Embed Size (px)
Citation preview

Preservation of the pattern of tyrosine phosphorylation
in human neutrophil lysates
II. A sequential lysis protocol for the analysis of tyrosine
phosphorylation-dependent signalling
Caroline Gilbert 1, Emmanuelle Rollet-Labelle 1, Paul H. Naccache *
Centre de Recherche en Rhumatologie et Immunologie, CIHR group on the Molecular Mechanisms of Inflammation,
Centre de Recherche du CHUL, and Department of Medicine, Faculty of Medicine, Laval University, Ste-Foy, Quebec, Canada
Received 17 September 2001; received in revised form 6 November 2001; accepted 6 November 2001
Abstract
In stimulated neutrophils, themajority of tyrosine phosphorylated proteins are concentrated in TritonX-100 or NP-40 insoluble
fractions. Most immunobiochemical studies, whose objective is to study the functional relevance of tyrosine phosphorylation are,
however, performed using the supernatants of cells lysed in non-ionic detergent-containing buffers (RIPA lysis buffers). This
observation prompted us to develop an alternative lysis protocol. We established a procedure involving the sequential lysis of
neutrophils in buffers of increasing tonicities that not only preserved and solubilized tyrosine phosphorylated proteins but also
retained their enzymatic activities. The sequential lysis of neutrophils in hypotonic, isotonic and hypertonic buffers containing
non-ionic detergents resulted in the solubilisation of a significant fraction of tyrosine phosphorylated proteins. Furthermore, we
observed that in monosodium urate crystals-stimulated neutrophils, Lyn activity was enhanced in the soluble fraction recovered
from the hypertonic fraction, but not from that of the first hypotonic lysis. The distribution of tyrosine phosphorylated proteins
between the NP-40 soluble and insoluble fractions was both substrate- and agonist-dependent. In neutrophils stimulated with
fMet-Leu-Phe, MSU crystals or by CD32 ligation, the tyrosine phosphorylated proteins were mostly insoluble. On the other hand,
in GM-CSF-treated cells, the phosphoproteins were more equally distributed between the two fractions. The results of this study
provide a new experimental procedure for the investigation of tyrosine phosphorylation pathways in activated human neutrophils
which may also be applicable to other cell types. D 2002 Elsevier Science B.V. All rights reserved.
Keywords: Cbl; CD32; Detergent insolubility; Fcg receptors; fMet-Leu-Phe; GM-CSF; Kinase activity; Lyn; Monosodium urate crystals;
Neutrophil; Phagocytosis; Signal transduction; Syk; Tyrosine phosphorylation
1. Introduction
Neutrophils play important roles in host defense
against injury and infection as well as in the inflam-
matory response (Smith, 1994). In order to accomplish
their function, neutrophils store in their granules var-
ious proteases and microbicidal agents (Borregaard
0022-1759/02/$ - see front matter D 2002 Elsevier Science B.V. All rights reserved.
PII: S0022-1759 (01 )00553 -1
Abbreviations: HLB, hypotonic buffer; HyperLB, hypertonic
buffer; ILB, isotonic buffer; LB, lysis buffer; MSU, Monosodium
urate; RIPA, rapid immunoprecipitation assay; SB, sample buffer.* Corresponding author. CHUL, Room T1-49, 2705 Boulevard
Laurier, Ste-Foy, Quebec, Canada G1V 4G2. Tel.: +1-418-654-
2772; fax: +1-418-654-2765.
E-mail address: [email protected]
(P.H. Naccache).1 The authors contributed equally to this study.
www.elsevier.com/locate/jim
Journal of Immunological Methods 261 (2002) 85–101

and Cowland, 1997) and are able to produce toxic
oxygen-derived metabolites (Rosen and Michel, 1997)
and lipids mediators (McDonald et al., 1991; Pouliot et
al., 1998) as well as to synthesize and secrete various
proteins involved in the regulation of the inflammatory
response (Hachicha et al., 1995; Cassatella, 1996).
Neutrophils respond to a wide variety of soluble
factors including cytokines (McDonald et al., 1991;
Pouliot et al., 1998) and chemoattractants (Torres et
al., 1993) as well as to particulate agonists (Gaudry et
al., 1993; Roberge et al., 1993). One of the earliest
events observed upon stimulation of neutrophils is an
increase in the level of tyrosine phosphorylation of a
number of proteins (Rollet et al., 1994). Several
neutrophil functional responses including adherence
(Gaudry et al., 1992; Naccache et al., 1994), locomo-
tion (Gaudry et al., 1992), and superoxide production
(Naccache et al., 1990) are inhibited by tyrosine kinase
inhibitors or enhanced by tyrosine phosphatase inhib-
itors (Grinstein et al., 1990; Cui et al., 1994). The
activity of multiple tyrosine kinases including Fgr
(Gutkind and Robbins, 1989; Hamada et al., 1993;
Krump et al., 1997), Lyn (Gaudry et al., 1995; Yan et
al., 1996; Ibarrola et al., 1997; Welch and Maridon-
neau-Parini, 1997), Hck (Welch et al., 1996), Jak2 (Al-
Shami et al., 1998), Pyk2 (Yan and Novak, 1999) and
Syk (Asahi et al., 1993; Yan and Berton, 1996; Yan et
al., 1997; Fernandez and Suchard, 1998) is modulated
after the stimulation of neutrophils. The presence of
tyrosine phosphatases such as SHP-1 (Brumell et al.,
1997), SHP-2 (Tidow et al., 1999), CD148 (Hundt and
Schmidt, 1997) and CD45 (Cui et al., 1996; Fialkow et
al., 1997) has been observed in human neutrophils.
Several tyrosine phosphorylated substrates such as Cbl
(Naccache et al., 1997; Willeke et al., 2000), CD32
(Rollet-Labelle et al., 2000), CD50 (Skubitz et al.,
1995), members of the MAP kinase family (Grinstein
and Furuya, 1992; Torres et al., 1993; Nahas et al.,
1996), paxillin (Fuortes et al., 1994; Fernandez and
Suchard, 1998), and SAM68 (Gilbert et al., 2001) have
also been identified.
The mechanisms linking neutrophil surface recep-
tors to tyrosine kinase signalling are, however, incom-
pletely understood. Immunoprecipitation coupled with
SDS-polyacrylamide gel electrophoresis is one of the
most commonly used immuno-chemical techniques
for studying tyrosine phosphorylation-dependent
pathways. This technique can potentially determine
several important characteristics of the antigen under
examination such as its presence and quantities, its
relative molecular weight, its rate of synthesis or
degradation, and the presence of certain post-transla-
tional modifications. It can also provide information
about interactions with proteins, nucleic acids and
other ligands. These protocols depend on lysing the
cells under mild conditions in RIPA buffers containing
cocktails of protease and phosphatase inhibitors.
These lysates are then centrifuged and the super-
natants are used for subsequent immunoprecipitations.
It is commonly observed, however, that the profile of
tyrosine phosphorylated proteins of whole neutrophils
is rapidly lost or artificially increased upon lysing the
cells in classical RIPA buffers. An alternative assay
involving lysis under denaturing conditions was pre-
viously developed to preserve the phosphorylation
levels in neutrophil lysates (Al-Shami et al., 1997b).
Although this protocol allows the identification of
tyrosine phosphorylated proteins (Al-Shami et al.,
1997b; Barabe et al., 1998; Khamzina and Borgeat,
1998; Rollet-Labelle et al., 2000; Gilbert et al., 2001),
it disrupts protein–protein interactions and inactivates
most enzymatic activities.
Most studies of the type discussed above analyze
the soluble fractions following lysis in RIPA buffers
while discarding the insoluble fractions. Several stud-
ies, however, have identified and focussed on the
formation of detergent-resistant membrane structures
(DRM, DIGs or lipids rafts) into which tyrosine
phosphorylated proteins and tyrosine kinases tend to
concentrate after receptor stimulation (Zhou et al.,
1995; Yan et al., 1996). The function of these deter-
gent-insoluble structures appears to be related to
signal transmission and/or membrane trafficking. Sev-
eral studies have shown that tyrosine kinase activities
were increased and concentrated in these insoluble
fractions upon ligation of phagocytic receptors in
adherent human neutrophils (Zhou et al., 1995; Yan
et al., 1996). Analysis of tyrosine phosphorylation
events in these structures depends on the preparation
of stable cell lysates that can be used as starting
material for their analysis by immunoprecipitation.
In the present study, the distribution and preserva-
tion of tyrosine phosphorylated substrates and of
several signalling-associated molecules was moni-
tored in human neutrophil lysates prepared under a
variety of native conditions. The results obtained
C. Gilbert et al. / Journal of Immunological Methods 261 (2002) 85–10186

illustrate the inadequacy of standard cell lysates
prepared under native conditions for use as starting
material in subsequent immunoprecipitation or co-
immunoprecipitation protocols. An alternative method
based on sequential lysis under conditions of increas-
ing tonicity is described. The use of this alternative
protocol underlined the critical importance of the
complete monitoring of the tyrosine phosphorylation
status as well as of the solubility and, when appli-
cable, the enzymatic activity of each individual target
molecule, in response to each and every agonist.
