenotypes in vivo[24–27]. However, there is no evidence that tissue-committedprogenitors residing in the bone marrow navigate to specificsites and develop according to an imprinted (genetic andepigenetic) program. True differentiation is exemplified bythe CD45-negative phenotype of the retina-engrafted cells andthe normal diploid genotype. Rare events of fusion betweenstem cells or the myeloid progeny of hematopoietic progeni-tors and injured tissue might result in lateral transfer of spe-cific markers without true differentiation [64]. An overzealousspeculation that fusion in culture negates developmentalplasticity [65] of bone marrow cells that differentiate into gliaand neurons [24–27] is not substantiated by careful genotypicexamination of diploid phenotypes of neural progenitorsconverting into endothelium [66] and of Fr25lin- cells con-verting into hepatocytes [34] and insulin-producing cells [35].The capacity of myeloid lineages to fuse, phagocytose, andengulf apoptotic and necrotic cells is irrelevant to the observedphenomena in the absence of CD45 expression in the hybrids[67]. The data reported here are likely to be an underestimateof cell engraftment in the retina [38], considering that only*50% of grafted cells express GFP [68] and the expression ofthis reporter protein is markedly downregulated duringneuronal differentiation [69].
Locating donor cells, especially long after transplantation,is technically difficult. Although the retina is considered animmunologically privileged site [70], researchers have re-ported GFP rejection in rabbits early after viral transfectionof retinal cells [71] or after subretinal administration of len-tiviral vectors [72]. The rejection was a humoral immune
1496 GOLDENBERG-COHEN ET AL.

response and not a cellular reaction [73,74]. There was norejection when the vector was injected without any trans-gene, indicating that it was not due to the viral vector [73]. Inthe present study, the donor cells were derived from GFP-transgeneic mice and were not virally transfected. We didnot find signs of GFP rejection in the eye, in accordance withnumerous other investigators who used GFP cells fromtransgenic GFP donors [74–77]. The cells were injected eitherintravenously or intravitreously, but not into the subretinalspace, and they survived the circulation and reached theinjured retina to incorporate into it and differentiate. The lackof rejection may be explained by species differences, thesmall number of cells injected, or the weaker expression ofthe transgenic proteins. To overcome the labeling difficultiesin the above techniques, we also identified donor cells byother methods: 42.1 and 42.2 and FISH for X-Y mismatch.
One of the major drawbacks of neural models of regen-eration is the difficult functional assessment of the organ.Unlike models of diabetes and hepatotoxicity [34,35], we donot know to what extent the observed contribution of thegrafted cells improves organ function. Preliminary studiesindicated better axonal transport following transplantation,but it is unlikely that axonal processes of donor cellsadopting neural phenotypes in the RGC were responsible forsuch distant transport. Rather, it may primarily rely on col-lateral support of endogenous repair through vascularizationand secretion of neuroprotective agents [40]. We assessed thepossibility that the grafted cells further contribute to tissueremodeling after injury by local secretion of neuroprotectiveagents. We detected increased levels of mRNA encodingIGF-1 [78,79], but no increased expression of TGF-b [80,81],FGF-2 [82–84], or the neurotrophic factor TNF-a [85–87].
In summary, a small-sized subset of adult bone marrowcells contributes to retinal remodeling after ischemic andmechanical injury by both direct contribution to cellular re-generation and support of endogenous repair. The majordirect cellular contribution is gliosis, which is the dominantinductive process within the physiological mode of repair ofthe retina as a neural tissue. Precursors of the various line-ages are not present in the grafted cells, which developphenotypes corresponding to the site of engraftment. Thecorrelation among stem cell location, morphology, pheno-type, and cooperation with endogenous cells in stabilizationof retinal architecture is strong evidence that those withflexible developmental potential receive site-specific induc-tive signals. A second mechanism is support of retinal re-modeling through angiogenesis in the immunomodulationand secretion of IGF-1. Monitoring of retinal function willdetermine the significance and potential implementation ofadult autologous bone marrow-derived cells in ischemic eyeinjury.
