Embed Size (px)
Citation preview

Primate Biogeography, Diversity, Taxonomy and Conservation of the
Coastal Forests of Kenya
Yvonne A. de Jong Thomas M. Butynski
Eastern Africa
Primate Diversity and Conservation Program
Report to the Critical Ecosystem Partnership Fund March, 2009

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 2
Primate Biogeography, Diversity, Taxonomy and Conservation of the Coastal Forests of Kenya. Yvonne A. de Jong & Thomas M. Butynski Eastern Africa Primate Diversity and Conservation Program, Nanyuki, Kenya. Report to the Critical Ecosystem Partnership Fund March, 2009.
Yvonne A. de Jong & Thomas M. Butynski Eastern Africa Primate Diversity and Conservation Program P.O. Box 149, 10400 Nanyuki, Kenya [email protected] & tbutynski&aol.com www.wildsolutions.nl
Cover photos Left: Adult male Chlorocebus pygerythrus hilgerti, west of Malindi, Kenya. Right: Galagoides cocos, Manda Island, Kenya. Citation: De Jong, Y.A. & Butynski, T.M. 2009. Primate Biogeography, Diversity, Taxonomy and Conservation of the Coastal Forests of Kenya. Report to the Critical Ecosystem Partnership Fund. Eastern Africa Primate Diversity and Conservation Program, Nanyuki, Kenya.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 3
CONTENTS
Executive summary 8 1. Introduction 11 2. Study area 13 3. Methods 15 3.1 Preparations 15 3.2 Field surveys 15 3.2.1 Diurnal primate surveys 15
3.2.2 Nocturnal primate surveys 18 3.2.3 Nocturnal listening survey 18
3.3 Data analysis 19 3.3.1 Galago vocalization analysis 19 3.3.2 Distribution mapping 19
3.4 Primate Photographic Maps 19 3.5 Definitions 21
4. Results 22 4.1 Survey site details, primate encounters and conservation 23
4.1.1 North Coast of Kenya 23 A. Lamu Archipelago 23
B. Kipini Conservancy and Witu Forest Reserve 25 C. Tana River Primate National Reserve 28
4.1.2 South Coast of Kenya 30 A. Kaya Rabai (Chijembeni), Mazeras 30 B. Diani 31 C. Mrima Hill Forest 32 D. Kaya Sega 33
4.1.3 Outside the coastal forests of Kenya 34 A. Mwea National Reserve 34 B. Kiboko Camp, Makindu 35 C. Tsavo West National Park 35 D. Tsavo East National Park 36 E. North Kilimangodo 37
4.2 Primates species of the coastal forests of Kenya 38 4.2.1 Otolemur garnettii lasiotis 38 4.2.2 Galago senegalensis braccatus 43 4.2.3 Galagoides cocos 46 4.2.4 Cercopithecus mitis albogularis 50 4.2.5 Cercopithecus mitis albotorquatus 57 4.2.6 Chlorocebus pygerythrus hilgerti 63 4.2.7 Chlorocebus pygerythrus excubitor 68 4.2.8 Papio cynocephalus ibeanus 71 4.2.9 Colobus angolensis palliates 77 4.2.10 Procolobus rufomitratus rufomitratus 80 4.2.11 Cercocebus galeritus 82 4.2.12 Hybrid, Chlorocebus pygerythrus hilgerti x
Cercopithecus mitis albogularis 84
5. Discussion 87 Acknowledgements 89 References 90

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 4
Appendix 1. Distinguishing character tables (draft) of Papio anubis, Papio cynocephalus,
Chlorocebus pygerythrus, Cercopithecus mitis, Colobus angolensis, Colobus guereza and their subspecies, in Kenya and Tanzania.
2. Primate groups/individuals encountered inside and outside the coastal forests of Kenya. 3. Diurnal road surveys conducted at the north coast of Kenya. 4. Diurnal road surveys conducted at the south coast of Kenya. 5. Diurnal road surveys, outside the coastal forests of Kenya. 6. Butynski, T.M., De Jong, Y.A., Perkin, A.W., Bearder, S.K. & Honess, P.E. 2006.
Taxonomy, distribution, and conservation status of three species of dwarf galagos (Galagoides) in eastern Africa. Primate Conservation 21: 63-79.
7. Culverwell, J., Feely, J., Bell-Cross, S., De Jong, Y.A. & Butynski, T. M. 2008. A new pig for Tsavo. Swara 31: 50-52.
8. De Jong, Y.A. Culverwell, J. & Butynski, T.M. 2009. Desert warthog Phacochoerus aethiopicus found in Tsavo East National Park and Tsavo West National Park, southern Kenya. Suiform Soundings 8: 4-6.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 5
Tables 1 Primate species and subspecies occurring in the coastal forest of Kenya,
number of encounters, population trend, current IUCN Red List degree of threat category, and abundance in the coastal forest of Kenya.
22
2 Rates of encounter with Otolemur garnettii lasiotis from 2005 – 2008 in the coastal forests of Kenya.
39
3 Rates of encounter with Galagoides cocos during 2005 – 2008 in the coastal forests of Kenya.
47
4 Encounter rates of Cercopithecus mitis albogularis during 2005 – 2008 in the coastal forests of Kenya and outside the Hotspot.
52
5 Field descriptions of C. m. albogularis Unguja Island, Tanzania, C. m. albogularis Usa River, Tanzania, C. m. albogularis Mrima Hill, Kenya, Cercopithecus mitis albogularis Diani, Kenya and C. m. albogularis Gedi Ruins, Kenya.
53
6 Encounter rates of Cercopithecus mitis albotorquatus during 2005 – 2008 in the Coastal Forests Kenya and while driving to the Hotspot.
59
7 Field descriptions of C. m. albogularis, Gedi Ruins, Kenya, C. m. albotorquatus, Witu Forest Reserve, Kenya and C. m. albogularis, Unguja Island, Tanzania.
60
8 Encounter rates (groups/hour and groups/km) of Chlorocebus pygerythrus hilgerti during 2005 – 2008 in the coastal forests of Kenya and travelling to the coastal area.
65
9 Field descriptions of Chlorocebus pygerythrus hilgerti obtained from the extremes of the coastal forests of Kenya (i.e., Diani and Mpeketoni) and from Shaba National Reserve in central Kenya.
66
10 Encounter rates of Papio cynocephalus ibeanus during 2005 – 2008 in the Kenya coastal forest Hotspot and while driving to the Hotspot.
73
11 Field descriptions of adult male P. c. ibeanus obtained from Manda Toto Island, Lamu Archipelago (close to the type locality of P. c. ibeanus), Tana River Primate National Reserve, Diani Beach (S coast).
76
12 Encounter rates (groups/hour and groups/km) of Colobus angolensis palliatus during 2005 – 2008 in the coastal forests of Kenya.
78
13 Phenotypic comparison of the Chlorocebus pygerythrus hilgerti x Cercopithecus mitis albogularis hybrid, Chlorocebus pygerythrus hilgerti and Cercopithecus mitis albogularis using field descriptions and photographs obtained at Diani Beach, Kenya.
85

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 6
Figures 1 Transects along which primate surveys were conducted from August 2005 through
July 2008 in the coastal forests of Kenya, and along roads leading to these forests. 16
2 Transects along which primate surveys were conducted from August 2005 through July 2008 in Kipini Conservancy, Witu Forest Reserve, and the Lamu Archipelago, northeast Kenya, and along roads leading to these sites.
17
3 Transects along which primate surveys were conducted from August 2005 through July 2008 in the coastal forests of southeast of the coastal forests of Kenya, and along roads leading to these sites.
18
4 Photo map for Chlorocebus pygerythrus. 20 5 Manda Toto Island. 25 6 Cercopithecus mitis albotorquatus in Witu Forest Reserve. 26 7 Otolemur garnettii lasiotis, Tana River Primate National Reserve, Kenya. 38 8 Approximate geographic distribution of Otolemur garnettii lasiotis in Kenya 38 9 Sonogram and oscillogram of the trailing advertising call of Otolemur garnettii
lasiotis from Diani Beach. 39
10 Colour of the tip of the tail of Otolemur garnettii lasiotis on the coast of Kenya. 41 11 Captive adult male Otolemur garnettii in Malindi, Kenya. 41 12 Galago senegalensis braccatus, Isiolo, central Kenya. 43 13 Approximate geographic distribution of Galago senegalensis braccatus in the
coastal area of Kenya. 44
14 Galagoides cocos, Takwa Ruins, Manda Island, Kenya. 46 15 Known distribution and encounters (2003-2008) with Galagoides cocos in Kenya. 47 16 Cercopithecus mitis albogularis, Diani, Kenya. 50 17 Approximate geographic distribution of Cercopithecus mitis albogularis in the
coastal area of Kenya. 51
18 Adult male C. m. albogularis at Jozani-Chwaka Bay National Park, Unguja Island, Tanzania.
55
19 Adult male C. m. albogularis at Gedi Ruins, Kenya. 55 20 Cercopithecus mitis albotorquatus male from Witu Forest, Kenya. 57 21 Known distribution and encounters (2003-2008) with Cercopithecus mitis
albotorquatus. 58
22 Chlorocebus pygerythrus hilgerti male at Diani Beach, Kenya. 63 23 Approximate geographic distribution of Chlorocebus pygerythrus hilgerti in the
coastal area of Kenya. 64
24 Suspected geographic distribution and evidence for Chlorocebus pygerythrus excubitor.
68
25 Adult male Papio cynocephalus ibeanus foraging on the beach of Manda Toto Island, Lamu Archipelago, Kenya.
71
26 Approximate geographic distribution of Papio cynocephalus ibeanus over the coast of Kenya.
72
27 Adult male Colobus angolensis palliatus at Mrima Hill, south coast of Kenya. 77 28 Known distribution and encounters (2003-2008) with Colobus angolensis palliatus
in the coastal forests of Kenya. 77
29 Cercocebus galeritus at Mchelelo Research Station, Tana River Primate National Reserve, Kenya.
82
30 Adult male Chlorocebus pygerythrus hilgerti x Cercopithecus mitis albogularis hybrid, Diani, Kenya.
84
31 Encounter site of C. p. hilgerti x C. m. albogularis hybrid. Diani, Kenya. 84

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 7
32 Dzombo Hill Forest Reserve, surrounded by degraded bushland, agriculture and human settlements.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 8
EXECUTIVE SUMMARY Although primates represent one of the best-known taxonomic groups found in the “Eastern Arc Mountains and Coastal Forests of Tanzania and Kenya Biodiversity Hotspot” (EACF Hotspot), numerous important questions remain concerning taxonomy, distribution, abundance, conservation status, and priorities for conservation actions. The objectives of this study were to 1) determine the distribution, diversity, and taxonomy of the primate fauna in the coastal forests of Kenya, to 2) determine the conservation (Red List Degree of Threat) status of all taxa of primates in the coastal forests of Kenya, and to 3) determine the primary threats to all taxa of primates in the coastal forests of Kenya.
Four survey areas were visited [1) Lamu Archipelago, 2) Kipini Conservancy and Witu Forest Reserve, 3) North Coast (Mombasa to Malindi), and 4) South Coast (Mombasa to Lunga Lunga)] in seven trips totalling 38 survey days. Additionally, regions to and from the survey areas, including sites outside the Hotspot, were surveyed. In total, 5439 km of diurnal and nocturnal surveys were conducted within and to/from the coastal forest of Kenya. Primates were encountered 239 times. Of these encounters, 178 were with groups and 61 were with solitary individuals. Of the 178 encounters with groups, 99 where in the coastal forests of Kenya, 79 were while travelling to or from these forests. Nine genera, nine species, and 11 subspecies of primate occur in the coastal forest of Kenya; Otolemur garnettii lasiotis, Galago senegalensis braccatus, Galagoides cocos, Cercopithecus mitis albogularis, Cercopithecus mitis albotorquatus, Chlorocebus pygerythrus hilgerti, Chlorocebus pygerythrus excubitor, Papio cynocephalus ibeanus, Colobus angolensis palliatus, Procolobus rufomitratus rufomitratus, Cercocebus galeritus. Three out of the eleven subspecies are nocturnal. Of the 11 subspecies, 55% (n=6) in are categorized by the 2008 IUCN Red List of Threatened Species as “Least Concern”, 9% (n=1) as “Vulnerable”, 18% (n=2) as “Endangered”, while 18% (n=2) were not assessed (Table x). The coastal forest of Kenya have been much reduced, fragmented and degraded, and what little forest remains is under increasing threat. The expansion of agriculture is the most critical threat, while the production of charcoal, taking of firewood and timber, and mining are additional serious causes of habitat loss and degradation (Obura, 2007). Papio cynocephalus ibeanus was the most often encountered primate in the coastal forests of Kenya (32 groups), followed by Cercopithecus mitis albogularis (24 groups), and Cercopithecus mitis albotorquatus (16 groups). Otolemur garnettii lasiotis and Galagoides cocos are common while Galago senegalensis braccatus is present but rare in this part of Kenya. This study found significant geographic range extensions for O. g. lasiotis, G. cocos and C. m. albotorquatus, and is the first to report a wild Cercopithecus mitis albogularis x Chlorocebus pygerythrus hilgerti hybrid. With data collected during this study,we reviewed (Butynski et al., 2006) the complicated nomenclatural history for the Kenya coast galago, Galagoides cf. cocos, and examined whether ‘cocos’ is the valid species name for this recently resurrected taxon. We concluded that Galagoides cocos is the name that should be applied to the Kenya coast galago---not Galagoides zanzibaricus. In Malindi, a phenotypically different looking captive O. g. lasiotis was observed. We observed considerable phenotypic differences among Cercopithecus mitis in the type locality

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 9
for C. m. albogularis (Unguja Island, Tanzania), the western-most locality for ‘C. m. albogularis’ that we visited (Usa River, close to the type locality of kibonotensis), the northeastern-most locality that we visited (Gedi Ruins), and those in between these localities (e.g., Mrima Hill, Vanga, Diani, Lunga Lunga). These phenotypic differences appear to be in a cline. C. m. albotorquatus is a poorly known subspecies. During this survey, C. m. albotorquatus were found to be common in the Witu Forest Reserve, Kipini Conservancy, and Tana River Primate National Reserve, but rare on Manda Island and on Lamu Island. The question of whether Cercopithecus mitis phylax is a valid subspecies remains unresolved.
Surprisingly little phenotypic variation was observed for C. p. hilgerti populations in the coastal forest of Kenya. We did, however, find variation in the intensity of coloration of the pelage. The question of whether C. p. excubitor is a valid subspecies remains unresolved. C. pygerythrus is present in the Lamu Archipelago but was not encountered during this study. If C. p. excubitor is a valid subspecies, than its geographical range is highly fragmented and it population size is critically low. P. c. ibeanus is a widespread, common and opportunistic primate in the coastal forests of Kenya. Outside the Hotspot it is even more abundant, occurring in and outside protected areas. Phenotypic variation was observed throughout its range, both inside and outside the coastal zone. During this study, an extensive hybrid zone between P. anubis and P. c. ibeanus across Kenya was observed: 1) from the northeast and east Mt. Kenya to the Lower Tana River, and 2) along the Nairobi – Mombasa Highway, starting at least at Makindu (north of the Chyulu Hills National Park), through Tsavo, to the coast. P. c. ibeanus is present along the coast, but differs phenotypically from the north coast to the south coast. Primate biodiversity is relatively high in the Tana River Primate National Reserve (seven species), Diani (six species), and Kipini Conservancy and Witu Forest Reserve (five species). Additional primate surveys in Boni and Dodori Forest Reserves, Patta Island, Kaya Gonja, Shimba Hills National Reserve, Buda Forest, and Mrima Hill (nocturnal surveys) are essential for compiling a list of ‘Primate Priority Conservation Sites in the coastal forest of Kenya. There can be no doubt, however, that the top priority site for primate conservation in the coastal forests of Kenya, indeed for all of Kenya, are the forests along the Lower Tana River. These forests not only hold the highest diversity of primate species in all of Kenya, they hold Kenya’s only two endemic species of primate. In addition, they are among the most threatened forests in all of Kenya and are of great importance for the conservation of many other taxa. During surveys driving to/from the coastal forests of Kenya, the desert warthog Phacochoerus aethiopicus was found west of Garissa. This is the first record west of the Tana River and extends the known geographic range to the northwest ca. 265 km. We also obtained the first record for P. aethiopicus in Tsavo East and Tsavo West National Parks (range extension of ca. 390 km to the southwest). In addition, common warthog Phacochoerus africanus and P. aethiopicus were found to be sympatric in Tsavo West National Park. This is the first site at which P. aethiopicus and P. africanus are known to be sympatric. To visually present the phenotypic diversity of the primates that occur in the coastal forests of Kenya, six photographic maps were developed (wildsolutions.nl; De Jong & Butynski, 2009). These comprise 288 of our photos taken of primates in Kenya and Tanzania, of which 72 are of Kenya coastal forest primates.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 10
Priority research questions for the primates of the coastal forests of Kenya include:
1) To what extent does phenotypic variation in O. g. lasiotis occur over its geographic range?
2) What is the southern boundary for O. g. lasiotis? 3) What is the geographical range of G. s. braccatus within and outside the coastal forests
of Kenya? , 4) How far up the Tana River does the range of G. cocos extent? 5) Which subspecies of C. mitis occurs between the Tana River/Delta and the Galana
River? 6) What subspecies of C. mitis occurs between the Galana River and Kilifi Creek? 7) Is C. m. phylax a valid subspecies and, if so, what is its geographic range? 8) What is the geographic range of C. p. hilgerti? 9) Is C. p. excubitor a valid subspecies and, if so, what is its geographic range? 10) What is the southern limit of the geographic range of C. m. albotorquatus? 11) Are there other reports of wild C. p. hilgerti x C. m. albogularis, and do they
phenotypically resemble the ‘Diani hybrid’? 12) Does Galagoides zanzibaricus occur in Kenya? 13) What primate taxa occur in the forests along the extreme northern coast of Kenya
(especially Boni-Dodori Forest)? 14) What primate taxa occur on Patta Island? 15) What is the taxonomic status of Papio in the forests along the Lower Tana River?

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 11
1. INTRODUCTION
Primates are of particular interest and importance because (1) they are essential components (often “keystone species”) of the ecosystems in which they occur, affecting the composition of the vegetation and accounting for a significant portion of the mammalian biomass; (2) they are vital to our understanding of human evolution and human diseases; (3) they are among the best indicators of the health of ecosystems, and (4) they are among the most important “flagship species” for those sites in which they are found. Indeed, the endemic and highly threatened primates have been highly instrumental in bringing the biological importance and plight of the Eastern Arc Mountains and the coastal forests of East Africa to the attention of the conservation community over the past 20 years.
For conservation purposes it is important to identify species and subspecies. With natural history data, together with carefully assessed geographical species and subspecies ranges, adequate and effective conservation measures can be taken. Primates are an especially important taxonomic group for the focus of conservation actions in the “Eastern Arc Mountains and Coastal Forests of Tanzania and Kenya Biodiversity Hotspot” (EACF Hotspot). The EACF Hotspot supports no fewer than nine endemic species and five endemic (or near-endemic) subspecies of primates are reported to occur in the Hotspot. Several of these primate taxa are listed in the IUCN/SSC Red List as “Critically Endangered” or “Endangered”. Others are listed as “Data Deficient”. This means that, “…there is inadequate information to make a direct, or indirect, assessment of its risk of extinction based on its distribution and/or population status”.
Although primates represent one of the best-known taxonomic groups found in the Hotspot, numerous important questions remain concerning taxonomy, distribution, abundance, conservation status, and priorities for conservation actions. In addition, field studies in which the phenotypic variation of primates species and subspecies is assessed are rare. “Undiscovered” species or subspecies of primates are still being found and the knowledge about how many species and subspecies of primates occur in the Hotspot, where they occur, and the level of extinction risk each taxon faces, is vital for primate conservation. In particular, there might well be additional unrecognized “cryptic” species in the genera Otolemur, Galago, and Galagoides. The main goal of this study is to prevent the loss of primate biodiversity in the coastal forests of Kenya. The objectives were to 1) determine the distribution, diversity, and taxonomy of the primate fauna in the coastal forests of Kenya, 2) determine the conservation (Red List Degree of Threat) status of all taxa of primates in the coastal forests of Kenya and 3) determine the primary threats to all taxa of primates in the coastal forests of Kenya.
With support from CEPF we (1) undertook primate-focused surveys throughout the Kenya coastal forest part of this Hotspot (onwards refered to as coastal forests of Kenya), (2) conducted a search of the literature along with much detailed communications with colleagues, (3) undertook Red List assessments for the 2008 IUCN Red List of Threatened Species of 24 species and subspecies, and (4) published ‘primate photographic maps’,(5) established a website to communicate our data and results, and (6) published three articles based on research collected under this project.
Primate surveys were conducted throughout the coastal forests of Kenya, as well as while driving to and from the Hotspot. The focus was on primate diversity, abundance, distribution, threats, and conservation status in the coastal forests of Kenya. The surveys served to provide information, for each taxon, on (1) phenotypic variation, (2) geographic range limits along the coast of Kenya, (3) a rough indication of abundance, (4) a baseline against which to monitor change, and (5) primary threats.
The Co-Project Leaders took advantage of their time in the field to collect opportunistic data on the distribution and abundance on other mammal taxa within and outside the Hotspot, in particular the desert warthog Phacochoerus aethiopicus, common warthog Phacochoerus africanus, and dik dik

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 12
Madoqua. Data on the taxonomy, abundance, distribution, and conservation status of some of Kenya’s ‘non-Hotspot’ primate taxa were also collected; particularly on the Somali galago Galago gallarum Thomas, 1901, Kenya lesser galago Galago senegalensis braccatus Elliot, 1907, and eastern patas monkey Erythrocebus patas pyrrhonotus (Hemprich & Ehrenberg, 1829).
This study did not focus on the Tana red colobus Procolobus rufomitratus rufomitratus (Peters, 1879) or on the Tana mangabey Cercocebus galeritus Peters, 1879. Although both of these species are threatened, they are relatively well known, and have received considerable recent attention under various other projects.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 13
2. STUDY AREA This study was conducted in the Kenya part of the Eastern Arc Mountains and Coastal Forests of Tanzania and Kenya Hotspot (in this study referred to as ‘coastal forests of Kenya’). This Hotspot stretches from the border of Kenya with Somalia, to the border of Tanzania with Mozambique, and includes Unguja (Zanzibar) Island, Mafia Island, and Pemba Island. In this study, we focused on the Kenya coastal area, a ca 600 km long sea front strip from Ishakani (1º 41’S) in the north to Vanga (4 º 40’S) in the south. The intervening habitats between the coastal forest patches are included in the Hotspot (CI, 2003). The vegetation in the Hotspot falls within the “Zanzibar-Inhambane Regional Mosaic Zone” (White, 1983). The Hotspot corresponds for a large part (with the exception of Somalia) with the WWF’s “Northern Zanzibar-Inhambane Coastal Forest Mosaic Ecoregion”. In 1990, 95 forest patches were recorded in the coastal forests of Kenya. These covered an area of 660 km² (Burgess et al., 2000a). The mean patch size was 6.7 km² in Kenya. The two largest coastal forests in Kenya are Arabuko-Sokoke (ca 370 km²) and the Shimba Hills National Reserve (ca 63 km2) (Younge et al., 2002). The coastal forests of Kenya support a high level of endemism, but are treatned due to clearance, fragmentation and degradation (by agriculture, settlement and tourism), illegal logging, charcoal burning, and other human activities. Climate is an important natural factor and influence on the vegetation structure and growth (Clarke, 2000). The climate at the Kenya Coast is mainly influenced by the large-scale pressure systems of the Western Indian Ocean and monsoon winds. During December through March the winds blow from the northeast and during May through October they blow southeast. Inbetween there are 1-2 month transition periods with variable and lower winds (Kairo & Bosire, 2007). The mean annual temperature at Mombasa is 26.3°C, with a mean annual maximum of 30.3°C and a mean annual minimum of 22.4°C (Irebelo, 2006). The north coast receives less rain which increases towards the south coast, and becomes less seasonal in the south. The north coast has two rainy seasons while the south coast one (Hawthorne, 1993 in Clarke, 2000). The Lamu Archipelago receives a mean annual total rainfall of 889 mm, 80% of which falls from April to June, with 345 mm in May (Irebelo, 2006). Malindi receives 1022 mm/yr. Mombasa receives1040 mm/yr, with an average of 240 mm in May. Vanga, the extreme southeast corner of Kenya on the Tanzanian border, is wet from May to December (Irebelo, 2006). The population of Kenya is estimated to be 32.0 million people, with ca 2.8 million (9.0%) residents in the coastal areas (Kairo & Bosire, 2007). The population growth rate at the coast is 3.1%, the national average is 2.9%. Population densities are highest in urban centers like Mombasa, Malindi, Lamu and Kilifi (Kairo & Bosire, 2007). Land use in the coastal region can be devided into four main categories, livestock ranches in the hinterland, agricultural settlement schemes, private land along the coastline, and un-alienated government land (Kamula & Ochiewo, 2007). Economic activities along the coast range from industries (e.g. salt industries and cement production), service (e.g. tourism), transport, fishing, agriculture and cottage industries (arts and crafts). The economic contribution of coastal activities to the national economy is ca 15% (of which 12.5% from tourism; Kairo & Bosire, 2007). The two main rivers that flow through and drain the coast are the Tana River and the Sabaki River. The Tana River (850 km in lenght) originates at Mt Kenya and enters the Indian Ocean at Kipini. The Sabaki River originates as the Athi River near Nairobi (Central Highlands) which when joined with the Tsavo River (close to Tsavo town), becomes the Galana River. A few kilometers north of Malindi the Sabaki River enters the Indian Ocean. The entire Athi-Galana-Sabaki system extends for 390 km (Kairo & Bosire, 2007).

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 14
National Parks and Reserves along the Kenya coast protect the Indian Ocean territorial waters and the areas that border the ocean (including the coral reefs, mangrove forests and beaches). On the north coast there are several parks and reserves, including Dodori Coastal Reserve, Boni National Reserve, Arabuko Sokoke Forest Reserve, Tana River Primate National Reserve, Kiunga Marine National Reserve, Malindi Marine National Park and Reserve, Watamu Marine National Park, and the Ras Tenewi Coastal Zone National Park (between the Tana River Delta and Lamu). On the south coast, the following protected areas are established, Shimba Hills National Reserve, Mombasa Marine National Park and Reserve, Diani-Chale Marine National Park and Reserve, Kisite Marine Park, and Mpunguti Reserve (Kenya Wildlife Service, 2009). For this study we divided the coastal area of Kenya in to four survey areas: 1) Lamu Archipelago, 2) Kipini Conservancy and Witu Forest Reserve, 3) North Coast (Mombasa to Malindi), and 4) South Coast (Mombasa to Lunga Lunga). Additionally, regions to and from the survey areas, including sites outside the Hotspot, were surveyed.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 15
3. METHODS
3.1 Preparations The literature was searched and consulted in order to (1) prepare a list of questions related to the taxonomy and distribution of the primates of the coastal forests of Kenya and Tanzania, (2) obtain all information related to the taxonomy and distribution of Kenya’s primates, and (3) compile the distinguishing (i.e., diagnostic) characters for each taxon of primate in the coastal forests of Kenya and Tanzania as presented by various authors. Distinguishing character (review and comparison) tables were drafted for Olive baboon Papio anubis (Lesson, 1827), yellow baboon Papio cynocephalus (Linnaeus, 1766), vervet monkey Chlorocebus pygerythrus (F. Cuvier, 1821), Sykes’s monkey Cercopithecus mitis Wolf, 1822, Angolan colobus Colobus angolensis Sclater, 1860, Colobus guereza Rüppel, 1835, and their subspecies, in Kenya and Tanzania (Appendix 1) for quick reference and comparison in the field. Additionally, these draft distinguishing character tables were distributed to several CEPF partners working in the Eastern Arc Mountains and Coastal Forests of Tanzania and Kenya Biodiversity Hotspot. The distinguishing character tables will, once finalized, be available on our website (www.wildsolutions.nl). A photo album was prepared and maintained for each primate taxon for quick reference and comparisons in the field. 3.2 Field surveys In order to confirm presence of diurnal and nocturnal primates, assess the relative abundance of primates, and meet the need to cover large areas in a limited time, rapid assessment survey methods were used. Differences in research conditions, constraints, and opportunities in the survey sites required a variety of methods and approaches. Field surveys were conducted from August 2005 through July 2008 by T. M. Butynski and Y. A. de Jong. A total of 302 survey hours were completed in 38 days, covering a distance of 5439 km. 3.2.1 Diurnal primate surveys A total of 218 h of diurnal surveys were completed during 38 surveys conducted during seven trips that covered a distance of 5265 km (Figures 1, 2 & 3). Surveys were conducted from a vehicle, boat, or on foot by two people. Surveys took place both in the Hotspot as well as during travels along roads to and from the Hotspot (referred to as ‘road surveys’ in this study). The number of primate groups encountered per kilometre and per hour were the indices used to assess relative abundance (Butynski & Koster, 1994; White & Edwards, 2000; Nekaris & Jayewardene, 2004). Information collected during each survey included date, weather conditions, start time, finish time, places surveys (GPS), walking/driving speed (GPS), and distance covered (GPS). When primate groups were encountered during a survey, the following data were collected: date, time, GPS coordinates (Garmin GPSmap 60Cx), altitude (by GPS), primate species/subspecies, vegetation type, and tree density. The focus during every primate encounter was on obtaining a detailed description of as many individuals in a group as time and visibility allowed. Photos were taken with a Nikon D70 digital camera fitted with a 400 mm Nikon lens, and with a Canon EOS 40D digital camera fitted with a Canon 75-300 mm lens. Most of the photos were shot in ‘raw’. As many of the individuals as possible in each group were photographed. Each primate group was appointed a unique number (hereafter referred to as the ‘group number’). The track of the complete survey was saved in a GPS and downloaded in a Dell Latitude notebook using Garmin MapSource software (Figure 1, 2 & 3).

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 16
Figure 1. Transects along which primate surveys were conducted from August 2005 through July 2008 in the coastal forests of Kenya, and along roads leading to these forests. Figure 2 and 3 show more details for northeast Kenya and the southeast of Kenya.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 17
Figure 2. Transects along which primate surveys were conducted from August 2005 through July 2008 in Kipini Conservancy, Witu Forest Reserve, and the Lamu Archipelago, northeast Kenya, and along roads leading to these sites.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 18
Figure 3. Transects along which primate surveys were conducted from August 2005 through July 2008 in the coastal forests of southeast of the coastal forests of Kenya, and along roads leading to these sites.
3.2.2 Nocturnal primate surveys The presence of galagos was recorded during nocturnal surveys. Nocturnal surveys were conducted for a total of 20 h, covering 175 km. These surveys were conducted from a vehicle and/or by foot at all study sites. These surveys were conducted between 18.45 - 23.00 h and 4.00 - 06.30 h. Reflection from the eyes of galagos can be observed at >100 m in suitably open habitats. Torches (Maglights and Petzl Tikka XP headlamps) were used to scan the vegetation. Walks and drives were conducted slowly (ca 0.5-1 km/h on foot and ca 5 -10 km/h by vehicle) with pauses to scan the vegetation, observe primates, and/or record vocalisations. The following were recorded: date, weather conditions, moon phase, start time, finish time, localities surveyed (GPS), walking/driving speed (GPS), and distance covered (GPS). When galagos were encountered, binoculars (Zeiss Victory 10x42 and Zeiss Dialyt 7x42B) were used. The following data were collected when primates were encountered: date, time, moon phase, GPS coordinates, altitude, primate species/subspecies, vegetation type, tree density, number of individuals, and height of individuals in vegetation. Additionally, phenotypic descriptions were obtained and photos were taken using a Canon EOS 40D digital camera with a 75-300 mm Canon lens combined with a Canon Speedlite 420EX flash. 3.2.3 Nocturnal listening survey A total of 64.0 listening surveys were completed during 38 survey days. The advertisement call of galagos provides species specific information that can be used for species identification (Zimmermann, 1994; Bearder et al, 1995). Listening from a fixed point can reveal the presence of galago species. Audio recordings of galago vocalizations (and other nocturnal mammals and birds), preferably the loud advertisement call, were made during surveys, or opportunistically, using two

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 19
Marantz Digital PMD660 recorders with Sennheiser Shot-Gun ME-66 microphones. The time and date of every recording is automatically saved within the audio file and notes were kept in our notebooks. Nocturnal listening surveys were mainly conducted from camp or from a higher point at dusk, dawn, and before and after nocturnal road or walking surveys. 3.3 Data analysis 3.3.1 Galago vocalization analysis Audio files were transferred from the Marantz digital audio recorder to a laptop. The best quality recordings were used to identify the species vocalizing. When confirmation was needed, vocalizations were send to S. K. Bearder and/or A. Perkin, Nocturnal Primate Research Group, Oxford Brookes University, Oxford, UK. Sonograms and spectrograms, as well as the numerical acoustic parameters, were produced from each vocalization using Avisoft-SASLab Pro software (R. Spect, Berlin; version 4.51). 3.3.2 Distribution mapping Locality records [date, time, locality name, latitude (decimal), longitude (decimal), altitude (m asl), primate species/subspecies, group code, vegetation type, notes] for all primates encountered during surveys were stored in a Microsoft Access database (XP). All records were plotted on a map using Garmin MapSource (6.10.2) and MapInfo Professional 8.0. 3.4 Primate Photographic Maps To present the phenotypic diversity of primates occurring in the coastal forests of Kenya, six photographic maps (‘photo maps’) were developed (Figure 4). These comprise 288 photos of out the thousands of photos taken of primates in Kenya and Tanzania. Of these, 72 are of primates in the coastal forests of Kenya. The photos were sorted into six taxonomic groups (Galagonidae, Papio, Cercopithecus mitis, Colobinae, Chlorocebus pygerythrus, and Chlorocebus pygerythrus hilgerti x Cercopithecus mitis albogularis hybrid). All photos were ‘geotaged’. The associated coordinates were either obtained automatically with a phototracker GPS (Gisteq PhotoTrackr) or a handheld GPS (Garmin, GPSmap 60Cx) using Picasa (version 2.7) software, and Google Earth (version 4.3.7284.3916 Beta) software to geotag the individual photos. The photos were uploaded to a Picasa Web Album which shows them automatically on a photo map (Figure 4). Viewers of the photo maps can select a road, a terrain, or a satellite map and then zoom in or out on specific areas. Photos can be enlarged and viewed separately on a detailed map. Additionally, viewers can change from “Map View” to “Album View” which gives an overview of all photos on the photo map. Each photo is labeled with the taxonomic name (genus, species, subspecies), name of the locality were the photo was taken, vegetation type, altitude, and various comments. Viewers are invited to leave their feedback and comments with each photo. The photo maps are available for download in Google Earth KML format. The first draft of the photo map for each of the six primate taxonomic groups is now available online. The ‘official version’ will be launched in March 2009. The web addresses will be distributed to a large number of primatologists. These primatologists will be invited to leave their comments with the photos and to discuss primate biogeography, diversity, taxonomy, and conservation. These six photo maps will serve as the beginning of a ‘living’ photo collection of eastern Africa’s primates. These six maps will be updated soon after new photos are available. Photo maps for other primate taxa in eastern Africa will gradually be added to this collection. This

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 20
resource can be used by anyone interested in the biogeography, diversity, taxonomy, and conservation of eastern Africa’s primates.
Figure 4. Screenshot of the ‘photo map’ for Chlorocebus pygerythrus. Overview of the six photo maps:
Galagonidae: Total of 28 photos uploaded of which seven were taken in the coastal forests of Kenya. Taxa photographed and mapped include: Otolemur garnettii lasiotis, Otolemur garnettii panganiensis, Otolemur garnettii garnettii, Otolemur monteiri monteiri, Galago senegalensis braccatus, Galago gallarum, Galagoides cocos, Galagoides granti. Web address: http://picasaweb.google.com/wildsolutions/Galagonidae?feat=email#
Papio: Total of 103 photos uploaded of which 17 were taken in the coastal forests of Kenya. Taxa photographed and mapped include: Papio cynocephalus ibeanus, Papio cynocephalus cynocephalus, Papio anubis, hybrids between P. cynocephalus and P. anubis. Web address: http://picasaweb.google.com/wildsolutions/PapioAnubisPapioCynocephalus?feat=email# Cercopithecus mitis: Total of 46 photos uploaded, of which 20 were taken in the coastal forests of Kenya. Taxa photographed and mapped include: Cercopithecus mitis albogularis, Cercopithecus mitis albotorquatus, Cercopithecus mitis kolbi. Web address: http://picasaweb.google.com/wildsolutions/CercopithecusMitis?feat=email#
Colobinae: Total of 17 photos uploaded, of which three were taken in the coastal forests of Kenya. Taxa photographed and mapped include: Colobus angolensis palliatus, Colobus

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 21
guereza kikuyuensis, Colobus guereza caudatus, Procolobus kirkii, Procolobus gordonorum. Web address: http://picasaweb.google.com/wildsolutions/Colobinae?feat=email#
Chlorocebus pygerythrus: Total of 89 photos uploaded, of which 20 were taken in the coastal forests of Kenya. The only taxon photographed and mapped is Chlorocebus pygerythrus hilgerti. Web address: http://picasaweb.google.com/wildsolutions/ChlorocebusPygerythrus?feat=email
Chlorocebus pygerythrus hilgerti x Cercopithecus mitis albogularis hybrid: Total of five photos uploaded, all from Diani, Kenya. Web address: http://picasaweb.google.com/wildsolutions/ChlorocebusPygerythrusHilgertiXCercopithecusMitisAlbogularisHybrid?authkey=Gv1sRgCMqKmI2R_d6zXA&feat=email#
All photo maps will become accessible in March 2009 through our website: www.wildsolutions.nl 3.5 Definitions In this study we apply the following definitions; Genus: “A genus is a monophyletic group of species (or a single species), which separated from other such groups earlier than the Miocene-Pliocene boundary.” (Groves, 2006). Species: “A species is a population (or group of populations), distinguished by the possession of one more consistent (fixed, absolute) heritable differences from other such populations.” (Groves, 2006). Subspecies: “A subspecies is a geographic segment of a species, distinguished by the possession at high frequencies, but not a much as 100%, of one or more heritable differences from other such segments.” (Groves, 2006). Extent of occurrence: “..the area contained within the shortest continuous imaginary boundary which can be drawn to encompass all the known, inferred or projected sites of present occurrence of a taxon, excluding cases of vagrancy” (IUCN, 2001). Area of occupancy: “..the area within its ‘extent of occurrence’ which is occupied by a taxon, excluding cases of vagrancy.” (IUCN, 2001). Kenya coastal forests: The Kenya coastal forest part of the Eastern Arc Mountains and Coastal Forests of Tanzania and Kenya Biodiversity Hotspot. The intervening habitats between the coastal forest patches are included.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 22
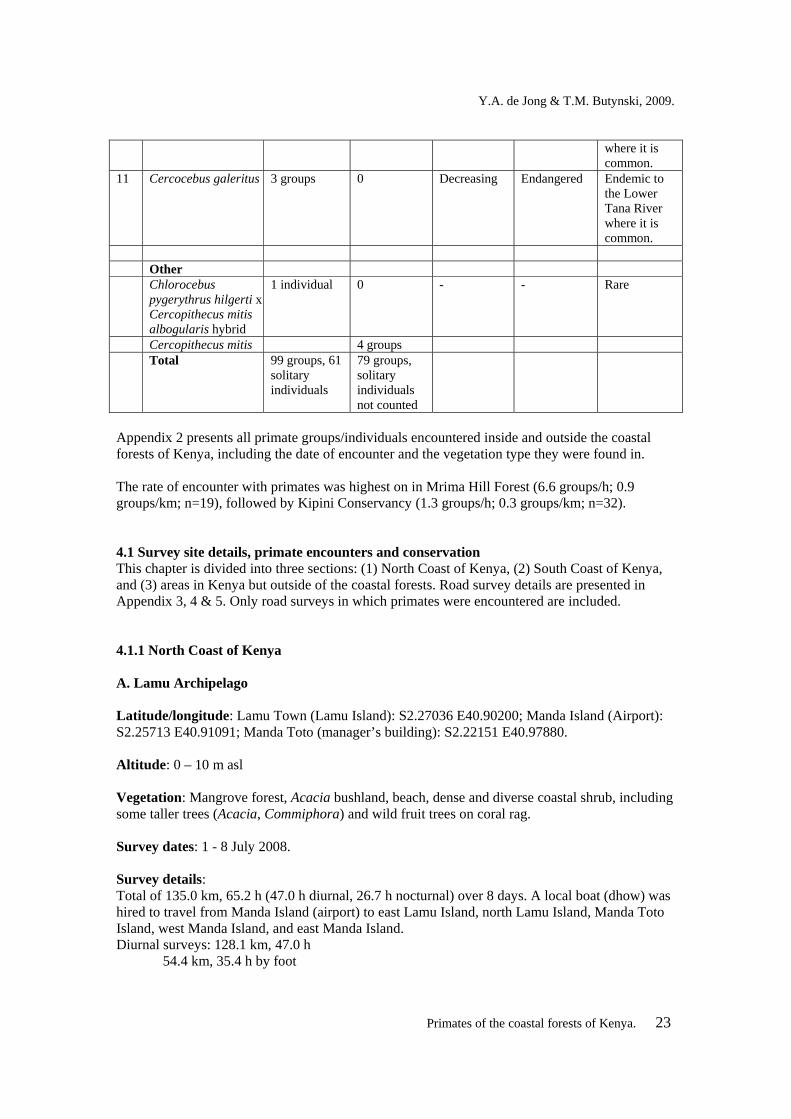
4. RESULTS During this project, primates were encountered 239 times during seven field trips that totalled 38 survey days. Of these encounters, 178 were with groups and 61 were with solitary individuals. Of the 178 encounters with groups, 99 where in the coastal forests of Kenya, 79 were while travelling to or from the coastal forests of Kenya (Table 1). Nine genera, nine species and 11 subspecies of primates occur in the coastal forest of Kenya. Eight of the subspecies are diurnal and three are nocturnal. Of the 11 subspecies, 45% (n=5) have more or less stable populations, 36% (n=4) have declining populations, and 18% (n=2) are data deficient (IUCN, 2008). Fifty-five percent (n=6) are categorized (IUCN, 2008) as “Least Concern”, 9% (n=1) as “Vulnerable”, and 18% (n=2) as “Endangered”, and 18% (n=2) were not assessed for the 2008 IUCN Red List of Threatened Species (Table 1). Table 1. Primate species and subspecies occurring in the coastal forest of Kenya, number of encounters, population trend, current IUCN Red List degree of threat category, and abundance in the coastal forest of Kenya. No. Subspecies Number
encounters in the coastal forests of Kenya
Number of encounters outside the coastal forestsof Kenya
Population trend (IUCN, 2008)
Degree of threat category (IUCN, 2008)
Abundance in the coastal forest of Kenya
Nocturnal primates
1 Otolemur garnettii lasiotis
>40 individuals
Present, not counted
Stable Least Concern Very common
2 Galago senegalensis braccatus
0 Present, not counted
Stable Least Concern Uncommon
3 Galagoides cocos >20 individuals
0 Stable Least Concern Common
Diurnal primates
4 Cercopithecus mitis albogularis
24 groups 3 groups Decreasing Least Concern Common
5 Cercopithecus mitis albotorquatus
16 groups
0 Decreasing Vulnerable Common in along the Lower Tana River and Witu area.
6 Chlorocebus pygerythrus hilgerti
11 groups 33 groups Stable Not assessed Common outside of the coastal forests.
7 Chlorocebus pygerythrus excubitor
0 0 ? Not assessed Rare
8 Papio cynocephalus ibeanus
32 groups
39 groups Stable Least Concern Very common
9 Colobus angolensis palliatus
11 groups 0 ? Least Concern Common in the coastal forest south of Mombasa
10 Procolobus rufomitratus rufomitratus
2 groups 0 Decreasing Endangered Endemic to the Lower Tana River
Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 23
where it is common.
11 Cercocebus galeritus 3 groups 0 Decreasing Endangered Endemic to the Lower Tana River where it is common.
Other Chlorocebus
pygerythrus hilgerti x Cercopithecus mitis albogularis hybrid
1 individual 0 - - Rare
Cercopithecus mitis 4 groups Total 99 groups, 61
solitary individuals
79 groups, solitary individuals not counted
Appendix 2 presents all primate groups/individuals encountered inside and outside the coastal forests of Kenya, including the date of encounter and the vegetation type they were found in. The rate of encounter with primates was highest on in Mrima Hill Forest (6.6 groups/h; 0.9 groups/km; n=19), followed by Kipini Conservancy (1.3 groups/h; 0.3 groups/km; n=32). 4.1 Survey site details, primate encounters and conservation This chapter is divided into three sections: (1) North Coast of Kenya, (2) South Coast of Kenya, and (3) areas in Kenya but outside of the coastal forests. Road survey details are presented in Appendix 3, 4 & 5. Only road surveys in which primates were encountered are included. 4.1.1 North Coast of Kenya A. Lamu Archipelago Latitude/longitude: Lamu Town (Lamu Island): S2.27036 E40.90200; Manda Island (Airport): S2.25713 E40.91091; Manda Toto (manager’s building): S2.22151 E40.97880. Altitude: 0 – 10 m asl Vegetation: Mangrove forest, Acacia bushland, beach, dense and diverse coastal shrub, including some taller trees (Acacia, Commiphora) and wild fruit trees on coral rag. Survey dates: 1 - 8 July 2008. Survey details: Total of 135.0 km, 65.2 h (47.0 h diurnal, 26.7 h nocturnal) over 8 days. A local boat (dhow) was hired to travel from Manda Island (airport) to east Lamu Island, north Lamu Island, Manda Toto Island, west Manda Island, and east Manda Island. Diurnal surveys: 128.1 km, 47.0 h
54.4 km, 35.4 h by foot

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 24
73.7 km, 11.6 h by boat (speed 4.9 – 6.3 km/h). Nocturnal surveys: 6.9 km, 3.7 h by foot Nocturnal listening survey: 14.7 h Primate species encountered: O. g. lasiotis, G. cocos, C. m. albotorquatus, P. c. ibeanus. Strong evidence was obtained for the presence of C. p. excubitor but this subspecies was never encountered. Encounter rates: Lamu Island: No diurnal or nocturnal primates seen. C. m. phylax: vocalizations heard from mangrove forest. Evidence was collected for the presence of C. p. excubitor. Residents stated that this taxon is present in very low numbers and occasionally hunted and kept/sold as pets. Manda Toto Island: Twice a group (3 individuals) of P. c. ibeanus was encountered on this small island. No nocturnal primates were heard or observed. Manda Island: The primate density on Manda Island is very low. C. mitis vocalizations (‘pyows’) heard once from the mangrove forest during a boat survey in the NW. In the SE one group of C. mitis was encountered (and ‘pyows’ heard) of which one individual was seen once during a ground survey. This group was in dense and diverse coastal shrub (including Acacia, Commiphora, baobab, and wild fruit trees) on coral rag on the edge of a mangrove forest. People report C. mitis to occur mainly in the mangrove forests. W Manda Island: G. cocos heard. Manda Island, at Takwa Ruins: G. cocos [4.1 individuals/h (n=6), 1.6 individuals/km (n=6)] and O. g. lasiotis [2.0 individuals/h (n=3), 0.8 individuals/km (n=3)]. Airport, Manda Island: several people said that C. p. excubitor visits the fresh water sources at the airport on a regular basis to drink. Audio recordings: Advertisement calls recorded of G. cocos and O. g. lasiotis Conservation The primate density in the Lamu Archipelago is very low. C. m. albotorquatus seems to be depended on the extensive mangrove forests surrounding the Islands. It is uncertain how much the other primate taxa depend on this ecosystem. Mangrove forests and other coastal wetlands occur for the most part on the north coast of Kenya, on the Lamu Archipelago, and at the tidal mouths of the Tana and Sabaki Rivers. Smaller wetlands occur in the mouths of semi perennial and seasonal coastal rivers on the south coast (Obura, 2007). The total area of mangroves forests in Kenya is estimated at 530 - 610 km². Sixty-sevel percent of Kenya’s mangroves occur in Lamu District, 10% in Kilifi District, and 10% in Kwale District. A total of 103 km² of mangrove forest has been lost [Abuodha & Kairo (2001) in Obura (2007)]. Mangrove forests have been extensively utilised for firewood, building poles (Rhizophora nzucronata is the preferred species), and masts (Heritiera littoralis) for Arab dhows (Irebelo, 2006). H.littoralis has been almost completely cut out from many sites. Mangrove swamps have been cleared, degraded and fragmented for human settlements and activities in several places (Irebelo, 2006; Obura, 2007).