2. Materials and methods
2.1. Antibodies
The polyclonal antibodies, anti-Cbl (sc-170), anti-
Lyn (sc-15) and anti-SHP-1 (sc-287, 1/1000) were
purchased from Santa Cruz Biotechnology (Santa
Cruz, CA, USA). The monoclonal antibodies anti-
Syk (MAB88906), anti-phosphotyrosine (UBI 05-
321, clone 4G10) were purchased from Chemicon
International (Missisauga, Ontario, Canada) and Up-
state Biotechnology (Lake Placid, NY, USA) respec-
tively. An anti-FcgRII polyclonal antiserum against the
cytoplasmic tail of CD32 (CT-10) was generated in our
laboratory as described previously (Ibarrola et al.,
1997). Monoclonal anti-FcgRII antibodies (IV.3) werepurified from the ascitic fluids of mice inoculated with
hybridoma HB 217 which was obtained from the
American Type Culture Collection (ATCC) (Rock-
ville, MD, USA). F(abV)2 fragments of the IV.3 anti-
body were prepared essentially as described in the
Pierce catalog (Rockford, IL, USA). Briefly, the anti-
bodies were digested with pepsin and intact antibodies
were eliminated by adding protein A and protein G
beads. The integrity and purity of the F(abV)2 frag-
ments was verified by their ability to label intact hu-
man neutrophils as determined by flow cytometry as
well as by their neutrophil activating properties (cal-
cium mobilization, superoxide generation, tyrosine
phosphorylation).
2.2. Reagents
Triton X100, trypsin inhibitor soybean, fMet-Leu-
Phe, and sodium orthovanadate (Na2VO4) were pur-
chased from Sigma Chemical (St-Louis, MO, USA).
Di-isopropylfluorophosphate (DFP), NP-40 10% sol-
ution and NP-40 were obtained from Calbiochem (La
Jolla, CA, USA). P-Nitrophenylphosphate, aprotinin
and leupeptin were purchased from ICN Pharmaceut-
icals (Costa Mesa, CA, USA). GM-CSF was supplied
by the Genetics Institute (Cambridge, MA, USA).
Triclinic MSU crystals were kindly provided by Drs.
R. de Medicis and A. Lussier (Universite of Sher-
brooke, Sherbrooke, Quebec, Canada) and prepared as
previously described (Gaudry et al., 1993). Dextran T-
500, Ficoll Paque, sephadex G-10, protein A sephar-
ose were purchased from Pharmacia Biotech (Dorval,
Quebec, Canada).
2.3. Cells
Neutrophils were obtained from healthy adult vol-
unteers as previously described (Rollet et al., 1994)
and were resuspended at a concentration of 4� 107
cells/ml in Hanks’ balanced salt solution (HBSS)
containing 1.6 mM calcium and no magnesium, pH
7.4.
2.4. Cell stimulation
Before all stimulations, neutrophil suspensions
(4� 107 cells/ml) were pre-incubated at room temper-
ature with 1 mM DFP (to minimize proteolysis) for 5
min. Neutrophils were then stimulated at the same
concentration at 37 �C with 3 mg/ml of MSU crystals
or 3 nM of GM-CSF for 10 min or with 100 nM of
fMet-Leu-Phe for 1 min. For CD32 cross-linking, the
cell suspensions were pre-incubated with 2.0 mg/ml of
anti-FcgRII (IV.3) antibodies for 1 min at 37 �C and
stimulated by cross-linking the cell-bound antibodies
with 20 mg/ml of F(abV)2 goat anti-mouse Fc antibody
(Jackson Immune Research, Mississauga, ON, Can-
ada) for 1 min at 37 �C or the indicated periods of
time. For immunoprecipitations, the F(abV)2 fragment
of IV.3 was used instead of the whole antibody.
2.5. Preparation of total cell lysates under denaturing
conditions
After stimulation, the reactions were stopped by
transferring 100 ml of the cell suspensions to an equal
volume of boiling 2� Laemmli sample buffer (SB)
C. Gilbert et al. / Journal of Immunological Methods 261 (2002) 85–101 87

(1� is 62.5 mM Tris–HCl, pH 6.8, 4% SDS, 5% b-mercaptoethanol, 8.5% glycerol, 2.5 mM orthovana-
date, 10 mM p-nitrophenylphosphate, 10 mg/ml leu-
peptin, 10 mg/ml aprotinin, 0.025% bromophenol
blue) and boiled for 7 min.
2.6. Hypotonic cell lysis
Neutrophils were pre-incubated and stimulated as
described above. After stimulation, the reactions were
rapidly stopped by transferring the cell suspensions in
pre-cooled (� 20 �C) 1.5-ml microcentrifuge tubes.
The cells were then centrifuged for 5–10 s at
6000� g in a micro-centrifuge and the pellets resus-
pended at a final concentration of 4� 107 cells/ml in a
hypotonic lysis buffer (HLB) (final concentrations:
0.1% NP-40, 20 mM Tris–HCl pH 7.5, 10 mM NaCl,
1 mM EDTA, 2 mM orthovanadate, 10 mg/ml aproti-
nin, 10 mg/ml leupeptin, 2 mM PMSF, 50 mg/ml
trypsin inhibitor soybean, and 3 mM DFP). For better
inhibition of phosphatases, p-nitrophenylphosphate 10
mM may be added in the lysis buffer. After 5 min of
incubation at 4 �C, the lysates were centrifuged at
600� g for 10 min at 4 �C. Aliquots of the super-
natants were added to an equal volume of boiling
2� SB or used for immunoprecipitation under native
or denaturing conditions or for the determination of
tyrosine kinase activities. The pellets were resus-
pended in HLB and then diluted in the same volume
of 2� SB or in an isotonic lysis buffer (ILB) or a
hypertonic lysis buffer (HyperLB) (see below for
composition) for immunoprecipitation or determina-
tion of tyrosine kinase activity. Nuclear integrity and
total cell lysis were routinely verified by light micro-
scopy.
2.7. Isotonic cell lysis
Neutrophils were pre-incubated and stimulated as
described above. After stimulation, the reactions were
rapidly stopped by transferring the cell suspensions
into precooled (� 20 �C) 1.5-ml microcentrifuge
tubes. The cell suspensions were then centrifuged
for 5–10 s at 6000� g in a micro-centrifuge. The
supernatants were removed and the cell pellets were
resuspended and incubated for 5 min at 4� 107 cells/
ml at 4 �C in cold isotonic lysis buffer (ILB) (final
concentrations: 1% NP-40, 20 mM Tris–HCl pH 7.5,
137 mM NaCl, 1 mM EDTA, 2 mM orthovanadate,
10 mg/ml aprotinin, 10 mg/ml leupeptin, 2 mM PMSF,
50 mg/ml trypsin inhibitor soybean, and 3 mM DFP).
The lysates were centrifuged at 600� g (Fig. 1) or
13,000� g (Fig. 5) for 10 min at 4 �C. Aliquots of thesupernatants were added to an equal volume of boil-
ing 2� SB. The pellets were resuspended in ILB and
then diluted in the same volume of 2� SB.
2.8. Sequential solubilisation
After stimulation, the soluble and insoluble frac-
tions of the hypotonic lysis were individually pro-
cessed for subsequent analysis. The soluble fraction
was centrifuged at 180,000� g at 4 �C for 45 min.
The supernatant was analyzed by transfer of an aliquot
into 2� SB. The pellet was dissolved in 1 volume of
HLB and transferred to boiling 2� SB. The insoluble
fraction of HLB was resuspended in ILB and centri-
fuged for 10 min at 13,000� g. The soluble material
of this second lysis was transferred into 2� SB and
the pellet was resuspended in the hypertonic lysis
buffer (HyperLB) (final concentrations: 1% NP-40,
1% Triton X100, 20 mM Tris–HCl pH 7.5, 400 mM
NaCl, 1 mM EDTA, 2 mM orthovanadate, 10 mg/ml
aprotinin, 10 mg/ml leupeptin, 2 mM PMSF, 50 mg/ml
trypsin inhibitor soybean, and 3 mM DFP). Two 5-s
sonication steps were necessary for maximal pellet
extraction. The lysates were centrifuged at 13,000� g
for 10 min at 4 �C. Aliquots of the supernatants weretransferred to an equal volume of boiling 2� SB. The
pellets were resuspended in HLB and then diluted in
the same volume of 2� SB. The samples were then
electrophoresed as described above.
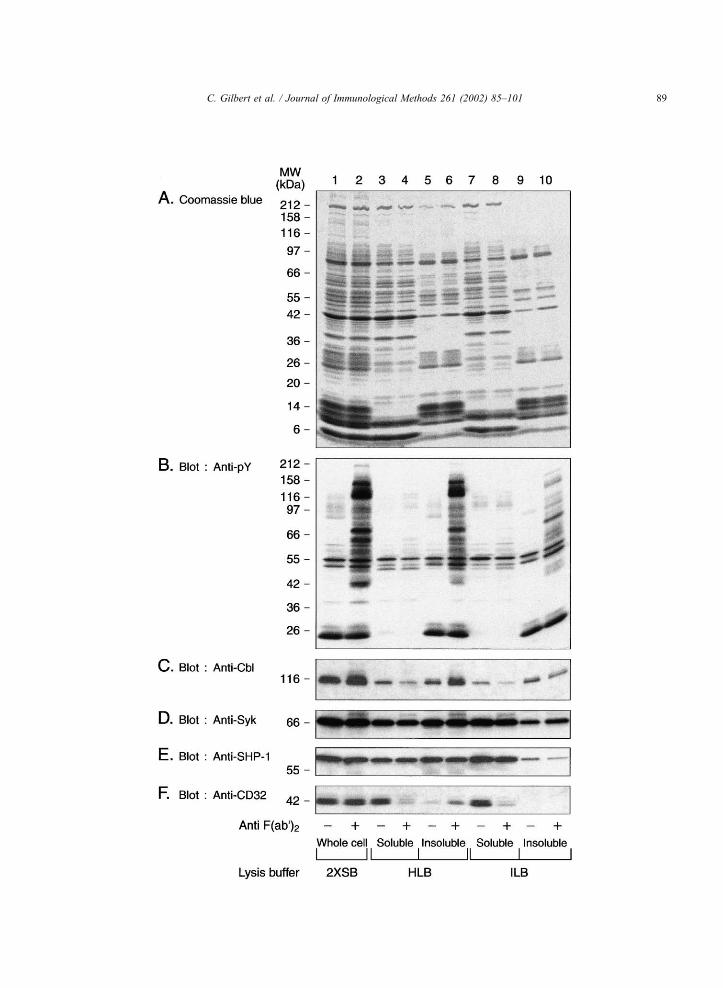
Fig. 1. Distribution and preservation of tyrosine phosphorylation profiles and signalling molecules in HLB and ILB. Neutrophils (4� 107 cells/
ml) were stimulated by cross-linking CD32 as indicated in Materials and methods. Briefly, the reactions were stopped by a rapid transfer of cell
aliquots into a precooled Eppendorf tube and a rapid centrifugation for lanes 3–10 or in the boiling 2� SB for lanes 1–2. The appropriate lysis
buffer was then added to the cell pellet, HLB for lanes 3–6 and ILB for lanes 7–10. Aliquots of soluble (lanes 3, 4 and 7, 8) and insoluble
(lanes 5, 6 and 9, 10) fractions were diluted in an equal volume of 2� SB. The samples were analysed by coomassie blue protein staining (panel
A) or by Western blot with anti-phosphotyrosine, anti-Cbl, anti-Syk, anti-SHP-1 or anti-CD32 antibodies (panels B–F, respectively).