Acknowledgments
The authors thank Dr. Saul Sharkis and Dr. Michael Col-lector for the outstanding support, discussion, and concep-tual contribution to this study. Funding was provided bygenerous grants from the Leah and Edward M. Frankel Trustfor bone marrow transplantation, the Zanvyl and IsabelleKrieger Fund, Israel Academy of Sciences and Humanities(ISF, NGC, 1371/08), the Eldor-Metzner Clinician ScientistAward, Chief Scientist, Israel Ministry of Health (NGC, 3-
3741), and the Lirot Foundation (NGC, 3-4538) and a pilotgrant from the North American Neuro-Ophthalmology So-ciety and by Tel Aviv University: The Adams Super Centerfor Brain Research (2004–2005, 2005–2007, NGC); the Clairand Ameddee Maratier Fund to prevent blindness (2004–2005; 2010–2011; NGC, 2008 BRAL); Walter Friendliest andHerman Shudder Fund (NGC 2006–2008); The Lions (BRAL2010) Mazritzky Fund (TS 2010).
Author Disclosure Statement
The results were presented in part at the Association forResearch in Vision and Ophthalmology, Fort Lauderdale, FL(ARVO), 2006, 2007; Meeting of the Israel Society for Eyeand Vision Research, Neve Ilan, Israel (ISVER), 2006, 2007,2008, 2009, 2010; Meeting of the North American Neuro-Ophthalmology Society (NANOS), Tucson, AZ, 2006, 2010;Meeting of the International Society for Stem Cell Research(ISSCR), Philadelphia, PA, 2008; the Second InternationalStem Cell Meeting, 2008, 2009, Tel Aviv, Israel (BIOMED);Tel Aviv Research Day, Tel Aviv, 2008, 2010; Rabin MedicalCenter Research Day 2006, Petach Tikva, Isreal; the NinthMichaelson Symposium 2007, Baltimore, MD; Meeting of theMacular Society, 2007, London, United Kingdom; the SecondYoung Investigators Stem Cell Meeting 2009, Bar Ilan Uni-versity, Ramat Gan, Israel; and ILANIT/FISEB Federation ofthe Israel Societies for Experimental Biology Meeting, Eilat,Isarel, February 2011.
References
1. Levkovitch-Verbin H. (2004). Animal models of optic nervediseases. Eye (Lond) 18:1066–1074.
2. Goldenberg-Cohen N, Y Guo, F Margolis, Y Cohen, NRMiller and SL Bernstein. (2005). Oligodendrocyte dysfunc-tion after induction of experimental anterior optic nerve is-chemia. Invest Ophthalmol Vis Sci 46:2716–2725.
3. Dratviman-Storobinsky O, M Hasanreisoglu, D Offen, YBarhum, D Weinberger and N Goldenberg-Cohen. (2008).Progressive damage along the optic nerve following induc-tion of crush injury or rodent anterior ischemic optic neu-ropathy in transgenic mice. Mol Vis 14:2171–2179.
4. Bonfanti L, E Strettoi, S Chierzi, MC Cenni, XH Liu, J-CMartinou, L Maffei and SA Rabacchi. (1996). Protection ofretinal ganglion cells from natural and axotomy-induced celldeath in neonatal transgenic mice overexpressing bcl-2. JNeurosci 16:4186–4194.
5. Selles-Navarro I, MP Villegas-Perez, M Salvador-Silva, JMRuiz-Gomez and M Vidal-Sanz. (1996). Retinal ganglion celldeath after different transient periods of pressure-inducedischemia and survival intervals. A quantitative in vivostudy. Invest Ophthalmol Vis Sci 37:2002–2014.
6. Chaudhary P, F Ahmed, P Quebada and SC Sharma. (1999).Caspase inhibitors block the retinal ganglion cell death fol-lowing optic nerve transection. Brain Res Mol Brain Res67:36–45.
7. Bernstein SL, Y Guo, SE Kelman, RW Flower and MAJohnson. (2003). Functional and cellular responses in a novelrodent model of anterior ischemic optic neuropathy. InvestOphthalmol Vis Sci 44:4153–4162.
8. Bernstein SL, Y Guo, BJ Slater, A Puche and SE Kelman.(2007). Neuron stress and loss following rodent anterior is-chemic optic neuropathy in double-reporter transgenic mice.Invest Ophthalmol Vis Sci 48:2304–2310.