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 25
Remarks In the past, the islands of the ‘Lamu Archipelago’ (i.e., Lamu, Manda, Manda Toto, and Patta Islands), have also been referred to as the ‘Witu Islands’ (e.g., Groves, 2001). The channels that separate the islands of the Lamu Archipelago Islands from the mainland are shallow and narrow (Patta Island ca 3 km; Manda Island ca 20 m; Lamu Island ca 150-200 m). As such, these islands are not particularly isolated. The extensive mangrove around all islands are probably a much greater barrier to primate movement than is the ocean (i.e., the channels). There are two endemic subspecies of primates described for the Lamu Archipelago, but the validity of both taxa is in question (Groves 2001, Grubb et al. 2003). Manda Island is the type locality for C. p. excubitor. The geographical range of C. p. excubitor is fragmented among the islands and its numbers appears to be extremely low (see Chapter 4.2.7). The type locality of C. mitis phylax is Patta Island. It is unclear if C. m. phylax is endemic to Patta Island (see Chapter 4.2.5). The density of diurnal primates on Lamu Island, Manda Toto Island, and Manda Island is very low. The diurnal primates that we observed were shy and the groups appeared to be small. C. mitis is present (apparently at low density) in the extensive mangrove forests of N Lamu Island (ca 150-200 m away from the mainland), and on NW and SW Manda Island. These are the first reported observation for C. mitis on these two islands. C. mitis were not detected on Manda Toto Island and are almost certainly absent from that tiny island (1 km²). It remains unknown as to which subspecies the C. mitis of Lamu Island and Manda Island belongs. The one C. mitis that we briefly observed on Manda Island could have been either C. m. albotorquatus or C. m. phylax. During this survey we collected the first record of G. cocos for the Lamu Archipelago. This record extended the range of this species 70 km to the east (Butynski et al., 2006; Chapter 4.2.3; Appendix 6; Figure 15). Further research: - Nocturnal and diurnal primate surveys on Patta Island to obtain presence/absence data, abundance data, and detailed field descriptions of all primate taxa. - The validity of C. m. phylax and of C. p. excubitor need to be assessed. This will not be easy given the low abundance and shyness of both species in the Lamu Archipelago. B. Kipini Conservancy and Witu Forest Reserve Latitude/longitude: S2.49910; E40.60710 (Kipini Conservancy headquarters) Altitude: 0-20 m asl Vegetation: Coastal thickets, open grassland with doum palm and bushes, patches of medium to dense moist and diverse coastal forest. Sandy soils.
Figure 5. Manda Toto Island on the left, Manda Island on the right. During low tide the two islands are almost connected.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 26
Survey dates: 21-25 June 2006 Survey details: Total of 132.5 km, 48.4 h (25.4 diurnal, 23.0 nocturnal) in 5 days. Diurnal surveys: 121.9 km;
113.9 km, 19.1 h by vehicle, 8.0 km, 6.3 h by foot
Nocturnal surveys: 11.6 km; 10.6 km, 2.0 h by vehicle, 1 km, 0.9 h by foot
Nocturnal listening hours: 20.1 h Primate species encountered: P. c. ibeanus, C. m. albotorquatus, C. p. hilgerti, G. cocos, and O. g. lasiotis. Encounter rates: Total of 32 diurnal primate groups encountered; 1.3 groups/h, 0.3 groups/km (n=32)
P. c. ibeanus: 0.8 groups/h, 0.2 groups/km (n=20). C. m. albotorquatus: 0.4 groups/h, 0.1 groups/km (n=10). C. p. hilgerti: 0.1 groups/h, 0.02 groups/km (n=2). O. g. lasiotis: 0.3 individual/h, 0.3 individual/km (n=3). unidentified nocturnal primate: 0.04 individual/h, 0.1 individual/km (n=1)
Audio recordings: Audio recordings of the advertisement calls of G. cocos and O. g. lasiotis were obtained. Conservation Kipini Conservancy was created in 2006. That same year, hundreds of illegal settlers arrived in the area, clearing a large portion of the natural vegetation. Other threats to the conservation of the area are illegal logging, tapping for palm wine, and poaching (Dowsett-Lemaire & Dowsett, 2007). In Witu Forest Reserve, a high level of illegal logging was observed. Some parts of the forest have been cleared of large trees. The biodiversity of Witu Forest Reserve and the water catchment of the forest are seriously threatened (Nielsen & Sick, 2008). Both G. cocos and O. garnettii are at low densities in Kipini Conservancy. P. c. ibeanus and C. p. hilgerti are common and often found in the vicinity of human settlements (in towns/villages, gardens). C. m. albotorquatus, a poorly-known subspecies, is common. Some authorities suggest state that C. m. albotorquatus is confined to the forests of the Lower Tana River (Chapter 4.2.5). That this is not the case as this subspecies is abundant in the Witu and Kipini forests. This is one
Figure 6. Cercopithecus mitis albotorquatus in Witu Forest Reserve

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 27
of the more significant finding of this project and one that bodes well for the long-term conservation of C. m. albotorquatus. In Kipini Forest we obtained the first solid evidence (good recordings of the advertisement call) for G. cocos north of the Tana River---extending the range ~60 km to the NE. This species is otherwise only known from the coastal forests of extreme northern Tanzania, and southern and central Kenya (Chapter 4.2.3, Appendix 6). The presence of G. cocos north of the Tana River/Tana Delta is of considerable conservation significance and suggests that this species may occur in extreme southern Somalia. Remarks Over the past 5 years, we have received reports of Procolobus in the Witu Forest Reserve and Kipini Forest. Kipini is ca. 90 km southeast of the known range of P. r. rufomitratus. Chapter 4.2.10 describes in more detail our search for Procolobus in Kipini Conservancy. If Procolobus are present at Witu and/or Kipini, they must be in very low numbers. This, coupled with the fact that the area of Kipini Forest in which Procolobus are said to occur has recently been given over by the Kenya Wildlife Service (KWS) and Kipini Conservancy (with funding from World Bank) for resettlement of people from the Lower Tana River, means that time is a critical factor as the small area of remaining (relatively species-rich) forest is likely to be destroyed in the near future. If Procolobus are present, then immediate actions will be necessary to prevent resettlement and to find an alternative solution for the people being resettled.
During February - March, 2007, members of the Danish Zoological Society (with CEPF funding) conducted biodiversity surveys in Witu Forest Reserve (Nielsen & Sick, 2008). Twenty mammal species, including four primates (Cercopithecus mitis, Papio cynocephalus, “Otolemur crassicaudatus” and ”Galago gallarum or senegalensis bracattus”, and eighty-five species of birds were recorded. Other recorded mammals include Lion Panthera leo, Bush Elephant Loxodonta africana, African Buffalo Syncerus caffer, Harveys Duiker Cephalophus harveyi, Suni Neotragus moschatus, and the Lesser Elephant Shrew Elephantulas rufescens. Duiker densities were low compared to other locations in East Africa. This was likely due to differences in habitat type and quality (Nielsen & Sick, 2008). Nielsen & Sick (2008) mention the possible presence of Galago gallarum in Witu Forest Reserve. Chapter 4.2.2 mentions that this is higly unlikely, given the habitat preference of G. gallarum (dry Acacia-Commiphora woodland/bushland and thorn scrub; Butynski & De Jong, 2004). It is more likely that the galago species they encountered during their surveys was either G. cocos or G. s. braccatus (Chapter 4.2.2), with G. cocos being the more likely. Nielsen & Sick (2008) report the presence of Otolemur crassicaudatus in Witu Forest Reserve. We encountered O. g. lasiotis in the area and strongly doubt the presence of O. crassicaudatus in the Reserve since this species is not known to occur in Kenya (Groves, 2001; T. M. Butynski & Y.A. de Jong, unpubl. data). The photos of Otolemur as presented in the report appear to be of O. g. lasiotis. Dowsett-Lemaire & Dowsett (2007) list the mammals they encountered during their vegetation, birds and mammals surveys in Kipini Conservancy during October-December, 2006. Five primate species are listed; Galagoides zanzibaricus, Otolemur garnettii, Papio cynocephalus, Cercopithecus mitis albogularis, and Cercopithecus aethiops. If we apply the taxonomy we use in this study to all five primate species, and add our knowlegde about their subspecies, the list becomes: G. cocos, O. g. lasiotis, P. c. ibeanus, C. m. albotorquatus and C. p. hilgerti. According to Dowsett-Lemaire & Dowsett (2007), all five primate species are common in the Conservancy.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 28
G. senegalensis was not recorded in Kipini Conservancy by Dowsett-Lemaire & Dowsett (2007). They observed G. senegalensis “in the bush country” of the Tana River Primate National Reserve (Andrews et al., 1975) and concluded that this species is probably not in Kipini Conservancy (in the absence of thornbush). Neither Nielsen & Sick (2008) nor Dowsett-Lemaire & Dowsett (2007) found Procolobus in the Witu - Kipini area, nor did we find evidence for Procolobus in this region. It now seems highly unlikely that Procolobus occur in this region. Further research There are no priorities for further primate research in the forests of the Witu - Kipini area.
C. Tana River Primate National Reserve Latitude/longitude: S1.87587; E40.13804 Altitude: 40 – 50 m asl Vegetation: Riverine evergreen gallery forest surrounded by grassland, wooded grassland, and arid medium-dense bushland on sandy soil (floodplain). Characteristic trees include Ficus spp, Phoenix reclinata, Acacia robusta, Populus ilicifolia, Blighia unijugata, Sorindeia madagascariensis, Diospyros mespiliformis, and Mimusops obtusifolia (Roberts & Luke, 1993 in Bennun & Njoroge, 1999). Survey dates: 6-7 August 2005 and 20-21 June 2006 Survey details: In total 30.3h [15.0 h (11.0 diurnal, 4.0 nocturnal) and 15.3 h (9.3 diurnal, 6.0 nocturnal)]. Diurnal surveys: ca 2 km, vehicle, along the edge of the forest Nocturnal surveys: ca 1 km, foot Nocturnal listening survey: 8 h. Recordings, photos, and phenotypic descriptions made of primates in the vicinity of Mchelelo Research Camp. Primate species encountered: P. c. ibeanus, C. m. albotorquatus, P. r. rufomitratus, C. galeritus, G. cocos, and O. g. lasiotis. Chlorocebus pygerythrus hilgerti and Galago senegalensis braccatus are present (Andrews et al., 1975; Butynski & Mwangi, 1994; Bearder et al., 2008), but were not encountered during this survey. Encounter rates 6-7 August 2005: Total of six diurnal primate groups encountered; 0.6 groups/h (n=6).
P. c. ibeanus: 0.1 groups/h (n=1). C. m. albotorquatus: 0.3 groups/h (n=3). P. r. rufomitratus: 0.1 groups/h (n=1). C. galeritus: 0.1 groups/h (n=1).

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 29
Encounter rates 20-21 June 2006: Total of seven diurnal primate groups encountered: 0.8 groups/h and 25 nocturnal primates; 4.2 individuals/h.
P. c. ibeanus encountered; 0.1 groups/h (n=1). C. m. albotorquatus encountered; 0.3 groups/h (n=3). P. r. rufomitratus encountered; 0.1 groups/h (n=1). C. galeritus encountered; 0.2 groups/h (n=2). O. g. lasiotis encountered; 15 individuals/h (n=15; estimate of 15 animals/ha). G. cocos encountered; 10 individuals/h (n=10; estimate of 15 animals/ha).
Audio recordings The adult male loud call (‘whoop-gobble’) of C. galeritus was recorded and send to Carolyn Ehardt, Project Leader of the CEPF-funded project ’Taxonomy and Conservation Genetics of the Threatened Mangabey Taxa of the Eastern Arc Mountains and Coastal Forests of Tanzania and Kenya Biodiversity Hotspot’ (a joint project with T. Butynski). The recordings were also sent to Dr. Jean-Pierre Gautier, the authority on the vocalizations of Cercocebus. Other species whose calls were recorded in the Tana River Primate National Reserve are O. g. lasiotis and G. cocos. Conservation Primate densities in Tana River Primate National Reserve are high. Six primate taxa were encountered and two more are known to occur in the Reserve. Two out of the eight primate taxa are listed as “Endangered” on the 2008 IUCN Red List of Threatened Species (C. galeritus and P. r. rufomitratus) and one is listed as “Vulnerable” (C. m. albotorquatus). The other taxa are listed as “Least Concern” (IUCN, 2008). Habitat loss due to agricultural clearing and extraction of forest products by local communities is the main threat to all primate taxa in the Tana River Primate National Reserve (Butynski & Mwangi, 1994, 1995; Mbora, 2003; Oates et al., 2008; Struhsaker & Grubb, in press). Additionally, there are major changes in the flow volume and flood cycles of the Tana River as a result of the construction of five hydroelectric power dams upriver (Butynski 1995). This is thought to have major impacts on the establishment and survival of the forests of the Lower Tana River. Recent agricultural developments in the area (e.g., two proposed large sugar cane plantations and one proposed large oil palm plantations) will result in a large influx of people to the region with a resultant increased demand for forest products and forest land. The proposed Tana Integrated Sugar Project in Tana River and Lamu Districts alone threatens more than 200 km² of natural habitat. Remarks During this study, we obtained photos and field descriptions for several primate taxa, including C. m. albotorquatus and P. c. ibeanus. Intra group phenotypic variation for P. c. ibeanus here is exceptionally great. Additionally, the phenotype of P. c. ibeanus here varies from that observed at Manda Toto Island, which is, apparently, close to the type locality of the subspecies (which is said to be Lamu Island). Chapter 4.2.8 describes the morphology of P. c. ibeanus in the Kenya coastal forests in more detail. Further research More individual P. c. ibeanus along the Lower Tana River need to be described, photographed, and compared with P. c. ibeanus in the coastal forests to the north and south of the Tana Delta and on the Lamu Archipelago.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 30
Presence-absence data of G. s. braccatus should be collected in or in the vicinity of the Reserve Details about the diurnal road surveys conducted at the north coast of Kenya are presented in Appendix 3 4.1.2 South Coast of Kenya
A. Kaya Rabai (Chijembeni), Mazeras Latitude/longitude: S3.94465 E39.58189 Altitude: 210 m asl Vegetation: Dry coastal forest, coconut palms Cocos nucifera, cashew nut Anacardium occi and mango Mangifera indica common on the forest edge. Agricultural land, human settlements and bush lands surround Kaya Rabai (Chijembeni). Survey date: 20-21 February 2006 Survey details: Nocturnal listening hours and audio recording: 10.3 h Primate species encountered: G. cocos, O. g. lasiotis. C. m. albogularis, C. p. hilgerti, and P. c. ibeanus are reported by residents to be present in the area but were not encountered during this survey. There are reports of C. a. palliates from Kaya Rabai (last report 1945; Anderson, 2007). Encounter rate: individuals G. cocos were heard very often and occasionally seen. Audio recordings: Digital and tape recordings were acquired of advertising calls and other vocalizations of G. cocos and O. g. lasiotis. Recordings were sent for analysis to the Nocturnal Research Group in Oxford. The sonogram and oscillogram of the vocalizations recorded were used to confirm that G. cocos is the name that should be applied to the Kenya coast galago---not Galagoides zanzibaricus (Butynski et al., 2006). Conservation: Kaya Rabai consists of a number of coastal forest fragments (totalling 8 km²) and is part of the Sacred Mijikenda Kaya Forests. In 2008 these forests received UNESCO World Heritage Site status [criteria (iii)(v)(vi); Unesco.org, 2009]. The forests in and around Mazeras are highly fragmented, degraded, and surrounded by agriculture. Some of the forests are sacred and at least somewhat protected. There are five primate taxa in and around these forests. The three forest-depended species (G. cocos, O. g. lasiotis, and C. m. albogularis) are likely declining as a result of habitat degradation and loss. Remarks: Data collected during this survey, together with earlier data from this area, have led to the following publication: Butynski, T.M., De Jong, Y.A., Perkin, A.W., Bearder, S.K. & Honess, P.E. 2006. Taxonomy, distribution, and conservation status of three species of dwarf galagos (Galagoides) in Eastern Africa. Primate Conservation 21: 63-79.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 31
Abstract: This paper reviews the complicated nomenclatural history for the Kenya coast galago, Galagoides cf. cocos, and examines whether ‘cocos’ is the valid species name for this recently resurrected taxon. This paper also reviews the phenotypic and vocal differences among G. cocos; the Zanzibar galago (Galagoides zanzibaricus zanzibaricus); the Udzungwa galago (Galagoides zanzibaricus udzungwensis); and the Mozambique galago (Galagoides granti), as well as their geographic ranges and conservation status. The following are among the findings: (1) Galagoides cocos’ is the name that should be applied to the Kenya coast galago; (2) in the field, the loud calls of these three species are diagnostic and remain the best means for identification; (3) there is a suite of phenotypic characters that, when taken together, can be used to distinguish among these three species when in the hand or viewed in the field in good light at close range; (4) G. z. zanzibaricus is phenotypically distinct from G. z. udzungwensis; (5) the three species are parapatric or, perhaps, narrowly sympatric; (6) the three species are endemic to the coastal forests of eastern Africa with G. cocos in the north (Kenya and northeastern Tanzania), G. zanzibaricus in Tanzania, and G. granti from southern Tanzania to southern Mozambique; and (7) none of the three species is threatened at this time, although G. z. zanzibaricus meets the IUCN Red List criteria for an Endangered subspecies. The complete article is attached in Appendix 6. Further research: The subspecies status of the C. mitis that occurs in Kaya Rabai needs to be determined. B. Diani Latitude/longitude: S4.33461; E39.56251 and S4.2848; E39.5913 Altitude: 0 to ca 20 m asl Vegetation: Gardens with tall exotic and indigenous trees, edge with patches of deciduous coral rag forest. Survey date: 23 April 2008 and 14 December 2008 Survey details: 23 April 2008 Nocturnal surveys: 0.8 km, 3.25 h on foot. Nocturnal listening survey: 2.72 h. Due to bad weather conditions further nocturnal surveys were cancelled 14 December 2008 Opportunistic observations: ca. 4 h. Primate species encountered: P. c. ibeanus, C. m. albogularis, C. a. palliates, C. p. hilgerti, C. p. hilgerti x C. m. albogularis hybrid, O. g. lasiotis, G. cocos. Encounter rates: 23 April 2008
O. g. lasiotis: 6.0 individuals/h, 3.6 individuals/km (n=3) G. cocos: 7.6 individuals/h, 4.8 individuals/km (n=4)

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 32
Audio recordings: The advertisement call and other vocalizations of O. g. lasiotis and G. cocos recorded. Conservation: Originally, Diani was one of the most diverse areas of forest along the Kenya coast with a rich coral rag flora (Rovertson & Luke, 1993 in Bennun & Njoroge, 1999). Six primate species occur in and around Diani and primate densities are high. Diani is a popular tourist destination with many large hotels and houses on large green compounds. All six primate species encountered in Diani were observed on hotel compounds, foraging on flowers, and sleeping in indigenous and exotic tall trees. Most primate species, but in particular P. c. ibeanus, C. m. albogularis, and, C. p. hilgerti, feed on hand-outs and raid tourist facilities and garbage bins for human food items. The destructive effects of tourist development on the natural vegetation is obvious in Diani. The Colobus Trust, an NGO concerned with primate (particular C. a. palliates) conservation along the south coast of Kenya, has established several practical conservation activities to protect the primates and the forests of the area. Coastal forest within the Diani-Chale Marine Reserve are being cleared, fragmented and degraded by big tourist companies and hotels, despite the protected status of the Reserve (colobus.wildlifedirect.org, 2008). Remarks: One adult male C. p. hilgerti x C. m. albogularis hybrid was encountered once (14 December 2008 at 12:45 h) during 4 days of opportunistic surveys. The hybrid was with two adult C. m. albogularis of which at least one was a male. Chapter 4.2.12 presents the details and photos of this hybrid. During a survey of C. a. palliates in 2001, Anderson et al. (2007) found 50 groups comprised of 332 individuals in Diani. This is the second biggest population in Kenya after Shimba Hills National Reserve. The biggest threat to their survival is at Diani is loss of habitat due to tourism activities. Other threats are illegal logging, charcoal production, firewood collection and hunting (Anderson et al., 2007). Further research: More surveys should be conducted in and around Diani in search of additional C. p. hilgerti x C. m. albogularis hybrids. The density, phenotypic characters, ecology and behaviour of these hybrids should be studied. C. Mrima Hill Forest
Latitude/longitude: S4.48714; E39.26224 Altitude: 100-300 m asl Vegetation: Degraded and secondary coastal forest. Road surveys on edge of forest and small agricultural settlements with trees such as mango, cashew, baobab and coconut palm). Survey dates: 7 February 2006, 17:00 - 18:50 h
20 February 2006, 11:00 - 12:02 h

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 33
Survey details: Diurnal surveys: 21.2 km, 2.9 h by vehicle, 10.0 km/h. Primate species encountered: C. m. albogularis and C. a. palliatus Encounter rates: Total of 19 primate groups encountered; 6.6 groups/h, 0.9 groups/km (n=19).
C. m. albogularis: 3.4 groups/h, 0.5 groups/km (n=10). C. a. palliates: 3.1 groups/h, 0.4 groups/km (n=9).
Audio recordings: None Conservation: Mrima Hill Forest is a Forest Reserve, Nature Reserve and National Monument. Despite its protected status, Mrima Hill Forest has been severely degraded due to selective logging and pole cutting and is surrounded by intensively cultivated farmland and a high human population density (e.g. Bennun & Njoroge, 1999). Mrima Hill Forest holds high densities of C. m. albogularis and C. a. palliatus. In 2001, Anderson et al. (2007) counted 13 groups (82 individuals) of C. a. palliatus in Mrima Hill Forest. Illegal logging, charcoal burning and firewood collection are reported to occur only on a low scale by Anderson et al. (2007) and are the only threats to C. a. palliatus in Mrima Hill Forest. This contradicts the information on conservation threats as given by Bennun & Njoroge (1999). Further information on the threats to the Mrima Hill Forest needs to be obtained. Remarks: The area needs to be revisited to conduct nocturnal primate surveys, surveys on foot within the forest (opposed to vehicle surveys from the edge of the forest), and to describe and survey the surrounding area to assess the conservation threats. Further research: - Conduct nocturnal surveys in Mrima Hill Forest - Assess conservation issues that threaten Mrima Hill Forest - The subspecies status of the C. mitis that occurs in Mrima Hill Forest needs to be determined. D. Kaya Sega Latitude/longitude: S4.54612; E39.09309 Altitude: 76 m asl Vegetation: Dense to medium-dense Commiphora bushland surrounded by patches of dry coastal forest. Seasonally dry river bed with tall trees. Survey date: 22 April 2008 Survey details: In total 3.8 h nocturnal surveys Nocturnal survey: 1.0 h, 1.6 km on foot Nocturnal listening hours and audio recording: 2.8 h

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 34
Primate species encountered: O. g. lasiotis Encounter rate: O. g. lasiotis: 1.9 individuals/h, 1.3 individuals/km (n=2) Several individuals were heard, observed, and recorded from camp. Audio recordings: Advertisement call and other vocalizations of O. g. lasiotis recorded from camp. Further research: Diurnal surveys should be conducted to obtain presence/absence and relative abundance figures of the diurnal primates. Details about the diurnal road surveys conducted at the south coast of Kenya are presented in Appendix 4 4.1.3 Outside the coastal forests of Kenya A. Mwea National Reserve Latitude/longitude: S0.80487; E37.59122 Altitude: 1015-1120 m asl Vegetation: Mostly Acacia-Commiphora bush-woodland but some dense, tall riverine vegetation. Survey dates: 3-4 August 2005, 28 April 08 Survey details: 3-4 August 2005 A total of 11.33 h, >26.4 km were surveyed in 1,5 days Diurnal surveys: >15.4 km; 5.63 h by vehicle Nocturnal surveys: 11 km; 1.18 h by vehicle Nocturnal listening hours: 4.52 h 28 April 2008 Diurnal surveys: 40.21 km; 2.95 h by vehicle Primate species encountered: C. mitis, Papio sp., and C. p. hilgerti Encounter rates: 3-4 August 2005 A total of 11 diurnal primate groups were encountered: 1.0 groups/h (n=11) C. mitis: 0.7 groups/h (n=4) C. p. hilgerti: 0.7 groups/h (n=3) Papio: 0.7 groups/h (n=4)

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 35
28 April 2008 Papio: 0.7 groups/h, 0.05 groups/km (n=2) Remark No galagos were encountered or heard in the 5.7 nocturnal hours we surveyed despite what appeared to be very suitable habitat for O. g. lasiotis or G. s. braccatus. Further research Photos and field descriptions of Papio taken in Mwea National Reserve need to be analyzed and compared with photos and field descriptions of P. c. ibeanus and P. anubis to determine if Mwea National Reserve is part of the hybrid zone for Papio. Photos and field descriptions of C. mitis taken in Mwea National Reserve need to be analyzed and compared with photos and field descriptions of C. m. albotorquatus and C. m. kolbi to determine if C. mitis occurring in Mwea National Reserve are intermediates (as or preliminary analysis suggests). Additional nocturnal surveys need to be conducted to determine which nocturnal primates occur in Mwea National Reserve---if any. B. Kiboko Camp, Makindu Latitude/longitude: S2.20346; E37.71383 Altitude: 956 m asl Vegetation: Acacia xanthophloea woodland, medium dense Acacia bushland. Survey dates: 6 February 2006 Survey details: A total of 5.79 h of surveys were completed in one night Nocturnal survey: 2.23 h Nocturnal listening survey: 1.33 h Primate species encountered: O. g. lasiotis and G. s. braccatus. Other primate species observed in this area are Papio and C. p. hilgerti (Y.A. de Jong & T. M. Butynski, pers. obs.). Encounter rates: A total of two nocturnal primates were encountered: 2.22 individuals/h (n=2) O. g. lasiotis: 1.11 individuals/h (n=1) G. s. braccatus: 1.11 individuals/h (n=1) C. Tsavo West National Park Latitude/longitude: S2.68341; E38.17749 (Mtito Andei, head quarters of the National Park)

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 36
Altitude: 700 – 1300 m asl Vegetation: Dense to open Acacia – Commiphora bushland, ground water forests, plains. Survey dates: 19-21 April 08 Survey details: Total of 150.7 km, 9.51 h (7.0 diurnal, 2.5 nocturnal) in 2 days. Diurnal survey: 150.7 km; 7.01 h by vehicle Nocturnal survey: 0.5 km; 0.50 h by foot Nocturnal listening hours: 2.0 h Primate species encountered: C. m. albogularis, C. p. hilgerti, Papio, and G. s. braccatus Encounter rates: A total of 15 diurnal primate groups were encountered: 1.8 groups/h, 0.1 groups/km (n=15). C. m. albogularis: 0.4 groups/h, 0.02 groups/km (n=3) C. p. hilgerti: 0.7 groups/h, 0.03 groups/km (n=5) Papio: 1.0 groups/h, 0.05 groups/km (n=7) G. s. braccatus: 4 individuals/h (n=2) Remarks: Desert warthog P. aethiopicus and common warthogs P. africanus were found sympatric in Tsavo West National Park. Data from this trip were published in Swara (Culverwell et al., 2008. A new pig for Tsavo Swara 31: 50-52) and in Suiform Soundings, the the newsletter of the IUCN/SSC Pigs, Peccaries, and Hippos Specialist Group (De Jong et al., 2009. Desert warthog Phacochoerus aethiopicus found in Tsavo East National Park and Tsavo West Pational Park, southern Kenya, Suiform Soundings 8: 4-6). Both articles are attached in Appendix 7 and 8. D. Tsavo East National Park Latitude/longitude: S3.36731; E38.56476 (Voi, head quarters of the National Park) Altitude: 150 – 1200 m asl Vegetation: Dense to open Acacia – Commiphora savannah bushland, open plains. Survey dates: 26-27 April 08 Survey details: Total survey hours: 10.7 h Diurnal surveys: 135 km; 7.7 h by vehicle Nocturnal listening hours: 3.0 h Primate species encountered: C. p. hilgerti, Papio and O. g. lasiotis

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 37
Encounter rates: Two groups of diurnal primates were encountered, 0.65 groups/h, 0.02 groups/km (n=2)
C. p. hilgerti: 0.3 groups/h, 0.01 groups/km (n=1) Papio: 0.3 groups/h, 0.01 groups/km (n=1)
Audio recordings: none E. North Kilimangodo Latitude/longitude: S4.28659; E39.05576 Altitude: 223 m asl Vegetation: Degraded Acacia bushland surrounded by agriculture and human settlements Survey dates: 21 April 08 Survey details: Nocturnal listening survey: 4 h Primate species encountered: Vocalizations heard of O. g. lasiotis and G. s. braccatus, no primates encountered. Audio recordings: Advertisement call recorded of G. s. braccatus. Details about the diurnal road surveys conducted outside the coastal forests of Kenya are presented in Appendix 5

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 38
4.2 Primates species of the coastal forests of Kenya 4.2.1 Otolemur garnettii lasiotis (Peters, 1876) Vernacular name: White-tailed greater galago Subspecies type locality: Mombasa, Kenya Synonym: hindei Elliot, 1907. Kitui, Kenya Field characters: Large galago with ears of intermediate size (relative to the other subspecies of O. garnettii). Muzzle often darker than rest of face. Inside of ears hairless, pinkish. Dorsum, sides and outer limbs vary from dark brown, brown, brownish-grey, to silvery grey. Ventrum paler than dorsum, varying from brown to grey or off- white. Tail bushy and similar to dorsum in colour as dorsum. Tail tip varies from black to off white. Distribution: Lower Jubba River, Somalia, south to Kenya. Along the Kenya coast inland to Kibwezi and the Taita Hills. South to Tanga, NE Tanzania (Figure 8; Schwarz, 1931; Olson, 1979; Groves, 2001). O. g. lasiotis is the most north-eastern subspecies of O. garnettii. Extent of occurrence: ca. 80,100 km². The area of occupancy is but a very small portion of the extent of occurrence as the habitat suitable for O. g. lasiotis is limited within its geographic range. Figure 8: Approximate geographic distribution of Otolemur garnettii lasiotis in Kenya with sites of encounters (2003-2008) also depicted. Based on Y. de Jong & T. Butynski (unpubl. data) and Butynski et al. (2008a).
Figure 7. Otolemur garnettii lastiotis present in good densities (>15 individuals/ha) in riverine forest in the Tana River Primate National Reserve, Kenya.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 39
Photographic map: Photo map for Galagonidae, contains 28 photographs, of which four are of O. g. lasiotis (T. M. Butynski & Y. A. de Jong). http://picasaweb.google.com/wildsolutions/Galagonidae?feat=email# or via: www.wildsolutions.nl Encounter sites: During this study, O. g. lasiotis was encountered in all six areas we conducted nocturnal surveys in the Coastal Forest of Kenya. Encounter sites in the coastal forests of Kenya and to the Hotspot were; Chyulu Hills National Park; Kiboko Camp, Makindu; Ndololo Camp, Tsavo East National Park; Mazeras; Kaya Sega; north Kilimangodo; Diani; Gedi Ruins; Watamu; Arabuko Sokoke Forest; Malindi; Tana River Primate National Reserve; Kipini Conservancy; NE of Witu Forest; Manda Island. Encounter rate: A total of at least 40 O. g. lasiotis were observed in the coastal forests of Kenya. Twenty-five individuals were observed during survey hours. Hundreds of O. g. lasiotis advertisement calls were heard (Table 2; Figure 8). Table 2. Rates of encounter with Otolemur garnettii lasiotis from 2005 – 2008 in the coastal forests of Kenya. Area Individuals/hour Individuals/km Vehicle/Foot Kaya Sega 1.9 (n=2) 1.3 (n=2) Foot Diani 6.0 (n=3) 3.6 (n=3) Foot Tana River PNR 15 (n=15) - Foot Kipini Conservancy 0.8 (n=1) 0.1 (n=1) Vehicle NE of Witu Forest 4.3 (n=2) 4.0 (n=2) Foot Manda Island 2.0 (n=3) 0.8 (n=3) Foot Vegetation: O. garnettii is a forest-depended species. During this study, O. g. lasiotis was encountered in dry coastal forest, evergreen forest, semi-evergreen riverine forest, secondary forest, fragmented forest, degraded coastal forest, gardens with tall trees, and on the forest edge. Altitude range: Sea level to ca 1300 m asl (Chyulu Hills National Park). Vocalizations: The advertisement call of O. g. lasiotis is a ‘trailing call’; a patterned sequence of units arranged into a phase with a loud beginning and which trails away over the last few units (Bearder et al., 1995; Honess, 1996). Calls are given singly both by males and females and are often replied to by other individuals (also Honess, 1996). Each trailing call comprises six to 10 loud cries reaching a crescendo and then trails off for the last three or four units (also Nash et al., 1989). The frequency of calling varies and depends on the population density (Bearder et al., 2003). The oscillogram and sonogram of the advertisement call of O. g. lasiotis are presented in Figure 9. Figure 9. Sonogram and oscillogram of the trailing advertising call of Otolemur garnettii lasiotis from Diani Beach, 34 km south of Mombasa, SE Kenya, the type locality of this subspecies. Call recorded by Y. de Jong and T. Butynski. A number of advertisement calls were heard in the beginning of

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 40
the evening before heavy weather made it impossible to record. The call follows the typical series of loud cries ending with a trailing off (last four units). For the above recording: call length = 3.9 seconds; frequency range = 0.38–4.41 kHz; fundamental frequency = 0.57 kHz; number of units = 9; unit duration: x= 0.27 sec (range: 0.209-0.360, n=9, sd=0.06). Conservation status: O. g. lasiotis occurs locally in high densities and is able to use secondary, fragmented and/or degraded vegetation. This subspecies occurs in a number of protected areas in Kenya, including the Tana River Primate National Reserve, Tsavo West National Park, Tsavo East National Park, Arabuko Sokoke Forest, Kipini Conservancy, Kaya Rabai, and other Mijikenda Kaya Forests (UNESCO World Heritage site since 2008; Unesco, 2009). No major treats for the species are reported (also IUCN, 2008), however, its fragmented distribution makes it vulnerable to local extinction due to vegetation clearing for agriculture, tourism, etc. (also Harcourt & Perkin, in press). IUCN Red List: Least Concern (Butynski et al., 2008a; www.iucnredlist.org/details/136900). Assessors: T.M. Butynski, S. Bearder, & Y.A. de Jong. Discussion: Otolemur garnettii is a common species in the coastal forests of Kenya. The taxonomy of Otolemur has been debated for many years. This study recognised O. g. lasiotis as the subspecies present in the coastal forests of Kenya. This is consistent with Hill & Meester (1974), Jenkins (1987), Groves (2001, 2005), Grubb et al. (2003), and Harcourt & Perkin (in press). Based on analysis of the geographic variation (phenotypic variation and skull measurements) found in O. garnettii, Olson (1979) considers lasiotis to be a synonym of Otolemur garnettii garnettii [Ogilby, 1838, type specimen from Unguja Island (Zanzibar), Tanzania]. According to Olson (1979), O. g. garnettii occupies the forests of the lower reaches of both the Juba and Webi Shabelle Rivers in Somalia, the lower Tana and Sabaki Rivers in Kenya, and isolated forests in the Chyulu and Taita Hills. In Tanzania, O. g. garnettii inhabits the entire length of the coastal region but is highly discontinuous (Olson, 1979). Jenkins (1987), Groves (2001), Harcourt & Perkin (in press) consider O. g. garnettii to be restricted to Unguja, Pemba, and Mafia Islands. The dorsum of this subspecies is rich reddish-brown and the ventrum is yellow with a slightly greenish tone. Tail brown or greyish-brown with a dark brown or blackish tail tip (Jenkins, 1987; Groves, 2001; Harcourt & Perkin, in press). Olson (1979) accepts that there are phenotypic differences between O. garnettii on Unguja and Pemba with those of the coastal mainland, but considered the differences to be no greater than those occurring between O. garnettii on the two islands. Several authors considered the tail tip in O. garnettii to be one of the diagnostic characters of the subspecies. A pale tail tip (in contrast to the colour of the dorsum and proximal part of the tail) is said to be typical for O. garnettii on the coast of the mainland. However, white tail tips are also found in the island populations (Olson, 1979). On both Pemba and Unguja Islands we only encountered O. g. garnettii with dark tail tips (n=10) during a brief survey of four nights (Butynski & De Jong, unpubl. data). Olson (1979) reports a high percentage of white tail tips in O. garnettii between the Pangani River, Tanzania, and Mombasa, Kenya. During our field studies, however, we observed seven O. garnettii in this region, six of which had a dark brown or blackish tail tip (Kaya Gonja, Kenya, and just a few kilometres south of the Kenya-Tanzania border) and one with an all brown tail (Diani, Kenya). Of all the 40 O. g. lasiotis that we described on the coast of Kenya, two had a whitish or greyish tail tip, 28 had a dark brown-blackish tail tip, and one had an all brown tail (Figure 10). In conclusion, in the coastal forests of Kenya, the colour of the tip of the tail O. g. lasiotis varies from whitish to greyish to blackish. Throughout the geographical range of O.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 41
garnettii in East Africa, outside the Kenya coastal area, we encountered whitish, greyish and blackish tail tips, sometimes at the same site (Butynski & De Jong, unpubl. data). We conclude that the colour of the tip of the tail cannot be used as a character for distinguishing among subspecies of O. garnettii. Figure 10: Colour of the tip of the tail of Otolemur garnettii lasiotis on the coast of Kenya (Y. de Jong & T. Butynski, unpubl. data; Butynski et al., 2008).
In April, 2008, we encountered a captive adult male Otolemur garnettii in Malindi. The individual looked phenotypically different (Figure 11) from any O. g. lasiotis we had observed along the coast of Kenya. The caretaker of this animal informed us that it had been brought in when a juvenile (2.5 years ago). Despite the unsuitable and unhygienic enclosure, the animal seemed healthy.
Figure 11. Captive adult male Otolemur garnettii in Malindi, Kenya. Field characters: Face grizzled silvery grey with a dusky silver wash on the cheeks and around the muzzle. Eye rings absent. Area around the eyes pinkish. Rhinarium dark grey. Area around rhinarium pinkish and sparsely haired. Inside and outside ears hairless and pink. Crown and dorsum silvery grey with mouse grey under fur. Lower back, outer hind legs, and tail dusky silver. Ventrum and inner limbs silvery white. Hands hairless and
pinkish. Finger tips brownish. Ear length ca. 41 mm.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 42
The mean ear length of O. g. lasiotis is 47 mm (range: 43-54 mm for males, n=3; Taita Hills, Kenya; A. Perkin, pers. obs. in Harcourt & Perkin, in press). The mean ear length of O. garnettii is 45 mm (range: 34-55 mm for both sexes combined, n= 356; Olson & Nash, 2002). The ear length of the Malindi Otolemur falls well within this range and is close to the measurements taken by Perkin in the Taita Hills. Olson (1979, p. 315) notes the considerable variation in the pelage colour of O. g. lasiotis and contrasts this to the consistency in colour of the highland subspecies (O. g. kikuyuensis). The National Museums of Kenya holds eight brown specimens collected in the Gedi area (Olson, 1979). Olson (1979) describes O. g. lasiotis as ”typically brown” but found five specimen (n = 29) with a greyish pelage, all collected along the Kenya coast in the vicinity of Gede (= Gedi, Polhill, 1988; ca. 15 km south of Malindi town). Three of those specimen are ”predominantly brown but possess a light greyish wash to the pelage on the sides of the body”. The other two specimen are ”unmistakably grey in colour with only the slightest hind of a brownish wash to the dorsal hairs”. According to Olson (1979), these specimens resemble Galago crassicaudatus monteiri (silver greater galago Otolemur monteiri in current taxonomy; Groves, 2001; Grubb et al., 2003) but the external and cranial dimensions, as well as the texture of the pelage, indicate that they are to O. garnettii. In 1963 (J. Buettner-Janusch & V. Buettner-Janusch, pers. comm. in Olson, 1979) reported high variation in pelage colour in the Sokoke Forest (Gedi area) population of O. garnettii. Olson (1979) suggests that further studies be conducted into the ”nature and source of this exceptionally wide range of variation” in O. garnettii for this part of Kenya. Several nocturnal primate specialists (including Simon Bearder, Leanne Nash, Caroline Harcourt and Andrew Perkin) were contacted during this study in order to identify the Malindi Otolemur. L. Nash (pers. comm.) commented that six out of the 12 O. garnettii that she described from Gedi were ”silvery”, ”grey with brown”, or ”grey-brown”. She analysed the photographs of the Malindi Otolemur and suggested that, on the basis of the face/ear relative proportions, that this is O. garnettii. S. Bearder (pers. comm.) suggested that this individual is an O. garnettii based on “the small ears, the shape of the face and the lack of black on the inside of the ears (which is a characteristic of Otolemur crassicaudatus).” He commented that colour variation is often found in Otolemur and describes this as an ”interesting variation”. On the basis of (1) the considerable colour variation observed for O. g. lasiotis, (2) ear size, (3) communication with nocturnal primate specialists, and (4) its geographical origin, we conclude that the captive Otolemur male observed in Malindi is an O. g. lasiotis. Nielsen and Sick (2008) report the presence of Otolemur crassicaudatus in Witu Forest Reserve. We encountered O. g. lasiotis in the area and strongly doubt the presence of O. crassicaudatus in the Reserve since this species is not known to occur in Kenya (Groves, 2001; T. M. Butynski & Y.A. de Jong, pers. comm.). Photos of Otolemur taken in the Reserve published in the report (Nielsen and Sick, 2008) resemble O. g. lasiotis. Future research priorities:
• Is the Pangani River, Tanzania, the southern boundary of O. g. lasiotis? • To what extent does phenotypic variation in O. g. lasiotis occurs throughout its
geographical range?

