Embed Size (px)
Citation preview


Presented by Mis Karimi (PhD student of medicine immunology)
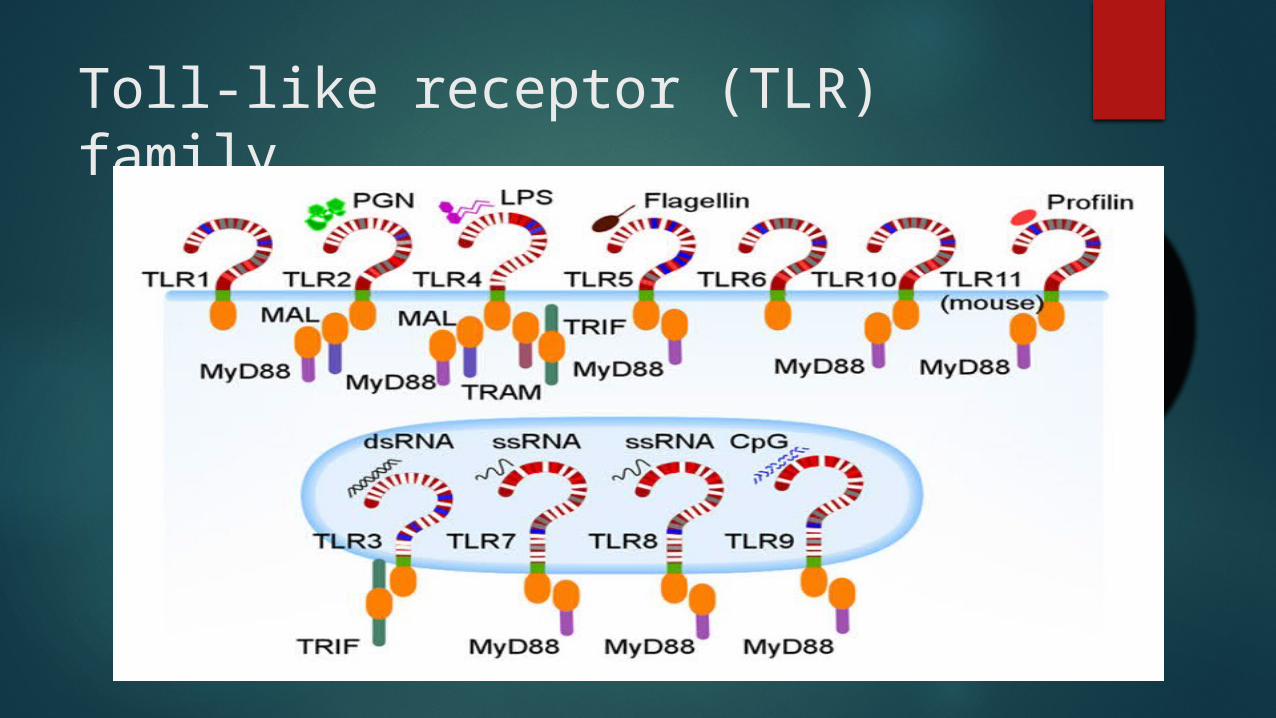
Toll-like receptor (TLR) family

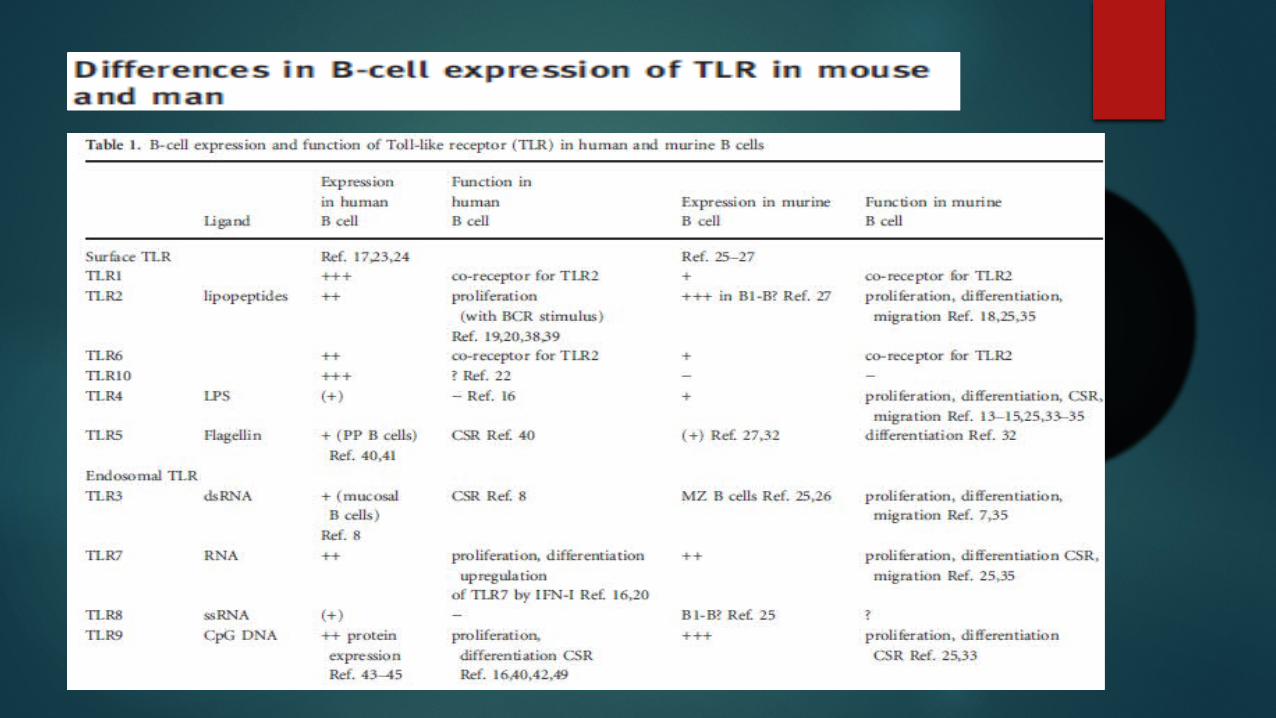
B-cell expression of TLRs
TLR expression patterns and their functions in different B-cell subsets
Expression of TLR on B lymphocytes varies depending on the mammalian species