BONE MARROW CELLS DIFFERENTIATE IN INJURED RETINA 1497

9. Chen CS, MA Johnson, RA Flower, BJ Slater, NR Miller andSL Bernstein. (2008). A primate model of nonarteritic ante-rior ischemic optic neuropathy. Invest Ophthalmol Vis Sci49:2985–2992.
10. Slater BJ, Z Mehrabian, Y Guo, A Hunter and SL Bernstein.(2008). Rodent anterior ischemic optic neuropathy (rAION)induces regional retinal ganglion cell apoptosis with a uniquetemporal pattern. Invest Ophthalmol Vis Sci 49:3671–3676.
11. Zhang C, Y Guo, NR Miller and SL Bernstein. (2009). Opticnerve infarction and post-ischemic inflammation in the ro-dent model of anterior ischemic optic neuropathy (rAION).Brain Res 1264:67–75.
12. Banin E, A Obolensky, M Idelson, I Hemo, E Reinhardtz, EPikarsky, T Ben-Hur and B Reubinoff. (2006). Retinal in-corporation and differentiation of neural precursors derivedfrom human embryonic stem cells. Stem Cells 24:246–257.
13. Parameswaran S, S Balasubramanian, N Babai, F Qiu, JDEudy, WB Thoreson and I Ahmad. (2010). Induced plurip-otent stem cells generate both retinal ganglion cells andphotoreceptors: therapeutic implications in degenerativechanges in glaucoma and age-related macular degeneration.Stem Cells 28:695–703.
14. Li Y, RG Reca, P Atmaca-Sonmez, MZ Ratajczak, ST Ildstad,HJ Kaplan and V Enzmann. (2006). Retinal pigment epi-thelium damage enhances expression of chemoattractantsand migration of bone marrow-derived stem cells. InvestOphthalmol Vis Sci 47:1646–1652.
15. Li Y, P Atmaca-Sonmez, CL Schanie, ST Ildstad, HJ Kaplanand V Enzmann. (2007). Endogenous bone marrow derivedcells express retinal pigment epithelium cell markers andmigrate to focal areas of RPE damage. Invest OphthalmolVis Sci 48:4321–4327.
16. Friedlander M, MI Dorrell, MR Ritter, V Marchetti, SKMoreno, M El-Kalay, AC Bird, E Banin and E Aguilar.(2007). Progenitor cells and retinal angiogenesis. Angiogen-esis 10:89–101.
17. Otani A and M Friedlander. (2005). Retinal vascular regen-eration. Semin Ophthalmol 20:43–50.
18. Lee IG, SL Chae and JC Kim. (2006). Involvement of circu-lating endothelial progenitor cells and vasculogenic factorsin the pathogenesis of diabetic retinopathy. Eye (Lond)20:546–552.
19. Grant MB, WS May, S Caballero, GA Brown, SM Guthrie,RN Mames, BJ Byrne, T Vaught, PE Spoerri, AB Peck andEW Scott. (2002). Adult hematopoietic stem cells providefunctional hemangioblast activity during retinal neovascu-larization. Nat Med 8:607–612.
20. Harris JR, R Fisher, M Jorgensen, S Kaushal and EW Scott.(2009). CD133 progenitor cells from the bone marrow con-tribute to retinal pigment epithelium repair. Stem Cells27:457–466.
21. Sengupta N, S Caballero, SM Sullivan, LJ Chang, A Afzal, SLi Calzi, JL Kielczewski, S Prabarakan, EA Ellis, et al. (2009).Regulation of adult hematopoietic stem cells fate for en-hanced tissue-specific repair. Mol Ther 17:1594–1604.
22. Atmaca-Sonmez P, Y Li, Y Yamauchi, CL Schanie, ST Ild-stad, HJ Kaplan and V Enzmann. (2006). Systemicallytransferred hematopoietic stem cells home to the subretinalspace and express RPE-65 in a mouse model of retinal pig-ment epithelium damage. Exp Eye Res 83:1295–1302.
23. Minamino K, Y Adachi, H Yamada, A Higuchi, Y Suzuki, MIwasaki, K Nakano, Y Koike, H Mukaide, et al. (2005). Long-term survival of bone marrow-derived retinal nerve cells inthe retina. Neuroreport 16:1255–1259.