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 43
4.2.2 Galago senegalensis braccatus Elliot, 1907 Vernacular name: Kenya lesser galago Subspecies type locality: Tsavo River, near Mt Kilimanjaro, Kenya Synonyms: None Field characters: Medium-sized galago with bony, short-haired tail (Figure 12). Long, thick, and woolly pelage. Back of ears brown or greyish. Front of ears hairless, pinkish. Muzzle pinkish or pale grey. Interocular stripe white. Eye rings dark grey. Sides of the face and lower cheeks off-white or pale grey. Crown, neck and shoulders grey or brownish-grey. Outer arms grizzled rufus-grey. Outer legs rufus with a sharp delineation between the grey upper thigh and dorsum. Ventrum, throat and chest, off-white with yellow wash. Tail grey to brown. Distribution: Highlands of Kenya and Tanzania, east of Eastern (Albertine) Rift Valley. In NE and NC Tanzania, south to Pangani River (T. M. Butynski & Y. A. de Jong, pers. obs.; Nash & Zimmermann, in press). In Tsavo region (Groves, 2001) and coastal area of Kenya (Jenkins, 1987). North to South Horr, Kenya (I. Mwenja, pers. comm.), were it is sympatric with Galago gallarum. The northern limit is unknown (Figure 13). Extent of occurrence: unknown Photographic map: Photo map for Galagonidae, contains 28 photographs, of which seven are of G. s. braccatus (T. M. Butynski & Y. A. de Jong). http://picasaweb.google.com/wildsolutions/Galagonidae?feat=email# or via: www.wildsolutions.nl Encounter sites: During this study, G. s. braccatus was not encountered in the coastal forests of Kenya between 2005 – 2008. Outside the hotspot it was encountered in the Chyulu Hills National Park; Kiboko Camp, Makindu; Komboyo Camp, Tsavo West National Park; north Kilimangodo; Mbuvu. Encounter rate: G. s. braccatus was not encountered in the coastal forests of Kenya. Vegetation: Outside the Hotspot, G. s. braccatus was encountered in dense to open Acacia or Acacia-Commiphora wood/bushland, thorn scrub, and savannah. Altitude range: About 100 – 1900 m asl (Laikipia Plateau, central Kenya; Y. A. de Jong & T. M. Butynski, pers. obs.) Vocalizations: The advertisement call of G. s. braccatus is a medium loud repetitive call comprised of single, low pitched units, repeated at regular intervals. The number of units in a call varies greatly and can last for up to 1 hr (also Harcourt, 1984; Nash et al., 1989; Bearder et al., 1995). The advertisement call is referred to as a ‘woo’ by Zimmerman (1990), a ‘bark’ by Harcourt (1984), but called a ‘honk’ in this study. Calls are given by males and females and made throughout the night with a peak right after dusk and right before dawn.
Figure 12. Galago senegalensis braccatus in Euphorbia tirucalli hedge around human settlement, Isiolo, central Kenya.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 44
Figure 13: Approximate geographic distribution of Galago senegalensis braccatus in the coastal area of Kenya with sites of encounters (2003-2008) depicted (Bearder et al., 2008; Y. de Jong & T. Butynski, unpubl. data). There is not enough data available to produce a more accurate map of the distribution of G. s. braccatus.
Conservation status: G. s. braccatus is most likely patchily distributed in the coastal forests of Kenya. There seem to be no serious threats to the survival of G. s. braccatus in the coastal area of Kenya, where this subspecies is present in low densities. Forest clearing for agriculture, degradation and fragmentation could result in an expansion of dryer bush/woodland and thorn bush which might support more G. s. braccatus in the coastal strip. IUCN Red List: Least Concern (Bearder et al., 2008; www.iucnredlist.org/details/136863). Assessors: S. K. Bearder, T.M. Butynski, & Y.A. de Jong Discussion: The limits of the geographical range of G. s. braccatus are not known. Jenkins (1987) described its range as from S Ethiopia to N and coastal Kenya. Groves (2001) only mentions the Tsavo Region as its range. Nash and Zimmermann (in press) mentions the highlands of Kenya and Tanzania east of the Eastern Rift Valley, south to Pangani River (Tsavo region) as the geographical range of G. s. braccatus. She notes that the northern limit is undetermined. G. s. braccatus is an uncommon species in the Coastal Forest of Kenya. During this study we did not encounter G. s. braccatus in the area. The main reason for the lack of encounters in SE Kenya is most likely due to the few nocturnal surveys we conducted in savannah, thorn bush, and open woodland in SE Kenya. G. s. braccatus does inhabit forests in certain areas but only in forest were other small- to medium-sized galago species are absent (Bearder et al., 2008). Five kilometers across the Kenya – Tanzania border, in NE Tanzania, we encounter eight G. s. braccatus in dense Acacia shrub (Figure 13) within a distance of ca 6 km (rates of encounter: 10.7 ind/h, 1.3 ind/km; travelling by vehicle; T. M. Butynski & Y. A. de Jong, unpubl. data). We

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 45
suspect that G. s. braccatus is present in low densities in the coastal region of Kenya, on the edge with coastal forest and in the semi-arid strip farther inland. G. senegalensis is a widespread species in Kenya. Its range is fragmented but it is locally common and occurs in high densities at many sites. G. s. braccatus was encountered at rates of: 0.1 ind/km (1.0 ind/h) by vehicle in the Ol Pejeta Conservancy, Laikipia Plateau (1,800-1,900 m asl), 1.1 ind/h by foot in Makindu (960 m asl), 1.2 ind/km (1.4 ind/h) by foot in Kajiado, S Kenya (1,500 m asl), and 1.1 ind/km (3.4 ind/h) by vehicle in Kora National Park (500 m asl). At Mpala Ranch, Laikipia Plateau, Off et al. (2008) estimated 0.4 and 2.4 animals/ha. The highest densities were in riverine vegetation, the lowest in dry bush areas. On the Laikipia Plateau, G. s. braccatus densities can locally be as high as 300 to 500 individuals/km² (Y. A. de Jong & T. M. Butynski, pers. comm.). In Tanzania, the encounter rates with G. s. braccatus were 0.1 ind/km (2.4 ind/h) by vehicle in Tanga (10 m asl), 0.9 ind/h by foot in Meia Meia (1,330 m asl), 4.1 individuals/km (3.5 individuals/h) by foot near Mikumi National Park (470 m asl), and 4.0 individuals/h by foot at Mto wa Mbu, Lake Manyara National Park (970 m asl; T. Butynski & Y. de Jong, pers. comm.). In Witu Forest Reserve, the Danish Zoological Society encountered either G. s. braccatus or Galago gallarum (Nielsen & Sick, 2008). During this study, we did not encounter any of the two lesser galagos when we visited Witu Forest Reserve but did find low densities of Galagoides cocos, a species not mentioned in their report. Given the fact that their surveys were conducted in a forest habitat, it is most likely that the species they encountered was either G. cocos (known to inhabit coastal forest) or G. s. braccatus (inhabiting mainly savannah, thorn bush, riverine vegetation, open woodland but occasionally found in forest were no other small- to medium-sized galagos are present; Bearder et al., 2008; Off et al., 2008; Nash, in press). G. gallarum inhabits dry Acacia-Commiphora wood/bushland and thorn bush (Butynski & De Jong, 2004). Dowsett-Lemaire and Dowsett (2007) did not encounter G. s. braccatus on Kipini Conservancy during their surveys in 2006. G. s. braccatus was not encountered in the Tana River Primate National Reserve during this study. This subspecies does, however, occur in this Reserve (Kingdon, 1971; Andrews et al., 1975; Homewood, 1976; Marsh, 1978; Wolfheim, 1983; Decker, 1994; Butynski & Mwangi, 1994; Bearder et al., 2008; T. M. Butynski, pers. obs.). We think that G. s. braccatus is present only in very low densities within this Reserve. Harcourt (1984) reports G. senegalensis in the coastal forest of Kenya but that this species is absent in the coastal forest of Diani were she studied Otolemur garnettii lasiotis and G. cocos (then referred to as Galago zanzibaricus) for 18 months. Future research priorities;
• What is the extent of occurrence or area of occupancy of G. s. braccatus? • Where in the coastal forests of Kenya does G. s. braccatus occur? • How will forest degradation and clearance affect G. s. braccatus?

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 46
4.2.3 Galagoides cocos (Heller, 1912) Vernacular name: Kenya coast galago Species type locality: Mazeras, Kenya. Synonyms: none Field characters: Small, reddish-brown or buffy-brown galago with mouse grey under fur (Figure 14). Ears relatively large (compared to Galago senegalensis and Galago gallarum), dark brown/grey inside. Ear tips slightly bent backwards. Muzzle pointed and turned slightly upwards at tip. Patch on either side of muzzle dark brown and prominent. Chin pinkish-grey. Nose stripe off white, extending to slightly above eyes. Eye rings grey. Throat, cheeks and ventrum off white to pale grey. Limbs, hands and feet buffy-brown. Dorsum buffy-brown. Proximal ca. 25% of tail same color as dorsum (i.e., buffy-brown) becoming gradually dark brown towards the tip of the tail (Butynski et al., 2006). Distribution: G. cocos is restricted to the coastal forests of Kenya and N Tanzania. Occurs along the coastal strip of Kenya, as far north as the Tana River Primate National Reserve, Kipini Conservancy (on the coast just north of the Tana Delta) and Manda Island southward to at least the Mgambo Forest Reserve in N Tanzania at the north end of the East Usambara Mountains (between 1º 43' and 4 º 49' S; Butynski et al., 2006; Figure 15). Reported to occur in S Somalia along the Webi Shabeelle River (Nash et al., 1989) but this requires confirmation. Extent of occurrence: ca. 13 900 km². Area of occupancy is very much less than this. Photographic map: Photo map for Galagonidae, contains 28 photographs, of which three are of G. cocos (T. M. Butynski & Y. A. de Jong). http://picasaweb.google.com/wildsolutions/Galagonidae?feat=email# or via: www.wildsolutions.nl Encounter sites: During this study, G. cocos was encountered in five out of the six areas we conducted nocturnal surveys in the coastal forests of Kenya; Kaya Rabai, Mazeras; Tana River Primate National Reserve; Kipini Conservancy; Manda Island; Diani.
Figure 14. Galagoides cocos in Acacia – baobab woodland at Takwa Ruins, Manda Island, Kenya.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 47
Figure 15: Known distribution and encounters (2003-2008) with Galagoides cocos in Kenya. The other source used for this map is Butynski et al. (2006).
Encounter rate: During this study, at least 20 G. cocos were encountered (excluding Kaya Rabai, Mazeras). All encounters were within the coastal forests of Kenya (Table 3; Figure 15). Table 3. Rates of encounter with Galagoides cocos during 2005 – 2008 in the coastal forests of Kenya. Area Individuals/hour Individuals/km Vehicle/Foot Note Kaya Rabai, Mazeras
- - Foot No counts conducted, many individuals heard and seen.
Tana River PNR 10 (n=10) Foot About 15 individuals/ha
Kipini Conser-vancy
0 0 Vocalisations heard only
Manda Island 4.1 (n=6) 1.6 (n=6) Foot Diani 7.6 (n=4) 4.8 (n=4) Foot Vegetation: G. cocos was encountered in evergreen and semi-evergreen riverine forest, dry coastal forest, fragmented, disturbed and degraded coastal vegetation, secondary forest, gardens and mosaics of mixed agriculture were some natural forest remains. Altitude range: Sea level to at least 350 m asl in the foothills of the East Usambara Mountains, Tanzania (Butynski et al., 2006).

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 48
Vocalizations: The advertisement call of G. cocos is an incremental call which is described as “A patterned phrase in which the units are arranged into distinct groups or ‘unit sequences’ which tend to get longer as the call progresses. Calls are given singly.” (Bearder et al., 1995). We describe the advertisement call of G. cocos in Butynski et al., (2006) as, “starts with a series of high-pitched, rapidly uttered, ’chirrups’ followed by units arranged in phrases that are high in frequency and amplitude, and that gradually become lower in amplitude. The number of units within each phrase increases incrementally until the end of the call. Often, as in this case, phrases with same number of units are repeated. The number of units per phrase rarely decreases. Units are often frequency modulated”. Butynski et al. (2006) presents the sonogram and oscillogram of G. cocos recorded in Diani, Kenya. Conservation status: G. cocos is the most abundant and widespread galago in the coastal forests of Kenya, with densities of approximately 170 –180 individuals/km² both at Gedi and Diani (Harcourt & Nash, 1986). G. cocos is able to persist in fragmented, disturbed and degraded coastal vegetation, secondary forest and mosaics of mixed agriculture were some natural vegetation remains. There is a need to better determine the edges of the geographical range of G. cocos along the coast of N Kenya, S Somalia and in the vicinity of Nairobi (Butynski et al., 2006). IUCN Red List: Least Concern (Butynski et al., 2008b; www.iucnredlist.org/details/136212). Assessors: T.M. Butynski, A. Perkin, S. Bearder, Y.A. de Jong, & P. Honess. G. cocos was only recently resurrected to a full species (Grubb et al., 2003; Butynski et al., 2006). Previously, ttaxonomists considered G. cocos as a subspecies or synonym of Galagoides zanzibaricus or Galago zanzibaricus (Butynski et al., 2006; Appendix 6). Discussion: Data on G. cocos, partly obtained during this survey, were published in Butynski et al. (2006). The abstract of this article is given below and the full article is presented in Appendix 6). This publication includes a map and presents a thorough review of the geographical range of G. cocos. After publication of the map, this study found two more localities for the species; Kipini Conservancy and Manda Island. The Manda Island locality extended the range 70 km to the east of the distribution shown in Butynski et al. (2006; Figure 15). Primate Conservation (2006) 21: 63 –79 Taxonomy, Distribution, and Conservation Status of Three Species of Dwarf Galagos (Galagoides) in Eastern Africa. Thomas M. Butynski, Yvonne A. de Jong, Andrew W. Perkin, Simon K. Bearder, and Paul E. Honess Abstract: This paper reviews the complicated nomenclatural history for the Kenya coast galago, Galagoides cf. cocos, and examines whether ‘cocos’ is the valid species name for this recently resurrected taxon. This paper also reviews the phenotypic and vocal differences among G. cocos; the Zanzibar galago (Galagoides zanzibaricus zanzibaricus); the Udzungwa galago (Galagoides zanzibaricus udzungwensis); and the Mozambique galago (Galagoides granti), as well as their geographic ranges and conservation status. The following are among the findings: (1) ‘Galagoides cocos’ is the name that should be applied to the Kenya coast galago; (2) in the field, the loud calls of these three species are diagnostic and remain the best means for identification; (3) there is a suite of phenotypic characters that, when taken together, can be used to distinguish among these three species when in the hand or viewed in the field in good light at close range; (4) G. z. zanzibaricus is phenotypically distinct from G. z. udzungwensis; (5) the three species are parapatric or, perhaps, narrowly sympatric; (6) the three species are endemic to the coastal forests of eastern Africa with

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 49
G. cocos in the north (Kenya and northeastern Tanzania), G. zanzibaricus in Tanzania, and G. granti from southern Tanzania to southern Mozambique; and (7) none of the three species is threatened at this time, although G. z. zanzibaricus meets the IUCN Red List criteria for an Endangered subspecies. The complete article can be found in Appendix 6. Future research priorities:
• Does G. cocos occur in Ololua Forest, Nairobi? • How for north, north west does G. cocos along the Tana River?

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 50
4.2.4 Cercopithecus mitis albogularis (Sykes, 1831) Vernacular name: Zanzibar Sykes’s monkey Species type locality: Zanzibar Island Synonyms: kibonotensis Lönnberg, 1908; maritime Heller, 1913; kima, Heller 1913 Field characters: Medium-size monkey. Face dark grizzled grey with hairless face mask (Figure 16). Eyes reddish-brown. Ear fringe narrow, pale grey. Brow band grizzled grey or grizzled grey-olive, directed forward and upward. Crown grizzled olive-grey with pale rufus wash. Cheeks grizzled pale grey, grizzled grey or grizzled olive-grey. Collar varying from pale grey to bright white, 55 - 80% complete. Collar line in Kenya populations well defined and extends from upper jaw, across the lower cheeks to ca 4-5 cm below the ears then down onto the sides of the neck, across the top of the shoulders to the nipples. Lower shoulders and upper outer arms dark grey. Lower outer arms black. Inner arms varying from grey to black. Outer legs dark grey, inner legs varying from pale grey (Kenya coast) to dark grey (Tanzania). Back of the upper legs varying from grizzled grey (Tanzania) to pale grey with rusty wash. Rusty wash most intense towards base of the tail in Kenya coast population. Dorsum grizzled grey between shoulders. Behind shoulders grizzled rusty red extending onto the upper tail. Ventrum varying from blackish or dark grey (Tanzania) to pale grey to off white (Kenya coast). Tail blackish or black. Base of the tail rusty all around. Distribution: From Gedi Ruins (central coast of Kenya), southwards along the coast to N Tanzania (including Unguja and Mafia Islands), west to Mount Kilimanjaro, Mount Meru, Tsavo West National Park, Chyulu Hills National Park, Taita Hills, and Mount Kasigau. Along the coast of Tanzania as far south as NE Mozambique (Figure 17). Extent of occurrence: ca. 209,000 km². The area of occupancy is but a very small portion of the extent of occurrence as the habitat suitable for C. m. albogularis is limited within its geographic range. Photographic map: Photo map of Cercopithecus mitis, contains 46 photographs, of which 15 in the coastal forests of Kenya (T. M. Butynski & Y. A. de Jong). http://picasaweb.google.com/wildsolutions/CercopithecusMitis?feat=email# or via: www.wildsolutions.nl
Figure 16. Cercopithecus mitis albogularis adult male at Diani, Kenya.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 51
Figure 17. Approximate geographic distribution of Cercopithecus mitis albogularis in the coastal area of Kenya with sites of encounters (2003-2008) also depicted. Based on Y. de Jong & T. Butynski (unpubl. data) and Kingdon et al. (2008c).
Encounter sites:
- Gedi Ruins (354, 355; Appendix 2) - Diani (346, 348, 350, 352; Appendix 2) - Gazi (185; Appendix 2) - Buda Forest (345; Appendix 2) - Mrima Hill (187, 190, 191, 194, 195, 197, 198, 248; Appendix 2) - Kaya Sega (343, Appendix 2) - Lunga Lunga (341, Appendix 2) - Vanga (342; Appendix 2) - Kaya Rabai (no number, Appendix 2) - Tsavo West National Park (330, 333, 338; Appendix 2)
Encounter rate: Twenty-two groups of C. m. albogularis were encountered in the coastal forests of Kenya. Four groups were encountered in Tsavo West National Park and Mazeras while travelling to the Hotspot (Table 4; Figure 17).

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 52
Table 4. Encounter rates (groups/hour and groups/km) of Cercopithecus mitis albogularis during 2005 – 2008 in the coastal forests of Kenya and outside the Hotspot. This table only includes those surveys during which C. m. albogularis was encountered (n=number of groups). Area Groups/hour Groups/km Vehicle/Foot Note Gedi Ruins One group seen
twice at the ruins Foot
Mrima Hill 3.4 (n=10) 0.5 (n=10) Vehicle Using road around forest edge.
Kaya Rabai, Mazeras 0 0 Foot Vocalisations heard
Tsavo West NP 0.4 (n=3) 0.02 (n=3) Vehicle Road Surveys Kiboko to Lunga Lunga (excluding Mrima Hill)
0 0 Vehicle Highway
Lunga Lunga to Mazeras (including Shimba Hills NR, excluding Mrima Hill)
0 0 Vehicle Main road and bush tracks
Mtito Andei to Kilimangodo North
0 0 Vehicle Bush track
Kilimangodo North to Kaya Sega (Vanga, Lunga Lunga, Kaya Gonja, Godo)
0.6 (n=3) 0.03 (n=3) Vehicle Using main roads
Kaya Sega to Diani (Vanga, Kaya Dzombo, Kaya Gonja, Buda Forest)
0.5 (n=4) 0.02 (n=4) Vehicle Using tracks and main roads. Three encounters in Diani town.
Diani, Tiwi to Nyali 0.4 (n=1) 0.02 (n=1) Vehicle Using main roads Nyali to Watamu 0.3 (n=1) 0.01 (n=1) Vehicle Using main roads Watamu to Mamburi (including Malindi)
0.2 (n=1) 0.01 (n=1) Vehicle Using main roads
Watamu to Tsavo East NP (including Malindi, Arabuko Sokoke Forest, Sala Gate Tsavo East NP)
0 0 Vehicle
Vegetation: C. m. albogularis is primarily a forest species and was encountered in coastal forest that varied from dry to moist coastal forest. This taxon is also encountered in miombo woodland, submontane, riverine, groundwater, secondary and plantation forests. Within populated coastal areas present on forest edged, in gardens, and in small patches of degraded coastal forest. Altitude range: 0 – 2,900 m asl (Mount Kilimanjaro, Tanzania; T. M. Butynski & J. M. Butynski, pers. comm.). Conservation status: C. m. albogularis is widespread and common in suitable habitat. Major threats to the survival of this taxa are habitat loss, degradation and fragmentation throughout those parts of its range that are outside protected areas.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 53
IUCN Red List: Cercopithecus mitis albogularis: Least Concern (Kingdon et al., 2008a; www.iucnredlist.org/details/40645) Assessors: Kingdon, J., Butynski, T.M. & De Jong, Y.A. Discussion: The taxonomy of Cercopithecus mitis (or Cercopithecus albogularis) has been debated for many years and is still not agreed on. In this study we consider Cercopithecus mitis albogularis to be the subspecies that occurs in the coastal forests of Kenya (Booth, 1963; Grubb, et al., 2003, Grubb, 2006; Kingdon et al., 2008a, and Lawes et al., in press). We consider C. m. kibonotensis to be a synonym of C. m. albogularis (contra Hill, 1966; Dandelot, 1968, 1974; Rahm, 1970;, Napier, 1981; Lernould, 1988; Kingdon, 1997; Groves, 2001) who accept C. m. kibonotensis as the name of the mainland subspecies (distribution: southern Kenya coast south of Kilifi Creek, and northern Tanzania inland to Mt Kilimanjaro and Mt Meru) and C. m. albogularis to be the island (Unguja & Mafia) equivalent, with the main diagnostics characters being the larger body size of C. m. kibonotensis and the banding of the hairs (Groves, 2001, 2006). Grubb (2006) analysed the body sizes of both subspecies and did not find significant differences---but the sample size was small. Booth (1968, p.46) considers all subspecies of C. m. albogularis east of the rift valley as one subspecies because there are no distinguishing differences between the forms/subspecies; there is a cline going from the Kenya highlands to Zanzibar. For this reason, she claims that kolbi, kibonotensis and monoides are synonyms of C. m. albogularis. Booth (1963) makes no comment on the status of C. m. albotorquatus. Table 5 present field descriptions obtained at five localities. The main phenotypic differences within C. m. albogularis is the colour of the ventrum and inner arms and legs which ranges from pale grey in the NE of the geographic range to blackish or dark grey in the Tanzania animals. The cheeks of the Kenya animals are more slender than for the Tanzania animals. The white collar of the Kenya animals is 60 - 80% complete, compared to 55-60% complete in the Tanzania population. Compared to the Tanzanian animals, the collar of the Kenya animals is more sharpely demarcated, runs through the cheeks, and closer to the ear. Unlike the Tanzanian animals, those in Kenya have red wash on the back of the upper hind legs. Table 5. Field descriptions of C. m. albogularis Unguja Island, Tanzania, C. m. albogularis Usa River, Tanzania, C. m. albogularis Mrima Hill, Kenya, Cercopithecus mitis albogularis Diani, Kenya and C. m. albogularis Gedi Ruins, Kenya. Variable C. m. albogularis
Unguja Island, Tanzania
C. m. albogularis Usa River, Tanzania
C. m. albogularis Mrima Hill, Kenya
Cercopithecus mitis albogularis Diani, Kenya
C. m. albogularis Gedi Ruins, Kenya
Altitude (m asl)
20 1195 80 16 20
Photo
Ears Narrow, pale grey, fringe
Pale grey tufts Narrow, pale grey, fringe
Narrow, pale grey, fringe (some whitish)
Fringe absent or narrow, pale grey.
Face Grizzled pale grey. Grizzled pale grey. Grizzled grey. Bare Grizzled grey. Bare Grizzled grey.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 54
Bare black skin around eyes.
Bare black skin around eyes
black skin around eyes.
black skin around eyes.
Bare black skin around eyes.
Cheeks Whiskers full, grizzled pale grey. No collar line crossing the lower cheeks.
Whiskers full, grizzled olive-grey. No collar line crossing the lower cheeks.
Whiskers not full, grey. Collar line through the cheeks.
Whiskers not full, grey. Collar line through the cheeks.
Whiskers not full, grey. Collar line through the cheeks.
Brow band Projects forward and upwards. Grizzled pale grey.
Projects forward and upwards. Grizzled olive-pale grey.
Projects forward and upwards. Grizzled grey.
Projects forward and upwards. Grizzled grey.
Projects forward and upwards. Grizzled grey.
Crown Grizzled olive-grey with rufus wash.
Grizzled olive-grey with rufus wash.
Grizzled olive-grey with pale rufus wash.
Grizzled olive-grey with pale rufus wash.
Grizzled olive-grey.
Collar Whitish-pale grey, ca 55-60% complete.
Absent. White throat patch present.
White, ca 75-80% complete. Collar line sharp, running from upper jaw, across cheeks ca 4 cm below the ears, onto neck, down the shoulders to include the throat and upper chest.
Not sharply defined. Around lower ca. 60% of neck. Throat off-white. Sides of neck light grey.
White, ca 75% complete. Collar line sharp, running from upper jaw, across cheeks to ca 4 cm below the ears onto neck, down shoulders, then grading into pale grey ventrum.
Throat Whitish White White Off white White Shoulders Grizzled dark grey Black Dark grey Grizzled dark grey Dark grey Outer arms
Black Black Black Black Black
Inner arms Dark grey Black Dark grey Blackish Dark grey Outer legs Grizzled grey, feet
blackish Dark grey Dark grey Grizzled grey Dark grey
Inner legs Grizzled grey Dark grey Upper part white, lower part dark grey
Pale grey Pale grey
Back of hind legs
Grizzled grey Grizzled grey Pale grey with rufus wash
Off white with rufus wash
Pale grey with rufus wash
Dorsum Grizzled grey between shoulders. Distal half rich red extending onto upper tail.
Grizzled olive-grey between shoulders. Behind shoulders rich red extending onto upper tail.
Grizzled grey between shoulders. Behind shoulders grizzled rusty red extending onto upper tail.
Grizzled grey between shoulders. Behind shoulders grizzled rusty red extending onto upper tail.
Grizzled grey between shoulders. Behind shoulders grizzled rusty red extending onto upper tail.
Ventrum Grizzled dark grey. Blackish Grey Grey. Center of the ventrum pale grey.
Pale grey
Sides Grizzled reddish-grey
Grizzled reddish-grey
Grizzled grey Grizzled grey Grizzled grey
Tail Blackish except pale red around base.
Blackish except rich red around base.
Blackish except bright rufus around base.
Black except bright rufus around base.
Blackish, except bright rufus around base.
Diagnostic Hairless face mask, Collar absent, Grey ventrum, Grey to pale grey Off white to pale

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 55
Heller (1913) provides a description of a specimen of C. m. albogularis from Mazeras (then referred to as Lasiopyga albogularis maritima) which notes the lighter coloration overall and the absence of a black lining or black tips to the hairs of the ventrum. Although we did not observe C. m. albogularis well enough in Mazeras to make a complete field description, the C. m. albogularis populations we found on the Kenya coast certainly have a pale grey or off-white ventrum. Groves (2001) notes in his description of C. a. albogularis and C. a. kibonotensis (which he accepts as a full subspecies of C. albogularis), that C. a. kibonotensis is very similar to C. a. albogularis (which he considers to be endemic to Unguja Island) except that C. a. albogularis has an almost complete collar, and the collar of C. a. kibonotensis is less extensive. Napier (1981) also describes C. m. kibonotensis with a less extensive collar compared to C. m. albogularis. However, during this study (which is based on field descriptions and photographs as opposed to the museum descriptions of Napier (1981) and Groves (2001), we found the opposite; the animals on Unguja Island have a whitish (males) or pale grey (juveniles and females) collar that is ca 55-60% complete; Figure 18) where as the animals on the Kenya coast have a collar with is ca 60-75% complete (white in males and off white or pale grey in juveniles and some females). In addition, the Kenya coast animals have a collar which forms a well demarcated line from the upper jaw, across the cheeks, under the ears onto the neck, and crosses the top of the shoulders down onto the chest (Figure 19).
characters dark grey ventrum, rich red dorsum, full cheek whiskers.
blackish ventrum, rich red dorsum, full cheek whiskers.
white upper inner legs.
ventrum grey ventrum.
Figure 18. Adult male C. m. albogularis at Jozani-Chwaka Bay National Park, Unguja Island, Tanzania. Note the grizzled grey cheeks and the whitish to pale grey collar which is ca 60% complete.
Figure 19. Adult male C. m. albogularis at Gedi Ruins, Kenya. Note that the white of the collar crosses the cheek and that the collar is >70% complete.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 56
In conclusion, we observed considerable phenotypic differences among the animals found in the type locality for C. m albogularis (Unguja Island, Tanzania), the western-most locality for ‘C. m. albogularis’ that we visited (Usa River, close to the type locality of kibonotensis), the northeastern-most locality that we visited (Gedi Ruins), and those in between these localities (e.g., Mrima Hill, Vanga, Diani, Lunga Lunga). These phenotypic differences appear to be in a cline. The phenotypic differences between the extremes of the geographic range of C. mitis from Gedi south to Unguja Island differ in degrees and seem to be part of the natural occurring variability within C. m. albogularis. Future research priorities:
• Which subspecies of C. mitis occurs between the Tana River and the Galana River?

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 57
4.2.5 Cercopithecus mitis albotorquatus Pousargues, 1896 Vernacular name: Pousargues’s white-collared monkey Subspecies type locality: not known (zoo animal) Synonyms: phylax Schwarz, 1927, Patta Island, Lamu Archipelago rufotinctus Pocock, 1907, “British East Africa (? Mombasa).” Field characters: Medium-size monkey (Figure 20). Appears to be slender as compared to the other subspecies of C. mitis. Face dark grizzled grey, slender, with short hair. Eyes reddish-brown. Ears visible with narrow grey fringe. Browband grizzled greyish-olive, directed far forward. Upper jaw dark grey. Collar bright white and almost (ca. 85%) complete Collar line very well defined and extends from upper jaw, across the cheeks to ca 2 cm below the ears then down onto the sides of the neck, across the top of the shoulders to the nipples. Crown grizzled greyish-olive with rufus teint. Lower shoulders and upper outer arms grizzled dark grey. Lower outer arms dark grey to blackish. Outer legs grey. Ventrum and inner limbs pale grey to off-white. Back of upper legs with long rusty hairs, most intense red towards the base of the tail. Dorsum grizzled grey between the shoulders becoming increasingly grizzled olive-russet towards the tail. Tail blackish. Base of the tail rusty all around. Distribution: Restricted to the forest along the north coast of Kenya and inland along the lower Tana River into Meru National Park. Perhaps along the coast in extreme Southern Somali but this needs confirmation (de Beaux, 1937; Hill, 1966; Gartland & Brain, 1968; Rahm, 1970; Gippoliti, 2002). Present on the Lamu Archipelago and patchily distributed throughout the Tana River forests as northwest as Meru National Park (Figure 21; De Jong & Butynski, 2004; Butynski & De Jong, 2008). Not known to occur south of the Tana River or Tana Delta where it appears to be replaced by C. m. albogularis. Along the Juba River and Shebelli River of S. Somalia it is replaced by C. m. zammaranoi. Extent of occurrence: ca. 11 500 km². Area of occupancy is very much less than this due to the severely fragmented geographic range and limited extent of forest in this semi-arid region.
Figure 20. Cercopithecus mitis albotorquatus male from Witu Forest, Kenya, where this subspecies is common.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 58
Figure 21. Known distribution and encounters (2003-2008) with Cercopithecus mitis albotorquatus. The other source used for this map is Butynski & De Jong (2008).
Photographic map: Photo map for Cercopithecus mitis, contains 46 photographs, of which five are of C. m. albotorquatus (T. M. Butynski & Y. A. de Jong). http://picasaweb.google.com/wildsolutions/CercopithecusMitis?feat=email# or via: www.wildsolutions.nl Encounter sites: During this study, C. m. albotorquatus was encountered in three out of the four areas we conducted diurnal surveys in the coastal forests of Kenya; - Tana River Primate National Reserve (17, 17A, 18A, 254, 254A, 255A; Appendix 2) - Witu Forest Reserve (269, 271, 271A; Appendix 2) - Kipini Conservancy (277A, 273, 278A to 278E, Appendix 2) - Takwa Ruins, Manda Island (367, Appendix 2) - Lamu Island (heard but not seen) Encounter rate: During this study, 16 groups of C. m. albotorquatus were encountered all in the coastal forests of Kenya (north coast; Table 6; Figure 17).

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 59
Table 6. Encounter rates (groups/h and groups/km) of Cercopithecus mitis albotorquatus during 2005 – 2008 in the Coastal Forests Kenya and while driving to the Hotspot. Only includes the surveys in which C. m. albotorquatus was encountered. Area Groups/hour Groups/km Vehicle/Foot Note Tana River PNR 0.30, n=6 Foot Witu Forest Reserve
7.14, n=3 0.6 n=3 Vehicle Using existing roads
Kipini Conservancy
1.60, n=5 1.2, n=5 Foot Vehicle surveys not included
Manda Island 0.16, n=1 0.1, n=1 Foot Calls heard from at least one other group
Lamu Island 0 0 Foot Calls heard from at least one group.
Vegetation: Evergreen and semi-evergreen riverine forests of the Tana River, surrounded by semi-arid medium-dense bush land on sandy soil (Tana River floodplain). Forest-grassland mosaic, coastal forest and dry coastal shrub. Mangrove forest. Altitude range: 25 – 400 m asl (De Jong & Butynski, 2004) Conservation status: C. m. albotorquatus is a poorly known subspecies. Threats to their conservation are habitat loss, degradation and fragmentation due to forest clearance and agriculture.
During this survey, C. m. albotorquatus were found to be common in the Witu Forest Reserve, Kipini Conservancy, and Tana River Primate National Reserve, but rare on Manda Island and on Lamu Island. Said to be present on Patta Island but we did not visit Patta Island. During primate surveys in Meru National Park and Kora National Park in 2003, C. m. albotorquatus were found to be common in the riverine forests of the Tana River (De Jong & Butynski, 2004). In short, during our study we found C. m. albotorquatus to be much more widespread than previously known and also that there are some sites where this subspecies is at high densities. These significant findings bode well for the long-term conservation and survival of C. m. albotorquatus.
IUCN Red List: Cercopithecus mitis albotorquatus: Vulnerable B1ab (i,ii,iii,v; Butynski & De Jong, 2008; www.iucnredlist.org/details/39990). Assessors: T. M. Butynski, & Y.A. de Jong Previous assessment: Data Deficient (DD; ver 2.3, 1994), year assessed, 2000 (Butynski & De Jong, 2008) Discussion: C. m. albotorquatus is a relatively poorly-known but distinctive subspecies that is , recognised as a subspecies by most primate taxonomist (Pocock, 1907; Hill, 1966; Rahm, 1970; Kingdon, 1971; Dandelot, 1974; Napier, 1981; Lernould, 1988; Kingdon, 1997; Groves, 2001, Grubb, 2001, Grubb et al., 2003). Several authors suggested that this subspecies was confined to the forests along the lower Tana River forests (Gartlan & Brain, 1968; Rahm, 1970; Kingdon, 1971; Andrews et al., 1975; Napier, 1981; Lernould, 1988; Groves, 2001). During a primate survey in 2003 by T. Butynski & Y. de Jong, this subspecies was found to be present and common as far inland as Meru National Park and Kora National Park. This is much farther inland (to the northwest) of the previous known range (De Jong & Butynski, 2004). This finding extended the known range ca 100 km to the west (Saka being the closest record; Napier, 1981).

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 60
The southern limit of the geographical range of C. m. albotorquatus is uncertain. Figure 21 shows a gap of 80 km between the ranges of C. m. albotorquatus and C. m. albogularis, based on the lack of reports from this area. The closest record for C. m. albotorquatus (Witu Forest Reserve) to C. m albogularis is for the Gedi Ruins (108 km SW of Witu Forest Reserve). We find, however, that the ‘C. m. albogularis’ at Gedi Ruins are phenotypically different from C. m. albotorquatus and from the description of the holotype for C. m. albogularis (which was collected on Zanzibar). Table 7 presents the field descriptions we obtained of C. m. albogularis from Gedi Ruins, Kenya, C. m. albotorquatus from Witu Forest Reserve, Kenya and C. m. albogularis from Unguja Island, Tanzania (type locality of C. m. albogularis). Table 7. Field descriptions of C. m. albogularis, Gedi Ruins, Kenya, C. m. albotorquatus, Witu Forest Reserve, Kenya and C. m. albogularis, Unguja Island, Tanzania. Description C. m. albogularis, Unguja
Island, Tanzania C. m. albogularis, Gedi RuinsKenya
C. m. albotorquatus, Witu Forest Reserve, Kenya
Altitude 20 m asl 20 m asl Foto
Ears Narrow pale grey fringe Absent or narrow pale
grey fringe. Narrow fringe, grey.
Face Grizzled pale grey. Bare black skin around eyes.
Grizzled grey. Bare black skin around eyes.
Triangular shape with short hair. Grizzled grey. Bare black skin around eyes.
Cheeks Full, grizzled pale grey. Grizzled grey, slender appearance. Collar line crossing the lower cheeks
Grizzled grey, slender appearance. Collar line crossing the lower cheeks
Brow band Projecting forward and upwards.Grizzled pale grey
Projecting forward and upwards. Grizzled grey.
Projects forward and upwards. Grizzled greyish-olive
Crown Grizzled olive-grey with rufus teint.
Grizzled olive-grey. Grizzled olive-grey with rufus teint..
Collar Whitish-pale grey, ca 55-60% complete.
Collar white, ca 75% completeCollar line sharp, running from upper jaw, across the cheeks to ca 4 cm below the ears into the neck, down crossing the top of the shoulders gradinginto the pale ventrum.
Bright white, almost (ca. 85%) complete. Collar line very well defined, extends from upper jaw, across the cheeks to ca 2 cm below the ears then down onto the sides of the neck, across the top of the shoulders to the nipples.
Throat Whitish White Bright white Shoulders Grizzled dark grey Dark grey Partly white from collar,
remainder grizzled grey

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 61
Outer arms Black Black Outer arms dark grey to blackish.
Inner arms Dark grey Dark grey Pale grey Outer legs Grizzled grey, feet
blackish Dark grey. Grey.
Inner legs Grizzled grey Pale grey Pale grey to off white. Back of hind limbs
Grizzled grey. Pale grey with rufus wash Long, off white, hairs with distinguishing rusty wash. Rust darkest around base of tail.
Dorsum Grizzled grey between shoulders. Half way the back onto the upper tail rich red.
Grizzled grey between shoulders. Below shoulders grizzled rusty red extending onto the upper tail.
Grizzled grey between shoulders. Behind shouldersgrizzled olive-grey-rusty becoming increasingly rusty towards the tail.
Ventrum Grizzled dark grey. Pale grey Pale grey to off white. Sides Grizzled reddish-grey Grizzled grey Grizzled grey Tail Pale red around base of
tail, rest of tail blackish. Bright rufus around the base ofthe tail, rest of blackish
Intense red around base of tail (especially ventrally). Terminal part blackish.
Overall The hairless face mask noticable, dark grey ventrum, rich red dorsum and full cheeks are distinguishable for this population
The off white-pale grey ventrum is distinguishable for this population
The short hair, triangular face and the bright and wide collar that is almost completeis distinguish-able for this subspecies.
C. mitis in the Lamu Archipelago The type locality of C. m. phylax is Patta Island. The distribution of C. m. phylax is the Lamu Archipelago (“Patta and Witu Islands.” (=Lamu Archipelago; Groves, 2001), however, Hill (1966) notes that this taxa is only known from the type locality (Patta Island). We found no published locality records of C. mitis for Manda and Lamu Islands to clarify this inconsistency. During a survey on Lamu Island, the density of primates was found to be very low. The observed primate groups were small and the animals were shy. We collected evidence that C. mitis is present in very low densities in the extensive mangrove forests on N Lamu Island (ca 150-200 m away from the mainland). Calls were heard from at least one group in the mangrove forest but we failed see these monkeys during our 2 day visit (mainly due to bad weather). The local people on this part of the Island are familiar with C. mitis and, we were told, they occasionally hunt and keep them as pets. During a survey on Manda Island, the density of C. mitis was found to be very low. As on Lamu, this is likely due to the lack of year round fresh water. On one occasion we heard C. mitis vocalizations (including ‘pyows’) from the mangrove forest during a boat survey on NW Manda Island. On SE Manda Island we encountered one group of C. mitis (and heard ‘pyows’). One individual was seen during our ground survey. The animal was in dense and diverse coastal shrub (including Acacia, Commiphora, and baobab) on coral rag on the edge of a mangrove forest. People here report that C. mitis to occur mainly in the mangrove forests. If Hill (1966) is correct in that phylax is restricted to Patta Island than this survey is the first to confirm that C. mitis is present both on Lamu Island and Manda Island.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 62
Hill (1966), Rahm (1970), Kingdon (1971), Napier (1981), Lernould (1988), Kingdon (1997) and Groves (2001) consider phylax to be valid subspecies. Dandelot (1968) considers C. m. phylax as a synonym of C. m. albotorquatus on the basis of the ‘similarity of characters and the proximity of the Island to Somalia’. Grubb (2001) agrees with this on the basis of skull measurements and the examination of pelage coloration. Like Dandelot (1968 in Groves, 2001, Grubb et al. (2003), Grubb (2006), and Lawes et al. (in press), we suspect that phylax is a synonym of C. m. albotorquatus. We suspect this for two reasons: 1) The channel that separates the Lamu Archipelago Islands from the mainland is shallow and narrow (Patta Island ca 3 km; Manda Island ca 20 m; Lamu Island ca 150-200 m). Therefore, these Islands are not particularly isolated. The extensive mangrove around all islands might be more of a barrier to primate movement than is the water. If C. mitis on the Lamu Archipelago is an endemic subspecies, then any differences from the mainland forms may be due more to the need to live in a very 'fresh water restricted' habitat and in mangrove forest, than to any isolation related to distance from the mainland. C. m. albotorquatus is known to occur on the mainland ca 45 km away (Witu Forest Reserve) from Lamu Island and ca 55 km away from Manda Island. It seems likely that in the past this taxon experienced gene flow with C. m. albotorquatus on the mainland on more than one occasion. 2) C. m. phylax is described as “having a dark olive head, upper parts dark reddish brown, russet tuft under base of tail, black arms, mouse grey legs, under parts white, and a white part-collar round neck” (Grubb, 2001). According to Groves (2001) phylax is “small in size”. Grubb (2001) compared the skull measurements of the female holotype to C. m. albotorquatus (female from Tana River) and found that the phylax holotype is slightly larger than the mainland specimen. He also notes that the pelage falls within the range of the Tana River specimens. Grubb (2001) states that “Some of these measurements may have been taken differently, but they are quite insufficient to suggest that insular and mainland populations are subspecifically different.” The animal we encountered on Manda Island looked similar to C. m. albotorquatus we encountered on the mainland. Unfortunately, we only (briefly) saw one C. mitis on Manda Island and we failed to obtain a photo. The group we encountered was shy and only the crown, face and under parts were seen. The crown of the individual we encountered on appeared no darker than the crown of C. m. albotorquatus. During this study we did not visit Patta Island. Future surveys should include a visit to Patta Island to further describe the C. mitis there and to collect preliminary ecological data on this taxon. Future research priorities:
• What is the southern limit of the geographical range of C. m. albotorquatus? • What is the geographical range of C. m. phylax? • How is C. m. phylax phenotypically different from C. m. albotorquatus (in Patta and
Manda Islands) and is it a valid subspecies? • Which subspecies of C. mitis occurs between the Tana River (west Meru National Park,
Ngaia Forest, Meru Forest and Mwea National Park) and the confirmed range of Cercopithecus mitis kolbi (Mt Kenya Forests).