Toll-like receptors in human B cells
Naïve human B cells do not express significant levels of TLRs unless they are pre-stimulated through the BCR
human memory B cells constitutively express TRL2, TLR6, TLR7, TLR9 and TLR10.
As in humans, TLRs are expressed differentially in B cell subsets.
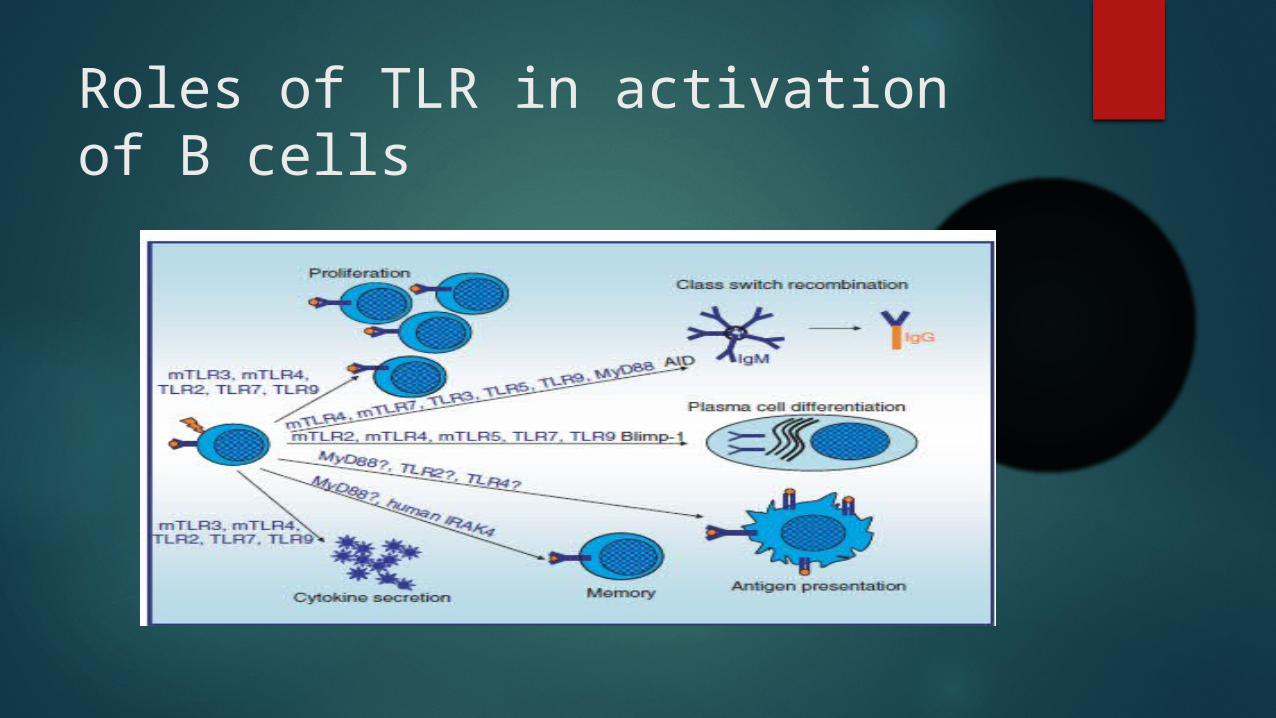
Roles of TLR in activation of B cells
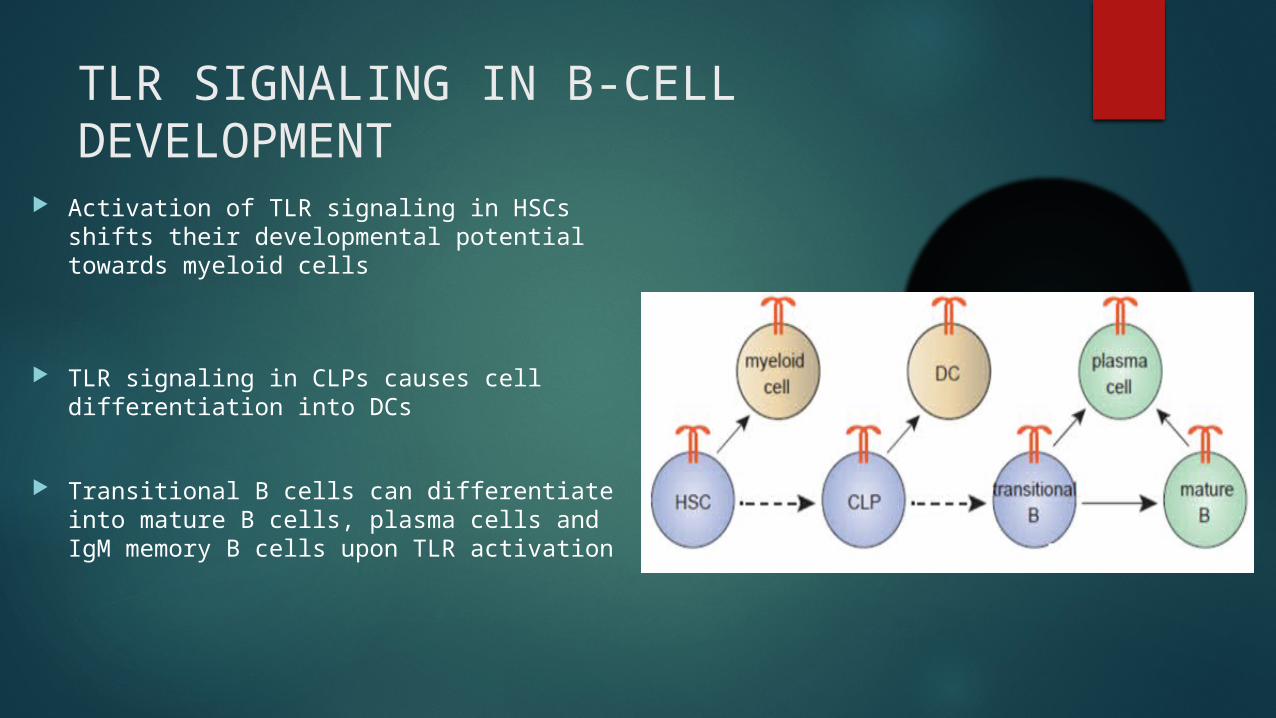
TLR SIGNALING IN B-CELL DEVELOPMENT
Activation of TLR signaling in HSCs shifts their developmental potential towards myeloid cells
TLR signaling in CLPs causes cell differentiation into DCs
Transitional B cells can differentiate into mature B cells, plasma cells and IgM memory B cells upon TLR activation
Cellular targets of TLR stimulation in human B cells
Interestingly, in the human body the local environment seems to shape the TLR repertoire
Memory B cells are more reactive and more prone to proliferate and differentiate into plasma blasts upon TLR stimulation
CpG-containing ODN, it is becoming more and more evident that only a few B-cell subsets respond to TLR9 stimulation with CpG-ODN