24. Eglitis MA and E Mezey. (1997). Hematopoietic cells dif-ferentiate into both microglia and macroglia in the brains ofadult mice. Proc Natl Acad Sci U S A 94:4080–4085.
25. Eglitis MA, D Dawson, KW Park and MM Mouradian.(1999). Targeting of marrow-derived astrocytes to the is-chemic brain. Neuroreport 10:1289–1292.
26. Brazelton TR, FM Rossi, GI Keshet and HM Blau. (2000).From marrow to brain: expression of neuronal phenotypesin adult mice. Science 290:1775–1779.
27. Mezey E, KJ Chandross, G Harta, RA Maki and SRMcKercher. (2000). Turning blood into brain: cells bearingneuronal antigens generated in vivo from bone marrow.Science 290:1779–1782.
28. Yodoi Y, M Sasahara, T Kameda, N Yoshimura and A Otani.(2007). Circulating hematopoietic stem cells in patients withneovascular age-related macular degeneration. Invest Oph-thalmol Vis Sci 48:5464–5472.
29. Weimann JM, CA Charlton, TR Brazelton, RC Hackman andHM Blau. (2003). Contribution of transplanted bone marrowcells to Purkinje neurons in human adult brains. Proc NatlAcad Sci U S A 100:2088–2093.
30. Kicic A, AC Shanley, CM Hall and PE Rakoczy. (2003).Marrow stromal cells (MSC): a species comparison. Adv ExpMed Biol 533:407–414.
31. Inoue Y, A Iriyama, S Ueno, H Takahashi, M Kondo, YTamaki, M Araie and Y Yanagi. (2007). Subretinal trans-plantation of bone marrow mesenchymal stem cells delaysretinal degeneration in the RCS rat model of retinal degen-eration. Exp Eye Res 85:234–241.
32. Jones RJ, P Celano, SJ Sharkis and LL Sensenbrenner. (1989).Two phases of engraftment established by serial bone mar-row transplantation in mice. Blood 73:397–401.
33. Lanzkron SM, MI Collector and SJ Sharkis. (1999). Hema-topoietic stem cell tracking in vivo: a comparison of short-term and long-term repopulating cells. Blood 93:1916–1921.
34. Jang YY, MI Collector, SB Baylin, AM Diehl and SJ Sharkis.(2004). Hematopoietic stem cells convert into liver cellswithin days without fusion. Nat Cell Biol 6:532–539.
35. Iskovich S, N Goldenberg-Cohen, J Stein, I Yaniv, I Fabian andN Askenasy. (2011). Elutriated stem cells derived from theadult bone marrow differentiate into insulin producing cellsin vivo and reverse chemical diabetes. Stem Cell Dev 2011 Apr4. [Epub ahead of print]; DOI: 10.1089/scd.2011.0057.
36. Krause DS, ND Theise, MI Collector, O Henegariu, SHwang, R Gardner, S Neutzel and SJ Sharkis. (2001). Multi-organ, multi-lineage engraftment by a single bone marrow-derived stem cell. Cell 105:369–377.
37. Pearl-Yafe M, J Stein, ES Yolcu, DL Farkas, H Shirwan, IYaniv and N Askenasy. (2007). Fas transduces dual apo-ptotic and trophic signals in hematopoietic progenitors.Stem Cells 25:3194–3203.
38. Iskovich S, N Goldenberg-Cohen, J Stein, I Yaniv, DL Farkasand N Askenasy. (2011). b-Cell neogenesis: experimentalconsiderations in adult stem cell differentiation. Stem CellsDev 20:569–582.
39. Hasanreisoglu M, S Dadon, BR Avraham, Y Cohen, DWeinberger and N Goldenberg-Cohen. (2009). Neuropro-tective effect of brimonidine in a mouse model of ischemicoptic neuropathy. Clin Exp Ophthalmol 37:718–729.
40. Mondal D, L Pradhan and VF LaRussa. (2004). Signaltransduction pathways involved in the lineage-differentiationof NSCs: can the knowledge gained from blood be used inthe brain? Cancer Invest 22:925–943.
1498 GOLDENBERG-COHEN ET AL.

41. Askenasy N and DL Farkas. (2003). In vivo imaging studiesof the effect of recipient conditioning, donor cell phenotypeand antigen disparity on homing of haematopoietic cells tothe bone marrow. Br J Haematol 120:505–515.