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 63
4.2.6 Chlorocebus pygerythrus hilgerti (Neumann, 1902) Vernacular name: Eastern vervet Type locality: Gobeli River, Webbi Shibeli, Harar, Ethiopia Synonyms: ellenbecki (Neumann, 1902), Lake Zwai, Suksuki and Maki Rivers, Ethiopia. johnstoni (Pocock, 1907), Moshi, S Mt Kilimanjaro, Tanzania. 1500 m asl callida (Heller, 1912), S Lake Naivasha, Kenya arenaria (Heller, 1913), Merille water holes, Marsabit Road, N Kenya contigua (Heller, 1920), Changamwe, 10 W Mombasa, Kenya luteus (Elliot, 1910), Wambugu, SW Mt Kenya, Kenya rubellus (Elliot, 1909), Fort Hall, Kenya. tumbili (Heller, 1913), Ndi, Taita District, S Kenya. Field characters: Medium-size, slender monkey (Figure 22). Face and ears blackish to jet black. Brow band, cheeks and throat off-white to white, forming a complete ring around the black face. Cheek whiskers medium to long sweeping upwards and backwards, covering part of the ears. Crown and dorsum grizzled olive-grey, intense olive-grey, or pale sandy brown. Ventrum and inner limbs off-white to white. Sides in some populations show a ‘white stripe’ between the olive-grey dorsum and the white ventrum. Upper outer arms and legs grizzled olive-grey. Lower outer arms and legs grizzled grey. Hands and feet blackish to jet black in males and grey to blackish in females and juveniles. Top of tail grizzled mouse grey, distal ca. 10% darker grey to jet black. Red patch under base of the tail of males in some populations. Underside of tail pale grey. Distribution: Throughout Kenya (excluding the Lamu Archipelago). Into E Uganda, S Ethiopia, S Somalia, and N Tanzania (Figure 23). Extent of occurrence: Not known. Photographic map: Photo map of Chlorocebus pygerythrus, contains 86 photographs, of which 20 in the coastal forests of Kenya (T.M. Butynski & Y.A. de Jong). http://picasaweb.google.com/wildsolutions/ChlorocebusPygerythrus?feat=email or via: www.wildsolutions.nl
Figure 22. Chlorocebus pygerythrus hilgerti male at Diani Beach, Kenya.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 64
Figure 23. Approximate geographic distribution of Chlorocebus pygerythrus hilgerti in the coastal area of Kenya with sites of encounters (2003-2008) depicted. Based on Y. de Jong & T. Butynski (unpubl. data) and Kingdon et al. (2008c). There is not enough data available to produce a more accurate distribution map of C. p. hilgerti.
Encounter sites:
- Wenje (15, 251A; Appendix 2) - Tana River Primate National Reserve north (256; Appendix 2) - Tana River Primate National Reserve north (252A; Appendix 2) - Kipini Conservancy (259; Appendix 2) - Mpeketoni (265; Appendix 2) - Tiwi (353; Appendix 2) - Nyali (353A; Appendix 2) - Malindi (356; Appendix 2) - Malindi West (358; Appendix 2) - Diani (373; Appendix 2) - Lamu Island (none seen but evidence collected for their presence (Appendix 2) - Manda Island (none seen but evidence collected for their presence (Appendix 2) Encounters while travelling to the Hotspot: - Tsavo West National Park (326, 328, 331, 334, 337; Appendix 2) - Tsavo East National Park (362; Appendix 2) - Voi south (249; Appendix 2) - Kora National Park (20, 21, 22, 23, 24, 25, 27A, 304; Appendix 2) - Meru National Park (27, 28, 307, 308A; Appendix 2) - Mwingi (12; Appendix 2) - Mwea National Reserve (1, 3, 5; Appendix 2)

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 65
Encounter rate: A total of 44 groups of C. p. hilgerti were encountered of which 11 groups were in the coastal forests of Kenya. Thirty-five groups were encountered while travelling to and from the Hotspot (Table 8; Figure 23). Table 8. Encounter rates (groups/hour and groups/km) of Chlorocebus pygerythrus hilgerti during 2005 – 2008 in the coastal forests of Kenya and travelling to the coastal area. Only included the surveys C. p. hilgerti was encountered. Area Groups/hour Groups/km Vehicle/Foot Note Nyali, Haller’s Park.
1 (n=1) 0.78 (n=1) Foot
Kipini Conservancy /Mpeketoni/ Witu Forest Reserve
0.1 (n=2) 0.02 (n=2) Vehicle
Tsavo West NP 0.7 (n=5) 0.03 (n=5) Vehicle 2 days survey Tsavo East NP 0.3 (n=1) 0.01 (n=1) Vehicle Kora NP 0.1 (n=1) 0.01 (n=1) Vehicle 2 days survey North Kitui NR/Kora NP
1. 7 (n=5) 0.08 (n=5) Vehicle
Meru NP (2005) 0.8 (n=2) 0.03 (n=2) Vehicle One additional group seen from Meru NP on Kora NP side of Tana River
Meru NP (2007) 0.4 (n=2) 0.02 (n=2) Vehicle Mwea NR 0.7 (n=3) Vehicle Road Surveys Diani – Nyali, south coast
0.4 (n=1) 0.02 (n=1) Vehicle, 25.2 km/h Travelling slowly on highway
Tana River PNR – Kipini Conservancy
0.2 (n=1) 0.01 (n=1) Vehicle, 27.5 km/h
Garissa – Tana River PNR
0.4 (n=2) 0.01 (n=2) Vehicle, 38.3 km/h
Watamu – N. Malindi – W. Malindi - Watamu
0.2 (n=1) 0.01 (n=1) Vehicle, 23.5 km/h
Tanga (Tz) – Kaya Rabai, Mazeras
0.4 (n=3) 0.01 (n=3) Vehicle Through Mrima Hill and Shimba Hills NR
Kaya Rabai, Mazeras - Nairobi
0.1 (n=1) 0.002 (n=1) Vehicle Highway
Mwea NR – Mwingi East
0.2 (n=1) 0.01 (n=1) Vehicle
Mwingi East – Tana River PNR
0.2 (n=2) 0.10 (n=2) Vehicle
Vegetation: Savannah, open to medium dense Acacia and Acacia-Commiphora woodland, close to permanent water sources containing tall Acacia trees (A. xanthophloea and A. tortilis). Found in miombo woodland, riverine vegetation and forest-grassland mosaic. Able to persist in degraded and fragmented vegetation. Throughout its range is able to live close to human

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 66
settlements and hotel compounds, attracted by water, agricultural crops, human food, and security from predators. Requires a year round source of water and tall trees in which to sleep and escape from predators. Altitude range: 0 – 2000 m asl (Naivasha, Kenya; Y. A. de Jong & T. M. Butynski, unpubl data) Conservation status: C. p. hilgerti is a very patchily distributed subspecies that is, however, often common in the open and medium dense Acacia woodlands of the coastal strip of Kenya. There are no threats to their conservation. Hunted in many places because of the damage it causes to crops. C. p. hilgerti can be especially common and abundant in the vicinity of tourist lodges and hotels on the coastal strip in Kenya.
IUCN Red List: Chlorocebus pygerythrus: Least Concern (Kingdon et al., 2008b; www.iucnredlist.org/details/136271). Assessors: Kingdon, J., Gippoliti, S., Butynski, T.M. & De Jong, Y.A. C. p. hilgerti is recognised but not assessed by the 2008 IUCN Red List of Threatened Species. Discussion: Chlorocebus pygerythrus is a patchily distributed but locally common species in the coastal forests of Kenya. The taxonomy of this taxon has been debated for many years. This study recognised C. p. hilgerti as the subspecies present in the coastal strip of Kenya. This is consistent with Groves (2001, 2005, 2006), and Kingdon et al., 2008b but at odds with the taxonomy of Hill (1966), Kingdon (1971, 1997), Napier (1981), and Grubb et al. (2003). We fully agree with Grubb et al. (2003) and Groves (2006) that more study and a full revision of the genus is required. During this study we found surprisingly little phenotypic variation between C. p. hilgerti populations in the extremes of their distribution in coastal forests of Kenya. We did find variation in the intensity of their colours in the region (and throughout Kenya and Tanzania; T. M. Butynski & Y. A. de Jong, unpubl data). Table 9 presents field description of C. p. hilgerti obtained from Mpeketoni (NE Kenya coast; number 265, Appendix 2), Diani (S Kenya coast; number 373, Appendix 2) and, as a comparison, Shaba National Reserve (central Kenya, outside the coastal area; number 370, Appendix 2). Table 9. Field descriptions of Chlorocebus pygerythrus hilgerti obtained from the extremes of the coastal forests of Kenya (i.e., Diani and Mpeketoni) and from Shaba National Reserve in central Kenya. Body Part Diani Mpeketoni Shaba NR Photo
Face Jet black, hairless Blackish, pale skin around Black

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 67
eyes Ears Black Blackish Blackish Whiskers White, medium length,
sweep upwards and backwards covering part of the ear.
White, long, sweep upwards and backwards covering the ears
White, medium long, sweep upwards and backwards covering part of the ears.
Brow band White, forming a complete white ring around face with cheeks and throat
White, forming a complete white ring around face with cheeks and throat
White, forming a complete white ring around face with cheeks and throat
Crown Grizzled olive-grey Grizzled olive-grey Grizzled olive-grey Throat Off white White White Dorsum Grizzled olive-grey. Grizzled olive-grey. Grizzled olive-grey. Ventrum Off white White White Sides Grizzled olive-grey.
Narrow off-white lateral stripe.
Intense olive in contrast with white ventrum. Narrow white lateral stripe.
Intense olive in contrast with white ventrum. Narrow white lateral stripe.
Hands and feet Black in males, grey to dark grey in juveniles and females.
Blackish in males, pale grey to dark grey in juveniles and females.
Blackish in males, pale grey to dark grey in juveniles and females.
Outer arms Upper arms grizzled olive-grey, lower arms grizzled grey
Upper arms grizzled olive- grey, lower arms grizzled grey
Upper arms grizzled olive-grey, lower arms grizzled grey
Inner arms Off-white White White Outer legs Grizzled mouse grey Upper legs grizzled olive-
grey. Lower legs grizzled grey.
Upper legs grizzled olive- grey. Lower legs grizzled grey.
Inner legs Off-white White White Tail Grizzled mouse grey,
distal ca. 10% darker grey. Red patch under base of the tail in males. Under tail slightly lighter grey.
Grizzled grey, terminal end black. Red patch under base of the tail in males. Under tail slightly lighter grey.
Grizzled grey, tail tip black. Under tail slightly lighter grey.
Overall Phenotypically like most other populations of C. p. hilgerti.
Relatively strong contrast between dorsum and ventrum due to more intense colours. Long whiskers stand out.
Phenotypically like most other populations of C. p. hilgerti.
The main phenotypic differences we found in the extremes of the geographical range of C. p. hilgerti in Kenya are the ‘white lateral stripes’ in some populations, the length of the whiskers, the intensity of fur colour (including the black and white body parts). Future research priorities:
• What is the extent of occurrence or area of occupancy of C. p. hilgerti?

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 68
4.2.7 Chlorocebus pygerythrus excubitor (Schwarz, 1926) Vernacular name: Manda vervet monkey Subspecies type locality: Manda Island, Witu Islands (=Lamu Archipelago), off coast of N Kenya. Synonyms: Chlorocebus voeltzkowi, Matschie, 1923 (nomen nudum). Field characters: No data Distribution: Lamu Archipelago, including Lamu Island, Manda Island, Patta Island, and probably Manda Toto Island (Figure 24). Extent of occurrence: < 200 km², based on the size of Lamu, Manda, and Patta Islands, excluding the extensive mangrove forests around the Islands which is unsuitable habitat for C. pygerythrus. The area of occupancy is highly fragmented and expected to be much smaller than this. Figure 24. Suspected geographic distribution and evidence for Chlorocebus pygerythrus excubitor. The actual area of occupancy is probably very much smaller than shown.
Photographic map: We have no photos of C. p. excubitor. Encounter sites: C. p. excubitor was not observed during this study but evidence was collected that this taxon is present in the following areas:
- Matondoni and the surrounding area, NE Lamu Island - Lamu Town and vicinity - Manda Airstrip and vicinity, E Manda Island

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 69
- SE Manda Island - Takwa Ruins, SE Manda Island - Manda Toto Island occasionally (reported by local fisherman, n=3). Said to sometimes
cross the narrow channel between Manda Island and Manda Toto Island during low tide. During this study we were not in the position to survey Patta Island. Encounter rate: C. p. excubitor was not encountered during this study. On all Islands C. p. excubitor occurs at very low densities Vegetation: No data. Evidence was collected, however, that C. p. excubitor occupies savannah wood/bushland and is attracted to farms and human settlements. Altitude range: sea level to ca 10 m asl. Conservation status: C. p. excubitor occurs in very low numbers in the Lamu Archipelago. One of the causes for this is likely the scarcity of year round fresh water sources on the Islands. Residents stated that C. p. excubitor is hunted and kept or sold as pets. It is unknown if the pet trade is a serious threat to this subspecies but this seems unlikely. The geographical range of C. p. excubitor is highly fragmented (between the Islands) and its total numbers must be very low. IUCN Red List: Chlorocebus pygerythrus: Least Concern (Kingdon et al., 2008b; www.iucnredlist.org/details/136271). Assessors: Kingdon, J., Gippoliti, S., Butynski, T.M. & De Jong, Y.A. C. p. excubitor is recognised but not assessed by the 2008 IUCN Red List of Threatened Species. The maximum possible area of occupancy for C. p. excubitor is < 200 km². Given this small and fragmented distribution, C. p. excubitor qualifies as Endangered (EN, B1) on the basis of the present criteria (IUCN, 2008).
Discussion: Chlorocebus pygerythrus excubitor (Schwartz, 1926) is accepted as a valid subspecies by Hill (1966), Kingdon (1971, 1997), Napier (1981), Groves (2001, 2005), Grubb et al. (2003), and Kingdon et al. (2008b). The holotype was collected on Manda Island. Hill (1966) describes this subspecies as “A small insular race of dull reddish pelage, very similar to Chlorocebus pygerythrus nesiotis but redder and, in the darker phase, heavily speckled with black.” During this study, we visited the Lamu Archipelago (Lamu, Manda, and Manda Toto Islands) for 8 days, collected evidence for the presence of C. pygerythrus, but did not see or hear this animal. Due to the absence of observations of C. pygerythrus in the Lamu Archipelago during this study, additional visits and surveys need to be conducted to either support or reject C. p. excubitor as a valid subspecies. If C. p. excubitor is indeed a valid subspecies, than its geographical range is highly fragmented and it population size is critically low. In this study we threat C. p. excubitor as a valid subspecies due to the lack of evidence to the contrary. However, having surveyed a part of the geographical range of C. p. excubitor, we have the following concerns about its current subspecies status:

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 70
- C. p. hilgerti occurs on the mainland only ca 30 km away (Mpeketoni) from Manda Island. It is likely that the geographical range extents even farther towards Lamu and Manda Islands.
- Manda Island is separated from the mainland by a very narrow and shallow channel that, in some places, is <20 m wide. During low tide, Manda Island is connected to the mainland. Lamu Island is ca. 15-200 m from the mainland. Therefore, both Islands are not particularly isolated. The extensive mangrove around all Islands might, however, be much more of a barrier to primate movement than is the open water.
Future research priorities:
• What is the geographical range of C. p. excubitor? • Obtain field observations, photos, and detailed descriptions of specimens of C.
pygerythrus from Lamu, Manda and Patta Islands and C. pygerythrus from the adjacent mainland to confirm the validity of C. p. excubitor.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 71
4.2.8 Papio cynocephalus ibeanus Thomas, 1893 Vernacular name: Ibean yellow baboon Subspecies type locality: Lamu Island, Lamu Archipelago, Kenya Synonyms: Papio ruhei Zukowsky, 1942. Banks of Webbi Shabeli, 40 km NW of Mogadishu, Somalia; Papio thoth ibeanus, Thomas 1893, Kenya: Lamu Field characters: Large, slender, mostly terrestrial primate (Figure 25). Males have a trace of mane, are larger and more robust than females. Muzzle almost hairless and pointed forward and slightly upwards with dark grey to blackish skin. In most populations, pale grey ‘muzzle-patches’ are present on either side of the muzzle just below the eyes. Cheeks and chest yellowish to off-white. Ears sparsely haired, dark grey to blackish, pointed in some populations. Crown grizzled yellowish-brown. Neck, dorsum, sides, and outer limbs grizzled yellowish-brown, yellowish-grey, or olive-brown. Ventrum and inner limbs pale yellowish-grey or off-white. Back of the upper legs pale yellow to golden-yellow. Hands and feet grizzled yellowish-brown to blackish. Tail grizzled yellowish-brown, base upwards, followed by a kink or a bend, than dropping down. Tail shape and length variable within groups. Callosities hairless, dark grey. Hair relatively wavy and long, especially on the shoulders, sides and upper legs. Distribution: S Somalia, SE and coastal Kenya (Figure 26; Jolly, 1993; Kingdon et al., 2008; Altmann et al., in press). Extent of occurrence: no data Photographic map: Photo map of Papio cynocephalus ibeanus, contains 103 photographs, of which 17 coastal forests of Kenya (T.M. Butynski & Y.A. de Jong). http://picasaweb.google.com/wildsolutions/PapioAnubisPapioCynocephalus?feat=email# or via: www.wildsolutions.nl
Figure 25. Adult male Papio cynocephalus ibeanus foraging on the beach of Manda Toto Island, Lamu Archipelago, Kenya.
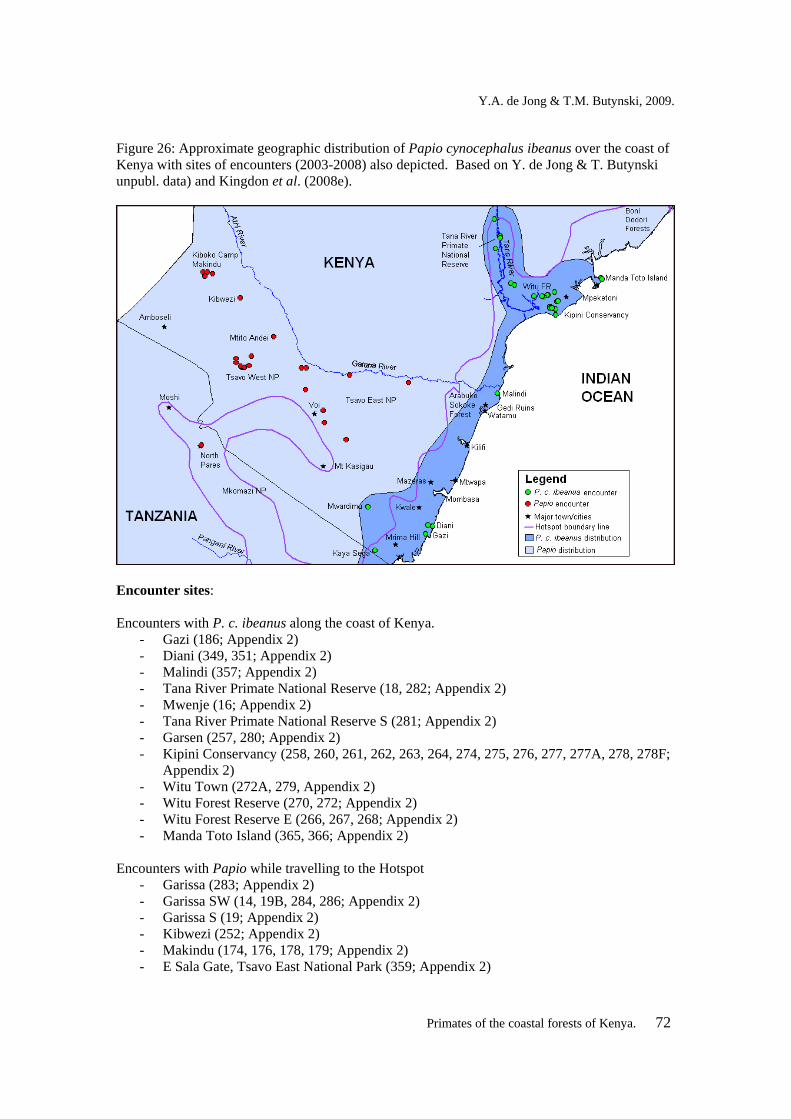
Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 72
Figure 26: Approximate geographic distribution of Papio cynocephalus ibeanus over the coast of Kenya with sites of encounters (2003-2008) also depicted. Based on Y. de Jong & T. Butynski unpubl. data) and Kingdon et al. (2008e).
Encounter sites: Encounters with P. c. ibeanus along the coast of Kenya.
- Gazi (186; Appendix 2) - Diani (349, 351; Appendix 2) - Malindi (357; Appendix 2) - Tana River Primate National Reserve (18, 282; Appendix 2) - Mwenje (16; Appendix 2) - Tana River Primate National Reserve S (281; Appendix 2) - Garsen (257, 280; Appendix 2) - Kipini Conservancy (258, 260, 261, 262, 263, 264, 274, 275, 276, 277, 277A, 278, 278F;
Appendix 2) - Witu Town (272A, 279, Appendix 2) - Witu Forest Reserve (270, 272; Appendix 2) - Witu Forest Reserve E (266, 267, 268; Appendix 2) - Manda Toto Island (365, 366; Appendix 2)
Encounters with Papio while travelling to the Hotspot - Garissa (283; Appendix 2) - Garissa SW (14, 19B, 284, 286; Appendix 2) - Garissa S (19; Appendix 2) - Kibwezi (252; Appendix 2) - Makindu (174, 176, 178, 179; Appendix 2) - E Sala Gate, Tsavo East National Park (359; Appendix 2)

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 73
- Meru NP P. anubis (26, 84, 301, 308, 309; Appendix 2) - Mtito Andei (180; Appendix 2) - Mwardimu (340; Appendix 2) - Mwea National Reserve (2, 6, 9, 11, 363, 364; Appendix 2) - Tsavo West National Park (182, 325, 327, 329, 332, 335, 336, 339; Appendix 2) - Tsavo East National Park (360, 361; Appendix 2) - Voi N (250, 251; Appendix 2) - Voi S (183, 184; Appendix 2)
Encounter rate: A total of 71 Papio groups were encountered of which 32 were P. c. ibeanus groups within the Kenya coastal forests. The remaining 39 groups were encountered while travelling to and from the Hotspot (Table 10; Figure 26). Table 10. Encounter rates (groups/hour and groups/km) of Papio cynocephalus ibeanus during 2005 – 2008 in the Kenya coastal forest Hotspot and while driving to the Hotspot. Only includes the surveys in which C. c. ibeanus was encountered. Area Groups/hour Groups/km Vehicle/Foot Tana River PNR 2005 0.1, n=1 Vehicle Tana River PNR 2006 0.1, n=1 Vehicle Kipini Conservancy & Witu Forest Reserve
0.8, n=20 0.16, n=20 Vehicle
Manda Toto Island 0.2, n=2 0.13, n=2 Foot Mwea NR 2005 0.7, n=4 Vehicle Mwea NR 2008 0.7, n=2 0.05, n=2 Vehicle, 13.6 km/h Tsavo West NP 1.0, n=7 0.05, n=7 Vehicle, 20.1 km/h Tsavo East NP 0.3, n=1 Vehicle, 23.3 km/h Road Surveys
Mwea NR – Tana River PNR
0.1, n=2 0.004 Vehicle, ca 45 km/h
Tana River PNR – Garissa
0.2, n=1 0.01, n=1 Vehicle, ca 45 km/h
Tana River Primate Reserve –Garsen – Witu
0.3, n=1 0.01, n=1 Vehicle, 30 km/h
Witu – Garsen – Tana River PNR
1.1, n=4 0.03, n=3 Vehicle, 40.2 km/h
Nairobi – Makindu – Mrima Hill
0.7, n=5 0.13, n=1 Vehicle, ca 65 km/h
Mazeras – Nairobi 0.4, n=3 0.01, n=1 Vehicle, 43.8 km/h Tana River PNR – Garissa – Nanyuki
0.3, n=3 0.01, n=3 Vehicle, ca 50 km/h
Garissa – Kora NP 0.1, n=1 0.003, n=1 Vehicle Kora NP – Meru NP 0.4, n=1 0.02, n=1 Vehicle Mtito Andei – Kilimangodo
0.2, n=1 0.004, n=1 Vehicle, 41.5 km/h
Kaya Sega – Diani 0.3, n=2 0.01, n=2 Vehicle, 27.9 km/h Mazeras – Nairobi 0.4, n=3 0.01, n=3 Vehicle Malindi – Tsavo East NP
0.3, n=2 0.01, n=2 Vehicle, 31.8 km/h
Vegetation: Savannah, open woodland, Brachystegia woodland (Miombo), dry bushland, and forest-grassland mosaic. Avoids forest but occupies forest edges. Encountered on beaches (Manda

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 74
Toto Island, Kenya, and on beaches in Saadani National Park, coastal Tanzania; T. M. Butynski & Y. A. de Jong, pers. obs.) and uses mangrove forests (also Kingdon et al., 2008e). P. c. ibeanus is able to persist in secondary and highly fragmented vegetation, including cultivated areas. Often encountered close to human settlements, attracted by water, human food, crops, and security from predators. Altitude range: Sea level to at least 1140 m asl Conservation status: P. c. ibeanus is a widespread, common and opportunistic primate in the coastal forests of Kenya. Outside the Hotspot it is even more abundant, occurring in and outside protected areas. No major threats to the survival of this subspecies are known. P. cynocephalus is listed as a vermin under the African Convention due to their tendency to raid crops and forage in and around human settlements. IUCN Red List: Papio cynocephalus ibeanus: Least Concern (Kingdon, 2008; http://www.iucnredlist.org/details/136862). Assessors: J. Kingdon Discussion: P. c. ibeanus is locally common in the coastal forests of Kenya where it is often found near human settlements to which it is attracted by water human food, crops and security from predators. This species appears to be limited mainly by year around supplies of water and suitable sites for sleeping (i.e., large, steep, cliffs and/or tall trees). The taxonomy of species with large geographical ranges and which occupy many different habitats are often a topic of debate. Often these widespread species show considerable phenotypic variation but are genetically linked due to intermediate populations (Sarmiento, 1998). During this study we found phenotypic variation among groups of P. c. ibeanus throughout its range inside and outside coastal forests of Kenya (see Table 11.). Groves (2001, 2005), Grubb et al. (2003), Kingdon et al. (2008e), and Altmann et al. (in press) recognize three subspecies for Papio cynocephalus: P. c. cynocephalus, P. c. ibeanus, and P. c. kindae. Table 11. shows some of the variation between P. cynocephalus groups at Diani [S coast; which Hill (1967) referred to P. c. cynocephalus] and groups on Manda Toto Island [N coast; which Hill (1967) referred to P. c. ibeanus]. The variation observed between Diani and Manda Toto Island (ca 300 km apart) is, however, less than the variation found within groups at Tana River Primate National Reserve (ca 100 km from Manda Toto Island), which indicates that P. c. ibeanus is present in Diani. Hill (1967) indicates that the Galana River is the boundary between P. c. ibeanus and P. c. cynocephalus (with P. c. cynocephalus occurring in the south part of the Kenya coastal strip and into Tanzania). He, thus, considers the Diani population to be P. c. cynocephalus, contradictive to our findings. P. c. ibeanus differs from P. c. cynocephalus in that its pelage is wavy---not straight (Jolly, 1993). Additionally, Jolly (1993) reports a trace of mane in P. c. ibeanus which is absent in P. c. cynocephalus. Zinner et al. (2008) also indicate that P. c. cynocephalus is present at Diani. Results of the molecular work on Papio in eastern Africa by Zinner et al. (2008) shows that Diani P. cynocephalus is genetically distinct from P. cynocephalus in Amboseli.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 75
P. c. ibeanus from S Somalia (once referred to as P. c. ruhei but now considered a synonym of P. c. ibeanus; Groves, 2001, Grubb et al., 2003, Zinner et al., 2008), is genetically closely related to P. anubis from Kenya and N Tanzania, and P. c. ibeanus from Amboseli. Zinner et al. (2008) speculate that P. c. ibeanus is a hybrid form between P. anubis and P. c. cynocephalus and that their P. cynocephalus sample from Diani represents P. c. ibeanus. Further research is needed along the south coast of Kenya and the north coast of Tanzania in order to determine where P. c. ibeanus gives way to P. c. cynocephalus, or whether there is a cline from one form into the other. Acknowledging that P. c. cynocephalus and P. c. ibeanus are hybridizing, we strongly suspect that no sharp boundary between these two subspecies exists and that what is present is a cline of intermediate forms. As such, the validity of P. c. ibeanus and of P. c. cynocephalus would be called into question as, indeed, would the validity of P. anubis. The morphological features displayed by each ‘subspecies’ of Papio includes pelage, body size, and dentition (Hill, 1967; Jolly & Brett, 1973, Hayes et al., 1990 in Kamilar, 2006). According to Kamilar (2006), there is no clear differentiation among the ecologies of savannah baboon subspecies. This is in support of the hypothesis by Jolly (1993) that non-hamadryas baboon taxa lack ecological separation. The morphological variation that is found in the subspecies of Papio are due to a latitudinal cline and that all savannah baboons are of one species (Jolly, 1993; Kamilar, 2006). We agree with Jolly (1993), Kamilar (2006) and Zinner (2008) on several points. However, we think it is useful in this study to classify the primates in the Kenya portion of this Hotspot as subspecies for the present time---pending further study. Therefore, we accept the taxonomy of Groves (2001, 2005), Grubb et al. (2003), Kingdon et al. (2008e), and Altmann et al. (in press), and recognise P. c. ibeanus to be the subspecies of P. cynocephalus present in the Kenya part of the Hotspot. Natural hybridization between P. anubis and P. c. ibeanus in the extensive contact zone running across Kenya is often reported (Maples & McKern, 1967; Kingdon, 1971, 1997; Maples, 1972; Napier, 1981; Samuels & Altmann, 1992; Jolly, 1993, 1998; Sarmiento, 1998; Alberts & Altmann, 2001; Groves, 2001; Grubb et al., 2003; Newman et al., 2003; Kamilar, 2006; Kingdon et al., 2008e; Zinner et al., 2008; Altmann et al., in press; Palombit, in press; Butynski & de Jong, pers. obs.). The coastal forests of Kenya is occupied by P. c. ibeanus, no P. c. ibeanus x P. anubis hybrids were encountered within to the Hotspot. Hybrids were however encountered over an extensive area while driving to the Hotspot from the Kenya Highlands where P. anubis occurs. Research has been conducted on hybrid groups in Amboseli National Park, Kenya. P. c. ibeanus is known to occur in Amboseli but P. anubis sometimes move into Amboseli from either the south (Mt Kilimanjaro), southeast (Namanga), or north (hybrid zone occurring around Shimba and Kiboko, along the Nairobi – Mombasa Highway; Maples & McKern, 1967). More research is needed to more clearly delineate the P. c. ibeanus x P. anubis hybrid zone. However, this study confirms at least sections of the hybrid zones: 1. NE & E of Mt. Kenya towards the Tana Delta. Papio across this >200 km wide area are readily recognized as phenotypically intermediate between P. anubis and P. c ibeanus. They also exhibit much intragroup variation in coat colour and body shape. From the interior of Kenya towards the coast, baboons become phenotypically more like the coastal P. c. ibeanus. 2. Hybrids occur along the Nairobi – Mombasa Highway, starting at least 35 km E of Makindu (Mapes & Mc Kern, 1967), N of the Chyulu Hills National Park , through Tsavo. Along this hybrid cline, baboons gradually become phenotypically more like P. c. ibeanus towards the coast.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 76
Table 11. Field descriptions of adult male P. c. ibeanus obtained from Manda Toto Island, Lamu Archipelago (close to the type locality of P. c. ibeanus), Tana River Primate National Reserve, Diani Beach (S coast). Field description
P. c. ibeanus Manda Toto Island
P. c. ibeanus Tana River PNR
P. c. ibeanus Diani Beach
Foto
Cheeks Yellowish to off-white Yellowish-white to pale grey
Pale yellowish-grey
Nostrils Directed upwards Directed upwards Directed upwards Pale grey muzzle patch
Present Present but vague Present
Head shape
Pointed Pointed Pointed
Mane Trace Absent Trace Dorsum Grizzled yellowish- grey Grizzled yellowish- grey.
Some with brighter yellow than others
Grizzled yellowish- grey
Ventrum and inner limbs
Greyish to off-white with yellow wash
Pale yellowish-white to pale grey
Pale yellowish-grey
Callosite shape
Medium U-shape Medium U-shape Medium U-shape
Tail Kinked Bent (arched) Kinked Body shape
Robust compared to Tana River PNR Papio
Slender (gracile) Robust compared to Tana River PNR Papio
Overall Phenotypically similar to population at Diani, SE Kenya.
Very different from other populations of Papio observed in East Africa. Also, more phenotypic variation seen within groups than observed for other populations of Papio in East Africa.
Phenotypically similar to population at Manda Toto Island (close to type locality)
Further research priorities:
• Where does P. c. ibeanus give way to P. c. cynocephalus, or is there a cline from one form into the other?
• What is the exact location and size is the P. c. ibeanus x P. anubis hybrid zone? • Are all savannah baboons part of one species?

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 77
4.2.9 Colobus angolensis palliatus Peters, 1868 Vernacular name: Peter’s Angola Colobus Subspecies type locality: East Africa, opposite Zanzibar (=Unguja Island). Synonyms: sharpei, Thomas, 1902. Fort Hill, Malawi. langheldi, Matschie, 1914. Ujiji, E Lake Tanganyika. Field characters: Medium-sized, arboreal, wholly black-and-white monkey with white cheeks, shoulders, and tail tip (Figure 27). Distribution: Discontinuously distributed in SE Kenya. Restricted to coastal forests of Kwale District. Northern boundary is the the border between Kwale District and Mombasa town. Southern Highlands, Eastern Arc Mountains, coastal forests and gallery forests of eastern Tanzania (Figure 28; Napier, 1985; Kingdon, 1997; Groves, 2001; Anderson et al., 2007; Kingdon et al., 2008d). Extent of occurrence: ca 272,000 km². Area of occupancy in Kenya ca. 249 km² (Anderson et al., 2007). Figure 28. Known distribution and encounters (2003-2008) with Colobus angolensis palliatus in the coastal forests of Kenya. The other sources used for this map are Kingdon et al., (2008d) and Anderson et al., (2007).
Figure 27. Adult male Colobus angolensis palliatus at Mrima Hill, south coast of Kenya, where this subspecies is common.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 78
Photographic map: The photo map for African Colobinae contains 17 photos, of which three are of C. a. palliatus (T. M. Butynski & Y. A. de Jong). http://picasaweb.google.com/wildsolutions/Colobinae?feat=email# or via: www.wildsolutions.nl Encounter sites:
- Mrima Hill (188, 189, 192, 193, 196, 199, 246, 247, 344; Appendix 2) - Diani (347, 375; Appendix 2)
Encounter rate: During this study, 11 groups of C. a. palliatus were encountered, all in the coastal forests of Kenya (Table 12; Figure 28). Table 12. Encounter rates (groups/hour and groups/km) of Colobus angolensis palliatus during 2005 – 2008 in the coastal forests of Kenya. This table only includes the surveys in which C. a. palliatus was encountered. Area Groups/hour Groups/km Vehicle/Foot Mrima Hill 3.1, n=9 0.42, n=9 Vehicle Kaya Sega - Diani, road survey
0.3, n=2 0.01 (n=2) Vehicle, 27.9 km/h
Diani Opportunistically, 1 group
Opportunistically, 1 group
Foot
Vegetation: C. a. palliatus was encountered in coastal forest, on the edge of forest and agricultural land, and in gardens of hotels and residencies. This taxon is able to survive in small patches of degraded and disturbed forest. Altitude range: sea level – at least 550 m asl (Amani Nature Reserve, W. Newmark, pers. comm.) Conservation: The major threat for the survival of C. a. palliates in Kenya is habitat loss and degradation due to clearance for agriculture, local settlements, tourism and coral block mining (Anderson et al., 2007; Bocian & Anderson, in press). The geographical range of C. a. palliates is highly fragmented. The largest forest, Shimba Hills National Reserve, is ca 150 km2 (Anderson et al., 2007). C. a. palliates is a flagship species for SE Kenya (Kahumbu, 1997; Cunneyworth & Rhys-Hurn, 2004, in Anderson et al., 2007).
C. a. palliates is present in several protected areas (e.g., Shimba Hills National Reserve) and in some of the Mijikenda Kaya Forests (UNESCO World Heritage site since 2008; Unesco, 2009). Anderson et al. (2007) reports that C. a. palliates does not raid crops.
IUCN Red List: Colobus angolensis palliates: Least Concern (Kingdon et al., 2008d; http://www.iucnredlist.org/details/5148). Assessors: J. Kingdon, T.T. Struhsaker, J. Oates, J.F. Hart, T.M. Butynski, Y.A. de Jong, & C.P. Groves. C. a. palliates was ‘Data Deficient’ in the 2000 Red List Assessment.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 79
Discussion: C. a. palliates is a locally common subspecies in the coastal forests of south Kenya where it sometimes occurs in very small patches of forest. In 2001, C. a. palliates was found in 55 populations in forest fragments in SE Kenya (Anderson et al., 2007). The total population in Kenya was estimated to be 3100-5000 individuals. Shimba Hills National Reserve supports the largest C. a. palliates population (17% of the animals) and is the largest protected forest in the geographical range (density estimate: 2.9±0.52 groups/km2 and 15.3±2.88 individuals/km² in the Reserve; Anderson et al., 2007). Density of C. a. palliates varies widely among sites and is significantly affected by forest area, forest loss over 12 years, and the availability of 14 major food tree species. Density in the Diani Forest is ca. 31 individuals/km2; in the Mwache F. R., ca. 6 individuals/km2. Are absent from some forest fragments (Anderson et al., 2007a). Historically, was present north of Mombasa to Arabuko Sokoke Forest (Anderson et al., 2007). Within Kenya, currently restricted to Kwale District, south of Mombasa.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 80
4.2.10 Procolobus rufomitratus rufomitratus (Peters, 1879) Vernacular name: Tana River Red Colobus Subspecies type locality: Muniuni, Tana River, Kenya Synonyms: Piliocolobus rufomitratus (Peters, 1879) Field characters: Medium-sized, arboreal monkey with russet crown and greyish-brown back and tail. Distribution: From Kipende in the north to Mitipani 60 km to the south, where the Lamu-Garsen road enters the Tana River floodplain, Lower Tana River, Kenya (Butynski et al., 2008c) Extent of occurrence: Sixty kilometres along the lower Tana River in riverine forest. Area of occupancy is considerably smaller as the forests here are highly fragmented and small (Butynski et al., 2008c). Map: Map of the geographical range of P. r. rufomitratus by Butynski et al. (2008c) can be found on http://www.iucnredlist.org/details/39993/rangemap Photographic map: No photos taken of P. r. rufomitratus Encounter sites:
- Mchelelo Research Station, (17B, 255; Appendix 2) Encounter rate: No data. Vegetation: Dependent upon riverine / gallery forest (Struhsaker & Grubb, in press). Altitude range: 20-40 m asl Conservation status: Habitat loss due to agricultural clearing and extraction of forest products by local communities is the main threat to P. r. rufomitratus (Butynski & Mwangi, 1994, 1995; Mbora, 2003; Mbora & Butynski, 2007; Butynski et al., 2008c; Struhsaker & Grubb, in press). Additionally, the alteration of river flow volume and cycles by five hydroelectric power dams upriver is likely to negatively affect the establishment, longevity and productivity of the forests along the lower Tana River (Butynski, 1995). Recently proposed agricultural ‘development’ projects (oil palm and sugar cane plantations) in the area are an indirect threat as they will greatly increase the number of people in the region. The proposed Tana Integrated Sugar Project in Tana River and Lamu Districts threatens more than 200 km² of semi-natural habitat in the area just to the south of the area in which the P. r. rufomitratus lives. The insecurity of the area and the lack of protection to the forests on which the P. r. rufomitratus depends could move this species into the Critically Endangered category in the near future (Butynski et al., 2008c; Wieczkowski & Butynski, in press). IUCN Red List: Endangered B1ab(iii,v) ver 3.1 (Butynski et al., 2008c; www.iucnredlist.org/details/39993) Assessors: T. M. Butynski, T. T. Struhsaker & Y.A. de Jong.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 81
Discussion: Since 2002, we have received reports of Procolobus in the Witu Forest Reserve and Kipini Conservancy on the north coast of Kenya. Kipini is ca. 90 km southeast of the known range of P. rufomitratus. Brief visits to the nearby Witu Forest Reserve by T. Butynski (pers. obs.) in 1994 and 2004 failed to reveal any Procolobus, but Procolobus was not the focus of his and no time was given to questioning local people about the presence of Procolobus. During this study, in June 2006, T. Butynski and Y. de Jong visited Kipini Conservancy for 1.5 days. Four residents of the Conservancy claimed to have seen Procolobus in Kipini. One of these people, Bakari Kawa Buya, was once a field assistant on a P. rufomitratus research project along the Tana River. As such, he is very familiar with P. rufomitratus. The informants independently claimed that there were at least two small groups of Procolobus in that part of the Conservancy that we visited, and that, unlike P. rufomitratus, these animals had reddish backs and tails. That is, their pelage color is noticeably different from P. rufomitratus. We found only three species of monkeys in Kipini Conservancy: yellow baboon Papio cynocephalus, Sykes’s monkey Cercopithecus mitis, and vervet Cercopithecus aethiops. All three are readily distinguished from Procolobus, and all three are well known to the local people---yet they claim that a fourth species, a Procolobus, is also present. These comments by the local people, together with the proximity of Kipini to the forests of the lower Tana River, strongly suggests that there is a fourth species of monkey in Kipini Forest.
We searched for Procolobus for 2.5 h on 24 June and for 3 h on 25 June. Although no Procolobus were encountered, we did find several sites with numerous green, compound, leaves on the ground, the petioles of which had been chewed off in a fashion characteristic of Procolobus feeding. Although we had considerable doubt as to the presence of Procolobus before we surveyed Kipini Conservancy, we realized that there was some chance that Procolobus are present---and that this was either a second population of P. rufomitratus or, more likely, a new taxon. If Procolobus are present, they must be in very low numbers. This, coupled with the fact that the area of Kipini Forest in which Procolobus are said to occur has recently been given over by the Kenya Wildlife Service (KWS) and Kipini Conservancy for resettlement of people from the lower Tana River area, means that time is a critical factor as the small area of remaining forest is likely to be destroyed in the near future.
As a result of our preliminary survey on Kipini Conservancy, we applied for a grant from the Margot Marsh Biodiversity Foundation which allowed us to follow up on the reports of Procolobus in Kipini Conservancy and Witu Forest Reserve. Additionally we will survey two other areas in East Africa for the presence of Procolobus. The primary objectives of the project are to (1) confirm or refute reports that populations of Procolobus occur in the Kipini Forest, Ngezi Forest, and Rufiji River Forests, and, if confirmed, (2) assess both the taxonomic and conservation status of these populations, and (3) make recommendations for conservation action.
During February and March, 2007, the Danish Zoological Society (with funds from CEPF) conducted surveys in Witu Forest Reserve (Nielsen & Sick, 2008). Aware of the P. rufomitratus reports they found no evidence of their presence and consider it unlikely that additional surveys will record P. r. rufomitratus in Witu forest Reserve due to the condition and isolated nature of Witu relative to other forest containing P. r. rufomitratus (Nielsen & Sick, 2008). . During this study no data was collected at the Tana River Primate National Reserve on P. r. rufomitratus.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 82
4.2.11 Cercocebus galeritus Peters, 1879 Vernacular name: Tana River Mangabey Subspecies type locality: Mitole, Tana River, Kenya Synonyms: Cercocebus galeritus ssp. galeritus, Peters, 1879 Field characters: A semi-terrestrial, medium-sized, greyish monkey with a long, lax, ‘crest’ of hair on the crown. Distribution: Endemic to a 60 km stretch of floodplain forests (in 27 patches) along the lower Tana River, SE Kenya, from Kanjonja in the north to Garsen in the south (01.40S-02.15S, 40.10E, 20-40 m asl; Butynski & Mwangi 1994, 1995; Butynski et al. 2008d). Extent of occurrence: Sixty square kilometres, but area of occupancy is considerably smaller (Butynski & Mwangi, 1994). Map: map of the geographical range of C. galeritus by Butynski et al. (2008d) can be found on http://www.iucnredlist.org/details/4200/rangemap Photographic map: none Encounter sites:
- Mchelelo Research Station, (16A, 19A, 253; Appendix 2) Encounter rate: No data. Found to be common within its small range (also Butynski et al., 2008d) Vegetation: restricted to riverine gallery forests and adjacent patches of bush (Butynski et al., 2008d) Altitude range: 20-40 m asl (Butynski et al., 2008d) Conservation status: C. galeritus only contains of one population of ca. 1,200 individuals. C. galeritus is severely threatened by deforestation for agricultural land, timber, burning of adjacent grasslands preventing forest regeneration, overgrazing of forest understory by livestock, and changes to the flow of the Tana River and water table by hydropower dams and irrigation projects. Recently proposed agricultural ‘development’ projects (oil palm and sugar cane plantations) in the area are an indirect threat as they will greatly increase the number of people in the region. The proposed Tana Integrated Sugar Project in Tana River and Lamu Districts threatens more than 200 km² of semi-natural habitat in the area just to the south of the area in which the mangabey lives. The insecurity of the area and the lack of protection to the forests on which the mangabey depends could move this species into the Critically Endangered category in the near future (Butynski et al., 2008d; Wieczkowski & Butynski, in press).
Figure 29. Cercocebus galeritus at Mchelelo Research Station, Tana River Primate National Reserve, Kenya.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 83
IUCN Red List: Endangered C2a(ii) ver 3.1 (Butynski et al., 2008d; www.iucnredlist.org/details/4200) Assessors: T.M. Butynski, T.T. Struhsaker, J. Kingdon & Y.A., de Jong Discussion: During this study there were no data collected for C. galeritus. During a primate survey in June, 2006, at Mchelelo Research Station, were adult male loud calls (‘whoop-gobble’) of C. galeritus were recorded and sent to Prof. Carolyn Ehardt, Co-Project Leader of the CEPF-funded project ’Taxonomy and Conservation Genetics of the Threatened Mangabey Taxa of the Eastern Arc Mountains and Coastal Forests of Tanzania and Kenya Biodiversity Hotspot’ (a joint project with T. M. Butynski). The recordings were also sent to Dr. Jean-Pierre Gautier, the authority on the vocalizations of Cercocebus.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 84
4.2.12 Hybrid Chlorocebus pygerythrus hilgerti x Cercopithecus mitis albogularis Field description: Slender, medium-size, greyish monkey (Figure 30). Bare, dark grey, skin around eyes and along top of muzzle. Cheeks and brow band pale grizzled grey. Ears dark grey with narrow pale grey. Throat patch and ventrum off white. Shoulders and outer limbs grizzled mouse grey. Lower outer front limbs slightly darker grey. Inner limbs light grey. Hands and feet dark grey. Crown, dorsum, sides and proximal ca. 10% upper part of tail grizzled greyish-olive with rufus wash. Remainder of tail grading into dark grey or blackish. Pelage around the ischial callosities pale grey with rusty wash. Scrotum dark grey to dark purple, partly hidden under the pelage. Distribution: Leopard Beach Hotel, Diani, Kenya (Figure 31). Figure 31. Encounter site of C. p. hilgerti x C. m. albogularis hybrid. Diani, Kenya (14 December, 2008).
Encounter sites: - Diani, S4.2848; E39.5913, 16 m asl Encounter rate: One adult male C. p. hilgerti x C. m. albogularis hybrid was encountered once (14th December 2008 at 12.45 h) during a four days period. The hybrid was with two adult C. m. albogularis of which at least one was a male.
Photographic map: Photo map contains five photographs (T. M. Butynski & Y. A. de Jong). http://picasaweb.google.com/wildsolutions/ChlorocebusPygerythrusHilgertiXCercopithecusMitisAlbogularisHybrid?authkey=Gv1sRgCMqKmI2R_d6zXA&feat=email# or via: www.wildsolutions.nl Vegetation: The hybrid was encountered in the gardens of the (large) Leopard Beach Hotel, about 100 m from the Indian Ocean. The vegetation includes many exotic and indigenous plants (including Terminalia sp, Cocos nucifera, Plumeria sp.). The hybrid and the two C. m. albogularis were feeding on flowers and human foods around the tourist rooms. Altitude: 16 m asl
Figure 30. Adult male Chlorocebus pygerythrus hilgerti x Cercopithecus mitis albogularis hybrid on a large beach hotel compound in Diani, Kenya.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 85
Discussion: Only one C. p. hilgerti x C. m. albogularis hybrid was encountered during this study. Although C. p. hilgerti and C. m. albogularis are sympatric at many sites, there are no reports of hybrids between these two species or between these two genera in the wild (Lernould, 1988; Erhart et al., 2004; Lawes et al., in press). C. pygerythrus and C. mitis are ecologically, morphologically, behaviourally and genetically very different from each other (e.g., Dutrillaux et al., 1980; Erhart et al., 2004). Table 13. presents a comparison of the phenotypic characters of C. p. hilgerti x C. m. albogularis, C. p. hilgerti, and C. m. albogularis. Bernstein (1971, cited in Gray, 1972) reported a dead full-term Cercopithecus aethiops x Cercopithecus mitis foetus at the Yerkes Regional Primate Research Center Field Station, Lawrenceville, Georgia, USA. Erhart et al. (2004) report two male Chlorocebus pygerythrus x Cercopithecus albogularis (in this study referred to as Cercopithecus mitis) hybrids, born in 1981 and 1986, which were the subject of their behavioural study at the University of Texas in Austin, USA. The parents originated in Uganda. One of the findings of their study is that the developmental trajectories of the hybrid males were more similar to C. albogularis than to C. pygerythrus (the mothers of the hybrids were C. pygerythrus). No behavioural observations were conducted on the Diani hybrid during our study but this particular hybrid was encountered with two C. m. albogularis. Table 13. Phenotypic comparison of the Chlorocebus pygerythrus hilgerti x Cercopithecus mitis albogularis hybrid, Chlorocebus pygerythrus hilgerti and Cercopithecus mitis albogularis using field descriptions and photographs obtained at Diani Beach, Kenya. Phenotypic charac-teristics
pygerythrus x mitis Chlorocebus pygerythrus hilgerti
Cercopithecus mitis albogularis
Photo
Ear fringe Narrow pale grey None Narrow, grey, some whitish
Cheeks Medium long hair, grizzled pale grey.
White, medium long whiskers that sweep upwards and backwards covering part of the ear.
Medium long hair, grizzled pale grey.
Face Triangular-shaped area of bare grey skin around eyes and nose. Grizzled pale grey pelage around the mask.
Jet black, hairless Grizzled grey. Bare black skin around eyes.
Eyes Mahogoney brown. Mahogoney brown. Light brown Brow band Projecting forward and
narrow. Grizzled pale grey.
White, narrow and short Projecting forward and upwards. Grizzled pale grey
Crown Grizzled olive-grey with pale rufus wash.
Grizzled olive-grey. Grizzled greyish-olive with pale rufus wash

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 86
Collar Vague and not sharply
defined. Around lower ca. 50% of the neck. Throat off white. Sides of neck mouse grey.
Absent Present but not distinct or sharply defined. Around lower ca. 50% of the neck. Throat off-white. Sides of neck light grey.
Throat Off white throat-patch Off white Off white throat-patch Shoulders Grizzled mouse grey Grizzled mouse grey Grizzled grey, becoming
blackish on the arms. Outer arms
Grizzled mouse grey. Lower arms slightly darker grey.
Grizzled mouse grey Black.
Inner arms Inner limbs light grey. Off-white Blackish. Outer legs Grizzled mouse grey. Grizzled mouse grey grizzled grey Inner legs Inner limbs light grey. Off-white pale grey. Hands and feet
Dark grey. Black in males. Dark grey in females and young.
Black
Back of legs
Hair slightly longer. Off white to pale grey. Rusty wash around ischial callosities.
Off white to pale grey. Red patch under base of tail in males.
Hair longer. Off white with rufus wash.
Dorsum Grizzled olive-grey with rufus wash extending unto the upper tail.
Grizzled olive-grey. Grizzled grey between shoulders. Below shoulders grizzled rusty extending onto the upper tail.
Ventrum Off white Off white Grey. Center of the ventrum pale grey.
Sides Grizzled olive-grey with rufus red wash.
Grizzled olive-grey. Narrow off-white stripe between side and ventrum
Mouse grey
Tail Grizzled olive-grey with rufus wash over the proximal 10% of upper tail (10%) grading into dark grey then blackish distally.
Grizzled mouse grey, distal 10% darker grey. Red patch under base of the tail in males
Proximally rusty red from dorsum. Distal part black. Intense red around base of tail.
Scrotum Purple to dark grey. Partly hidden behind fur.
Bright turquoise to blue. Dark grey, largely hidden under fur.
Overall Very grey animal. Body shape most resembles C. p. hilgerti. Generally intermediate in colour between C. p. hilgerti and C. m. albogularis.
Phenotypically like other populations of C. p. hilgerti.
Phenotypically like other populations of C. m. albogularis.
Future research priorities:
• Are there other reports of wild hybrids between C. p. hilgerti and C. m. albogularis and do they phenotypically resemble the ‘Diani hybrid’?