C. Gilbert et al. / Journal of Immunological Methods 261 (2002) 85–10188
C. Gilbert et al. / Journal of Immunological Methods 261 (2002) 85–101 89

The soluble fractions of HyperLB were also used
under native conditions for immunoprecipitation and
tyrosine kinase activity. These fractions (soluble
HyperLB) were also obtained by direct addition of
HyperLB to the HLB pellets.
2.9. Immunoprecipitation under denaturing condi-
tions
Neutrophils were incubated and stimulated as
described above. Aliquots (500 ml) of the cells were
lysed by direct transfer to an equal volume of boiling
2� lysis buffer (2�LB) (1� is 62.5 mM Tris–HCl,
pH 6.8, 2% SDS, 1.5% b-mercaptoethanol, 8.5%
glycerol, 2.5 mM orthovanadate, 10 mg/ml leupeptin,
10 mg/ml aprotinin, 0.025% bromophenol blue) and
boiled for 7 min. Immunoprecipitations were per-
formed as previously described (Al-Shami et al.,
1997b). Briefly, lysates were filtered through sepha-
dex G-10 columns to remove the denaturing agents.
The filtered lysates were precleared with protein A-
sepharose at 4 �C for 30 min in the presence of 1%
NP-40, 2 mM orthovanadate, 10 mg/ml leupeptin, and
10 mg/ml aprotinin. The samples were then immuno-
precipitated using 2 mg of anti-Cbl, 2 mg of anti-Syk or
6 mg of anti-CD32 previously bound to protein A-
sepharose for 90 min at 4 �C on a rotator platform
with constant end-over-end mixing. The beads were
collected and washed four times with a lysis buffer
containing 137 mM NaCl, 1% NP-40 but no SDS, b-mercaptoethanol or bromophenol blue. Sample buffer
(40 ml, 2� ) was added to the beads which were
boiled for 7 min. The proteins in the samples were
then separated by electrophoresis as described above.
The membranes were blotted with anti-phosphotyro-
sine or with the immunoprecipitating antibodies (anti-
Syk, anti-Cbl or anti-CD32) for visualizing the
amounts of precipitated protein.
2.10. Native immunoprecipitation and tyrosine kinase
activity
The aliquots (200 ml or 8� 106 cell equivalents) of
HLB or HyperLB supernatant lysates were mixed
with HyperLB or HLB, respectively, to bring back
the NaCl, NP-40 and Triton X-100 concentrations to
137 mM, 0.75%, 0.75%, respectively. These isotonic
lysates were immunoprecipitated using 1.5 mg of anti-
Lyn for 90 min at 4 �C on a rotator platform with
constant end-over-end mixing. Fifty microliters (30%
slurry) of protein A-sepharose was then added and the
samples were incubated for 1 h at 4 �C. The beads
were collected and washed four times with ILB buffer
without EDTA. The beads were incubated at 4 �C in
kinase buffer (1�KB, final concentrations: 500 mMHepes pH 7.5, 10 mM MgCl2, 3 mM MnCl2 with or
without to 50 mM ATP) and transferred to 37 �C for
the indicated times. The reactions were stopped by a
quick spin and the supernatants were removed. Sam-
ple buffer (40 ml, 2�) was added to the beads which
were boiled for 7 min. The proteins in the samples
were then separated by electrophoresis as described
above. The membranes were blotted with the anti-
phosphotyrosine, or with the immunoprecipitating
antibodies for visualizing the amounts of precipitated
protein. Immunoprecipitation with control rabbit IgG
followed by kinase activity was simultaneously under-
taken to verify the specificity of the precipitations.
2.11. Electrophoresis and immunoblotting
Before electrophoresis, all samples in 2� SB were
boiled for 7 min. The samples (equivalent cell num-
bers) were then subjected to 7.5–20% SDS-PAGE
and the proteins transferred to Immobilon PVDF
membranes (Millipore, Bedford, MA, USA). Immu-
noblotting was performed using the appropriate anti-
bodies (anti-Cbl (1/1000), anti-Lyn (1/2000), anti-
SHP-1 (1/1000), anti-CD32 (1/1000), anti-Syk (1/
2000) and anti-phosphotyrosine (1/4000)) and re-
vealed using the renaissance detection system (NEN
Life Science, Boston, MA.) using HRP-conjugated
secondary anti-mouse or anti-rabbit antibodies (Jack-
son, Immune Research, Mississauga, ON, Canada) at
a dilution of 1/20,000 as previously described (Al-
Shami et al., 1997a).
3. Results
3.1. Effect of tonicity of the lysis buffers on the
preservation and distribution of tyrosine phosphor-
ylation patterns and signalling-associated molecules
The difficulties associated with the preservation of
phosphotyrosine profiles and protein integrity in
C. Gilbert et al. / Journal of Immunological Methods 261 (2002) 85–10190

human neutrophils have recently been described (Al-
Shami et al., 1997b; Naccache et al., 1997). The
extraction of tyrosine phosphorylated proteins is
known to be affected by the composition of the lysis
buffer (Woods Ignatoski, 1996). The pH, presence of
phosphate, salt concentration, cell concentration, pres-
ence of cations and protease and phosphatase inhib-
itors can all modify the detergent behavior and the
solubilisation of individual proteins. Previous studies
have reported the solubilisation of proteins without
affecting the structure of the nuclei using a hypotonic
lysis buffer coupled with low concentrations of deter-
gents (0.1% NP-40) (Pouliot et al., 1996; McDonald
et al., 1998; Gilbert et al., 2001). The data in Fig. 1
compare this hypotonic buffer (HLB) with a more
classical isotonic RIPA buffer (ILB). The soluble and
insoluble fractions of the two lysis buffers (lanes 3–
10) were compared to the whole cell lysates (lanes 1–
2). The protein and tyrosine phosphorylation profiles
as well as the preservation of several proteins are
illustrated in panels A–F for the soluble (lanes 3–4,
7–8) and insoluble (lanes 5–6, 9–10) fractions
derived from these cell lysis protocols. To study the
tyrosine phosphorylation profile, neutrophils were sti-
mulated by cross-linking CD32 for 1 min at 37 �C as
described in Materials and methods. The samples
were analyzed by Coomassie blue protein staining
(panel A) or by immunoblotting with anti-phospho-
tyrosine, anti-Cbl, anti-Syk, anti-SHP-1 or anti-CD32
antibodies (panels B–F, respectively).
The percentage of lysed cells and the integrity of
the nuclei were routinely verified by light microscopy
(data not shown) or by immunoblot analysis with
nuclear markers such as p62 nucleoporin (a constitu-
ent of nuclear membranes). This confirmed that the
cells were completely lysed in the two buffers and that
nuclear integrity was better preserved in HLB than in
ILB (data not shown). Coomassie blue staining
(panel A) of these samples indicated that the amount
of protein extracted with the two lysis buffers was
roughly similar.
The tyrosine phosphorylation profile induced by
CD32 cross-linking, as monitored after the direct
transfer of the cells into sample buffer, is illustrated
in the first two lanes of panel B and served as a
reference point for the evaluation of the adequacy of
the native cell lysates. Two major conclusions can be
drawn from monitoring the tyrosine phosphorylation
profiles in the various fractions (panel B). Firstly,
tyrosine phosphorylated proteins were concentrated
in the insoluble fractions of both lysis protocols (lanes
5–6 and 9–10). Secondly, the tyrosine phosphoryla-
tion profiles were significantly better preserved in
HLB than in ILB (lanes 4–6 vs. lanes 8–10).
Immunoblotting of the various fractions with anti-
Cbl, anti-Syk, anti-SHP-1 or anti-CD32 antibodies
(panels C–F, respectively) further illustrated the dif-
ferences between the HLB and ILB lysis procedures.
The anti-Cbl blots showed that Cbl was better pre-
served in the soluble and insoluble fractions of HLB
(lanes 3–6) than ILB (7–10). Additionally, CD32
cross-linking induced a translocation of Cbl to the
insoluble fractions which was more evident in HLB
than ILB. The tyrosine kinase Syk was somewhat
more soluble in ILB than HLB and was relatively
insensitive to proteolysis. SHP-1 was also well-pre-
served in both lysis buffers though it was more easily
extracted in ILB than in HLB. Ligation of CD32
induced a translocation of CD32 from the soluble to
the insoluble fractions which was more readily appa-
rent in HLB than ILB. Hence, ILB is adequate for
studies requiring an optimal solubilisation of Syk and
SHP-1. However, based on the three criteria, (i)
prevention of proteolysis, (ii) preservation of tyrosine
phosphorylation and (iii) day-to-day reproducibility of
tyrosine phosphorylation levels (data not shown), the
data in Fig. 1 indicate that HLB is the preferred buffer
for the preparation of starting material from which to
study tyrosine phosphorylation-dependent signalling
events in human neutrophils.