42. Yaniv I, J Stein, DL Farkas and N Askenasy. (2006). The taleof early hematopoietic cell seeding in the bone marrowniche. Stem Cells Dev 15:4–16.
43. Johnson TV, ND Bull and KR Martin. (2010). Identificationof barriers to retinal engraftment of transplanted stem cells.Invest Ophthalmol Vis Sci 51:960–970.
44. Nishida A, M Takahashi, H Tanihara, I Nakano, JB Taka-hashi, A Mizoguchi, C Ide and Y Honda. (2000). In-corporation and differentiation of hippocampus-derivedneural stem cells transplanted in injured adult rat retina.Invest Ophthalmol Vis Sci 41:4268–4274.
45. Young MJ, J Ray, SJ Whiteley, H Klassen and FH Gage.(2000). Neuronal differentiation and morphological integra-tion of hippocampal progenitor cells transplanted to theretina of immature and mature dystrophic rats. Mol CellNeurosci 16:197–205.
46. Meyer JS, ML Katz and MD Kirk. (2005). Stem cells for ret-inal degenerative disorders. Ann N Y Acad Sci 1049:135–145.
47. Taylor AW. (2002). Neuroimmunomodulation and immuneprivilege: the role of neuropeptides in ocular immunosup-pression. Neuroimmunomodulation 10:189–198.
48. Singh T, S Prabhakar, A Gupta and V Anand. (2011). Re-cruitment of stem cells into the injured retina after laser in-jury. Stem Cell Dev [Epub ahead of print]; DOI: 10.1089/scd.2011.0002.
49. Otani A, K Kinder, K Ewalt, FJ Otero, P Schimmel and MFriedlander. (2002). Bone marrow-derived stem cells targetretinal astrocytes and can promote or inhibit retinal angio-genesis. Nat Med 8:1004–1010.
50. Otani A, MI Dorrell, K Kinder, SK Moreno, S Nusinowitz, EBanin, J Heckenlively and M Friedlander. (2004). Rescue ofretinal degeneration by intravitreally injected adult bonemarrow-derived lineage-negative hematopoietic stem cells. JClin Invest 114:765–774.
51. Ortiz M, JW Wine, N Lohrey, FW Ruscetti, SE Spence and JRKeller. (1999). Functional characterization of a novel hemato-poietic stem cell and its place in the c-Kit maturation pathwayin bone marrow cell development. Immunity 10:173–182.
52. Sharkis SJ, MI Collector, JP Barber, MS Vala and RJ Jones.(1997). Phenotypic and functional characterization of thehematopoietic stem cell. Stem Cells 15(Suppl 1):41–45.
53. Sharkis SJ, S Neutzel and MI Collector. (2001). Phenotypeand function of hematopoietic stem cells. Ann N Y Acad Sci938:191–195.
54. Orkin SH and LI Zon. (2002). Hematopoiesis and stem cells:plasticity versus developmental heterogeneity. Nat Im-munol 3:323–328.
55. Askenasy N, I Yaniv, J Stein and SJ Sharkis. (2006). Ourperception of developmental plasticity: esse est percipi (to beis to be perceived)? Curr Stem Cell Res Ther 1:85–94.
56. Iskovich S, A Kaminitz, M Pearl-Yafe, K Mizrahi, J Stein, IYaniv and N Askenasy. (2007). Participation of adult bonemarrow-derived stem cells in pancreatic regeneration: neo-genesis versus endogenesis. Curr Stem Cell Res Ther 2:272–279.
57. Qiu G, MJ Seiler, BB Thomas, K Wu, M Radosevich and SRSadda. (2007). Revisiting nestin expression in retinal pro-genitor cells in vitro and after transplantation in vivo. ExpEye Res 84:1047–1059.
58. Laywell E, P Rakic, VG Kukekov, EC Holland and DASteindler. (2000). Identification of a multipotent astrocytic
stem cell in the immature and adult mouse brain. Proc NatlAcad Sci U S A 97:13883–13888.
59. Namba T, H Mochizuki, M Onodera, Y Mizuno, H Namikiand T Seki. (2005). The fate of neural progenitor cells ex-pressing astrocytic and radial glial markers in the postnatalrat dentate gyrus. Eur J Neurosci 22:1928–1941.