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 87
5. DISCUSSION During this study we encountered 99 groups of diurnal primates and more than 60 nocturnal primates in the coastal forest of Kenya. We recognise nine species and 11 primate subspecies in the coastal forests of Kenya. All 11 subspecies are described and discussed in separate sections in Chapter 4.2. Papio cynocephalus ibeanus was the most often encountered diurnal primate in the coastal forests of Kenya (32 groups), followed by Cercopithecus mitis albogularis (24 groups) and Cercopithecus mitis albotorquatus (16 groups). Three nocturnal primates species occur in the coastal forest of Kenya, Otolemur garnettii lasiotis and Galagoides cocos are common, Galago senegalensis braccatus is present but rare in this part of Kenya. This study is the first to report a wild Cercopithecus mitis albogularis x Chlorocebus pygerythrus hilgerti hybrid. This study resulted in additional insights into the geographical range of all 11 subspecies. Considerable geographical range extensions for O. g. lasiotis, G. cocos and C. m. albotorquatus were found. Field descriptions were obtained of almost all subspecies and these have been compared with the descriptions of individuals encountered in parts of the geographic ranges outside of the coastal forests of Kenya. Data collected during this study, combined with museum and literature research, will be used to write additional articles and notes on the distribution, diversity, taxonomy, and conservation status of six primate species (Papio anubis, Papio cynocephalus, Cercopithecus mitis, Chlorocebus pygerythrus, Otolemur garnetttii, and Galago senegalensis) in Kenya and Tanzania, and in the ‘Kenya Primate Atlas’ that we are now compiling. Primate encounters during this project formed the basis of our distribution maps, and species databases. In addition, locality data for all primate taxa occurring in Kenya have been obtained from a thorough search of the literature and museum specimens, as well as from communications with other field workers, and entered into these databases. Each Microsoft Access database holds one primate species and its subspecies. The databases will be used to compile and maintain distribution maps for each primate taxon. These databases will be updated soon after new information is received. As such, they will serve as ‘living’ databases to support primate research and conservation initiatives, such as the periodic IUCN Red List degree of threat assessments that are undertaken by the IUCN/SSC Primate Specialist Group. These databases will be made accessible on our website (wildsolutions.nl). Priority research questions for the primates of the coastal forests of Kenya include:
1) To what extent does phenotypic variation in O. g. lasiotis occur over its geographic range?
2) What is the southern boundary of O. g. lasiotis? 3) What is the geographical range of G. s. braccatus within and outside the coastal forests
of Kenya? , 4) How far up the Tana River does the range of G. cocos extent? 5) Which subspecies of C. mitis occurs between the Tana River/Delta and the Galana
River? 6) What subspecies of C. mitis occurs between the Galana River and Kilifi Creek? 7) What is the geographical range of C. m. phylax? 8) Is C. m. phylax a valid subspecies? 9) What is the geographic range of C. p. hilgerti? 10) Is C. p. excubitor a valid subspecies? 11) What is the geographic range of C. p. excubitor? 12) What is the southern limit of the geographic range of C. m. albotorquatus?

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 88
13) Are there other reports of wild hybrids between C. p. hilgerti and C. m. albogularis and do they phenotypically resemble the ‘Diani hybrid’?
14) Does Galagoides zanzibaricus occur in Kenya? 15) What primate taxa occur in the forests along the extreme northern coast of Kenya
(especially Boni-Dodori Forest)? 16) What primate taxa occur on Patta Island? 17) What is the taxonomic status of Papio in the forests along the Lower Tana River?
We established a website – www.wildsolutions.nl; De Jong & Butynski (2009), ‘Primate Biogeography, Diversity, Taxonomy and Conservation in Eastern Africa’ to communicate our photos, audio recordings, species databases, publications, and other data/information to colleagues and institutions. This website will be updated on a regular basis, and will be used by T. M. Butynski and Y.A. de Jong as a tool and resource for their research and conservation projects in eastern Africa. The combination of gradual climatic desiccation and increasing human activities accounts for much of the forest loss along the coast of East African (Clarke & Karoma, 2000). The majority of the coastal forests in Kenya are gazetted as Forest Reserves (Rodgers & Burgess, 2000a), however, the problems that occur in forest management and protection often go hand-in-hand with conflicts of interests among a) the local people who utilised the forest resourches in the past, b) commercial companies who exploit the forest, and c) the Forest Department which is responsible for forest protection and management (Burgess & Mbwana, 2000). As a result, the coastal forest of Kenya are under continuous and increasing threat. These forests are being degraded, fragmented, shrinking and disappearing. The expansion of agriculture is recognised as the most critical threat, while the production of charcoal, taking of firewood and timber, and the mining of limestone, lead, marble and several minerals is causing additional serious habitat loss (Obura, 2007). The diurnal, forest–dependent, primates are the most threatened primate taxa in the coastal forests of Kenya. Of the 11 primate subspecies present, 55% (n=6) are categorized as “Least Concern”, 9% (n=1; C. m. albotorquatus) is “Vulnerable”, 18% (n=2; P. r. rufomitratus and C. galeritus) are “Endangered”, and 18% (n=2) were not assessed for the 2008 IUCN Red List of Threatened Species. The conservation threats to all primate taxa and survey sites in the coastal forests of Kenya are described in Chapter 4.1 and 4.2. Conservation activities should focus on the areas with the highest biodiversity or with endemic taxa (e.g. Rodgers & Burgess, 2000b). Of the sites visited during this study, primate densities were very low in the Lamu Archipelago, but these Islands hold five species of primate and up to two endemic subspecies. Primate biodiversity is relatively high in the Tana River Primate National Reserve (seven species), Diani (six species), and Kipini Conservancy and Witu Forest Reserve (five species). Additional primate surveys in Boni and Dodori Forest Reserves, Patta Island, Kaya Gonja, Shimba Hills National Reserve, Buda Forest and Mrima Hill (nocturnal surveys) are essential for compiling a list of ‘Primate Priority Conservation Sites in the coastal forest of Kenya. There can be no doubt, however, that the top priority site for primate conservation in the coastal forests of Kenya, indeed for all of Kenya, are the forests along the Lower Tana River. These forests not only hold the highest diversity of primate species in all of Kenya, they hold Kenya only two endemic species of primate. In addition, they are among the most threatened forests in all of Kenya.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 89
ACKNOWLEDGEMENTS We are grateful to Conservation International and CEPF for their financial support. We thank the Kenyan Ministry of Education, Science & Technology for permission to conduct field work in Kenya, and John Salehe, Ian Gorden, Lorna Depew, John Watkin, Russ Mittermeier, Stephen Nash, Andrew Perkin, Nike Doggart, Kristina Razon, Farouk Sherman, Quentin Luke, Trish Luke, Jean-Pierre Dekker, Juliet King, Bernard Agwanda, Jake Butynski, Tine van de Velde, George Eshiamwata, Simon Bearder, Todd Olson, Carolyn Harcourt, Carolyn Ehardt, Arnoud de Jong, Leanne Nash, David Mbora, Julie Wieczkowski, Noel Rowe, Jonathan Kingdon, Dennis Milewa, Paul Honess, James Culverwell, Jim Feely, Sheila Bell-Cross, Iregi Mwenja, Helen Dufresne and Tom Struhsaker for their support to this Project.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 90
REFERENCES Alberts, S.C. & Altmann, J. 2001. Immigration and hybridization patterns of yellow and anubis baboons in and around Amboseli, Kenya. American Journal of Primatology 53: 139-154.
Altmann, J., Combes, S.L. & Alberts, S.C. in press. Papio cynocephalus. In: The Mammals of Africa (eds T.M. Butynski, J. Kingdon & J. Kalina). Elsevier, Amsterdam, Netherlands.
Anderson, J., Cowlishaw, G. & Rowcliffe, J.M. 2007. Effects of forest fragmentation on the abundance of Colobus angolensis palliatus in Kenya’s coastal forests. International Journal of Primatology 28: 637-655. Andrews P., Groves C.P. & Horne J.F.M. 1975. Ecology of the lower Tana River flood plain (Kenya). Journal of East African Natural History Society 151: 1-31. Gray, A. P. 1972. Mammalian Hybrids: A Check-list with Bibliography (Revised). Commonwealth Bureau of Animal Breeding and Genetics (Edinburgh). Technical Communication No. 10. Bearder, S.K., Butynski, T.M. & De Jong, Y.A. 2008. Galago senegalensis. In: 2008 IUCN Red List of Threatened Species. <www.iucnredlist.org>. Downloaded on 08 February 2009. Bearder, S.K., Honess, P.E. & Ambrose, L. 1995. Species diversity among galagos with special reference to mate recognition. In Creatures of the Dark: The Nocturnal Prosimians (eds L. Alterman, G.A. Doyle & M.K. Izard, M.K.), Plenum, New York. Pp. 331-52.
Bearder, S.K., Ambrose, L., Harcourt, C., Honess, P., Perkin, A.W., Pimley, E., Pullen, S. & Svoboda, N. 2003. Species-typical patterns of infant contact, sleeping site use and social cohesion among nocturnal primates in Africa. Folia Primatologica 74: 337-54. Bennun, L. & Njoroge, P. 1999. Important Bird Areas in Kenya. Nature Kenya (East Africa Natural History Society), Nairobi. Bocian, C.M. & Anderson, J. in press. Colobus angolensis. In: The Mammals of Africa (eds T.M. Butynski, J. Kingdon & J. Kalina). Elsevier, Amsterdam, Netherlands. Booth, C.P. 1963. Taxonomic studies of Cercopithecus mitis Wolf (East Africa). National Geographic Society, Research Report. Washington D.C. USA. Burgess, N.D., Clarke, G.P., Madgwick, J., Robertson, S.A. & Dickinson, A. 2000a. 3.1 Distribution and status. In: Coastal Forests of Eastern Africa (eds N.D. Burgess & C.P. Clarke). IUCN, Gland, Switzerland and Cambridge, UK. Pp 71-81. Burgess, N.D., Matthews, P., Evers, Y. & Woodcock, K. 2000b. Non-timber uses, threats and local attitudes. In: Coastal Forests of Eastern Africa (eds N.D. Burgess & C.P. Clarke). IUCN, Gland, Zwitserland and Cambridge, UK. Pp 281-302. Burgess, N.D & Mbwana, S.B. 2000. Forestry. In: Coastal Forests of Eastern Africa (eds N.D. Burgess & C.P. Clarke). IUCN, Gland, Zwitserland and Cambridge, UK. Pp 263-279.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 91
Butynski, T.M. 1995. Report says dam could threaten Kenya’s endangered primates. African Primates 1: 14-17. Butynski, T.M., Bearder, S.K. & De Jong, Y.A. 2008a. Otolemur garnettii lasiotis. In: IUCN 2008. 2008 IUCN Red List of Threatened Species. <www.iucnredlist.org>. Downloaded on 07 January 2009. Butynski, T.M. & De Jong, Y.A., 2004. Natural history of the Somali lesser galago (Galago gallarum). Journal of East African Natural History 93: 23-38. Butynski, T.M. & De Jong, Y.A. 2008. Cercopithecus mitis albotorquatus. In: 2008 IUCN Red List of Threatened Species. <www.iucnredlist.org>. Downloaded on 07 January 2009. Butynski, T.M., De Jong, Y.A., Perkin, A.W., Bearder, S.K. & Honess, P.E. 2006. Taxonomy, distribution, and conservation status of three species of dwarf galagos (Galagoides) in eastern Africa. Primate Conservation 21: 63-79. Butynski, T.M. & Koster, S.H. 1994. Distribution and conservation status of primates in Bioko Island, Equatorial Guinea. Biological Conservation 3: 893-909. Butynski, T.M. & Mwangi, G. 1994. Conservation status and distribution of the Tana River red colobus and crested mangabey. Unpublished report to Kenya Wildlife Service and Zoo Atlanta, Nairobi. Butynski, T.M. & Mwangi, G. 1995. Census of Kenya’s endangered red colobus and crested mangabey. African Primates 1: 8-10. Butynski, T.M., Perkin, A.W., Bearder, S.K., De Jong, Y.A. & Honess, P.E. 2008b. Galagoides cocos. In: 2008 IUCN Red List of Threatened Species. <www.iucnredlist.org>. Downloaded on 04 January 2009. Butynski, T.M., Struhsaker, T. & De Jong, Y.A. 2008c. Procolobus rufomitratus rufomitratus. In: 2008 IUCN Red List of Threatened Species. <www.iucnredlist.org>. Downloaded on 16 January 2009. Butynski, T.M., Struhsaker, T.T., Kingdon, J. & De Jong, Y.A. 2008d. Cercocebus galeritus. In: 2008 IUCN Red List of Threatened Species. <www.iucnredlist.org>. Downloaded on 15 January 2009. Clarke, G.P. 2000. Climate and climate history. In: Coastal Forests of Eastern Africa (eds N.D. Burgess & C.P. Clarke). IUCN, Gland, Switzerland and Cambridge, UK. Pp 47-67. Clarke, C.P. & Karoma, N.J. 2000. History of anthropic disturbance. In: Coastal Forests of Eastern Africa (eds N.D. Burgess & C.P. Clarke). IUCN, Gland, Switzerland and Cambridge, UK. Pp 251-261. Colobus Trust. 2009. Colobus Trust Blog. http://colobus.wildlifedirect.org/2008/04/04/destruction-of-the-diani-forest-update. Downloaded on 1 February 2009.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 92
Culverwell, J., Feely, J., Bell-Cross, S., De Jong, Y.A. & Butynski, T.M. 2008. A new pig for Tsavo, Swara 31: 50-52. Dandelot, P. 1968. The Primates. Preliminary Identification Manual for African Mammals. Smithsonian Institution, Washington, D. C. Dandelot, P. 1974. Order Primates. Part 3. In The Mammals of Africa: An Identification Manual (eds J. Meester & H.W. Setzer). Smithsonian Institution Press, Washington, D.C. De Beaux, O. 1937. Spedizione zoological del Marchese Saverio Patrizi nel Basso Giuba e nell’Oltregiuba. Mammiferi. Annali Museo Civica Storia Naturale Genova 58: 150-173. De Jong, Y.A. & Butynski, T.M. 2004. Surveys of the Somali lesser bushbaby (Galago gallarum) and northern lesser bushbaby (Galago senegalensis) in Kenya and Ethiopia. Unpublished report to Primate Action Fund and Primate Conservation Inc., Washington, D.C.
De Jong, Y.A. & Butynski, T.M. 2009. Eastern Africa Primate Platform, Biogeography, Taxonomy, Diversity, and Conservation Status of Primates in Eastern Africa. www.wildsolutions.nl.
De Jong, Y.A., Culverwell, J. & Butynski, T. M. 2009. Desert warthog Phacochoerus aethiopicus found in Tsavo East National Park and Tsavo West National Park, southern Kenya. Suiform Soundings 8: 4-6. d’Huart, J. & Grubb, P. 2001. Distribution of the common warthog (Phacochoerus africanus) and the desert warthog (Phacochoerus aethiopicus). African Journal of Ecology 39: 156-169. Dowsett-Lemaire, F. & Dowsett, R.J. 2007. Surveys of the vegetation, birds and mammals of Kipini Conservancy (Lamu and Tana River districts), Kenya, October-December 2006. Report no. 55. Dutrillaux, B., Couturier, J., Chauvier., G. 1980. Chromosomal evolution of 19 species or subspecies of Cercopithecinae. Annales de Génétique 23: 133-143. Erhart, E.M., Bramblett, C.A., Overdorff, D.J. 2004. Behavioral development of captive male hybrid cercopithecine monkeys. Folia Primatologica 868: 1-11. Gartlan, J.S. & Brain, C.K. 1968. Ecological and social variability in Cercopithecus aethiops and C. mitis. In: Primates: Studies in Adaptation and Variability (ed. P.C. Jay). Holt, Rinehart, & Winston, New York. pp. 253-292. Gippoliti, S. 2002. Zammarano’s monkey Cercopithecus mitis zammaranoi De Beaux, 1923: The forgotten monkey of Somalia. African Primates 6: 26-32. Groves, C.P. 2001. Primate Taxonomy. Smithsonian Institution Press. Washington and London. Groves, C.P. 2005. Order Primates. In Mammal Species of the World (eds D.E. Wilson & D.M. Reeder) Johns Hopkins University Press.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 93
Groves, C.P. 2006. Taxonomy and biogeography of the primates of western Uganda. In: Primates of Western Uganda. Developments in Primatology: Progress and Prospects. (eds N.E. Newton-Fisher, H. Notman, J.D. Paterson & V. Reynolds) Springer. Berlin. Pp 3-20. Grubb, P. 2001. Synonyms reduce the number of subspecies in the guenon Cercopithecus mitis. African Primates 5: 24-32. Grubb, P., Butynski, T.M., Oates, J.F., Bearder, S.K., Disotell, T.R., Groves, C.P. & Struhsaker, T.T., 2003, Assessment of the diversity of African Primates. International Journal of Primatology 24: 1301-1357. Harcourt, C.S. 1984. The behaviour and ecology of galagos in Kenyan coastal forest. Ph.D dissertation, University of Cambridge, Cambridge, UK. Harcourt, C.S. & Nash, L.T. 1986. Species differences in substrate use and diet between sympatric galagos in two Kenyan coastal forests. Primates 27: 41–52. Harcourt, C.S. & Perkin, A. W. in press. Otolemur garnettii. In: The Mammals of Africa (eds T.M. Butynski, J. Kingdon & J. Kalina). Elsevier, Amsterdam, Netherlands. Heller, E. 1913. New races of ungulates and primates from British East Africa. Smithsonian institution 61: 1-13. Hill, W.C.O. 1966. Primates Comparative Anatomy and Taxonomy. VI Catarrhini Cercopithecoidea Cercopithecinae. Edinburgh, University Press. UK. Hill, W.C.O. 1967. Taxonomy of the baboon. In: The Baboon in Medical Research. Vol. 2 (ed. H. Vagtborg). University of Texas, Austin. pp. 4-11. Hill, W.C.O. & Meester, J. 1974. Infraorder Lorisformes. In: The Mammals of Africa: An Identification Manual (eds J. Meester & H.W. Setzer) Smithsonian Institution Press, Washington, D.C. Homewood, K. 1976. Ecology and behaviour of the Tana mangabey (Cercocebus galeritus galeritus). Ph.D. dissertation. University of London. Honess, P. E. 1996. Speciation among galagos (Primates, Galagidae). PhD dissertation, Oxford Brookes University, Oxford, UK. Isaac, N.J.B., Mallet, J. & Marce, G.M. 2004. Taxonomic inflation: Its influence on macroecology and conservation. Trends in Ecology and Evolution 19: 464-469. Irebelo. 2006. 2.5 Kenya. http://ramsar.wetlands.org/Portals/15/KENYA.pdf. Downloaded on 2 February 2009. IUCN 2008. 2008 IUCN Red List of Threatened Species. <www.iucnredlist.org>. Downloaded on 07 October 2008. IUCN 2001. IUCN Red List Categories and Criteria, Version 3.1. IUCN The World Conservation Union, Gland, Switzerland.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 94
Jenkins, P.D. 1987. Catalogue of Primates in the British Museum (Natural History) and elsewhere in the British Isles. Part IV: Suborder Strepsirrhini, including the subfossil Madagascan lemurs and Family Tarsiidae. British Museum, London. Jolly, C.J. & Brett, F.L. 1973. Genetic markers and baboon biology. Journal of Medical . Primatology 2: 85-99 Jolly, C.J. 1993. Species, subspecies, and baboon systematics. In: Species, Species Concepts, and Primate Evolution (eds. W. H. Kimbel & L. B. Martin). Plenum Press, New York. Pp. 67-107. Jolly, C.J. 1997-1998. Baboon nomenclature. African Primates 3: 47-48. Kairo, J.G. & Bosire, J. 2007 General introduction. In: UNEP, Kenya, State of the Coast 2007 Report. Nairobi, Kenya. Pp 1-7. Kamula, J. & Ochiewo, J. 2007. Coastal communities. In: UNEP, Kenya, State of the Coast 2007 Report. Nairobi, Kenya. Pp 37-47. Kamilar, J. M. 2006. Geographic variation in savanna baboon (Papio) ecology and its taxonomic and evolutionary implications. In: Primate Biogeography (eds. M.L. Shawn & J.G. Fleagle) Springer, New York. Pp. 169-200. Kingdon, J. 1971. East African Mammals: An Atlas of Evolution in Africa. Volume I. Academic Press, London. Pp. 446. Kingdon, J. 1997. The Kingdon Field Guide to African Mammals. Academic Press, San Diego, CA. Pp. 464. Kingdon, J. 2008. Papio cynocephalus ibeanus. In: 2008 IUCN Red List of Threatened Species. <www.iucnredlist.org>. Downloaded on 08 February 2009. Kingdon, J., Butynski, T.M. & De Jong, Y.A. 2008a. Cercopithecus mitis albogularis. In: 2008 IUCN Red List of Threatened Species. <www.iucnredlist.org>. Downloaded on 08 February 2009. Kingdon, J., Butynski, T.M. & De Jong, Y.A. 2008e. Papio cynocephalus. In: 2008 IUCN Red List of Threatened Species. <www.iucnredlist.org>. Downloaded on 10 February 2009. Kingdon, J., Gippoliti, S., Butynski, T.M. & De Jong, Y.A 2008b. Chlorocebus pygerythrus. In: 2008 IUCN Red List of Threatened Species. <www.iucnredlist.org>. Downloaded on 08 February 2009. Kingdon, J., Gippoliti, S., Butynski, T.M., Lawes, M.J., Eeley, H., Lehn, C. & De Jong, Y.A. 2008c. Cercopithecus mitis. In: 2008 IUCN Red List of Threatened Species. <www.iucnredlist.org>. Downloaded on 07 January 2009. Kingdon, J., Struhsaker, T., Oates, J., Hart, J.F., Butynski, T. M., De Jong, Y. A. & Groves, C.P. 2008d. Colobus angolensis ssp. palliatus. In: 2008 IUCN Red List of Threatened Species. <www.iucnredlist.org>. Downloaded on 10 February 2009.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 95
Kenya Wildlife Service. 2009. http://www.kws.go.ke, downloaded on 31 January, 2009. Lawes, M.J., Cords, M. & Lehn, C. (in press). Cercopithecus mitis. In: The Mammals of Africa (eds T.M. Butynski, J. Kingdon & J. Kalina). Elsevier, Amsterdam, Netherlands. Lernould, J.M. 1988. Classification and distribution of guenons. In: A Primate Radiation: Evolutionary Biology of the African Guenons (eds. A. Gautier-Hion, G. Bourlier, J.P. Gautier & J. Kingdon ), pp 54-77. Cambridge University Press, Cambridge. Maples, W.R. 1972. Systematic reconsideration and revision of the nomenclature of Kenya baboons. American Journal of Physological Anthropology: 36: 9-20. Maples, W. & McKern, T. 1967. A preliminary report on classification of the Kenya baboon. In: The Baboon in Medical Research vol. II (ed. H. Vagtborg). University of Texas Press, Austin. Pp. 12-22. Marsh, C. W. 1978. Ecology and social organization of the Tana River red colobus (Colobus badius rufomitratus). Ph.D. dissertation, University of Bristol. Mbora, D.N.M. 2003. Habitat quality and fragmentation and the distribution and abundance of the Tana River red colobus monkey, Procolobus rufomitratus, in eastern Kenya (p. 145). Oxford, Ohio: Ph.D. thesis, Miami University. Mbora, D.N.M. & Butynski, T.M. 2007. Tana red colobus. In: Primates in peril: The world’s 25 most endangered primates (eds. Mittermeier, R. et al.). Primate Conservation 22: 1-40. Napier, P.H. 1981. Catalogue of Primates in the British Museum (Natural History) and elsewhere in the British Isles. Part II: Family Cercopithecidae, Subfamily Cercopithecinae. British Museum, London. Nash, L.T., Bearder, S.K. & Olson, T.R. 1989. Synopsis of galago species characteristics. International Journal of Primatology 10: 57–80. Nash, L.T. & Zimmermann, E. in press. Galago senegalensis. In: The Mammals of Africa (eds T.M. Butynski, J. Kingdon & J. Kalina). Elsevier, Amsterdam, Netherlands. Nekaris, K.A.I. & Jayewardene, J. 2004. Survey of the slender loris (Primates, Lorisidae Gray, 1821: Loris tardigradus Linnaeus, 1758 and Loris lydekkerianus Cabrera, 1908) in Sri Lanka. Journal of Zoology 262: 1-12. Newman, T.K., Jolly, C.J. & Rogers, J. 2003. Mitochondrial phylogeny and systematics of baboon (Papio). American Journal of Physical Anthropology 124: 17-27. Nielsen, M.R. & Sick, C. 2008. Conservation and use of Witu Forest, Kenya. Biodiversity and disturbance survey and management recommendations. The Danish Zoological Society, Forlaget BIOS, Copenhagen, Denemark. Obura, D. 2007. Coastal Ecosystems. In: UNEP, Kenya, State of the Coast 2007 Report. Pp 8-24.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 96
Off, E.C., Isbell, L.A. & Young, T.P. 2008. Population density and habitat preferences of the Kenya lesser galago (Galago senegalensis braccatus) along the Ewaso Nyiro River, Laikipia, Kenya. Journal of East African Natural History 97: 109-116. Olson, T.R. 1979. Studies on aspects of the morphology and systematics of the genus Otolemur Coquerel, 1859 (Primates: Galagidae). Ph.D. thesis, University of London, U.K. Olson, T.R. & Nash, L.T. 2002. Galago (Galagidae) body measurements and museum collections data. African Primates 6: 50-53. Palombit, R.A. in press. Papio anubis In: The Mammals of Africa (eds T.M. Butynski, J. Kingdon & J. Kalina). Elsevier, Amsterdam, Netherlands. Pocock, R.I. 1907. A monographic revision of monkeys of the genus Cercopithecus. Proceedings of the Zoological Society of London: 677-746. Polhill, D. 1988. Flora of Tropical East Africa. Index of Collecting Localities. Royal Botanic Gardens, Kew. Great Britain. Rahm U. 1970. Ecology, zoogeography and systematics of some African forest monkeys. In: Old World Monkeys. Evolution, Systematics and Behavior (eds J.R. Napier & P.H. Napier). Academic Press, London and New York: pp 591-626. Rodgers, W.A. & Burgess, N.D. 2000a. Coastal forest conservation problems. In: Coastal Forests of Eastern Africa (eds N.D. Burgess & G.P. Clarke). IUCN, Gland, Zwitserland and Cambridge, UK. Pp 303-315. Rodgers, W.A. & Burgess, N.D. 2000b. Taking conservation action. In: Coastal Forests of Eastern Africa (eds N.D. Burgess & G.P. Clarke). IUCN, Gland, Zwitserland and Cambridge, UK. Pp 317-334. Samuels, A. & Altmann, J. 1991. Baboons of the Amboseli Basin: Demographic stability and change. International Journal of Primatology 12: 1-19. Sarmiento, E. 1998. Current problems with Papio taxonomies. African Primates 3: 48-52. Schwarz, E. 1926. Die meerkatzen der Cercopithecus aethiops-Gruppe. Z. Saugetierkunde. 1: 28–47. Schwarz, E. 1931. On the African short-tailed lemurs or pottos. Annals and Magazine of Natural History (London), Ser. 10: 249-256. Struhsaker, T.T. & Grubb, P. In press. Procolobus rufomitratus. In: The Mammals of Africa (eds T.M. Butynski, J. Kingdon & J. Kalina). Elsevier, Amsterdam, Netherlands.
Unesco, 2009. Sacred Mijikenda Kaya Forests. http://whc.unesco.org/en/list/1231, downloaded on 2 February 2009.

Y.A. de Jong & T.M. Butynski, 2009.
Primates of the coastal forests of Kenya. 97
Wieczkowski, J.A. & Butynski, T.M. in press. Cercocebus galeritus. In: The Mammals of Africa (eds T.M. Butynski, J. Kingdon & J. Kalina). Elsevier, Amsterdam, Netherlands. White, F. 1983. The Vegetation of Africa. A Descriptive Memoir to Accompany the UNESCO/AETFAT/UNSO Vegetation Map of Africa. UNESCO, Paris, France. White, L. & Edwards, A. 2000. Methods for assessing the status of animal populations. In: Conservation Research in the African Rain Forests: A Technical Handbook (eds L. White & A. Edwards). Wildlife Conservation Society, New York. Pp. 191-201. Wolfheim, J.H. 1983. Primates of the World, Distribution, Abundance and Conservation. University of Washington Press, Washington. Younge, A, Negussie, G. & Burgess, N. 2002. Eastern Africa Coastal Forest Programme. Regional Workshop Report, Nairobi, Kenya. Zimmermann, E. 1990. Differentiation of vocalizations in bushbabies (Galaginae, Prosimiae, Primates) and the significance for assessing phylogenetic relationships. Zeitschrift fuer zoologische Systematik und Evolutionsforschung 28: 217-239. Zimmermann, E. 1995. Loud calls in nocturnal Prosimians: structure, evolution and ontogeny. In: Current Topics in Primate Vocal Communication (eds Zimmermann et al.) Plenum Press, New York. Pp 47-72). Zinner, D., Kraft, R. & Roos, C. 2008. The phylogenetic position of “Papio ruhei” – a unique baboon taxon from Somalia. Zoological Garten N.F. 77: 303-311.

Appendix 1. Distinguishing character tables (draft) of Papio anubis, Papio cynocephalus, Chlorocebus pygerythrus, Cercopithecus mitis, Colobus angolensis, Colobus guereza and their subspecies, in Kenya and Tanzania. Distinguishing characters in Papio anubis - DRAFT P. anubis P.a. furax P.a. neumanni Face
- Colour cheek - Colour naked
skin
Purple-black (1), naked, dark grey to black (3), ruffs of hair at cheeks (3), dark grey (4), black (5), hair on cheeks prominent (5)
Eastern Uganda and western Kenya (above the Chanler Falls on the Northern Ewaso Ngiro).
Below the Chanler Falls on the Ewaso Ngiro, then southward through Kenya and Tanzania.
Mane Short to medium (1), male large manes (4), Upper parts Olive-brown (1), dark grey to olive-brown sometimes grading into olive or
light brown (khaki or grey) (3), Darker than neumanni (6) Pattern and colour generally similar (6)
Under parts Olive-brown (1), dark grey to olive-brown sometimes grading into olive or light brown (khaki or grey) (3), dark (5)
Tail Broken (1), hairy, sometimes bent at acute angle midway appearing as if broken (3), directed up for proximal one-fourth then falls away as if broken (4), blackish (5)
Limbs Hands and feet black in some populations (3), feet olive-brown (1), forelimbs blackish (5)
Pelage overall Olive-brown (1), dark grey to olive-brown sometimes grading into olive or light brown (khaki or grey) (3), olive brown (4), furax darker than neumanni (?), dark greyish brown, fading into black (5)
Male - Female Canines smaller in females (3) Nostrils Protruding beyond plane of upper lip (6) Other Shorter limbed, more robust and muscular (5) Shorter muzzle than neumanni (6) Literature 1. Sarmiento, E. 1997. Current problems with Papio taxonomies. African Primates 3: 48-52 2. MOA ms, Papio cynocephalus draft 2002 3. MOA ms, Papio anubis draft 2003 4. Groves, C. 2001. Primate Taxonomy p.238-240. 5. Maples, W.R. 1994. Systematic reconsideration and a revision of the nomenclature of Kenya Baboons. Am. J. Phys. Anthrop. 36: 9-20 6. Dandelot 1971. An identification manual Primates In: Meester, J, & Setzer, H.W. The mammals of Africa. p.

Distinguishing characters in Papio cynocephalus - DRAFT P. cynocephalus P.c. typical P.c. ibeanus P.c. kinda Face
- Colour cheek - Colour naked
skin
Purple-black (1), muzzle greyish to black (2), cheeks almost white and hair sparse (2), cheeks white (4), cheeks very pale approaching a yellow-white(5), hair short on top of head and lays flat (5)
Mikumi (T), Ruaha (T) Amboseli (K), Tana River (K) Angola, Zambia (west of Typical and Ibean)
Mane None, nape and neck yellow-fringed (1), no manes (4) Upper parts Brownish-yellow (1), light brown to yellow brown (2), yellow to yellow-brown (4). Juvenile
with whitish coat reddish-brown (2)
Under parts Almost white and hair sparse (2), white (4) Tail Variably arched (1), light brown to yellow brown (2), long tail tapered and sharp bend or
hook, very variable (2), varying from simply curved in P. c. Kindae to more usually broken but held low (4)
Curved (4)
Limbs Longer limbs than anubis (2), outer limbs light brown to yellow brown (2), inner limbs almost white and hair sparse (2)(4).
Pelage overall Brownish-yellow (1), long (2), straight and soft, silky pelage (2)
wavier and coarser pelage (2)
straight and soft, silky pelage (2)
Male - Female Females 50% of male body size and 80% of limb length (2), Other Armpit and crotch colour ranges from pinkish to almost white (2). More variable in colour
than anubis, from fairly bright yellow with black bands on the hairs to a rufus brown (5), fur long but less dense than anubis.
Literature 1. Sarmiento, E. 1997. Current problems with Papio taxonomies. African Primates 3: 48-52 2. MOA ms, Papio cynocephalus draft 2002 3. MOA ms, Papio anubis draft 2003 4. Groves, C. 2001. Primate Taxonomy p.238-240. 5. Maples, W.R. 1994. Systematic reconsideration and a revision of the nomenclature of Kenya Baboons. Am. J. Phys. Anthrop. 36: 9-20

Distinguishing characters for Chlorocebus pygerythrus – DRAFT.
1. Groves, 2001. Primate Taxonomy, p. 202-203, 2. MOA ms Cercopithecus (a) pygerythrus, draft 03, 3. Lonnberg, 1913 Mammals collected by the Swedish zoological expedition to East Africa. K. Svenska Vetensk Aixad, Handl. 48(5): 37-39, 4. Dandelot 1971. An identification manual Primates In: Meester, J, & Setzer, H.W. The mammals of Africa. P. 27-29., 5. Hill, O.W.C. 1953. Primates. Comparative Anatomy and Taxonomy. 6. Pocock, R.I. 1907. A revision of the monkeys of the genus Cercopithecus, Proc. Zool. Soc London
Species C. tantalus C.p. excubitor C. aethiops C.p. johnstoni Distribution Volta River to Sudan, Uganda and L. Turkana
district. Witu Islands, Kenya Sudan & Ethiopia NE Tanzania & Kenya
Limbs Grey (1). Lateral sides mouse grey Arms: forearm to elbow and on legs from hindlimb to knee purely mouse grey. Upperarm speckled grey with black and the tight white (5)
Laterally more grey than body (5) tights yellowish wash (5). Greyer than body (6) Legs sometimes washed with yellow (6)
Hand/Feet Dorsum of h/f darker Black (5) Wrists and h/f black. Feet less black than hands (5). Black, also the wrists, sharply defined (6) Feet less black than hands (6)
Under parts White (1) Cream (5) Pale olive-yellow (4) Dirty white (5)(6) Upper parts Grizzled, gold to greenish (1). Golden green (5) Resembling hilgerti (1) Grey-green, frequently with yellowish
tinge. Lighter bands of dorsal hairs between dark olive-buff and old gold (5)
Washed out tawny grey (5)(6)
Tail Creamy white tip, white basal tuft (1). Long shiny hairs (5). Whitish/yellow upper side and lightly speckled with black tips underside. Terminal end creamy white (5) white tuft at root (5)
Above dark grey with basal part showing extension of back colour, terminal end black, beneath yellowish white with red patch at root (5)
End of tail whitish (4) Grey above, pure black at tip, rusty red at base (5). Grey above, scarcely washed with yellow black at tip (6) Large rusty rusty-red basal patch
Face Black (1) Black line from corner eye running back along temples (1)
Whitish moustache Jet black (5)(6)
Brow band & frontal band
White (1). Frontal band distinct, narrow in the centre and white (5) separate from the whiskers (5). Long black brow hairs border it in front (5)
Distinct but narrow (5) Frontal band greyish white to tawny white (5). Greyish-white or tawny-white browband (6)
Cheek whiskers
Long, stiff and yellowish with short black tips (1). White with yellow at tips, directed upwards and backwards conceiling the ears (5) Ends of longer hairs speckled black & yellow, tips black (5)
Whiskers long, falciform, pure white (4) Long whiskers passing gradually above into colour of crown (5). Greyish-white or tawny-white extending from browband (6) Long cheek hairs overlaying ears (6)
Crown Grizzled, gold to greenish (1). Golden green (5) Washed out tawny grey (5) Neck Ears Body size Smaller than hilgerti (2)(4) Scrotum Sky blue (1) Turquoise blue Overall Sides golden green (5) rusty patch on pubic and
perineal area and around scrotum (5) Differs from hillgerti in that there is no
rufous in the pelage (5)

Distinguishing characters in Chlorocebus pygerythrus - DRAFT
1. Groves, 2001. Primate Taxonomy, p. 202-203, 2. MOA ms Cercopithecus (a) pygerythrus, draft 03, 3. Lonnberg, 1913 Mammals collected by the Swedish zoological expedition to East Africa. K. Svenska Vetensk Aixad, Handl. 48(5): 37-39, 4. Dandelot 1971. An identification manual Primates In: Meester, J, & Setzer, H.W. The mammals of Africa. P. 27-29., 5. Hill, O.W.C. 1953. Primates. Comparative Anatomy and Taxonomy. 6. Pocock, R.I. 1907. A revision of the monkeys of the genus Cercopithecus, Proc. Zool. Soc London
Species C.p. arenarius C.p. callidus C.p. centralis Distribution N. Kenya, from Tana to L. Rudolf, Somalia, central
Ethiopia Tanzania, Kenya between L. Victoria and Rift Valley to Mt Elgon and Mt Debasien; Elgeyo and Mau esc.
NW Tanzania, SW Uganda, Rwanda, Burundi, DRC.
Limbs Deep mouse grey, upper arms tinge of dorsal colour (5)
Laterally iron grey (5) Light iron grey laterally, sharp demarcation with dorsal (5). Forearm below elbow and hindleg below knee ashy grey speckled (6)
Hand/Feet Deep brownish-black (5) Black (4). Almost completely black (5) Wrist, H/F black mixed with grey (5). Wrist and hand black, ankle grey, foot blackish (6)
Under parts Well haired dirty white (5) Brownish fawn (4) Upper parts Pale olive green (5) Paler than johnstoni (5) Greenish (5). Greenish speckled with black (6) Tail Brownish-red at root tail (5) Bicolour, grey above
with greenish tinge; below white or yellowish white. Top black (5)
Grey speckled above. Greenish of back continues on tail (5). Terminal end black (5)
Grey with yellow speckling near root dorsally. Terminal end black (5). Few rusty hairs at root (5)(6)
Face Black (5) Chin black (5) Nose & muzzle with black hairs Brow band & frontal band Black brow hairs (5) Cheek
- whiskers
White (5) Not white, individual variation (5) Short, not concealing ears (5)(6)
Crown Pale olive green (5) Greenish (5) Neck Ears Body size Scrotum Surrounded with white hairs (5) Overall Long pelage esp. on shoulders (5). Pubic area tinged with red
(6)

Distinquishing characters in Cercopithecus mitis - DRAFT
Species albotorquatus kibonotensis stuhlmanni elgonis L. l. mauae C. l. phylax C. kolbi hindei Distribution Tana River (Kenya) S. Kenya & N. Tanz to Kilifi,
Taitas, Mazeras W. Kenya & Uganda W. Kenya & Highlands & U Mau (Kenya) Patta & Witu Island (K) Tutha, Kenya district ?
Under base of tail Red patch extending to thighs sometimes present (2)(3)
Russet tuft (2)
Under parts Dark ashy grey, buffy, grey with buffy median line, grey with tufts of buff (2), grey (4), ashy grey or cream (7). Anterior part of chest snow white (9)
olive grey, ashy grey or black speckled, grey speckled with white, black (2), grey white (6). Blackish chest (10)
Black (2)(3) White (2)
Upper parts Orange suffusion rump (3), speckled grey with a wide band of russet (4), pale yellow (5), dark olive (7). Speckled yellow (9)
Darker than under parts (3), frosty grey (6), steel blue grey sometimes faintly greenish (7)
Pale (2) Olive (2) dark reddish-brown (2)
Crown Very yellow or not yellow (2), dark olive (7). Head speckled yellowish-grey with black (9)
Black (3)(7) Black, distinctive with brow band(10). Back of the head black (10)
Dark (3) Reddish-yellow (9)
Brow-band Dark olive (7) Grey – black, varies a lot(3), grey (7), grey white (2) Speckled as cheeks (10)
Ear - tufts
White (2), ear tufts yellowish, pale-yellowish-white, no tufts, short grey (2) usually white (3) Almost bare ears, little hair (10)
No ear tufts (2)(3) Yellowish and banded (9)
Cheeks Whitish whiskers, face black (6). Speckled and sides of neck as brow band(10)
Face dark olive (2)
Collar broad & white (almost complete (2)(3), ¾ (4) not always present (7). Snow white half collar (9)
small white (2), not complete (3) No collar (10) Black (3) white & partly (2) Less less white and less distinctive than kolbi (9)
Throat White (4)(7). Snow white contrasting with grey of cheeks (9)
Less white on throat than Kolbi , it does not reach down to neck (good indicator)(8)
Grey white (6), white (7) Greyish-white patch (10)
black patch (2)
Shoulders Dove-grey (5), dark olive (7). Speckled yellowish-grey with black (9). Nape speckled yellowish-grey with black (9)
wholly black from crown to nape, speckled, unspeckled (2) thick mass of speckled hair on shoulders from cheeks (10)
speckled (2)
Limbs sometimes dark grey (2), dark grey (4), dove-grey (5). Inside fore leg ashy grey becoming sooty grey towards wrists. Outer limbs speck smokey grey (9). Inner limbs pale grey to white(9)
Hindlegs are much blacker than kolbi (8)
sometimes speckled down to feet, black, dark grey (2), partly black (3)
dark hind limbs (2)(3) Legs mouse grey (2), arms black (2)
Tail Dark grey (4). Base speckled yellow for 2 inch, gradually rufouslaterally and inferiorly, rest dark speckled. Term end black (9) Rusty red at root above and outside (9)
Terminal half black (3) 0-60% of distal black (2), frosty grey (6)
less black & speckled (2)
Other Inner thighs may be reddish-brown (7). Lumbar region dark brown, red brown, deep ochre, reddisch ochre, olive speckled (2). Cheeks speckled yellowish-grey with black (little greyer than head) (9)
Very like albogularis (3), neck black (7). No brown color in fur but grey black (10) Eyes dul muddy brown (10)
Smaller in size (3) Cheeks reddish-yellow (9). Co