Furthermore, CpG-ODN supported productive CSR in naive human B cells in the presence of recombinant interleukin-10
In a recent report Rita Carsetti’s group characterized transitional human B cells as the main non-memory B-cell subset responsive to TLR9 activation
Recently, studies have described human IgM+ CD27+ B cells as the main target cells for immunostimulatory DNA ODN
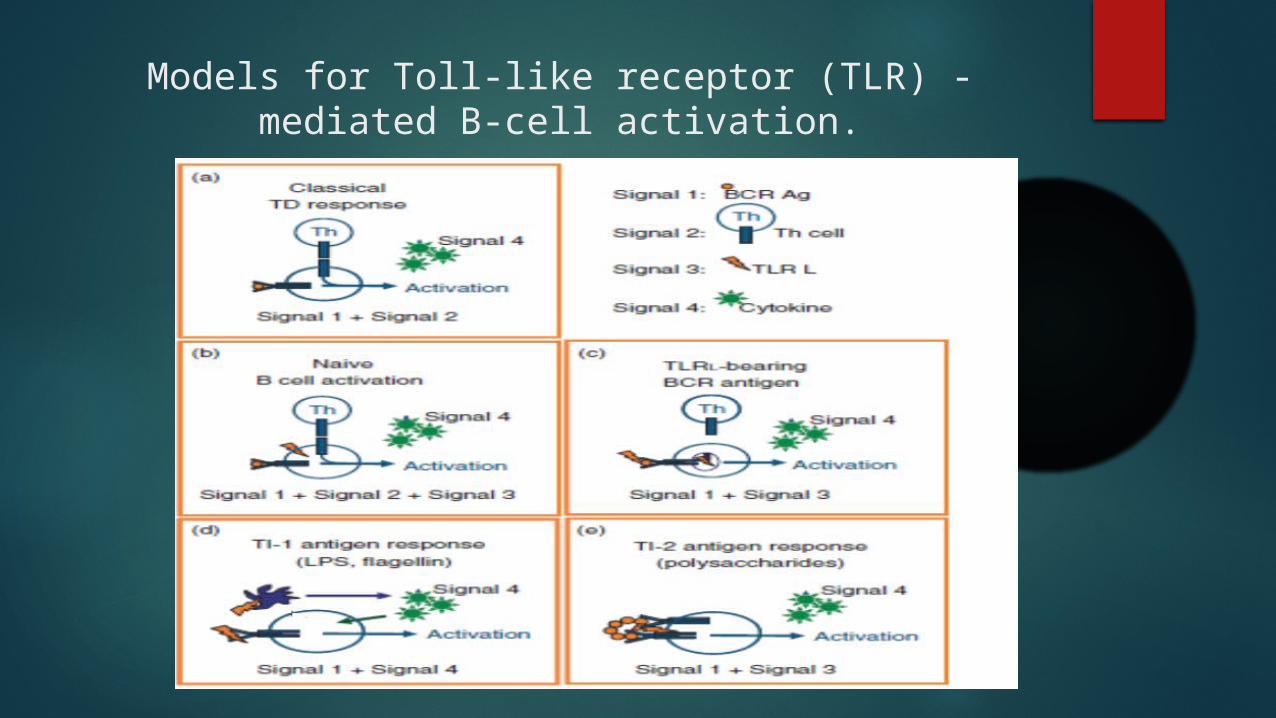
Models for Toll-like receptor (TLR) -mediated B-cell activation.