60. Sergent-Tanguy S, DC Michel, I Neveu and P Naveilhan.(2006). Long-lasting coexpression of nestin and glial fi-brillary acidic protein in primary cultures of astroglial cellswith a major participation of nestin( + )/GFAP(-) cells in cellproliferation. J Neurosci Res 83:1515–1524.
61. Locatelli F, S Corti, C Donadoni, M Guglieri, F Capra, SStrazzer, S Salani, R Del Bo, F Fortunato, A Bordoni and GPComi. (2003). Neuronal differentiation of murine bonemarrow Thy-1- and Sca-1-positive cells. J Hematother StemCell Res 12:727–734.
62. Wislet-Gendebien S, P Leprince, G Moonen and B Rogister.(2003). Regulation of neural markers nestin and GFAP ex-pression by cultivated bone marrow stromal cells. J Cell Sci116:3295–3302.
63. Mizobe T, K Kidoguchi, M Tamaki, T Sasayama, T Kondohand E Kohmura. (2006). Sca-1 and Thy-1 accelerate neuron-like differentiation in bone marrow stromal cells. Kobe JMed Sci 52:119–125.
64. Wurmser AE and FH Gage. (2002). No stem cells: cell fusioncauses confusion. Nature 416:485–487.
65. Terada N, T Hamazaki, M Oka, M Hoki, DM Mastalerz, YNakano, EM Meyer, L Morel, BE Petersen and EW Scott.(2002). Bone marrow cells adopt the phenotype of other cellsby spontaneous cell fusion. Nature 416:542–545.
66. Wurmser AE, K Nakashima, RG Summers, N Toni, KAD’Amour, DC Lie and FH Gage. (2004). Cell fusion-independent differentiation of neural stem cells to the en-dothelial lineage. Nature 430:350–356.
67. Camargo FD, M Finegold and MA Goodell. (2004). Hema-topoietic myelomonocytic cells are the major source of he-patocyte fusion partners. J Clin Invest 113:1266–1270.
68. Biankin SA, MI Collector, AV Biankin, LJ Brown, W Klee-berger, WL Devereux, CA Zahnow, SB Baylin, DN Watkins,SJ Sharkis and SD Leach. (2007). A histological survey ofgreen fluorescent protein expression in ‘green’ mice: impli-cations for stem cell research. Pathology 39:247–251.
69. Vroemen M, N Weidner and A Blesch. (2005). Loss of geneexpression in lentivirus- and retrovirus-transduced neuralprogenitor cells is correlated to migration and differentiationin the adult spinal cord. Exp Neurol 195:127–139.
70. Gregerson DS and J Yang. (2003). CD45-positive cells of theretina and their responsiveness to in vivo and in vitrotreatment with IFN-gamma or anti-CD40. Invest Ophthal-mol Vis Sci 44:3083–3093.
71. Doi K, J Kong, J Hargitai, SP Goff and P Gouras. (2004).Transient immunosuppression stops rejection of virus-transduced enhanced green fluorescent protein in rabbitretina. J Virol 78:11327–11333.
72. Auricchio A, G Kobinger, V Anand, M Hildinger, EO’Connor, AM Maguire, JM Wilson and J Bennett. (2001).Exchange of surface proteins impacts on viral vector cellularspecificity and transduction characteristics: the retina as amodel. Hum Mol Genet 10:3075–3081.
73. Andersson G, M Denaro, K Johnson, P Morgan, A Sullivan,S Houser, C Patience, ME White-Scharf and JD Down.(2003). Engraftment of retroviral EGFP-transduced bonemarrow in mice prevents rejection of EGFP-transgenic skingrafts. Mol Ther 8:385–391.
BONE MARROW CELLS DIFFERENTIATE IN INJURED RETINA 1499

74. Takahashi M, H Miyoshi, IM Verma and FH Gage. (1999).Rescue from photoreceptor degeneration in the rd mouse byhuman immunodeficiency virus vector-mediated genetransfer. J Virol 73:7812–7816.
