Embed Size (px)
Citation preview

Presynaptic calcium-channel currents in normal and cspmutant Drosophila peptidergic terminals
Miguel Morales,* Alberto FerruÂs and Manuel MartõÂnez-PadroÂnInstituto Cajal (CSIC) Ave. Doctor Arce 37, Madrid 28002, Spain
Keywords: calcium channels, cysteine string protein, Drosophila melanogaster, neuropeptide, release, synapse
Abstract
The study of regulated vesicle exocytosis, which underlies neurotransmitter and neuropeptide release, has bene®ted from aconvergence of several independent approaches. These include the use of genetically tractable organisms and model preparationsthat allow a direct characterization of presynaptic ionic currents. Aiming for a comprehensive analysis of release, we had alreadydeveloped a Drosophila preparation in which electrophysiological recordings from peptidergic terminals are feasible. Here, we reporton the characterization of the Ca2+-channel currents present in these terminals. With Ba2+ as the charge carrier, the presynapticmembrane expresses a current type with high-activation threshold and little inactivation. This current is blocked by verapamil anddiltiazem at micromolar concentrations, it is relatively insensitive to nifedipine and completely resistant to non-L-type Ca2+-channelantagonists. As a comparison, we also analysed the pharmacology of high-threshold Ba+2 currents on muscle ®bres. A high-activation threshold Ca2+-channel current is also present in muscle ®bres, albeit with a distinct pharmacological pro®le. Thus,peptidergic terminals and muscle ®bres exhibit different subtypes of voltage-gated Ca2+ channels. The putative role of cysteine stringprotein (CSP) as a neuronal Ca2+-channel modulator was tested by examining the peptidergic presynaptic current in csp nullmutants. We show that CSP is expressed in peptidergic boutons and abolished in the mutant. Direct recordings, under conditions thatinhibit calcium in¯ux into glutamatergic terminals, show that Ca2+-currents in peptidergic csp terminals are entirely normal. This resultindicates that CSP is not a generic Ca2+-channel modulator and it might perform different functions in fast versus slow forms ofrelease.
Introduction
Voltage-gated Ca2+ channels have been identi®ed as a ubiquitous
component of animal cells of virtually all phyla. They play a critical
role in depolarization±secretion coupling and, hence, their character-
ization is a requirement in the study of synaptic function. The
application of electrophysiological techniques to study presynaptic
currents has been hindered by the extremely small size of nerve
endings and, with some exceptions (e.g. LlinaÂs et al., 1981; Lemos &
Nowycky, 1989; Stanley & Goping, 1991), the nature of presynaptic
calcium channels has been inferred from the pharmacological
sensitivity of neurotransmitter release to selective antagonists. Based
on biophysical and pharmacological criteria, vertebrate calcium
channels have been classed into types T, N, L, P, Q and R. Thus, L-
type channels are highly sensitive to inorganic compounds, including
phenylalkylamines (PA), benzothiazepines (BTZ) and dihydropyr-
idines (DHP), while the peptide toxins w-Conotoxin GVIA and w-
Agatoxin IVA appear rather selective for channel types N and P,
respectively.
Transmitter release is triggered by a sudden increase in Ca2+
concentration restricted to the vicinity of the release machinery. Slow
exocytosis, as it occurs in neuropeptide release, has many features in
common with hormone release from endocrine cells. High-frequency
stimulation is usually required for optimal peptide release (Iverfeldt
et al., 1989), presumably due to the need for a sustained increase in
bulk calcium concentration at presynaptic terminals (Neher &
Zucker, 1993). It has been suggested that Ca2+ entry via strategically
located N- and/or P-type channels dominates the release of small
clear synaptic vesicles that contain `fast' neurotransmitters, whereas
L-type channels are often linked to the release of neuropeptide-
containing large dense-core vesicles (Dunlap et al., 1995). Larval
muscle ®bres in Drosophila are contacted by several types of synaptic
terminals (Atwood et al., 1993; Jia et al., 1993). Among these, type I
boutons contain small clear vesicles and mediate glutamatergic
release (Jan & Jan, 1976; Johansen et al., 1989), whereas peptidergic
type III boutons contain large-dense core vesicles and exhibit insulin-
like and proctolin immunoreactivity (Anderson et al., 1988; Gorczyca
et al., 1993).
As triggers of synaptic transmission, presynaptic calcium channels
interact with synaptic proteins of the docking-fusion machinery, such
as syntaxin and SNAP-25 (Wiser et al., 1996; Rettig et al., 1997), and
this interaction might depend on the channel type (Bezprozvanny
et al., 1995). Cysteine string proteins (CSP) are a major component of
all synaptic vesicle membranes (Mastrogiacomo et al., 1994; Pupier
et al., 1997). They have been proposed to modulate the activity of
presynaptic calcium channels (Gundersen & Umbach, 1992).
Drosophila csp mutants exhibit a temperature-dependent loss of
nerve-impulse evoked transmission, this defect being attributed to a
failure in depolarization-secretion coupling (Umbach et al., 1994).
Indirect evidence suggested that Ca2+ in¯ux into presynaptic
terminals could be impaired in csp mutants (Umbach & Gundersen,
Correspondence: Dr A. FerruÂs, as above. E-mail: [email protected]
*Present address: UCSD Department of Biology, 9500 Gillman Drive, LaJolla, CA 92093±0346, USA
Received 17 November 1998, accepted 13 January 1999
European Journal of Neuroscience, Vol. 11, pp. 1818±1826, 1999 Ó European Neuroscience Association

1997; Ranjan et al., 1998). This hypothesis has received support from
Ca2+ imaging experiments on csp mutant terminals (Umbach et al.,
1998). We have developed a preparation that allows patch-clamp
recordings from type III terminals in vivo (MartõÂnez-PadroÂn & FerruÂs,
1997; 1998). Here, we combine this technical achievement with the
genetic capabilities of Drosophila to characterize presynaptic Ca2+-
channel currents in these terminals, as well as to study their proposed
modulation by CSP. We show that this protein, readily expressed in
type III boutons, does not appear to be a generic Ca2+ channel
modulator.
Materials and methods
Mutants and experimental preparation
The ecdysone (ecd1) allele is a temperature sensitive recessive lethal
(Garner et al., 1977). When kept at 30 °C, ecd1 mutant larvae fail to
pupate and develop enlarged type III synaptic boutons. Although
electrophysiological recordings are feasible in wild type strains and
csp mutants, they are much easier in ecd1 mutants, and we have
systematically used this mutant to characterize presynaptic type III
Ca2+-currents, and as controls for comparison with csp mutants at
restrictive temperatures. Nevertheless, we have obtained a limited
number of recordings from the Oregon-R wild type strain in order to
ensure that ecd1 currents are very much like wild type. The
Drosophila cspR1 allele represents a deletion of the entire csp gene.
cspR1 is a temperature-sensitive paralytic mutant. About 4% of the
animals survive to adulthood at 25 °C, but none at 29 °C (Zinsmaier
et al., 1994); this temperature-sensitive paralysis is caused by a
reversible failure of evoked synaptic transmission (Umbach et al.,
1994; Zinsmaier et al., 1994).
The experimental preparation to record ionic currents from type III
synaptic boutons has been previously described (MartõÂnez-PadroÂn &
FerruÂs, 1997, 1998). Brie¯y, third instar larvae were pinned down
onto a Sylgard-coated experimental chamber, dissected open along
the dorsal midline in a solution containing (in mM): NaCl, 100; KCl,
5; MgCl2, 20; HEPES, 5; and Sucrose, 115 (pH 7.3), and digested for
8±10 min in the same solution with 200 U/mL collagenase (Type IA;
Sigma, St Louis, USA). Type III terminals were unambiguously
identi®ed using a 3 40 Nomarski water immersion objective (Nikon).
Unless otherwise indicated, all electrophysiological recordings were
performed at room temperature (21±23 °C) on type III synaptic
terminals innervating ventro-longitudinal muscle ®bre 12 (segments
A2±A6).