3.2. The presence of tyrosine kinase cofactors in the
lysis buffers artefactually increases the tyrosine
phosphorylation levels in the soluble and insoluble
fractions
The composition of a lysis buffer can modify the
solubility, activity and phosphorylation status of pro-
teins and their intramolecular associations. Mg2 + is a
known tyrosine kinase cofactor which is present in the
lysis buffers used in various studies. The effects of the
addition of Mg2 + to HLB were examined and the
results of these experiments are summarized in Fig. 2.
Panel A presents the patterns of tyrosine phosphor-
ylation observed in whole cell lysates (lanes 1 and 2),
and in the soluble (lanes 3–4, 7–8) and insoluble
C. Gilbert et al. / Journal of Immunological Methods 261 (2002) 85–101 91
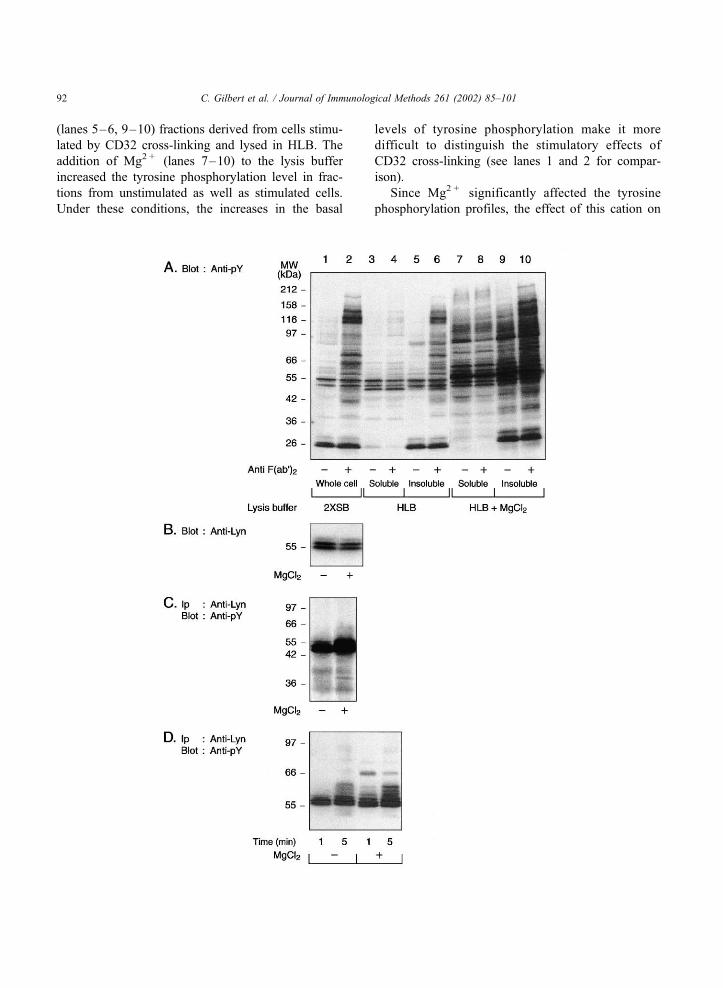
(lanes 5–6, 9–10) fractions derived from cells stimu-
lated by CD32 cross-linking and lysed in HLB. The
addition of Mg2 + (lanes 7–10) to the lysis buffer
increased the tyrosine phosphorylation level in frac-
tions from unstimulated as well as stimulated cells.
Under these conditions, the increases in the basal
levels of tyrosine phosphorylation make it more
difficult to distinguish the stimulatory effects of
CD32 cross-linking (see lanes 1 and 2 for compar-
ison).
Since Mg2 + significantly affected the tyrosine
phosphorylation profiles, the effect of this cation on
C. Gilbert et al. / Journal of Immunological Methods 261 (2002) 85–10192

the activity of a tyrosine kinase known to be activated
by CD32 cross-linking, namely Lyn, was examined.
The data in panels B and C show that while equal
amounts of Lyn were solubilized in the absence or
presence of Mg2 + (panel B), the kinase recovered by
immunoprecipitation was more heavily tyrosine phos-
phorylated when Mg2 + was present during lysis and
precipitation (panel C). Hyperphosphorylation of
SHP-1 was also observed in the presence of Mg2 +
in the lysis buffer (data not shown). The effect of
Mg2 + on the catalytic activity of Lyn was then tested
in an in vitro kinase assay. The results obtained (panel
D) show that Lyn was able to autophosphorylate itself
under both sets of conditions but that the autophos-
phorylation capacity of Lyn was increased when
Mg2 + was present in the lysis buffer.
3.3. Tyrosine phosphorylated proteins are concen-
trated in the insoluble fractions
The results described in Figs. 1 and 2 revealed a
heterogeneous distribution of phosphorylated proteins
among the soluble and insoluble fractions derived
from the lysis protocols after CD32 engagement. We
therefore monitored more closely three major tyrosine
phosphorylated substrates previously identified fol-
lowing CD32 cross-linking, namely, Cbl, Syk and
CD32 itself (Rollet-Labelle et al., 2000) (Fig. 3).
Briefly, aliquots of the soluble fractions derived from
the lysis of control and CD32-stimulated cells in HLB
were resuspended in an equal volume of boiling
2�LB as described in Materials and methods. The
insoluble material was resuspended in the same vol-
ume of HLB as the soluble fractions and subsequently
in an equal volume of boiling 2�LB. Cbl, Syk and
CD32 were immunoprecipitated from both fractions
Fig. 3. Immunoprecipitation of tyrosine phosphorylated proteins
from the HLB soluble and insoluble fractions. Neutrophils (4� 107
cells/ml) were stimulated by cross-linking CD32 as indicated in
Materials and methods. The reactions were stopped by a rapid
transfer of cell aliquots in a precooled Eppendorf tube and a rapid
centrifugation. The cell pellets were resuspended in HLB and the
soluble and insoluble fractions were diluted by 1/2 in boiling
2�LB. Immunoprecipitations under denaturing conditions were
performed as described in Materials and methods with the anti-Cbl,
anti-Syk or anti-C32 antibody. Phosphorylated proteins were
revealed with an anti-phosphotyrosine antibody (upper panels)
and the amount of immunoprecipitated protein was visualized by
immunoblot with the respective antibodies (lower panels).
Fig. 2. Effect of the presence of Mg2 + in HLB on tyrosine phosphorylation and tyrosine kinase activity levels. Neutrophils (4� 107 cells/ml)
were stimulated by cross-linking CD32 as indicated in Materials and methods. The reactions were stopped by transferring cell aliquots into the
boiling 2� SB for lanes 1–2 or a rapid transfer into a precooled Eppendorf tube and a rapid centrifugation for lanes 3–10. HLB with (lanes 3–
6) or without (lanes 7–10) MgCl2 was then added to the cell pellet. Aliquots of soluble (lanes 3–4 and 7–8) and insoluble (lanes 5–6 and 9–
10) fractions were diluted in an equal volume of 2� SB. Samples were analysed by immunoblot with an anti-phosphotyrosine antibody. In
panels B–D, the unstimulated cells were lysed with the appropriate HLB buffer. In panel B, the soluble fraction or starting material of both lysis
protocols was immunoblotted with anti-Lyn antibody. For immunoprecipitation in panels C and D, 200 ml of soluble material (8� 106 of cells)
was pre-incubated with an anti-Lyn antibody. Lyn activity was determined in the presence of ATP for 1 or 5 min in a kinase buffer. Precipitates
were analysed by Western blot with an anti-phosphotyrosine antibody.
C. Gilbert et al. / Journal of Immunological Methods 261 (2002) 85–101 93
as described in Materials and methods. The precip-
itates were probed with antiphosphotyrosine antibod-
ies as well as with anti-Cbl, anti-Syk or anti-CD32
antibodies (panels A, B, C, respectively). The results
obtained showed that phosphorylated Cbl, Syk and
CD32 were highly concentrated in the insoluble frac-
tions in agreement with the observations of Fig. 1.
Reblots with anti-Cbl, anti-Syk or anti-CD32 anti-
bodies (lower panels) indicated that cross-linking of
CD32 did not grossly modify the amounts of Cbl and
Syk present in each fraction. On the other hand,
significant decreases of CD32 were observed in the
soluble fractions which were compensated, in part at
least, by increases of CD32 in the insoluble frac-
tions.