TLR signaling in B-cell differentiation
Toll-like receptors were proposed to play a role in B-cell differentiation at early stages of B-cell development
B-cell precursor maturation in vitro is supported by the early exposure to TLR4 ligands but arrested upon stimulation of TLR2
contrast, there is strong evidence that TLR regulate the differentiation of transitional and mature B cells.
TLR-mediated activation of the transcription factor NF-kB is believed to be sufficient to promote the expression of Blimp-1, a transcription factor crucial for plasma cell and pre-plasma memory B-cell differentiation.

TLR signaling in B-cell differentiation
Additionally, a recent study suggested a role for TRAF3 in TLR mediated
and antigen-mediated plasma cell differentiation and IgG secretion.
There are no publications available on the persistence
of TLR expression and the role of TLR in plasma cells.

Modulation of antibody production in vivo
TLR engagement in B cells mediates TI and TD antibody responses
Dual BCR and TLR engagement in antigen-specific B cells induces T cell-independent CSR or primes B cells for T cell-dependent CSR and SHM
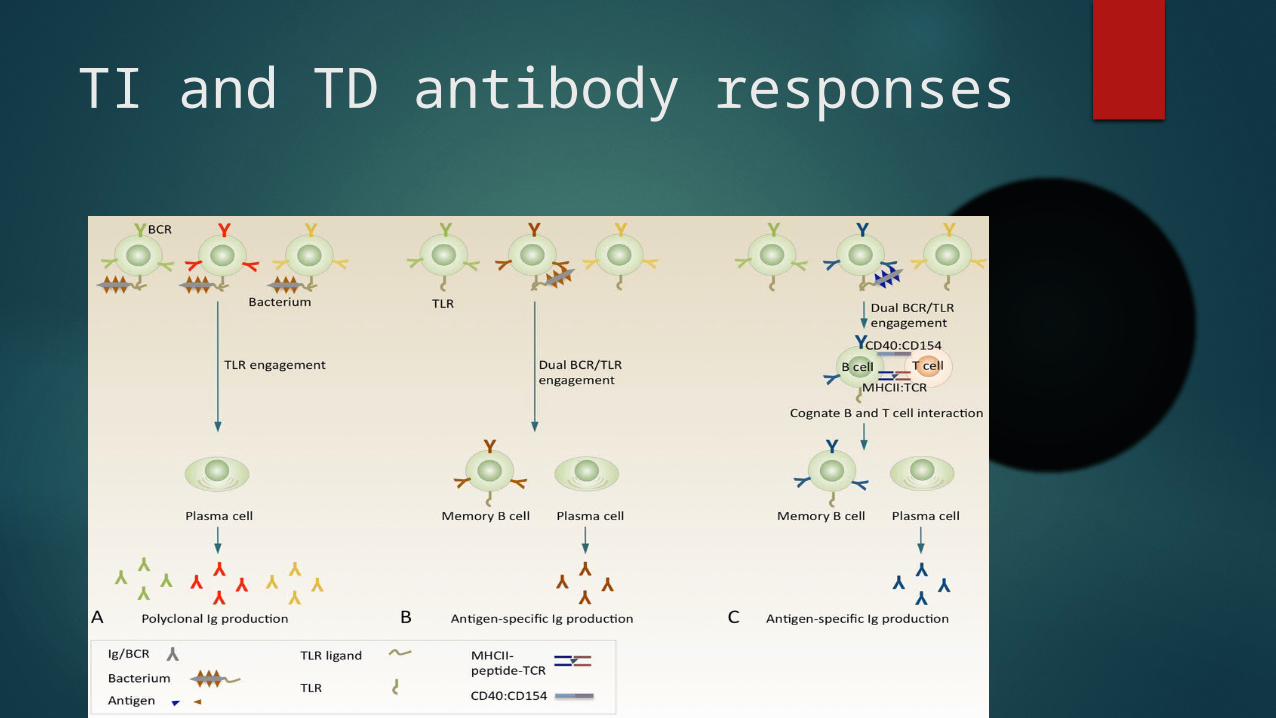
TI and TD antibody responses
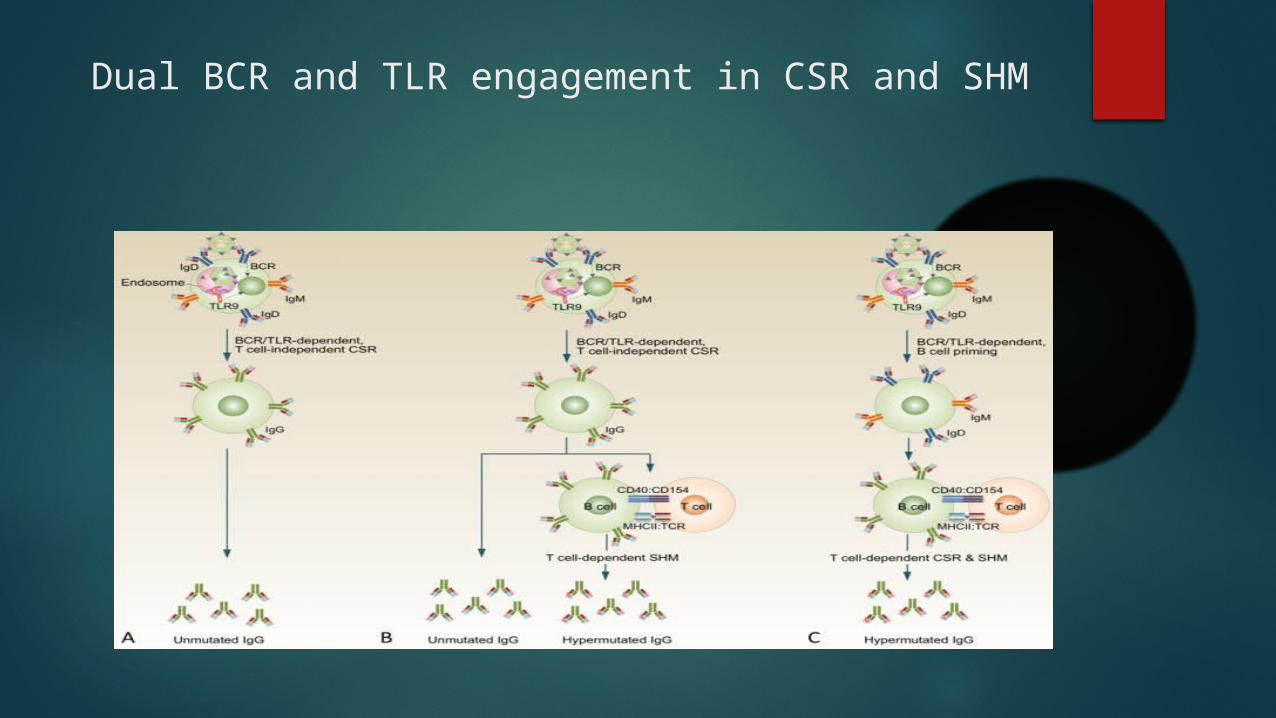
Dual BCR and TLR engagement in CSR and SHM