Electrophysiological recordings
The perforated patch-clamp technique (Horn & Marty, 1988) was
used to record whole-terminal currents as previously described
(MartõÂnez-PadroÂn & FerruÂs, 1997). Brie¯y, high resistance pipettes
(10±20 MW were pulled from thin-wall borosilicate glass (World
Precision Instruments). The pipette tip was ®lled with an internal
solution containing (in mM): CsCl, 140; MgCl2, 4; HEPES, 10;
EGTA, 5±10 (pH 7.3), and then back-®lled with the same solution to
which a ®nal concentration of 200 mg/mL Nystatin (Sigma) was
added. Whole-terminal currents were recorded with a patch-clamp
ampli®er (Axopatch 1D), ®ltered at 2 kHz, digitized and stored in a
computer for further analysis using PClamp Software (Axon
instruments, CA, USA). Upon seal formation and membrane
perforation, the pipette tip resistance increases 3±4 times, which
represents a voltage error of < 5 mV for a current magnitude of
100 pA (see MartõÂnez-PadroÂn & FerruÂs, 1997) and » 1 mV with 80%
series resistance compensation. Currents were leak-corrected by
subtracting the current produced by a pulse protocol of the same
magnitude but opposite polarity. Pooled data in the text are presented
as mean 6 SEM.
In most experiments (see below), the bath solution used to record
calcium-channel currents from synaptic terminals contained (in mM):
NaCl, 100; KCl, 5; BaCl2, 5; MgCl2, 20; HEPES, 5; Sucrose, 115, and
5mM TTX (pH 7.3). Because Ba2+ does not support muscle contrac-
tion, it yields more stable current recordings. Also, Ba2+ acts as a K+
channel blocker and helps to reduce contaminating K+ currents, which
are further blocked by the presence of Cs+ in the pipette solution.
Muscle calcium currents were recorded from muscle ®bre 6 (segments
A2±A3), using an Axoclamp-2 A ampli®er in the two-electrode
voltage-clamp mode. Short-shank micropipettes were pulled to a ®nal
resistance of 10±20 MW when ®lled with a 1 m KCl solution. The
external solution for muscle current recordings was the same as for
synaptic recordings, except that it included 100 mM quinidine and
10 mM 3,4-diaminopyridine to reduce contaminating K+ currents. In
those experiments addressing the function of CSP, the temperature of
the bath was controlled by a junction Peltier (HC10A controller) with a
HC10AD headstage (Dagan Corporation, Minneapolis, USA).
Space-clamp considerations
As type III terminals are connected by ®ne axonal processes, we were
concerned that current generated at poorly space-clamped adjacent
terminals could signi®cantly contaminate our recordings. In order to
assess the severity of this problem we performed simultaneous
recordings from two adjacent boutons. The results from one
successful experiment are shown (Fig. 1). The lower traces corre-
spond to Ba2+-inward currents recorded in one terminal (A) under
voltage-clamp conditions, in response to depolarizing voltage steps
from ±50 mV; while the upper traces represent the membrane
potential change recorded in an adjacent terminal (B) » 12 mm away.
These currents show a particularly poor clamp, and they are typical of
those recorded immediately after seal formation, but they have been
selected to illustrate the electrical relationship between the two
terminals. Voltage steps below ±20 mV caused a small, progressive
depolarization of terminal B, and no detectable inward current at
terminal A. However, voltage commands to ±20 mV and above,
trigger a much larger depolarization in B; its temporal course is
closely matched by the current pro®le at terminal A, and we attribute
it to Ba2+ in¯ux through Ca2+ channels in terminal B. Thus, Ba2+-
currents recorded at type III boutons under these conditions, are
signi®cantly contaminated by Ba2+ in¯ux at nearby terminals. As a
compromise solution, we sought to restrict activation of barium
currents to a single terminal by using focal application of 10 mM Ba2+
(or Ca2+), in extracellular solution, delivered from an unpolished
patch pipette located in close proximity to the bouton. Although
records using this procedure were often contaminated by current from
poorly clamped nearby boutons, as suggested by the sudden, all-or-
none activation of the current (Fig. 2A) and a progressive reduction in
latency to peak current with increasing depolarization, a number of
successful cases showed inward currents with progressive activation
and constant time to peak, suggestive of little or no contamination
(Fig. 2B). This latter set of data was used to derive the biophysical
parameters of the synaptic Ca2+-channel current.
Drugs
Verapamil hydrochloride and diltiazem hydrochloride (Roig Farma,
Barcelona, Spain) were dissolved in saline solution. Nifedipine
(Sigma) was dissolved in ethanol at a stock concentration of 10 mM.
Synthetic Funnel Web spider toxin was a gift from Dr R. LlinaÂs (New
York University, USA). All peptide toxins (Alomone Lab.,
Drosophila presynaptic calcium currents 1819
Ó 1999 European Neuroscience Association, European Journal of Neuroscience, 11, 1818±1826

Jerusalem, Israel) were dissolved in stock solution containing (in
mM): NaCl, 100; Tris, 10; EDTA, 1, and 0.1% BSA (pH 7.5), and
kept stored at ±20 °C. Bath solution exchanges were made by ¯ushing
the recording chamber with a solution volume 10 times larger at a
perfusion rate of 4 mL/min. The toxin peptides were added directly to
the recording chamber to obtain the desired ®nal concentration.
Immunolabelling
Third instar larvae were ®xed for 30 min in freshly made 4%
paraformaldehyde in Ca2+-free PBS containing 5 mM EGTA. Speci-
mens were incubated in mouse monoclonal anti-CSP antibody
(mAb49, 1 : 200, Buchner et al., 1988) overnight at 4 °C in PBT
(0.1% Triton in PBS), thoroughly washed in PBT and then incubated
in secondary antimouse antibody conjugated to Cy2 for 1 h at room
temperature. Preparations were mounted on PBS:glycerol (1 : 1) and
viewed under confocal microscopy.
Results
Calcium-channel currents in type III synaptic terminals
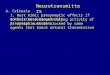
A typical family of Ba2+ currents, activated from a holding potential
of ±50 mV in 10 mV steps, where lack of space-clamp control
becomes evident, is depicted in Fig. 2A (left panel). In this particular
case, 10 mM Ba2+ solution was applied from an unpolished patch
pipette placed close to the bouton, but similar currents are obtained if
Ba2+ is present in the bath instead. Membrane steps below ±20 mV
did not elicit any inward current. In some cases, the current became
active in an all-or-none fashion at » ±15 to ±10 mV, whereas in other
cases some gradation in the activation could be observed, presumably
depending on the degree of coupling between boutons. On average,
the macroscopic current thus recorded reached a maximum peak at
» 0 mV, with little inactivation throughout the test pulses. Bath
application of 200 mM Cd2+caused a complete blockade of the
macroscopic inward current (not shown), indicating that it is, in all
likelihood, due to Ba2+ in¯ux through Ca2+ channels. With further
membrane depolarization, the current amplitude declines as the
electrochemical driving force for Ba2+ diminishes. A limited number
of Ca2+ current recordings (n = 3) was obtained by delivering Ca2+
(10 mM) from a nearby pipette (Fig. 2A, right traces). The currents
thus recorded, showed two important features when compared with
those recorded in Ba2+: (i) the current exhibited a more inactivating
pro®le; and (ii) current activation also occurred in an all-or-none
fashion, but at a more positive potential (0 mV) than with Ba2+
(Fig. 2C).
Using the procedure of focal delivery of divalent cations, and
restricting the voltage range to relatively lower values, we could
record a limited number of Ba2+ (n = 3) and Ca2+ (n = 1) currents
showing no contamination, as judged from their progressive
activation and constant time to peak (Fig. 2B). From a holding
potential of ±50 mV, and using Ba2+as the charge carrier, depolariz-
ing voltage steps above ±30 mV produced a relatively noninactivating
terminal inward current, which peaked at » 10 mV (Fig. 2B, left
traces). With extracellular Ca2+ (Fig. 2B, right traces), however, the
current showed considerable time-dependent inactivation. Also, the
threshold for current activation was 15±20 mV more depolarized than
in Ba2+. Membrane depolarization to the same potentials, before
delivery of the divalent cation from the pipette, produced only a small
outward leak current. This small current which was used for leak
subtraction, further indicates that the inward current is probably due
to the in¯ux of the cation used. The corresponding current±voltage
(I±V) relationships are shown in Fig. 2C.