Distinquishing characters in Cercopithecus mitis - DRAFT
Literature 1. Dandelot 1971 In: Meester, J, & Setzer, H.W. The Mammals of Africa. p. 18-21., 2. Grubb, P. 2001. Synonyms reduce the number of subspecies in the guenon Cercopithecus mitis. African Primates 5: 24-33., 3. Groves, C. 2001. Primate Taxonomy p.206-211., 4. Harcourt, C.S. 1974. Ecology & social organisation of the Sykes monkey in the Tana River Gallery Forest (MS). 5. Kingdon, J. 1984. East African Mammals. Pp. 234-247., 6. Rudran, R. 1978. Socioecology of the Blue Monkeys (C.m. stuhlmanni) of the Kibale Forest, Uganda. 7. Lawes, Cords & Lehn (ms) Cercopithecus mitis, Mammals of Africa., 8. Lonnberg, 1913 Mammals collected by the Swedish zoological expedition to East Africa. K. Svenska Vetensk Aixad, Handl. 48(5): 37-39. 9. Pocock, R.I. 1907. A revision of the monkeys of the genus Cercopithecus, Proc. Zool. Soc London 10. Booth, 1963 Nat. Geo soc. Research report.
Species neumani moloneyi monoides kolbi C.a. albogolaris doggetti
Distribution Nyando Valley, 30 miles east of Kisumu
Mtn region of S. Tanz., Zambia, Malawi
Coastal forest T (SE) Kenya highlands east of rift Zanzibar (T) NE Tanzania, DRC, SW Uganda, S Burundi, Rwanda
Under base of tail Reddish hair (3) Reddish hair (3) No red-brownish patch in adults, younger animals do show this patch (8)
Russet tuft (3) Reddish (3)
Under parts speckled grey (2) Dark (1), dark slate grey (7). Smokey grey (9)
Dark (7) Not black (1)
Upper parts bright russet (1), dark red saddle (7). Ruddy chestnut brown (9)
Reddish-brown (3)(7). speckled and rich reddish orange-yellow (9)
Redder than albogularis (3)(7), dark red (5), russet (7)
Reddish-brown (3) Light-silvery brown (3), grizzled grey (5), light grey brown (7)
Crown Dark grey (3). Blackish speckled with yellow (9)
Yellowish grey (3), yellowish-olive (7). Head thickly speckled with black (9)
Darker than body (3) Black (3)(5)(7)
Brow-band narrowly banded, dusky grey (2)
Light speckled grey (3). White above brow (9)
Yellowish gray (3) Brow fringe (3) Pale & grizzled (5)
Ear - Tufts
Grey (2) Tufts present (8) Tufts Long & bright (5)(7), white (8)(9)
Cheeks Whiskers light speckled grey (3). Cheeks speckled greyish yellow (9)
Cheek whiskers yellowish grey (3), cheeks yellowish-olive (7)
Greyish (2), cheek whiskers (3) Long grizzled whiskers (5)
Collar No collar (10) Creamy (3) No whitish half colar (9) Nearly complete (3), broad snow-white (5)(7) Less sharp definitions between dark and white compared to albotorquatus (9)
Nearly complete (3)
Throat Patch creamy (3)(7) White (1), little patch (3), varies in size (7)
White (2) (3)
Shoulders Speckled with greyish yellow (9). In front of shoulders black (9)
Yellowish-olive (7). Behind shoulders speckled and rich reddish orange-yellow (9)
Black (5) Greyish (2), speckled yellowish grey (3)
Limbs slightly speckled (2) Thighs speckled grey (9). Black upperarms (9). Grey speckled (9) H/F black speckled (9)
Thighs dark grey speckled (9) Hind legs speckled, looking dark grey (8) Reddish tinge on hindlegs
Dark grey, shiny black (5), hindlimbs blackish grey (7)
Tail 66% of distal half black (3), ventral surface reddish (7) Black, speckled towards base, reddish above root (9)
40% of terminal end black (3) No red on on anal region and base tail (9)
60% distal part black 30% terminal end black
Other Speckled gray with dark reddish saddle (3), nape dark grey (3), sides and thighs light grey (7)(1). Sides speckled grey (9)
Neck yellowish-olive (7). Neck thickly speckled with black (9) Closely resembling C. a. albogularis (9)
Longer fur than albotorquatus (9) Nape black (3)(7). Buff speckled frontal band (3). Possibly the closest resemblance with ancestral type (5)
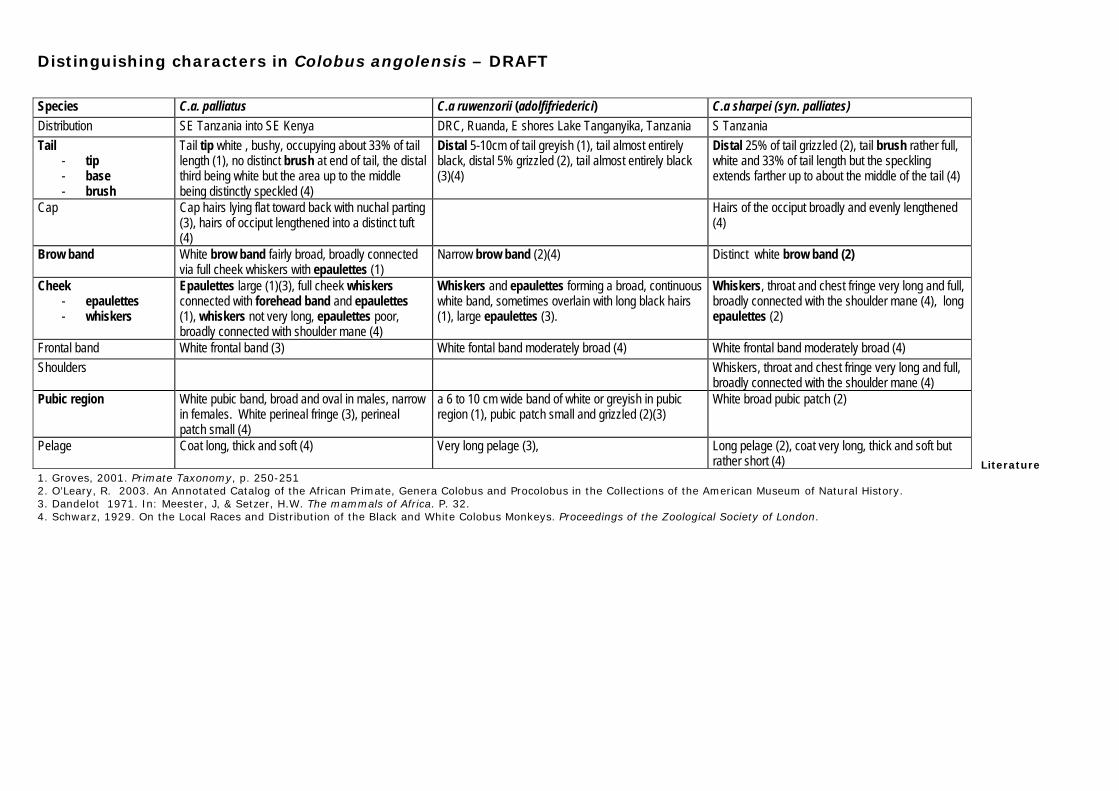
Distinguishing characters in Colobus angolensis – DRAFT
Literature
1. Groves, 2001. Primate Taxonomy, p. 250-251 2. O’Leary, R. 2003. An Annotated Catalog of the African Primate, Genera Colobus and Procolobus in the Collections of the American Museum of Natural History. 3. Dandelot 1971. In: Meester, J, & Setzer, H.W. The mammals of Africa. P. 32. 4. Schwarz, 1929. On the Local Races and Distribution of the Black and White Colobus Monkeys. Proceedings of the Zoological Society of London.
Species C.a. palliatus C.a ruwenzorii (adolfifriederici) C.a sharpei (syn. palliates) Distribution SE Tanzania into SE Kenya DRC, Ruanda, E shores Lake Tanganyika, Tanzania S Tanzania Tail
- tip - base - brush
Tail tip white , bushy, occupying about 33% of tail length (1), no distinct brush at end of tail, the distal third being white but the area up to the middle being distinctly speckled (4)
Distal 5-10cm of tail greyish (1), tail almost entirely black, distal 5% grizzled (2), tail almost entirely black (3)(4)
Distal 25% of tail grizzled (2), tail brush rather full, white and 33% of tail length but the speckling extends farther up to about the middle of the tail (4)
Cap
Cap hairs lying flat toward back with nuchal parting (3), hairs of occiput lengthened into a distinct tuft (4)
Hairs of the occiput broadly and evenly lengthened (4)
Brow band White brow band fairly broad, broadly connected via full cheek whiskers with epaulettes (1)
Narrow brow band (2)(4) Distinct white brow band (2)
Cheek - epaulettes - whiskers
Epaulettes large (1)(3), full cheek whiskers connected with forehead band and epaulettes (1), whiskers not very long, epaulettes poor, broadly connected with shoulder mane (4)
Whiskers and epaulettes forming a broad, continuous white band, sometimes overlain with long black hairs (1), large epaulettes (3).
Whiskers, throat and chest fringe very long and full, broadly connected with the shoulder mane (4), long epaulettes (2)
Frontal band White frontal band (3) White fontal band moderately broad (4) White frontal band moderately broad (4) Shoulders Whiskers, throat and chest fringe very long and full,
broadly connected with the shoulder mane (4) Pubic region White pubic band, broad and oval in males, narrow
in females. White perineal fringe (3), perineal patch small (4)
a 6 to 10 cm wide band of white or greyish in pubic region (1), pubic patch small and grizzled (2)(3)
White broad pubic patch (2)
Pelage Coat long, thick and soft (4) Very long pelage (3), Long pelage (2), coat very long, thick and soft but rather short (4)

Distinguishing characters in Colobus guereza - DRAFT
Literature 1. Groves, 2001. Primate Taxonomy, p. 250-251 2. Oates, J.F. & Trocco, T.F. 1983. Taxonomy and Phylogeny of Black-and-White Colobus Monkeys. Folia Primatologica: 40:83-113. 3. Schwarz, 1929. On the Local Races and Distribution of the Black and White Colobus Monkeys. Proceedings of the Zoological Society of London. 4. Dandelot 1971. An identification manual Primates In: Meester, J, & Setzer, H.W. The mammals of Africa. P. 31. 5. O’Leary, R. 2003. An Annotated Catalog of the African Primate, Genera Colobus and Procolobus in the Collections of the American Museum of Natural History. 6. MOA ms Colobus guereza, draft 04 7. Thomson 1885, Report on the mammals obtained and observed by Mr H.H. Johnston and Mt Kilimanjaro. Proc. Zool. Soc. Lond. p. 219 8. Heller, 1913. Smiths. Misc. Coll. lxi, No. 17, p11.
Species C.g. caudatus C.g. matschiei C.g. kikuyensis C.g. percivali C. abyssinicus roosevelti Distribution Mt Kilimanjaro, Mt Meru, Momela
Lakes and Kahe. Kenya, west of Rift and in the Rift, west to Mt Elgon, south to Ngorongoro Crater and the Grumeti River Tanzania
From Ngong Escarpment to Mt Kenya and Aberdares, Kenya
Mount Gargues, Mathews Range, Kenya
Mau Forest (8)
Tail - Length - Tuft - Base
White tail tuft making up over 80% of tail length, only extreme proximal part of tail black (1)(6). Base of tail black, slightly shorter than in kikuyensis (3). Tail tuft longer than kikuyensis (3), tail completely white, often yellow in colour (4). 3 to 4 inches of base black, the rest white tuft (20-21 inch)(7)
White tail tuft short (45% of tail length), rest black (1)(6), tail longer than head and body (1), tuft copious, 50% of the length of tail (3)(4). Tail base black, tail tuft full covering 33% of total length (5).
White tail tuft very bushy, extending over 72% of tail length (1), tail proximally gray (1), tail short, about equal to head plus body (1), Base of tail grey, slightly longer than in caudatus (4). Tail completely white with tuft almost from base (4). Base of tail black, white tuft voluminous forming about 66% of the length of tail (5)(6).
White tail tuft extends over 63% of tail (1), tail as long as head plus body (1), tuft full (3)(6) and bushy, occupies 66% of the whole tail (3). Tail tuft smaller than caudatus, longer tail, larger body than caudatus (8)
More extensively black tail than C. g. matschiei (8). White terminal end of tail 25%, other 75% black short hair (8)
Veil - length
Flank veil longer than kikuyensis (1)(3). Veil covers black part of tail completely
Flank veil much yellowed, not extending up onto back but covering base of tail (1), posterior portion of veil about as long as the tail tuft, extending to the basal portion of the tail (3)(4)(5), occasionally as far down as the beginning of the tuft (3).
Flank veil very long extending well up on back and beyond base of tail tuft (1)(3), veil covering 30% of the tail (4). Veil covering up to a quarter of the tail (5)
Flank veil very long, creamy yellow, only just reaching tail base (1)(3).
Shoulders White hairs sprinkled over shoulders, nearly linking veil to white of throat (1)
No white sprinkling on shoulders below veil (1)
Throat White (1) Under parts Fur less woolly than kikuyensis
(1)(3)
Other Longer and distinct thigh-band (compared to kikuyensis) (3)
Hair short (1) Hair long; thigh stripe short (1), Thigh-band short and pale (3), fur soft and woolly (3)
Hair very long (1). Temporal ridges into sagittal crest (8)
Resembling C. g. matschiei but smaller. Sagittal cres on the skull when aged (8)
Appendix 2. Encounters with Primate groups within and outside the coastal forest of Kenya (2005-2008). Encounters with Cercopithecus mitis albogularis and Cercopithecus mitis albotorquatus within and outside the coastal forests of Kenya (2005-2008). Number Species Locality name Latitude Longitude Altitude m Vegetation Date Time 185 C. m. albogularis Gazi -4.40361 39.50647 9 Along road 07-02-06 16.37 187 C. m. albogularis Mrima Hill -4.49444 39.26333 81 Secondary forest edge, coastal and shamba mango cashew
baobab cocos palm 07-02-06 17.16
190 C. m. albogularis Mrima Hill -4.48444 39.27057 110 Secondary forest edge, coastal and shamba mango cashew baobab cocos palm
07-02-06 17.27
191 C. m. albogularis Mrima Hill -4.48361 39.27133 82 Secondary forest edge, coastal and shamba mango cashew baobab cocos palm
07-02-06 17.29
194 C. m. albogularis Mrima Hill -4.49069 39.25439 101 Secondary forest edge, coastal and shamba mango cashew baobab cocos palm
07-02-06 17.55
195 C. m. albogularis Mrima Hill -4.48881 39.25431 124 Secondary forest edge, coastal and shamba mango cashew baobab cocos palm
07-02-06 18.07
197 C. m. albogularis Mrima Hill -4.48656 39.25418 130 Secondary forest edge, coastal and shamba mango cashew baobab cocos palm
07-02-06 18.19
198 C. m. albogularis Mrima Hill -4.48428 39.25394 146 Secondary forest edge, coastal and shamba mango cashew baobab cocos palm
07-02-06 18.20
248 C. m. albogularis Mrima Hill -4.48694 39.27056 79 Secondary forest edge, coastal and shamba mango cashew baobab cocos palm
20-02-06 11.13
- C. m. albogularis Kaya Rabai, Mazeras -3.94494 39.58164 212 Coastal forest edge including Coconut palms cashew nut (Anacardium occidentale), mango (Mangifera indica)
330 C. m. albogularis Tsavo West NP, Finch Hatton camp
-2.93328 37.90464 798 Ground water forest 20-4-08 12.10
333 C. m. albogularis Tsavo West NP, Finch Hatton camp
-2.93171 37.90537 806 Ground water forest 20-4-08 13.40
338 C. m. albogularis Tsavo West NP, Mzima Springs
-2.98343 38.02153 804 Ground water forest including Raphia farinifera and Phoenix reclinata palms.
20-4-08 17.25
341 C. m. albogularis Lunga lunga -4.55784 39.12630 48 In tall fig tree in riverine forest, close to village along the highway
22-4-08 10.30
342 C. m. albogularis Vanga -4.66056 39.21556 0 Mangrove forest 22-4-08 12.15 343 C. m. albogularis Vanga W -4.54278 39.06028 123 Dense dry bushland 22-4-08 15.37 345 C. m. albogularis Buda Forest -4.46444 39.41497 77 Forest edge with highway 23-4-08 15.15
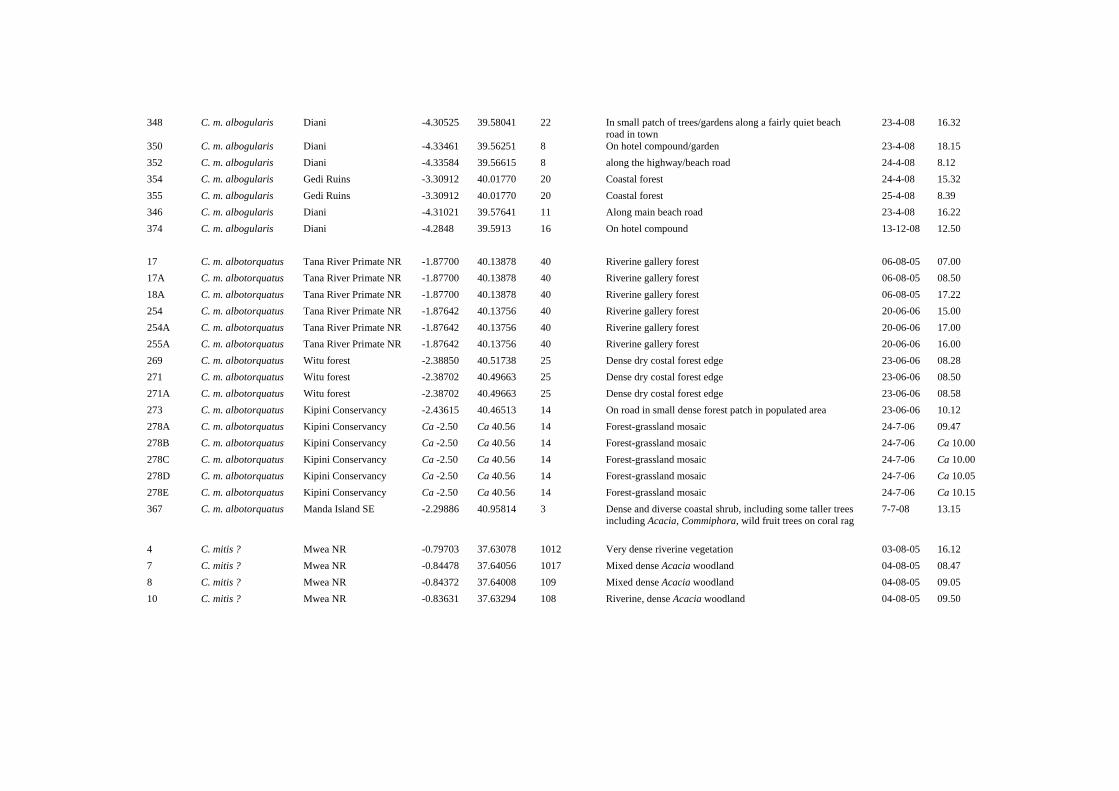
348 C. m. albogularis Diani -4.30525 39.58041 22 In small patch of trees/gardens along a fairly quiet beach road in town
23-4-08 16.32
350 C. m. albogularis Diani -4.33461 39.56251 8 On hotel compound/garden 23-4-08 18.15 352 C. m. albogularis Diani -4.33584 39.56615 8 along the highway/beach road 24-4-08 8.12 354 C. m. albogularis Gedi Ruins -3.30912 40.01770 20 Coastal forest 24-4-08 15.32 355 C. m. albogularis Gedi Ruins -3.30912 40.01770 20 Coastal forest 25-4-08 8.39 346 C. m. albogularis Diani -4.31021 39.57641 11 Along main beach road 23-4-08 16.22 374 C. m. albogularis Diani -4.2848 39.5913 16 On hotel compound 13-12-08 12.50 17 C. m. albotorquatus Tana River Primate NR -1.87700 40.13878 40 Riverine gallery forest 06-08-05 07.00 17A C. m. albotorquatus Tana River Primate NR -1.87700 40.13878 40 Riverine gallery forest 06-08-05 08.50 18A C. m. albotorquatus Tana River Primate NR -1.87700 40.13878 40 Riverine gallery forest 06-08-05 17.22 254 C. m. albotorquatus Tana River Primate NR -1.87642 40.13756 40 Riverine gallery forest 20-06-06 15.00 254A C. m. albotorquatus Tana River Primate NR -1.87642 40.13756 40 Riverine gallery forest 20-06-06 17.00 255A C. m. albotorquatus Tana River Primate NR -1.87642 40.13756 40 Riverine gallery forest 20-06-06 16.00 269 C. m. albotorquatus Witu forest -2.38850 40.51738 25 Dense dry costal forest edge 23-06-06 08.28 271 C. m. albotorquatus Witu forest -2.38702 40.49663 25 Dense dry costal forest edge 23-06-06 08.50 271A C. m. albotorquatus Witu forest -2.38702 40.49663 25 Dense dry costal forest edge 23-06-06 08.58 273 C. m. albotorquatus Kipini Conservancy -2.43615 40.46513 14 On road in small dense forest patch in populated area 23-06-06 10.12 278A C. m. albotorquatus Kipini Conservancy Ca -2.50 Ca 40.56 14 Forest-grassland mosaic 24-7-06 09.47 278B C. m. albotorquatus Kipini Conservancy Ca -2.50 Ca 40.56 14 Forest-grassland mosaic 24-7-06 Ca 10.00 278C C. m. albotorquatus Kipini Conservancy Ca -2.50 Ca 40.56 14 Forest-grassland mosaic 24-7-06 Ca 10.00 278D C. m. albotorquatus Kipini Conservancy Ca -2.50 Ca 40.56 14 Forest-grassland mosaic 24-7-06 Ca 10.05 278E C. m. albotorquatus Kipini Conservancy Ca -2.50 Ca 40.56 14 Forest-grassland mosaic 24-7-06 Ca 10.15 367 C. m. albotorquatus Manda Island SE -2.29886 40.95814 3 Dense and diverse coastal shrub, including some taller trees
including Acacia, Commiphora, wild fruit trees on coral rag 7-7-08 13.15
4 C. mitis ? Mwea NR -0.79703 37.63078 1012 Very dense riverine vegetation 03-08-05 16.12 7 C. mitis ? Mwea NR -0.84478 37.64056 1017 Mixed dense Acacia woodland 04-08-05 08.47 8 C. mitis ? Mwea NR -0.84372 37.64008 109 Mixed dense Acacia woodland 04-08-05 09.05 10 C. mitis ? Mwea NR -0.83631 37.63294 108 Riverine, dense Acacia woodland 04-08-05 09.50

Encounters with Chlorocebus pygerythrus hilgerti within and outside the coastal forests of Kenya (2005-2008). Number Species Locality name Latitude Longitude Altitude m Vegetation Date Time 1 C. p. hilgerti Mwea NR -0.82114 37.64000 1032 03-08-05 13.10 3 C. p. hilgerti Mwea NR -0.81444 37.62541 Dense Acacia- Commiphora bushland 03-08-05 15.57 5 C. p. hilgerti Mwea NR -0.79703 37.63078 Dense Acacia- Commiphora bushland 03-08-05 16.12 12 C. p. hilgerti Mwingi E -0.91331 38.12856 04-08-05 13.45 13 C. p. hilgerti Garissa SW -0.66986 39.12436 Acacia- Commiphora woodland 05-08-05 08.43 15 C. p. hilgerti Wenje (Mwiti) -1.78528 40.09735 45 Riverine vegetation 05-08-05 15.40 20 C. p. hilgerti Kora NP -0.24972 38.58889 Acacia- Commiphora bushland/woodland 08-08-05 16.08 21 C. p. hilgerti Kora NP -0.20778 38.63444 08-08-05 16.41 22 C. p. hilgerti Kora NP -0.07250 38.61083 08-08-05 17.22 23 C. p. hilgerti Kora NP -0.03750 38.61667 Bushland close to river 08-08-05 17.39 24 C. p. hilgerti Kora NP -0.03306 38.59194 Bushland close to river 08-08-05 17.49 25 C. p. hilgerti Kora NP -0.02553 38.57444 Bushland close to river 08-08-05 17.59 27A C. p. hilgerti Kora NP -0.07161 38.42039 Riverine vegetation 09-08-05 09.12 27 C. p. hilgerti Meru NP -0.07161 38.42039 875 Acacia- Commiphora bushland/woodland 09-08-05 08.58 28 C. p. hilgerti Meru NP 0.93360 37.21028 538 Riverine vegetation 09-08-05 11.00
85 C. p. hilgerti Meru NP 0.16597 38.20756 600 Campsite, riverine forest and dry bushland 13-10-05
175 C. p. hilgerti Makindu, Hunters lodge -2.20999 37.71606
Woodland, including Acacia xanthophloea, ficus. Short grass undergrowth. 06-02-06 15.13
177 C. p. hilgerti Makindu, Kiboko camp -2.19331 37.67158 927
Woodland, including A. xanthophloea, ficus. Short grass undergrowth. 06-02-06 17.06
178A C. p. hilgerti Makindu, Kiboko camp -2.19331 37.67158 927
Woodland, including A. xanthophloea, ficus. Short grass undergrowth. 07-02-06 06.40
181 C. p. hilgerti Tsavo -2.99344 38.45839 462 Along highway 07-02-06 10.31 249 C. p. hilgerti Voi S -3.57500 38.77694 249 Along highway, Commiphora-Acacia bush-woodland 21-02-06 08.22 251A C. p. hilgerti Wenje (Mwiti) -1.78122 40.10539 25 Acacia bushland, sandy soil, close to village 20-06-06 10.08
252A C. p. hilgerti Tana River Primate NR N -1.84272 40.06139 42 Acacia bushland, sandy soil, close to village 20-06-06 10.47
256 C. p. hilgerti Tana River Primate NR S -1.93767 40.08487 46 Acacia bushland sandy soil 21-06-06 10.48
259 C. p. hilgerti Kipini Conservancy -2.49178 40.53170 14 Grassland with doumpalm, greenbush whith some taller trees 22-06-06 10.07
265 C. p. hilgerti Mpeketoni -2.39103 40.69322 14 Village 22-06-06 17.02 285 C. p. hilgerti Garissa SW -0.67555 39.10205 339 26-06-06 8.03 287 C. p. hilgerti Garissa SW -0.78003 38.73337 26-06-06 10.00
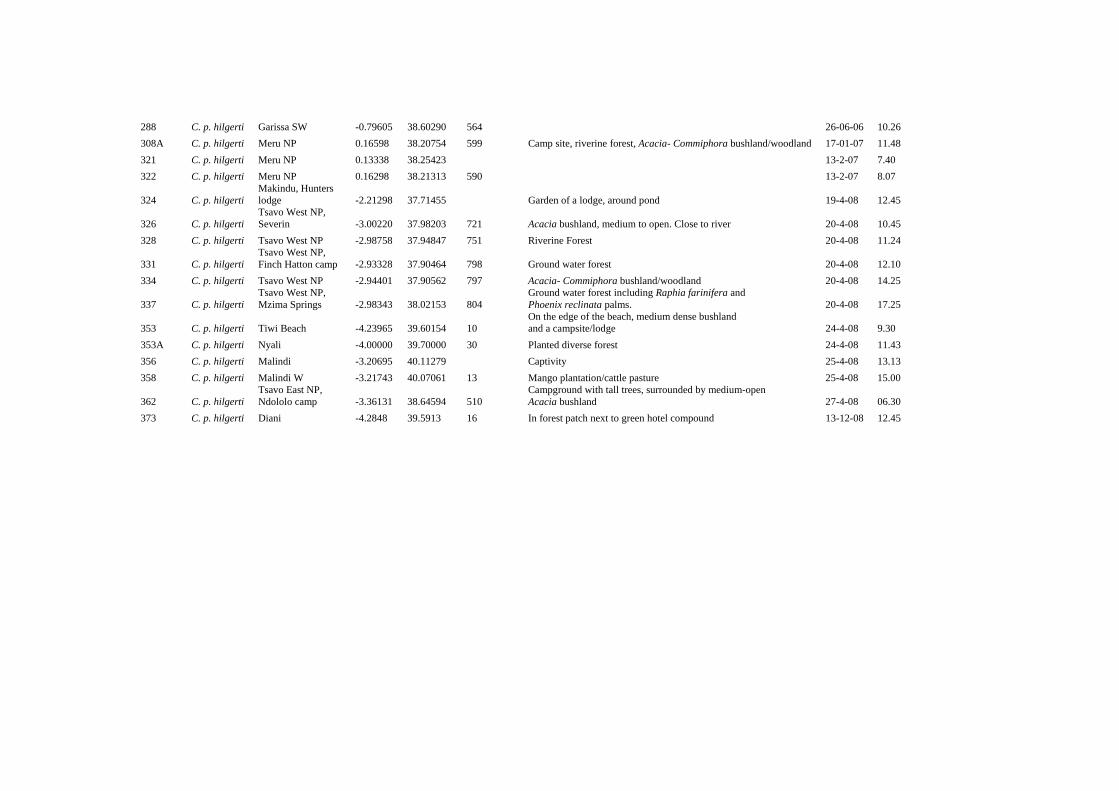
288 C. p. hilgerti Garissa SW -0.79605 38.60290 564 26-06-06 10.26 308A C. p. hilgerti Meru NP 0.16598 38.20754 599 Camp site, riverine forest, Acacia- Commiphora bushland/woodland 17-01-07 11.48 321 C. p. hilgerti Meru NP 0.13338 38.25423 13-2-07 7.40 322 C. p. hilgerti Meru NP 0.16298 38.21313 590 13-2-07 8.07
324 C. p. hilgerti Makindu, Hunters lodge -2.21298 37.71455 Garden of a lodge, around pond 19-4-08 12.45
326 C. p. hilgerti Tsavo West NP, Severin -3.00220 37.98203 721 Acacia bushland, medium to open. Close to river 20-4-08 10.45
328 C. p. hilgerti Tsavo West NP -2.98758 37.94847 751 Riverine Forest 20-4-08 11.24
331 C. p. hilgerti Tsavo West NP, Finch Hatton camp -2.93328 37.90464 798 Ground water forest 20-4-08 12.10
334 C. p. hilgerti Tsavo West NP -2.94401 37.90562 797 Acacia- Commiphora bushland/woodland 20-4-08 14.25
337 C. p. hilgerti Tsavo West NP, Mzima Springs -2.98343 38.02153 804
Ground water forest including Raphia farinifera and Phoenix reclinata palms. 20-4-08 17.25
353 C. p. hilgerti Tiwi Beach -4.23965 39.60154 10 On the edge of the beach, medium dense bushland and a campsite/lodge 24-4-08 9.30
353A C. p. hilgerti Nyali -4.00000 39.70000 30 Planted diverse forest 24-4-08 11.43 356 C. p. hilgerti Malindi -3.20695 40.11279 Captivity 25-4-08 13.13 358 C. p. hilgerti Malindi W -3.21743 40.07061 13 Mango plantation/cattle pasture 25-4-08 15.00
362 C. p. hilgerti Tsavo East NP, Ndololo camp -3.36131 38.64594 510
Campground with tall trees, surrounded by medium-open Acacia bushland 27-4-08 06.30
373 C. p. hilgerti Diani -4.2848 39.5913 16 In forest patch next to green hotel compound 13-12-08 12.45

Encounters with Papio within and outside the coastal forests of Kenya (2005-2008) Number Species Locality name Latitude Longitude Altitude m Vegetation Date Time 26 P. anubis Meru NP -0.07333 38.41361 09-08-05 08.36 301 P. anubis Meru NP 0.26747 38.12800 757 Meru NP entrance 14-01-07 13.22 308 P. anubis Meru NP -0.13125 38.25040 540 Grass, edge with riverine forest 17-01-07 11.27 309 P. anubis Meru NP 0.13923 38.17225 Meru HQ 18-01-07 8.00 84 P. anubis Meru NP 0.16597 38.20756 599 On and around campsite, riverine forest and dry bushland
(including Acacia, commiphora) 13-10-05
19B P. c. ibeanus Garissa SW -0.73556 38.97806 Medium dense Acacia bushland, 08-08-05 08.21 14 P. c. ibeanus Garissa SW -0.64844 39.27019 Medium dense Acacia - Commiphora woodland 05-08-05 09.19 16 P. c. ibeanus Wenje (Mwiti) N -1.73361 40.08889 05-08-05 17.38 18 P. c. ibeanus Tana River Primate
NR -1.87700 40.13878 35 Dry bushland, near riverine forest 06-08-05 11.29
19 P. c. ibeanus Garissa S -0.78694 39.66833 Degraded open Acacia bushland 07-08-05 15.25 186 P. c. ibeanus Gazi -4.40361 39.50647 9 Along road 07-02-06 16.37 257 P. c. ibeanus Garsen -2.27600 40.22712 43 Dense Acacia coastal shrub, Ficus sycamorous
woodland 21-06-06 12.54
258 P. c. ibeanus Kipini Conservancy -2.54205 40.60223 13 Forest edge with agricultural land and small village 21-06-06 16.42 260 P. c. ibeanus Kipini Conservancy -2.48272 40.55528 Grassland with doumpalm, greenbush with
some taller trees 22-06-06 10.45
261 P. c. ibeanus Kipini Conservancy -2.47582 40.55717 13 Grassland with doumpalm, greenbush with some taller trees, on edge with agricultural land
22-06-06 10.55
262 P. c. ibeanus Kipini Conservancy -2.47265 40.55675 Grassland, open bush/woodland with doumpalm bordering agricultural land
22-06-06 10.58
263 P. c. ibeanus Kipini Conservancy -2.42517 40.61950 Grassland, open bush/woodland with doumpalm bordering agricultural land
22-06-06 14.35
264 P. c. ibeanus Kipini Conservancy -2.42258 40.62205 16 Grassland, open bush/woodland with doumpalm bordering agricultural land
22-06-06 14.39
266 P. c. ibeanus Witu forest E -2.34858 40.59468 16 Coastal forest clearing, moving into agricultural land and tall grass
23-06-06 07.48
267 P. c. ibeanus Witu forest E -2.37585 40.53937 Open woodland with doumpalm 23-06-06 08.08 268 P. c. ibeanus Witu forest E -2.37767 40.53607 23-06-06 08.13 270 P. c. ibeanus Witu forest -2.38817 40.50380 25 Dense dry costal forest edge 23-06-06 08.45 272 P. c. ibeanus Witu forest -2.38475 40.49023 25 Dense dry costal forest edge 23-06-06 09.13 274 P. c. ibeanus Kipini Conservancy -2.48328 40.55850 Forest-grassland mosaic 23-06-06 11.11 275 P. c. ibeanus Kipini Conservancy -2.48420 40.56627 Forest-grassland mosaic 23-06-06 11.20

276 P. c. ibeanus Kipini Conservancy -2.48767 40.58597 Forest-grassland mosaic 23-06-06 11.29 277 P. c. ibeanus Kipini Conservancy -2.49470 40.60373 Forest-grassland mosaic 23-06-06 11.41 278 P. c. ibeanus Kipini Conservancy -2.48517 40.57365 Forest-grassland mosaic 24-06-06 11.00 279 P. c. ibeanus Witu town -2.38812 40.42112 25 Forest-grassland mosaic, on edge with town 25-06-06 11.57 280 P. c. ibeanus Garsen E -2.29057 40.25813 25-06-06 12.26 281 P. c. ibeanus Tana River Primate
NR S -1.98548 40.09865 Town 25-06-06 13.40
282 P. c. ibeanus Tana River Primate NR
-1.87642 40.13892 Riverine gallery forest edge with dry bushland 25-06-06 14.35
283 P. c. ibeanus Garissa -0.46300 39.62180 Close to human settlement 26-06-06 6.48 284 P. c. ibeanus Garissa SW -0.58888 39.34577 Open woodland 26-06-06 7.27 286 P. c. ibeanus Garissa SW -0.76335 38.80568 471 Medium dense bushland, red soil with little
to no undergrowth 26-06-06 9.18
340 P. c. ibeanus Mwardimu -4.17304 39.01811 251 Close to agricultural, outside medium dense Acacia bushland
21-4-08 17.06
349 P. c. ibeanus Diani -4.33785 39.56533 Along beachroad 23-4-08 17.01 351 P. c. ibeanus Diani -4.33461 39.56251 8 Hotel compound/garden 23-4-08 18.15 357 P. c. ibeanus Malindi -3.20695 40.11279 Captivity 25-4-08 13.13 365 P. c. ibeanus Manda toto Island -2.22719 40.98672 9 Beach, mangrove forest 4-7-08 10.40 366 P. c. ibeanus Manda toto Island -2.22719 40.98672 9 Beach, mangrove forest 5-7-08 8.35 272A P. c. ibeanus Witu town S -2.49906 40.52739 25 Mainroad 25-06-06 11.57 277A P. c. ibeanus Kipini Conservancy -2.45803 40.63002 9 23-06-06 11.41 278F P. c. ibeanus Kipini Conservancy -2.49180 40.56206 15 Open woodland with tall grass, dense forest 23-06-06 11.41 2 Papio Mwea NR -0.80556 37.62556 Dense Acacia – Commiphora bushland 03-08-05 15.30 6 Papio Mwea NR -0.79508 37.61667 Open Acacia tortolis woodland, lots of undergrowth 03-08-05 16.48 9 Papio Mwea NR -0.83939 37.63725 Dense Acacia woodland 04-08-05 09.19 11 Papio Mwea NR -0.84075 37.62903 Riverine, medium Acacia woodland 04-08-05 10.03 174 Papio Makindu,
Kiboko camp -2.19331 37.67158 927 Highway 06-02-06 13.10
176 Papio Makindu, Hunters lodge
-2.20999 37.71606 Lodge compound with large yellow fever 06-02-06 15.54
178 Papio Makindu, Kiboko camp
-2.19331 37.67158 927 A. xanthophloea, ficus woodland, tall trees, short grass undergrowth.
06-02-06 17.45
179 Papio Makindu, Kiboko camp
-2.19331 37.67158 927 A. xanthophloea, ficus woodland, tall trees, short grass undergrowth.
07-02-06 08.25
180 Papio Mtito Andei -2.74000 38.23000 710 Highway 07-02-06 09.50 182 Papio Tsavo West NP -3.00180 38.46244 471 Highway, close to garbage dump 07-02-06 10.55

183 Papio Voi S -3.46458 38.65333 537 Highway 07-02-06 11.44 184 Papio Voi S -3.61031 38.83939 444 Highway, Acacia - Commiphora bushland 07-02-06 12.23 250 Papio Voi N -3.18639 38.49639 590 Highway 21-02-06 09.00 251 Papio Voi N -3.00444 38.46889 490 Highway 21-02-06 09.18 252 Papio Kibwezi -2.41139 37.94861 937 Highway 21-02-06 10.24 325 Papio Tsavo West NP,
Chyulu Gate -2.90508 38.03984 787 Acacia - Commiphora bushland with dense tall grass 19-4-08 16.16
327 Papio Tsavo West NP, Severin
-3.00218 37.97029 725 Open and low bush with short grass and bare ground. Edge with riverine vegetation.
20-4-08 11.13
329 Papio Tsavo West NP -2.96189 37.91249 787 20-4-08 11.52 332 Papio Tsavo West NP,
Finch Hatton camp -2.92463 37.91076 808 Garden of lodge. Ground water forest and water
around 20-4-08 12.39
335 Papio Tsavo West NP -2.98965 37.95239 751 Open Acacia - Commiphora bushland 20-4-08 15.10 336 Papio Tsavo West NP -3.00074 37.98199 721 20-4-08 15.45 339 Papio Tsavo West NP,
Mzima Springs -2.98343 38.02153 804 Ground water forest including Raphia farinifera
and Phoenix reclinata palms. 20-4-08 17.25
359 Papio Tsavo East NP, Sala Gate E
-3.12033 39.36525 145 Acacia bushland, medium 26-4-08 11.08
360 Papio Tsavo East NP -3.05814 38.86868 268 Open grass and bushland 26-4-08 15.45 361 Papio Tsavo East NP,
Ndololo camp -3.36131 38.64594 510 Tall trees, campground, medium/open Acacia
bushland 27-4-08 06.30
363 Papio Mwea NR -0.78226 37.63596 1018 Riverine Forest, edge with dense Acacia bushland and agricultural land
28-4-08 10.45
364 Papio Mwea NR -0.81176 37.60661 1075 Dense Acacia woodland with tall grass 28-4-08 13.50

Encounters with Procolobus rufomitratus rufomitratus within the coastal forests of Kenya (2005-2008) Number Species Locality name Latitude Longitude Altitude m Vegetation Date Time
17B P. r. rufomitratus Tana River Primate NR -1.87641 40.13891 46 Riverine gallery forest
255 P. r. rufomitratus Tana River Primate NR -1.87641 40.13891 46 Riverine gallery forest 20-06-06 15.30
Encounters with Cercocebus galeritus within the coastal forests of Kenya (2005-2008) Number Species Locality name Latitude Longitude Altitude m Vegetation Date Time 16A C. galeritus Tana River Primate NR -1.87642 40.13756 46 Riverine gallery forest 6-8-05 05.00 19A C. galeritus Tana River Primate NR -1.87642 40.13756 46 Riverine gallery forest 7-8-05 07.30 253 C. galleritus Tana River Primate NR -1.87642 40.13756 46 Riverine gallery forest 20-6-06 14.10
Encounters with Colobus angolensis palliates within and outside the coastal forests of Kenya (2005-2008) Number Species Locality name Latitude Longitude Altitude m Vegetation Date Time 188 C. a. palliatus Mrima Hill -4.49231 39.26725 80 Secondary forest edge, coastal and shamba mango cashew baobab cocos palm 07-02-06 17.23 189 C. a. palliatus Mrima Hill -4.48667 39.27094 86 Secondary forest edge, coastal and shamba mango cashew baobab cocos palm 07-02-06 17.26 192 C. a. palliatus Mrima Hill -4.48361 39.27133 82 Secondary forest edge, coastal and shamba mango cashew baobab cocos palm 07-02-06 17.29 193 C. a. palliatus Mrima Hill -4.49378 39.25539 81 Secondary forest edge, coastal and shamba mango cashew baobab cocos palm 07-02-06 17.50 196 C. a. palliatus Mrima Hill -4.48881 39.25431 124 Secondary forest edge, coastal and shamba mango cashew baobab cocos palm 07-02-06 18.07 199 C. a. palliatus Mrima Hill -4.48428 39.25394 146 Secondary forest edge, coastal and shamba mango cashew baobab cocos palm 07-02-06 18.20 246 C. a. palliatus Mrima Hill -4.49222 39.26722 72 Secondary forest edge, coastal and shamba mango cashew baobab cocos palm 20-02-06 11.10 247 C. a. palliatus Mrima Hill -4.48944 39.26917 80 Secondary forest edge, coastal and shamba mango cashew baobab cocos palm 20-02-06 11.12 344 C. a. palliatus Mrima Hill -4.49423 39.25727 83 Secondary forest edge, coastal and shamba mango cashew baobab cocos palm 23-4-08 11.24 347 C. a. palliatus Diani -4.30525 39.58041 22 In small patch of trees/gardens along a fairly quiet beach road in town 23-4-08 16.32 375 C. a. palliatus Diani -4.2848 39.5913 16 In forest patch next to green hotel compound 13-12-08 12.45

Appendix 3. Diurnal road surveys conducted at the north coast of Kenya. Only road surveys in which primates were encountered are included. A. Tana River Primate National Reserve to Garsen to Witu, Kenya. Survey date: 21 June 2006 Survey hours: 3.7 h Trip distance: 129 km Road: Partly tarmac road (B8, C112), 30.0 km/h Primate encounter rates: Total of two diurnal primate groups encountered; 0.5 groups/h, 0.02 groups/km (n=2)
C. p. hilgerti: 0.3 groups/h, 0.01 groups/km (n=1). P. c. ibeanus: 0.3 groups/h, 0.01 groups/km (n=1).
B. Witu to Garsen to Tana River Primate National Reserve, Kenya. Survey date: 25 June 2006 Survey hours: 3,7 h Trip distance: 130 km Road: Partly tarmac road (B8, C112), 40.0 km/h Primate encounter rates: P. c. ibeanus: 1.1 groups/h, 0.03 groups/km (n=4). C. Nyali to Watamu (including Gedi Ruins) Survey date: 24 April 2008 Survey hours: 4.4 h Trip distance: 129.4 km Road: Main highway (B8), 42 km/h. Primate encounter rate:
C. m. albogularis: 0.3 groups/h (n=1), 0.01 groups/km (n=1) D. Watamu to north Malindi to Watamu Survey date: 25 April 2008 Survey hours: 6.6 h Trip distance: 163.6 km Road: Main highway and several side roads (B8 & C103), 24 km/h. Primate encounter rate: Two groups of diurnal primates encountered, 0.3 groups/h (n=2), 0.01 groups/km (n=2)
C. m. albogularis: 0.2 groups/h, 0.01groups/km (n=1) C. p. hilgerti: 0.2 groups/h, 0.01groups/km (n=1)
P. c. ibeanus, C. p. hilgerti and O. g. lasiotis found in captivity in Malindi. All three individuals said to come from the vicinity of Malindi. Photos and descriptions obtained.