Modulation of CSR in vitro
In murine B cells the TLR4 ligand LPS promotes the CSR of naive B cells to IgG2b
and IgG3, and to IgG1 and IgE when combined with IL-4.
Additionally, in murine B cells, TLR9 stimulation can inhibit LPS + IL-4-
mediated CSR to IgG1 and IgE and promote IgG2a, IgG2b and IgG3 production
Modulation of CSR in vitro
In the human immune system Andrea Cerutti’s group was the first to find that CSR to IgG is induced when naive B cells are cultured in the presence of IL-10 and stimulated with CpG-ODN.
CSR to IgG and IgA was observed when human B cells isolated from
tonsils were challenged with TLR3 ligand Poly I : C and BAFF.

Modulation of CSR in vitro
Moreover, the same group reported that T-cell-independent CSR to IgA2 is most efficient in human B cells treated with the TLR5 ligand flagellin or TLR9 ligand CpG DNA in the presence of APRIL.

TLR SIGNALING IN AUTOANTIBODY PRODUCTION
Marshak-Rothstein’s group first demonstrated that a synergistic engagement of BCR and TLR9 signaling could effectively activate autoreactive B cells in vitro.
Genetic deficiency of TLR7, TLR9 or MyD88 often leads to reduced production of autoantibody, and increased expression of TLR7 causes susceptibility to autoimmune diseases.

TLR SIGNALING IN AUTOANTIBODY PRODUCTION
Shlomchick’s group generates a model of SLE, they found that the autoreactive B cells actually proliferate and undergo somatic hyper mutation in the so-called ‘extra follicular’ sites, not in GCs.
When used B cell MyD88-deficient mice crossed to a different SLE mouse model (lyn-/-) and found a strong dependence on B-cell TLR signaling for autoantibody level and associated pathological change.

TLR SIGNALING IN ANTIBODY RESPONSE TO IMMUNIZATION
In cases of antigens containing defined TLR ligands, TLR signaling does play important roles in many aspects of antibody response.
For proteins conjugated with TLR ligands, which are typical TD type antigens, the antigen-specific IgM response is also dependent on B-cell TLR signaling to some degree
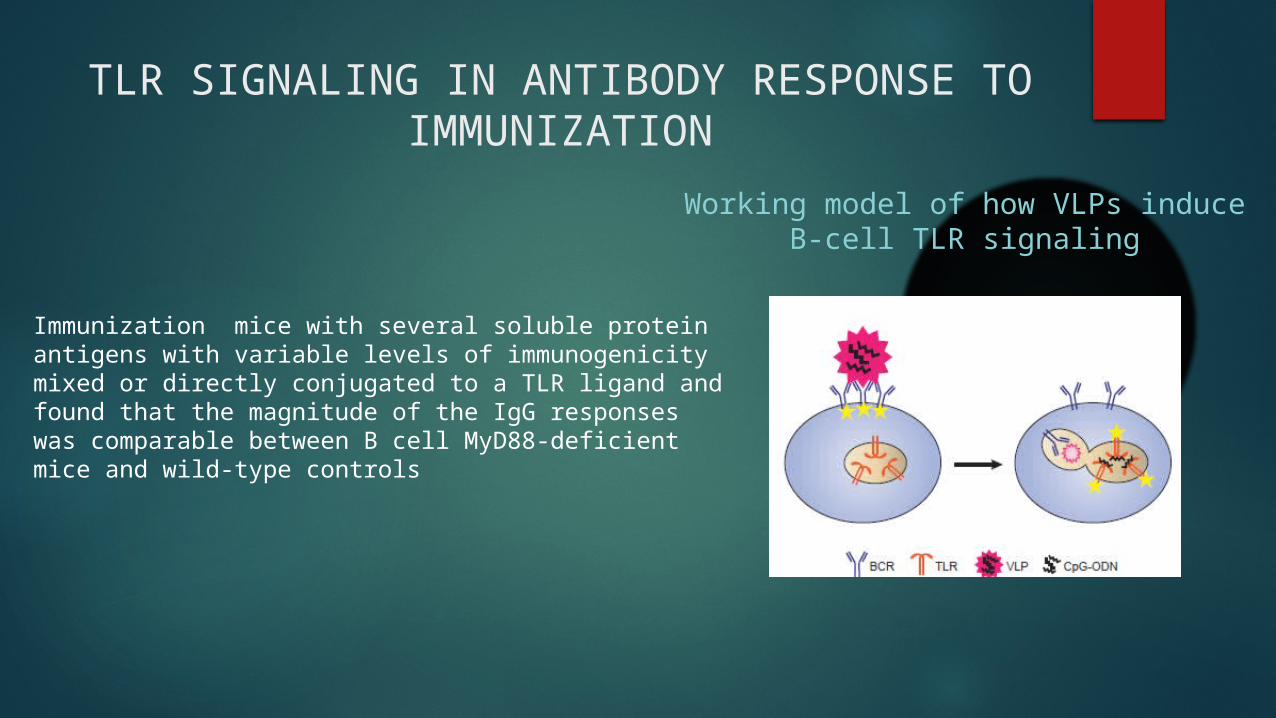
TLR SIGNALING IN ANTIBODY RESPONSE TO IMMUNIZATION
Immunization mice with several soluble protein antigens with variable levels of immunogenicity mixed or directly conjugated to a TLR ligand and found that the magnitude of the IgG responses was comparable between B cell MyD88-deficient mice and wild-type controls
Working model of how VLPs induce B-cell TLR signaling