75. Bennett J, AM Maguire, AV Cideciyan, M Schnell, E Glover,V Anand, TS Aleman, N Chirmule, AR Gupta, et al. (1999).Stable transgene expression in rod photoreceptors after re-combinant adeno-associated virus-mediated gene transfer tomonkey retina. Proc Natl Acad Sci U S A 96:9920–9925.
76. Miyoshi H, M Takahashi, FH Gage, IM Verma. (1997). Stableand efficient gene transfer into the retina using an HIV-basedlentiviral vector. Proc Natl Acad Sci U S A 94:10319–10323.
77. Surace EM, A Auricchio, SJ Reich, T Rex, E Glover, S Pineles,W Tang, E O’Connor, A Lyubarsky, et al. (2003). Delivery ofadeno-associated virus vectors to the fetal retina: impact ofviral capsid proteins on retinal neuronal progenitor trans-duction. J Virol 77:7957–7963.
78. Araki M, H Suzuki and P Layer. (2007). Differential en-hancement of neural and photoreceptor cell differentiationof cultured pineal cells by FGF-1, IGF-1, and EGF. DevNeurobiol 67:1641–1654.
79. van Wijngaarden P, HM Brereton, IL Gibbins, DJ Coster andKA Williams. (2007). Kinetics of strain-dependent differen-tial gene expression in oxygen-induced retinopathy in therat. Exp Eye Res 2007;85:508–517.
80. Misumi S, TS Kim, CG Jung, T Masuda, S Urakawa, Y Isobe,F Furuyama, H Nishino and H Hida. (2008). Enhancedneurogenesis from neural progenitor cells with G1/S-phasecell cycle arrest is mediated by transforming growth factorbeta1. Eur J Neurosci 28:1049–1059.
81. Park SM, JS Jung, MS Jang, KS Kang and SK Kang. (2008).Transforming growth factor-beta1 regulates the fate of cul-tured spinal cord-derived neural progenitor cells. Cell Prolif41:248–264.
82. Li Q and D Stephenson. (2002). Postischemic administrationof basic fibroblast growth factor improves sensorimotorfunction and reduces infarct size following permanent focalcerebral ischemia in the rat. Exp Neurol 177:531–537.
83. Sapieha PS, M Peltier, KG Rendahl, WC Manning and A DiPolo. (2003). Fibroblast growth factor-2 gene delivery stim-
ulates axon growth by adult retinal ganglion cells after acuteoptic nerve injury. Mol Cell Neurosci 24:656–672.
84. Zaverucha-do-Valle C, F Gubert, M Bargas-Rega, JL Coronel,LA Mesentier-Louro, A Mencalha, E Abdelhay, MF Santiagoand R Mendez-Otero. (2011). Bone-marrow mononuclear cellsincrease retinal ganglion-cell survival and axon regenerationin the adult rat. Cell Transplant 20:391–406.
85. Fontaine V, S Mohand-Said, N Hanoteau, C Fuchs, K Pfi-zenmaier and U Eisel. (2002). Neurodegenerative and neu-roprotective effects of tumor necrosis factor (TNF) in retinalischemia: opposite roles of TNF receptor 1 and TNF receptor2. J Neurosci 22:RC216.
86. Bernardino L, F Agasse, B Silva, R Ferreira, S Grade andJO Malva. (2008). Tumor necrosis factor-alpha modulatessurvival, proliferation, and neuronal differentiation inneonatal subventricular zone cell cultures. Stem Cells 26:2361–2371.
87. Taoufik E, E Petit, D Divoux, V Tseveleki, M Mengozzi,ML Roberts, S Valable, P Ghezzi, J Quackenbush, et al.(2008). TNF receptor I sensitizes neurons to erythropoietin-and VEGF-mediated neuroprotection after ischemic andexcitotoxic injury. Proc Natl Acad Sci U S A 105:6185–6190.
Address correspondence to:Dr. Nitza Goldenberg-Cohen
The Krieger Eye Research LaboratoryDepartment Ophthalmology, Pediatric Unit
Schneider Children’s Medical Center of IsraelPetach Tikva 49202
Israel
E-mail: [email protected]
Received for publication July 11, 2011Accepted after revision September 9, 2011
Prepublished on Liebert Instant Online September 9, 2011
1500 GOLDENBERG-COHEN ET AL.





