We have explored the possibility that synaptic terminals could
contain a Ca2+ current component that would undergo steady-state
inactivation at the resting potential. Figure 2D shows superimposed
current traces activated by a test potential to 10 mV from a variety of
holding potentials (between ±100 mV and ±30 mV). Using this
voltage protocol, we observed no differences in either peak amplitude
or time course of the Ba2+ current evoked by the test pulse.
Notwithstanding the space-clamp problems, these results suggest that
no low-voltage activated component is present in type III terminals.
In conclusion, the single voltage-dependent Ca2+-channel current
detected exhibits a high activation threshold (above ±30 mV) and may
undergo Ca2+-dependent inactivation.
Pharmacology of synaptic and muscle Ca2+ currents
Accepting the fact that the Ca2+ channel current will be contaminated
by in¯ux at nearby terminals, these experiments were carried out
using 5 mM Ba2+ in the bath. Consequently, we aimed at a qualitative,
rather than a quantitative, pharmacological analysis. Presynaptic
Ba2+-currents, elicited from a holding potential of ±50 mV, were
reduced by verapamil in a concentration-dependent manner (Fig. 3A).
Bath application of 10 mM verapamil did not have any effect, whereas
50 and 100 mM reduced the current by 60% and 90%, respectively
(n = 3). These data indicate that 50% blockade would require » 30 mM
FIG. 1. Simultaneous recordings from two adjacent type III boutons. TerminalA (lower traces) was voltage-clamped at ±50 mV while terminal B (uppertraces) was recorded under current-clamp. Increasing depolarizing voltagecommands applied to terminal A in 10 mV steps are detected in terminal B,®rst as small depolarizations, which increase linearly, and then as much largerstepwise increases coincidental with the all-or-none activation of the inwardcurrents at terminal A. The time courses of terminal B membrane potentialsand terminal A currents are closely related. This is more obvious for the pulseto ±20 mV. In this particular case, the terminal B membrane potential at thebeginning of the pulse behaves very much the same as for morehyperpolarized pulses, re¯ecting electrotonic transmission from terminal A.However, later in the pulse, the membrane potential suddenly increases,presumably due to opening of terminal B Ca2+ channels, which is detected interminal A as a large, and also sudden, inward current.
1820 M. Morales et al.
Ó 1999 European Neuroscience Association, European Journal of Neuroscience, 11, 1818±1826
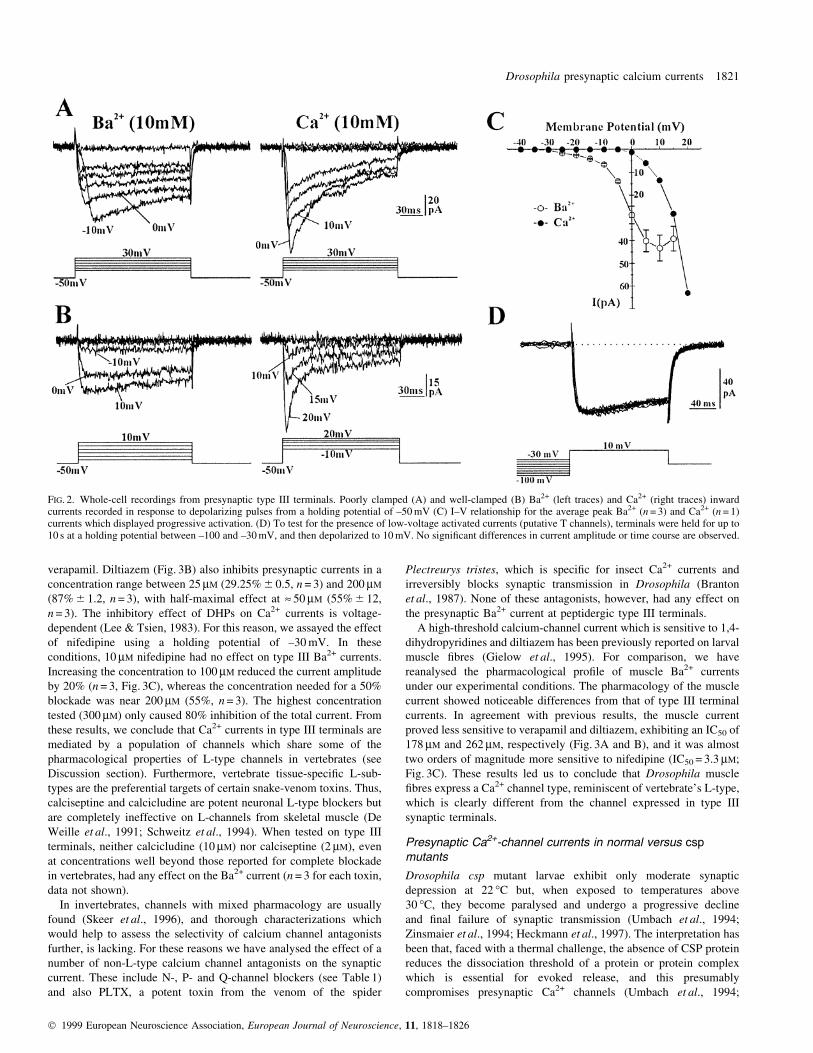
verapamil. Diltiazem (Fig. 3B) also inhibits presynaptic currents in a
concentration range between 25 mM (29.25% 6 0.5, n = 3) and 200 mM
(87% 6 1.2, n = 3), with half-maximal effect at » 50 mM (55% 6 12,
n = 3). The inhibitory effect of DHPs on Ca2+ currents is voltage-
dependent (Lee & Tsien, 1983). For this reason, we assayed the effect
of nifedipine using a holding potential of ±30 mV. In these
conditions, 10 mM nifedipine had no effect on type III Ba2+ currents.
Increasing the concentration to 100 mM reduced the current amplitude
by 20% (n = 3, Fig. 3C), whereas the concentration needed for a 50%
blockade was near 200 mM (55%, n = 3). The highest concentration
tested (300 mM) only caused 80% inhibition of the total current. From
these results, we conclude that Ca2+ currents in type III terminals are
mediated by a population of channels which share some of the
pharmacological properties of L-type channels in vertebrates (see
Discussion section). Furthermore, vertebrate tissue-speci®c L-sub-
types are the preferential targets of certain snake-venom toxins. Thus,
calciseptine and calcicludine are potent neuronal L-type blockers but
are completely ineffective on L-channels from skeletal muscle (De
Weille et al., 1991; Schweitz et al., 1994). When tested on type III
terminals, neither calcicludine (10 mM) nor calciseptine (2 mM), even
at concentrations well beyond those reported for complete blockade
in vertebrates, had any effect on the Ba2+ current (n = 3 for each toxin,
data not shown).
In invertebrates, channels with mixed pharmacology are usually
found (Skeer et al., 1996), and thorough characterizations which
would help to assess the selectivity of calcium channel antagonists
further, is lacking. For these reasons we have analysed the effect of a
number of non-L-type calcium channel antagonists on the synaptic
current. These include N-, P- and Q-channel blockers (see Table 1)
and also PLTX, a potent toxin from the venom of the spider
Plectreurys tristes, which is speci®c for insect Ca2+ currents and
irreversibly blocks synaptic transmission in Drosophila (Branton
et al., 1987). None of these antagonists, however, had any effect on
the presynaptic Ba2+ current at peptidergic type III terminals.