The Otolemur garnettii found in captivity was an adult male. The individual looked phenotypically different from any O. g. lasiotis we had observed along the coast of Kenya (Chapter 4.2.1). E. Watamu to Tsavo East National Park (Ndololo Camp). Survey date: 26 April 2008 Survey hours: 8.1 Trip distance: 260 Road: secondary road (C103), 32 km/h. Primate encounter rate: Two groups of diurnal primates encountered, 0.3 groups/h (n=2), 0.01 groups/km (n=2)
Papio: 0.3 groups/h, 0.01 groups/km (n=2)

Appendix 4. Diurnal road surveys conducted at the south coast of Kenya A. Nairobi to Makindu to Mrima Hill Forest Survey date: 6-7 February 2006 Survey hours: 7.6 h Trip distance: 535 km Road: Highway (A109 & A14), ca 65 km/h Primate encounter rate: Total of seven diurnal primate groups encountered; 0.9 groups/h, 0.01 groups/km (n=7).
C. p. hilgerti: 0,1 groups/h, 0.002 groups/km (n=1). Papio: 0.7 groups/h, 0.01 groups/km (n=5). C. m. albogularis: 0.1 groups/h, 0.002 groups/km (n=1).
Remarks: Phenotypic characters of Papio change from ‘good’ Papio anubis in Nairobi to Papio cynocephalus ibeanus on the Kenya coast, with animals with intermediate characteristics in between Nairobi and the coast. The extent of the P. anubis x P cynocephalus hybrid zone needs to be determined (Chapter 4.2.8) B. Mrima Hill Forest to Mazeras (including Shimba Hills National Reserve) Survey date: 20 February 2006 Survey hours: 8.2 h Trip distance: 311.0 km Road: Secondary roads and highways (A14, C106, C107, A109), 44 km/h Primate species encountered: None Remark During this road survey we drove through and partly around Shimba Hills National Reserve. No primates were encountered during this survey. In 2001 the Shimba Hills National Reserve is reported to hold the largest population of C. a. palliates in Kenya (458 groups, 2436 individuals; Anderson et al., 2007). C. North Kilimangodo to Kaya Sega Survey date: 22 April 2008 Survey hours: 5.3 h Trip distance: 107.7 km Road: Secondary roads (including C106), 19 km/h. Primate encounter rate: C. m. albogularis: 0.6 groups/h, 0.03 groups/km (n=3). Remarks During this road survey we drove through Kaya Gonja, a 4.2 km² Forest Reserve close to Kaya Sega. In 2001, Kaya Gonja held seven groups (33 individuals) of C. a. palliates (Anderson et al., 2007). Main threats to this population are illegal logging and hunting. Other threats are charcoal making, firewood collection and encroachment (Anderson et al., 2007). D. Kaya Sega to Diani

Survey date: 23 April 2008 Survey hours: 7.5 h Trip distance: 221.1 km Road: Secondary roads and partly highway (including C106, C108, A14), 28 km/h. Primate encounter rate: In total eight diurnal primate groups encountered, 1.1 groups/h, 0.04 group/km (n=8)
C. m. albogularis: 0.5 groups/h, 0.02 groups/km (n=4) C. a. palliates: 0.3 groups/h, 0.01 groups/km (n=2) P. cynocephalus: 0.3 groups/h, 0.01 groups/km (n=2)
Remarks During this road survey we briefly visited Dzombo Hill Forest, a 5 km² Forest Reserve and National Monument which reaches an altitude of 470 m asl. The Hill is covered with coastal mixed forest but is surrounded by degraded bushland, agriculture and human settlements (Figure 32). Despite its protected status, the forest covering Dzombo Hill is under threat from timber and pole extraction, bark stripping and agricultural encroachment (Bennun & Njoroge, 1999; Anderson et al., 2007). Anderson et al. (2007) encountered four groups (23 individuals) of C. a. palliates in the forest. Hunting is a serious threat to this small population (Anderson et al., 2007). During this road survey we passed Buda Forest Reserve (6.8 km²) and encountered C. m. albogularis on the edge of the forest. In 2001, Anderson et al. (2007) counted eight groups (52 individuals) of C. a. palliates in Buda Forest. The main threat to Buda Forest is illegal logging. Other threats are charcoal making, firewood collection, and hunting (Anderson et al., 2007). Further research Nocturnal and diurnal surveys should be conducted in Dzombo Hill Forest and Buda Forest to assess primate presence/absence and densities.
E. Diani to Tiwi to Nyali Survey date: 24 April 2008 Survey hours: 2.2 h Trip distance: 64.9 km Road: Highway (A14) and some secondary side roads, 28 km/h. Primate encounter rate: In total, two groups of diurnal primates encountered, 0.8 groups/h, 0.03 groups/km (n=2)
C. m. albogularis: 0.4 groups/h, 0.02 groups/km (n=1) C. p. hilgerti: 0.4 groups/h, 0.02 groups/km (n=1)
Figure 32. Dzombo Hill Forest Reserve, surrounded by degraded bushland, agriculture and human settlements.

Appendix 5. Diurnal road surveys, outside the coastal forests of Kenya A. Mwea National Reserve to Tana River Primate National Reserve. Survey date: 4-5 August 2005 Survey hours: 15.3 h (13.8 diurnal, 1.5 nocturnal) Trip distance: 547 km (541 diurnal, 5.8 nocturnal) Road: highways and dirt roads (B7, A3, B8), ca 45 km/h Primate encounter rates: A total of five diurnal primate groups encountered; 0.36 groups/h, 0.01 groups/km (n=5). A total of two nocturnal primates were encountered; 1.33 individuals/h, 0.34 individuals/km (n=2) Papio: 0.14 groups/h, 0.004 groups/km (n=2).
C. p. hilgerti: 0.22 groups/h, 0.006 groups/km (n=3) G. s. braccatus: 0.67 individuals/h, 0.17 individuals/km (n=1). Galago sp: 0.67 individuals/h, 0.17 individuals/km (n=1).
Remarks: Papio changes from ‘good’ P. anubis to the east of Mwea National Reserve to ‘good’ P. c. ibeanus at Kipini on the Kenya coast with intermediate (hybrid) individuals in between. The size of the Papio hybrid zone needs to be determined. Chapter 4.2.8 presents more details on the Papio hybrid zone. Two solitary desert warthogs P. aethiopicus were encountered 15 km and 80 km west of Garissa town in medium-dense Acacia bushland. These are the first records west of the Tana River and extend the geographic range to ca. 265 km northwest of Mkokoni, the nearest locality (d’Huart & Grubb, 2001). These encounteres are published in two publications (Culverwell et al., 2008; Appendix 6, and De Jong et al., 2009; Appendix 7). B. Tana River Primate National Reserve to Garissa. Survey date: 8 August 2005 Survey hours: 5.13 h Trip distance: 202 km Road: highways and dirt roads (B8, A3, B7), ca 45 km/h Primate encounter rates:
Papio encountered; 0.19 groups/h, 0.01 groups/km (n=1) C. Mazeras to Nairobi Survey date: 21 February 2006 Survey hours: 7.0 h Trip distance: 440 km Road: Highway (A109) Primate encounter rates: Four diurnal primate groups encountered; 0.57 groups/h, 0,01 groups/km (n=4).
C. p. hilgerti: 0.14 groups/h, 0.002 groups/km (n=1). Papio: 0.43 groups/h, 0.01 groups/km (n=3).
Remarks:

Papio changes from ‘good’ Papio anubis in Nairobi to P. c. ibeanus on the Kenya coast with intermediate (hybrid) individuals in between. The size of the Papio hybrid zone needs to be determined. Chapter 4.2.8 presents more detail on the Papio hybrid zone. D. Nanyuki to Garissa to Tana River Primate National Reserve Survey date: 19-20 June 2006 Survey hours: 15.0 h (12.1 diurnal, 2.9 nocturnal) Trip distance: 614 km Road: Highways and dirt roads (A2, C74, B7, A3, B8); 57.7 km/h and 38.3 km/h Primate encounter rates:
C. p. hilgerti: 0.16 groups/h, 0.003 groups/km (n=2). E. Tana River Primate National Reserve to Garissa to Nanyuki Survey date: 25-26 June 2006 Survey hours: 11.1 h Trip distance: 650 km Road: Highways and dirt roads (B8, A3, B7, C74, A2); 45.0 km/h & 57.0 km/h Primate encounter rates: Total of six diurnal primate groups encountered; 0.54 groups/h, 0.01 groups/km (n=6)
C. p. hilgerti: 0.27 groups/h, 0.01 groups/km (n=3). Papio: 0.27 groups/h, 0.01 groups/km (n=3).
Remark: Papio changes from ‘good’ P. anubis in Nanyuki to P. c. ibeanus at Kipini on the Kenya coast. About 100 km west of Garissa we encountered Papio hybrids with a striking golden yellow ventrum, cheeks, sides, inner limbs and back of their hind legs. The hybrid zone between the two species is obviously very wide with much phenotypic variation within and among groups between Garissa to at least Mwingi. The size of the Papio hybrid zone needs to be determined. Chapter 4.2.8 presents more detail on the Papio hybrid zone. F. Nanyuki to Mtito Andei Survey date: 19 April 2008 Survey hours: 7.4 h Trip distance: 451.5 km Road: Highways (A2, A109); 59 km/h Primate encounter rates:
C. p. hilgerti: 0.14 groups/h, 0.002 groups/km (n=1)
G. Mtito Andei to north Kilimangodo Survey date: 21 April 2008 Survey hours: 6.5 h Trip distance: 285.8 km Road: Secondary roads and highway (A109,), 42 km/h Primate encounter rates:
P. cynocephalus: 0.15 group/h, 0.004 (n=1).

63
Abstract: This paper reviews the complicated nomenclatural history for the Kenya coast galago, Galagoides cf. cocos, and exam-ines whether ‘cocos’ is the valid species name for this recently resurrected taxon. This paper also reviews the phenotypic and vocal differences among G. cocos; the Zanzibar galago (Galagoides zanzibaricus zanzibaricus); the Udzungwa galago (Galagoides zanzibaricus udzungwensis); and the Mozambique galago (Galagoides granti), as well as their geographic ranges and conserva-tion status. The following are among the findings: (1) ‘Galagoides cocos’ is the name that should be applied to the Kenya coast galago; (2) in the field, the loud calls of these three species are diagnostic and remain the best means for identification; (3) there is a suite of phenotypic characters that, when taken together, can be used to distinguish among these three species when in the hand or viewed in the field in good light at close range; (4) G. z. zanzibaricus is phenotypically distinct from G. z. udzungwensis; (5) the three species are parapatric or, perhaps, narrowly sympatric; (6) the three species are endemic to the coastal forests of eastern Africa with G. cocos in the north (Kenya and northeastern Tanzania), G. zanzibaricus in Tanzania, and G. granti from southern Tanzania to southern Mozambique; and (7) none of the three species is threatened at this time, although G. z. zanzibaricus meets the IUCN Red List criteria for an Endangered subspecies.Key Words: Dwarf galagos, Galagoides, cocos, granti, zanzibaricus, udzungwensis, taxonomy, conservation
Taxonomy, Distribution, and Conservation Status of Three Species of Dwarf Galagos (Galagoides) in Eastern Africa
Thomas M. Butynski1, Yvonne A. de Jong2, Andrew W. Perkin3, Simon K. Bearder3, and Paul E. Honess4
1Eastern Africa Regional Program, Conservation International, Nairobi, Kenya2Kenya Institute of Primate Research, Ol Kalou, Kenya
3Nocturnal Primate Research Group, Anthropology Department, Oxford Brookes University, Oxford, UK4Department of Veterinary Services, University of Oxford, Oxford, UK
Introduction
Many of the species and subspecies of the family Galagi-dae (galagos or bushbabies) have been subjected to repeated taxonomic revisions and name changes over the past century (for example, Elliot 1913; Allen 1939; Hill 1953; Groves 1977, 1993, 2001, 2005; Grubb et al. 2003). The Kenya coast galago (or Diani small galago), Galagoides cocos, is no exception (Figs. 1 and 2). Galagoides cocos is a recently revived, highly cryptic species of the coastal forest of eastern Africa.
In this paper we (1) review the nomenclatural history for the Kenya coast galago, (2) provide new information that con-firms that ‘cocos’ is the valid name for this recently revived species, (3) summarize the phenotypic and the main qualitative vocal differences among G. cocos, the Zanzibar galago (Gala-goides zanzibaricus), and the Mozambique or Grant’s galago (Galagoides granti), (4) review their geographic ranges, and (5) examine their conservation status (Figs. 1–7).
Nomenclatural History for the Kenya Coast Galago
On 16 December 1911, Edmund Heller (1912) col-lected an adult male dwarf galago (one of 10 specimens) at Mazeras, Kenya, which he named Galago moholi cocos. This taxon was raised to species status (Galago cocos) by Elliot (1913), but later placed as a subspecies of the Somali galago (Galago gallarum cocos), as a subspecies of the Zanzibar galago (Galago zanzibaricus cocos), or simply not recognized as a valid taxon and placed as a junior synonym of the Zanzibar galago (Galago senegalensis zanzibaricus or Galago zanzibaricus zanzibaricus or Galago zanzibari-cus or Galagoides zanzibaricus) (Table 1). Most recently, this taxon has been provisionally referred to as ‘Galagoi-des cf. cocos’ (Bearder et al. 2003; Grubb et al. 2003). This binomial is ‘provisional’ because the validity of the use of the name ‘cocos’ requires confirmation.
Primate Conservation 2006 (21): 63 –79

64
Butynski et al.
Figure 1. Adult (sex not known) Kenya coast galago (Galagoides cocos) from Arabuko-Sokoke Forest Reserve, southeastern Kenya (near Gedi). Note the muzzle patches and buffy-brown dorsum. Photograph by Harald Schuetz.
Figure 2. Adult female Kenya coast galago (Galagoides cocos) from Diani, southeastern Kenya. Note the muzzle patches. Photograph by Andrew Perkin.
Figure 3. Adult male Zanzibar galago (Galagoides zanzibaricus udzungw-ensis) from Pande Game Reserve, Tanzania (near Dar es Salaam). Note the absence of muzzle patches. Photograph by Nike Doggart.
Figure 4. Adult (sex not known) Udzungwa (or Matundu) galago (Galagoides zanzibaricus udzungwensis) from Matundu Forest Reserve, Udzungwa Moun-tains, south-central Tanzania (near Ifakara). Note that the hairs of the tail are of even length, sparse, and wiry, that the bone of the tail is visible, and that the tip of the tail is dusky. Photograph from Honess (1996).

65
Galagoides in Eastern Africa
Many of the more recent taxonomic studies on Galagi-dae make no mention of cocos, but presumably they consider cocos to be a synonym of Galago zanzibaricus (for example, Groves 1977; Nash et al. 1989; Masters 1998; Zimmermann
Figure 6. Typical adults of three Galagoides spp. at the British Museum of Natural History, London. Bottom to top: Kenya coast galago (Galagoides cocos) from Gande, Kenya, Udzungwa galago (Galagoides zanzibaricus ud-zungwensis) from Kissarawe, Tanzania, and Mozambique galago (Galagoides granti) from Coguno, Mozambique. Note the great similarity in the color of the dorsum, and that G. granti is the largest of the three species and has a wider, fuller, tail. Photograph by Tom Butynski.
1990) or of Galagoides zanzibaricus (for example, Honess 1996; Anderson 1999, 2000; DelPero et al. 2000; Masters and Bragg 2000; Masters and Brothers 2002).
The Need for Confirmation of the Name ‘cocos’ for the Kenya Coast Galago
Until recently, G. zanzibaricus was considered a poly-typic species of the coastal forests from southern Somalia through Kenya and Tanzania (including Unguja Island, Zanzi-bar, Tanzania) to southern Mozambique, and inland to central
Table 1. Summary of the nomenclature changes for the Kenya coast galago (Galagoides cocos).
Authority Latin name
Heller (1912) Galago moholi cocosElliot (1913), Hollister (1924) Galago cocos
Allen and Loveridge (1927) Galago gallarum cocos
Schwarz (1931), Hill (1953), Allen (1939), Hill and Meester (1977)
cocos a synonym of Galago senegalensis zanzibaricus
Jenkins (1987)cocos a synonym of Galago zanzibaricus zanzibaricus
Kingdon (1971, 1997), Groves (2005)cocos a synonym of Galago zanzibaricus
Groves (1993)cocos a synonym of Galagoides zanzibaricus
Groves (2001) Galago zanzibaricus cocos
Bearder et al. (2003), Grubb et al. (2003)
Galagoides cocos
Figure 5. Adult (sex not known) Mozambique galago (Galagoides granti) from Rondo Forest Reserve, southeastern Tanzania (near Lindi). This individual is emerging from a tree hole and, thus, its tail is not visible. Note the relatively large, blackish ears and buffy-brown dorsum. Photograph by Simon Bearder.
Figure 7. Two adult Zanzibar galagos (Galagoides zanzibaricus zanzibaricus) (left) from Unguja Island, Zanzibar, Tanzania, and two adult Udzungwa galagos (Galagoides zanzibaricus udzungwensis) (right) from Kissarawe, Tanzania. All four specimens are at the British Museum of Natural History (BMNH), Lon-don. The two specimens of G. z. zanzibaricus represent the extremes in pelage coloration among the 10 adult specimens at the BMNH. Note that the dorsum, tail, and outer front limbs are medium to bright cinnamon in G. z. zanzibaricus and buffy-brown in G. z. udzungwensis. Photograph by Tom Butynski.

66
Butynski et al.
Tanzania, Malawi, and extreme eastern Zimbabwe (Hill 1953; Groves 1977; Hill and Meester 1977; Smithers and Wilson 1979; Jenkins 1987; Courtenay and Bearder 1989; Skinner and Smithers 1990; Groves 2001, 2005; Bearder et al. 2003; Grubb et al. 2003). The ecology, behavior, and vocal repertoire of the mainland subspecies, G. z. cocos, is well known, hav-ing been the focus of detailed field studies at Diani and Gedi Forests, Kenya (Harcourt 1986; Harcourt and Nash 1986a, 1986b). Far less well known is the nominotypical subspecies, G. z. zanzibaricus, an endemic of Unguja Island, Zanzibar. It was not until A. Perkin visited Unguja Island in 1998 and recorded the species-specific advertising call of topotypical G. zanzibaricus (Fig. 8) that it became clear that these two forms were different:
(1) The species-specific advertising call of G. z. zanzibari-cus is very different from that of G. z. cocos. Galagoi-des z. zanzibaricus has a ‘single unit rolling call’ (Fig. 8), and G. z. cocos has an ‘incremental call’ (Fig. 9) (Hon-ess 1996; Honess and Bearder 1996; Perkin et al. 2002; Grubb et al. 2003).
(2) The species-specific advertising call of G. z. zanzibaricus is identical, or nearly so, to the species-specific advertis-ing call of the recently named Udzungwa (or Matundu) galago, Galagoides udzungwensis (see A. Perkin unpubl. data, cited in Bearder 1999). This led to the realization that G. udzungwensis may not be a new species, but rather synonymous with, or a subspecies of, G. zanzibaricus (see Perkin et al. 2002; Bearder et al. 2003; Grubb et al. 2003). Here we treat the Udzungwa galago as a distinct mainland subspecies (G. z. udzungwensis), but emphasize that the taxonomic status of the Udzungwa galago is far from resolved (see below).
Based on a considerable body of knowledge concerning the species-specific advertising calls of the Galagidae, and their wide use and acceptance as a robust species recogni-
tion and taxonomic tool (Zimmermann et al. 1988; Courtenay and Bearder 1989; Harcourt and Bearder 1989; Nash et al. 1989; Zimmermann 1990; Masters 1991; Bearder et al. 1995, 2003; Honess 1996; Honess and Bearder 1996; Butynski et al. 1998; Ambrose 1999, 2003; Bearder 1999; Groves 2001; Perkin et al. 2002), it was judged that the level of difference between the advertising calls of G. z. cocos and G. z. zanzi-baricus/G. z. udzungwensis is far greater than can be accom-modated at the subspecies level. In fact, the advertising call of G. z. cocos is far more similar to the advertising ‘incremental’ call of G. granti (formerly G. zanzibaricus granti) (Fig. 10) than it is to the ‘single unit rolling’ call of G. z. zanzibaricus (see Bearder et al. 1995). As such, G. z. cocos was reassigned species status, G. cf. cocos (Bearder et al. 2003; Grubb et al. 2003). Interestingly, the geographic range of G. zanzibaricus is located between the geographic ranges of G. cocos and G. granti (see below).
Grubb and co-authors give a succinct overview of this complex situation.
“Galagos at Diani, which were thought to be Galagoides zanzibaricus (Harcourt and Nash, 1986a, b) are vocally distinct from true or topotypical G. zan-zibaricus of Zanzibar. They are provisionally identi-fied as Galagoides cf. cocos, and we assign them to the G. granti group. Galagos from the Udzungwa Mtns and other localities in Tanzania have been named Galagoides udzungwensis (Honess, 1996) but, on the basis of their vocalization, do not differ from those of topotypical G. zanzibaricus of Zanzibar (A. Perkin unpubl. data, cited in Bearder, 1999). The form udzungwensis may prove to be a valid taxon at the subspecific level but until the systematics is clari-fied, we relegate it to the synonymy of Galagoides zanzibaricus.” (Grubb et al. 2003, pp.1315–1316).
And below“The galago recorded from Diani is vocally dis-
tinct from Galagoides zanzibaricus and has been
Figure 8. Sonogram and oscillogram of the ’single unit rolling’ advertising call of the Zanzibar galago (Galagoides zanzibaricus zanzibaricus) from Unguja Island, Zanzibar, Tanzania, the type locality for this species. Call recorded by Andrew Perkin. This call is comprised of a series of ‘rolling’ trill units that, after a few units, increase in frequency and amplitude before reaching a mild crescendo and then trailing off with trill units of lower amplitude and frequency. The lowering of the frequency is achieved by eliminating the higher frequency elements. The lowest frequency elements remain constant. Each trill unit is made up of a very rapid series of trill subunits. The number of units per ‘single unit rolling’ call varies considerably (Honess 1996; A. Perkin, pers. obs.). For the above recording: Call length = 10.8 seconds. Frequency range = 0.62–11.12 kHz. Fundamental frequency = 0.75 kHz. Range of unit frequency modulation = 0.81–3.57 kHz. Number of phrases = 0. Number of units = 19. For the Udzungwa galago (Galagoides zanzibaricus udzungwensis) population in the Matundu Forest Reserve, south-central Tanzania, the type locality for this subspecies: Mean number of units per single unit rolling call = 14 (SE = 0.17, range = 1– 46, n = 2,122). Mean unit interval = 0.28 seconds (n = 181). Mean unit length = 0.22 seconds (n = 196). Range of fundamental frequency = 0.95 to 1.00 kHz (Honess 1996, Honess and Bearder 1996, A. Perkin unpubl. data). Oscillograms of the single unit rolling call of G. z. udzungwensis are presented in Bearder et al. (1995), Honess (1996), Honess and Bearder (1996), and King-don (1997).

67
Galagoides in Eastern Africa
Figure 9. Sonogram and oscillogram of the ’incremental’ advertising call of the Kenya coast galago (Galagoides cocos) from Diani Beach, southeastern Kenya. Call recorded by Simon Bearder. This call often, but not always, starts with a series of high-pitched, rapidly uttered, “chirrups” followed by units arranged in phrases that are high in frequency and amplitude, and that gradually become lower in amplitude. The number of units within each phrase increases incrementally until the end of the call. Often, as in this case, phrases with same number of units are repeated. The number of units per phrase rarely decreases. Units are often frequency modulated. For the above recording: Call length = 4.7 seconds. Frequency range = 0.65–11.15 kHz. Fundamental frequency = 0.98 kHz. Range of unit frequency modulation = 0.68–10.37 kHz. Number of phrases = 6 (with three introductory “chirrup” units and one incipient unit at the end). Mean number of units per phrase = 2.8. For the G. cocos population at Diani: Mean call length = 4.3 seconds (range = 1.7–8.6 seconds, n = 12). Frequency range = 0.8–9.3 kHz. Fundamental frequency = 0.8–1.2 kHz. Mean number of phrases = 6 (range = 3 –11, n = 13). Mean number of units per phrase = 2.5 (range 1–11, n = 60). Mean unit interval = 0.35 seconds (range 0.20 –0.59 seconds, n = 27). Mean unit length = 0.41 seconds (range 0.15–0.57 seconds, n = 33) (Courtenay and Bearder 1989). See also the acoustic measurements presented in Zimmermann (1990). Additional sonograms and oscillograms of the incremental advertising call, and other calls of G. cocos, are presented in Courtenay and Bearder (1989), Harcourt and Bearder (1989), Zimmermann (1990), Bearder et al. (1995), and Kingdon (1997).
Figure 10. Sonogram and oscillogram of the incremental advertising call of the Mozambique galago (Galagoides granti) from Rondo Forest Reserve, Rondo Plateau, southeastern Tanzania (near Lindi). Call recorded by Paul Honess. This sonogram is an example of a full incremental call in which the numbers of units in each phase gradually increase incrementally. Compared with the incremental call of G. cocos, the incremental call of G. granti maintains relatively consistent amplitude, has more units per phrase, and is more staccato. In the above example, the amplitude increases slightly during the middle phrases and decreases slightly during the last phrase. For the above recording: Call length = 4.9 seconds. Frequency range = 0.56 –11.18 kHz. Fundamental frequency = 0.75 kHz. Range of unit frequency modulation = 1.03 –5.08 kHz. Number of phrases = 6. Mean number of units per phrase = 3.6 (range 3 –5). In the G. granti population of the Rondo Forest Reserve: Mean number of phrases per incremental call = 5.8 (SE = 0.2, range = 1–17, n = 211). Mean unit interval = 0.55 seconds (n = 41). Mean unit length = 0.41 seconds (n = 53) (Honess 1996, Honess and Bearder 1996). Additional oscillograms of the incremental advertising call, and other calls, of G. granti are presented in Honess (1996), Honess and Bearder (1996), and Kingdon (1997).

68
Butynski et al.
recorded elsewhere in Kenya and in Tanzania, where A. Perkin (in litt.) reported it from the northern tip of the East Usambara Mtns, seemingly close to G. zanzibaricus (Table IV). The form Galago moholi cocos Heller, 1912, was described from Mazeras (Manzeras), relatively close to Diani (Table IV). Groves (2001) recognized cocos as a mainland form of Galagoides zanzibaricus. Vocalizations typical of G. zanzibaricus have not been recorded in Kenya. Therefore, it seems highly likely that the Diani galago is a separate species, Galagoides cf. cocos. Never-theless it is important to confirm this. Vocalizations recorded from as near to the type locality of Gala-goides cocos as possible should be compared with the voice of the Diani galago. The forest at the type locality (Kaya Mazeras) has been destroyed, but for-ests 5 km and 20 km distant (Kaya Mtswakana and
Kaya Fungo respectively) and some others might be visited (Butynski unpubl., including information from Q. Luke). Museum specimens of Galagoides cocos should be compared with the Diani galago further to ensure that we are dealing with a single taxon. We pro-visionally consider the Diani galago to be conspecific with Galagoides cocos, under the vernacular name Kenya coast galago.” (Grubb et al. 2003, p.1317).
In other words, in considering the Kenya coast galago once again a valid species, one important action remains: the confirmation of ‘cocos’ as the species name. This is necessary because the tape recordings used to describe the loud call for the Kenya coast galago were made at Diani, Kenya (04°18′S, 39°35′E) (Zimmermann 1990; Bearder et al. 1995), c.40 km south of the type locality for G. cocos (i.e., Mazeras). There are three rivers between these two sites that are potential barri-
Figure 11. Sonogram and oscillogram of the incremental advertising call of the Kenya coast galago (Galagoides cocos) from Kaya Chijembeni (Rabai), 4 km north-east of Mazeras, southeastern Kenya, the type locality for this species. Call recorded by Yvonne de Jong and Tom Butynski. Although numerous full incremental calls were heard and recorded at Kaya Chijembeni during two nights, there was much background noise and wind, and none of the recordings of the full incremental call were suitable for the production of a clear sonogram. The sonogram presented here is of an incomplete or incipient incremental call. Nonetheless, the first five phrases of incremental units are present and identifiable as the species-specific advertising call of G. cocos. In this case, there are no “chirrup” phrases and the number of units does not increase incrementally but the call still follows the typical high-pitched, rapid or staccato pattern of the G. cocos incremental call. For the above recording: Call length = 2.8 seconds. Frequency range = 0.98–12.4 kHz. Fundamental frequency = 0.77 kHz. Number of phrases = 5. Number of units per phrase = 2. Frequency modulation is not detectable, probably due to the low amplitude of the call.

69
Galagoides in Eastern Africa
ers to dispersal (Maji ya Chumvi, Mambome, and Pemba/Cha Shimba). A visit to Mazeras was, therefore, required to deter-mine whether the species-specific loud call of G. cocos at the type locality is the same as that of the dwarf galago at Diani.
Note that, in the above quotation, Grubb et al. (2003) assume that the holotype of G. cocos was collected in the for-est of ‘Kaya Mazeras’, which has since been destroyed. What Heller (1912) actually says about the type locality is:
“The Mazeras specimens were all obtained on the brushy borders of the cocoa-palm groves. These groves are the dominant feature in the landscape of the coast belt, and extend almost unbroken from the sea beaches inland a distance of ten or fifteen miles. They mark the tropical littoral zone more precisely than any other plant growth.” (Heller 1912, p.2).
In short, Heller obtained the holotype (and nine other specimens) of G. cocos in an area of coconut palms (Cocos nucifera) and bushland, and not in forest.
Confirmation of the Name ‘cocos’ for the Kenya Coast Galago
T. Butynski and Y. de Jong visited the Mazeras area on 10 –12 February 2004. The first night was spent searching for G. cocos in the Mazeras Botanical Garden (03°57′58′′S, 39°33′05′′E, 134 m a.s.l.) in Mazeras town. No dwarf galagos were heard or seen, although the small-eared greater galago, (Otolemur garnettii), was common (about eight individuals seen or heard).
The second night was spent searching for G. cocos on the edge of Kaya Chijembeni (Rabai) (03°56′42′′S, 39°34′54′′E, 210 m a.s.l.), a relatively large coastal forest located about 4 km northeast of Mazeras town, 17 km from the Indian Ocean, and 40 km north of Diani. Coconut palms are an extremely common species there on the forest edge and, with cashew nut (Anacardium occidentale) and mango (Mangifera indica), are scattered throughout the farmlands and bush lands that surround Kaya Chijembeni. Many dwarf galagos were heard and seen at this site (both on the forest edge and inside the forest), and tape recordings were acquired of ‘incremental’ advertising calls and other vocalizations. To our ears, the advertising call recorded in Kaya Chijembeni matched that recorded for the dwarf galago at Diani, as well as the advertising call that T. Butynski has heard many hun-dreds of times (and recorded) at others sites on the coast of Kenya (for example, Kilifi, Watamu, Gedi), and in forests along the lower Tana River (02°30′S, 40°30′E), c.150 km to the north of Mazeras.
That the incremental advertising call recorded from G. cocos at Kaya Chijembeni is the same as that recorded from the dwarf galago at Diani is confirmed through compari-sons of the sonograms and oscillograms of these calls (Figs. 9 and 11). The advertising calls from these two sites have a very similar incremental structure pattern, frequency range, and call duration.
T. Butynski and Y. de Jong heard about 100 G. cocos incremental calls during one night at Kaya Chijembeni. As at other sites where C. cocos is similarly common, there was a distinct ‘dusk chorus’ (c. 19:05–19:15h) of incremental calls, a much lower rate of incremental calls throughout the night, and a slight ‘dawn chorus’ (c. 05:45–05:55h) of incremental calls.
The only other species of galago heard at Kaya Chi-jembeni was O. garnettii. Thus, only two species of galagos were seen or heard in the Mazeras/Kaya Chijembeni/Rabai area — G. cocos and O. garnettii. Y. de Jong and T. Butynski returned to Kaya Chijembeni on 20 February 2006 to obtain better (digital) recordings of the dwarf galago (Fig. 11). Again, the only galagos heard were G. cocos and O. garnettii.
Incremental calls identical to those of topotypical G. cocos at Mazeras have been recorded (by A. Perkin, T. Butynski, Y. de Jong, S. Bearder, N. Cordiero, N. Svoboda, A. Kempson and S. Gregory) at several localities along the Kenya coast both north and south of Mazeras, as well as in the northern lowland coastal forests of the East Usambara Mountains in northeastern Tanzania. These calls were analyzed by A. Perkin and S. Bearder.
Visual examination of the type G. cocos and eight other adult specimens of G. cocos from Mazeras (housed at the United States National Museum, Washington, DC) reveal that they are not phenotypically different from the three spec-imens of G. cocos at the National Museums of Kenya that were collected along the Tana River (NMK 992), and on the Kenya coast at Gedi (NMK 5351) and Kipendi (no specimen number). One of the specimens at the National Museums of Kenya (MR14) was collected at Mrima Hill, Kenya, very near the border with Tanzania. This specimen has an intact penis, the morphology of which helps to identify this population as G. cocos (see below).
We conclude that there is now no doubt that the type of G. cocos that Heller (1912) described from Mazeras is conspecific with the dwarf galago found along much of the coast of Kenya and into northern Tanzania. In short, the name ‘cocos’ can correctly be used as the species name of the Kenya coast galago.
This clarification of the correct name for the Kenya coast galago has at least three important implications for previ-ous research conducted on G. cocos, G. zanzibaricus, and G. granti. First, most of the field research that has been con-ducted on the distribution, abundance, behavior, and ecology of ‘G. zanzibaricus’ was, in fact, conducted on G. cocos (for example, Harcourt 1984, 1986; Harcourt and Nash 1986a, 1986b; Harcourt and Bearder 1989). Second, the distinc-tive rolling calls of galagos on mainland Tanzania that led Honess (1996) and Honess and Bearder (1996) to name this form G. udzungwensis, belong to G. zanzibaricus (i.e., G. z. udzungwensis). Third, those researchers who obtained data from specimens initially assigned to ‘G. zanzibaricus’ have sometimes, unknowingly, combined data from two species (G. cocos and G. zanzibaricus), and, on occasion, from a third species (G. granti). This is especially the case for specimens collected from coastal Kenya, from coastal Tanzania south of

70
Butynski et al.
the East Usambara Mountains, and from Unguja Island, Zan-zibar. This means that (1) the results and conclusions of some previous studies of ‘G. zanzibaricus’ may need to be reviewed and reevaluated, and (2) that the providence of each and every specimen labeled ‘G. zanzibaricus’ must be known in order to help ensure that the specimen is not, in fact, G. cocos or G. granti.
Morphological Differences Among G. cocos, G. zanzibari-cus and G. granti
Galagoides cocos, G. zanzibaricus, and G. granti are among the most cryptic of primate species. That they are simi-lar phenotypically and morphologically is demonstrated by the inability of some of the foremost primate taxonomists of their time to differentiate among them. For example, Schwarz (1931), in reference to G. senegalensis zanzibaricus, states (p. 56):
“There can be no doubt that Heller’s cocos is identical with this race. The size, coloration, and the large upper M³ are found both in the series at Berlin and the one of cocos studied by Heller and Hollister. By the identification of the two the known range of zanzibaricus is considerably extended. There is no difference between the island and coast specimens.”
What is needed next is a detailed comparison of large numbers of G. cocos and G. zanzibaricus specimens to deter-mine their morphological differences, coupled with ecologi-cal, behavioral, acoustic, and molecular studies. Other than the highly distinctive species-specific advertising call, one of the differences noted thus far is that G. cocos is slightly larger than G. z. zanzibaricus. For example, the mean length of the head+body for G. cocos from southeast Kenya and northeast Tanzania is 158 mm (n = 46, range = 142–183 mm) (Appendix I, Tables A and B), while the mean length of the head+body for G. z. zanzibaricus from Unguja Island, Zanzibar, is 143 mm (n = 11, range 125–150) (Appendix I, Table C). This size dif-ference extends to body weight; G. cocos has a mean body weight of 144 g (n = 78, range = 117–172), whereas G. z. zanzibaricus has a mean body weight of 127 g (n = 10, range = 104–172).
Of the two subspecies of G. zanzibaricus, it appears that the island form, G. z. zanzibaricus, is smaller than the main-land form, G. z. udzungwensis (Appendix I, Tables C and D). Mean length of the head+body for G. z. udzungwensis is 162 mm (n = 17, range = 139–180) and mean body weight is 145 g (n = 6, range = 118–105). The data available suggest that G. granti is larger than G. z. zanzibaricus, and very simi-lar in size to G. z. udzungwensis and G. cocos (Appendix I, Tables E, F and G).
Color of the nose stripe, chin, throat, cheeks and ven-trum, length of the nose stripe, and length of the tail relative to length of the head+body, have all been proposed as useful for distinguishing among G. cocos, G. z. zanzibaricus, G. z. udzungwensis, and G. granti (for example, Elliot 1913; Nash et al. 1989; Honess 1996; Groves 2001). However, our stud-
ies lead us to conclude that there is (1) too much intraspecific variation and, especially, (2) too much interspecific overlap for these characters to serve as diagnostic features.
The full range of phenotypic variation present in G. cocos, G. z. zanzibaricus, G. z. udzungwensis, and G. granti remains unknown. Although they need to be examined quantitatively, and with sample sizes far larger than those currently available, the following phenotypic characters hold promise for distin-guishing among G. cocos, G. z. zanzibaricus, G. z. udzungwen-sis, and G. granti, especially when taken together:
Muzzle patchesG. cocos – patch on either side of muzzle dark, blackish, and prominent (Figs. 1 and 2).G. z. zanzibaricus – patch on either side of muzzle less dark, grayish, and less prominent.G. z. udzungwensis – patch on either side of muzzle less dark, grayish, and less prominent (Figs. 3 and 4).G. granti – patch on either side of muzzle less dark, grayish, and less prominent (Fig. 5).
EarsG. cocos – seldom longer than 38 mm, dusky behind.G. z. zanzibaricus – seldom longer than 35 mm, dusky behind.G. z. udzungwensis – seldom longer than 33 mm, dusky behind.G. granti – seldom shorter than 37 mm, blackish behind. Ears not only long but also relatively broad (Fig. 5).
DorsumG. cocos – hairs c.10 mm long, tipped buffy-brown (Figs. 1 and 6). G. z. zanzibaricus – hairs c.8 mm long, tipped cinnamon or rufous-cinnamon (Fig. 7).G. z. udzungwensis – hairs c.9 mm long, tipped buffy-brown (Figs. 6 and 7).G. granti – hairs c.12 mm long, tipped buffy-brown with slight pinkish tint (Figs. 5 and 6).
TailG. cocos – even length hairs over tail; hairs dense, c.14 mm long, soft. Proximal c.25% of tail same color as dorsum (i.e., buffy-brown); distal c.33% dark buffy-brown in some (Mazeras) specimens, but same color as dorsum in other specimens (Fig. 6).G. z. zanzibaricus – even length hairs over tail; hairs sparse, c.13 mm long, wiry, rufous-cinnamon, cinnamon, or dusky-cinnamon (highly variable). Tail darker/brighter cinnamon than dorsum, and either evenly colored or with gradual darkening to reddish or dusky toward tip (Fig. 7).G. z. udzungwensis – even length hairs over tail; hairs sparse, c.11 mm, wiry. Proximal c.75% of tail same color as dorsum (i.e., buffy-brown); distal c.25% slightly darker brown or dusky. Some with tail tipped white (Figs. 4, 6, and 7).G. granti – bushy, wider over distal c.80%; hairs dense, c.15 mm long, soft. Tail darker than dorsum with distal c.10 – 60% blackish-brown. Some with tail tipped white (Fig. 6).
Of these four taxa, G. z. zanzibaricus and G. granti are phenotypically the most distinctive. The dorsum of G. z. zan-zibaricus is cinnamon and the tail has at least some rufous, whereas the dorsum of the other three taxa is buffy-brown and all lack rufous in the tail (Fig. 7). The color of the dorsum of G. cocos, G. z. udzungwensis, and G. granti is virtually identical, although there is a slight pinkish tint to the dorsum of G. granti when seen in good light (Fig. 6).
When observed at close range in the field, G. granti is distinguished from G. cocos, G. z. zanzibaricus, and G. z. udzungwensis by its relatively large, broad, round, and black-ish (behind) ears, and by the very full, bottlebrush-shaped tail, which is blackish-brown over the distal part (Figs. 5 and 6).

71
Galagoides in Eastern Africa
The particularly large ears of G. granti have been noted previ-ously (for example, Honess 1996; Masters and Bragg 2000).
As a species, G. zanzibaricus is probably best distin-guished phenotypically from G. cocos and G. granti by the relatively short, wiry, stiff hairs over the tail. This hair type makes it relatively easy to see the skin of the tail through the pelage (Fig. 4). Galagoides cocos and G. granti have rela-tively long, soft, lax hairs over the tail.
Of the four taxa considered here, G. cocos and G. z. udzungwensis are, phenotypically, the most difficult to distin-guish from one another (Fig. 6). The presence in G. cocos of a prominent dark, blackish patch on either side of the muzzle is probably the best phenotypic character available for distin-guishing C. cocos and G. z. udzungwensis in the field (Figs. 1–4) (A. Perkin pers. obs.). As already stated, however, all of the characters listed above are in need of detailed quantitative study in order to determine their reliability, both in the field and in the museum.
The penile morphology of G. cocos, G. zanzibaricus, and G. granti is diagnostic (Fig. 12). For details, see Hon-
ess (1996), Honess and Bearder (1996), Anderson (2000), and Perkin (in press). See sketches in Kingdon (1997).
Vision plays an important role in the life histories of all galagos, perhaps especially for species recognition. Detailed study of the light and dark facial markings of these four taxa may reveal that they are species-specific and, therefore, a useful diagnostic tool. This is not only a priority topic for research related to the search for species-typical differences among G. cocos, G. z. zanzibaricus, G. z. udzungwensis, and G. granti, but also among the many other cryptic taxa within the Galagidae (Bearder 1999; Bearder et al. 2006).
The differences noted here among G. cocos, G. zanzibari-cus, and G. granti in their species-specific advertising calls, body measurements, and phenotypic characters are consistent with the species-level differences observed for other species in the Galagidae (Honess 1996; Honess and Bearder 1996; Masters and Bragg 2000; Masters and Brothers 2002).
In contrast to the great similarity among G. cocos, G. zan-zibaricus, and G. granti, these three species are readily distin-guished from the other seven species of galagos with which one or all are sympatric or parapatric. These are O. garnet-tii, the thick-tailed (or large-eared) greater galago (Otolemur crassicaudatus), Somali lesser galago (Galago gallarum), northern lesser galago (Galago senegalensis), southern lesser galago (Galago moholi), mountain dwarf galago (Galagoides orinus), and Rondo dwarf galago (Galagoides rondoensis). The main morphological characters for distinguishing among G. cocos, G. gallarum, and G. senegalensis are summarized in Butynski and De Jong (2004).
Geographic Ranges of G. cocos, G. zanzibaricus and G. granti
Galagoides cocos occurs in evergreen forest all along the coastal strip (plain) of Kenya, south of the Tana River (Nash et al. 1989; Bearder et al. 2003; Grubb et al. 2003) southward to at least the Mgambo Forest Reserve in northern Tanzania at the north end of the East Usambara Mountains (A. Perkin unpubl. data) (Fig. 13). Galagoides cocos is reported to occur as far north as the Webi Shabeelle River in southern Somalia (Nash et al. 1989), but this needs confirmation.
Thomas Butynski recorded the advertising call of a galago in the Ololua Forest, Nairobi, that S. Bearder identi-fied as that of the G. cocos. Ololua Forest is c.390 km inland from the coast of Kenya and, at 1,850 m a.s.l., well above the known altitudinal range for G. cocos elsewhere (0 –350 m). This record for Ololua Forest requires confirmation.
Galagoides cocos and G. z. udzungwensis are parapatric or, perhaps, sympatric at a few sites c.2–8 km to the north of the East Usambara Mountains in the coastal strip of north-eastern Tanzania (Figs. 13 and 14) (A. Perkin in litt. in Grubb et al. 2003). Although there is a complex mosaic of habitat types in this region, preliminary observations indicate that G. cocos is present in the dry mixed coastal forests and mixed woodland of the northernmost forests of Tanzania’s coastal strip (for example, Bombo East I and Bombo East II For-
Figure 12. Schematic drawings showing the penile morphology of: 1 – Ga-lagoides cocos; 2 – Galagoides granti; 3 – Galagoides zanzibaricus udzun-gwensis. A – ventral view; B – dorsal view; C – lateral view (dorsum right). Scale is indicated on the right. See Perkin (in press) for details. Adapted from Perkin (in press).