3.4. Distribution of tyrosine phosphorylated proteins
between the soluble and insoluble fractions in
neutrophils stimulated with various agonists
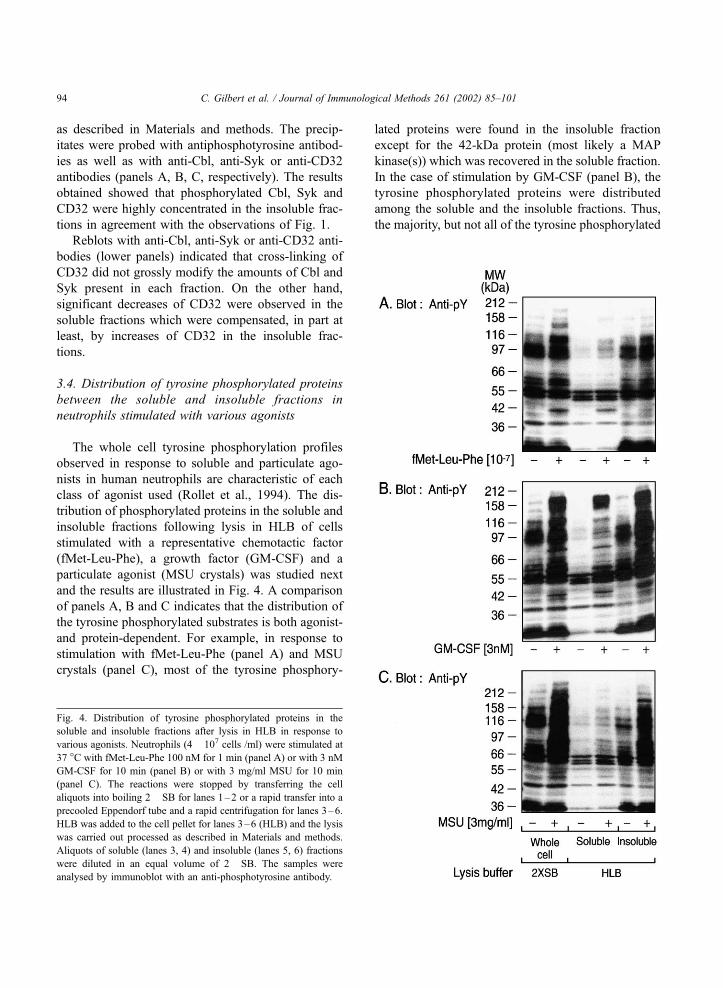
The whole cell tyrosine phosphorylation profiles
observed in response to soluble and particulate ago-
nists in human neutrophils are characteristic of each
class of agonist used (Rollet et al., 1994). The dis-
tribution of phosphorylated proteins in the soluble and
insoluble fractions following lysis in HLB of cells
stimulated with a representative chemotactic factor
(fMet-Leu-Phe), a growth factor (GM-CSF) and a
particulate agonist (MSU crystals) was studied next
and the results are illustrated in Fig. 4. A comparison
of panels A, B and C indicates that the distribution of
the tyrosine phosphorylated substrates is both agonist-
and protein-dependent. For example, in response to
stimulation with fMet-Leu-Phe (panel A) and MSU
crystals (panel C), most of the tyrosine phosphory-
lated proteins were found in the insoluble fraction
except for the 42-kDa protein (most likely a MAP
kinase(s)) which was recovered in the soluble fraction.
In the case of stimulation by GM-CSF (panel B), the
tyrosine phosphorylated proteins were distributed
among the soluble and the insoluble fractions. Thus,
the majority, but not all of the tyrosine phosphorylated
Fig. 4. Distribution of tyrosine phosphorylated proteins in the
soluble and insoluble fractions after lysis in HLB in response to
various agonists. Neutrophils (4� 107 cells /ml) were stimulated at
37 �C with fMet-Leu-Phe 100 nM for 1 min (panel A) or with 3 nM
GM-CSF for 10 min (panel B) or with 3 mg/ml MSU for 10 min
(panel C). The reactions were stopped by transferring the cell
aliquots into boiling 2� SB for lanes 1–2 or a rapid transfer into a
precooled Eppendorf tube and a rapid centrifugation for lanes 3–6.
HLB was added to the cell pellet for lanes 3–6 (HLB) and the lysis
was carried out processed as described in Materials and methods.
Aliquots of soluble (lanes 3, 4) and insoluble (lanes 5, 6) fractions
were diluted in an equal volume of 2� SB. The samples were
analysed by immunoblot with an anti-phosphotyrosine antibody.
C. Gilbert et al. / Journal of Immunological Methods 261 (2002) 85–10194
proteins in response to the four agonists investigated
(CD32, fMet-Leu-Phe, GM-CSF and MSU crystals),
were recovered in the insoluble fraction. Furthermore,
the stimulatory effects of these agonists were pre-
served in this lysis protocol as the sum of the tyrosine
phosphorylated proteins present in the soluble and the
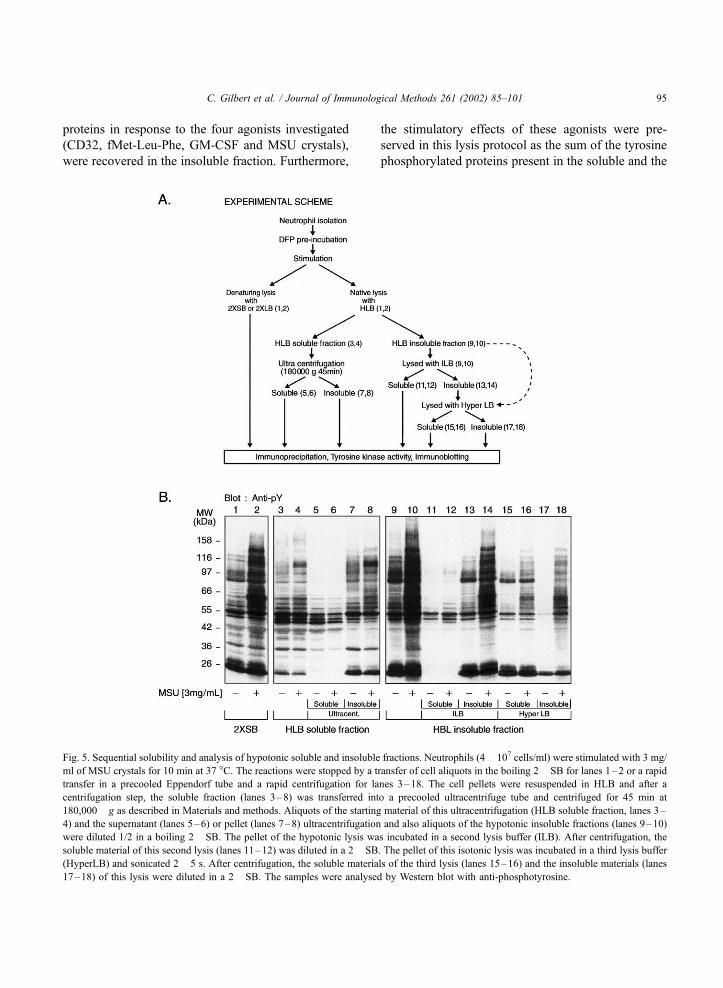
Fig. 5. Sequential solubility and analysis of hypotonic soluble and insoluble fractions. Neutrophils (4� 107 cells/ml) were stimulated with 3 mg/
ml of MSU crystals for 10 min at 37 �C. The reactions were stopped by a transfer of cell aliquots in the boiling 2� SB for lanes 1–2 or a rapid
transfer in a precooled Eppendorf tube and a rapid centrifugation for lanes 3–18. The cell pellets were resuspended in HLB and after a
centrifugation step, the soluble fraction (lanes 3–8) was transferred into a precooled ultracentrifuge tube and centrifuged for 45 min at
180,000� g as described in Materials and methods. Aliquots of the starting material of this ultracentrifugation (HLB soluble fraction, lanes 3–
4) and the supernatant (lanes 5–6) or pellet (lanes 7–8) ultracentrifugation and also aliquots of the hypotonic insoluble fractions (lanes 9–10)
were diluted 1/2 in a boiling 2� SB. The pellet of the hypotonic lysis was incubated in a second lysis buffer (ILB). After centrifugation, the
soluble material of this second lysis (lanes 11–12) was diluted in a 2� SB. The pellet of this isotonic lysis was incubated in a third lysis buffer
(HyperLB) and sonicated 2� 5 s. After centrifugation, the soluble materials of the third lysis (lanes 15–16) and the insoluble materials (lanes
17–18) of this lysis were diluted in a 2� SB. The samples were analysed by Western blot with anti-phosphotyrosine.
C. Gilbert et al. / Journal of Immunological Methods 261 (2002) 85–101 95
insoluble fractions closely corresponded to those
detected in the whole cell lysates.
3.5. Sequential solubilisation of the tyrosine phos-
phorylated proteins
Since most of the tyrosine phosphorylated proteins
were recovered in the pellets of the hypotonic lysis, a
further extraction of this pellet was warranted in order
to be able to carry out immunoprecipitations from this
fraction. On the other hand, we also wanted to better
characterize the soluble fraction of the hypotonic
lysis. The soluble and insoluble fractions of the
hypotonic lysis buffer were therefore individually
processed for subsequent analysis as indicated in
panel A of Fig. 5.
After stimulation with MSU crystals, the soluble
fraction (lanes 3–4) of the hypotonic lysis was cen-
trifuged for 45 min at 180,000� g. The resulting
soluble and insoluble fractions (lanes 5–8) were
analysed by immunoblotting with an anti-phosphotyr-
osine antibody. Two conclusions can be derived from
this analysis. First, the tyrosine phosphorylated pro-
teins were concentrated in the pellets of the ultra-
centrifugation step (lanes 7–8 vs. 5–6). Secondly,
these data confirm the stability of the tyrosine phos-
phorylation signals in the HLB soluble fractions.
The HLB insoluble fraction was then sequentially
lysed in buffers of increasing tonicities, the composi-
tion of which is described in Materials and methods.