TLR-induced cytokine secretion in B cell
Murine IL-10 secretion was detected in MZ B cells in response to single TLR2, TLR4, TLR9 or combined TLR stimulation but was absent in splenic follicular B cells whereas IL-6 was detectable in both populations.
Among the murine B-cell subpopulations tested, only follicular B cells synthesized IFN-γ in response to stimulation with TLR9 ligands combined with TLR2, TLR4 or TLR7 agonists.
TLR-induced cytokine secretion in B cell
Several reports further indicate that murine B cells produce TNF, IL-6,
IL-12, IL-23 and IL-27 in response to TLR engagement.
Human B cells are generally considered poor cytokine producers. Stimulation
with TLR7 and TLR9 ligands results in the secretion of IL-10, IL-6 and IL-8.
Further, IL-1β and IL-2 secretion have been detected in
response to CpG-ODN.
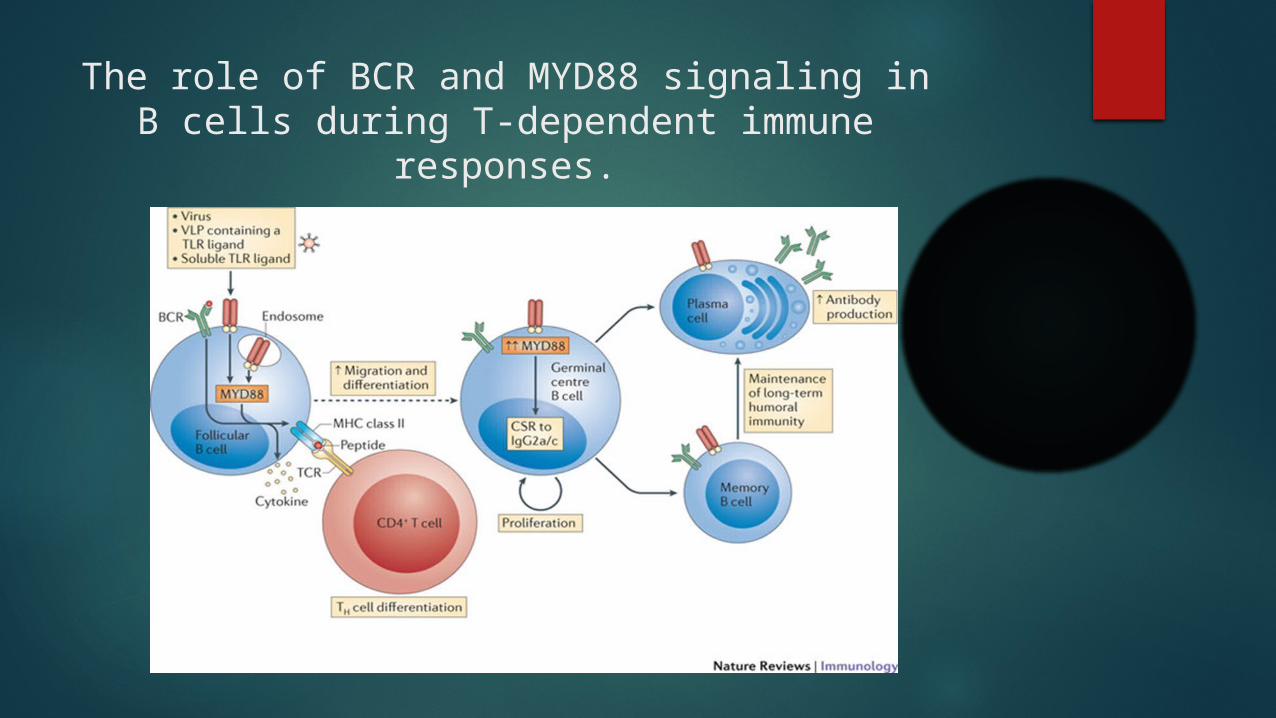
The role of BCR and MYD88 signaling in B cells during T-dependent immune responses.
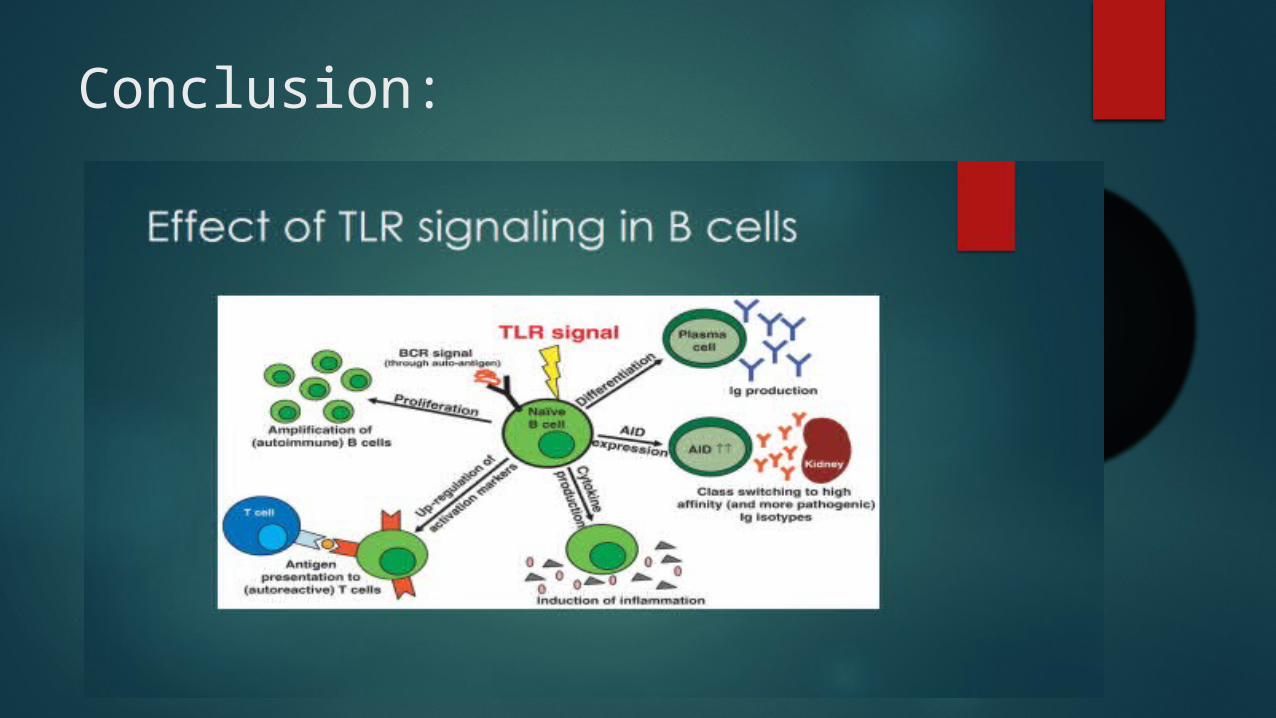
Conclusion:

Perspective•
• Understanding the mechanisms of how innate signaling contributes to humoral responses will provide a platform for developing applications, including vaccines, for human diseases
• TLRs are a potential target for therapeutic intervention in autoimmune diseases.

Thank you for
attention

References 1- Bekeredjian-Ding I, Jego G. Toll-like receptors--sentries in the B-cell
response. Immunology. 2009; 128:311–323.
2- Pone EJ, Zan H, Zhang J, Al-Qahtani A, Xu Z, Casali P. Toll-like receptors and B-cell receptors synergize to induce immunoglobulin class-switch DNA recombination: relevance to microbial antibody responses. Critical Reviews™ in Immunology. 2010;30(1).
3- Hua Z, Hou B. TLR signaling in B-cell development and activation. Cellular & molecular immunology. 2012;10(2):103-6.
4- Meyer-Bahlburg A, Rawlings DJ. B cell autonomous TLR signaling and autoimmunity. Autoimmunity reviews. 2008;7(4):313-6.

5- Hou B, Saudan P, Ott G, Wheeler ML, Ji M, Kuzmich L et al. Selective utilization of Toll-like receptor and MyD88 signaling in B cells for enhancement of the antiviral germinal center response. Immunity 2011; 34: 375–384.
6- Herlands RA, Christensen SR, Sweet RA, Hershberg U, Shlomchik MJ. T cell-independent and toll-like receptor-dependent antigendriven activation of autoreactive B cells. Immunity 2008; 29: 249–260.
7- He B, Qiao X, Cerutti A. CpG DNA induces IgG class switch DNA recombination by activating human B cells through an innate pathway that requires TLR9 and cooperates with IL-10. J Immunol 2004; 173:4479–91.