A high-threshold calcium-channel current which is sensitive to 1,4-
dihydropyridines and diltiazem has been previously reported on larval
muscle ®bres (Gielow et al., 1995). For comparison, we have
reanalysed the pharmacological pro®le of muscle Ba2+ currents
under our experimental conditions. The pharmacology of the muscle
current showed noticeable differences from that of type III terminal
currents. In agreement with previous results, the muscle current
proved less sensitive to verapamil and diltiazem, exhibiting an IC50 of
178 mM and 262 mM, respectively (Fig. 3A and B), and it was almost
two orders of magnitude more sensitive to nifedipine (IC50 = 3.3 mM;
Fig. 3C). These results led us to conclude that Drosophila muscle
®bres express a Ca2+ channel type, reminiscent of vertebrate's L-type,
which is clearly different from the channel expressed in type III
synaptic terminals.
Presynaptic Ca2+-channel currents in normal versus cspmutants
Drosophila csp mutant larvae exhibit only moderate synaptic
depression at 22 °C but, when exposed to temperatures above
30 °C, they become paralysed and undergo a progressive decline
and ®nal failure of synaptic transmission (Umbach et al., 1994;
Zinsmaier et al., 1994; Heckmann et al., 1997). The interpretation has
been that, faced with a thermal challenge, the absence of CSP protein
reduces the dissociation threshold of a protein or protein complex
which is essential for evoked release, and this presumably
compromises presynaptic Ca2+ channels (Umbach et al., 1994;
FIG. 2. Whole-cell recordings from presynaptic type III terminals. Poorly clamped (A) and well-clamped (B) Ba2+ (left traces) and Ca2+ (right traces) inwardcurrents recorded in response to depolarizing pulses from a holding potential of ±50 mV (C) I±V relationship for the average peak Ba2+ (n = 3) and Ca2+ (n = 1)currents which displayed progressive activation. (D) To test for the presence of low-voltage activated currents (putative T channels), terminals were held for up to10 s at a holding potential between ±100 and ±30 mV, and then depolarized to 10 mV. No signi®cant differences in current amplitude or time course are observed.
Drosophila presynaptic calcium currents 1821
Ó 1999 European Neuroscience Association, European Journal of Neuroscience, 11, 1818±1826
Umbach & Gundersen, 1997). In this respect, Umbach et al. (1998),
using calcium imaging, have recently shown that nerve-stimulated
Ca2+ in¯ux into glutamatergic synaptic boutons is strongly reduced in
csp mutants at restrictive temperature. CSP protein, however, has
been reported to associate with both small clear and large dense-core
vesicle membranes (Mastrogiacomo et al., 1994; Pupier et al., 1997).
The protein is also abundant in non-neuronal secretory tissues
(Koham et al., 1995), suggesting that it may be present in all vesicles
targeted for exocytosis.
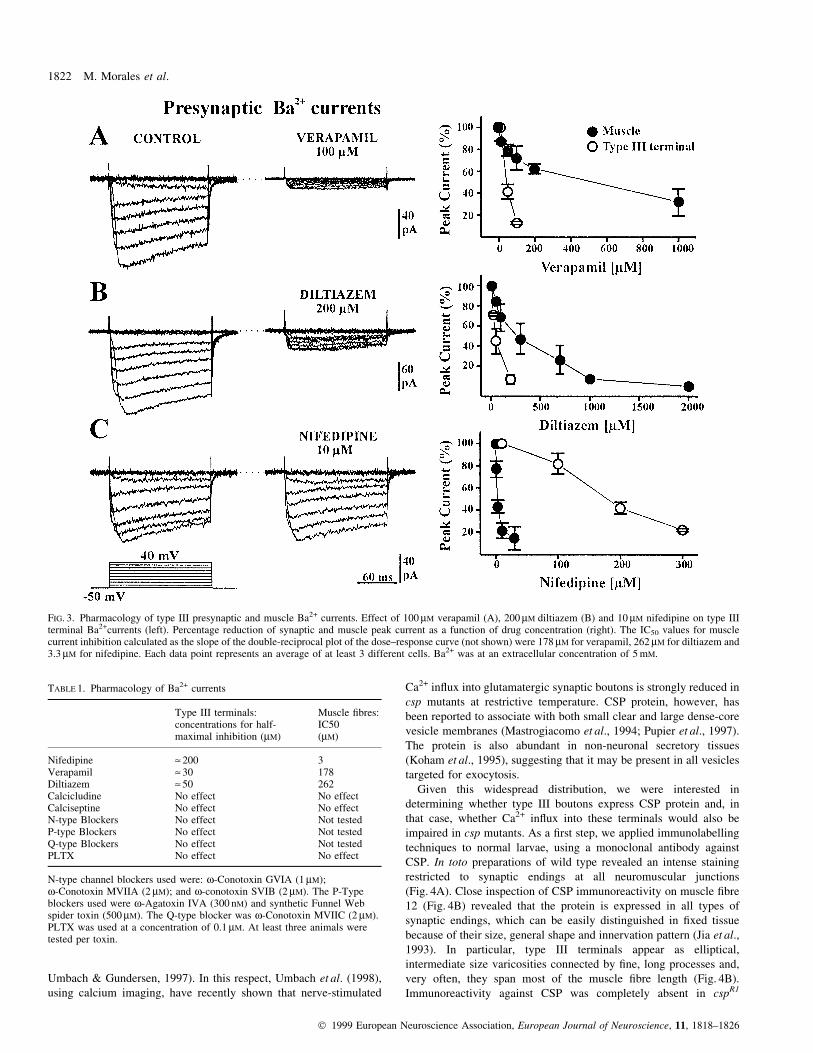
Given this widespread distribution, we were interested in
determining whether type III boutons express CSP protein and, in
that case, whether Ca2+ in¯ux into these terminals would also be
impaired in csp mutants. As a ®rst step, we applied immunolabelling
techniques to normal larvae, using a monoclonal antibody against
CSP. In toto preparations of wild type revealed an intense staining
restricted to synaptic endings at all neuromuscular junctions
(Fig. 4A). Close inspection of CSP immunoreactivity on muscle ®bre
12 (Fig. 4B) revealed that the protein is expressed in all types of
synaptic endings, which can be easily distinguished in ®xed tissue
because of their size, general shape and innervation pattern (Jia et al.,
1993). In particular, type III terminals appear as elliptical,
intermediate size varicosities connected by ®ne, long processes and,
very often, they span most of the muscle ®bre length (Fig. 4B).
Immunoreactivity against CSP was completely absent in cspR1
FIG. 3. Pharmacology of type III presynaptic and muscle Ba2+ currents. Effect of 100 mM verapamil (A), 200 mM diltiazem (B) and 10 mM nifedipine on type IIIterminal Ba2+currents (left). Percentage reduction of synaptic and muscle peak current as a function of drug concentration (right). The IC50 values for musclecurrent inhibition calculated as the slope of the double-reciprocal plot of the dose±response curve (not shown) were 178 mM for verapamil, 262 mM for diltiazem and3.3 mM for nifedipine. Each data point represents an average of at least 3 different cells. Ba2+ was at an extracellular concentration of 5 mM.
TABLE 1. Pharmacology of Ba2+ currents
Type III terminals: Muscle ®bres:concentrations for half- IC50maximal inhibition (mM) (mM)
Nifedipine » 200 3Verapamil » 30 178Diltiazem » 50 262Calcicludine No effect No effectCalciseptine No effect No effectN-type Blockers No effect Not testedP-type Blockers No effect Not testedQ-type Blockers No effect Not testedPLTX No effect No effect
N-type channel blockers used were: w-Conotoxin GVIA (1 mM);w-Conotoxin MVIIA (2 mM); and w-conotoxin SVIB (2 mM). The P-Typeblockers used were w-Agatoxin IVA (300 nM) and synthetic Funnel Webspider toxin (500 mM). The Q-type blocker was w-Conotoxin MVIIC (2 mM).PLTX was used at a concentration of 0.1 mM. At least three animals weretested per toxin.
1822 M. Morales et al.
Ó 1999 European Neuroscience Association, European Journal of Neuroscience, 11, 1818±1826
mutant larvae (Fig. 4C), both in type III and other terminals, thereby
con®rming that cspR1 is a null mutation for the csp gene.