Briefly, the HLB insoluble material was resuspended
in ILB and an aliquot was removed for direct analysis
(lanes 9–10). ILB soluble and insoluble fractions
were then prepared (lanes 11–14). The ILB insoluble
material was then resuspended in HyperLB and solu-
ble and insoluble fractions were isolated (lanes 15–
18). The relevant fractions were analyzed by immu-
noblotting with anti-phosphotyrosine antibodies
(panel B). The whole cell lysates (lanes 1–2) served
as the reference point for the extent of stimulation as
far as the global tyrosine phosphorylated profile was
concerned. As shown in Fig. 5, the majority of the
tyrosine phosphorylated proteins present in the pellet
of the HLB step remain insoluble in ILB (lanes 11 and
12 vs. lanes 13 and 14). It should be noted that the
profile of tyrosine phosphorylated proteins in the ILB
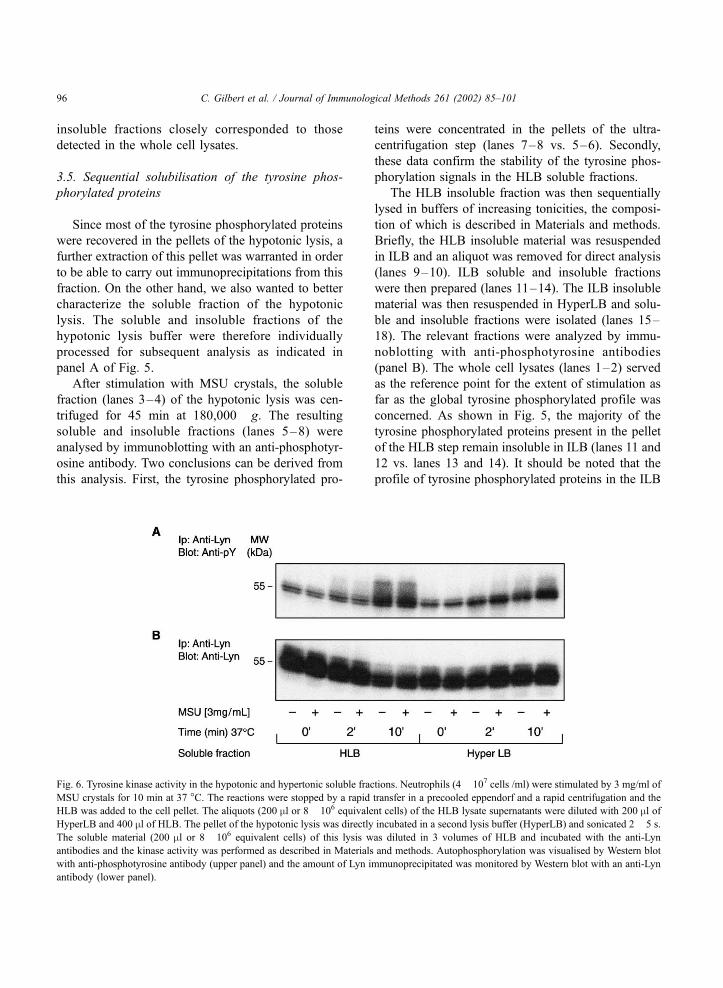
Fig. 6. Tyrosine kinase activity in the hypotonic and hypertonic soluble fractions. Neutrophils (4� 107 cells /ml) were stimulated by 3 mg/ml of
MSU crystals for 10 min at 37 �C. The reactions were stopped by a rapid transfer in a precooled eppendorf and a rapid centrifugation and the
HLB was added to the cell pellet. The aliquots (200 ml or 8� 106 equivalent cells) of the HLB lysate supernatants were diluted with 200 ml ofHyperLB and 400 ml of HLB. The pellet of the hypotonic lysis was directly incubated in a second lysis buffer (HyperLB) and sonicated 2� 5 s.
The soluble material (200 ml or 8� 106 equivalent cells) of this lysis was diluted in 3 volumes of HLB and incubated with the anti-Lyn
antibodies and the kinase activity was performed as described in Materials and methods. Autophosphorylation was visualised by Western blot
with anti-phosphotyrosine antibody (upper panel) and the amount of Lyn immunoprecipitated was monitored by Western blot with an anti-Lyn
antibody (lower panel).
C. Gilbert et al. / Journal of Immunological Methods 261 (2002) 85–10196

pellet still closely resembled that of the original
material (lanes 1 and 2). Finally, resuspension in
HyperLB did result in the solubilisation of some
proteins in the ILB pellet (lanes 15 and 16). However,
several tyrosine phosphorylated proteins remained
insoluble even under these more stringent conditions
(lanes 17–18).
A partial characterization (by immunoblotting) of
the fractions obtained during the sequential lysis
protocol followed herein was carried out (data not
shown). Cytosolic markers (myeloid related protein-
14 (MRP-14) and lactate dehydrogenase (LDH)) were
found predominantly in the soluble fraction of the first
lysis with HLB and in the soluble fractions of the
ultracentrifugation steps. The distribution of VLA2,
CD16, CD11b, CD45 or CD32 between the soluble
and insoluble fractions of HLB and HyperLB lysis
step was specific for each protein tested. Of relevance,
the receptor for cholera toxin (GM1) (Hart, 1975), a
major constituent of lipid rafts, was totally insoluble
in HLB buffer but was solubilized in HyperLB. Early
endosome-associated protein-1 (EEA-1), an endoso-
mal marker, was equally distributed between the
soluble and insoluble fractions of HLB. The HyperLB
step completely solubilized EEA-1. Lysosome-asso-
ciated membrane protein-1 (LAMP-1) was entirely
recovered in the HLB soluble fraction. Cytoskeletal
markers were also analyzed. A significant transloca-
tion of actin to the insoluble fraction was evident after
CD32 cross-linking as monitored by Coomassie blue
staining (Fig. 1). Paxillin and ezrin were equally
distributed between the soluble and the insoluble
fractions of the hypotonic lysis. Extraction with the
hypertonic lysis buffer solubilized these proteins.
Finally, nuclear markers (p62 nucleoporin) remained
insoluble and were found in the pellets of the
HyperLB step.
3.6. Lyn activity following its extraction from Hy-
perLB
In order to assess the state of the proteins extracted
in HyperLB, the immunoreactivity and enzymatic
activity of the Src kinase Lyn were tested next in
the soluble fractions of the HLB and HyperLB lysates.
We previously showed that Lyn is activated in re-
sponse to MSU crystals (Gaudry et al., 1995). We
wanted to confirm this result following the sequential
lysis described here. Neutrophils were exposed to 3
mg/ml of MSU crystals for 10 min before being lysed
in HLB. HLB soluble and insoluble fractions were
prepared as described in Fig. 5. Since we observed in
Fig. 5 that the HLB insoluble material was also
insoluble in ILB, the HLB insoluble fractions were
directly resuspended in HyperLB. Soluble and insolu-
ble fractions were prepared and Lyn was immuno-
precipitated from the HLB and HyperLB soluble
fractions. The results shown in Fig. 6 demonstrate
that Lyn immunoprecipitated from the HyperLB solu-
ble fractions retained in vitro kinase activity. How-
ever, a stimulatory effect of MSU crystals on the auto-
kinase activity of Lyn was only detectable in the
HyperLB lysates (Fig. 6, upper panel, right side).
Reblots indicated that equal amounts of Lyn were
immunoprecipitated and loaded in the control and
MSU-stimulated cells (Fig. 6, lower panel). The
results of preliminary experiments (data not shown)
indicated that the auto-kinase activity of Hck was
also increased in the soluble material of the hyper-
tonic lysis following stimulation with MSU crys-
tals.
4. Discussion
The results obtained during the course of these
studies provide a framework for the immunobiochem-
ical investigation of tyrosine phosphorylation-depend-
ent signalling pathways in human neutrophils. A
sequential lysis protocol in buffers of increasing toni-
cities is described that allows the preparation of
neutrophil lysates that retain their characteristic tyro-
sine phosphorylation profile and enzymatic activities.
This procedure also established that the detergent
solubility of individual tyrosine phosphorylated sub-
strates differed, not only according to the protein
under investigation, but also in response to the spe-
cific agonist used.
The basis of this method is an initial lysis in a
hypotonic buffer. Under these conditions, the overall
tyrosine phosphorylation profile is preserved to a
major extent. The two fractions thus obtained (soluble
and insoluble) can then be further analyzed, either
directly (in the case of the soluble fraction) or follow-
ing its solubilization in buffers of increasing tonicities
(in the case of the original insoluble fraction). Addi-
C. Gilbert et al. / Journal of Immunological Methods 261 (2002) 85–101 97

tionally, an ultra-centrifugation step can be utilized to
further characterize the fraction soluble in the hypo-
tonic lysis buffer. Each of the fractions can be ana-
lyzed by immunoblotting or by monitoring enzymatic
activities. This general protocol can be adapted to suit
individual experimental requirements.
The severe experimental problems associated with
the preparation of neutrophil lysates (proteolysis,
dephosphorylation or hyperphosphorylation) are
widely acknowledged but poorly documented. The
data presented in this report confirm and extend
previously published observations concerning the
lability of neutrophil lysates (Al-Shami et al., 1997b;
McDonald et al., 1998). The characteristic tyrosine
phosphorylation profiles observed upon direct transfer
of suspensions of neutrophils into boiling sample
buffers are grossly altered upon lysis in classical RIPA
buffers (see Al-Shami et al., 1997b; and also Fig. 1).
A previous attempt to preserve the original patterns of
tyrosine phosphorylation relied on a denaturing lysis
protocol which, while effective, eliminated the possi-
bility of studying protein–protein interactions and
enzymatic activities (i.e., kinases and phosphatases)
in the lysates (Al-Shami et al., 1997b). The sequential
lysis protocol described in the present manuscript
overcomes these limitations in that lysates prepared
under native conditions maintain to a significant
extent their profile of tyrosine phosphorylation as well
as their enzymatic activities.
Furthermore, whereas tyrosine phosphorylation is
transient in neutrophils, we observed that tyrosine
phosphorylation in the insoluble fraction is well con-
served even 60 min following cell lysis. This can be
explained first by the technique used to stop the
stimulation, i.e. rapid transfer of the cells in pre-
cooled tubes followed by rapid centrifugation. This
approach immediately eliminates the incubation me-
dium which may contain degradative products (pro-
teases, phosphatases) released by the cells during
stimulation.
Cross-linking of CD32 induces its insolubility as
described in Figs. 1 and 3. Insolubility in non-ionic
detergent coupled with concentration of tyrosine
phosphorylated specific substrates (Cbl, Syk, and
CD32; Figs. 1 and 3) in the HLB insoluble fractions
suggests the implication of the lipid raft structures or
detergent resistant membranes (DRMs). Insolubilisa-
tion of membrane receptors coupled to kinase activ-
ities was also observed in T cell lines (Solomon et al.,
1998) and in various others cells (Zhou et al., 1995).