72
Butynski et al.
Figure 13. Approximate geographic distribution of the Kenya coast galago (Galagoides cocos). The shaded area extends out 5 km from the center of each locality point. The distribution of this species remains poorly known. Names of the sites plotted on this map, and the sources of these data, are available from Yvonne de Jong (e-mail: <[email protected]>).
Nairobi
Machakos
Arusha
Kiboko
Mwingi
Kitui
Same
Voi
AmaniTanga
Diani
Mazeras
Mombasa
Hola
Malindi
Garsen
Gedi
0 50
kilometers100
0
kilometers
50 100
KENYA
TANZANIAIN
DIAN
OCEAN
LEGEND
G. cocos
TanaR
iver
Athi
Rive
r
Pangani River
distribution
G. cocos unconfirmed
G. cocos locality
Sabaki River
Galana
River
Pemba
- 3°
37° 38° 39° 40°
- 4°
- 5°
- 2°
- 1°
Zanzibar
(Unguja)

73
Galagoides in Eastern Africa
Figure 14. Approximate geographic distributions of the Zanzibar galago (Galagoides zanzibaricus) The shaded area extends out 5 km from the center of each locality point. The distribution of this species remains poorly known. Names of the sites plotted on this map, and the sources of these data, are available from Yvonne de Jong (e-mail: <[email protected]>).
Dodoma
Farkwa
Kondoa
Iringa
Kilosa
Kihansi
Ifakara
Mang'ula
Mikumi
Mahenge
Same
Babati
Morogoro
Handeni
Amani
Utete
Tanga
Dar es Salaam
Kisiju
Kilwa Kivinje
Mombasa
0
kilometers50 100
KENYA
TANZANIA
IND
IAN
OC
EAN
Zanzibar
Pangani River
Wami River
Ruvu
Rive
r
Rufiji River
LEGEND
G. zanzibaricus distribution
G. zanzibaricus locality
Pemba
Mafia
(Unguja)
G. zanzibaricus unconfirmed
Great Ruaha River
Rufiji
River
37°36° 38° 39° 40°
- 8°
- 7°
- 6°
- 5°
- 4°
- 3°

74
Butynski et al.
est Reserves), as well as in the more moist (but relatively tree-species and bird-species poor) groundwater forests and adjacent woodlands on the lower northern slopes of the East Usambara Mountains (for example, Mgambo Forest Reserve) (A. Perkin and N. Cordeiro unpubl. data). In this region, G. z. udzungwensis appears to be confined to the more moist (and relatively tree-species and bird-species rich) forest on the eastern slopes of the East Usambara Mountains (for example, Kambai, Segoma, Manga, and Marimba forest reserves) (A. Perkin unpubl. data).
Galagoides z. udzungwensis and G. granti appear to be parapatric at the Kilombero-Rufiji River with G. z. udzun-gwensis reaching its southern limit on the north (left) bank (Fig. 14) and G. granti reaching its northern limit on the south (right) bank (Fig. 15) (Honess 1996; A. Perkin in litt. in Grubb et al. 2003). In addition, the geographic ranges of G. z. udzungwensis and G. granti either approach one another or meet at the southern end of the Udzungwa Mountains in south-central Tanzania. Here, G. granti is present at 1,500 to 1,800 m a.s.l. in the Lulanda Forest Reserve at the headwa-ters of the Kilombero River (A. Perkin in litt. in Grubb et al. 2003; A. Perkin unpubl. data), and G. z. udzungwensis is pres-ent from 400 to 1,070 m a.s.l. at Kihanzi (which is c.24 km to the east of Lulanda) (Honess 1996; Butynski et al. 1998; N. Cordeiro pers. comm.). Kihanzi is the known southwestern limit for G. z. udzungwensis, and Lulanda is the known north-western limit for G. granti.
David Moyer and E. Mulungu (pers. comm.) tape recorded the loud call of a Galagoides sp. at three sites in extreme western Tanzania: Mbala Forest, Sitebe-Sifuta Mountains (6º04′40′′S, 30º32′10′′E, 1,700 m a.s.l., 16 August 2005), Mahale Mountains National Park at Mfitwa Mountain (6º07′55′′S, 29º47′38′′E, 2,440 m a.s.l., 20 November 2005), and at Pasagulu Mountain (6°03′47′′S, 29°45′14′′E, 1,500 m a.s.l., 6 February 2006). The species recorded may have been G. granti. If so, this extends its geographic range c.700 km to the northwest (Fig. 15).
Groves (2001) identified three specimens collected in sub-montane forest at Bagilo (800 –1,000 m a.s.l.) in the Uluguru Mountains (south of Morogoro, Fig. 15) as G. cf. granti (Grubb et al. 2003). If these are G. granti, then the Uluguru Mountains represent the northernmost site for this species, and the only known site for G. granti that is north of the Rufiji River. These three specimens had earlier been identified by Lawrence and Washburn (1936) as G. senegalensis zanzibaricus. A. Perkin has since examined them at Harvard University’s Museum of Comparative Zoology (specimen numbers: 22450, 22449, 22451) and also ascribes them to G. zanzibaricus. In 1993, P. E. Honess and S. K. Bearder (Honess 1996) visited the forest at Bagilo but did not find G. granti there — only G. orinus was present. Unfortunately, there is no longer any forest at Bagilo (Perkin 2000; Doggart et al. 2004) but A. Perkin (unpubl. data) found G. zanzibaricus at 900 m a.s.l. on the edge of the Ulu-guru North Forest Reserve (c.2 km west of Bagilo Village) and up to 700 m a.s.l. in the Uluguru Mountains.
The known southern limit for G. granti is the Limpopo River in southern Mozambique. The western limit in the south-ern part of the range appears to be extreme eastern Zimbabwe (Smithers and Lobão Tello 1976; Smithers and Wilson 1979; Skinner and Smithers 1990).
In summary, present information indicates that G. cocos, G. zanzibaricus, and G. granti are parapatric, or narrowly sym-patric, species of the evergreen forests of the coastal strip of eastern Africa from northern Kenya (perhaps southern Somalia) to extreme southern Mozambique and extreme eastern Zimba-bwe. Galagoides cocos is the northern species, G. zanzibaricus is the central species, and G. granti is the southern species.
Conservation Status of G. cocos, G. zanzibaricus, and G. granti
Galagoides cocos, G. zanzibaricus, and G. granti now survive in highly fragmented, probably declining, populations as a result of the extensive (>65%) loss of eastern Africa’s original coastal forest cover. More than 90% of the original coastal forest of Kenya and Tanzania has either been destroyed or degraded (Burgess et al. 2004). Fortunately, all three species are able to persist in secondary forest and in mosaics of mixed agriculture where some forest remains.
As a recently resurrected species (Grubb et al. 2003), G. cocos does not yet appear on the IUCN Red List. It is the most abundant and widespread galago in the coastal forests of Kenya, with densities of approximately 170 –180 individuals/km² both at Gedi and Diani (Harcourt and Nash 1986a). Gala-goides cocos is found at elevations from sea level to at least 210 m a.s.l. over the coastal zone of Kenya (T. Butynski and Y. de Jong unpubl. data), and to at least 350 m a.s.l. in the foothills of the East Usambara Mountains, Tanzania (A. Perkin unpubl. data). The information available indicates that G. cocos, when assessed for the IUCN Red List using the 2001 criteria (IUCN 2001), will be placed in the Least Concern category.
G. zanzibaricus is listed as Lower Risk/Near Threatened on the 2006 IUCN Red List (IUCN 2006), but assessed only using the 1994 criteria (IUCN 1994). G. zanzibaricus is the most abundant and widespread galago in the coastal forests of Tanzania, including ‘coastal’ forest sites located at least 370 km inland (for example, Udzungwa Mountains) to c.1,100 m a.s.l. The density of G. zanzibaricus varies greatly from site to site. In the Udzungwa Mountains (for example, Matundu Forest Reserve), G. z. udzungwensis is estimated to occur at densities of more than 500 individuals/km² (Butynski et al. 1998), whereas <100 individuals/km² occur at many other sites (T. Butynski and A. Perkin pers. obs.). Although G. z. zanzi-baricus is confined to Unguja Island, it is widespread over the eastern and southern parts of the island (Lumsden and Masters 2001), and is common in at least some places (e.g., >200 indi-viduals/km² in Jozani–Chwaka Bay National Park (T. Butynski and Y. de Jong pers. obs). The information available indicates that G. zanzibaricus, when reassessed for the IUCN Red List, should be placed in the ‘Least Concern’ category.

75
Galagoides in Eastern Africa
Figure 15. Approximate geographic distributions of the Mozambique galago (Galagoides granti). The shaded area extends out 20 km from the center of each locality point. The distribution of this species remains poorly known. Names of the sites plotted on this map, and the sources of these data, are available from Yvonne de Jong (e-mail: <[email protected]>).
??Kabala
Bulawago
Francistown
Lusaka
Kasama
Ndola
Lubumbashi
Maputo
Pietersburg
Kigoma
LakeTanganyika
Harare
Masvingo
Chimoio
Tambarara
Tabora
Mbeya
Lilongwe
Songo
Chipata
Beira
PembeCoguno
Songea
Lake
Mal
awi
Dodoma
Iringa
Lulanda
CuambaNacala
Quelimane
Blantyre
Tanga
Morogoro
Mang'ula
Mahenge
Lindi
MombasaSame
Dar es Salaam
0 200
kilometers400
INDI
ANO
CEAN
TANZANIA
MOZAMBIQUE
ZIMBABWE
ZAMBIA
MA
LAW
IRuvuma River
Rufiji River
Zambezi River
Limpopo
River
LEGEND
G. granti distribution
G. granti locality
G. granti unconfirmed
Pemba
Zanzibar(Unguja)
DRC
SOUTH AFRICA
- 14°
- 16°
- 18°
- 20°
- 22°
- 24°
- 12°
- 10°
- 8°
- 6°
- 4°
- 26°
28° 30° 32° 34° 36° 38° 40° 42° 44°
Mafia

76
Butynski et al.
The two subspecies, G. z. zanzibaricus and G. z. udzun-gwensis, have not as yet been assessed for the IUCN Red List. While G. z. udzungwensis is a widespread subspecies that will likely be assessed as Least Concern, G. z. zanzibaricus has a far smaller ‘extent of occurrence’, being endemic to Unguja Island, Zanzibar (c.2,000 km²) where rates of habi-tat degradation, loss, and fragmentation are particularly high. As such, G. z. zanzibaricus is expected to be assessed as an Endangered taxon.
Galagoides granti, assessed under the 1994 criteria (IUCN 1994), is listed as Data Deficient in the 2006 IUCN Red List (IUCN 2006). Galagoides granti is present in coastal and submontane evergreen forest, gallery forest, and species-rich woodlands, including some hilly miombo woodlands (e.g., Mahenge foothills). In the southern part of its range, G. granti is present between the coast and about 200 km inland (i.e., eastern Zimbabwe) up to 360 m a.s.l. (Smithers and Wilson 1979; Skinner and Smithers 1990). According to the specimen tags, C. H. B. Grant collected this species up to 400 m a.s.l. at Tambarara in central Mozambique. In the northern part of its range, G. granti occurs from the coast to at least 310 km inland (i.e., Lulanda, Tanzania) up to at least 1,800 m a.s.l. Galagoides granti occurs over a much greater range of habitat types, altitudes, and climates than does G. cocos or G. zanzibaricus. There is now enough information available for an IUCN Red List assessment of this species. The available data indicate that G. granti, once reassessed, will be placed in the Least Concern category. Galagoides cocos, G. zanzibari-cus, and G. granti are all currently CITES Appendix II species (http://www.cites.org).
Acknowledgments
We thank David Moyer and Esteban Sarmiento for their unpublished data, the curatorial staff at the United States National Museum, British Museum of Natural History, and American Museum of Natural History for providing access to their collections; Harald Schuetz for the photograph in Fig-ure 1; and Nike Doggart for the photograph in Figure 3. We appreciate the support given to our research on primates in eastern Africa by the Margot Marsh Biodiversity Foundation, Conservation International, Zoo Atlanta, the National Muse-ums of Kenya, the IUCN Eastern Africa Regional Office, the Kenya Institute of Primate Research, the Critical Ecosystem Partnership Fund, the Primate Action Fund, Primate Conser-vation Inc., the Leverhulme Trust, Oxford Brookes University, WWF-Tanzania, the Tanzanian Commission for Science and Technology, Tanzania Forestry Division, Tanzania National Parks, Kenya Wildlife Service, and Kenya Ministry of Edu-cation, Science and Technology. Norbert Cordeiro, Nadine Svoboda, Alex Kempson, and Stephen Gregory provided tape recordings of the advertising calls of G. cocos. Norbert Cor-deiro assisted with vegetation descriptions and unpublished data. Julie Anderson and the Colobus Trust helped with sur-veys funded by the Born Free Foundation.
Literature Cited
Allen, G. M. 1939. A checklist of African mammals. Bull. Mus. Comp. Zool. Harvard College 83: 1–763.
Allen, G. M. and A. Loveridge. 1927. Mammals from the Uluguru and Usambara Mountains, Tanganyika Territory. Proc. Boston Soc. Nat. Hist. 38: 425– 426.
Ambrose, L. 1999. Species diversity in West and Central Afri-can galagos (Primates, Galagonidae): The use of acous-tic analysis. PhD thesis, Oxford Brookes University, Oxford.
Ambrose, L. 2003. Three acoustic forms of Allen’s galagos (Primates: Galagonidae) in the Central African region. Primates 44: 25–39.
Anderson, M. J. 1999. The use of hand morphology in the taxonomy of galagos. Primates 40: 469– 478.
Anderson, M. J. 2000. Penile morphology and classification of bush babies (subfamily Galagoninae). Int. J. Primatol. 21: 815–836.
Bearder, S. K. 1999. Physical and social diversity among noc-turnal primates: A new view based on long-term research. Primates 40: 267–282.
Bearder, S. K., P. E. Honess and L. Ambrose. 1995. Species diversity among galagos with special reference to mate recognition. In: Creatures of the Dark: The Nocturnal Prosimians, L. Alterman, G. A. Doyle and M. K. Izard (eds.), pp.331–352. Plenum Press, New York.
Bearder, S., L. Ambrose, C. Harcourt, P. Honess, A. Perkin, E. Pimley, S. Pullen and N. Svoboda. 2003. Species-typi-cal patterns of infant contact, sleeping site use and social cohesion among nocturnal primates in Africa. Folia Pri-matol. 74: 337–354.
Bearder, S. K., K. A. I. Nekaris and D. J. Curtis. 2006. A re-evaluation of the role of vision in the activity and com-munication of nocturnal primates. Folia Primatol. 77: 50 –71.
Burgess, N, J. Salehe, N. Doggart, G. P. Clark, I. Gordon, P. Sumbi and A. Rodgers. 2004. Coastal forests of east-ern Africa. In: Hotspots Revisited, R. A. Mittermeier, P. Robles Gil, M. Hoffmann, J. Pilgrim, T. Brooks, C. G. Mittermeier, J. Lamoreux and G. A. B. da Fonseca (eds.), pp.230 –239. CEMEX, Agrupación Serra Madre, SC, México.
Butynski, T. M., C. L. Ehardt and T. T. Struhsaker. 1998. Notes on two dwarf galagos (Galagoides udzungwensis and Galagoides orinus) in the Udzungwa Mountains, Tanzania. Primate Conserv. (18): 69–75.
Butynski, T. M. and Y. A. de Jong. 2004. Natural history of the Somali lesser galago (Galago gallarum). J. E. Afr. Nat. Hist. 93: 24–38.
Courtenay, D. O. and S. K. Bearder. 1989. The taxonomic sta-tus and distribution of bushbabies in Malawi with empha-sis on the significance of vocalizations. Int. J. Primatol. 10: 17–34.
DelPero, M., J. C. Masters, D. Zuccon, P. Cervella, S. Crov-ella and G. Ardito. 2000. Mitochondrial sequences as

77
Galagoides in Eastern Africa
indicators of generic classification in bush babies. Int. J. Primatol. 21: 889–904.
Doggart, N., J. Lovett, B. Mhoro, J. Kiure, A. Perkin and N. D. Burgess. 2004. Biodiversity surveys in the forest reserves of the Uluguru Mountains: Descriptions of the biodiver-sity of individual forest reserves. Report, Tanzania Forest Conservation Group, Dar es Salaam, Tanzania.
Elliot, D.G. 1913. A Review of the Primates. Vol. 3. American Museum of Natural History, New York.
Groves, C. P. 1977. Taxonomy and phylogeny of prosim-ians. In: Prosimian Anatomy, Biochemistry and Evolu-tion, R. D. Martin, G. A. Doyle and A. C. Walker (eds), pp.449– 473. Duckworth, London.
Groves, C. P. 1993. Order Primates. In: Mammal Species of the World: A Taxonomic and Geographic Reference, 2nd Ed., D. E. Wilson and D. M. Reeder (eds.), pp.243 –277. Smithsonian Institution Press, Washington, DC.
Groves, C. P. 2001. Primate Taxonomy. Smithsonian Institu-tion Press, Washington, DC.
Groves, C. P. 2005. Order Primates. In: Mammal Species of the World: A Taxonomic and Geographic Refer-ence, 3rd edi. D. E. Wilson and D. M. Reeder (eds.), pp.111–184.. Johns Hopkins University Press, Baltimore, Maryland.
Grubb, P., T. M. Butynski, J. F. Oates, S. K. Bearder, T. R. Disotell, C. P. Groves and T. T. Struhsaker. 2003. Assess-ment of the diversity of African primates. Int. J. Primatol. 24: 1301–1357.
Harcourt, C. S. 1984. The behaviour and ecology of galagos in Kenyan coastal forest. PhD thesis, University of Cam-bridge, Cambridge, UK.
Harcourt, C. S. 1986. Galago zanzibaricus: Birth seasonal-ity, litter size and perinatal behaviour of females. J. Zool., Lond. 210: 451– 457.
Harcourt, C. S. and S. K. Bearder. 1989. A comparison of Galago moholi in South Africa with Galago zanzibaricus in Kenya. Int. J. Primatol. 10: 35– 45.
Harcourt, C. S. and L. T. Nash. 1986a. Species differences in substrate use and diet between sympatric galagos in two Kenyan coastal forests. Primates 27: 41–52.
Harcourt, C. S. and L. T. Nash 1986b. Social organization of galagos in Kenyan coastal forests. 1. Galago zanzibari-cus. Am. J. Primatol. 10: 339–355.
Heller, E. 1912. New races of insectivores, bats, and lemurs from British East Africa. Smithson. Misc. Coll. 60: 1–13.
Hill, W. C. O. 1953. Primates: Comparative Anatomy and Taxonomy. I. Strepsirhini. University of Edinburgh Press, Edinburgh, UK.
Hill, W. C. O. and J. Meester. 1977. Suborder Prosimii, Infra-order Lorisformes. In: The Mammals of Africa: An Iden-tification Manual, Part 3.2., J. Meester and H. W. Setzer (eds.). Smithsonian Institution Press, Washington, DC.
Hollister, N. 1924. East African mammals in the United States National Museum. Part III. Primates, Artiodactyla, Pro-boscidea, and Hyracoidea. Bull. U. S. Natl. Mus. 99: 1–164.
Honess, P. E. 1996. Speciation among galagos (Primates: Galagidae) in Tanzanian forests. PhD Thesis, Oxford Brookes University, Oxford, UK.
Honess, P. E. and S. K. Bearder. 1996. Descriptions of the dwarf galago species of Tanzania. African Primates 2: 75–79.
IUCN. 1994. IUCN Red List Categories. IUCN Species Sur-vival Commission, Gland, Switzerland.
IUCN. 2001. IUCN Red List Categories and Criteria: Version 3.1. IUCN Species Survival Commission, Gland, Swit-zerland.
IUCN. 2006. 2006 IUCN Red List of Threatened Species. Website: www.iucnredlist.org. Accessed 15 May 2006.
Jenkins, P. D. 1987. Catalogue of Primates in the British Museum (Natural History) and Elsewhere in the British Isles. Part IV. Suborder Strepsirrhini, Including the Sub-fossil Madagascan Lemurs and Family Tarsiidae. British Museum (Natural History), London.
Kingdon, J. 1971. East African Mammals. Vol. 1. Academic Press, London.
Kingdon, J. 1997. The Kingdon Field Guide to African Mam-mals. Academic Press, London.
Lawrence, B. and S. L. Washburn. 1936. A new eastern race of Galago demidovii. Occ. Pap. Boston Soc. Nat. Hist. 8: 255–266.
Lumsden, W. H. R. and J. C. Masters. 2001. Galago (Galago-nidae) collections in East Africa (1953 –1955): Ecology of the study areas. African Primates 5: 37– 42.
Masters, J. C. 1991. Loud calls of Galago crassicaudatus and G. garnettii and their relation to habitat structure. Pri-mates 32: 149–174.
Masters, J. C. 1998. Speciation in the lesser galagos. Folia Primatol. 69(Suppl.1): 357–370.
Masters, J. C. and N. P. Bragg. 2000. Morphological corre-lates of speciation in bush babies. Int. J. Primatol. 21: 793 –813.
Masters, J. C. and D. J. Brothers. 2002. Lack of congruence between morphological and molecular data in recon-structing the phylogeny of the Galagonidae. Am. J. Phys. Anthropol. 117: 79–93.
Nash, L. T., S. K. Bearder and T. R. Olson. 1989. Synopsis of galago species characteristics. Int. J. Primatol. 10: 57–80.
Perkin, A. W. 2000. The taxonomic status of bushbabies (gala-gos) in the Uluguru Mountains. Report for the Wildlife Conservation Society of Tanzania, Dar es Salaam. Web-page: www.africanconservation.com/uluguru. Accessed 8 May 2006.
Perkin, A. W. In press. Comparative penile morphology of East African galagos of the genus Galagoides (Family Galagidae): Implications for taxonomy. Amer. J. Prima-tol.
Perkin, A. W., S. K. Bearder, T. M. Butynski, B. Agwanda and B. Bytebier. 2002. The Taita mountain dwarf galago Galagoides sp.: A new primate for Kenya. J. E. Afr. Nat. Hist. 91: 1–13.

78
Butynski et al.
Schwarz, E. 1931. On the African long-tailed lemurs or gala-gos. Ann. Mag. Nat. Hist. 7: 41– 66.
Skinner, J. D. and R. H. N. Smithers. 1990. The Mammals of the Southern African Subregion. University of Pretoria, Pretoria.
Smithers, R. H. N. and J. L. P. Lobão Tello. 1976. Check list and atlas of the mammals of Mocambique. Museum Mem-oir, Natl. Museums and Monuments of Rhodesia No. 8.
Smithers, R. H. N. and V. J. Wilson. 1979. Check list and atlas of the mammals of Zimbabwe Rhodesia. Museum Mem-oir, Natl. Museums and Monuments of Rhodesia No. 9.
Zimmermann, E. 1990. Differentiation of vocalisations in bush-babies (Galaginae, Prosimiae, Primates) and the signifi-cance for assessing phylogenetic relationships. Z. Zoolo-gische Systematik und Evolutionsforschung 28: 217–239.
Zimmermann, E., S. K. Bearder, G. A. Doyle and A. B. Ander-sson. 1988. Variations in vocal patterns of lesser bushba-bies (Galago senegalensis and Galago moholi) and their implications for taxonomic relationships. Folia Primatol. 51: 87–105.
Authors’ addresses:Thomas M. Butynski, Eastern Africa Regional Program, Conservation International, c/o IUCN EARO, P.O. Box 68200, 00200 Nairobi, Kenya.Yvonne A. de Jong, Kenya Institute of Primate Research, P.O. Box 38, Ol Kalou 20303, Kenya.Andrew W. Perkin and Simon K. Bearder, Nocturnal Pri-mate Research Group, Anthropology Department, School of Social Sciences and Law, Oxford Brookes University, Oxford OX3 0BP, UK.Paul E. Honess, Department of Veterinary Services, Univer-sity of Oxford, Oxford OX1 3PT, UK.
Received for publication: February 2006Revised: May 2006
Appendix I
Body measurements for adults of the Kenya coast galago (Gala-goides cocos), the Zanzibar galago (Galagoides zanzibaricus), and the Mozambique (galago (Galagoides granti). All measurements are either known to be, or believed to be, from living or fresh adult specimens.
Table A. Measurements for eight Galagoides cocos collected at Mazeras, Ke-nya, the type locality for G. cocos (Hollister 1924). All measurements taken from the tags of specimens collected by E. Heller and housed at the United States National Museum (specimen numbers: 181810, 184218, 184219, 184220, 184221, 184222, 184223, and 184225). Because adult male and adult female body linear measurements for G. cocos are not significantly different (Harcourt and Nash 1986b), the data for males and females are combined in this sample.
Measurement Mean SD RangeSample size (n)
Head+body length 155 mm 6 145–165 mm 8Tail length 213 mm 11 200 –230 mm 8Hindfoot length 57 mm 3 53 – 60 mm 7Ear length 35 mm 2 33 –38 mm 7
Table B. Measurements for live specimens of Galagoides cocos. These data derive from the following two sources: (1) Harcourt and Nash (1986b) for G. cocos at Gedi (c.100 km northeast of Mombasa) and Diani (30 km south of Mombasa) on the coast of Kenya; (2) A. Perkin (unpubl. data) for G. cocos at Arabuko-Sokoke Forest (c.100 km northeast of Mombasa, n = 2), Diani (n = 3), and East Usambara Mountains (c.150 km southwest of Mombasa (n = 1). Because adult male and adult female body linear measurements for G. cocos are not significantly different, the data for males and females are combined in this sample (Harcourt and Nash 1986b). Because body weights of adult male and adult (non-pregnant) female G. cocos are significantly different (Harcourt and Nash 1986b), the body weight data are presented separately for each sex.
Measurement Mean RangeSample size
(n)
Head + body length 159 mm 142–183 mm 38Tail length 212 mm 182–230 mm 30Hindfoot length 54 mm 40 –57 mm 49Ear length 30 mm 23 – 40 mm 32
Body weight (males) 150 g 135–167 g 36Body weight (females) 138 g 117–172 g 42
Table C. Combined measurements for male and female Galagoides zanzibari-cus zanzibaricus from Unguja Island, Zanzibar, Tanzania, the type locality for G. zanzibaricus. Measurements taken from one specimen captured and released by A. Perkin and from the tags of 10 specimens housed at the British Muse-um of Natural History. Nine of these collected by W. H. R. Lumsden (speci-men numbers 1964.971, 1964.972, 1964.974, 1964.975, 1964.977, 1964.978, 1964.979, 1964.980, and 1964.981) and one obtained by an unknown collector (specimen number: 1955.331).
Measurement Mean SD RangeSample size
(n)
Head+body length 143 mm 8 125–150 mm 11Tail length 214 mm 12 198–235 mm 11Hindfoot length 56 mm 3 51–59 mm 11Ear length 32 mm 2 30 –35 mm 11
Body weight 127 g 20 104–172 g 10
Table D. Combined measurements for male and female Galagoides zanzibari-cus udzungwensis obtained from the following sites in Tanzania; two from Matundu Forest Reserve (Honess 1996); two from Kissarawe (housed at the British Museum of Natural History); two from Pugu Forest Reserve, three from Pande Game Reserve (A. Perkin, unpubl. data); four from Bagilo, Uluguru Mountains; and four from Amboni, near Tanga. These last eight specimens were collected by A. Loveridge and are housed at the Museum of Comparative Zoology, Harvard University.
Measurement Mean SD RangeSample size
(n)
Head+body length 162 mm 11 139–180 mm 17Tail length 222 mm 16 202–270 mm 17Hindfoot length 58 mm 6 50 –70 mm 17Ear length 31 mm 3 25–37 mm 17
Body weight 145 g 27 118–195 g 6

79
Galagoides in Eastern Africa
Table E. Combined measurements for male and female Galagoides granti collected at Coguno (type locality) and Tambarara, Mozambique, by C. H. B. Grant during the Rudd Expedition. Coguno is the type locality. Measurements taken from specimen tags. All 12 specimens housed at the British Museum of Natural History (specimen numbers: 906.11.8.5, 1906.11.8.6, 1906.11.8.7, 1906.11.8.8, 1906.11.8.9, 1906.11.8.10, 1908.1.1.12, 1908.1.1.13, 1908.1.1.14, 1908.1.1.15, 1908.1.1.16, and 1908.1.1.129).
Measurement Mean SD RangeSample size
(n)
Head+body length 153 mm 6 140 –160 mm 12Tail length 230 mm 6 216 –237 mm 12Hindfoot length 58 mm 3 54– 63 mm 12Ear length 38 mm 2 36 – 43 mm 12
Table F. Combined measurements for male and female Galagoides granti from eastern Zimbabwe (Smithers & Wilson 1979).
Measurement Mean RangeSample size
(n)
Head+body length 162 mm Not provided 10Tail length 232 mm 214–254 mm 10Hindfoot length 62 mm 59– 63 mm 10Ear length 40 mm 38– 41 mm 9
Weight 165 g 139–178 mm 6
Table G. Combined measurements for two male and one female Galagoides granti from Tanzania, two from Kichi Hills Forest Reserve, and one from Lu-lunda, Udzungwa Mountains (A. Perkin unpubl. data).
Measurement Mean SD RangeSample size
(n)
Head+body length 164 mm 15 154–181 mm 3Tail length 214 mm 7 208–222 mm 3Hindfoot length 58 mm 1 58–59 mm 3Ear length 38 mm 1 37–38 mm 2
Body weight 136 g 25 110 –160 g 3

50 SWARA July – September 2008
TsavoBy James Culverwell, Jim Feely, Sheila Bell-Cross, Yvonne A. de Jong and Thomas M. Butynski
There are only two spe-cies of warthog, and both occur in Kenya; the common warthog Phacochoerus afri-canus and the desert
warthog Phacochoerus aethiopicus. Little is known about the natural history of the desert warthog—in-deed, the desert warthog might well
be Africa’s least-known, non-forest, large mammal. The morphological differences between the two species of warthog are described by Grubb (1993) and by d’Huart & Grubb (2005). The most diagnostic and noticeable characteristics for both species are summarized in Box 1.
d’Huart & Grubb (2001) compiled a map depicting the geographical range of both the common warthog and the desert warthog in the Horn of Africa (Er-itrea, Djibouti, Somalia, Ethiopia,
Uganda and Kenya). Their find-ings were summarized in 2002 by Boy in Swara 25—1: 20-21. The authors found the desert warthog to occur from Puntland (northern Somalia) southwestward through Somalia and southeastern Ethio-pia to central and eastern Kenya. The only sites shown on the map for Kenya were Moyale, El Wak, Merelle (‘midway between Arch-er’s Post and Mt Marsabit’), and Mkokoni (in the Kiunga Ma-rine Reserve). The southernmost
How to most readily differentiate the desert warthog from the common warthog in the field (d’Huart & Grubb 2005). See Figures 1 and 2.
1. Warts—Adult desert warthog has a hook-shaped wart under the eye. Adult common warthog has a cone-shaped wart under the eye.
2. Ears—Ear tips of desert warthog are bent backwards. Ear tips of common warthog are erect.
3. Suborbital area—Suborbital area (ie, under the eye) appears swollen in adult desert warthog. In adult common warthog this swelling is absent.
4. Head—Head of desert warthog is ‘egg-shaped’ when viewed from the front. Head of common warthog is slightly ‘diabolo-shaped’ when viewed from the front.
A new pig for
Figure 1. Adult male common warthog Phacochoerus africanus on the open plains of the Laikipia Plateau, central Kenya. Note the cone shaped warts, pointed ears, and the diabolo-
shaped head. Photograph by T. M. Butynski & Y. A. de Jong.
Figure 2. Adult male desert warthog Phacochoerus aethiopicus in medium dense shrub in Tsavo West National Park, southeast Kenya. Note the flipped-back ears, hooked warts, the
broad, egg-shaped head, and the swollen suborbital area (area under the eyes). Photograph by T. M. Butynski & Y. A. de Jong.

51SWARA July – September 2008
record for Kenya was from the Kiunga Marine Reserve, about 60 kilometres northeast of Lamu. All records were from north of the Ewaso Ng’iro River and east of the Tana River. They speculated that the two species would be found to overlap in some parts of their range, but found no evidence for this; their data show the shortest distance between common warth-og and desert warthog records as 25 kilometres (in Puntland).
During a primate survey in 2005, TMB and YDJ encountered two solitary desert warthogs, 15 and 80 kilometres southwest of Garissa respectively, in medium dense acacia bushland. These en-counters are important, as they are the first records of desert warthog west of the Tana River. Two ques-tions remained, however. Are the desert warthog and common
warthog sympatric? How far south does the desert warthog occur?
On 7 June 2007, JC, JF and SBC traversed Tsavo East Na-tional Park. Briefed a few days earlier about desert warthog char-acteristics by TMB and YDJ, they travelled through the high-density tourism areas south of the Voi River, far to the southwest of the nearest confirmed desert warthog locations. Late that afternoon they were startled to find two adult desert warthogs in low scrub on the edge of the Dika Plains, just 13 kilometres north of Buchuma Gate, and managed to take a few photographs before the warthogs turned tail. They had just recorded a new large mammal species for Tsavo East National Park, and a major range extension of over 300 kilometres for the desert warthog!
The next day they returned to the area and located four more desert warthogs only nine kilo-metres from the Kenya Wildlife Service headquarters at Voi; one adult and three subadults stood their ground long enough for posi-tive identification to be made and
for more photographs to be taken. What struck JC, JF and SBC im-mediately during these encounters was the overall shape of the ani-mals’ heads, the hooked warts, and the flipped-back tip of the ears (see Box 1), all of which are diagnos-tic, easily-seen field characters of the desert warthog. During this trip, no common warthogs were seen. All photographs were sent to experts for confirmation. Some of these, together with some of our other warthog photographs, are available on a digital map at www.tinyurl.com/warthogMap.
In July 2007, JC returned briefly to Tsavo East National Park for two days. He observed two
sounders of desert warthogs—each of four animals—in open shrub-land north and south of the Voi River in the vicinity of Satao Safari Camp. Once again, no common warthogs were seen.
In October 2007, JC visited Tsavo West National Park for two days, where desert warthogs were found to be fairly common around Severin Safari Camp and Kitani
Figure 3. Desert warthog
Phacochoerus aethiopicus and
common warthog Phacochoerus
africanus localities in Tsavo East
National Park and Tsavo West
National Park, southeast Kenya.
FINDINGS

52 SWARA July – September 2008
Safari Lodge, just to the north of the Tsavo River. Three desert warthogs were seen feeding in an open area along a major tourism circuit, permitting lengthy obser-vation. One of these animals was a subadult male. Subadult desert warthogs, like adults, have ears that are flipped-back at the tips, and broader-based genal warts, al-lowing them to be differentiated in the field from common warthogs despite them lacking the hooked warts of adult male desert warth-ogs. During this visit JC also found common warthogs in Tsavo West National Park only a few hundred metres from where he observed desert warthogs.
In April, 2008, TMB and YDJ visited Tsavo West National Park for three days, and encountered common warthogs around Kilagu-ni Serena Safari Lodge, Komboyo Camp, Kitani Safari Lodge and Severin Safari Camp (including
areas where desert warthogs were seen during the earlier trip by JC). About six kilometres of Finch Hat-ton’s Camp, in low bush on the edge of riverine forest, one group of desert warthogs (one adult male and three subadults) was encoun-tered only 150 metres from six common warthogs. This sight-ing is not only the farthest range extension for desert warthog (390 kilometres) from Mikoni, the most southern record of d’Huart & Grubb, but it is also the first evidence that the two species of warthogs are sympatric.
Finding desert warthog in Tsavo, and finding them sympatric with common warthog, is excit-ing news for the species and for Kenya. Tsavo West National Park and Tsavo East National Park can add one more species to their list of large mammals. What further surprises can we expect from this long-overlooked pig?
Have you seen warthogs in Africa?
To better understand the distribution of Africa’s warthogs, we would like to know if you have seen desert warthogs or common warthogs in Eritrea, Ethiopia, Djibouti, Somalia, Uganda, Kenya or Tanzania. Details and/or a photograph of your record would be highly appreciated.
The following information is most important
date of sighting •species •name of nearest village or town •latitude and longitude •elevation •habitat•
Please send your information and/orphotograph to: Yvonne de Jong PO Box 149 Nanyuki 10400, Kenya [email protected] Thank you! Your help is much appreciated!
ReferencesBoy, G. 2002. The whole hog. •Swara 25 (1): 20-21.D’Huart, J. & Grubb, P. 2001. Dis-•tribution of the common warthog (Phacochoerus africanus) and the desert warthog (Phacochoerus aethiopicus). African Journal of Ecology 39: 156-169.Grubb, P. 1993. The Afrotropical •suids Phacochoerus, Hylochoerus, and Potamochoerus: taxonomy and description. In: Pigs, Peccaries and Hippos. Status Survey and Conservation Action Plan (ed. W. I. R. Oliver). IUCN/SSC, Gland, Switzerland iucn.org/themes/ssc/sgs/pphsg/•Contents.htm D’Huart, J. & Grubb, P. 2005. • A photographic guide to the differenc-es between the common warthog (Phacochoerus africanus) and the desert warthog (Ph. aethiopicus). Suiform Soundings 5 (2): 4-8iucn.org/themes/ssc/sgs/pphsg/•Suiform%20soundings/Newslet-ter%205(2)pdf
The East African Wild Life Society’s
2009 CALENDARS
January 2009Su Mo Tu We � Fr Sa 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31FEBRUARY 2009Su Mo Tu We � Fr Sa 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28
DECEMBER 2008Su Mo Tu We � Fr Sa 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31
Flamingo at sunset in Lake Nakuru National Park
TZ
UG
March 2009Su Mo Tu We � Fr Sa
1 2 3 4 5 6 7 8 9 10 11 12 13 14
15 16 17 18 19 20 21
22 23 24 25 26 27 28
29 30 31FEBRUARY 2009Su Mo Tu We � Fr Sa
1 2 3 4 5 6 7
8 9 10 11 12 13 14
15 16 17 18 19 20 21
22 23 24 25 26 27 28
APRIL 2009Su Mo Tu We � Fr Sa
1 2 3 4
5 6 7 8 9 10 11
12 13 14 15 16 17 18
19 20 21 22 23 24 25
26 27 28 29 30
Elephants in the Mara headed for a sun downer of their own
UG
SUNDAY
FEBRUARY 2009MONDAY TUESDAY WEDNESDAY THURSDAY FRIDAY SATURDAY
1 2 3 4 5 6 7
8 9 10 11 12 13 14
15 16 17 18 19 20 21
22 23 24 25 26 27 28
MARCH 2009Su Mo Tu We Th Fr Sa 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31
These Giraffe in the magnificent evening Mara light
SUNDAY
APRIL 2009MONDAY TUESDAY WEDNESDAY THURSDAY FRIDAY SATURDAY
1 2 3 4
5 6 7 8 9 10 11
12 13 14 15 16 17 18
19 20 21 22 23 24 25
26 27TZ 28 29 30
MAY 2009Su Mo Tu We Th Fr Sa
31 1 2
3 4 5 6 7 8 9
10 11 12 13 14 15 16
17 18 19 20 21 22 23
24 25 26 27 28 29 30
Zebra in flight on the Mara Plains
Good Friday
Easter Monday
Member Retail Prices
Ksh 300US$ 5.00UK£ 3.0€ 4.00
Non-Member Retail Prices
Ksh 350US$ 5.80UK£ 3.20€ 4.60
Member
Retail Prices
Ksh 250
US$ 5.00
UK£ 2.50
€ 3.20
Non-Member
Retail Prices
Ksh 270
US$ 5.80
UK£ 3.00
€ 3.50
Extra fee for
Postage
-
US$ 3.40
UK£ 2.00
€ 2.70
Extra fee for Postage
-US$ 6.30UK£ 3.40€ 4.90
Wall Calendar - retailDesk Calendar - retail
OVERPRINT AREA
OVERPRINT AREA

SUIFORM SOUNDINGS VOL 8(2) PAGE 4
Desert warthog Phacochoerus aethiopicus found in Tsavo East �ational
Park and Tsavo West �ational Park, southern Kenya Yvonne A. de Jong¹, James Culverwell² and Thomas M. Butynski³
¹P.O. Box 149, 10400 6anyuki, Kenya, [email protected]
²8 Manga Gardens, Kitisuru Rd, 6airobi, Kenya, [email protected]
³P.O. Box 149, 10400 6anyuki, Kenya, [email protected]
Both species of warthog, the common warthog Phacochoerus africanus and the desert warthog Phacochoerus aethiopicus, occur in Kenya. The desert warthog may be Africa’s least known non-forest large mammal as its distribution is poorly understood and it has never been the focus of an ecological or behavioural study. None of the earlier books and field guides on the mammals of eastern Africa mention the desert warthog (e.g., Dorst & Dandelot 1969, Haltenorth & Diller 1977, Kingdon 1979), and no game laws recognise this taxon (I. Parker, pers. comm.). Kingdon (1997) is the first major work to recognize the desert warthog as a full species and to bring this species to the attention of a large audience. The preliminary distribution map for the desert warthog compiled by d’Huart & Grubb (2001) presents only four localities for Kenya. They show the southern-most locality as Mkokoni, 60 km northeast of Lamu Island (north coast of Kenya). d’Huart & Grubb found no evidence for desert warthog south of the Ewaso Ng’iro River in central Kenya or west of the Tana River in eastern Kenya. They questioned whether the common warthog and the desert warthog might be sympatric at some sites. d’Huart & Grubb (2005) produced a photographic guide that highlights the diagnostic differences be-tween the common warthog and the desert warthog. Some of the main diagnostic phenotypic characters used to identify the two species of warthog in the field are as follows: common warthogs have pointed ear tips, cone-shaped genal warts, a ‘diabolo-shaped’ head (when viewed from the front), and the suborbital areas are not swollen (fig. 1); desert warthogs have ear tips that are lax and flipped back, hook-shaped genal warts, an ‘egg-shaped’ head (when viewed from the front), and swollen suborbital areas (fig. 2) In 2005, we started to opportunistically collect distribution data for both species of warthog in Kenya. TMB and YdJ found desert warthogs 15 km and 80 km west of Garissa town in 2005 when they encoun-tered two solitary individuals in medium-dense Acacia bushland during a primate survey. These are the first records west of the Tana River and extend the geographic range to ca. 265 km northwest of Mkok-oni, the nearest locality mentioned by d’Huart & Grubb (2001). In 2007, JC, J. Feely, and S. Bell-Cross visited Tsavo East National Park south of the Voi River. Al-though they encountered no common warthogs during this trip, they did observe two sounders of desert warthogs in low bush on the edge of the Dika Plains, ca 13 km north and northwest of Buchuma Gate.
Papers and
communications

SUIFORM SOUNDINGS VOL 8(2) PAGE 5
Photographs were taken and sent to experts for confirmation. Some of these photographs, together with some of our other warthog photographs, are available for viewing on an online digital map at: http://picasaweb.google.com/wildsolutions/WarthogSightingsInKenya/photo?authkey=WigT2oFY78k#map. These observations considerably extend the known geographical range for the desert warthog (ca. 310 km south from the nearest Garissa sighting and ca. 320 km southwest from Mkokoni) (Fig. 3). JC made two further visits to the Tsavos in 2007. He found both species of warthog in Tsavo West Na-tional Park and desert warthog north of the Voi River in Tsavo East National Park. In 2008, TMB and YdJ visited Tsavo West National Park and observed several sounders both of common warthogs and desert warthogs. In the northwest of the Park, in low bush on the edge of riverine forest, they found a sounder of six common warthogs only 150 m from a sounder of four desert warthogs. This locality represents not only the south western-most site in the range for desert warthog (ca. 390 km from Mkokoni, the southern-most point of d’Huart & Grubb 2001), but it also provided the first evidence that common warthog and desert warthog are at least narrowly sympatric over this part of their geographic ranges.
Figure 1: Adult male common warthog Phacochoerus africanus on the plains of the Lai-kipia Plateau, central Kenya. Note the pointed ears, the cone-shaped warts, the ‘diabolo-shaped’ head, and the lack of swelling of the suborbital area.
Figure 2: Adult male desert warthog Phacochoe-rus aethiopicus in medium-dense shrub in Tsavo West National Park, southern Kenya. Note the flipped-back ear tips, the hooked warts, the broad, ‘egg-shaped’, head, and the swollen suborbital area

SUIFORM SOUNDINGS VOL 8(2) PAGE 6
Although we have yet to find the common warthog in Tsavo East National Park, it would be surprising if this species were not present there. If not present, however, the common warthog would need to be de-leted from the list of large mammals known for Tsavo East National Park. As concerns the desert wart-hog, Tsavo East National Park and Tsavo West National Park can now add one more species to their al-ready impressive list of large mammals.
Figure 3: Locations of common warthog Phacochoerus africanus and desert warthog Phacochoerus aethiopicus sightings in Tsavo East National Park and Tsavo West National Park, southern Kenya. References
d’Huart J and Grubb P. 2001. Distribution of the common warthog (Phacochoerus africanus) and the de-sert warthog (Phacochoerus aethiopicus). African Journal of Ecology 39: 156-169.
d’Huart J & Grubb P. 2005. A photographic guide to the differences between the common warthog (Phacochoerus africanus) and the desert warthog (Ph. aethiopicus). Suiform Soundings 5: 4-8. < http://iucn.org/themes/ssc/sgs/pphsg/Suiform%20soundings/Newsletter%205(2)pdf >
Dorst J & Dandelot P. 1969. A Field Guide to the Larger Mammals of Africa. Houghton Mifflin Co., Boston, USA.
Haltenorth T & Diller H. 1977. A Field Guide to the Mammals of Africa including Madagascar. Collins, London, UK.
Kingdon J. 1979. East African Mammals. An Atlas of Evolution in Africa. Vol.III B. University of Chi-cago Press, Chicago, USA.
Kingdon J. 1997. The Kingdon Field Guide to African mammals. Academic Press, New York, USA.





