As temperature-induced paralysis is the characteristic feature of
csp mutants, we tested our laboratory stock for temperature-
dependent paralysis. All animals (n = 30) within a group of csp third
instar larvae exposed to 32 °C, became paralysed within 18 min (not
shown); in like manner, a similar group exposed to 34 °C, became
completely paralysed within 12 min. Control groups of ecd1 larvae
run in parallel in each case, were not affected by these procedures. In
addition, all csp larvae recovered within 1 h upon returning to room
temperature.
Proposals about the role of CSP, suggest that the thermal paralysis
results from a corresponding thermal dependence of a Ca2+ current in
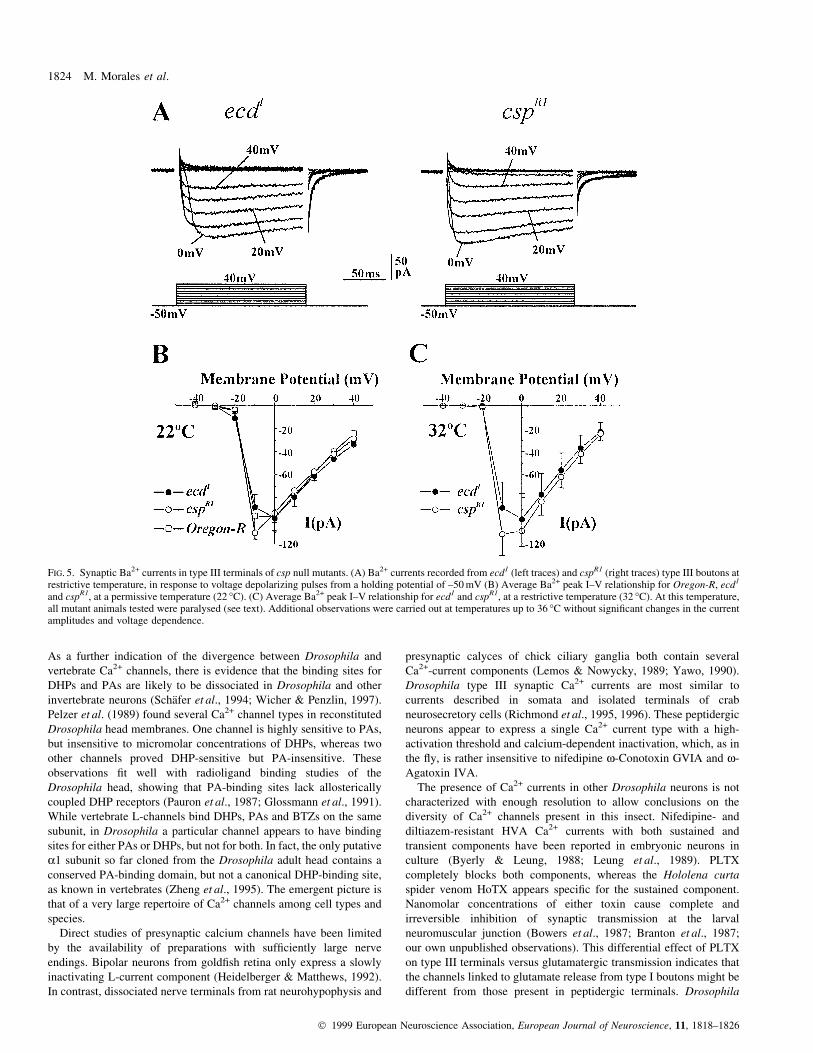
the mutant. We examined type III synaptic Ba2+ currents in cspR1
mutants and compared them with those in ecd1 and wild type as
controls. Figure 5B represents average Ba2+ current peak-voltage
relationships for ecd1, Oregon-R and cspR1 larvae at room
temperature (22 °C). No major differences appear between the three
genotypes. The peak current amplitude measured at 0 mV was
97.3 6 5.4 pA (n = 19) for ecd1, 96.6 6 11.9 (n = 9) for Oregon-R and
93.2 6 7.9 (n = 5) for cspR1 larvae, suggesting that ecd1 Ba2+ currents
are entirely similar to wild type and that lack of CSP protein per se,
does not affect this current at the type III terminals.
Further, increasing the temperature from 22 °C to 32 °C had no
major effect on the average current peak amplitude of either ecd1
(98.6 6 22.2 pA at 0 mV, n = 5) or cspR1 larvae (108.7 6 13.4 pA at
0 mV, n = 9) (Fig. 5A and C). That is, temperature-dependent
paralysis of csp mutant larvae does not correlate with any signi®cant
reduction of type III terminal Ba2+ currents. Considering that type III
boutons do express CSP protein, our results suggest that CSP must
play an alternative role in these terminals.
Discussion
This study represents the ®rst attempt to characterize directly
presynaptic Ca2+-channel currents in peptidergic terminals of
Drosophila. Poor space-clamping posed a major problem which
prevented us from doing a precise quantitative study. We tried to
restrict Ca2+-channel activation to a single bouton by applying
divalent cations locally. The procedure yielded a modest rate of
success, and a large number of recordings was required to obtain a
few well-clamped currents, as judged by their progressive activation
and constant latency to peak.
Type III terminals express a high-voltage activated (HVA) Ca2+-
channel current with no evidence of a low-voltage activated (T-type)
component. With Ba2+ as charge carrier, the current shows little
inactivation during the test pulse. The I±V plot shows an activation
threshold at » ±20 mV and a maximal current of » +10 mV. The limited
data we have obtained with Ca2+ perfusion showed a clearly
inactivating current pro®le which suggests that these channels may
undergo Ca2+-dependent inactivation. Also, it would appear that Ba2+
causes a leftward shift of the I±V relationship as has been observed in
other preparations (e.g. Branchaw et al., 1997). Pharmacological tools
have been used in many studies to distinguish between HVA channel
types. Synaptic Ba2+ currents were not sensitive to drugs or to toxins
considered highly speci®c for vertebrates Ca2+-channel types. The
Drosophila synaptic current was blocked by PAs and BTZs only,
compounds which are considered less selective for L-type channels.
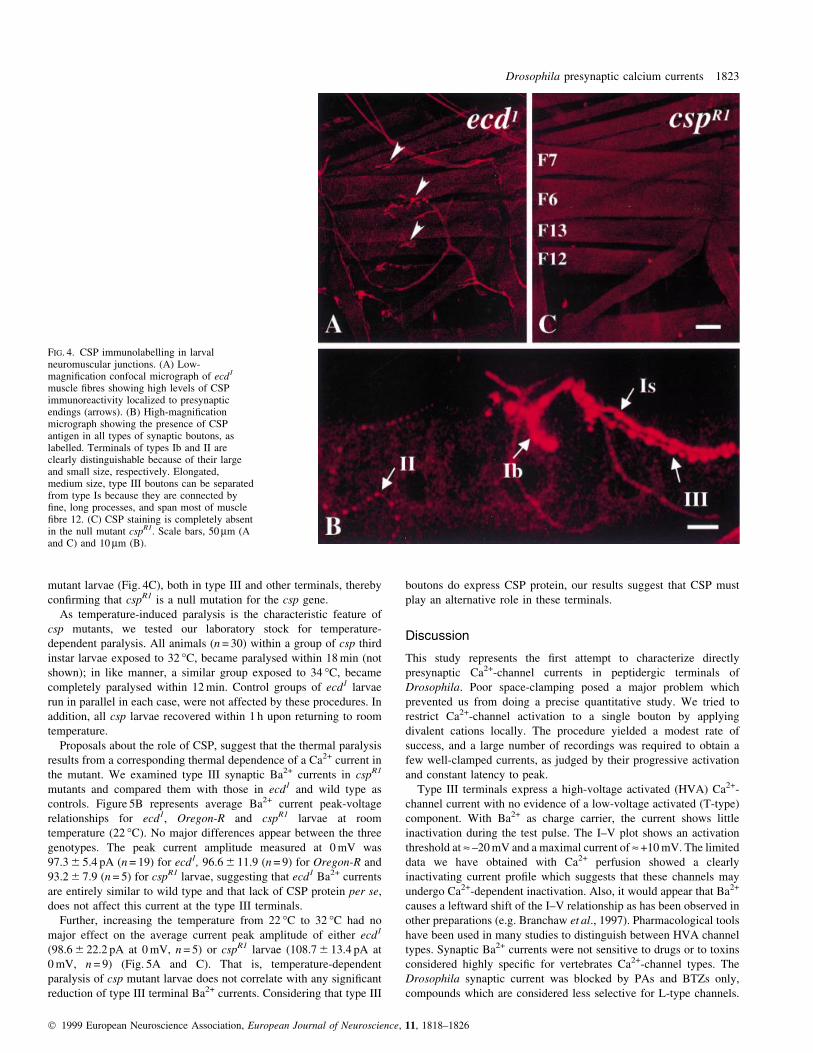
FIG. 4. CSP immunolabelling in larvalneuromuscular junctions. (A) Low-magni®cation confocal micrograph of ecd1
muscle ®bres showing high levels of CSPimmunoreactivity localized to presynapticendings (arrows). (B) High-magni®cationmicrograph showing the presence of CSPantigen in all types of synaptic boutons, aslabelled. Terminals of types Ib and II areclearly distinguishable because of their largeand small size, respectively. Elongated,medium size, type III boutons can be separatedfrom type Is because they are connected by®ne, long processes, and span most of muscle®bre 12. (C) CSP staining is completely absentin the null mutant cspR1. Scale bars, 50 mm (Aand C) and 10 mm (B).