One of the main findings of the present investiga-
tion was that the detergent extractability of the tyro-
sine phosphorylated substrates was both substrate and
agonist dependent. Nevertheless, a large percentage of
the tyrosine phosphorylated proteins was insoluble in
regular RIPA buffers, of hypo- or iso-tonicity. This
point raises questions about the interpretation of
immunoprecipitation studies in which the soluble
fraction is used as starting material in the absence of
a detailed picture of the distribution of the protein
being examined. Furthermore, the level of phospho-
rylation of various proteins can be artefactually in-
creased by the presence of Mg2 + in the lysis buffers, a
procedure sometimes adopted to help preserve mem-
brane and cytoskeletal integrity. Once again, this may
impact on the interpretation of the functional signifi-
cance of apparent stimulation, or lack of stimulation,
of the tyrosine phosphorylation of specific substrates
in those cases where the profiles of tyrosine phos-
phorylation of the lysates used as starting material for
the immunoprecipitation differ significantly from that
of whole cells.
The importance of knowing the distribution of
proteins in detergent extracts of the cells is strikingly
illustrated by the behavior of CD32, Cbl and Syk in
response to stimulation by CD32 ligation. All three
proteins were rapidly tyrosine phosphorylated follow-
ing CD32 cross-linking (Marcilla et al., 1995; Nacc-
ache et al., 1997; Rollet-Labelle et al., 2000) and, as
shown in Fig. 3, they were recovered in both the
soluble and the insoluble fractions. However, in all
three cases, the tyrosine phosphorylated proteins were
highly concentrated in the detergent-insoluble frac-
tions. The physiological relevance of the soluble form
of these proteins is therefore likely to differ from that
present in the insoluble fraction.
The functional relevance of the above considera-
tions is also highlighted by the results of the experi-
ments in which the kinase activity of Lyn was
monitored in the soluble fractions of both the hypo-
tonic and hypertonic lysis buffers. The enzyme
remained active in both fractions, an indication of
preservation of structure and function in the lysates.
Of more functional relevance, however, was the
observation that the stimulatory effects of MSU crys-
tals on the activity of Lyn were only readily detectable
C. Gilbert et al. / Journal of Immunological Methods 261 (2002) 85–10198

in the hypertonic buffer lysate. A study of the soluble
fraction of the hypotonic lysate would have missed
this effect, while a lysis in an isotonic buffer would be
likely to result in a significant level of signal distor-
tion (proteolysis, dephosphorylation). The sequential
lysis protocol characterized in the present study over-
comes several of these problems by preserving the
original phosphorylation status and by sequentially
giving access to the various fractions. These observa-
tions indicate that both fractions must always be
examined in this kind of studies. These data are
consistent with the results of Zhou et al. (1995) who
reported previously a partitionning of Src kinase
activities between the soluble and the insoluble frac-
tions derived from adherent neutrophils. These en-
zymes are involved in the early steps of several neu-
trophil responses and are differentially regulated
depending on their intracellular distribution (Welch
and Maridonneau-Parini, 1997). Our results also indi-
cate that, depending on the agonist used, the distribu-
tion of the tyrosine phosphorylated substrates varies
and must be individually characterized. Furthermore,
the addition of cofactors in the lysis buffer must be
carefully controlled.
In conclusion, we demonstrated that the detergent
insoluble fractions cannot be excluded in the bio-
chemical studies of stimulated human neutrophils.
The concentration of signalling molecules in this
fraction may explain some of the controversial results
concerning tyrosine phosphorylated proteins, tyrosine
kinase activities or protein associations which have
been reported in stimulated neutrophils. The present
approach, which is based on the examination of all
cell fractions, may help to understand the biochemical
mechanisms of neutrophil responses. This method
may also be extended to the monitoring of phospha-
tase activities if NaVO4 is omitted from the lysis
buffers allowing the evaluation of the balance of
kinase and phosphatase activities in stimulated cells
which is most likely to be a key to the regulation of
several functional responses in many cells.
Acknowledgements
Supported in part by grants and fellowships from the
Canadian Institutes of Health Research and from the
Arthritis Society of Canada. C. Gilbert was supported
by fellowships from the K.M. Hunter Charitable
Foundation, the Canadian Institutes of Health Research
and the Fonds pour la Formation de Chercheurs et
l’Aide a la Recherche and the Fonds de la Recherche en
Sante du Quebec.
The authors wish to thank Mr. Sylvain Levasseur
for his expert help with the hybridoma cultures and
the purification of antibodies and Dr. Maria Fernandes
for carefully reading this manuscript.
References
Al-Shami, A., Bourgoin, S.G., Naccache, P.H., 1997a. Granulocyte-
macrophage colony-stimulating factor-activated signaling path-
ways in human neutrophils: I. Tyrosine phosphorylation-depend-
ent stimulation of phosphatidylinositol 3-kinase and inhibition
by phorbol esters. Blood 89, 1035–1044.
Al-Shami, A., Gilbert, C., Barabe, F., Gaudry, M., Naccache, P.H.,
1997b. Preservation of the pattern of tyrosine phosphorylation in
human neutrophil lysates. J. Immunol. Methods 202, 183–191.
Al-Shami, A., Mahanna, W., Naccache, P.H., 1998. Granulocyte-
macrophage colony-stimulating factor-activated signaling path-
ways in human neutrophils. Selective activation of Jak2, Stat3,
and Stat5b. J. Biol. Chem. 273, 1058–1063.
Asahi, M., Taniguchi, T., Hashimoto, E., Inazu, T., Maeda, H.,
Yamamura, H., 1993. Activation of protein-tyrosine kinase
p72syk with concanavalin A in polymorphonuclear neutrophils.
J. Biol. Chem. 268, 23334–23338.
Barabe, F., Gilbert, C., Liao, N., Bourgoin, S.G., Naccache, P.H.,
1998. Crystal-induced neutrophil activation: VI. Involvement of
FcgammaRIIIB (CD16) and CD11b in response to inflammatory
microcrystals. FASEB J. 12, 209–220.
Borregaard, N., Cowland, J.B., 1997. Granules of the human neu-
trophilic polymorphonuclear leukocyte. Blood 89, 3503–3521.
Brumell, J.H., Chan, C.K., Butler, J., Borregaard, N., Siminovitch,
K.A., Grinstein, S., Downey, G.P., 1997. Regulation of Src
homology 2-containing tyrosine phosphatase 1 during activation
of human neutrophils. Role of protein kinase C. J. Biol. Chem.
272, 875–882.
Cassatella, M.A., 1996. Cytokines Produced by Polymorphonuclear
Neutrophils: Molecular and Biological Aspects. Chapman &
Hall, Austin.
Cui, Y., Harvey, K., Alcard, L., Jansen, J., Hughes, C., Siddiqui,
R.A., English, D., 1994. Regulation of neutrophil responses by
phosphotyrosine phosphatase. J. Immunol. 152, 5420–5428.
Cui, Y., Harvey, K.A., Siddiqui, R.A., Jansen, J., Akard, L.P.,
Thompson, J.M., Garcia, J.G., English, D., 1996. Cytosolic in-
activation of translocated neutrophil plasma membrane protein
tyrosine phosphatase. Blood 87, 341–349.
Fernandez, R., Suchard, S.J., 1998. Syk activation is required for
spreading and H2O2 release in adherent human neutrophils. J.
Immunol. 160, 5154–5162.
Fialkow, L., Chan, C.K., Downey, G.P., 1997. Inhibition of CD45
during neutrophil activation. J. Immunol. 158, 5409–5417.
C. Gilbert et al. / Journal of Immunological Methods 261 (2002) 85–101 99

Fuortes, M., Jin, W.W., Nathan, C., 1994. Beta 2 integrin-dependent
tyrosine phosphorylation of paxillin in human neutrophils trea-
ted with tumor necrosis factor. J. Cell Biol. 127, 1477–1483.
Gaudry, M., Caon, A.C., Gilbert, C., Lille, S., Naccache, P.H., 1992.
Evidence for the involvement of tyrosine kinases in the loco-
motory responses of human neutrophils. J. Leukocyte Biol. 51,
103–108.
Gaudry, M., Roberge, C.J., de Medicis, R., Lussier, A., Poubelle,
P.E., Naccache, P.H., 1993. Crystal-induced neutrophil activa-
tion: III. Inflammatory microcrystals induce a distinct pattern
of tyrosine phosphorylation in human neutrophils. J. Clin. Invest.
91, 1649–1655.
Gaudry, M., Gilbert, C., Barabe, F., Poubelle, P.E., Naccache, P.H.,
1995. Activation of Lyn is a common element of the stimulation
of human neutrophils by soluble and particulate agonists. Blood
86, 3567–3574.
Gilbert, C., Barabe, F., Rollet-Labelle, E., Bourgoin, S.G., McColl,
S.R., Damaj, B.B., Naccache, P.H., 2001. Evidence for a role for
SAM68 in the responses of human neutrophils to ligation of
CD32 and to monosodium urate crystals. J. Immunol. 166,
4664–4671.
Grinstein, S., Furuya, W., 1992. Chemoattractant-induced tyrosine
phosphorylation and activation of microtubule-associated pro-
tein kinase in human neutrophils. J. Biol. Chem. 267, 18122–
18125.
Grinstein, S., Furuya, W., Lu, D.J., Mills, G.B., 1990. Vanadate
stimulates oxygen consumption and tyrosine phosphorylation
in electropermeabilized human neutrophils. J. Biol. Chem. 265,
318–327.
Gutkind, J.S., Robbins, K.C., 1989. Translocation of the FGR pro-
tein-tyrosine kinase as a consequence of neutrophil activation.
Proc. Natl. Acad. Sci. U. S. A. 86, 8783–8787.