Drosophila presynaptic calcium currents 1823
Ó 1999 European Neuroscience Association, European Journal of Neuroscience, 11, 1818±1826
As a further indication of the divergence between Drosophila and
vertebrate Ca2+ channels, there is evidence that the binding sites for
DHPs and PAs are likely to be dissociated in Drosophila and other
invertebrate neurons (SchaÈfer et al., 1994; Wicher & Penzlin, 1997).
Pelzer et al. (1989) found several Ca2+ channel types in reconstituted
Drosophila head membranes. One channel is highly sensitive to PAs,
but insensitive to micromolar concentrations of DHPs, whereas two
other channels proved DHP-sensitive but PA-insensitive. These
observations ®t well with radioligand binding studies of the
Drosophila head, showing that PA-binding sites lack allosterically
coupled DHP receptors (Pauron et al., 1987; Glossmann et al., 1991).
While vertebrate L-channels bind DHPs, PAs and BTZs on the same
subunit, in Drosophila a particular channel appears to have binding
sites for either PAs or DHPs, but not for both. In fact, the only putative
a1 subunit so far cloned from the Drosophila adult head contains a
conserved PA-binding domain, but not a canonical DHP-binding site,
as known in vertebrates (Zheng et al., 1995). The emergent picture is
that of a very large repertoire of Ca2+ channels among cell types and
species.
Direct studies of presynaptic calcium channels have been limited
by the availability of preparations with suf®ciently large nerve
endings. Bipolar neurons from gold®sh retina only express a slowly
inactivating L-current component (Heidelberger & Matthews, 1992).
In contrast, dissociated nerve terminals from rat neurohypophysis and
presynaptic calyces of chick ciliary ganglia both contain several
Ca2+-current components (Lemos & Nowycky, 1989; Yawo, 1990).
Drosophila type III synaptic Ca2+ currents are most similar to
currents described in somata and isolated terminals of crab
neurosecretory cells (Richmond et al., 1995, 1996). These peptidergic
neurons appear to express a single Ca2+ current type with a high-
activation threshold and calcium-dependent inactivation, which, as in
the ¯y, is rather insensitive to nifedipine w-Conotoxin GVIA and w-
Agatoxin IVA.
The presence of Ca2+ currents in other Drosophila neurons is not
characterized with enough resolution to allow conclusions on the
diversity of Ca2+ channels present in this insect. Nifedipine- and
diltiazem-resistant HVA Ca2+ currents with both sustained and
transient components have been reported in embryonic neurons in
culture (Byerly & Leung, 1988; Leung et al., 1989). PLTX
completely blocks both components, whereas the Hololena curta
spider venom HoTX appears speci®c for the sustained component.
Nanomolar concentrations of either toxin cause complete and
irreversible inhibition of synaptic transmission at the larval
neuromuscular junction (Bowers et al., 1987; Branton et al., 1987;
our own unpublished observations). This differential effect of PLTX
on type III terminals versus glutamatergic transmission indicates that
the channels linked to glutamate release from type I boutons might be
different from those present in peptidergic terminals. Drosophila
FIG. 5. Synaptic Ba2+ currents in type III terminals of csp null mutants. (A) Ba2+ currents recorded from ecd1 (left traces) and cspR1 (right traces) type III boutons atrestrictive temperature, in response to voltage depolarizing pulses from a holding potential of ±50 mV (B) Average Ba2+ peak I±V relationship for Oregon-R, ecd1
and cspR1, at a permissive temperature (22 °C). (C) Average Ba2+ peak I±V relationship for ecd1 and cspR1, at a restrictive temperature (32 °C). At this temperature,all mutant animals tested were paralysed (see text). Additional observations were carried out at temperatures up to 36 °C without signi®cant changes in the currentamplitudes and voltage dependence.
1824 M. Morales et al.
Ó 1999 European Neuroscience Association, European Journal of Neuroscience, 11, 1818±1826

larval muscle ®bres also exhibit a voltage-dependent HVA Ba2+
current selectively blocked by L-type channel antagonists. This
current, however, proved almost two orders of magnitude more
sensitive to nifedipine, but less affected by PAs and BTZs (see also
Gielow et al., 1995), which agrees with the reported dissociation of
DHP and PA binding sites, as mentioned earlier.
The study of csp mutants is justi®ed because of the proposed role
of this protein as a presynaptic Ca2+ channel modulator. Thus,
coexpression in Xenopus oocytes of torpedo CSP cDNA positively
regulates the expression of N-type currents, and this expression is
selectively reduced by coinjection with a cloned antisense CSP
cRNA, which prompted the hypothesis that CSPs could act as
subunits or regulators of Ca2+ channels (Gundersen & Umbach,
1992). On the other hand, Drosophila null mutations for the csp gene
are semilethal during development and death is premature (Zinsmaier
et al., 1994). When exposed to high temperature, csp mutant larvae
exhibit a progressive decrement leading to ®nal blockade of nerve-
evoked, but not spontaneous, quantal release (Umbach et al., 1994).
Further characterization of the temperature-sensitive phenotype,
revealed that exocytosis can be promoted using methods that bypass
presynaptic Ca2+ channels, such as a-latrotoxin and Ca2+ ionophores,
but not by experimental procedures that rely on Ca2+ channel opening
(Umbach & Gundersen, 1997; Ranjan et al., 1998), suggesting that
calcium in¯ux might be impaired in csp mutants. A direct
demonstration of blocked Ca2+ entry in csp mutants at restrictive
temperature was recently provided by Umbach et al. (1998) using
imaging techniques. These authors, have also shown that calcium
in¯ux is blocked in vesicle-depleted shibiri synaptic terminals, in line
with the proposal that the interaction between CSP and Ca2+ channels
would take place during synaptic vesicle docking. Nonetheless,
evidence for a direct link between CSP and a Ca2+ channel is still
pending.
We have shown that type III boutons do express CSP protein, and
that this expression is abolished in cspR1 mutants. The electro-
physiological results, however, indicate that lack of CSP may not
affect type III terminal Ca2+ currents. Current peak amplitudes in csp
mutants measured at permissive (22 °C) versus restrictive (32 °C)
temperature were not signi®cantly different, and they were also
similar to those of control larvae at both temperatures. That is, Ba2+
currents at type III terminals stay the same in otherwise temperature-
sensitive-paralysed csp larvae. As Ca2+ in¯ux into csp type I
terminals is blocked at 32 °C (Umbach et al., 1998), these results
support the idea that absence of CSP has differential effects on
peptidergic versus glutamatergic release.
As glutamate (type I) and peptide (type III) release appear to
depend on different Ca2+ channel types, the interaction with CSP
might be channel-type speci®c. Alternatively, the putative CSP±Ca2+-
channel interaction might require the channels to be tightly coupled to
the docking complex (Sheng et al., 1996) which, under the present
hypothesis for CSP, is implied by the vesicular location of CSP. In
fact, one appealing aspect of this model is that only those channels in
close proximity to docked vesicles, and therefore in a prime position
to elicit effective exocytosis, would be allowed to open. However,
considering the widespread distribution of CSPs (see Buchner &
Gundersen, 1997) which includes non-neuronal tissues (Chamberlain
& Burgoyne, 1996), it is unlikely that presynaptic calcium channel
regulation is the only function of CSPs. In this respect, over-
expression of CSP enhances Ca2+-dependent and GTPgS-dependent
exocytosis from permeabilized PC12 cells, without affecting
depolarization-induced cytosolic Ca2+ signals (Chamberlain &
Burgoyne, 1998). An alternative hypothesis for CSP function
emphasizes the presence of a J-domain. These motives are known
to interact with heat-shock protein 70 (Hsp70) as chaperones to
regulate the folding of substrate proteins (Cyr et al., 1994). At the
synaptic terminal, Hsp70 is thought to participate in the uncoating of
clathrin-coated vesicles (Rothman & Schmid, 1986). In this context,
the function of CSP might be to contribute to rapid vesicle decoating
in conjunction with Hsp70 (Sudhof, 1995). In fact, CSP enhances
Hsc70 ATPase activity in a dose-dependent manner (Braun et al.,
1996). However, Ranjan et al. (1998) directly examined endocytosis
using the ¯uorescent probe FM1±43, and found no vesicle recycling
defect in Drosophila csp mutants. Taken together, the available data
on CSP indicate that there is a wider range of biological roles for this
protein than presented hereto.
Acknowledgements
We appreciate the critical comments on earlier versions of the manuscript ofDrs J. Lerma, W. BunÄo, A. Villarroel and O. Herreras. This research has beenfunded by grants DGICYT 93-0149 and 96-006 from the Spanish Ministry ofCulture
Abbreviations
CSP, cystein string protein; BTZ, benzothioazepines; DHP, dihydropyridines;Hsp70, Heat Shock Protein 70; HVA, high voltage activated; PA, phenylalk-ylamines; PBT, 0.1% Triton in PBS; PLTX, a potent toxin from the venom ofthe spider Plectreurys tristes.
References
Anderson, M.S., Halpern, M.E. & Keshishian, H. (1988) Identi®cation of theneuropeptide transmitter proctolin in Drosophila larvae: characterization of®ber-speci®c neuromuscular endings. J. Neurosci., 8, 242±255.
Atwood, H.L., Govind, C.K. & Wu, C.F. (1993) Differential ultrastructure ofsynaptic terminals on ventral longitudinal abdominal muscles in Drosophilalarvae. J. Neurobiol., 24, 1008±1024.
Bezprozvanny, I., Scheller, R.H. & Tsien, R.W. (1995) Functional impact ofsyntaxin on gating of N-type and Q-type calcium channels. Nature, 378,623±626.
Bowers, C.W., Phillips, H.S., Lee, P., Jan, Y.N. & Jan, L.Y. (1987)Identi®cation and puri®cation of an irreversible presynaptic neurotoxinfrom the venom of the spider Hololena curta. Proc. Natl Acad. Sci. USA,84, 3506±3510.
Branchaw, J.L., Banks, M.I. & Jackson, M.B. (1997) Ca2+- and voltage-dependent inactivation of Ca2+ channels in nerve terminals of theneurohypophysis. J. Neurosci., 17, 5772±5781.
Branton, W.D., Kolton, L., Jan, Y.N. & Jan, L.Y. (1987) Neurotoxins fromPlectreurys Spider venom are potent presynaptic blockers in Drosophila. J.Neurosci., 7, 4195±4200.
Braun, J.E.A., Wilbanks, S.M. & Scheller, R.H. (1996) The cysteine stringsecretory vesicle protein activates Hsc70 ATPase. J. Biol. Chem., 271,25989±25993.
Buchner, E., Bader, R., Buchner, S., Cox, J., Emson, P.C., Flory, E.,Heizmann, C.W., Hemm, S., Hofbauer, A. & Oertel, W.H. (1988) Cell-speci®c immuno-probes for the brain of normal and mutant Drosophilamelanogaster. I. Wildtype visual system. Cell Tissue Res., 253, 357±370.
Buchner, E. & Gundersen, C.B. (1997) The Dna-J like cysteine string proteinand exocytotic neurotransmitter release. Trends Neurosci., 20, 223±227.
Byerly, L. & Leung, H.T. (1988) Ionic currents of Drosophila neurons inembryonic cultures. J. Neurosci., 8, 4379±4393.
Chamberlain, L.H. & Burgoyne, R.D. (1996) Identi®cation of a novel cysteinestring protein variant and expression of cysteine string proteins in non-neuronal cells. J. Biol. Chem., 271, 7320±7323.
Chamberlain, L.H. & Burgoyne, R.D. (1998) Cysteine string protein functionsdirectly in regulated exocytosis. Mol. Biol. Cell, 9, 2259±2267.
Cyr, D.M., Langer, T. & Douglas, M.G. (1994) DnaJ-like proteins: molecularchaperones and speci®c regulators of Hsp70. Trends Biochem. Sci., 19, 176±181.
De Weille, J.R., Schweitz, H., Maes, P., Tartar, A. & Lazdunski, M. (1991)Calciseptine, a peptide isolated from black mamba venom, is a speci®cblocker of the L-type calcium channel. Proc. Natl Acad. Sci. USA, 88,2437±2440.
Drosophila presynaptic calcium currents 1825
Ó 1999 European Neuroscience Association, European Journal of Neuroscience, 11, 1818±1826

Dunlap, K., Luebke, J.I. & Turner, T.J. (1995) Exocytotic Ca2+ channels inmammalian central neurons. Trends Neurosci., 18, 89±98.
Garner, A., Kauvar, L. & Lepesant, J.A. (1977) Roles of ecdysone inDrosophila development. Proc. Natl Acad. Sci. USA, 74, 5099±5103.
Gielow, M.L., Gang-Guo, G. & Singh, S. (1995) Resolution andpharmacological analysis of the voltage dependent calcium channels ofDrosophila larval muscles. J. Neurosci., 15, 6085±6093.
Glossmann, H., Zech, C., Striessnig, J., Staudinger, R., Hall, L., Greenberg, R.& Armah, B.I. (1991) Very high af®nity interaction of DPI 201±106 andBDF 8784 enantiomers with the phenylalkylamine-sensitive Ca2+-channelin Drosophila head membranes. Br. J. Pharmacol., 102, 446±452.
Gorczyca, M.G., Augart, C. & Budnik, V. (1993) Insulin-like receptor andinsulin-like peptide are localized at neuromuscular junctions in Drosophila.J. Neurosci., 13, 3692±3704.
Gundersen, C.B. & Umbach, J.A. (1992) Suppression cloning of the cDNA fora candidate subunit of a presynaptic calcium channel. Neuron, 9, 527±537.
Heckmann, M., Adelsberger, H. & Dudel, J. (1997) Evoked transmitter releaseat neuromuscular junctions in wild type and cysteine string protein nullmutant larvae of Drosophila. Neurosci. Lett., 228, 167±170.
Heidelberger, R. & Matthews, G. (1992) Calcium in¯ux and calcium currentin single synaptic terminals of gold®sh retinal bipolar neurons. J. Physiol.(Lond.), 447, 235±256.
Horn, R. & Marty, A. (1988) Muscarinic activation of ionic currents measuredby a new whole-cell recording method. J. Gen. Physiol., 92, 145±159.
Iverfeldt, K., SerfoÈzoÈ, P., Diaz-Arnesto, L. & Bartfai, T. (1989) Differentialrelease of coexisting neurotransmitters: frequency dependence of the ef¯uxof substance P, thyrotropin releasing hormone and (3H (-serotonin fromtissue slices of rat ventral spinal cord. Acta Physiol. Scand., 137, 63±71.