Hachicha, M., Naccache, P.H., McColl, S.R., 1995. Inflammatory
microcrystals differentially regulate the secretion of macrophage
inflammatory protein 1 and interleukin 8 by human neutrophils:
a possible mechanism of neutrophil recruitment to sites of in-
flammation in synovitis. J. Exp. Med. 182, 2019–2025.
Hamada, F., Aoki, M., Akiyama, T., Toyoshima, K., 1993. Associ-
ation of immunoglobulin G Fc receptor II with Src-like protein-
tyrosine kinase Fgr in neutrophils. Proc. Natl. Acad. Sci. U. S. A.
90, 6305–6309.
Hart, D., 1975. Evidence for the non-protein nature of the receptor
for the enterotoxin in Vibrio cholerae on murine lymphoid cells.
Infect. Immun. 11, 742–747.
Hundt, M., Schmidt, R.E., 1997. Functional characterization of re-
ceptor-type protein tyrosine phosphatase CD148 (HPTP eta/
DEP-1) in Fc gamma receptor IIa signal transduction of human
neutrophils. Eur. J. Immunol. 27, 3532–3535.
Ibarrola, I., Vossebeld, P.J., Homburg, C.H., Thelen, M., Roos, D.,
Verhoeven, A.J., 1997. Influence of tyrosine phosphorylation on
protein interaction with FcgammaRIIa. Biochim. Biophys. Acta
1357, 348–358.
Khamzina, L., Borgeat, P., 1998. Correlation of alpha-fetoprotein
expression in normal hepatocytes during development with ty-
rosine phosphorylation and insulin receptor expression. Mol.
Biol. Cell 9, 1093–1105.
Krump, E., Nikitas, K., Grinstein, S., 1997. Induction of tyrosine
phosphorylation and Na + /H + exchanger activation during
shrinkage of human neutrophils. J. Biol. Chem. 272, 17303–
17311.
Marcilla, A., Rivero-Lezcano, O.M., Agarwal, A., Robbins, K.C.,
1995. Identification of the major tyrosine kinase substrate in
signaling complexes formed after engagement of Fc gamma
receptors. J. Biol. Chem. 270, 9115–9120.
McDonald, P.P., McColl, S.R., Naccache, P.H., Borgeat, P., 1991.
Studies on the activation of human neutrophil 5-lipoxygenase
induced by natural agonists and Ca2 + ionosphere A23187. Bio-
chem. J. 280, 379–385.
McDonald, P.P., Bovolenta, C., Cassatella, M., 1998. Activation of
distinct transcription factors in neutrophils by bacterial LPS,
interferon gamma and GM-CSF and necessity to overcome the
action of endogenous proteases. Biochemistry 37, 13173–
13265.
Naccache, P.H., Gilbert, C., Caon, A.C., Gaudry, M., Huang, C.K.,
Bonak, V.A., Umezawa, K., McColl, S.R., 1990. Selective in-
hibition of human neutrophil functional responsiveness by erb-
statin, an inhibitor of tyrosine protein kinase. Blood 76, 2098–
2104.
Naccache, P.H., Jean, N., Liao, N.W., Bator, J.M., McColl, S.R.,
Kubes, P., 1994. Regulation of stimulated integrin surface ex-
pression in human neutrophils by tyrosine phosphorylation.
Blood 84, 616–624.
Naccache, P.H., Gilbert, C., Barabe, F., Al-Shami, A., Mahana, W.,
Bourgoin, S.G., 1997. Agonist-specific tyrosine phosphoryla-
tion of Cbl in human neutrophils. J. Leukocyte Biol. 62, 901–
910.
Nahas, N., Molski, T.F., Fernandez, G.A., Sha’afi, R.I., 1996. Ty-
rosine phosphorylation and activation of a new mitogen-acti-
vated protein (MAP)-kinase cascade in human neutrophils
stimulated with various agonists. Biochem. J. 318, 247–253.
Pouliot, M., McDonald, P.P., Krump, E., Mancini, J.A., McColl,
S.R., Weech, P.K., Borgeat, P., 1996. Colocalization of cytosolic
phospholipase A2, 5-lipoxygenase, and 5- lipoxygenase-activat-
ing protein at the nuclear membrane of A23187-stimulated hu-
man neutrophils. Eur. J. Biochem. 238, 250–258.
Pouliot, M., Gilbert, C., Borgeat, P., Poubelle, P.E., Bourgoin, S.,
Creminon, C., Maclouf, J., McColl, S.R., Naccache, P.H., 1998.
Expression and activity of prostaglandin endoperoxide synthase-
2 in agonist-activated human neutrophils. FASEB J. 12, 1109–
1123.
Roberge, C.J., Gaudry, M., de Medicis, R., Lussier, A., Poubelle,
P.E., Naccache, P.H., 1993. Crystal-induced neutrophil activa-
tion: IV. Specific inhibition of tyrosine phosphorylation by col-
chicine [see comments]. J. Clin. Invest 92, 1722–1729.
Rollet, E., Caon, A.C., Roberge, C.J., Liao, N.W., Malawista, S.E.,
McColl, S.R., Naccache, P.H., 1994. Tyrosine phosphorylation
in activated human neutrophils. Comparison of the effects of
different classes of agonists and identification of the signaling
pathways involved. J. Immunol. 153, 353–363.
Rollet-Labelle, E., Gilbert, C., Naccache, P.H., 2000. Modulation of
human neutrophil responses to CD32 cross-linking by serine/
threonine phosphatase inhibitors: cross-talk between serine/
threonine and tyrosine phosphorylation. J. Immunol. 164, 1020–
1028.
C. Gilbert et al. / Journal of Immunological Methods 261 (2002) 85–101100

Rosen, H., Michel, B.R., 1997. Redundant contribution of myelo-
peroxidase-dependent systems to neutrophil-mediated killing of
Escherichia coli. Infect. Immun. 65, 4173–4178.
Skubitz, K.M., Ahmed, K., Campbell, K.D., Skubitz, A.P., 1995.
CD50 (ICAM-3) is phosphorylated on tyrosine and is associated
with tyrosine kinase activity in human neutrophils. J. Immunol.
154, 2888–2895.
Smith, J.A., 1994. Neutrophils, host defense, and inflammation: a
double-edged sword. J. Leukocyte Biol. 56, 672.
Solomon, K.R., Mallory, M.A., Finberg, R.W., 1998. Determination
of the non-ionic detergent insolubility and phosphoprotein asso-
ciations of glycosylphosphatidylinositol-anchored proteins ex-
pressed on T cells. Biochem. J. 334, 325–333.
Tidow, N., Kasper, B., Welte, K., 1999. SH2-containing protein
tyrosine phosphatases SHP-1 and SHP-2 are dramatically in-
creased at the protein level in neutrophils from patients with
severe congenital neutropenia (Kostmann’s syndrome). Exp.
Hematol. 27, 1038–1045.
Torres, M., Hall, F.L., O’Neill, K., 1993. Stimulation of human
neutrophils with formyl-methionyl-leucyl- phenylalanine indu-
ces tyrosine phosphorylation and activation of two distinct mi-
togen-activated protein-kinases. J. Immunol. 150, 1563–1577.
Welch, H., Maridonneau-Parini, I., 1997. Lyn and Fgr are activated
in distinct membrane fractions of human granulocytic cells. On-
cogene 15, 2021–2029.
Welch, H., Mauran, C., Maridonneau-Parini, I., 1996. Nonreceptor
protein-tyrosine kinases in neutrophil activation. Methods 9,
607–618.
Willeke, T., Behrens, S., Scharffetter-Kochanek, K., Gaehtgens, P.,
Walzog, B., 2000. Beta2 integrin (CD11/CD18)-mediated sig-
naling involves tyrosine phosphorylation of c-Cbl in human
neutrophils. J. Leukocyte Biol. 68, 284–292.
Woods Ignatoski, K.M., Verderame, M., 1996. Lysis buffer compo-
sition dramatically affects extraction of phospho-tyrosine-con-
taining proteins. BioTechniques 20, 794–796.
Yan, S.R., Berton, G., 1996. Regulation of Src family tyrosine
kinase activities in adherent human neutrophils. Evidence that
reactive oxygen intermediates produced by adherent neutrophils
increase the activity of the p58c-fgr and p53/56lyn tyrosine
kinases. J. Biol. Chem. 271, 23464–23471.
Yan, S.R., Novak, M.J., 1999. Beta2 integrin-dependent phosphor-
ylation of protein-tyrosine kinase Pyk2 stimulated by tumor
necrosis factor alpha and fMLP in human neutrophils adherent
to fibrinogen [In Process Citation]. FEBS Lett. 451, 33–38.
Yan, S.R., Fumagalli, L., Berton, G., 1996. Activation of SRC
family kinases in human neutrophils. Evidence that p58C-
FGR and p53/56LYN redistributed to a Triton X-100-insoluble
cytoskeletal fraction, also enriched in the caveolar protein cav-
eolin, display an enhanced kinase activity. FEBS Lett. 380,
198–203.
Yan, S.R., Huang, M., Berton, G., 1997. Signaling by adhesion in
human neutrophils: activation of the p72syk tyrosine kinase and
formation of protein complexes containing p72syk and Src fam-
ily kinases in neutrophils spreading over fibrinogen. J. Immunol.
158, 1902–1910.
Zhou, M.J., Lublin, D.M., Link, D.C., Brown, E.J., 1995. Distinct
tyrosine kinase activation and Triton X-100 insolubility upon Fc
gamma RII or Fc gamma RIIIB ligation in human polymorpho-
nuclear leukocytes. Implications for immune complex activation
of the respiratory burst. J. Biol. Chem. 270, 13553–13560.
C. Gilbert et al. / Journal of Immunological Methods 261 (2002) 85–101 101





