Jan, L.Y. & Jan, Y.-N. (1976) Properties of the larval neuromuscular junctionin Drosophila melanogaster. J. Physiol. (Lond.), 262, 189±214.
Jia, X.-X., Gorczyca, M. & Budnik, V. (1993) Ultrastructure of neuromuscularjunctions in Drosophila: Comparison of wild type and mutants with increaseexcitability. J. Neurobiol., 24, 1025±1044.
Johansen, J., Halpern, M.E., Johansen, K.M. & Keshishian, H. (1989)Stereotypic morphology of glutamatergic synapses on identi®ed musclecells of Drosophila larvae. J. Neurosci., 9, 710±725.
Koham, S.A., Pescatori, M., Brecha, N.C., Mastrogiacomo, A., Umbach, J.A.& Gundersen, C.B. (1995) Cysteine string protein immunoreactivity in thenervous system and adrenal gland of rat. J. Neurosci., 15, 6230±6238.
Lee, K.S. & Tsien, R.W. (1983) Mechanism of calcium channel blockade byverapamil, D600, diltiazem and nitrendipine in single dialysed heart cells.Nature, 302, 790±794.
Lemos, J.R. & Nowycky, M.C. (1989) Two types of calcium channels coexistin peptide-releasing vertebrate nerve terminals. Neuron, 2, 1419±1426.
Leung, H.T., Branton, W.D., Phillips, H.S., Jan, L. & Byerly, L. (1989) Spidertoxins selectively block calcium currents in Drosophila. Neuron, 3, 767±772.
LlinaÂs, R., Steinberg, I.Z. & Walton, K. (1981) Presynaptic calcium currents insquid giant synapse. Biophys. J., 33, 289±322.
MartõÂnez-Padron, M. & FerruÂs, A. (1997) Presynaptic recordings fromDrosophila. Correlation of macroscopic and single-channel K+ currents. J.Neurosci., 17, 3412±3424.
MartõÂnez-PadroÂn, M. & FerruÂs, A. (1998) Patch-clamp recordings fromDrosophila: presynaptic terminals. In: de Pablo, F., FerruÂs, A. & Stern, C.D.(eds.), Cellular and Molecular Procedures in Developmental Biology:Current Topics in Developmental Biology, Vol. 36. New York: AcademicPress, pp. 303±312.
Mastrogiacomo, A., Parsons, S.M., Zampighi, G.A., Jenden, D.J., Umbach,J.A. & Gundersen, C.B. (1994) Cysteine string proteins: a potential linkbetween synaptic vesicles and presynaptic Ca2+ channels. Science, 263,981±982.
Neher, E. & Zucker, R.S. (1993) Multiple calcium dependent processes relatedto secretion in bovine chromaf®n cells. Neuron, 10, 21±30.
Pauron, D., Qar, J., Barhanin, J., Fournier, D., Cuany, A., Pralavorio, M.,Berge, J.B. & Lazdunski, M. (1987). Identi®cation and af®nity labeling ofvery high af®nity binding sites for the phenylalkylamine series of Ca2+
channel blockers in the Drosophila nervous system. Biochemistry, 26,6311±6315.
Pelzer, S., Barhanin, J., Pauron, D., Trautwein, W., Lazdunski, M. & Pelzer,D. (1989) Diversity and novel pharmacological properties of Ca2+ channelsin Drosophila brain membranes. EMBO J., 8, 2365±2371.
Pupier, S., Leveque, C., Marqueze, B., Kataoka, M., Takahashi, M. & Seagar,M.J. (1997) Cysteine string proteins associated with secretory granules ofthe rat neurohypophysis. J. Neurosci., 17, 2722±2727.
Ranjan, R., Bronk, P. & Zinsmaier, K.E. (1998) Cysteine string protein isrequired for calcium secretion coupling of evoked neurotransmission inDrosophila but not for vesicle recycling. J. Neurosci., 18, 956±964.
Rettig, J., Heinemann, C., Ashery, U., Sheng, Z.H., Yokoyama, C.T., Catterall,W.A. & Neher, E. (1997) Alteration of Ca2+ dependence of neurotransmitterrelease by disruption of Ca2+ channel/syntaxin interaction. J. Neurosci., 17,6647±6656.
Richmond, J.E., Penner, R., Keller, R. & Cooke, I.M. (1996) Characterizationof the Ca2+ current in isolated terminals of crustacean peptidergic neurons.J. Exp. Biol., 199, 2053±2059.
Richmond, J.E., Sher, E. & Cooke, I.M. (1995) Characterization of the Ca2+
current in freshly dissociated crustacean peptidergic neuronal somata. J.Neurophysiol., 73, 2357±2368.
Rothman, J.E. & Schmid, S.L. (1986) Enzymatic recycling of clathrin fromcoated vesicles. Cell, 46, 5±9.
SchaÈfer, S., Rosenboom, H. & Menzel, R. (1994) Ionic currents of Kenyoncells from the mushroom body of the honeybee. J. Neurosci., 14, 4600±4612.
Schweitz, H., Heurteaux, C., Bois, P., Moinier, D., Romey, G. & Lazdunsky,M. (1994) Calcicludine, a venom peptide of the Kunitz-type proteaseinhibitor family, is a potent blocker of high-threshold Ca2+ channels with ahigh af®nity for L-type channels in cerebellar granule neurons. Proc. NatlAcad. Sci. USA, 91, 878±882.
Sheng, Z.-H., Rettig, J., Cook, T. & Catterall, W.A. (1996) Calcium dependentinteraction of N-type calcium channels with the synaptic core complex.Nature, 379, 451±454.
Skeer, J.M., Norman, R.I. & Sattelle, D.B. (1996) Invertebrate voltage-dependent calcium channel subtypes. Biol. Rev., 71, 137±154.
Stanley, E.F. & Goping, G. (1991) Characterization of a calcium current in avertebrate cholinergic presynaptic nerve terminal. J. Neurosci., 11, 985±993.
Sudhof, T.C. (1995) The synaptic vesicle cycle: a cascade of protein±proteininteractions. Nature, 375, 645±653.
Umbach, J.A. & Gundersen, C.B. (1997) Evidence that cysteine string proteinsregulate an early step in the Ca2+-dependent secretion of neurotransmitter atDrosophila neuromuscular junctions. J. Neurosci., 17, 7203±7209.
Umbach, J.A., Minoru, S., Kidokoro, Y. & Gundersen, C.B. (1998) Attenuatedin¯ux of calcium ions at nerve endings of csp and shibire mutant drosophila.J. Neurosci., 18, 3233±3240.
Umbach, J.A., Zinsmaier, K.E., Eberle, K.K., Buchner, E., Benzer, S. &Gundersen, C.B. (1994) Presynaptic dysfunction in Drosophila csp mutants.Neuron, 13, 899±907.
Wicher, D. & Penzlin, H. (1997) Ca2+ currents in central insect neurons:electrophysiological and pharmacological properties. J. Neurophysiol., 77,186±199.
Wiser, O., Bennett, M.K. & Atlas, D. (1996) Functional interaction of syntaxinand SNAP-25 with voltage-sensitive L- and N-type Ca+2 channels. EMBOJ., 15, 4100±4110.
Yawo, H. (1990) Voltage-activated calcium currents in presynaptic nerveterminals of the chick ciliary ganglion. J. Physiol. (Lond.), 428, 199±213.
Zheng, W., Feng, G., Ren, D., Eberl, D.F., Hannan, F., Dubald, M. & Hall,L.M. (1995) Cloning and characterization of a calcium channel (1 subunitfrom Drosophila melanogaster with similarity to the rat brain type Disoform. J. Neurosci., 15, 1132±1143.
Zinsmaier, K.E., Eberle, K.K., Buchner, E., Walter, N. & Benzer, S. (1994)Paralysis and early death in cysteine string protein mutants of Drosophila.Science, 263, 977±980.
1826 M. Morales et al.
Ó 1999 European Neuroscience Association, European Journal of Neuroscience, 11, 1818±1826